Embed Size (px)
Citation preview

Plant Molecular Biology 28: 189-194, 1995. © 1995 Kluwer Academic Publishers. Printed in Belgium.
Short communication
189
Structural organization and differential expression of carrot fl-frnctofuranosidase genes: identification of a gene coding for a flower bud-specific isozyme
Kathrin Lorenz, Susanne Lienhard and Arnd Sturm* Friedrich Miescher-Institut, Postfach 2543, CH-4002 Basel Switzerland," (* author for correspondence)
Received 2 September 1994; accepted in revised form 5 January 1995
Key words: fl-fructofuranosidase, invertase, gene expression, gene structure, flower buds, Daucus carota
Abstract
Three genomic clones (Inv*Dcl, Inv*Dc2 and Inv*Dc3) were isolated by using the cDNA for carrot cell wall fl-fructofuranosidase as a probe. The expression patterns of the three genes differed markedly. High levels of Inv*Dcl transcripts were found in leaves and roots of young carrot, whereas in plants with developing tap roots no transcripts were detected. A high level of mRNA of Inv*Dcl was also present in suspension-cultured cells. In developing reproductive organs, only low levels of transcripts of Inv*Dcl were found in flower buds and flowers and none at later stages of development. In contrast, Inv*Dc2 and Inv*Dc3 were not expressed in vegetative plant organs. Invbl *Dcl was exclusively and strongly expressed in flower buds, and Inv*Dc3 at a very low level in suspension-cultured cells.
Acid fl-fructofuranosidase (13 F) (invertase), which hydrolyses sucrose into glucose and fructose, is of ubiquitous occurrence in the plant kingdom. Most higher plants contain several forms of this en- zyme, which can be distinguished by their iso- electric points, solubilities and subcellular loca- tions. Carrot contains at least three different acid flF isoenzymes. Two of them accumulate as soluble proteins in the vacuole [6, 16] and one is ionically bound to the cell wall [6, 13]. A com- parison of their cDNA-derived amino acid se- quences reveals that all three enzymes are pre-
proproteins with signal peptides, N-terminal propeptides, and long domains constituting the mature polypeptides [15]. In contrast to the se- quence of the cell wall enzyme, the sequences of the vacuolar acid flFs include short C-terminal extensions which may contain the information for vacuolar targeting. Marked differences among the three nucleotide sequences as well as different hybridization patterns on genomic DNA gel blots prove that the three isoenzymes of carrot acid flF are encoded by different genes [15].
The cell wall-bound enzyme is N-glycosylated
The nucleotide sequence data reported will appear in the EMBL, GenBank and DDBJ Nucleotide Sequence Databases under the accession numbers X69321, X78423 and X78424.

190
like the vacuolar isoforms [6, 12]. It has an ap- parent molecular mass of 63 kDa, a pH optimum of 4.6 and a basic pI of 9.9 [6]. The function of the cell wall flF is not clear. It has been suggested that this enzyme plays a key role in photoassimi- late partitioning between source and storage sink tissues [3 ]. The lack of both transcripts and en- zyme activity in developing tap roots contradicts this hypothesis and suggests a different function [141.
Three genomic clones, which strongly hybrid- ized to the coding-sequence derived from the 5'- half of the carrot cell wall/~F cDNA [13], were isolated from carrot DNA and sequenced. The clones were named Inv*Dcl, Inv*Dc2 and Inv*Dc3 according to Jansson [ 5 ]. The structure and the promoter of the genomic clone Inv*Dcl, which codes for cell wall /~F purified from suspension-cultured carrot cells [13], have al- ready been described by Ramloch-Lorenz [8]. The intron sequences were deduced by compar- ing the protein-coding sequence of the cDNA with the sequence of Inv*Dcl. They have the bound- ary sequences GT/AG typical of dicotyledonous plant introns [4]. The coding region of this gene is organized into 6 exons and 5 introns. The des- ignation of the exon sequences of the two other genomic clones (Inv*Dc2 and Inv*Dc3) was based on their high level of sequence identity to
the exon sequences of Inv*Dcl. The structure of Inv*Dcl together with the structures of Invb *Dcl and Inv*Dc3 are shown in Fig. 1.
