Embed Size (px)
Citation preview

THE JCURNAL ~~BIOLOGICAL CHEMISTRY Vol. 247, No. 24, Issue of December 25, pp. 8078-8084, 1972
Printed in U.S.A.
Steroid Binding Properties of Some Peptide Fragments of
Bovine Serum Albumin Obtained on Peptic Digestion*
(Received for publication, August 28, 1972)
WILLIAM H. PEARLMANAND I.F.F.FoNG
From the Department of Pharmacology, School of Medicine, University of North Carolina, Chapel Hill, North Carolina 27514
SUMMARY
To elucidate the structural characteristics of bovine serum albumin (BSA) which allow for steroid binding, BSA was digested with pepsin under conditions which afford maximal retention of steroid binding activity consistent with a maxi- mal degree of peptide fragmentation. Digestion with pepsin- BSA (w/w), 1:3000, pH 3.0, for 2 hours at 25” afforded a complex mixture of peptide fragments with molecular weights of about 14,000 (a major class of components), and 27,000 and 34,000 (minor classes) estimated on sodium dodecyl sulfate gel electrophoresis of the digest. As much as 40 to 50% of the initial steroid binding activity was retained in the BSA digest; the steroids studied were progesterone, testosterone, and 17/3-estradiol. Steroid binding activity was empirically defined as the product of the ratio of bound to unbound steroid times the reciprocal of the total protein or peptide concentration (gram per liter) in an assay system containing Sephadex G-25 or G-10 under conditions of equilibrium.
The crude BSA digest was treated successively with 2% and 10% trichloroacetic acid; the respective trichloroacetic acid-precipitable peptides were chromatographed on Sepha- dex G-75. The 2% trichloroacetic acid chromatographic fractions exhibited a relatively high progesterone binding activity, and also appreciable testosterone and estradiol binding, whereas successive chromatographic fractions of the 10 % trichloroacetic acid fraction exhibited diminishing steroid binding activity. Binding activity correlated with the absorbance ratio, 280:258 nm, of the respective chro- matographic fractions, suggesting that peptide fragments which are richer in tyrosine tend to retain steroid binding activity to a greater degree. Although the bulk of the pep- tide material remains to be resolved, two peptide fragments, KL and VI, were isolated from the 2% trichloroacetic acid and 10 % trichloroacetic acid fractions, respectively. Pep- tide KL (mol wt 10,050) is rich in tyrosine. The equilibrium constant, nk, for the formation of a complex of peptide KL with progesterone, testosterone, or 17&estradiol at 25” was about 0.44, 0.18, or 0.33 x lo4 M-I, respectively. Peptide VT (mol wt 2766) appears to be identical with the NHz-
* This investigation was supported by Research Grant GB-27492 from the National Science Foundation and Research Grant CA- 11628 from the National Cancer Institute, United States Public Health Service. A preliminary account of this investigation has been presented (1).
terminal or Asp fragment of BSA previously isolated by Peters and Hawn ((1967) J. Biol. Chem. 242, 1566). Pep- tide Vl contains no tyrosine and exhibits very little steroid- binding affinity.
The binding of steroid sex hormones to plasma proteins (2) constitutes an important mechanism for regulating the phys- iologically effective level of these hormones in the peripheral circulation. Binding is due largely to serum albumin (3) which has a low binding affinity (nk N- lo4 I@) but a high binding capacity (nP) since the molar concentration (P) of this protein is high, i.e. 0.5 x 10e3 M. A significant role in steroid binding has also been ascribed to a testosterone (and estradiol) binding component (4) of serum and to corticosteroid-binding globulin (which also binds progesterone (5)); these proteins, albeit present in low concentration (< 10d6 M) in human serum, exhibit a high binding affinity (- lo8 I@).
The precise structure and conformation of the steroid mole- cule which allow formation of specific steroid-macromolecular complexes are well established (3). There is, on the other hand, little information similarly regarding the protein molecule in such interactions. Bovine serum albumin appears to be emi- nently suitable as a model protein for such study because it closely resembles human serum albumin in important respects and is abundantly available in crystalline form. Moreover, the chemical structure and conformation of serum albumin have been studied intensively, particularly in connection with the remarkable ability of this protein to form reversible complexes with a variety of smaII ions and molecuIes of diverse configura- tion (6, 7). I f BSA,l a single polypeptide chain (7) containing about 600 ammo acids and 17 disulfide bonds, could be frag- mented into relatively small polypeptides and yet retain an ap- preciable degree of steroid binding activity, such peptide frag- ments might be more amenable to further investigation. In this experimental approach we found guidance in the reports of Weber and Young (8) on the fragmentation of BSA by pepsin and the physicochemical properties of such digests, including binding of the polycyclic anion, anihnonaphthalenesulfonic acid; and especially, the closely related studies of Peters and CO-
workers (9, 10) which led to the isolation of an NHQ-terminal peptide fragment with the copper binding properties of intact
i The abbreviations used are: BSA, bovine serum albumin; SDS, sodium dodecyl sulfate.
