Embed Size (px)
Citation preview

Biotechnology Letters 24: 1455–1459, 2002.© 2002 Kluwer Academic Publishers. Printed in the Netherlands.
1455
Stabilization of trypsin by chemical modification with β-cyclodextrinmonoaldehyde
Michael Fernandez1, Alex Fragoso2, Roberto Cao2, Maysa Baños2, Maria L. Villalonga1 &Reynaldo Villalonga1,∗1Enzyme Technology Group, Center for Biotechnological Studies, University of Matanzas, Matanzas, C.P. 44740,Cuba2Laboratory of Bioinorganic Chemistry, Faculty of Chemistry, University of Havana, Havana 10400, Cuba∗Author for correspondence (Fax: +53 45 253101; E-mail: [email protected])
Received 28 March 2002; Revisions requested 10 April 2002; Revisions received 20 June 2002; Accepted 21 June 2002
Key words: β-Cyclodextrin, enzyme stability, modified enzyme, trypsin
Abstract
The monoaldehyde derivative of β-cyclodextrin was attached to trypsin via reductive alkylation with NaBH4. Thethermostability was enhanced from 49.5 ◦C to 60 ◦C for modified trypsin. The activation free energy of thermalinactivation at 50 ◦C was increased by 3.2 kJ mol−1. The conjugated enzyme retained 100% of its initial activityafter 3 h incubation at pH 9.
Introduction
Several chemical (Roig & Kennedy 1992, Yamazakiet al. 1999, Darias & Villalonga 2001) and chemoen-zymatic (Longo & Combes 1995, 1999) proceduresfor attaching sugar moieties to enzymes have beenreported for their increased stability. A variety ofcarbohydrate compounds, including ionic (Gómez& Villalonga 2000, Gómez et al. 2000) and non-ionic polysaccharides (Srivastava 1991, Sundaram &Venkatesh 1998), as well as low-molecular weightsugars (Marshall & Rabinowitz 1975), has been usedto prepare such neoglycoenzymes.
Cyclodextrins (CDs) are cyclic oligosaccharidescontaining six (αCD), seven (βCD) or eight (γ CD)α-1,4-linked D-glucopyranose units, with hydrophilichydroxyl groups on their outer surface and a hy-drophobic cavity in the center (Szejtli 1998). CDscan form inclusion complexes with many hydrophobiccompounds (Szejtli 1998), but little has been done toapply them to neoglycoenzymes. Although Morand &Biellmann (1991) reported the intramolecular cross-linking of α-amylase with a polyaldehyde derivative ofβCD using periodate oxidation, the extreme chemicalconditions that were needed to activate the oligosac-
charide destroyed the structure of the βCD, and con-sequently, the resulting modified enzyme can not betaken as a proper βCD derivative.
The present paper reports the reductive alkylationof bovine pancreatic trypsin (EC 3.4.21.4) with mono-6-formyl-βCD in which one primary hydroxyl grouphas been replaced by a formyl group. The effect ofthis transformation on the stability properties of theenzyme is also reported.
Materials and methods
Materials
Fractogel EMD BioSEC (S), NaBH4, N-α-benzoyl-L-arginine ethyl ester hydrochloride (BAEE) and bovinepancreatic trypsin [30 U mg−1 vs BAEE (Schwert& Takenaka 1955), 3 × 10−2 katal kg−1 vs casein(Laskowski 1955)] were obtained from Merck. βCDwas purchased from Amaizo. All other chemicals wereof analytical grade.

1456
Synthesis of monoaldehyde derivative of βCD
The monoaldehyde derivative of βCD (mono-6-formyl-βCD) was prepared in 70% yield by DMSOoxidation of the corresponding βCD monotosylate(Zhong et al. 1998) in the presence of 2,4,6-trimethylpyridine (collidine) at 130 ◦C under N2,followed by precipitation with ethanol (Yoon et al.1995). The product was characterized by conventionalNMR techniques and also by conversion into its oximederivative (Yoon et al. 1995).
Preparation of trypsin-βCD conjugate
NaBH4, 30 mg, was added to a reaction mixture con-taining 10 mg trypsin dissolved in 5 ml 50 mM sodiumphosphate buffer, pH 6, and 50 mg mono-6-formyl-βCD. The solution was stirred for 16 h in the darkat 4 ◦C, further dialyzed against 2.5 l 20 mM sodiumacetate buffer, pH 5, and then applied to a gel filtrationcolumn Fractogel EMD BioSEC (S) (2.6 × 60 cm),equilibrated with 20 mM sodium acetate buffer, pH 5,100 mM NaCl. The active fractions containing carbo-hydrates were pooled and kept at 4 ◦C.
