Embed Size (px)
Citation preview

Struktur und Dynamik
heterotropher Bakteriengemeinschaften im Wattenmeer
und der Deutschen Bucht
Structure and dynamics of heterotrophic bacterial
communities in the German Wadden Sea
and the German Bight
Dissertation
zur Erlangung des akademischen Grades einer
Doktorin der Naturwissenschaften (Dr. rer. nat.)
der Fakultät V Mathematik und Naturwissenschaften
der Carl von Ossietzky Universität Oldenburg
vorgelegt von
Beate Rink
geboren am 23.01.1974 in Bremerhaven

Erstgutachter : Prof. Dr. Meinhard Simon
Zweitgutachter: Prof. Dr. Heribert Cypionka
Eingereicht am:
Disputation am:

Für Rosemarie

Erklärung
Teilergebnisse dieser Arbeit sind als Beiträge bei den genannten Fachzeitschriften eingereicht
oder werden eingereicht. Mein Beitrag an der Erstellung der verschiedenen Manuskripte wird
im Folgenden erläutert:
Rink, B., Seeberger, S., Martens, T., Duerselen, C. D., Simon, M., und Brinkhoff, T. (2006)
Effects of a phytoplankton bloom in a coastal ecosystem on the composition of bacterial
communities (Eingereicht bei Aquat. Microb. Ecol.)
Etablierung und Spezifitätstest der Roseobacter spezifischen PCR durch S. S. unter Anleitung
von B. R. und T. B (Diplomarbeit, 2003). Durchführung der spezifischen PCR und DGGE,
der Klonierung und Sequenzierung durch B. R. Statistische Auswertung und Erstellung der
phylogenetischen Stammbäume durch B. R. Erstellung der ersten Fassung des Manuskripts
durch B. R., Überarbeitung durch T. B., B. R. und M. S.
Rink, B., Martens, T., Fischer, D., Lemke, A., Grossart, H. P., Simon, M., und Brinkhoff, T.
(2006) Tidal effects on coastal bacterioplankton (In Vorbereitung zum Einreichen bei
Limnol. Oceanogr.)
Planung und Durchführung der Probenahme 2005 durch B. R. Durchführung der spezifischen
PCR und DGGE sowie der RNA Untersuchungen und CARD-FISH durch B. R. Statistische
Auswertung und Erstellung der phylogenetischen Stammbäume durch B. R. Erstellung der
ersten Fassung des Manuskripts durch B. R., Überarbeitung durch T. B., B. R. und M. S.
Stevens, H., Brinkhoff, T., Rink, B., Vollmers, J., und Simon, M. (2006) Diversity and
abundance of Gram-positive bacteria in a tidal flat ecosystem (Eingereicht bei Environ.
Microbiol.)
Durchführung der spezifischen CARD-FISH und DGGE Untersuchungen von J. V. unter
Anleitung von B. R. und T. B (Leistungsnachweis, 2005). Überarbeitung des Manuskriptes
von B. R., T. B. und M. S.

Rink, B., Brinkhoff, T., Ziegelmüller, K., und Simon, M. (2006) High regional variability of
bacterial communities in the German Bight, North Sea (Eingereicht bei Aquat. Microb.
Ecol.)
Planung und Durchführung der Probenahme 2002 von Mirko Lunau und B. R. Planung und
Durchführung der Probenahme 2003 von B. R. Molekularbiologische Untersuchungen (PCR,
DGGE, Klonierung, Sequenzierung), statistische Auswertung und Erstellung der
phylogenetischen Stammbäume durch B. R. Erstellung der ersten Fassung des Manuskripts
durch B. R., Überarbeitung durch T. B., B. R. und M. S.
Tagungsbeiträge
Rink B, Stevens H, Simon M, Brinkhoff T (2006) Stability of Microbial Communities Within
Different Time Scales in a Tidal Flat Ecosystem. Posterbeitrag, International
Symposium Microbial Ecology (ISME-11), Wien, Österreich, 20-25 August
Rink B, Brinkhoff T, Simon M (2004) Bacterial communities reflect different regional
properties of the German Bight. Vortrag, VAAM-Meeting Braunschweig, 28-31 March
Rink B, Kruse M, Seeberger S, Stevens H, Brinkhoff T, Simon M (2004) Seasonal and spatial
differences in the composition and abundance of bacterial communities in the German
Bight of the North Sea. Posterbeitrag, International Symposium Microbial Ecology
(ISME-10), Cancun, Mexico, 22-27 August
Simon M, Selje N, Schledjewski R, Rink B, Grossart HP (2004) Diversity and substrate
turnover of bacterioplankton communities in the Gulf of Aqaba, Red Sea. Posterbeitrag,
International Symposium Microbial Ecology (ISME-10), Cancun, Mexico, 22-27
August
Rink B, Lunau M, Seeberger S, Stevens H, Brinkhoff T, Grossart H-P, Simon M (2003)
Diversity patterns of aggregate-associated and free-living bacterial communities in the
German Wadden Sea. In Rullkötter J. (ed.), BioGeoChemistry of Tidal Flats -
Proceedings of a Workshop held at the Hanse Institute of Advanced Study, Delmenhorst
(Germany), 14- 17 May. Forschungszentrum Terramare, Wilhelmshaven, Berichte Nr.
12, 96-98. ISSN 1432-797X.

Lunau M, Rink B, Grossart H-P, Simon M (2003) How to sample marine microaggregates in
shallow and turbid environments? - Problems and solutions. In Rullkötter J. (ed.),
BioGeoChemistry of Tidal Flats - Proceedings of a Workshop held at the Hanse
Institute of Advanced Study, Delmenhorst (Germany), 14-17 May. Forschungszentrum
Terramare, Wilhelmshaven, Berichte Nr. 12, 85-88. ISSN 1432-797X.
Rink B, Brinkhoff T, Simon M (2002) Completing the picture of natural habitats: The use of
specific Primersets in DGGE. Posterbeitrag, VAAM Meeting, Berlin, 23-26 March

Zusammenfassung
Im Wattenmeer unterliegen die Organismen hochdynamischen Prozessen. Eine flache
Wassersäule und der Einfluß der Gezeiten sorgen für starke Strömungen und hohe
Resuspensionsraten. Auch der tidale Ein- und Ausstrom von Wassermassen aus der Nordsee
in das Rückseitenwatt beeinflusst das System. Während in Herbst- und Wintermonaten
sedimentologische Faktoren überwiegen, ist im Frühjahr und Sommer ein deutlicher Einfluss
biologischer Größen nachweisbar. Im Rahmen des interdisziplinären Forschungsprojekts
„Biogeochemie des Watts“, in das diese Arbeit eingebunden ist, wurden große Varianzen
innerhalb des Schwebstoffaufkommens sowie in bakterieller Aktivität und Abundanz auf
saisonaler Ebene sowie im Tidenzyklus beschrieben.
In der vorliegenden Arbeit wurde untersucht, inwiefern tidale und saisonale Faktoren die
Struktur der ansässigen Bakteriengemeinschaften in der Wassersäule beeinflussen.
Weiterführend wurde untersucht, ob die im Wattenmeer detektierten Phylotypen
standortspezifisch oder auch in anderen Gebieten der Deutschen Bucht nachweisbar sind.
Im Wattenmeer fand die Beprobung in der Otzumer Balje im Rückseitenwatt von Spiekeroog
statt. Im ersten Teil dieser Arbeit wurden zur Untersuchung des Zusammenhangs von
Bakteriengemeinschaften und Phytoplankton wöchentlich Proben genommen und mittels
gruppenspezifischer DGGE (Denaturierende Gradienten Gelelektrophorese) und statistischer
Methoden untersucht. Im zweiten Teil wurden neben saisonalen auch tidale Vorgänge
beleuchtet. Die Probenahme fand im Herbst, Frühjahr und Sommer in einstündigem und
dreistündigem Probenahmeraster statt. Die Bakteriengemeinschaften wurden mittels
gruppenspezifischer DGGE für alpha-Proteobakterien, Bacteroidetes und Roseobacter
sowohl DNA- als auch RNA basiert untersucht. Zusätzlich wurden FISH (Fluoreszens in situ
Hybridisierung) und die hoch sensitive CARD-FISH (Catalyzed Reporter Deposition-FISH)
eingesetzt und somit erstmalig die Abundanzen einzelner Bakteriengruppen in der
Wassersäule des Wattenmeeres dargestellt.
In einer vorangegangenen Arbeit wurden im Wattenmeer bemerkenswert viele gram-positive
Bakterien isoliert, was zu der Annahme führte, dass diese Bakteriengruppe eine besondere
Stellung in diesem Habitat einnimmt. Zur Vervollständigung der Daten wurde im dritten Teil
dieser Arbeit die CARD-FISH eine Actinobakterien-spezifische Sonde eingesetzt und
zusätzlich eine spezifische DGGE entwickelt, um Abundanz und phylogenetische Vielfalt der
Actinobakterien im Watt zu untersuchen. Die Probenahme hierzu wurde an verschiedenen
Standorten im Spiekerooger Rückseitenwatt durchgeführt.