8078
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

BSA. In this paper, we describe experimental conditions for peptic digestion of BSA which afford peptide fragments with steroid binding properties. Although the bulk of this complex peptide material remains to be resolved, two small peptide fragments, one of which (“KL”) exhibits an appreciable binding affinity for progesterone, were isolated and the amino acid com- position determined.
EXPERIMENTAL PROCEDURES
Materials
Bovine serum albumin (crystalline) and pepsin (three times crystallized) were obtained from Pentex. [ 1,2-3H]Progesterone,
loo R\ pepsin : BSA (w/w) I :3000 -
..n , : 300 .__...__..___
0 20 40 60 80 100 120 140 160 180 DIGESTION TIME (MINUTES)
FIG. 1. Relative steroid binding activity during the course of peptic digestion of 5% BSA, pH 3.0, at 25’. Portions of the digest were removed at stated intervals and assayed for steroid binding activity relative to that of intact BSA. For further details, see “Experimental Procedures.”
8079
[l ,2-3H]testosterone, and 17/3-[6,7-3H]estradio1 were obtained from New England Nuclear or Amersham-Searle; the specific activity was 50, 50, and 40 Ci per mmole, respectively.
Peptic Digestion of BSA
Preliminary Studies-One-gram amounts of BSA were digested in preliminary studies, see Figs. 1 to 5, under experimental con- ditions previously described (8). BSA (1 g) was dissolved in 20 ml of distilled water, and after adjusting the solution with about 0.45 ml of 2 N HCl to pH 3.0, 1 ml of a 0.2 M sodium cit- rate-phosphate buffer, pH 3.0, was added. To this solution maintained at 25” was added 0.1 ml of a pepsin solution in the same buffer; the ratio, pepsin-BSA (w/w) was 1:3000 and, in some experiments as noted, 300. The incubation period was variable as noted. Digestion was stopped by adjusting the in- cubation mixture with 1 M K2HP04 (0.4 ml per ml of digest) to pH 7.4; and, to ensure denaturation of pepsin (lo), this pH was maintained for 1 hour at 25”.
Preliminary Fractionation of Peptic Digest with Trichloroacetic Acid-The BSA digest was treated with 400/, trichloroacetic acid as previously described (10) to furnish a 2aJ, trichloroacetic acid-precipitable fraction; the concentration of the supernatant solution was increased to obtain a IO’%-precipitable fraction. The precipitates were individually suspended in a small volume of distilled water and brought into solution by adding N NaOH to about pH 6.5. A portion of this solution was diluted with 0.155 M sodium phosphate buffer, pH 7.4, and the absorbance read at 280 nm. Comparison with BSA, E:%, = 6.54 at 280 nm, afforded a crude estimate of the peptide content of the 2% and 10% trichloroacetic acid fractions (see Fig. 5).
Chrorndographic Studies-From the preliminary studies de- scribed above, it appeared that digestion of BSA with pepsin in a ratio of 3OOO:l (w/w); pH 3.0, for 2 hours at 25’ would furnish suitable peptide material for chromatographic study. Ten grams of BSA were accordingly digested. Preliminary fraction- ation of the peptic digest was made with trichloroacetic acid, as described above. The yield of the 2% and 10% fractions,
FIG. 2. Polyacrylamide gel elect,rophoresis, pH 8.3, of a peptic digest of 5yo BSA (the same as that, shown in Fig. l), pepsin-BSA (w/w), 1:3000; 100 pg of peptide material were applied to each tube. Portions of the peptic digest were removed at the stated intervals.