A comparison of the 5'-upstream sequences of the three genes revealed no regions of significant similarity. Whereas a TATA and a CAAT box have been identified in Inv*Dcl 99 bp upstream of the translation initiation codon [8], such cis elements have not been detected in the upstream sequences of lnv*Dc2 and Inv*Dc3 (780 bp and 920 bp were analysed, respectively). Because we only determined the transcription start site of Inv*Dcl [8], we cannot rule out the possibility that the other two genes contain upstream intron sequences.
The two genomic clones lnv*Dc2 and Inv*Dc3 code for polypeptides highly related to the puri- fied cell wall enzymes (see below). Their genomic structures are very similar to that oflnv*Dcl with two exceptions: their first introns are much smaller (318 bp and 517 bp, respectively) and in both genes exon 2 of the Inv*Dcl gene is sepa- rated by an additional intron into two protein- coding domains, the first of which is very short (9 bp). The structures of the genes for tomato vacu- olar flF (accession number Z12027) [2] and Ara- bidopsis cell wall flF (accession number X74514) [9], the only other plant flF genes so far isolated, are very similar to the structures of Inv*Dc2 and
o 1 I I
Z n v ~ D o l
2 I
K S Y i i i
+1 r i
3 4 5 6 I I I I
E X I !
z KV
m
H X S ! I i
Y EE BE H E I I# I I ' I
m m . I
!
B E B H ! I I I
Z n ~ D c ' 2 i : m m , , m mm m m m m m
I I
Y K E B S E B E E B X H I I I II I I I ! I I I
EE ! I
Z n ~ 3 I ! I I _ _ 1 __1 I I | I
! I
Fig. 1. Restriction and structural maps of the genomic subclones for Inv*Dcl, Inv*Dc2 and lnv *Dc3. Three different genomic clones (6.6 kb, 3.5 kb and 4.6 kb, respectively) were isolated from DNA of wild carrot cv. Queen Anne's Lace and sequenced. The upper line represents the scale in kilobases. Signs on the restriction maps are as follows: B, Barn HI; E, Eco RI; H, Hind III; K, Kpn I; S, Sph I; V, Sst I; X, Xba I; Y, Sal I; Z, Sma I. On the structural maps, the coding regions are represented by filled boxes with introns shown as a line. The sequences used as probes are marked by bars below the structural maps.

1 50 Inv3 MART- - -KILVFSSDSSLFLLS IFSF IF- - -LNINGVDSTHRVFPELQS
: I I: : " I:::11 III :11 I1: I I I I I I I I inv2 ML IRCFHIKMALVTCFHSMLFLSAVVFIFSLDVNIRGVEASHQVFPELQS
: : : : I I:::: ::I llll I': :Ill invl MGWr I ~ ' ~ Y D H G S L P F ~ S L ~ I LLWPI"PI~H I N G V ' ~ I ~ I I I Y N I ~
51 100 i n v 3 I SAVD~VIIR'£~YIIFQPQI~m I I~I~I'GP~YKGYYltLFYQYI~KGSVWG
:111:1 I I I I I I l l l / I I I I l l l l l l l l : l l l : l l l l l l l l l l l I I I inv2 VSAVI~/QLVHR~'ntFQPKI~gIINDPIIG~HId~YQYNPKGAVWG
