Embed Size (px)
Citation preview

Eur. J. Biochem. 267, 1837±1846 (2000) q FEBS 2000
Structural analysis of two glycosphingolipids from thelipopolysaccharide-lacking bacterium Sphingomonas capsulata
Kazuyoshi Kawahara1, Hermann Moll2, Yuriy A. Knirel2,3, Ulrich Seydel2 and Ulrich ZaÈ hringer2
1Department of Bacteriology, The Kitasato Institute, Tokyo, Japan; 2Forschungszentrum Borstel, Zentrum fuÈr Medizin und Biowissenschaften,
Borstel, Germany; 3N.D. Zelinsky Institute of Organic Chemistry, Russian Academy of Sciences, Moscow, Russia
Two glycosphingolipids, GSL-1 and GSL-3, were isolated from Sphingomonas capsulata and studied by
methylation analysis, laser desorption mass spectrometry, and 1H and 13C NMR spectroscopy, including
two-dimensional 1H,1H COSY and heteronuclear 13C,1H COSY experiments. GSL-1 and GSL-3 differ in
their carbohydrate part, their structures being a-d-GlcpA-(1!1)-Cer and a-d-Galp-(1!6)-a-d-GlcpN-
(1!4)-a-d-GlcpA(1!1)Cer, respectively. Variations occur in the ceramide of GSL-1 and GSL-3, both
having the same long-chain bases, erythro-2-amino-1,3-octadecanediol (sphinganine), (13Z)-erythro-2-amino-
13-eicosene-1,3-diol and (13Z)-erythro-2-amino-13,14-methylene-1,3-eicosanediol, in the ratios 2.6 : 1 : 3.5 in
GSL-1 and 1 : 1.2 : 1.5 in GSL-3. All bases are quantitatively substituted by amide-linked (S)-2-hydroxymyristic
acid.
Keywords: chemical structure; glycosphingolipids; Sphingomonas capsulata; lipopolysaccharide-free
Gram-negative bacteria.
Glycosphingolipids (GSL) are amphiphilic molecules present inthe plasma membrane of eukaryotic cells, where they play arole in cell±cell recognition and interaction [1]. Manystructural variations were reported in GSL of mammalian andother vertebrate tissues, shellfishes and insects [2], whereas inbacteria GSL are uncommon.
The bacterial cell envelope is known to consist of acytoplasmic membrane, a peptidoglycan layer, and, in Gram-negative bacteria, an outer membrane that is extremelyasymmetric with respect to lipid distribution. The outermembrane is characterized by the presence of lipopoly-saccharide (LPS), which is located almost exclusively on theouter leaflet [3]. No exception from this architectural principlehad been reported for Gram-negative bacteria until wedemonstrated that Sphingomonas paucimobilis lacks LPS [4].This bacterium replaced LPS by two kinds of GSL. One ofthem had a-d-GlcA as the only sugar component and wascalled GSL-1. A similar GSL had been previously isolated fromthe bacterium Flavobacterium devorans [5], which was latertransferred to a new genus Sphingomonas [6]. The otherglycosphingolipid, GSL-4A, contained an a-d-Manp-(1!2)-a-d-Galp-(1!6)-a-d-GlcpN-(1!4)-a-d-GlcpA-(1!tetrasaccharidechain, thus sharing a minimal glucuronosyl ceramide structurewith GSL-1.
Sphingomonas capsulata GIFU 11526 [6] was depositedin the American Type Culture Collection as Flavobacteriumcapsulatum [7], which later transferred to the genus
Sphingomonas [6]. However, recent studies on its 16SrRNA sequence have shown that S. capsulata is phylo-genetically far from S. paucimobilis [8,9]. Therefore, althoughthese two Sphingomonas species are taxonomically related,they might have different GSL. Furthermore, it has beendemonstrated that both GSL from S. paucimobilis are localizedin the outer membrane and their carbohydrate moiety isexposed on the cell surface [10]. This and other findingsindicated that Sphingomonas GSL may share structural featuresand possess functional and physicochemical similarities withLPS [11]. The GSL of S. capsulata might be anotherrepresentative of the family of GSL from Gram-negativebacteria, which are thought to replace LPS in the outermembrane.
In this paper, we report on the elucidation of the completestructure of two GSL isolated from S. capsulata. These GSLshow both structural similarities with and differences fromtwo glycosphingolipids, GSL-1 and GSL-4A, found inS. paucimobilis [4,10].
M A T E R I A L S A N D M E T H O D S
Bacterial strain and growth
S. capsulata GIFU 11526 (originally F. capsulatum ATCC14666, Type strain) was cultivated in liquid medium containing0.5% yeast extract (Difco, Detroit, MI, USA), 0.5% casaminoacids (Difco), 0.2% (NH4)2SO4, 0.2% K2HPO4, and 0.1%MgSO4´7H2O, using a 30 L jar fermenter at 30 8C for 24 h. Thebacteria were killed by heating (100 8C, 30 min) and harvestedby centrifugation (10 000 g, 10 min).
Extraction and purification of GSL
Bacterial cells were washed with distilled water and lyo-philized. Dried cells were homogenized with chloroform/methanol (2 : 1, v/v) to remove phospholipids, and GSL were
Correspondence to U. ZaÈhringer, Forschungszentrum Borstel, Zentrum fuÈr
Medizin und Biowissenschaften, Parkallee 22, D-23845 Borstel, Germany.
Fax: 1 49 4537188612, Tel.: 1 49 4537188462,
E-mail: [email protected]
Abbreviations: CI, chemical ionization; EI, electron impact; GlcA,
glucuronic acid; GSL, glycosphingolipid; LD, laser desorption;
LPS, lipopolysaccharide; 14:0(2-OH), (S )-2-hydroxymyristic acid.
(Received 20 December 1999, revised 27 January 1999, accepted
28 January 1999)
1838 K. Kawahara et al. (Eur. J. Biochem. 267) q FEBS 2000
extracted twice from the residual cells with chloroform/methanol (1 : 3, v/v) at 80 8C for 1 h. The crude GSLpreparation was concentrated and applied to a column of silicagel 60 (70±230 mesh, Merck, Darmstadt, Germany) forpurification. Lipids were eluted using a stepwise gradient ofchloroform/methanol from 4 : 1 to 1 : 3 (v/v). The elution wasmonitored by TLC using aluminium plates of silica gel 60(Merck) and a solvent system of chloroform/methanol/aceticacid/water (25 : 15 : 4 : 2, v/v/v/v). Lipids were visualized byspraying with 10% sulfuric acid in ethanol and heating. GSLfractions were dialysed against dilute aqueous triethylaminepH 8.5 (0.005%, v/v) and lyophilized.