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8080
FIG. 3. SDS gel electrophoresis, pH 7.2, of a peptic digest of 5% BSA, the same as that shown in Fig. 2. The origin (0), direc- tion of migration (- pole to + pole), and location of the eleetrophoretic bands are shown in Figs. 2 and 3; the estimated molecular weight is stated in Fig. 3. About 12 pg and 24 pg of peptide material were applied to tubes, 0 to 60 min, and to tubes, 120 and 180 min, respectively.
pepsin : BSA (W/W) 1:3000 - 1:300 -_ __-____
S = TCA supernatant
i 604 -ios P%TCA PPT I
0 60 120 180 24.0 300 360 DIGESTION TIME (MINUTES)
FIG. 4. Peptic digestion of 5% BSA as a function of pepsin concentration and digestion time: yield of precipitable peptide material (PPT) in the 2% and 10% trichloroacetic acid fractions and of nonprecipitable material in the supernatant fraction (S). Yield of precipitable peptide material is based on absorbance at ‘280 nm relative to that of intact BSA; the yield of S is based on the difference between the absorbance of intact BSA and the sum of the absorbancies of the 2% and lOTi, trichloroacetic acid frac- tions.
based on the weight of salt-free, lyophilized peptide material, was 4601, and 30%, respectively.
Two Per Cent Trichloroacetic Acid Fraction
Chromatography and Isolation of BSA Peptide Fragment KL- The 2% fraction was desalted in batches, 60 ml each of a solu- tion, pH 6.5, of peptides (2.48 g based on absorbance at 280 nm with BSA as a standard protein) ; a column (5.08 X 45.5 cm)
D = total peptic digest TCA = trichloroacetic acid ppt
M 60 .s -a .r
m 40 0) .g
z 20 nz
0 Testosterone Estradiol Progesterone
FIN. 5. Relative steroid binding activity of 2% trichloroacetic acid and 10% fractions of peptic digests of 5% BSA. Digestion proceeded for 2 hours at 25”, with pepsin-BSA (w/w), 1:3000 (clear urea) or 1:300 (hatched urea), pH 3.0. For further details, see fractionation with trichloroacetic acid under “Experimental Procedures.”
of Sephadex G-10, previously equilibrated in distilled water, was used. The salt-free peptide material was chromatographed at 4” on Sephadex G-75 in a volatile buffer, 0.6 N acetic acid- 0.01 N pyridine, pH 3.0, in a manner similar to that previously described (9). Chromatography was accomplished in batches (50 ml each of a solution adjusted with acetic acid to pH 3.0 and containing 1.4 g of lyophilised peptides) on two columns (2.5 X 90 cm, each) in tandem. The total hold-up volume (Vn) was 244 ml; the flow rate was 21 ml per hour. The col- umn effluent was monitored at 280 nm; the elution curve in- creased gradually except for a single (terminal) peak representing Fraction KL (Table I). The effluent fractions (5 ml, each) were examined by cellulose acetate electrophoresis, pH 5.0; like frac- tions were pooled and lyophilized, see Table I. Fraction KL
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

TABLE I
Sephadex G-76 chromatographic fractions of 2% trichtoroacetic acid-precipitable peptide material: yield and molecular species A batch of 1.4 g of peptide material was chromatographed; for
further details, see “Experimental Procedures.”
Fraction
ml m&T x 10-s
A 286390 110 36.5; 28.5;16.0
BC 391-560 321 34.5; 26.0; 15.0
DEF 561-725 465 24.5;13.5b
GHI 726-780 215 15.4
KL 781-826 161 9.6
a Estimated on SDS gel electrophoresis. b A minor component.
TABLE II
Sephadex G-75 chromatographic fractions of iO% trichloroacetic acid-precipitable material: yield and molecular species
Fraction
I II III IV
v VI
Effluent volume Weight
ml mx x 10-s
201-260 163 36.0; 16.0
261-330 134 34.0; 15.4 331450 183 16.0
451-590 237 15.0; 11.5
591-720 332 10.3b 721-820 71 <5.oc
Molecular weighta
Major components Minor
(or trace) components
79
76 77 81; 28
a Estimated on SDS gel electrophoresis. b Fraction V was purified by rechromatography on Sephadex
G-75 prior to estimation of the molecular weight.
c Estimation of the molecular weight of peptides below this limit is not precise.
was purified by preparative electrophoresis (9) and submitted for amino acid analysis.
Ten Per Cent Trichloroaeetic Acid Fraction
Chromatography and Isolation of BSA Peptide Fragment VI-
A portion of a solution, pH 6.5, of the 10% fraction was diluted in a large volume of 0.6 N acetic acid-O.01 N pyridine, pH 3.0, and concentrated by passage through a UM-2 membrane in a Diaflo ultrafilter (Amicon) as previously described (10) ; the procedure was repeated to remove trichloroacetic acid. A concentrated solution (25 ml) was thus obtained containing 756 mg of peptide material according to the absorbance at 280 nm with BSA as a standard protein. It was passed on a column (5.08 X 42 cm) of Sephadex G-75 in the above buffer, pH 3.0, at 4”. The hold-up volume (V,) was 225 ml; the flow rate was 83 ml per hour. The column effluent was monitored at 280 nm; only Fractions I and VI (Table II) represented peaks in the elution curve. Appropriate effluent fractions were pooled and lyophilized. A total of 1120 mg of peptide material was thus recovered, see Table II. The absorbance at 280 nm of the chromatographic fractions was appreciably lower than that of BSA, which may account for the apparent discrepancy in the estimated amount of peptide material applied to the column and the amount actually recovered from the effluent. Fraction VI was dissolved in a minimal volume of distilled water and passed
8081
through a short column of Sephadex G-10 previously equilibrated with distilled water; the effluent fraction containing peptide material was lyophilized and submitted for amino acid analysis.