I:1 II I I I I I I I I / 1 1 : l l l l l l l l l l l l l I I I I I /111111111 i n v l VGAENVKQVII~GYltFQPK01mII~PNGPI4YYKGWZHLF YQ~KG&VWG
101 150 inv3 NI~AHSVSKDL I ~ I ~ E P A I FPSI~FDQYGOWSGSATILPGNKPVILY
I I I I I I l : l l l l l l l : l l l l l l l l l l l l l I l l l l l l l l : l l l I I I1 :1 inv2 NIVWAHS ISKDL IliWVALEPAIFPSI~PFDKYGOISGSATVLPGGKPVIMY
I I I I I I 1 : 1 I I I I I : l l l l l l l l l l l l l l l l : l l l l l : l l l I I I1 :1 i n v l NI~WAHSV STDL I liWI~LEPAI FP SKPFDI~/GCR SGSAT ILPG~I~VI LY
151 200 inv3 TG I VS PDPENAQVQNYAVPANYSDPFLREWVKPDNNPLVGV- HTENP SAF
fill I I I llll:[Illlllll:llll-'lllllI-'l : II I I inv2 TGIVTPSPVNTQVQNFAVPANYSDPYLREWZKPDNNP IVRA-RSENSS SF
I I I I I I I I I l : l : l l l I I I I I I I I I I I I I 1 : 1 I :11 I i n v l TGIVEGPPI~P/~/QI~/AI PANLS DP~ Paoli ~DNI~LW/dlNGENATAF
201 250 ~nv3 RDPTTAWFD- GGHWKMLVGS SRKHRG IAYLYRSKDFKKWKRS PHP I HTKA
IIIIIIIII I III:IIII lllllllllIll.':l II : II:I I inv2 RDPTTAWFD - GVHWK I LVGS RRKHRG IAYLYRS RNF LKWTKAKHP LHS KD
[IIIIII:I : lll:llIl: :IIIIIIIII::I.'IIIIIIII II invl RDPTTAWLDKS GHWIOfLVGSKRNRRGI AYLYRS KDF IKWTKAEHP I H S QA
251 300 inv3 ETGMWECPDFYPVSPRSEDGLDNSKMGRGI KHVLKVSLNSTRYEYYT I GR
l l l l l l I I I I I I : : : I I I I I : I I I I I I I I I l l l l l l l l : l inv2 RTGMWECLDFYPVAPKGMNGLDTSVTGQD IKHVLKVSLYSTRYEYYTVGE
IIIIII ]I:II II:IIIIIIIII: :LIIIIIII IIIIIIIII invl NTGMWECPDFFPVSLKGLNGLDTSVTGE SVKHVLKVSL DLTRYEYYTVGT
301 ~ 350 inv3 YNRVRDFYVPDNTSVDGWAGLRYDYGNFYASKTFYDP I KKRRI LWGWANE
I:I l:llIllllllllllllllllllllllll:l[ I IIllllllll inv2 YDRDND I YVPDNTSVDGWAGLRYDYGNFYASKTFFDPDKQRR I LWGWANE
I I I l'lllIllllllllll[lllllllllllll I IIIIIIIIII Invl YLTDKDRY I PDNTSVDGWAGLRYDYGNFYASKTFFDP SEI~RR I LWGWANE
351 400 Inv3 SDSQIDDVQKGWAGIQLIPRRIWLDPSGRQLVQWPIEEVEGLRGSELHM-
I I I III/111111/1111::1111 I:/1:111111:: I I I ::11 inv2 SDSKQDDVQKGWAGIQLIPRKLWLDPNGKQLIQWPIEEIQLLRGQNVHM-
III II IIIIIIIIIII IIIII IIII:IIIIII:: Ill I : Invl SD STAHDVAKGWAG I QL I pRTLWLDP SGKQLMQWP I EELETLRGS KVKF S
401 450 inv3 RNQKLDMGVHVEVTGI TAAQADVDATF S FKS LDKAE S FDP EWINLDAQDV
! i: I I:II l:lllllllllllllllll-'I] III:I III II Inv2 GSQVLNTGEH I EVKGVTAAQADVDATF S FK S LDRAEWFDPNWAKLDALDV
/ I I :1111:1111111: I I I I I I I : I I I I I II : i n v l RKQDL S KGI LVEVKGITAAQADVEVTF S FK S LAKRE pFDPKWLEYDAEKI
451 500 ~nv3 CDSMGSTIQGGLGPFGLLTLASKDLEEYTPVFFRIFKAEDQKLKVLMCSD
II IIII: IIIIIII:IIIII lllillllll:ll I lllllIlll Inv2 CDWMGS TVRGGLGPFGFLTLAS EKLEEYTPVFFR~TKD- KLKVLI~C S D