NMR spectroscopy
Spectra (360 MHz 1H-NMR and 90.5 MHz 13C-NMR) wererecorded on an AM-360 spectrometer (Bruker, Germany) at32 8C. Samples were dissolved in a mixture ofCDCl3 : CD3OD : D2O 2 : 2 : 0.2 (v/v). Chemical shiftsare referenced to internal chloroform (dH 7.26 p.p.m.,dC 77.0 p.p.m.). Standard Bruker software (disnmr, version8911 01.0) was used to run two-dimensional 1H,1H COSY andheteronuclear 13C,1H COSY experiments.
GLC, GLC-MS and laser-desorption MS
GLC was performed on a GC-14 A instrument (Shimadzu,Kyoto, Japan) equipped with a chemically bonded capillarycolumn (25 m � 0.2 mm) of CBP-1 or CBP-10 (Shimadzu).
GLC-MS was performed on a Hewlett-Packard HP 5985instrument (Palo Alto, CA) equipped with a chemically bondedfused silica capillary column (25 m � 0.32 mm) of SE-54(Weeke, MuÈhlheim, Germany). Electron impact (EI) massspectra were recorded at 70 eV. Ammonia was used as areactant gas in chemical ionization (CI) MS.
Laser-desorption (LD-MS) was performed on a LAMMA500 instrument (Leybold-Heraeus, KoÈln, Germany) underconditions described previously [12]. CsI and NaI were usedas cationization reagents.
Sphingolipid analyses
Fatty acids were analysed by GLC after hydrolysis with4 m HCl (100 8C, 5 h) and esterification with etherealdiazomethane.
The glycosyl moiety of GSL-3 was oxidized by stirring inaqueous triethylamine 0.005% (v/v) containing 0.025 mNaIO4 at 4 8C for 120 h in the dark. An excess of NaIO4
was destroyed with ethylene glycol, and the oxidized sugarswere reduced with NaBH4 at room temperature for 3 h.After dialysis and lyophilization, the product was methano-lysed (1 m HCl/methanol, 100 8C, 5 h) to liberate sphingoidand fatty acid derivatives, which were peracetylated [Ac2O/pyridine 1 : 1 (v/v), 100 8C, 30 min] and analysed by GLC andGLC-MS.
GSL-1 and GSL-3 (1 mg of each) were freed from sugarsand the amide-linked fatty acid (S )-2-hydroxymyristic acid[14:0(2-OH)] by methanolysis (0.2 m HCl/methanol, 65 8C,5 h). After neutralization and evaporation, the residue wassuspended in chloroform (2 mL) and oxidized with Pb(OAc)4
(1 mg) at room temperature for 2 h. The product was extractedtwice with chloroform (2 mL), the organic phases werecombined, dried over Na2SO4 and evaporated. The resultingaldehyde was dissolved in diethyl ether (2 mL) and reduced tothe corresponding alcohol with LiAlH4 or LiAlD4 (< 5 mg) for1.5±2 h with stirring. The solvent was evaporated in a stream ofnitrogen, water (1 mL) was added, and the product wasextracted three times with chloroform (3 mL). The combinedorganic layers were dried and the residue was conventionallytrimethylsilylated or esterified with freshly prepared nicotinicacid chloride in pyridine (3.5 mg in 500 ml) in a glass-tube at85 8C for 1 h. After evaporation of pyridine, the product wasextracted three times with a 1 : 1 (v/v) mixture of hexane/water(2 mL) and the organic layers were combined, dried, andanalysed by GLC using a temperature gradient 150 (3 min) to320 8C at 5 8C´min21.
Sugar and methylation analyses
GlcN was determined by the method of Strominger et al. [13]after hydrolysis with 4 m HCl (100 8C, 16 h). Neutral sugars
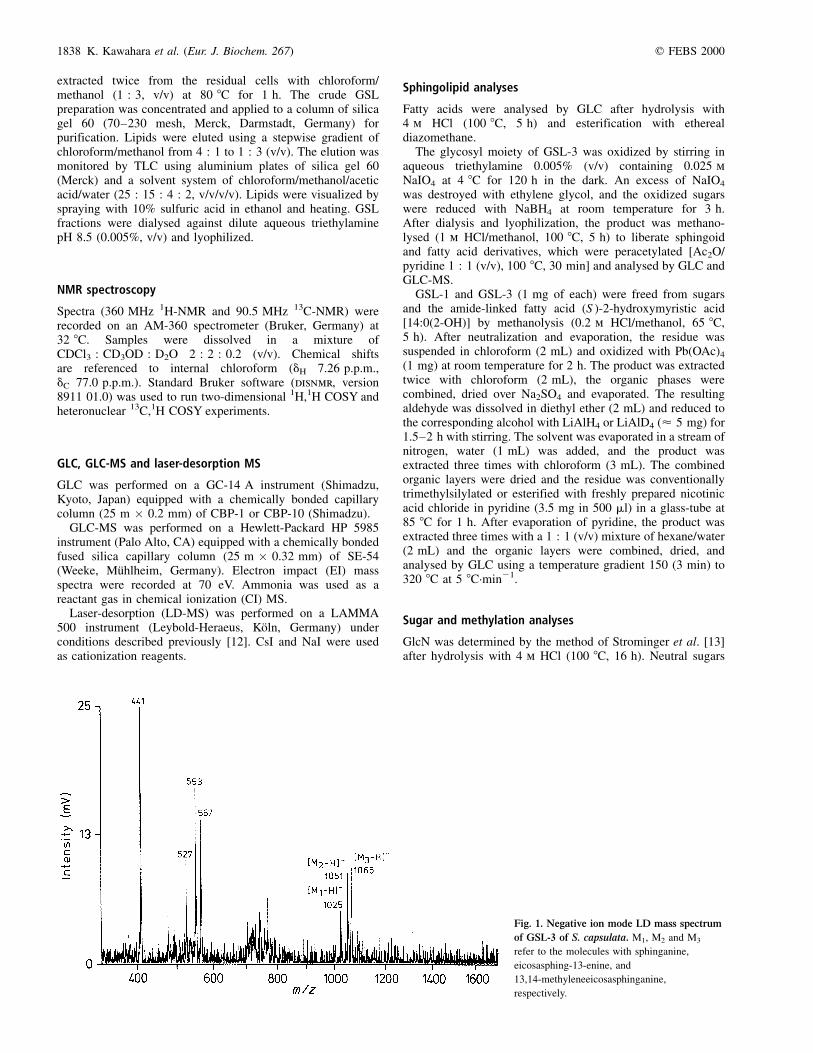
Fig. 1. Negative ion mode LD mass spectrum
of GSL-3 of S. capsulata. M1, M2 and M3
refer to the molecules with sphinganine,
eicosasphing-13-enine, and
13,14-methyleneeicosasphinganine,
respectively.