Electrophoresis
Polyacrylamide gel electrophoresis was carried out at pH 8.3 in a Canalco apparatus as described by Davis (11). SDS gel electrophoresis was performed with the same apparatus as de- scribed by Weber and Osborn (12). Cellulose acetate (Sepra- phore III) electrophoresis was performed with a Gelman ap- paratus using the buffer, 0.04 N pyridine adjusted with acetic acid to pH 5.0, instead of 0.05 M sodium acetate, pH 5.0, as de- scribed by Peters and Hawn (9) ; the peptides were fixed in 5% trichloroacetic acid and stained with Ponceau S (Allied Chemical) in our study. Preparative electrophoresis (9) was carried out at 4” on a bed of Sephadex G-25 (fine) in a volatile buffer, 0.05 N acetic acid-O.048 N pyridine, pH 5.0.
Amino Acid Analysis
The peptides were hydrolyzed with 6 N HCl for 20 hours at 110”; the hydrolysates were assayed on an automatic amino acid analyzer, Beckman model 116, as previously described (13).
Measurement of Steroid Binding
The method for measuring steroid binding by batchwise use of Sephadex G-25 has been previously described (4). The technique is comparable in principle to that of equilibrium di- alysis. Protein, P, and steroid bound to protein, &, are excluded by the gel (200 mg of Sephadex G-25) in a final volume (a) of 2 ml of 0.155 M phosphate buffer, pH 7.4, at 25”; unbound steroid, X,, partitions freely between the external phase (ue = 1.5 ml) and the internal gel phase (vi = 0.5 ml). The steroid is to some extent reversibly adsorbed to the gel matrix. Adsorption is taken into account when routinely determining the partition factor, K’, i.e. the ratio of the total amount of steroid in the phase external to the gel to that in the total gel (internal volume plus gel matrix) in the absence of protein. Mean values & S.D. (n assays) for K’ were 0.979 i 0.071 (n = la), 1.10 & 0.045 (n = 9), and 0.448 =t 0.013 (n = 9) for progesterone, testoster- one, and 17&estradiol, respectively. The theoretical value for K’ would be 3 if adsorption were nil since v&i = 3.
Sephadex G-10 was used instead of Sephadex G-25 to measure steroid binding to BSA peptide fragment VI, because peptides of molecular weight less than 5000 may not be completely ex- cluded by Sephadex G-25. The assay system was modified so that v, v,, and vi were 1.5, 1.36, and 0.14 ml, respectively. Steroid adsorption to the gel matrix was considerable and so this system may be better described as one of competitive adsorption (14). Mean K’ values for progesterone, testosterone, and 170.estradiol were 0.360,0.472, and 0.055, respectively. The theoretical value for K’ would be 9.7 if adsorption were nil since V&Q = 9.7. Values obtained for the binding constants for steroid-BSA in- teraction were similar in assays with Sephadex G-10 and G-25.
Calculation of Steroid Binding Activity and Binding Constant
Steroid binding activity (previously designated (4) as the binding affinity) is empirically defined as the value for (&/xu) (l/P) where & and S, denote the molar concentrations of bound and unbound steroid, respectively, and P is the concentration (gram per liter) of total protein in the external volume (vJ of the assay system. Tritium-labeled progesterone, testosterone, or 17/%estradiol in fixed amounts of about 1.4, 0.8, or 1.2 ng, respectively, were added to the assay tube. The concentration of protein in the assay system was not varied in the present
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from
8082
study. The concentration of BSA was such as to afford an &/SU value of about 1.0. The steroid binding activity of in- tact BSA and BSA digest (or purified fractions thereof) was de- termined simultaneously. The ratio of the binding activity of the BSA digest (or peptide fragments) to that of intact BSA times 100 is defined as the relative binding affinity.