I : IIII II:IIII:IIIIIIIIIIIIIIIIIIII : IIIIIII ~nvl CSLKGS TVQGGVGPFGLLTLASEKLEEYTPVFFRVFKAQNTH- KVLMC S D
501 550 inv3 AKRSS-LAEGLYKPSFRGFVDVDLS -DKKI SLRSLZ DNSVVESFGAQRKN
I I I I I I I ] I I I I I I - ' I I I I I I l l l l l l l l l l l l l l i l l l i l : : inv2 AKRSSTTAEGLYKPPFAGYVDVDLS -DKKI SLRSLIDNSVVESFGAHGRT
I III IIII:I IIl:lllll lllll[IIIIIlllIlllIl I:I invl ATRS S-LKEGLYRPSFAGFVDVDLATDKKI SLRSLIDNSVVESFGAKGKT
551 597 inv3 L I S SRVYPTLAIYNNAHLFVFNNGTEP ITVDNLDAWS~SPSE~
[ I I I I I : l l l l l l l : l l l l l l l l : l i ' I I I I I I I II II inv2 CI TSRVYPKIAIYNNAHVFVFNNGTEAIT IDSLDAWSMKAPSLMNNN
II IIIII :l-'l:lll::IIlll I II:: IIIIIII I II invl C I S S RVYPTLAVYEIqAHLYVFNNGS ET I TVENLDAW SMKKPLI~4N
Fig. 2. Comparison of the amino acid sequence deduced for Inv*Dcl (invl) with the amino acid sequences deduced for Inv*Dc2 (inv2) and lnv*Dc3 (inv3). The amino acid sequences are in the one-letter code and have been aligned by introducing gaps (---) to maximize identities. Vertical bars indicate iden-
191
Inv*Dc3 and also contain a second exon of 9 bp. This short exon encodes part of a highly con- served region (NDPNG; flF motif, indicated in Figure 2) found in all known flF proteins [ 13,9]. The coding sequences of the tomato and the Ara- bidopsis flF gene are 50-60 ~o identical to the cod- ing sequences of In v*Dcl, Inv*Dc2 and Inv*Dc3, whereas the coding sequences of the three carrot flF genes share 71-76~o identity.
The deduced amino acid sequences derived from the three genomic clones were compared (Fig. 2). The polypeptides encoded by Inv*Dcl, Inv*Dc2 and Inv*Dc3 have calculated molecular masses of 66.8 kDa, 67.4 kDa and 66.4 kDa, and isoelectric points of 9.9, 8.9 and 7.4, respectively. In analogy to cell wall flF [13] encoded by Inv*Dcl, the two other proteins may also be pre- proproteins with signal sequences, N-terminal propeptides and long domains constituting the mature polypeptides. The three protein sequences contain several potential N-glycosylation sites (Asn-X-Ser/Thr) in addition to the 'flF motif and a conserved cysteine residue, which are hallmarks offlF protein sequences [ 13 ]. The three sequences are ca. 75~o identical. In contrast to the cDNA- deduced amino acid sequence of the carrot vacu- olar flFs [15], the deduced amino acid sequences of the three carrot flF genes lack C-terminal ex- tensions. Because these C-terminal sequences may contain the information for vacuolar target- ing [7,1], it is likely that Inv*Dc2 and Inv*Dc3 code for two further cell wall flF isoforms.