q FEBS 2000 Glycosphingolipids from Sphingomonas capsulata (Eur. J. Biochem. 267) 1839
were determined as alditol acetates by GLC after acidhydrolysis (0.1 m HCl, 100 8C, 48 h), reduction with NaBH4
and peracetylation. Determination of uronic acid was per-formed by the carbazole-sulfuric acid method [14] afterhydrolysis with 1 m H2SO4 (100 8C, 5 h). In addition, GlcAwas identified by GLC after carboxyl-reduction, hydrolysis(1 m CF3COOH, 120 8C, 2 h), carbonylreduction (NaBH4) andperacetylation.
GSL-3 was N-acetylated (1.6% Ac2O in methanol, v/v) andpermethylated by the method of Hakomori [15] with somemodifications [16,17]. The product was hydrolysed with 1 mCF3COOH at 120 8C for 2 h, and liberated sugars wereconventionally reduced with NaBH4, peracetylated (Ac2O/pyridine) and analysed by GLC-MS.
R E S U LT S
Isolation and characterization of GSL
The crude extract from S. capsulata contained two kinds ofGSL having Rf 0.48 and 0.18 in TLC and also significantamounts of contaminating phospholipids. Purification by silicagel column chromatography afforded two GSL, one with amonosaccharide and the other with a trisaccharide carbo-hydrate chain (GSL-1 and GSL-3, respectively). GSL-1 fromS. capsulata with Rf 0.48 was identical to GSL-1 from
S. paucimobilis [4,10], whereas GSL-3 with Rf 0.18migrated faster than GSL4A from S. paucimobilis having Rf
0.11 [4,10].Negative ion mode LD-MS showed a heterogeneity in the
sphingolipid moiety of GSL-1 and GSL-3. The latter wascharacterized by the molecular masses of 1026, 1052 and1066 Da (Fig. 1). Fragment ions with m/z 527, 553 and567, which were derived from the ceramide moiety,showed the same heterogeneity pattern. LD-MS analysisof GSL-1 revealed the molecular masses of 703, 729 and743 Da and the same fragment ions as in GSL-3.Therefore, the heterogeneity occurs in the sphingolipidmoiety and is similar in both GSL. These data showedalso that GSL-1 and GSL-3 have monosaccharide andtrisaccharide carbohydrate moieties, respectively.
Studies on the sphingolipid moiety
GLC analysis of the methyl ester revealed the presence of14:0(2OH) as the only fatty acid in both GSL-1 and GSL-3.GLC of the l-phenylethylamide derivative showed that 14:0(2-OH) has the (S)-configuration.
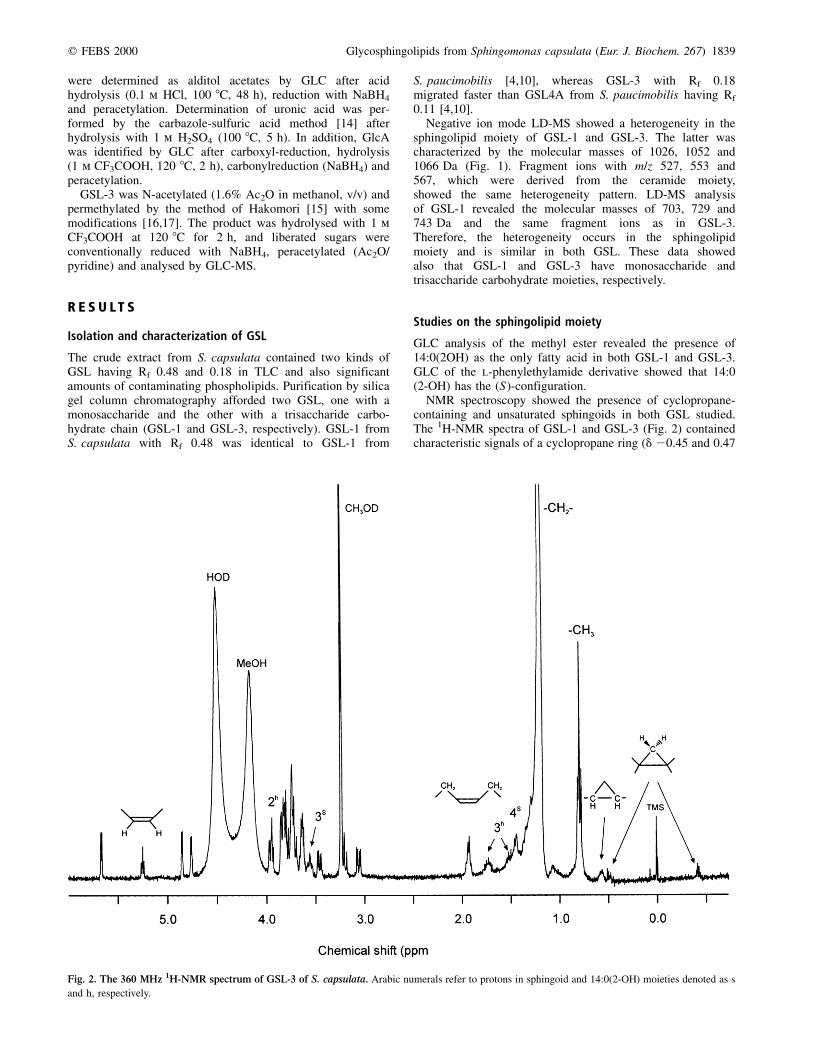
NMR spectroscopy showed the presence of cyclopropane-containing and unsaturated sphingoids in both GSL studied.The 1H-NMR spectra of GSL-1 and GSL-3 (Fig. 2) containedcharacteristic signals of a cyclopropane ring (d 20.45 and 0.47
Fig. 2. The 360 MHz 1H-NMR spectrum of GSL-3 of S. capsulata. Arabic numerals refer to protons in sphingoid and 14:0(2-OH) moieties denoted as s
and h, respectively.