The value for the binding constant, nk, for the association of a steroid with a pure BSA peptide fragment (or with BSA) of known molecular weight may be obtained (4) from the Scatchard equation, (&/su) (l/P) = k (n - &,/P), where k is the intrinsic association constant for each binding site and n is the number of binding sites in the peptide molecule; the symbols, X,, &, and P, have been defined above and are expressed in molar concen- tration in each instance. The term 86/P, is negligible since & < P in the present study. An approximate value for nk was thus obtained for intact BSA and BSA Peptide Fragments KL and VI from &/sU at a single level (P) of the protein or peptide in the Sephadex assay system.
RESULTS AND DISCUSSION
According to Fig. 1, as much as 40 to 50% of the initial steroid binding activity remains after 2 or 3 hours of peptic digestion of BSA (ratio, pepsin-BSA (w/w), 1:3000) ; and, when the pepsin concentration is increased lo-fold, 10 to 20% of the binding activity remains. Progesterone, testosterone, and 17@-estradiol were selected for study to represent the major categories of steroid sex hormones; cortisol, a representative of the adreno- cortical hormones, was not included because it binds weakly to serum albumin. The binding activity of BSA peptide fragments of molecular weight considerably less than 5000 would go un- detected in our assay which depends on the molecular exclusion properties of the dextran gel, Sephadex G-25. The binding activity shown in Fig. 1 may therefore be attributed to BSA peptide fragments of molecular weight greater than 5000. The steroid binding activity of smaller peptide fragments, e.g. chro- matographic Fraction VI (see below) was determined with the aid of the more highly cross-linked dextran, Sephadex G-10 (upper molecular exclusion limit, 700).
That the binding activity of the peptic digest (Fig. 1) cannot be due to BSA which had escaped fragmentation may be con- cluded on viewing Fig. 2 and especially, Fig. 3. According to the latter figure (SDS gel electrophoresis), there is no BSA re- maining after 30 min of digestion, pepsin-BSA (w/w), 1:3000; and, after 2 or 3 hours of digestion, the major class of BSA frag- ments consists of peptides of molecular weight, 14,000, and minor classes, similarly of molecular weight, 27,000 and 34,000. Al- though there appear to be only a few classes of peptide compo- nents in the 2-hour digest, more components may be seen on polyacrylamide gel electrophoresis (Fig. 2).
Preliminary studies were undertaken to determine the experi- mental conditions for peptic digestion of BSA affording a maxi- mal yield of peptide fragments, especially small fragments with appreciable steroid binding activity. Following the example of Bradshaw and Peters (lo), trichloroacetic acid was used to fractionate the BS4 digest. Fig. 4 describes the yield of the 2% trichloroacetic acid and 10% trichloroacetic acid fractions as a function of pepsin concentration and digestion time. The yield of total precipitable material after 2 hours of digestion with pepsin-BSA (w/w), 1:3000, represented the bulk of the starting material, correlating with a 40 to 50% yield of steroid binding activity in the unfractionated digest (Fig. 1). The yield, simi- larly but with pepsin-BSA (w/w), 1:300, was considerably lower with respect to total precipitable peptides (Fig. 4) and steroid
2% TCA PPT 10% TCA PPT
Testosterone -
A BC DEF GHI KL I II Ill IV v VI
EFFLUENT FRACTION