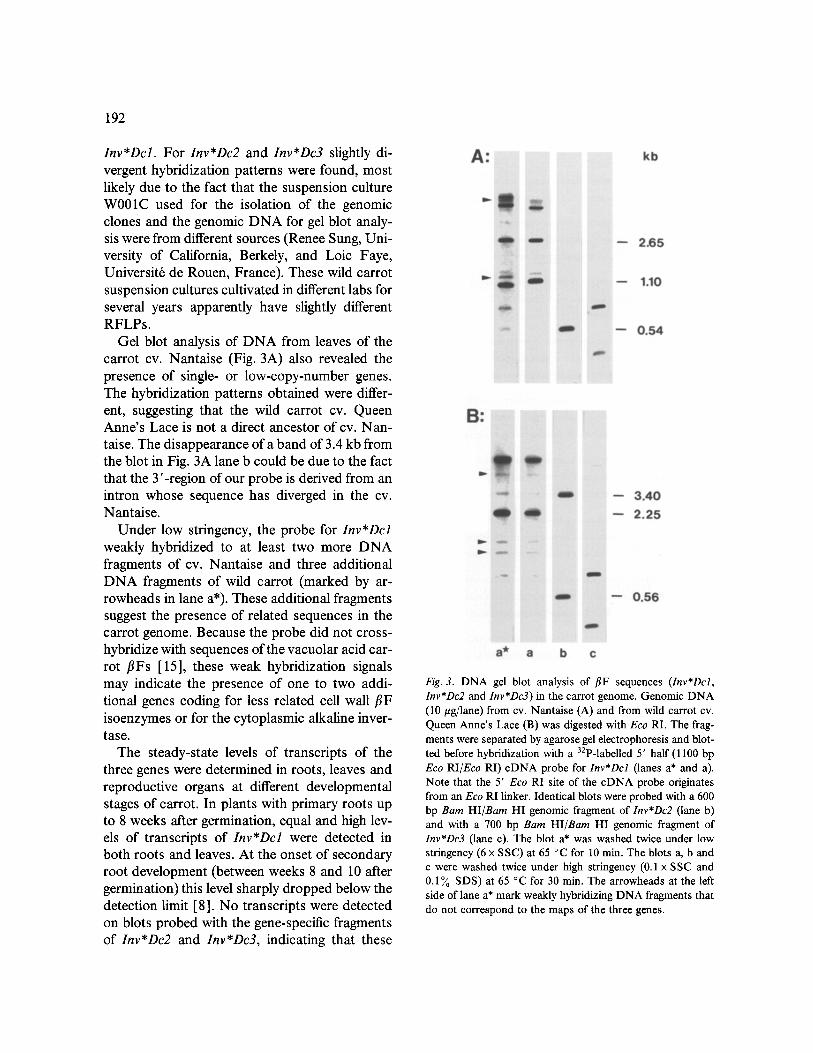
DNA gel blot analyses [11] were performed with Eco RI-restricted DNA from wild carrot (Fig. 3B). The probes used are marked in Fig. 1 and share about 70 ~o identity. Under high strin- gency, the probes hybridized to only a few restric- tion fragments and very little cross-hybridization was observed. The lengths of the fragments to which the probes hybridized only correspond to the lengths predicted from the gene map for
tical residues and colons indicate conservative replacements. The first amino acid residue of the mature polypeptide en- coded by Inv*Dcl [13] is marked by a black dot. The '/~F motif' (underlined) and the cysteine residue (arrow) are con- served elements among bacterial, yeast and plant flFs [13].
192
Inv*Dcl. For Inv*Dc2 and Inv*Dc3 slightly di- vergent hybridization patterns were found, most likely due to the fact that the suspension culture W001C used for the isolation of the genomic clones and the genomic DNA for gel blot analy- sis were from different sources (Renee Sung, Uni- versity of California, Berkely, and Loic Faye, Universit6 de Rouen, France). These wild carrot suspension cultures cultivated in different labs for several years apparently have slightly different RFLPs.
Gel blot analysis of DNA from leaves of the carrot cv. Nantaise (Fig. 3A) also revealed the presence of single- or low-copy-number genes. The hybridization patterns obtained were differ- ent, suggesting that the wild carrot cv. Queen Anne's Lace is not a direct ancestor of cv. Nan- taise. The disappearance of a band of 3.4 kb from the blot in Fig. 3A lane b could be due to the fact that the 3'-region of our probe is derived from an intron whose sequence has diverged in the cv. Nantaise.
Under low stringency, the probe for Inv*Dcl weakly hybridized to at least two more DNA fragments of cv. Nantaise and three additional DNA fragments of wild carrot (marked by ar- rowheads in lane a*). These additional fragments suggest the presence of related sequences in the carrot genome. Because the probe did not cross- hybridize with sequences of the vacuolar acid car- rot flFs [15], these weak hybridization signals may indicate the presence of one to two addi- tional genes coding for less related cell wall flF isoenzymes or for the cytoplasmic alkaline inver- tase.