1840 K. Kawahara et al. (Eur. J. Biochem. 267) q FEBS 2000
for the methylene protons and d 0.56 for the methine protons inGSL-3). Similar signals were present in the spectra of GSLfrom S. paucimobilis, containing (13Z )-erythro-2-amino-13,14-methylene-1,3-octadecanediol [4]. In addition, the spectra ofGSL-1 and GSL-3 showed two coupled multiplets for theolefinic and allylic protons of another, monounsaturatedsphingoid (d 5.23 and 1.93, respectively; data from the 1H,1HCOSY spectrum of GSL-3). The same minor signals werepresent also in the 1H-NMR spectrum of GSL-4A fromS. paucimobilis, although the corresponding unsaturated basehas not been identified in the course of chemical studies [4]owing to its low content. The 13C-NMR spectrum of GSL-3(Fig. 3) confirmed the presence of a cyclopropyl ring (signals atd 10.3 for CH2 and d 15.2 for two CH groups; data from theDEPT spectrum) and a double bond (signals at d 129.3 and129.4). The position of the signals for allylic carbons at d 26.6and 26.7 (data from the 13C,1H COSY spectrum) indicated thecis configuration of the olefinic group (the signals for the transisomer would appear at d 32±33) [18].
Methanolysis of GSL-3 afforded only small amounts ofsphingoids, most likely owing to the attachment of GlcA. WhenGSL-3 was oxidized by periodate prior to methanolysis, threelong-chain bases were released smoothly and analysed byGLC-MS in the EI and CI modes as peracetylated derivatives.They had respective relative retention times of 1, 1.14 and 1.22,
relative peak areas of 1 : 1.2 : 1.5 and molecular masses of427, 453 and 467 Da (CI MS data). The same compounds in theratios 2.6 : 1 : 3.5 were derived also from GSL-1. One of them,having the lowest molecular mass, was identified as peracetyl-ated erythro-2-amino-octadecane-1,3-diol (sphinganine). Thecompound with the highest molecular mass was indistinguish-able by the retention time and the EI mass spectrum from thederivative of (13Z )-erythro-2-amino-13,14-methylene-eicosane-1,3-diol, which has been identified earlier in GSL-4A fromS. paucimobilis [4]. The molecular mass of 453 Da for the thirdcompound pointed to an eicosasphingenine, which wasidentified as (13Z )-erythro-2-amino-13-eicosene-1,3-diol.
Long-chain alcohols were prepared from GSL-3 bymethanolysis and oxidation with Pb(OAc)4 followed byreduction with LiAlH4 and analysed by GLC-MS as trimethyl-silyl derivatives. As a result, the following compounds wereidentified: hexadecanol (from sphinganine) (11Z )-11,12-methyleneoctadecanol [from (13Z )-erythro-2-amino-13,14-methylene-1,3-eicosanediol], and (11Z )-11-octadecenol [from(13Z )-erythro-2-amino-13-eicosene-1,3-diol]. The last com-pound had the same retention time as the authentic sampleprepared by carboxyl reduction of (11Z)-11-octadecenoic acidand differed from the (11E )-isomer.
In order to confirm the position of the cyclopropyl ring andthe double bond, the long-chain alcohols were converted to
Fig. 3. The 90 MHz 13C-NMR spectrum of GSL-3 of S. capsulata. Arabic numerals refer to carbons in sphingoid and 14:0(2-OH) residues denoted as s and
h, respectively.

q FEBS 2000 Glycosphingolipids from Sphingomonas capsulata (Eur. J. Biochem. 267) 1841
nicotinates. GLC-MS analysis of the 11,12-methyleneocta-decanol derivative with the molecular mass of 387 Da showed aseries of fragments differing by 14 units, which was interruptedafter m/z 262 (the C1±C10 fragment) and continued from m/z302 (the C1±C12 fragment) (Fig. 4). Thus, the cyclopropyl ringis located between C11 and C12. A similar pattern for the11-octadecenol derivative with the molecular mass of373 Da, characterized by the interruption of the homologseries between m/z 262 and 288, confirmed the location of thedouble bond between the same carbons.
Studies on the carbohydrate moiety
GSL-1 was found to contain GlcA and no other sugarcomponent. Sugar analysis of GSL-3 showed that it containedd-Gal, d-GlcN, and d-GlcA. Methylation analysis of GSL-3revealed 1,5-di-O-acetyl-2,3,4,6-tetra-O-methyl-d-galactitoland 1,5,6-tri-O-acetyl-2-deoxy-3,4-di-O-methyl-2-(N-methyl-acetamido)-d-glucitol, indicating that Gal was at the terminalend and GlcN was substituted at position 6. When GSL-3was hydrolysed, N-acetylated, borohydride reduced andpermethylated, a disaccharide derivative was identified byGLC-MS, in addition to methylated monosaccharide alditols. Ithad a molecular mass of 526 Da determined by CI-MS andgave characteristic fragment ions in EI-MS at m/z 250 and260!228, corresponding to the reducing (GlcA-ol) andnonreducing (GlcNAc) sugar moieties, respectively. This
derivative had the identical retention time and the identicalMS fragmentation pattern to those of the carbonyl-reduced andpermethylated GlcNAc-(1!4)-GlcA-ol disaccharide derivedfrom GSL-4A of S. paucimobilis [4].