FIG. 6. Relative steroid binding activity of Sephadex G-75 chromatographic fractions of 2% and 10% -fractions of a peptic digest of 5Y0 BSA: the relative bindincr activitv of BSA is taken
I ” ,
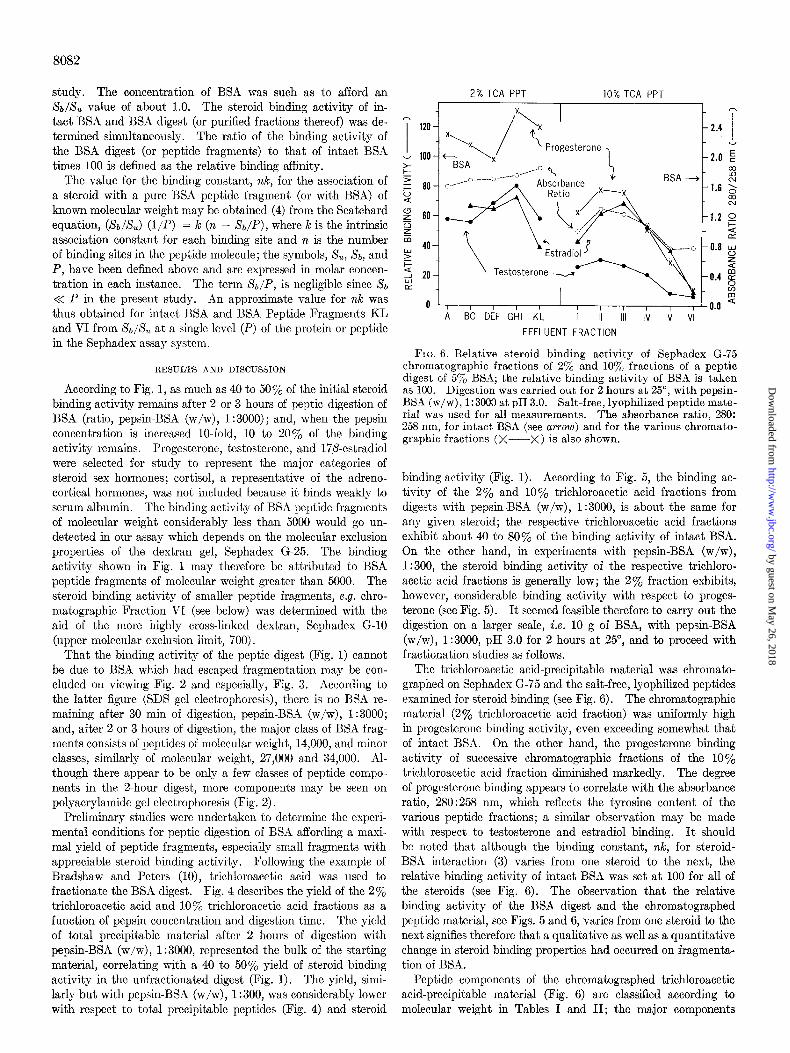
as 100. Digestion was carried out for 2 hours at 25”, with pepsin- BSA (w/w), 1:3000 at pH 3.0. Salt-free, lyophilized peptide mate- rial was used for all measurements. The absorbance ratio, 280: 258 nm, for intact BSA (see arrow) and for the various chromato- graphic fractions (X---X) is also shown.
binding activity (Fig. 1). According to Fig. 5, the binding ac- tivity of the 2% and 10% trichloroacetic acid fractions from digests with pepsin-BSA (w/w), 1:3000, is about the same for any given steroid; the respective trichloroacetic acid fractions exhibit about 40 to 80% of the binding activity of intact BSA. On the other hand, in experiments with pepsin-BSA (w/w), 1: 300, the steroid binding activity of the respective trichloro- acetic acid fractions is generally low; the 2yo fraction exhibits, however, considerable binding activity with respect to proges- terone (see Fig. 5). It seemed feasible therefore to carry out the digestion on a larger scale, i.e. 10 g of BSA, with pepsin-BSA (w/w), 1:3000, pH 3.0 for 2 hours at 25”, and to proceed with fractionation studies as follows.
The trichloroacetic acid-precipitable material was chromato- graphed on Sephadex G-75 and the salt-free, lyophilized peptides examined for steroid binding (see Fig. 6). The chromatographic material (2% trichloroacetic acid fraction) was uniformly high in progesterone binding activity, even exceeding somewhat that of intact BSA. On the other hand, the progesterone binding activity of successive chromatographic fractions of the 10% trichloroacetic acid fraction diminished markedly. The degree of progesterone binding appears to correlate with the absorbance ratio, 280:258 nm, which reflects the tyrosine content of the various peptide fractions; a similar observation may be made with respect to testosterone and estradiol binding. It should be noted that although the binding constant, nk, for steroid- BSA interaction (3) varies from one steroid to the next, the relative binding activity of intact BSA was set at 100 for all of the steroids (see Fig. 6). The observation that the relative binding activity of the BSA digest and the chromatographed peptide material, see Figs. 5 and 6, varies from one steroid to the next signifies therefore that a qualitative as well as a quantitative change in steroid binding properties had occurred on fragmenta- tion of BSA.
Peptide components of the chromatographed trichloroacetic acid-precipitable material (Fig. 6) are classified according to molecular weight in Tables I and II; the major components
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

TABLE III TABLE IV Amino acid composition of isolated jragments Pepticle fragments of bovine serum albumin: number of residues of
each kind in 100 residues analyzed
Amho acid
Lysine ..................... Histidine .................. Arginine .................. Aspartic acid. ............. Threonine .................. Serine ..................... Glutamic acid .............. Proline. ................... Glycine .................... Alanine .................... Half -cystine ............... Valine ..................... Methionine ................ Isoleucine .................. Leucine. ................... Tyrosine ................... Phenylalanine. ............. Tryptophan. ...............
Total residues .............. Molecular weight@. .........