Assays
Esterolytic activity of native and modified trypsin wasdetermined at 25 ◦C in 67 mM Tris/HCl buffer, pH 8,using BAEE as substrate (Schwert & Takenaka 1955).One unit of esterolytic activity was defined as theamount of enzyme that hydrolyses 1 µmol BAEE perminute at 25 ◦C. Proteolytic activity was determinedas described by Laskowski (1955) using milk caseinas substrate. Total carbohydrates were determined bythe phenol/sulfuric acid method using glucose as stan-dard. The degree of modification of amino groups wasdetermined by measuring the amount of free aminogroups with o-phthalaldehyde using glycine as stan-dard (Bruneel & Schacht 1993). Protein concentrationwas estimated by the Lowry method using bovineserum albumin as standard.
Results and discussion
The modifying agent used in this work was synthe-sized by oxidizing mono-6-O-tosyl-βCD, preparedaccording to Zhong et al. (1998), with DMSO inthe presence of collidine (Yoon et al. 1995). Thismethod afforded a cyclodextrin derivative contain-ing one formyl group per molecule at the 6-position
of the glucopyranose ring, thus keeping the conicalshape and the complexing properties of the molecule.The reactivity of the formyl group was checked byreaction with hydroxylamine. The activated oligosac-charide was further attached to the free amino groupsof bovine pancreatic trypsin by reductive alkylation inpresence of NaBH4. The overall synthetic method isillustrated in Scheme 1.
Scheme 1. Preparation of βCD-trypsin conjugate. Tryp representsthe enzyme
By this synthetic procedure, 4 out of the 15 NH2groups of trypsin reacted with the monoaldehydederivative of βCD as revealed by the o-phthalaldehydeassay (Bruneel & Schacht 1993). This result agreeswith the amount of carbohydrate quantified in theconjugated enzyme. The sugar content for the βCD-trypsin conjugate was determined as 20% by weightof transformed protease, representing an average of4 mol βCD attached to each mol of protein.
The esterolytic and proteolytic activity retainedfor trypsin after modification with the oligosaccharidewas determined as 60% and 57%, respectively. Thisreduction could be originated by the steric hindranceproduced by the attached βCD residues to the diffu-sion of substrate to the active site of trypsin. Othercauses could be the chemical conditions used in thecoupling reaction, e.g. alkaline pH and reductive con-ditions. However, the reduction in enzymatic activityshowed by the modified enzyme was compensatedby the improved stability properties showed by thisadduct.