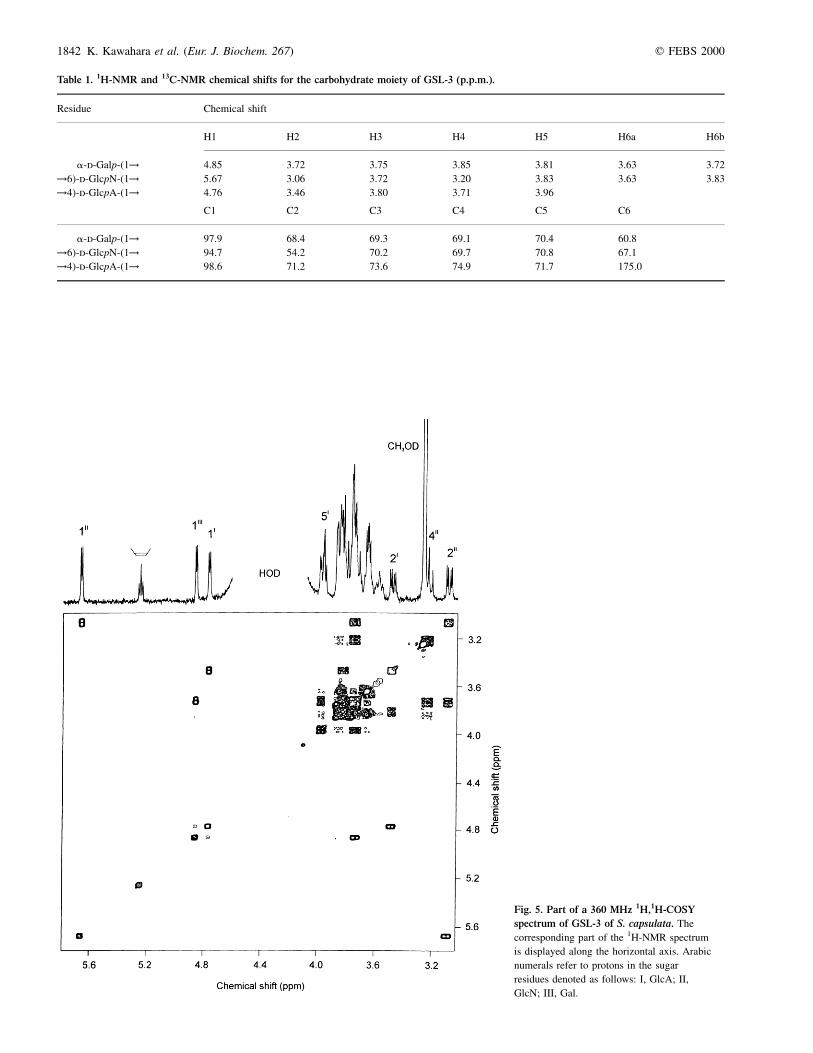
The 1H-NMR and 13C-NMR spectra of GSL-3 were assignedusing two-dimensional 1H,1H COSY (Fig. 5) and heteronuclear13C,1H COSY (Fig. 6) experiments (Table 1). When theunambiguous assignment was complicated by close positionof proton signals in the 1H-NMR spectrum (e.g. as in Gal), the13C-NMR chemical shift data of the corresponding mono-saccharides [19] and the glycosylation effects [20], aswell as data of a related model compound, GSL-4A fromS. paucimobilis with known structure [4], were used.Relatively small J1,2 coupling constant values of 3.2±3.8 Hz for the signals of the anomeric protons at d 4.76,4.85 and 5.67 indicated that all three sugar constituents inGSL-3 are a-linked.
The 13C-NMR chemical shift data of GSL-3 demonstratedsubstitution of GlcA at position 4 and GlcN at position 6, thelatter finding being consistent with the methylation analysisdata. Indeed, the signals for C4 of GlcA and C6 of GlcN wereshifted downfield to d 74.9 and 67.1, respectively, as comparedto their position in the spectra of the corresponding nonsub-stituted monosaccharides [19]. The chemical shifts for C2±C6of Gal were similar to those in nonsubstituted a-galacto-pyranose, thus confirming the terminal position of thismonosaccharide in GSL-3.
Fig. 4. EI mass spectrum of
(11Z)-11-octadecenylnicotinate derived
from GSL-3 of S. capsulata.
1842 K. Kawahara et al. (Eur. J. Biochem. 267) q FEBS 2000
Table 1. 1H-NMR and 13C-NMR chemical shifts for the carbohydrate moiety of GSL-3 (p.p.m.).
Residue Chemical shift
H1 H2 H3 H4 H5 H6a H6b
a-d-Galp-(1! 4.85 3.72 3.75 3.85 3.81 3.63 3.72
!6)-d-GlcpN-(1! 5.67 3.06 3.72 3.20 3.83 3.63 3.83
!4)-d-GlcpA-(1! 4.76 3.46 3.80 3.71 3.96
C1 C2 C3 C4 C5 C6
a-d-Galp-(1! 97.9 68.4 69.3 69.1 70.4 60.8
!6)-d-GlcpN-(1! 94.7 54.2 70.2 69.7 70.8 67.1
!4)-d-GlcpA-(1! 98.6 71.2 73.6 74.9 71.7 175.0
Fig. 5. Part of a 360 MHz 1H,1H-COSY
spectrum of GSL-3 of S. capsulata. The
corresponding part of the 1H-NMR spectrum
is displayed along the horizontal axis. Arabic
numerals refer to protons in the sugar
residues denoted as follows: I, GlcA; II,
GlcN; III, Gal.

q FEBS 2000 Glycosphingolipids from Sphingomonas capsulata (Eur. J. Biochem. 267) 1843
Combining the MS and NMR data obtained, it could beconcluded that the carbohydrate moiety of GSL-3 has thestructure of a a-d-Galp-(1!6)-a-d-GlcpN-(1!4)-a-d-GlcpAtrisaccharide.