Peptide fragment
Ia I I
VI
I I Aspa,
Integer integer
Integer
2.77 1.14 1.01 3.17 0.75 1.35 4.49 1.10 0.80 2.86 1.54” 2.00 0.0 0.12 3.46 1.76 1.45 0.00”
2.77 3 3 2.08 2 3 0.79 1 1 2.21 2 2 1.09 1 1 0.96 1 1 3.74 4 3 0.37 0 0 1.61 2 2 1.21 1 1 0.0 0 0 0.99 1 1 0.0 0 0 0.96 1 1 2.84 3 3 0.46 0 0 1.88 2 2 0.00~ 0 0
24 24 2766 2808
- 0 Peptide fragment Asp was isolated by Peters and Hawn (9). b After performic acid oxidation. c Tryptophan estimated from molecular extinction coefficient
at 280 nm and tyrosine content. d A minimal estimate.
listed correspond fairly well to the three major classes of com- ponents seen in Fig. 3. The resolution obtained on Sephadex G-75 chromatography was crude; interaction of the peptide fragments had probably occurred. Nevertheless, Fractions GHI and KL (2% trichloroacetic acid fraction, Table I) appear to contain one class of peptides with respect to molecular weight, i.e. 15,400 and 9,600, respectively. It is interesting that these chromatographic fractions exhibit a relatively high degree of progesterone binding and an appreciable degree of testosterone and estradiol binding (see Fig. 6). Fraction GHI (and the preceding fractions) exhibited several components on polyacryl- amide gel electrophoresis (not shown) but Fraction KL was homogeneous. Fraction KL was further purified by electropho- resis on Sephadex G-25 and submitted for amino acid analysis, see Table III. The minimal molecular weight was estimated to be 3350, i.e. about one-third of that estimated on SDS gel elec- trophoresis. Peptide Fragment KL contains no methionine, isoleucine, or tryptophan but it is rich in tyrosine. In other respects, Peptide KL resembles BSA in the frequency of indi- vidual amino acids per 100 residues analyzed (see Table IV). Peptide KL and intact BSA have about the same mobility on cellulose acetate electrophoresis, pH 5.0; on the other hand, KL moves very fast, i.e. with the tracking dye, on polyacrylamide gel electrophoresis, pH 8.3 (see Fig. 7).
Only one peptide fragment appeared in chromatographic Fraction VI (lOTo fraction, Table II). Peptide VI is similar in molecular weight and in electrophoretic behavior (Fig. 7) to the NH&err&al or Asp fragment of BSA previously isolated by Peters and Hawn (9). Both peptides were isolated from the
8083
Amino acid Peptide fragment
BSA“
KL VI --
Lysine ....................... Histidine .................... Arginine .................... Aspartic acid. ............... Threonine .................... Serine ....................... Glutamic acid ................ Proline. ..................... Glycine ...................... Alanine ...................... Half-cystine ................. Valine ....................... Methionine .................. Isoleucine. ................... Leucine. ..................... Tyrosine ..................... Phenylalanine. ............... Tryptophan. .................
10.4 11.5 10.1 3.45 7.69 3.00 3.45 3.85 4.00
10.4 7.69 9.50 3.45 3.85 5.67 3.45 3.85 4.67
13.8 15.4 13.2 3.45 0.0 4.83 3.45 7.69 2.83
10.4 3.85 8.17 6.90 0.0 6.33 6.90 3.85 5.83 0.0 0.0 0.67 0.0 3.85 2.33
10.4 11.5 10.8 6.90 0.0 3.33 : 3.45 7.69 4.67 0.0 0.0 0.33
Total residues ................ 29 24 602
-
-
-
a Calculated from data cited in a recent review (7).
FIG. 7. Electrophoresis of BSA and Peptide Fragments KL and VI. The origin (0) and direction of migration (- pole to + pole) are indicated. The tracking dye, bromphenol blue, appears as a narrow zone at the bottom of the polyacrylamide gel patt,ern of Fragment VI, but it is not seen in the BSA pattern because intact BSA adsorbs the dye. A faint zone representing Peptide Frag- ment VI may be seen at the bottom of the cellulose acetate pat- tern. BSA, KL, and VI were applied in amounts of 75, 50, and 100 rg, respectively, in gel electrophoresis; and, in amounts of 200 gg each in cellulose acetate electrophoresis.