Im vierten Teil wurden im Sommer 2002 und 2003 verschiedene Standorte der Deutschen
Bucht an der Küstenzone, vor Helgoland und in der offenen Nordsee beprobt. Die
Bakteriengemeinschaften wurden mit spezifischer DGGE für alpha-Proteobakterien und
Bacteroidetes untersucht. Zur weiteren Beschreibung der Ökologie an den untersuchten
Standorten wurden zusätzlich hydrologische, mikrobiologische und partikuläre Parameter
bestimmt.
Zusammenfassend ergaben sich aus diesen Arbeiten folgende Hauptaussagen:
• Im Wattenmeer sind die Bakteriengemeinschaften in der Wassersäule im
Wesentlichen aus alpha- und gamma-Proteobakterien sowie Bacteroidetes
zusammengesetzt. Darüber hinaus sind beta-Proteobakterien abundant auf
Aggregaten. Hierbei bilden frei lebende und Aggregat-assoziierte Bakterien distinkte
Gemeinschaften sowohl im Wattenmeer als auch in der Deutschen Bucht. Die Struktur
der frei lebender Bakteriengemeinschaften besteht hauptsächlich aus wenigen
dominanten Phylotypen der Roseobacter Gruppe. Ihre Zusammensetzung ist saisonal
und räumlich stabil. Die Struktur der Aggregat-assoziierten Bakterien zeigt grössere
Artenvielfalt als bei frei lebenden Bakterien und unterliegt deutlicher räumlich-
zeitlichen Einflussfaktoren. Hier dominieren Phylotypen innerhalb der Bacteroidetes,
gamma- und delta-Proteobakterien.
• Saisonale Einflüsse auf die Bakteriengemeinschaften sind in den produktiven
Frühjahrs- und Sommermonaten erkennbar. Insbesondere Aggregat-assoziierte
Bakterien der Roseobacter-Gruppe und Bacteroidetes unterliegen biologischen
Einflussfaktoren wie Phytoplanktonblüten. Tidale Einflüsse auf bakterielle Aktivität
und Abundanz werden nur geringfügig und nicht systematisch durch Änderungen in
der Zusammensetzung der Bakteriengemeinschaften reflektiert.
• Actinobakterien stellen knapp 5% des Bakterioplanktons im Wattenmeer. Ihre
Abundanz und Zusammensetzung im Süßwasserbereich unterscheidet sich von den
marinen Standorten, wobei frei lebende und Aggregat-assoziierte Actinobakterien
distinkte Gemeinschaften bilden. Aus dem Wattenmeer isolierte Stämme zeigen hohe
Anpassungsfähigkeit anhand breiter Substrat- und Salinitätsspektren.
• Insgesamt wird das organische Material im Wattenmeer von wenigen dominanten
Bakterienarten umgesetzt, die ganzjährig auftreten und hoch angepasst sind. In
produktiven Jahreszeiten treten darüber hinaus weitere, spezialisierte Bakterienarten
auf, die in kurzen Zeitskalen von Änderungen der Zusammensetzung des organischen
Materials, z. B. durch absterbendes Phytoplanton, profitieren.

Summary
In the German Wadden Sea, organisms are influenced by highly dynamic processes. A
shallow water column and tidal impact cause strong currents and high resuspension rates. The
introduction of North Sea water masses also influences the Wadden Sea System. While
sedimentological factors prevail in autumn and winter months, biological processes dominate
in spring and summer. Within the research group “Biogeochemistry of tidal flats”, in which
this thesis is included, tidal and seasonal variations of suspended matter appearance and
bacterial activity and abundance were described.
Hence, the focus of this thesis was to investigate the extend of tidal and seasonal impacts on
the structure of resident bacterial communities in the water column. Furthermore, we
determined if phylotypes detected in the German Wadden Sea are site-specific or detectable at
other locations in the German Bight as well.
Sampling was performed in the backbarrier tidal flat system of Spiekeroog in the German
Wadden Sea. In the first part of this work, samples were taken weekly to investigate
correlations of the bacterial communities and phytoplankton by group-specific DGGE
(Denaturing gradient gel electrophoresis) and statistical methods. In the second part, in
addition to seasonal also tidal processes were focussed. Sampling was performed in autumn,
spring and summer hourly and in three hour intervals. The bacterial communities were
investigated by group-specific DGGE (Denaturing gradient gel electrophoresis) for alpha-
Proteobacteria, Bacteroidetes and the Roseobacter group. In addition, FISH (Fluorescense in
situ hybridization) and the highly sensitive CARD-FISH (Catalyzed reporter deposition-
FISH) were applied to determine abundances of individual bacterial groups in the water
column of the German Wadden Sea.
In a former study, remarkably high numbers of different gram-positive Bacteria were isolated
which led to the assumption that this bacterial group exhibits an exceptional position in this
habitat. To complete these data, CARD-FISH with Actinobacteria-specific probes was
applied and a specific DGGE was established to determine abundances and phylogenetic
variety of Actinobacteria in the Wadden Sea. Samples were taken at different sites in the
backbarrier tidal flat system of Spiekeroog.
In the last part of this work, different locations at the coastal line, near Helgoland and offshore
were investigated in the German Bight in summer 2002 and 2003. The bacterial communities
were analysed by specific DGGE for alpha-Proteobacteria and Bacteroidetes. To describe the
ecology of the sampling sites hydrological, microbiological and particulate parameters were
determined additionally.

The major findings of this thesis can be summarized as follows:
• The bacterial communities in the water column of the German Wadden Sea are mainly
composed of alpha- and gamma-Proteobacteria and Bacteroidetes. In addition, beta-
Proteobacteria are abundant on aggregates. Free-living and aggregate-associated
bacteria form distinct communities in the German Wadden Sea and in the German
Bight as well. The structure of free-living bacterial communities is mainly composed
of few dominant phylotypes affiliated to the Roseobacter group. Their composition is
stable on seasonal and spatial scales. The structure of aggregate-associated bacteria
shows higher richness compared to free-living bacteria and is influenced by spatial-
temporal impacts to a greater extend. Aggregate-associated bacteria are dominated by
bacteria affiliated to the Bacteroidetes phylum, gamma- and delta-Proteobacteria.
• Seasonal influences on the bacterial communities are detectable in the highly
productive spring and summer months. Especially aggregate-associated Roseobacter
and the Bacteroidetes follow biological impacts e. g. phytoplankton blooms. Tidal
influences on bacterial activities and abundances are only marginally and not
systematically reflected by changes of the bacterial community composition.
• Actinobacteria represent about 5% of the Wadden Sea bacterioplankton. Their
abundance and composition differs between the fresh water and marine sites, and free-
living and aggregate-associated bacteria form distinct communities. Strains isolated
from the Wadden Sea show high adaptation qualities on the basis of broad substrate
and salinity ranges.
• The organic matter in the Wadden Sea is mediated by few dominant bacterial species
which are present throughout the year and are highly adapted. In productive seasons,
specialised bacteria appear additionally which benefit from the changes of the organic
matter composition, e. g. decaying phytoplankton, on small time-scales.