D I S C U S S I O N
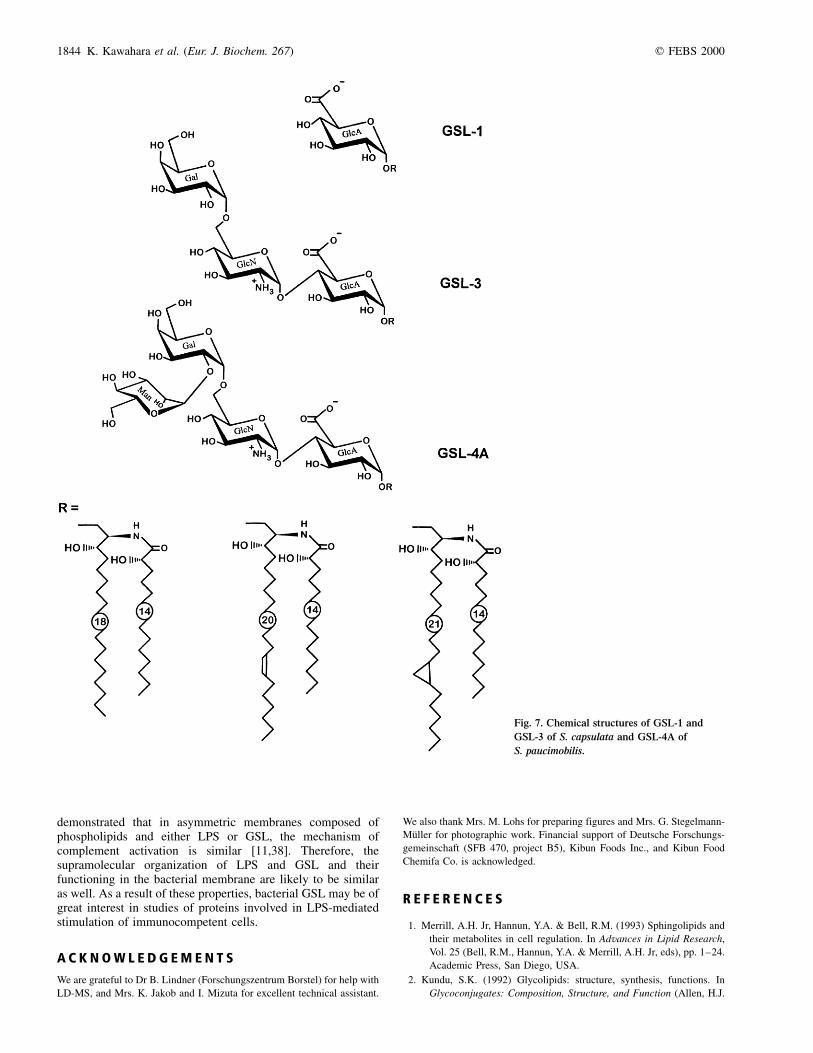
As a result of this work, we propose the chemical structures fortwo GSL isolated from S. capsulata (Fig. 7). Both GSL-1 andGSL-3 represent a mixture of three molecules, differing in thenature of the long-chain base. Two of the sphingoids present inS. capsulata have been previously reported as components ofGSL from S. paucimobilis [4], in particular of GSL-4A(Fig. 7). The third long-chain base, (13Z )-erythro-2-amino-13-eicosene-1,3-diol, has been characterized for the firsttime in S. capsulata as in GSL from S. paucimobilis it ispresent only as a minor component. Thus, the molar ratiosof erythro-2-amino-1,3-octadecanediol (13Z )-erythro-2-amino-13-eicosene-1,3-diol and (13Z )-erythro-2-amino-13,14-methyl-ene-1,3-eicosanediol were estimated as 1 : 1.2 : 1.5 and12.5 : 1 : 11.5, respectively, in GSL-3 from S. capsulata andGSL-4A from S. paucimobilis reinvestigated in this work. Allsphingolipids from both species contain (S )-2-hydroxymyristicacid as the only fatty acid component. GSL-3 from S. capsulatahas a truncated carbohydrate chain lacking the terminal Manresidue as compared to that of S. paucimobilis [4]. With respectto the carbohydrate moiety, the monoglycosyl GSL-1 fromS. capsulata is identical to GSL-1 from S. paucimobilis [4].
These data show that both glycosyl and sphingolipid partsof GSL are closely related by structure in the twoSphingomonas species studied. GSL from some otherspecies of Sphingomonas may vary not only in the length ofthe carbohydrate chain, but also in the sugar sequence [21]. Asis the case for S. paucimobilis [4], S. capsulata is devoid ofLPS and is thus the second Gram-negative bacterium, thatreplaces LPS in the outer membrane by GSL. Although GSL
have recently been found also in Chlorobium limicola [22] andZymomonas mobilis [23], they cannot be considered as commoncomponents of bacterial cells. It is worthy of noting that someother bacteria, such as Bacteroides [24±26], Bdellovibrio [27],Acetobacter [28], Flavobacterium (Sphingobacterium) [29],and the aquatic bacteria Arcocella aquatica NO-502 andFlectobacillus major FM [30] contain sphingolipids as cellconstituents.
Biosynthetic pathways of GSL in bacteria remain unknown.In yeast [31] and animal tissues [32], sphinganine issynthesized from palmitoyl-CoA and l-Ser. As palmitic acidand (11Z )-11-octadecenoic acid (cis-vaccenic acid) are themain cellular fatty acids of Sphingomonas [6], and the pathwayof the cyclopropyl ring formation from the correspondingunsaturated fatty acid is well known in Gram-negative bacteria[33], it is reasonable to hypothesize that sphingoids found inSphingomonas GSL are synthesized through pathways similarto those in eukaryotic cells, using fatty acyl-CoA available inbacterial cells. However, sphingosine, a 4,5-dehydrogenatedsphinganine, which is common in eukaryotic cells, has not yetbeen found in bacteria.
Remarkably, like enterobacterial LPS GSL-4A fromS. paucimobilis can induce interleukin-1 and interleukin-6in, and release tumour necrosis factor-a from, human mono-cytes, although the doses needed to reach the inducing potencyof LPS (1 ng´mL21) were 1000±10 000-fold higher [34].However, studies using mAbs against CD14 (MEK 18) havedemonstrated that the stimulation of monocytes by GSL is notmediated by CD14 and the mechanism of the GSL action isthus different from LPS [35]. An alternative way of tumournecrosis factor-a release and interleukin-1 stimulation washypothesized by Kolesnick and colleagues [36,37]. Theysuggested also that a structural similarity between lipid A andceramide may account for the ability of lipid A to stimulateceramidase in monocytes [37,38]. Furthermore, comparisonof physicochemical properties of Re LPS and GSL-1
Fig. 6. Part of a 90 MHz 13C,1H-COSY
spectrum of GSL-3 of S. capsulata. The
corresponding part of the 1H-NMR and 13C-NMR
spectra are displayed along the vertical and
horizontal axes, respectively. Arabic numerals
refer to atoms in the residues denoted as follows:
I, GlcA; II, GlcN; III, Gal; s, sphingoid.