The thermal stability profile of native and βCD-modified trypsin was determined by incubation during10 min at different temperatures, as illustrated in Fig-ure 1. Under these conditions, the conjugated enzyme
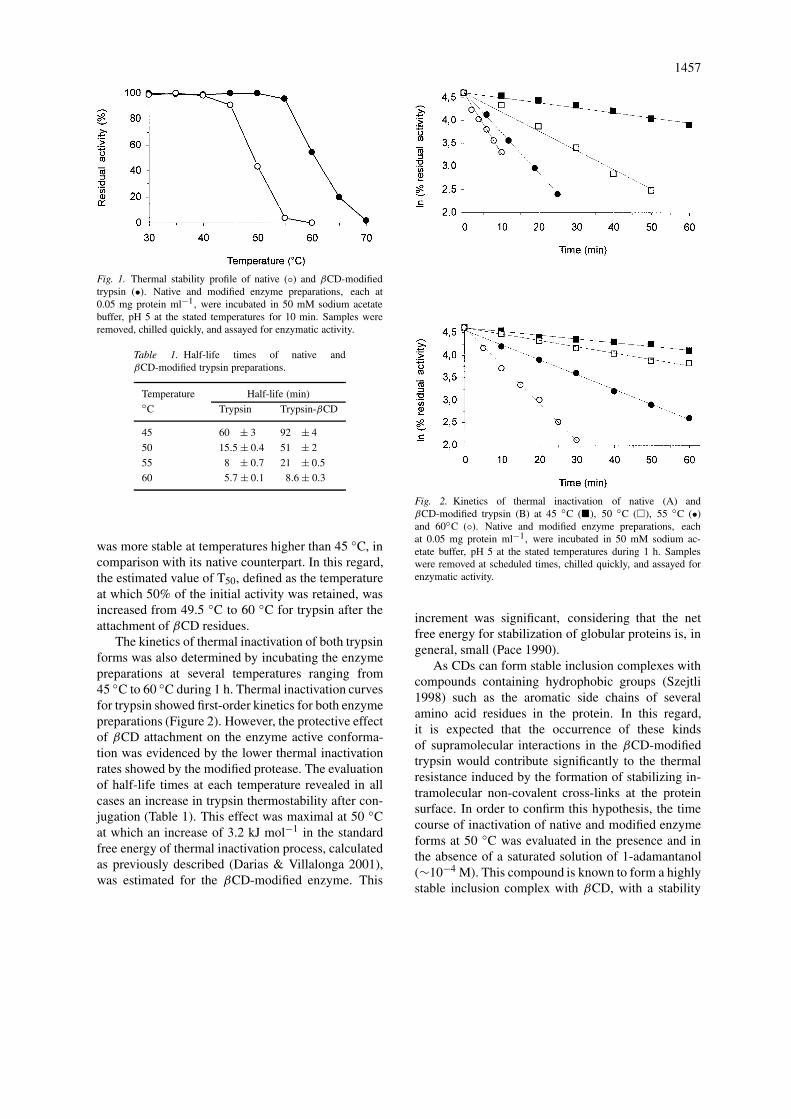
1457
Fig. 1. Thermal stability profile of native (◦) and βCD-modifiedtrypsin (•). Native and modified enzyme preparations, each at0.05 mg protein ml−1, were incubated in 50 mM sodium acetatebuffer, pH 5 at the stated temperatures for 10 min. Samples wereremoved, chilled quickly, and assayed for enzymatic activity.
Table 1. Half-life times of native andβCD-modified trypsin preparations.
Temperature Half-life (min)◦C Trypsin Trypsin-βCD
45 60 ± 3 92 ± 4
50 15.5 ± 0.4 51 ± 2
55 8 ± 0.7 21 ± 0.5
60 5.7 ± 0.1 8.6 ± 0.3
was more stable at temperatures higher than 45 ◦C, incomparison with its native counterpart. In this regard,the estimated value of T50, defined as the temperatureat which 50% of the initial activity was retained, wasincreased from 49.5 ◦C to 60 ◦C for trypsin after theattachment of βCD residues.
The kinetics of thermal inactivation of both trypsinforms was also determined by incubating the enzymepreparations at several temperatures ranging from45 ◦C to 60 ◦C during 1 h. Thermal inactivation curvesfor trypsin showed first-order kinetics for both enzymepreparations (Figure 2). However, the protective effectof βCD attachment on the enzyme active conforma-tion was evidenced by the lower thermal inactivationrates showed by the modified protease. The evaluationof half-life times at each temperature revealed in allcases an increase in trypsin thermostability after con-jugation (Table 1). This effect was maximal at 50 ◦Cat which an increase of 3.2 kJ mol−1 in the standardfree energy of thermal inactivation process, calculatedas previously described (Darias & Villalonga 2001),was estimated for the βCD-modified enzyme. This
Fig. 2. Kinetics of thermal inactivation of native (A) andβCD-modified trypsin (B) at 45 ◦C (�), 50 ◦C (�), 55 ◦C (•)and 60◦C (◦). Native and modified enzyme preparations, eachat 0.05 mg protein ml−1, were incubated in 50 mM sodium ac-etate buffer, pH 5 at the stated temperatures during 1 h. Sampleswere removed at scheduled times, chilled quickly, and assayed forenzymatic activity.
increment was significant, considering that the netfree energy for stabilization of globular proteins is, ingeneral, small (Pace 1990).