Inhaltsverzeichnis
Zusammenfassung
Summary
I. Einleitung 1
I.1 Kleine Lebewesen, große Wirkung – Marine heterotrophe Bakterien im
globalen Stoffkreislauf 2
I.2 Who´s who – Die Zusammensetzung der Bakteriengemeinschaften 5
I. 3 Geographie und Ökologie der Untersuchungsgebiete 8
I.3.1 Die Nordsee und die Deutsche Bucht 8
I.3.2 Das Wattenmeer 11
I.4 Zielsetzungen der Arbeit 15
I.5 Literatur 16
II. Effects of a phytoplankton bloom in a coastal ecosystem on the composition
of bacterial communities 20
Abstract 22
Introduction 23
Materials and Methods 24
Results 28
Discussion 32
Literature cited 36
III. Tidal effects on coastal bacterioplankton 48
Abstract 51
Introduction 52
Materials and Methods 54
Results 56
Discussion 60
References 66

IV. Diversity and abundance of Gram-positive bacteria in a tidal flat ecosystem 81
Abstract 83
Introduction 84
Results 85
Discussion 88
Experimental procedures 93
References 98
V. High regional variability of bacterial communities in the German Bight,
North Sea 114
Abstract 116
Introduction 117
Materials and Methods 118
Results 121
Discussion 124
References 130
VI. Schlussbetrachtung und Ausblick 144
Danksagung
Kurzbiographie

Abkürzungsverzeichnis
CARD-FISH catalyzed reporter deposition-FISH
Chl a Chlorophyll a
CO2 Kohlendioxid
DAPI 4´,6´-Diamidino-2-phenylindol hydrochlorid
DGGE Denaturierende Gradienten Gelelektrophorese
DNA desoxy ribonucleic acid
DOC dissolved organic carbon
DOM dissolved organic matter
DSM, DSMZ Deutsche Sammlung von Mikroorganismen und Zellkulturen
et al. et alii
FISH fluorescence in situ hybridization
FL free living
HT high tide
LT low tide
ml Milliliter
MT mean tide
NCBI National Center for Biotechnology Information
n. a. not available
n. d. not determined
PA particle attached
PCR polymerase chain reaction
PIC particulate inorganic carbon
POC particulate organic carbon
psu practical salinity unit
rRNA ribosomal ribonucleic acid
SPM suspended particulate matter

1
I.
Allgemeine Einleitung

Kapitel I Einleitung
2
I.1 Kleine Lebewesen, große Wirkung – Marine heterotrophe Bakterien im globalen
Stoffkreislauf
Bei der Betrachtung der Gesamtgröße der Weltmeere erscheint es zunächst kaum vorstellbar,
dass mikroskopisch kleine Lebewesen den Großteil des Umsatzes organischen Materials im
Wasser bewirken sollen. Berücksichtigt man allerdings, dass in einem tausendstel Liter
bereits durchschnittlich 1-3 Mio. Bakterien vorhanden sind, ist offenbar, warum sich die
Forschung seit mehr als zwei Jahrzehnten bemüht, diese höchst bemerkenswerten Lebewesen
besser kennen zu lernen. Bakterieller Abbau und Remineralisierung wirken sich auf den
Stoffkreislauf aller Elemente aus (Schlegel, 1992; Madigan et al., 2003), wobei der
Kohlenstoffkreislauf große Bedeutung nicht zuletzt für klimatische Veränderungen besitzt. So
hat die Aktivität mariner phototropher und heterotropher Bakterien sowohl durch die
Fixierung als auch durch den Ausstoß von CO2 Einfluss auf das Weltklima (Smith und
Hollibaugh, 1993; Wollast, 1993; Falkowski et al., 1998), so dass der marinen mikrobiellen
Ökologie im Zuge der globalen Erwärmung immer größere Bedeutung beigemessen wird.
Auch das genetische Potential der marinen Bakterien, das durch moderne Methoden zwar
detektiert, aber bei weitem noch nicht entschlüsselt wurde, gibt der Wissenschaft Rätsel auf
(Venter et al., 2004). Die zukunftsträchtige und viel versprechende Vision einer möglichen
medizinischen oder biotechnologischen Nutzung mariner Mikroorganismen bietet daher ein
weiteres großes Interessensgebiet der ökologischen Forschung.
Die Nahrungsquelle heterotropher Bakterien, organischer Kohlenstoff, liegt in der
Wassersäule in gelöster (dissolved organic carbon, DOC) oder in partikulär gebundener Form
(particulate organic carbon, POC) vor.
DOC umfasst bis zu 95% des Gesamtkohlenstoffs der Weltmeere (Hedges, 1992) und wird
nach Zusammensetzung und bakterieller Verfügbarkeit in eine labile und eine refraktäre
Fraktion unterteilt (Søndergaard und Middelboe, 1995). Die Entstehung von gelöstem
organischen Material (dissolved organic matter, DOM) ist durch einen komplexen Kreislauf
gekennzeichnet, in dem im Wesentlichen Phytoplankton, Bakterien und Viren eine Rolle
spielen (Abb. 1). Anhand der Darstellung wird deutlich, dass Bakterien DOM direkt
aufnehmen und verwerten und somit die größte Bedeutung für die Umsetzung des DOC
besitzen. Durch das daraus resultierende Wachstum und den Fraß durch Protozoen gelangt der
Kohlenstoff dann indirekt in das Nahrungsnetz höherer Organismen. Die Entstehung des
DOM ist abhängig von verschiedenen Faktoren, da die am Kreislauf beteiligten Organismen

Kapitel I Einleitung
3
räumlich-zeitlichen Gegebenheiten unterliegen. Phytoplankton bildet bei ausreichender
Nährstoffversorgung und günstigen Licht- und Temperaturverhältnissen Blüten aus, die in
direkter Form zu der Absonderung („Leakage“) von DOM führen kann (Bjørnsen, 1988).
Nach Absterben der Blüte werden durch Lysis der Zellen ebenfalls gelöste Stoffe freigesetzt.
Darüber hinaus entsteht bei diesem Vorgang Detritus (partikuläre Zellreste), der durch
Bakterien hydrolysiert und somit dem DOM Pool zugeführt wird.
Neben dem DOC stellt in Aggregaten angereicherter POC eine weitere wichtige organische
Kohlenstoffquelle dar (Alldredge, 1979) und bildet die Grundlage für komplexe Lebens-
gemeinschaften, die sich im Weiteren aus Phytoplankton, Protozoen, Bakterien und Pilzen
zusammensetzen (Alldredge and Silver, 1988). An Aggregate angeheftet, weisen sie,
verglichen mit frei suspendierten Mikroorganismen, wesentlich höhere Zelldichten auf
(Simon et al., 2002). Je nach Ursprung des partikulären Materials variieren der organische
Anteil sowie dessen Zusammensetzung aus Kohlenhydraten und Proteinen, die den Bakterien
als Substrat dienen (Azam und Cho, 1987; Smith et al., 1995, Azam und Cho, 1987; Biddanda
und Benner, 1997). Auch hier ist das Phytoplankton, abhängig von seiner
Artenzusammensetzung, hauptsächlicher Nährstofflieferant (Smith et al., 1995); weitere
Bestandteile von Aggregaten sind hochrefraktäres oder auch anorganisches Material, z. B.
resuspendiertes Sediment (Eisma, 1993). Die Freisetzung der Nährstoffe durch Bakterien
erfolgt durch die Ausscheidung hydrolytischer Ektoenzyme, die partikulär gebundene
Abb. 1: Kreislauf gelösten organischen Kohlenstoffs in der Wassersäule (dissolved organic matter, DOM; modifiziert nach Riemann, 2001)

Kapitel I Einleitung
4
Makromoleküle in Oligo- und Monomere spalten (Madigan, 2003). Die hydrolysierten Stoffe
werden teils von den Aggregat-assoziierten Bakterien selbst verwertet, teils diffundieren sie
jedoch auch in das Umgebungswasser und stehen somit den frei suspendierten Bakterien und
anderen planktischen Organismen zur Verfügung (Smith et al., 1992). Durch die
Substrataufnahme wachsen die Bakterien und bilden somit Biomasse, die wiederum
Zooplankton als Nahrungsquelle dient. Durch diesen Stoffkreislauf, der als microbial loop
bezeichnet wird, werden Nährstoffe aus abgestorbenen Tier- und Pflanzenresten (Detritus) für
höhere Trophiestufen wieder verfügbar (Azam et al., 1983).
In flachen Küstenzonen, Wattsystemen und Ästuaren unterscheidet sich die Situation im
Vergleich zu den offenen Ozeanen durch ein sehr hohes Schwebstoffaufkommen. Die
Schwebstoffe werden hier durch Flüsse oder von den angrenzenden Landgebieten eingetragen
und sind zum Teil hohen Scherkräften ausgesetzt, die durch die flache Wassersäule und
Tidenhub entstehen. Dadurch sind die Aggregate in den Küstenzonen wesentlich kleiner und
häufiger (Lunau et al., 2006) und besitzen, verglichen mit Schwebstoffen in küstenfernen
Gebieten, einen geringeren organischen Anteil (Postma, 1981; Lunau et al., 2006). Durch
ständige Turbulenz werden die Aggregate fortwährend resuspendiert und somit in Schwebe
gehalten, was ebenfalls Auswirkungen auf die angehefteten Bakterien hat. So weisen
Aggregat-assoziierte Bakterien in schwebstoffreichen Gewässern wesentlich höhere
Enzymaktivitäten und Biomasse auf, und können sogar bis zu 95% der Gesamtaktivität der
suspendierten Bakterien ausmachen (Crump et al., 1998; Crump & Baross, 2000).
Auch die Bakteriengemeinschaften können in Küstenzonen anders zusammengesetzt sein als
in zulaufenden Flüssen oder im offenen Meer. So bilden sich entweder Mischformen von
Süßwasser- und marinen Bakteriengemeinschaften (Rappé et al., 2000), oder auch distinkte
Bakteriengemeinschaften von Süßwasser, Brackwasser und marinem Milieu aus (Selje et al.,
2003, Crump et al. 1999).
Diese Zusammenhänge verdeutlichen, in welchem Umfang Bakterien das gesamte
Nahrungsnetz beeinflussen und dass freilebende und Aggregat-assoziierte Bakterien
vollkommen unterschiedliche Lebensbedingungen vorfinden. Daher ist eine differenzierte
Untersuchung beider Lebensgemeinschaften essentiell, um die ökologischen Zusammenhänge
in der Wassersäule verstehen zu können.