1844 K. Kawahara et al. (Eur. J. Biochem. 267) q FEBS 2000
demonstrated that in asymmetric membranes composed ofphospholipids and either LPS or GSL, the mechanism ofcomplement activation is similar [11,38]. Therefore, thesupramolecular organization of LPS and GSL and theirfunctioning in the bacterial membrane are likely to be similaras well. As a result of these properties, bacterial GSL may be ofgreat interest in studies of proteins involved in LPS-mediatedstimulation of immunocompetent cells.
A C K N O W L E D G E M E N T S
We are grateful to Dr B. Lindner (Forschungszentrum Borstel) for help with
LD-MS, and Mrs. K. Jakob and I. Mizuta for excellent technical assistant.
We also thank Mrs. M. Lohs for preparing figures and Mrs. G. Stegelmann-
MuÈller for photographic work. Financial support of Deutsche Forschungs-
gemeinschaft (SFB 470, project B5), Kibun Foods Inc., and Kibun Food
Chemifa Co. is acknowledged.
R E F E R E N C E S
1. Merrill, A.H. Jr, Hannun, Y.A. & Bell, R.M. (1993) Sphingolipids and
their metabolites in cell regulation. In Advances in Lipid Research,
Vol. 25 (Bell, R.M., Hannun, Y.A. & Merrill, A.H. Jr, eds), pp. 1±24.
Academic Press, San Diego, USA.
2. Kundu, S.K. (1992) Glycolipids: structure, synthesis, functions. In
Glycoconjugates: Composition, Structure, and Function (Allen, H.J.
Fig. 7. Chemical structures of GSL-1 and
GSL-3 of S. capsulata and GSL-4A of
S. paucimobilis.

q FEBS 2000 Glycosphingolipids from Sphingomonas capsulata (Eur. J. Biochem. 267) 1845
& Kisailus, E.C., eds), pp. 203±262. Marcel Dekker, New York,
USA.
3. Rietschel, E.T., Brade, H., Holst, O., Brade, L., MuÈller-Loennies, S.,
Mamat, U., ZaÈhringer, U., Beckmann, F., Seydel, U., Brandenburg,
K., Ulmer, A.J., Mattern, T., Heine, H., Schletter, J., Loppnow, H.,
SchoÈnbeck, U., Flad, H.-D., Hauschildt, S., Schade, U.F., Di Padova,
F., Kususmoto, S. & Schumann, R.R. (1996) Bacterial endotoxin:
chemical constitution, biological recognition, host response, and
immunological detoxification. Curr. Top. Microbiol. Immunol. 216,
39±81.
4. Kawahara, K., Seydel, U., Matsuura, M., Danbara, H., Rietschel,
E.T. & ZaÈhringer, U. (1991) Chemical structure of glyco-
sphingolipids isolated from Sphingomonas paucimobilis. FEBS
Lett. 292, 107±110.
5. Yamamoto, A., Yano, I., Masui, M. & Yabuuchi, E. (1978) Isolation of
a novel sphingoglycolipid containing glucuronic acid and 2-hydroxy
fatty acid from Flavobacterium devorans ATCC 10829. J. Biochem.
(Tokyo) 83, 1213±1216.
6. Yabuuchi, E., Yano, I., Oyaizu, H., Hashimoto, Y., Ezaki, T. &
Yamamoto, H. (1990) Proposals of Sphingomonas paucimobilis
gen. nov. & comb. nov., Sphingomonas parapaucimobilis sp.
nov., Sphingomonas yanoikuyae sp. nov., Sphingomonas adhae-
siva sp. nov., Sphingomonas capsulata comb. nov. & two
genospecies of the genus Sphingomonas. Microbiol. Immunol. 34,
99±119.
7. Leifson, E. (1962) The bacterial flora of distilled and stored water. III.
New species of the genera Corynebacterium, Flavobacterium,
Spirillum and Pseudomonas. Int. Bull. Bacteriol. Nom. Tax. 12,
161±170.
8. Takeuchi, M., Sawada, H., Oyaizu, H. & Yokota, A. (1994)
Phylogenetic evidence for Sphingomonas and Rhizomonas as
nonphotosynthetic members of the alpha-4 subclass of the Proteo-
bacteria. Int. J. Syst. Bacteriol. 44, 308±314.
9. Balkwill, D.L., Drake, G.R., Reeves, R.H., Fredrickson, J.K., White,
D.C., Ringelberg, D.B., Chandler, D.P., Romine, M.F., Kennedy,
D.W. & Spadoni, C.M. (1997) Taxonomic study of aromatic-
degrading bacteria from deep-terrestrial-subsurface sediments and
description of Sphingomonas aromaticivorans sp. nov., Sphingo-
monas subterranea sp. nov. & Sphingomonas stygia sp. nov. Int. J.
Syst. Bacteriol. 47, 191±201.
10. Kawasaki, S., Moriguchi, R., Sekiya, K., Nakai, T., Ono, E., Kume, K.
& Kawahara, K. (1994) The cell envelope structure of the
lipopolysaccharide-lacking gram-negative bacterium Sphingomonas
paucimobilis. J. Bacteriol. 176, 284±290.
11. Wiese, A., Reiners, J.O., Brandenburg, K., Kawahara, K., ZaÈhringer, U.
& Seydel, U. (1996) Planar asymmetric lipid bilayers of glyco-
sphingolipid or lipopolysaccharide on one side and phospholipids on
the other: membrane potential, porin function, and complement
activation. Biophys. J. 70, 321±329.
12. Seydel, U., Lindner, B., Wollenweber, H.-W. & Rietschel, E.T. (1984)
Structural studies on the lipid A component of enterobacterial
lipopolysaccharides by laser desorption mass spectrometry. Location
of acyl groups at the lipid A backbone. Eur. J. Biochem. 145,
505±509.
13. Strominger, J.L., Park, J.T. & Thompson, R.E. (1959) Composition of
the cell wall of Staphylococcus aureus: its relation to the mechanism
of action of penicillin. J. Biol. Chem. 234, 3263±3267.
14. Bitter, T. & Muir, H.M. (1962) A modified uronic acid carbazole
reaction. Anal. Biochem. 4, 330±334.
15. Hakomori, S. (1964) A rapid permethylation of glycolipid, and
polysaccharide catalyzed by methylsulfinyl carbanion in dimethyl
sulfoxide. J. Biochem. (Tokyo) 55, 205±208.