The steady-state levels of transcripts of the three genes were determined in roots, leaves and reproductive organs at different developmental stages of carrot. In plants with primary roots up to 8 weeks after germination, equal and high lev- els of transcripts of Inv*Dcl were detected in both roots and leaves. At the onset of secondary root development (between weeks 8 and 10 after germination) this level sharply dropped below the detection limit [8]. No transcripts were detected on blots probed with the gene-specific fragments of Inv*Dc2 and Inv*Dc3, indicating that these
Fig. 3. DNA gel blot analysis of flF sequences (Inv*Dcl, Inv*Dc2 and Inv*Dc3) in the carrot genome. Genomic DNA (10 #g/lane) from cv. Nantaise (A) and from wild carrot cv. Queen Anne's Lace (B) was digested with Eco RI. The frag- ments were separated by agarose gel electrophoresis and blot- ted before hybridization with a 32p-labelled 5' half (1100 bp Eco RI/Eco RI) cDNA probe for Inv*Dcl (lanes a* and a). Note that the 5' Eco RI site of the cDNA probe originates from an Eco RI linker. Identical blots were probed with a 600 bp Bam HI~Barn HI genomic fragment of Inv*Dc2 (lane b) and with a 700 bp Barn HI/Bam HI genomic fragment of Inv*Dc3 (lane c). The blot a* was washed twice under low stringency (6 x SSC) at 65 °C for 10 min. The blots a, b and c were washed twice under high stringency (0.1 x SSC and 0.1% SDS) at 65 °C for 30 min. The arrowheads at the left side of lane a* mark weakly hybridizing DNA fragments that do not correspond to the maps of the three genes.

193
genes were not or only poorly expressed in the organs analysed. Figure 4 shows as an example the steady-state mRNA levels for Inv*Dcl and Inv*Dc2 in 4- and 14-week-old leaves and roots.
Gel blot analyses of RNA from flower buds, flowers, developing seeds and mature seeds showed a low level of Inv*Dcl mRNA in flower buds and flowers, and none in seeds. In contrast, a high level of Inv*Dc2 transcripts was found in flower buds and none in flowers or seeds. Tran- scripts oflnv*Dc3 were not detected in reproduc- tive organs.
The gene for the purified cell wall flF was shown to be transiently expressed in response to wound- ing and infection [ 13]. In contrast, no wound- or pathogen-induced accumulation of mRNA of the other two genes was observed (data not shown). The expression of the three genes in suspension- cultured carrot cells also differed. Whereas the transcript levels of lnv*Dcl were high [ 13], only very low levels oflnv*Dc3 were detected, mRNA of Inv*Dc2 was not found (data not shown).
Cell wall ~F may control the exit of sucrose from young leaves and the transport of the dis-
accharide into developing primary roots (utiliza- tion sinks). Hydrolysis of some of the sucrose by cell wall flF may retain assimilates in the leaves, thereby promoting leaf growth and development. The remaining part of the sucrose may be loaded into the phloem and subsequently used for peti- ole and root growth. Developing petioles and roots as well as the reproductive organs are strong utilization sinks which are dependent on the im- port of assimilates from leaves. High transcript levels oflnv*Dcl in petioles and young roots [ 14], which also correlate with high activity of a cell wall/~F in these organs, indicate that this enzyme may play a role in sucrose partitioning between leaves and some of the utilization sinks of the plant. Hydrolysis of sucrose by the cell wall/~F may provide the developing organs with hexoses and may also prevent reloading of sucrose into the phloem [3]. Furthermore, cell wall/3F activ- ity may also be involved in cell growth and dif- ferentiation. In carrot tissue cultures, for example, marked changes in the activities of the different /~Fs, as well as their distribution between cell fractions, appear to be intimately related to the process of cell differentiation [10]. Cell wall/~F also seems to be involved in the response of plants to wounding and infection [ 13 ]. The homology of the carrot enzyme to the levan hydrolysing en- zyme levanase suggests that it may hydrolyse the slime coat of bacterial pathogens, thereby inhib- iting bacterial growth directly or making the pathogen susceptible to further defense reactions. Thus, the presence of extracellular/~F in flower buds may have a role in protecting this organ against pathogen attack.