Kapitel I Einleitung
5
I.2 Who´s who – Die Zusammensetzung der Bakteriengemeinschaften
Da Bakterien unter dem Mikroskop und in der Kultivierung nur sehr wenige Unterschiede
anhand von Zellmorphologie und Wachstum aufweisen, wurde die Artenvielfalt von
natürlichen Bakteriengemeinschaften lange Zeit unterschätzt. Darüber hinaus bot die
Kultivierung nur bedingt Einblick in das Vorkommen und die Häufigkeit von Bakterienarten,
da die Bedingungen, die Bakterien im Labor vorfinden, nicht den natürlichen Gegebenheiten
entsprachen. Einige Bakterienstämme oder auch phylogenetische Gruppen konnten leicht
unter künstlichen Bedingungen angereichert werden und wurden somit auch häufiger in
verschiedenen Habitaten nachgewiesen, während sich andere Bakterien nur unter bestimmten
Voraussetzungen kultivieren ließen oder bis heute unkultiviert bleiben. Daher ergaben sich
große Unterschiede zwischen mikroskopisch und durch Kultivierungsansätze ermittelte
Zellzahlen („great plate count anomaly“, Staley & Konopka, 1985).
So brachte die Einführung molekularbiologischer Methoden, die auf dem Vergleich des
Erbguts anhand der ribosomalen RNA beruhten, neue Einblicke in die mikrobielle Ökologie
und die phylogenetischen Zusammenhänge (Woese et al., 1987). Bis heute stellt die
hochkonservierte 16S rRNA bzw. der 16S rRNA Genabschnitt eine wesentliche Grundlage
für die Untersuchung von Bakteriengemeinschaften dar. Die Vervielfältigung und
Sequenzierung von Genen ermöglichte es, Bakteriengenome und Phylogenie unabhängig von
Kultivierungserfolgen zu erforschen (Saiki et al, 1988; Sanger et al., 1977). Gängige
Methoden zur Detektion sind z.B. die Denaturierende Gradienten Gelelektrophorese (DGGE;
Muyzer et al., 1993), Restriktionsfragment Längen-Polymorphismus (RFLP; Marsh, 1999),
oder die rDNA Intergenic Spacer Analysis (RISA). Die Quantifizierung von
Bakteriengruppen oder auch –arten kann durch Fluoreszenz in situ Hybridisierung (FISH;
Giovannoni et al, 1988; Amann et al., 1990) bzw. Catalyzed Reporter Deposition-FISH
(CARD-FISH; Pernthaler et al., 2002) sowie mittels Realtime PCR (Heid et al., 1996)
erfolgen. Heute werden Kultivierungsansätze und kultivierungsunabhängige Methoden sowie
Aktivitätsmessungen kombiniert, um möglichst viele Informationen über die Mikrobiologie
eines Habitats zu gewinnen.
Durch den Einsatz dieser Methoden konnte die Struktur der am Stoffumsatz beteiligten
Bakterien, die vorher als „Black Box“ betrachtet wurden, weiter aufgeklärt werden
(Giovannoni & Rappé 2000). So stellte sich heraus, dass insbesondere die gram-negativen
Proteobakterien sowie Bacteroidetes bedeutende Gruppen innerhalb des marinen

Kapitel I Einleitung
6
heterotrophen Bakterioplanktons bilden. Darüber hinaus wurden u.a. methylotrophe
Bakterien, Planctomycetales und die gram-positiven Actinobakterien in marinen Habitaten
nachgewiesen.
Innerhalb der Proteobakterien wurden die gamma-Proteobakterien lange Zeit als die
dominanteste Gruppe des marinen Bakterioplanktons angenommen, da sich Vertreter dieser
Gruppe leicht unter Laborbedingungen isolieren ließen. Über kultivierungsunabhängige
Methoden fand man jedoch heraus, dass die meisten weltweit nachgewiesenen Phylotypen
distinkte Cluster bildeten, die wiederum keine Isolate beinhalteten (Giovannoni und Rappé,
2000). Mittlerweile konnten teilweise auch für diese Cluster mit gezielten
Anreicherungsversuchen einzelne Isolate gewonnen werden (Cho und Giovannoni, 2004), so
dass die Erforschung der ökologischen Funktion dieser Organismen weiter voranschreiten
kann. Physiologisch betrachtet sind gamma-Proteobakterien fakultativ anaerobe und
chemoheterotrophe Organismen, die häufig Oberflächen-assoziiert vorkommen und somit im
Sediment sowie auf Aggregaten eine zentrale Rolle einnehmen.
Die alpha-Proteobakterien sind ebenfalls weltweit verbreitet und zumeist durch die
Subgruppen Sphingomonas und Roseobacter vertreten. Weitere große Bedeutung besitzen die
hoch spezialisierten Cluster SAR 11 (Rappé et al. 2002) und SAR 116 innerhalb der alpha-
Proteobakterien. Die chemoorganotrophen Roseobacter wurden bisher ausschließlich im
marinen Milieu nachgewiesen und stellen dort habitatabhängig bis zu 50% der gesamten
alpha-Proteobakterien. Einige Vertreter gehören zu den aeroben anoxygenen phototrophen
Bakterien und sind somit auch in der Lage, Photosynthese zu betreiben. Aktuell werden große
Forschungsprojekte zur Genomentschlüsselung dieser Organismen1 durchgeführt, die das
Potential dieser hoch interessanten und vielfältigen Gruppe weiter aufklären sollen.
Die aeroben oder fakultativ anaeroben, chemoorganotrophen Bacteroidetes bilden die dritte
große Gruppe innerhalb des marinen Bakterioplankton. Sie sind hoch divers und leben in der
Wassersäule sowohl frei suspendiert als auch Aggregat-assoziiert. Ihre
Stoffwechselphysiologie ist äußerst vielfältig, doch es hat sich gezeigt, dass besonders schwer
abbaubare, hochmolekulare Substanzen bevorzugt von Bacteroidetes abgebaut werden
können, z. B. Chitin oder Cellulose (Cottrell und Kirchman, 2000). Darüber hinaus sind sie
häufig beweglich und können auf Oberflächen gleiten, so dass durch diese Eigenschaften
1 Auch andere Bakteriengruppen, die in marinen Habitaten von Bedeutung sind, werden derzeit durch große Genomprojekte erforscht (z. B. Moran et al., 2004). Die Untersuchung des genetischen Potentials von Organismen führt neben der Entschlüsselung bisher unbekannter Gene auch zur Entdeckung neuer Stoffwechselwege oder biotechnologisch nutzbarer Substanzen (Fusetani, 2000). Man kann daher annehmen, dass die Ozeane ein riesiges Potential bisher unentdeckter Ressourcen bietet, deren Erforschung im Zuge interdisziplinärer Projekte immer mehr in den Vordergrund tritt.

Kapitel I Einleitung
7
angenommen wurde, dass sie besonders auf Aggregaten eine große Bedeutung für
Stoffumsatzprozesse einnehmen.
Diese Zusammenhänge zeigen, dass das Verständnis über die Vorgänge der
Remineralisierung durch die Strukturaufklärung der beteiligten Bakteriengemeinschaften
wesentlich verbessert werden konnte. Darüber hinaus können lokale Gegebenheiten einzelner
Habitate jedoch übergeordnet Einfluss auf die Zusammensetzung der
Bakteriengemeinschaften und somit auch auf die Effektivität des Stoffumsatzes nehmen.