As CDs can form stable inclusion complexes withcompounds containing hydrophobic groups (Szejtli1998) such as the aromatic side chains of severalamino acid residues in the protein. In this regard,it is expected that the occurrence of these kindsof supramolecular interactions in the βCD-modifiedtrypsin would contribute significantly to the thermalresistance induced by the formation of stabilizing in-tramolecular non-covalent cross-links at the proteinsurface. In order to confirm this hypothesis, the timecourse of inactivation of native and modified enzymeforms at 50 ◦C was evaluated in the presence and inthe absence of a saturated solution of 1-adamantanol(∼10−4 M). This compound is known to form a highlystable inclusion complex with βCD, with a stability
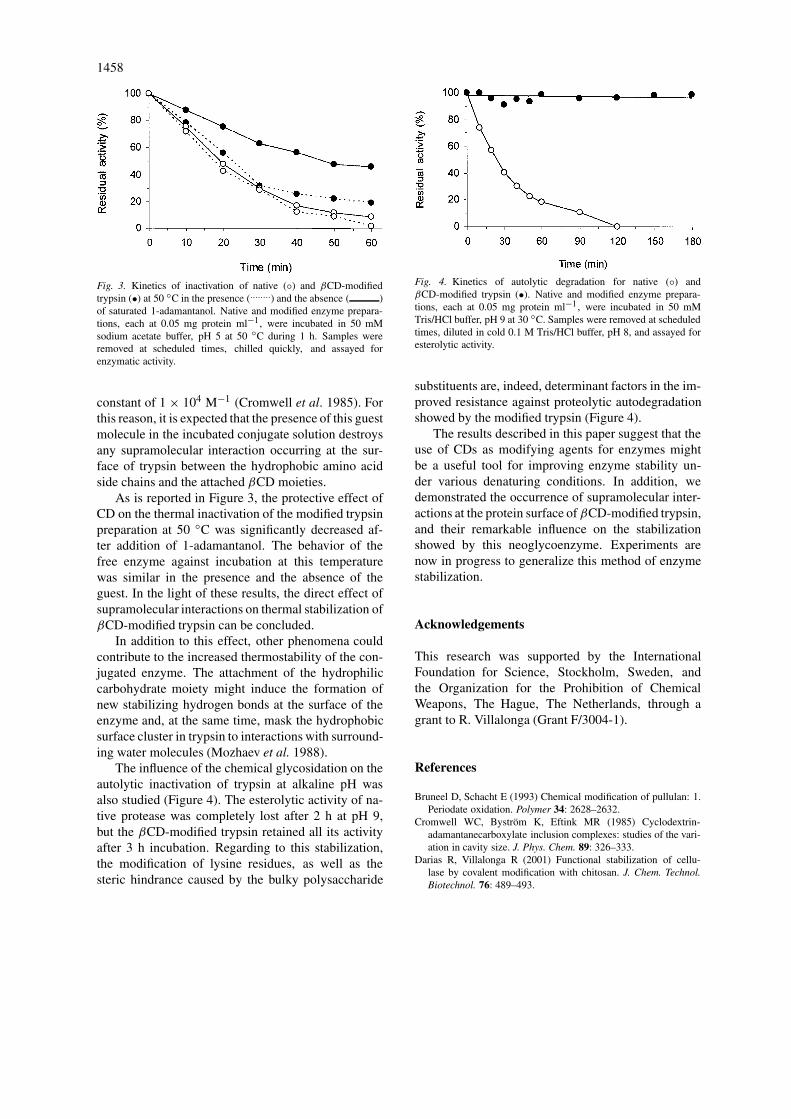
1458
Fig. 3. Kinetics of inactivation of native (◦) and βCD-modifiedtrypsin (•) at 50 ◦C in the presence (........) and the absence ( )of saturated 1-adamantanol. Native and modified enzyme prepara-tions, each at 0.05 mg protein ml−1, were incubated in 50 mMsodium acetate buffer, pH 5 at 50 ◦C during 1 h. Samples wereremoved at scheduled times, chilled quickly, and assayed forenzymatic activity.
constant of 1 × 104 M−1 (Cromwell et al. 1985). Forthis reason, it is expected that the presence of this guestmolecule in the incubated conjugate solution destroysany supramolecular interaction occurring at the sur-face of trypsin between the hydrophobic amino acidside chains and the attached βCD moieties.
As is reported in Figure 3, the protective effect ofCD on the thermal inactivation of the modified trypsinpreparation at 50 ◦C was significantly decreased af-ter addition of 1-adamantanol. The behavior of thefree enzyme against incubation at this temperaturewas similar in the presence and the absence of theguest. In the light of these results, the direct effect ofsupramolecular interactions on thermal stabilization ofβCD-modified trypsin can be concluded.
In addition to this effect, other phenomena couldcontribute to the increased thermostability of the con-jugated enzyme. The attachment of the hydrophiliccarbohydrate moiety might induce the formation ofnew stabilizing hydrogen bonds at the surface of theenzyme and, at the same time, mask the hydrophobicsurface cluster in trypsin to interactions with surround-ing water molecules (Mozhaev et al. 1988).
The influence of the chemical glycosidation on theautolytic inactivation of trypsin at alkaline pH wasalso studied (Figure 4). The esterolytic activity of na-tive protease was completely lost after 2 h at pH 9,but the βCD-modified trypsin retained all its activityafter 3 h incubation. Regarding to this stabilization,the modification of lysine residues, as well as thesteric hindrance caused by the bulky polysaccharide
Fig. 4. Kinetics of autolytic degradation for native (◦) andβCD-modified trypsin (•). Native and modified enzyme prepara-tions, each at 0.05 mg protein ml−1, were incubated in 50 mMTris/HCl buffer, pH 9 at 30 ◦C. Samples were removed at scheduledtimes, diluted in cold 0.1 M Tris/HCl buffer, pH 8, and assayed foresterolytic activity.
substituents are, indeed, determinant factors in the im-proved resistance against proteolytic autodegradationshowed by the modified trypsin (Figure 4).