Acknowledgements
Fig. 4. Steady-state mRNA levels of lnv*Dcl and Inv*Dc2 in leaves and roots of two different developmental stages and in reproductive organs of carrot plants cv. Nantaise. Upper panel: total RNA from flower buds (B), flowers (F), develop- ing seeds (G), mature seeds (S) and 4- and 14-week-old leaves and roots was analysed on a RNA gel blot (10 #g/lane). The blot was probed with a 32p-labelled gene-speciflc DNA frag- ment of Inv*Dcl. Lower panel: an identical blot was probed with a 32p-labelled gene-specific DNA fragment of Inv*Dc2.
We thank Roland Beffa, Patrick King, Orturn Mittelsten-Scheid and Christoph Unger for criti- cal reading of the manuscript.

194
References
1. Bednarek SY, Raikhel NV: The barley lectin carboxyl- terminal propeptide is a vacuolar protein sorting deter- minant in plants. Plant Cell 3:1195-1206 (1991).
2. Elliott KJ, Butler WO, Dickinson CD, Konno Y, Vedvick TS, Fitzmaurice L, Mirkov TE: Isolation and character- ization of fruit vacuolar invertase genes from two tomato species and temporal differences in mRNA levels during fruit ripening. Plant Mol Biol 21:515-524 (1993).
3. Eschrich W: Free space invertase, its possible role in phloem unloading. Ber Deut Bot Ges 93:363-378 (1980).
4. Goodall GJ, Kiss T, Filipowicz W: Nuclear mRNA splic- ing and small nuclear RNAs and their genes in higher plants. Oxford Surv Plant Mol & Cell Biol 7:255-296 (1991).
5. Jansson S, Pichersky E, Bassi R, Green BR, Ikeuchi M, Melis A, Simpson DJ, Spangfort M, Staehelin LA, Thornber JP: A nomenclature for the genes encoding the chlorophyll a/b-binding proteins of higher plants. Plant Mol Biol Rep 10:242-253 (1992).
6. Laurirre C, Laurirre M, Sturm A, Faye L, Chrispeels MJ: Characterization of fl-fructosidase, an extracellular gly- coprotein of carrot cells. Biochimie 70:1483-1491 (1988).
7. Neuhaus JM, Sticher L, Meins F Jr, Boiler T: A short C-terminal sequence is necessary and sufficient for the targeting of chitinases to the plant vacuole. Proc Natl Acad Sci USA 88:10362-10366 (1991).
8. Ramloch-Lorenz K, Knudsen S, Sturm A: Molecular characterization of the gene for carrot cell wall fl-fructosidase. Plant J 4:545-554 (1993).
9. Schwebel-Dugu6 N, E1 Mtili N, Krivitzky M, Jean- Jacques I, Williams JHH, Thomas M, Kreis M, Lecharny A: Arabidopsis gene and cDNA encoding cell-wall inver- tase. Plant Physiol 104:809-810 (1994).
10. Silva MP, Ricardo CPP: fl-Fructosidases and in vitro dedifferentiation-redifferentiation of carrot cells. Phy- tochemistry 31:1507-1511 (1992).
11. Southern E: Detection of specific sequences among DNA fragments separated by gel electrophoresis. J Mol Bio198: 503-517 (1975).
12. Sturm A: Heterogeneity of the complex N-linked oligo- saccharides at specific glycosylation sites of two secreted carrot glycoproteins. Eur J Biochem 199: 169-179 (1991).
13. Sturm A, Chrispeels M: cDNA cloning of carrot extra- cellular fl-fructosidase and its expression in response to wounding and infection. Plant Cell 2:1107-1119 (1990).
14. Sturm A, Sebkovgt V, Lorenz K, Hardegger M, Lienhard S, Unger C: Development- and organ-specific expression of the genes for sucrose synthase and three isoenzymes of acid fl -fructofuranosidases in carrot. Planta 195, 601-610 (1994).
15. Unger C, Hardegger M, Lienhard S, Sturm A: cDNA cloning of carrot soluble acid fl-fructofuranosidases and comparison with the cell wall isoenzyme. Plant Physiol 104:1351-1357 (1994).
16. Unger C, Hofsteenge J, Sturm A: Purification and char- acterization of a soluble fl-fructofuranosidase from Dau- cus carota. Eur J Biochem 204:915-921 (1992).





