Kapitel I Einleitung
8
I. 3 Geographie und Ökologie der Untersuchungsgebiete
I.3.1 Die Nordsee und die Deutsche Bucht
Die Nordsee liegt auf dem europäischen Kontinentalschelf und wird begrenzt von den
Britischen Inseln und dem Europäischen Kontinent (Abb. 2). Sie ist mit einer
durchschnittlichen Tiefe von 93 m ein flaches Schelfmeer und durch verschiedene
angrenzende Land- und Wasserregionen beeinflusst. Salines Atlantikwasser dringt im Norden
zwischen der schottischen und der norwegischen Küste sowie südlich durch den Ärmelkanal
in die Nordsee. Der größte Eintrag von Süßwasser erfolgt über den Skagerrak aus der Ostsee
und durch verschiedene große Flüsse, die in die Nordsee münden (z. B. von deutscher Seite
die Flüsse Rhein, Ems, Weser, Elbe und Eider). Der mittlere Salzgehalt ist demnach mit
durchschnittlich 15 – 25 Promille an den Küstengebieten geringer als in der offenen Nordsee,
wo durchschnittlich 32 – 35 Promille vorherrschen (Alongi, 1997). Aufgrund der
Amphidromie in der südlichen Nordsee (Defant, 1923) fließt das Wasser in der deutschen
Bucht entgegen dem Uhrzeigersinn, wodurch auch der Transport von partikulären und
gelösten Stoffen sowie von planktischen Organismen beeinflusst wird.
Die deutsche Bucht, der südliche Teil der Nordsee, reicht von Jütland in Dänemark über die
Friesischen Inseln (Nord-, Ost-, und Westfriesische Inseln) bis zur niederländischen Grenze
im Westen. Im Nordwesten wird sie begrenzt von der Doggerbank, einer flachen Sandbank
Abb. 2: Geographische Lage der Nordsee und der Deutschen Bucht sowie des Nationalparks Deutsches Wattenmeer von der ostfriesischen zur nordfriesischen Küste (modifiziert nach http://www.bsh.de)

Kapitel I Einleitung
9
innerhalb der Nordsee, die durch große Fischvorkommen insbesondere für die Fischerei eine
wesentliche Rolle spielt. Das Küstengebiet der Deutschen Bucht bildet das Deutsche
Wattenmeer, eine einzigartige Flachwasserzone, die sich hinter den Friesischen Inseln
erstreckt (vgl. I.3.2).
Grundlage des Nahrungsnetzes in der Nordsee bilden einzellige Algen, das Phytoplankton,
welches im Jahresverlauf mehrere Blüten ausbildet (Alongi, 1997). Im Frühjahr (März bis
April) führen steigende Temperatur- und Lichteinstrahlung sowie hohe
Nährstoffkonzentrationen dazu, dass sich eine Blüte aus Kieselalgen (Diatomeen,
Bacillariophyceae) ausbildet (Drebes, 1974). Durch Nährstofflimitierung und sukzessiven
Fraßdruck durch Zooplankton endet die Blüte zumeist im Juni, bis im Spätsommer eine
zweite, meist weniger intensive Phytoplanktonblüte entsteht (Alongi, 1997). Während der
Blüte scheiden die Diatomeen gelösten organischen Kohlenstoff aus, der von Bakterien
genutzt werden kann (Ittekott et al. 1981). Auch nach dem Zusammenbruch einer
Phytoplanktonblüte profitieren heterotrophe Bakterien vom nährstoffhaltigen Lysat. Dies
kann sich in vermehrter bakterieller Aktivität, Abundanz und Veränderungen in der
Artenzusammensetzung ausdrücken (Reinthaler et al., 2005; Smith et al, 1995, Riemann et al.,
2000, Fandino et al., 2001).
Seit Mitte der 1950er Jahre wurde in der Nordsee ein stetiger Anstieg von Nährstoffen
bedingt durch anthropogene Einflüsse gemessen, der dazu geführt hat, dass die Nordsee stark
eutrophiert ist (Alongi, 1997). Dies führte zu Verschiebungen sowohl in der
Artenzusammensetzung als auch in der Biomasse des Phytoplanktons und resultierte in der
Einschränkung der gesamten Artenvielfalt in der Nordsee. Da die Nährstoffe zumeist über die
Zuflüsse in die Nordsee eingetragen werden, sind erhöhte Konzentrationen von Phosphat,
Nitrat oder auch von Schwermetallen als Gradient von der Küste in die offenen Gewässer zu
beobachten. Darüber hinaus ist durch die globale Erwärmung auch eine Erwärmung der
Wassersäule in der Deutschen Bucht um durchschnittlich 1,1°C beobachtet worden (Wiltshire
and Manly, 2004). Die damit verbundene Verschiebung der Algenblüten könnte durch eine
Veränderung der temperaturabhängigen Rahmenbedingungen entstanden sein. Der Zustand
der Nordsee wird daher seit Jahrzehnten durch verschiedene Institutionen in Monitoringserien
untersucht, um die Nutzung und Belastung der Gewässer zu überwachen und den Lebensraum
zu schützen (Bundesamt für Seeschiffahrt und Hydrographie, BSH; http://www.bsh.de).

Kapitel I Einleitung
10
Untersuchungen zu den Bakteriengemeinschaften in der Nordsee liegen von verschiedenen
Autoren vor. So untersuchten Eilers et al. (2001) die Kultivierbarkeit von Nordseebakterien
bei Helgoland. Es konnten hauptsächlich alpha- und gamma-Proteobakterien sowie
Bacteroidetes isoliert werden. FISH-Zählungen mit spezifischen Sonden ergaben, dass diese
Gruppen ebenfalls einen großen Teil der Bakteriengemeinschaften in der Nordsee darstellen.
Große saisonale Unterschiede in der Hybridisierbarkeit waren erkennbar, die zeigten, dass in
den biologisch hoch produktiven Sommermonaten wesentlich höhere Effizienz erreicht wurde
als in den Wintermonaten. Gerdts et al. (2004) gaben eine Übersicht der Aktivität, Abundanz
und saisonale Veränderungen bakterieller Gemeinschaften, die mit verschiedenen Methoden
als Langzeitmonitoring bei Helgoland durchgeführt wurden. Besonders in den produktiven
Sommermonaten ergaben sich deutliche Änderungen in der Aktivität und Zusammensetzung.
In der südlichen Nordsee wurden saisonal bakterielle Respiration, Artenreichtum
(„Richness“) und Biomasseproduktion entlang von Transekten (Reinthaler et al., 2005) sowie
in Abhängigkeit von Phytoplanktonblüten untersucht (Reinthaler & Herndl., 2005). Es zeigte
sich, dass die bakterielle Biomasseproduktion saisonal stark variiert und korreliert ist mit der
Primärproduktion.
Diese Zusammenhänge verdeutlichen die Notwendigkeit, durch weitere Erforschung der
relevanten Bakterienarten in der Nordsee Schlüsselorganismen zu erkennen und zu
beschreiben. Anhand solcher Indikatororganismen könnten sowohl Änderungen der
ökologischen Gegebenheiten sowie detaillierte Aussagen über Stoffumsatz und äußere
Einflüsse möglich sein. Die Erforschung der Wechselwirkungen zwischen physiko-
chemischen und biologischen Kräften stellt eine essentielle Brücke dar zum Verständnis der
Umwelt und der Bedeutung für das gesamte Ökosystem.

Kapitel I Einleitung
11
I.3.2 Das Wattenmeer
Die südöstliche Nordseeküste besteht aus einem besonderen ökologischen Lebensraum, dem
europäischen Wattenmeer (Abb. 2). Es bildet mit einer Gesamtfläche von ca. 7500 m2 und
einer Gesamtlänge von 500 km die größte zusammenhängende Wattfläche der Welt und reicht
von Den Helder (Niederlande) bis Esbjerg (Dänemark). Es ist gekennzeichnet durch eine hohe
Artenvielfalt und wurde daher 1985 zum Nationalpark erklärt. Der deutsche Teil des
Wattenmeeres wird unterteilt in das Niedersächsische-, das Hamburgische- und das
Schleswig-Holsteinische Wattenmeer. Die Friesischen Inseln sind dem Wattenmeer in
Richtung Nordsee vorgelagert und bilden so eine natürliche Begrenzung.
Als Lebensraum ist das Wattenmeer stark durch die Gezeiten geprägt, wodurch große Teile
des Watts in regelmäßigem Abstand trocken fallen und daher extreme Lebensbedingungen
bieten. Nach Lozan et al. (1994) kann das Watt in vier Ablagerungsbereiche unterteilt werden:
a) Das Sublitoral: Ständig von Salzwasser bedeckte Flächen, z. B. Seegat, Wattrinnen
b) Das Eulitoral: Bereiche, die bei Hochwasser überflutet sind und bei Niedrigwasser
trockenfallen, z. B. Wattflächen zwischen Inseln und Festland
c) Das Supralitoral: Nur bei hochauflaufender Flut von Salzwasser bedeckt, z. B.
Salzmarschen der Inseln und des Festlands
d) Die Dünen: Keine Überspülung mit Salzwasser
Die im Watt lebenden Organismen müssen daher eine hohe Anpassungsfähigkeit besitzen, da
durch die zeitweise Exponierung der Wattfläche und eine insgesamt flache Wassersäule starke
Temperaturschwankungen entstehen. Auch Salinitätsschwankungen sind sehr ausgeprägt,
insbesondere bei starken Regenfällen und an Flußmündungen. Der Austausch der
Wasserkörper zwischen der Nordsee und dem Wattenmeer geschieht über die Seegatten,
Durchlässe zwischen den Inseln, in denen bei jeder Ebbe und Flut sehr hohe
Strömungsgeschwindigkeiten von bis zu 2 m s-1 erreicht werden.
Durch die Eutrophierung der Nordsee insbesondere an den Küsten (vgl. Abschnitt I.3.1) sind
seit den 70er Jahren diverse Projekte zur Beobachtung der Stoffflüsse im Wattenmeer
durchgeführt worden (Baretta and Ruardij, 1988; Cadée, 1984; de Wilde und Beukema,
1984). Um die komplexen Zusammenhänge zwischen der Hydrographie, der Biologie und