10% trichloroacetic acid fraction of the BSA digest; the digestion time in the previous study (9) was, however, 33 min, instead of 2 hours. The amino acid composition of Peptide VI and the Asp peptide (see Table III), were almost identical; there was a slight difference in the integer values estimated for the histidine and glutamic acid residues. Estimates for the minimal molecu- lar weight of Peptides VI and Asp were 2766 and 2808, respec- tively. The molecular weights for the purified BSA Peptide Fragments KL and VI having been established, the binding
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8084
TABLE V
Binding constants (nk) for the association of various steroid hormones with pure BSA peptide fragments
For method of calculation, see “Experimental Procedures.”
Peptide or protein Molecular weight
~;E Testos-
I I terone Es:%01
liter/mole x 10’
KL 10,050” 0.44 0.18 0.33 VI 2,766 0.014
I I
0.008 0.010 BSA* 69,000 2.8 2.4 6.1
a Calculated as 3 times the minimal molecular weight, 3350 (see Table III).
* The nk values for BSA may be compared to those reviewed by Westphal (3) which vary considerably depending on the albumin preparation used, e.g. prior removal of bound fatty acid results in an increased nk for steroid binding.
constants, nk, were calculated for comparison with those for in- tact BSA (see Table V). It is noneworthy that Peptide VI con- tains no tyrosine and exhibits little steroid binding activity (see Fig. S), whereas Peptide KL is rich in tyrosine and exhibits con- siderable binding activity; however, proline and cyst&e which are also absent in Peptide VI are present in Peptide KL. The frequency of ammo acid residues in Peptide VI thus differs in im- portant respects from that of Peptide KL and intact BSA (see Table IV).
The tyrosine residues in Peptide KL may be important in the mechanism for steroid binding. A similar suggestion but with respect to intact BSA was made by Levedahl and Perlmutter (15) from studies on the pH dependency of testosterone-BSA interaction. Swaney and Klotz (16) have recently suggested that the amino acids clustered around the lone tryptophan of
human serum albumin may be important for steroid binding. However, BSA (which has two tryptophan groups) affords the Peptide Fragment KL which contains no tryptophan and yet it binds progesterone.
Acknowledgment-We are indebted to Dr. David A. Ontjes for the determination of the amino acid composition of Peptide Fragments KL and VI.
REFERENCES
1. PEARLMAN, W. H., AND FONG, I. F. F. (1972) Fed. Proc. 31,430 2. ANTONIADES, H. N., ed (1960) Hormones in Human Plasma,
p. 476, Little, Brown and Company, Boston 3. WESTPHAL, U. (1971) Steroid-Protein Interactions, pp. 10%
113, Springer-Verlag, New York 4. PEARLMAN, W. H., AND CR~PY, 0. (1967) J. Biol. Chem. 242,
182-189 5. SEAL, U. S., AND DOE, R. P. (1966) in Steroid Dynamics (PIN-
cus, G., NAKAO, T., AND TAIT, J. F., eds) p. 63, Academic Press, New York
6. STEINHARDT, J., AND REYNOLDS, J. A. (1969) Multiple Equi- libria in Proteins, pp. 85-124, Academic Press, New York
7. PUTNAM, F. W. (1965) in The Proteins (NEURATH, H., ed) Vol. III, pp. 187-199, Academic Press, New York
8. WEBER, G., AND YOUNG, L. B. (1964) J. Biol. Chem. 239,1424- 1431
9. PETERS, T., JR., AND HAWN, C. (1967) J. Biol. Chem. 242,1566- 1573
10. BRADSHAW, R. A., AND PETERS, T., JR. (1969) J. Biol. Chem. 244, 5582-5589
11. DAVIS, B. J. (1964) Ann. N. Y. Acad. SC!. 121,404 12. WEBER, K., AND OSBORN, M. (1969) J. Biol. Chem. 244, 4406-
4412 13. SPACKMAN, D. H., STEIN, W. H., AND MOORE, S. (1958) Anal.
Chem. 36, 1190 14. HEYNS, W., VAN BAELEN, H., AND DE MOOR, P. (1968) Ann.
Endocrinol. 29, 123 15. LEVEDAHL, B. H:, AND PERLMUTTER, R. (1956) Arch. Biochem.
Biophys. 61,442 16. SWANEY, J. B., AND KLOTZ, I. M. (1970) Biochemistry 9, 2570
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from

William H. Pearlman and I. F. F. FongObtained on Peptic Digestion
Steroid Binding Properties of Some Peptide Fragments of Bovine Serum Albumin
1972, 247:8078-8084.J. Biol. Chem.
http://www.jbc.org/content/247/24/8078Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/247/24/8078.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on May 26, 2018
http://ww
w.jbc.org/
Dow
nloaded from





