16. Helander, I., Lindner, B., Brade, H., Altmann, K., Lindberg, A.A.,
Rietschel, E.T. & ZaÈhringer, U. (1988) Chemical structure of the
lipopolysaccharide of Haemophilus influenzae strain I-69 Rd-/b1.
Description of a novel deep-rough chemotype. Eur. J. Biochem. 177,
483±492.
17. Kawahara, K., Brade, H., Rietschel, E.T. & ZaÈhringer, U. (1987)
Studies on the chemical structure of the core-lipid A region of the
lipopolysaccharide of Acinetobacter calcoaceticus NCTC 10305.
Detection of a new 2-octulosonic acid interlinking the core
oligosaccharide and lipid A component. Eur. J. Biochem. 165,
489±495.
18. Gunstone, F.D. (1993) High resolution 13C NMR spectroscopy of
lipids. In Advances in Lipid Methodology 2 (Christie, W.W., ed.),
pp. 1±68. The Oily Press, Dundee, UK.
19. Bock, K. & Pedersen, C. (1983) Carbon-13 nuclear magnetic resonance
spectroscopy of monosaccharides. Adv. Carbohydr. Chem. Biochem.
41, 27±66.
20. Lipkind, G.M., Shashkov, A.S., Knirel, Y.A., Vinogradov, E.V. &
Kochetkov, N.K. (1988) A computer-assisted structural analysis of
regular polysaccharides on the basis of 13C-NMR data. Carbohydr.
Res. 175, 59±75.
21. Kawahara, K., Kuraishi, H. & ZaÈhringer, U. (1999) Chemical
structure and function of glycosphingolipids of Sphingomonas
spp. and their distribution among members of the a-4
subclass of Proteobacteria. J. Ind. Microbiol. Biotech. 23,
408±413.
22. Jensen, M.T., Knudsen, J. & Olson, J.M. (1991) A novel aminogly-
colipid found in Chlorobium limicola f. thiosulfatophilum 6230. Arch.
Microbiol. 156, 248±254.
23. Tahara, Y. & Kawazu, M. (1994) Isolation of glucuronic acid-
containing glycosphingolipid from Zymomonas mobilis. Biosci.
Biotech. Biochem. 58, 586±587.
24. Riazza, V., Tucker, A.N. & White, D.C. (1970) Lipids of Bacteroides
melaninogenicus. J. Bacteriol. 101, 84±91.
25. Kunsman, J.E. (1973) Characterization of the lipids of six strains of
Bacteroides ruminicola. J. Bacteriol. 113, 1121±1126.
26. Miyagawa, E., Azuma, R., Suto, T. & Yano, I. (1979) Occurrence
of free ceramides in Bacteroides fragilis. J. Biochem. (Tokyo) 86,
311±320.
27. Steiner, S., Conti, S.F. & Lester, R.L. (1973) Occurrence of
phosphonosphingolipids in Bdellovibrio bacteriovorus strain UKi2.
J. Bacteriol. 106, 1199±1211.
28. Tahara, Y., Nakagawa, A. & Yamada, Y. (1986) Occurrence of
free ceramide in. Acetobacter xylinum. Agric. Biol. Chem. 50,
2949±2950.
29. Yabuuchi, E., Kaneko, T., Yano, I., Moss, C.W. & Miyoshi,
N. (1983) Sphingobacterium gen. nov., Sphingobacterium
spiritivorum comb. nov., Sphingobacterium multivorum comb.
nov., Sphingobacterium mizutae sp. nov. & Flavobacterium
indologenes sp. nov. glucose-nonfermenting Gram-negative rods
in CDC group IIK-2 and Iib. Int. J. Syst. Bacterol. 33,
580±598.
30. Batrakov, S.G., Sheichenko, V.I. & Nikitin, D.I. (1999) A novel
glycosphingolipid from Gram-negative aquatic bacteria. Biochim.
Biophys. Acta 1440, 163±175.
31. Braun, P.E. & Snell, E.E. (1968) Biosynthesis of sphingolipid bases II.
Keto intermediates in synthesis of sphingosine and dihydrosphingo-
sine by cell-free extracts of Hansenula ciferri. J. Biol. Chem. 243,
3775±3783.
32. Braun, P.E., Morell, P. & Radin, N.S. (1970) Synthesis of C18- and
C20-dihydrosphingosines, ketodihydrosphingosines, and ceramides
by microsomal preparations from mouse brain. J. Biol. Chem. 245,
335±341.
33. Wilkinson, S.G. (1988) Distribution of lipids. 7. Gram-negative
bacteria. In Microbial Lipids, Vol. 1 (Ratledge, C. & Wilkinson,
S.G., eds), pp. 299±488. Academic Press, London, UK.
34. Krziwon, C., ZaÈhringer, U., Kawahara, K., Weidemann, B., Kusumoto,
S., Rietschel, E.T., Flad, H.-D. & Ulmer, A.J. (1995) Glyco-
sphingolipids from Sphingomonas paucimobilis induce monokine
production in human mononuclear cells. Infect. Immun. 63,
2899±2905.
35. Hailman, E., Lichenstein, H.S., Wurfel, M.M., Miller, D.S.,
Johnson, D.A., Kelley, M., Busse, L.A., Zukowski, M.M. &
Wright, S.D. (1994) Lipopolysaccharide (LPS)-binding protein
accelerates the binding of LPS to CD14. J. Exp. Med. 179,
269±277.

1846 K. Kawahara et al. (Eur. J. Biochem. 267) q FEBS 2000
36. Joseph, C.K., Wright, S.D., Bornmann, W.G., Randolph, J.T., Kumar,
E.R., Bittman, R., Liu, J. & Kolesnick, R.N. (1994) Bacterial
lipopolysaccharide has structural similarity to ceramide and stimu-
lates ceramide-activated protein kinase in myeloid cells. J. Biol.
Chem. 269, 17606±17610.
37. Kolesnick, R. & Golde, D.W. (1994) The sphingomyelin pathway
in tumor necrosis factor and interleukin-1 signaling. Cell 77,
325±328.
38. MuÈnstermann, M., Wiese, A., Brandenburg, K., ZaÈhringer, U., Brade,
L., Kawahara, K. & Seydel, U. (1999) Complement activation by
bacterial surface glycolipids: a study with planar bilayer membranes.
J. Membr. Biol. 167, 223±232.





