Kapitel I Einleitung
12
anthropogenen Einflüssen im Wattenmeer zu studieren, wurden darüber hinaus umfassende
interdisziplinäre Forschungsprojekte ins Leben gerufen. Diese sind zusammenfassend für das
nordfriesische Wattenmeer von Gätje und Reise (1998) sowie für das Spiekerooger
Rückseitenwatt von Dittmann (1999) veröffentlicht worden und bilden die Grundlage für das
Verständnis der Wattenmeerökologie. In beiden Werken wurden Entstehung, Geologie,
Nährstoffkonzentrationen und Stoffflüsse sowie Flora und Fauna untersucht. Eine wesentliche
Rolle nahmen Phyto- und Zooplankton ein; das Bakterioplankton wurde zwar als relevant
erachtet, erschien jedoch als „Black Box“, da nur die Abundanz des gesamten
Bakterioplanktons gemessen wurde. Die Artenzusammensetzung wurde in diesen Arbeiten
nicht berücksichtigt.
Im Rahmen der DFG-geförderten interdisziplinären Forschergruppe „BioGeoChemie des
Watts“, die 2001 ins Leben gerufen wurde, sind weitere Erkenntnisse über die Vorgänge im
Wattenmeer gewonnen worden. Hierzu wurde ein Messpfahl im Spiekerooger Rückseitenwatt
(Standort Otzumer Balje; http://www.icbm.de/watt) errichtet, über den ein fortwährendes
Monitoring der physiko-chemischen Parameter durchgeführt wird. Zusätzlich wurden
konzertierte Meßkampagnen sowie regelmäßige Beprobungen von Tagesgängen
durchgeführt. Durch diese umfangreichen Datensätze manifestierte sich die Bedeutung
biologischer Vorgänge auch im sediment-dominierten Wattenmeer. Besonders zu
Niedrigwasser während des Tages waren biologische Einflüsse erkennbar, sowie saisonal zu
produktiven Jahreszeiten, in denen Phytoplanktonblüten auftraten (Grossart et al., 2004;
Lunau et al., 2006). So wurden im Gegensatz zu den Herbst- und Wintermonaten
beispielsweise im Mai und Juni erhöhte bakterielle Biomasseproduktion und Abundanz
gemessen. Partikelabundanz und –größe verhielt sich gegenläufig, indem in den
Wintermonaten höhere Abundanzen kleinerer Partikel detektiert wurden, im Frühjahr und
Sommer jedoch größere Aggregate in kleinerer Anzahl. Es ist daher anzunehmen, dass der
Einfluß von Phyto- und Bakterioplankton auf Aggregation und Disaggregation durch
Ausscheidung klebriger Substanzen (TEP, transparente Exopolymere; EPS,
Exopolysaccharide; Passow, 2002; Bhaskar et al., 2005) auch in Wattsystemen von
elementarer Bedeutung ist.
Auf tidaler Ebene konnten regelmäßig wiederkehrende Signaturen des suspendierten
partikulären Materials (SPM) nachgewiesen werden, die mit weiteren partikulären Parametern
wie partikulärem organischem Kohlenstoff (POC) und Chlorophyll a korrelierten. Die
Abundanz partikel-assoziierter Bakterien verhielt sich in der Gesamtprobe weitestgehend

Kapitel I Einleitung
13
konstant, wie auch schon in anderen Arbeiten gezeigt (Stevens et al., 2005). Die Auftrennung
in eine absinkende und eine frei schwebende Fraktion zeigte jedoch deutliche Unterschiede in
der Besiedelung (Lunau et al., 2004). Diese Hinweise deuten darauf hin, dass sich die
distinkten Bakteriengruppen unterschiedlich verhalten und Einfluss nehmen.
Erste Untersuchungen zur Zusammensetzung der bakteriellen Gemeinschaft im Watt wurden
1998 durch Llobet-Brossa und Kollegen im Wattenmeersediment durchgeführt. Mittels
Fluoreszenz In Situ Hybridisierung (FISH) wurden sulfatreduzierende Bakterien sowie
Bacteroidetes im Jadebusen nachgewiesen (Llobet-Brossa et al. 1998, 2002). Im Rahmen der
Forschergruppe „BioGeoChemie des Watts“ wurden umfangreiche Untersuchungen auch in
tieferen Sedimentschichten an mehreren Probenahmeorten im Spiekerooger Rückseitenwatt
durchgeführt (Mußmann et al., 2005; Köpke et al., 2005; Willms et al., 2006).
Kultivierungsansätze ergaben eine hohe Artenvielfalt, und die Isolate konnten den
phylogenetischen Gruppen der Proteobakterien, Bacteroidetes, Fusobakterien,
Actinobakterien und Firmicutes zugeordnet werden. Die molekularbiologischen
Untersuchungen ergaben ein ähnliches Spektrum innerhalb der phylogenetischen Gruppen
und die zusätzliche Detektion methanogener Archaeen. Beide Untersuchungen ergaben
Zusammenhänge sowohl zu Aktivitätsmessungen als auch zu sedimentologischen Parametern
und geben somit deutliche Hinweise auf eine ökologische Bedeutung der nachgewiesenen
Stämme und Phylotypen.
Auch in der Wassersäule des Spiekerooger Rückseitenwatts wurden umfassende
Untersuchungen über Bakteriengemeinschaften von Stevens et al. (2005a, b) durchgeführt. In
den Jahren 1999 bis 2000 wurden monatlich Proben genommen, in denen mittels DGGE
nachgewiesen werden konnte, dass hier distinkte Bakteriengruppen existieren: Auf dem
Wattsediment, auf Schwebstoffen, und frei lebend in der Wassersäule. In jedem dieser
Kompartimente waren Phylotypen nachgewiesen worden, die sich ausschließlich in diesem
Lebensraum befanden, sowie Schnittmengen zwischen den einzelnen Gruppen. Vor allem auf
Schwebstoffen bildeten die nachgewiesenen Phylotypen eine Mixtur aus Sediment- und
freilebenden Bakterien. Im Allgemeinen wurden Phylotypen verschiedener Proteobakterien
(alpha-, beta-, gamma- und delta-Proteobakterien), Bacteroidetes und Gram-positive
Bakterien gefunden. Saisonale Veränderungen wurden vorwiegend in den Sommermonaten
während oder nach Phytoplanktonblüten beobachtet. Ein parallel durchgeführter umfassender

Kapitel I Einleitung
14
Kultivierungsansatz ergab nur wenige Übereinstimmungen zu den molekularbiologischen
Ergebnissen.
Die monatliche Probenahme in der Wassersäule war somit geeignet, einen ersten Einblick in
den mikrobiologischen Lebensraum Wattenmeer zu geben, gab jedoch keinen Aufschluss
über die Zeitskala, in der sich Veränderungen der bakteriellen Zusammensetzung ereigneten.
Ebenfalls waren bestimmte Bakteriengruppen unterrepräsentiert, von denen sich in
Kultivierungsansätzen (z. B. Gram-positive Bakterien) sowie anhand von FISH-Zellzahlen
(Bacteroidetes) gezeigt hat, dass sie einen großen Anteil der Bakteriengemeinschaft im
Wattenmeer bilden.
Die weitere Aufklärung der Zusammensetzung der Bakteriengemeinschaften auf Ebene
einzelner, als häufig erkannter phylogenetischer Gruppen ist daher dringend erforderlich.
Ebenso stellt sich die Frage, wie die Bakteriengemeinschaften in kleineren Zeitskalen
beeinflusst werden und inwiefern sich die oszillierenden SPM-Signaturen im Tidenzyklus auf
einzelne Bakteriengruppen auswirken. Es ergaben sich daher folgende Zielsetzungen für die
vorliegende Arbeit:

Kapitel I Einleitung
15
I.4 Zielsetzungen der Arbeit
Ziel der Arbeit war die detaillierte Analyse der frei lebenden und aggregat-assoziierten
Bakteriengemeinschaften im Wattenmeer und der Deutschen Bucht mittels DGGE und FISH
anhand von 16S rRNA und 16S rRNA Genabschnitten. Um auch unterrepräsentierte
Bakteriengruppen erfassen zu können, wurden in allen Arbeiten nicht nur Bacteria-, sondern
auch gruppenspezifische Oligonukleotide (PCR-Primer) verwendet.
Ein Teilaspekt dieser Zielsetzung war die Untersuchung der Zusammenhänge zwischen
Phytoplanktonblüten und Veränderungen in den Bakteriengemeinschaften beider
Kompartimente im ostfriesischen Wattenmeer mit einem engen Probenahmeraster (Kapitel
II).
Einen weiteren Aspekt stellte die Untersuchung tidaler Einflüsse auf die
Bakteriengemeinschaften in Abhängigkeit von saisonalen Aspekten dar (Kapitel III).
Aufgrund der extremen Verhältnisse, die im Wattenmeer herrschen (vgl. Abschnitt I.3.2), war
die Anwendung hoch sensitiver Methoden erforderlich, die zunächst etabliert und getestet
werden mussten. So wurde die DGGE nicht nur DNA, sondern auch RNA basiert
durchgeführt; neben der FISH wurde für einen Vergleich ebenfalls die CARD-FISH Methode
angewendet.
Anhaltspunkte über außergewöhnlich hohe Abundanzen von Gram-positiven Bakterien im
Wattenmeer (Stevens, 2004) erforderten molekularbiologische Untersuchungen, um die
Relevanz dieser Aussage zu untermauern (Kapitel IV). Mit Hilfe einer spezifischen DGGE
sowie CARD-FISH Untersuchungen konnten zusätzliche Hinweise über Zusammensetzung
und Abundanz von Gram-positiven Bakterien an verschiedenen Standorten im Wattenmeer
nachgewiesen werden.
Eine weitere Zielsetzung stellte die Untersuchung von Bakteriengemeinschaften im Watten-
meer und der Deutschen Bucht dar, um den Austausch von Wassermassen und die räumliche
Verteilung der dominierenden Bakteriengruppen darzustellen (Kapitel V). Hierzu wurden
zwei Messkampagnen im Sommer 2002 und 2003 mit umfassenden Probenahmen
durchgeführt, um lokale Gegebenheiten und ihren Einfluss auf die Bakteriengemeinschaften
zu erfassen.

Kapitel I Einleitung
16
I.5 Literatur
Alldredge, A. L. 1979. The chemical composition of macroscopic aggregates in two neritic seas. Limnol Oceanogr 24:855-866
Alldredge, A. L., and M. W. Silver. 1988. Characteristics, dynamics, and significance of marine snow. Prog Oceanogr 20:41-82
Amann, R. I., L. Krumholz, and D. A. Stahl. 1990. Fluorescent-oligonucleotide probing of whole cells for determinative, phylogenetic, and environmental studies in microbiology. J Bacteriol 172: 762-770
Azam, F., T. Fenchel, J. G. Field, J. S. Gray, L. A. Meyer-Reil, and T. F. Thingstad. 1983. The ecological role of water-column microbes in the sea. Mar Ecol Prog Ser 10:257-263
Azam, F., and B. C. Cho. 1987. Bacterial utilization of organic matter in the sea. In Ecology of Microbial Communities, Cambridge University Press, 261-281
Baretta, J., and P. Ruardij. 1988. Tial flat estuaries. Berlin: Springer-Verlag.
Bhaskar, P., H. P. Grossart, N. Bhosle, and M. Simon. 2005. Production of macroaggregates from dissolved exopolymeric substances (EPS) of bacterial and diatom origin. FEMS Microbiol Ecol 53: 255-264
Biddanda, B. A., and R. Benner. 1997. Carbon, nitrogen and carbohydrate fluxes during the production of particulate and dissolved organic matter by marine phytoplankton. Limnol Oceanogr 42:506-518
Bjørnsen, P. K. 1988. Phytoplankton exudation of organic matter: Why do healthy cells do it? Limnol Oceanogr 33:151-154
Cadée, G. C. 1984. Has input of organic matter into the western part of the Dutch Wadden Sea increased during the last decades? Neth Inst Sea Res Publ Ser 10:71-82
Cho, J. C., and S. J. Giovannoni. 2004. Cultivation and growth characteristics of a diverse group of oligotrophic marine Gammaproteobacteria. Appl Environ Microbiol 70: 432-440
Cottrell, M., and D. L. Kirchman. 2000. Natural assemblages of marine Proteobacteria and members of Cytophaga-Flavobacter cluster consuming low- and high-molecular-weight dissolved organic matter. Appl Environ Microbiol 66:1692-1697
Crump, B. C., J. A. Baross, and C. A. Simenstad. 1998. Dominance of particle-attached bacteria in the Columbia River estuary, USA. Aquat Microb Ecol 14:7-18
Crump, B. C., E. V. Armbrust, and J. A. Baross (1999) Phylogenetic analysis of particle-attached and free-living bacterial communities in the Columbia River, its estuary and the adjacent coastal ocean. Appl Environ Microbiol 65:3192-3204
Crump, B. C., and J. A. Baross. 2000. Characterization of the bacterially-active particle fraction in the Columbia River estuary. Mar. Ecol. Prog. Ser. 206: 13-22
De Wilde, P. A. W. J., and J. J. Beukema. 1984. The role of zoobenthos in the consumption of organic matter in the Dutch Wadden Sea. Neth Inst Sea Res Publ Ser 10:145-158
Dittmann, S. 1999. The Wadden Sea ecosystem: stability, properties and mechanisms. New York: Springer-Verlag.

Kapitel I Einleitung
17
Eilers, H., J. Pernthaler, J. Peplies, F. O. Glöckner, G. Gerdts, and R. Amann. 2001. Isolation of novel pelagic bacteria from the German Bight and their seasonal contributions to surface picoplankton. Appl Environ Microbiol 67:5134-5142
Eisma, D. 1993. Suspended matter in the aquatic environment. Sprimger, Heidelberg
Falkowski, P. G., R. T. Barber, and V. V. Smetacek. 1998. Biogeochemical controls and feedbacks on ocean primary production. Science 281: 200-207
Fusetani, N. 2000. Drugs from the Sea. Basel: Karger-Verlag, S. 1ff
Gätje, C., and K. Reise. 1998. Ökosystem Wattenmeer. Austausch-, Transport- und Stoffumwandlungsprozesse. New York: Springer-Verlag.
Gerdts, G., A. Wichels, H. Döpke, K. W. Klinge, W. Gunkel, und C. Schütt. 2004. 40-year long-term study of microbial parameters near Helgoland (German Bight, North Sea): historical view and future perspectives. Helgol Mar Res 58: 230-242
Giovannoni, S. J., E. F. DeLong, G. J. Olsen, and N. R. Pace. 1988. Phylogenetic group-specific oligodeoxynucleotide probes for identification of single microbial cells. J Bacteriol 170: 720-726
Giovannoni, S. J., and M. Rappé. 2000. Evolution, diversity, and molecular ecology of marine prokaryotes. In: D Kirchman (ed) Microbial Ecology of the Oceans, Wiley-Liss, Inc, 47-84
Hedges, J. I. 1992. Global biogeochemical cycles: progress and problems. Mar Chem 39:67-93
Heid C. A., J. Stevens, K. J. Livak, and P. M. Williams. 1996. Real-time quantitative PCR. Genome Res 6:986-994.
Ittekott, V., U. Brockmann, W. Michaelis, und E. T. Degenes. 1981. Dissolved free and combined carbohydrates during a phytoplankton bloom in the Northern North Sea. Mar Ecol Prog Ser 4: 299-305
Köpke, B., R. Willms, B. Engelen, H. Cypionka, and H. Sass. 2005. Microbial diversity in coastal subsurface sediments: A cultivation approach using various electron acceptors and substrate gradients. Appl Environ Microbiol 71: 7819-7830
Llobet-Brossa, E., R. Rossello-Mora, and R. Amann. 1998. Microbial community composition of Wadden Sea sediments as revealed by fluorescence in situ hybridization. Appl Environ Microbiol 64:2691-2696
Llobet-Brossa, E., R. Rabus, M. E. Böttcher, M. Könneke, N. Finke, A. Schramm, R. L. Meyer, S. Grotzschel, R. Rossello-Mora, and R. Amann. 2002. Community structure and activity of sulfate-reducing bacteria in an intertidal surface sediment: a multi-method approach. Aquat Microb Ecol 29:211-226
Lozan, J. L., E. Rachor, K. Reise, H. v. Westernhagen, und W. Lenz. 1994. Warnsignale aus dem Wattenmeer. Blackwell Wissenschafts-Verlag, Berlin.
Lunau, M., A. Sommer, A. Lemke, H. P. Grossart, und M. Simon. 2004. A new sampling device for microaggregates in turbid aquatic systems. Limnol Oceanogr: Methods 2:387-397
Lunau, M., A. Lemke, O. Dellwig, und M. Simon. 2006. Physical and biogeochemical controls of microaggregate dynamics in a tidally affected coastal ecosystem. Limnol Oceanogr 51: 847-859
Marsh, T. L. (1999) Terminal restriction length polymorphism (T-RFLP): an emerging