The results described in this paper suggest that theuse of CDs as modifying agents for enzymes mightbe a useful tool for improving enzyme stability un-der various denaturing conditions. In addition, wedemonstrated the occurrence of supramolecular inter-actions at the protein surface of βCD-modified trypsin,and their remarkable influence on the stabilizationshowed by this neoglycoenzyme. Experiments arenow in progress to generalize this method of enzymestabilization.
Acknowledgements
This research was supported by the InternationalFoundation for Science, Stockholm, Sweden, andthe Organization for the Prohibition of ChemicalWeapons, The Hague, The Netherlands, through agrant to R. Villalonga (Grant F/3004-1).
References
Bruneel D, Schacht E (1993) Chemical modification of pullulan: 1.Periodate oxidation. Polymer 34: 2628–2632.
Cromwell WC, Byström K, Eftink MR (1985) Cyclodextrin-adamantanecarboxylate inclusion complexes: studies of the vari-ation in cavity size. J. Phys. Chem. 89: 326–333.
Darias R, Villalonga R (2001) Functional stabilization of cellu-lase by covalent modification with chitosan. J. Chem. Technol.Biotechnol. 76: 489–493.

1459
Gómez L, Villalonga R (2000) Functional stabilization of invertaseby covalent modification with pectin. Biotechnol. Lett. 22: 1191–1195.
Gómez L, Ramírez HL, Villalonga R (2000) Stabilization of inver-tase by modification of sugar chains with chitosan. Biotechnol.Lett. 22: 347–350.
Laskowski M (1955) Trypsinogen and trypsin. Meth. Enzymol. 2:26–36.
Longo MA, Combes D (1995) A novel chemoenzymatic glycosyla-tion strategy: application to lysozyme modification. FEBS Lett.375: 63–66.
Longo MA, Combes D (1999) Thermostability of modified en-zymes: a detailed study. J. Chem. Technol. Biotechnol. 74:25–32.
Marshall JJ, Rabinowitz ML (1975) Enzyme stabilization by cova-lent attachment of carbohydrate. Arch. Biochem. Biophys. 167:777–779.
Morand P, Biellman J-F (1991) Modification of α-amylase fromBacillus licheniformis by the polyaldehyde derived from β-cyclodextrine and α-amylase thermostability. FEBS Lett. 289:148–150.
Mozhaev VV, Šikšnis VA, Melik-Nubarov NS, Galkantaite NZ, De-nis GJ, Butkus EP, Zaslavsky BY, Mestechkina NM, Martinek K(1988) Protein stabilization via hydrophilization. Covalent mod-ification of trypsin and α-chymotrypsin. Eur. J. Biochem. 173:147–154.
Pace CN (1990) Conformational stability of globular proteins.Trends Biochem. Sci. 15: 14–17.
Roig MG, Kennedy JF (1992) Perspectives for chemical modifica-tions of enzymes. CRC Crit. Rev. Biotechnol. 12: 391–412.
Schwert GB, Takenaka Y (1955) A spectrophotometric determina-tion of trypsin and chymotrypsin. Biochim. Biophys. Acta 16:570–575.
Srivastava RAK (1991) Studies on stabilization of amylase bycovalent coupling to soluble polysaccharides. Enzyme Microb.Technol. 13: 164–170.
Sundaram PV, Venkatesh R (1998) Retardation of thermal and ureainduced inactivation of α-chymotrypsin by modification withcarbohydrate polymers. Protein Eng. 11: 691–698.
Szejtli J (1998) Introduction and general overwiew of cyclodextrins.Chem. Rev. 98: 1743–1754.
Yamazaki T, Tsugawa W, Sode K (1999) Increased thermal stabilityof glucose dehydrogenase by cross-linking chemical modifica-tion. Biotechnol. Lett. 21: 199–202.
Yoon J, Hong S, Martin KA, Czarnik AW (1995) A general methodfor the synthesis of cyclodextrinyl aldehydes and carboxylicacids. J. Org. Chem. 60: 2792–2795.
Zhong N, Byun HS, Bittman R (1998) A convenient methodfor the monotosylation of β-cyclodextrin on the primary side.Tetrahedron Lett. 38: 2919–2920.





