Kapitel I Einleitung
18
method for characterization diversity among homologous populations of amplification products. Curr. Opin. Microbiol. 2: 323-327.
Moran, M. A., und Kollegen. 2004. Genome sequence of Silicibacter pomeroyi reveals adaptations to the marine environment. Nature 432: 910-913
Mussmann, M., K. Ishii, R. Rabus, und R. Amann. 2005. Diversity and vertical distribution of cultured and uncultured Deltaproteobacteria in an intertidal mud flat of the Wadden Sea. Environ Microbiol 7: 405-418
Muyzer, G., E. C. de Waal, and A. G. Uitterlinden. 1993. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl Environ Microbiol 59:695-700
Passow, U. 2002. Production of transparent exopolymer particles (TEP) by phyto- and bacterioplankton. Mar Ecol Prog Ser 238: 1-12
Pernthaler, A., J. Pernthaler, and R. Amann. 2002. Fluorescence in situ hybridization and catalyzed reporter deposition for the identification of marine bacteria. Appl. Environ. Microbiol. 68: 3094-3101
Postma, H. 1981. Exchange of materials between the North Sea and the Wadden Sea. Mar Geol 40: 199-213
Rappé, M. S., K. Vergin, und S. J. Giovannoni. 2000. Phylogenetic comparisons of a coastal bacterioplankton community with its counterparts in open ocean and freshwater systems. FEMS Microbiol Ecol 33: 219-232
Rappé, M. S., S. A. Connon, K. L. Vergin, and S. J. Giovannoni. 2002. Cultivation of the ubiquitous SAR11 marine bacterioplankton clade. Nature 418:630-633
Reinthaler, T., and G. Herndl. 2005. Seasonal dynamics of bacterial growth efficiencies inr elation to phytoplankton in the southern North Sea. Aquat Microb Ecol 39:7-16
Reinthaler, T., C. Winter, and G. Herndl. 2005. Relationships between bacterioplankton richness, respiration, and production in the southern North Sea. Appl Environ Microbiol 71:2260-2266
Saiki R. K, D. H. Gelfand, S. Stoffel, S. J. Scharf, R. Higuchi, G. T. Horn, K. B. Mullis, and H. A. Ehrlich. 1988. Primer-directed Enzymatic amplification of DNA with a thermostable DNA Polymerase. Science 239:487-491
Sanger, F., G. M. Air, B. G. Barrell, N. L. Brown, A. R. Coulson, C. A. Fiddes, C. A. Hutchinson, P. M. Slocombe, and M. Smith. 1977. Nucleotide sequence of bacteriophage phi X174 DNA. Nature 265:687-695
Simon, M., H. P. Grossart, B. Schweitzer, and H. Plough. 2002. Microbial ecology of organic aggregates in aquatic ecosystems. Aquat Microb Ecol 28:175-211
Smith, D. C., M. Simon, A. L. Alldredge, and F. Azam. 1992. Intense hydrolytic enzyme activity on marine aggregates and implications for rapid particle dissolution. Nature 359:139-142
Smith, S. V., and J. T. Hollibaugh. 1993. Coastal metabolism and the oceanic organic carbon balance. Rev. Geophy. 31:75.
Smith, D. C., G. F. Steward, R. A. Long, and F. Azam. 1995. Bacterial mediation of carbon fluxes during a diatom bloom in a mesocosm. Deep Sea Res 42:75-97
Staley, J. T., und A. Konopka. 1985. Measurements of in situ activities of nonphotosynthetic microorganisms in aquatic and terrestrial habitats. Annu Rev Microbiol 39: 321-346

Kapitel I Einleitung
19
Stevens, H. 2004. Heterotrophe Bakteriengemeinschaften des Deutschen Wattenmeeres – Diversität, Dynamik und Abundanz
Stevens, H., T. Brinkhoff, and M. Simon. 2005a. Composition of free-living, aggregate-associated and sediment surface-associated bacterial communities in the German Wadden Sea. Aquat Microb Ecol 38:15-30
Stevens, H., M. Stübner, M. Simon, and T. Brinkhoff. 2005b. Phylogeny of Proteobacteria and Bacteroidetes from oxic habitats of a tidal flat system. FEMS Microb Ecol 54:351-365
Søndergaard, M., and M. Middelboe. 1995. A cross-system analysis of labile dissolved organic carbon. Mar Ecol Prog Ser 118:283-294
Willms, R., H. Sass, B. Köpke, J. Köster, H. Cypionka, and B. Engelen. 2006. Specific bacterial, archaeal, and eukaryotic communities in tidal-flat sediments along a vertical profile of several meters. Appl Environ Microbiol 72: 2756-2764
Woese, C., R. 1987. Bacterial evolution. Microbiol Rev 51:221-271
Wollast, R. 1993. Interactions of carbon and nitrogen cycles in the coastal zone. In Interactions of C, N, P and S biogeochemical cycles and global change. Wollast, R., Mackenzie, F. T., and Chou, L. (eds.) Springer-Verlag, Berlin

20
II.
Effects of a phytoplankton bloom in a coastal ecosystem
on the composition of bacterial communities

Kapitel II Effects of a phytoplankton bloom on bacterial communities
21
Effects of a phytoplankton bloom in a coastal ecosystem on the composition
of bacterial communities
Beate Rink 1, Susanne Seeberger 1, Torben Martens 1, Claus-Dieter Duerselen 2,
Meinhard Simon 1, Thorsten Brinkhoff 1*
1 Institute for Chemistry and Biology of the Marine Environment (ICBM), University of
Oldenburg, P.O. Box 2503, D-26111 Oldenburg, Germany
2 AquaEcology, Marie-Curie-Str. 1, D-26129 Oldenburg, Germany
*Corresponding author. E-mail: [email protected]
Running head: Effects of a phytoplankton bloom on bacterial communities
KEY WORDS: Free-living and particle-attached bacteria, Bacteroidetes, Roseobacter,
phytoplankton, DGGE

Kapitel II Effects of a phytoplankton bloom on bacterial communities
22
ABSTRACT: We studied the composition of free-living and aggregate-associated bacterial
communities during the course of the phytoplankton succession in spring and early summer in
the German Wadden Sea, a tidal flat ecosystem in the southern North Sea. We applied the
DGGE approach based on PCR amplified 16S rRNA gene fragments, and, in addition to
Bacteria-specific primers, used primers specific for alpha-Proteobacteria, the Roseobacter
clade, and the Bacteroidetes phylum. Even though the application of Bacteria- and alpha-
Proteobacteria-specific primers detected some changes, changes were most pronounced with
the Roseobacter- and Bacteroidetes-specific primer sets. They were supported by a
correspondence analysis, which showed a highly significant correlation of the DGGE banding
patterns of the Roseobacter specific PCR with the composition of the phytoplankton. This
indicates that changes of the phytoplankton composition in this habitat are not reflected by the
patterns of the most abundant or most readily amplifiable phylotypes. The findings rather
suggest that few, specialized heterotrophic bacteria are most responsive to the organic matter
supplied by senescent phytoplankton and that the main part of organic matter in the German
Wadden Sea is utilized by generalists. Sequence analyses of excised bands revealed a high
diversity for the Bacteria- and Bacteroidetes-specific approaches. The bacterial community
detected by the alpha-Proteobacteria-specific primer set, however, was mainly composed of
bacteria affiliated to the Roseobacter clade.

Kapitel II Effects of a phytoplankton bloom on bacterial communities
23
INTRODUCTION
Today it is well established that heterotrophic bacteria are an important component of and
key players in the biogeochemical cycling of elements and the flux of energy in aquatic
ecosystems. Depending on the ecosystem and on various environmental and biotic factors the
composition of the bacterial communities involved may exhibit distinct differences and
variations in time and space. Temperature preferences certainly select for certain bacterial
taxa but little direct information is available within this context. The most important factor for
selecting specific bacterial groups is supply by specific monomeric and polymeric
components of the dissolved organic matter (DOM) pool and of inorganic nutrients such as
phosphate, ammonium or nitrate. It has been shown that alpha-Proteobacteria prefer
monomers such as amino acids and N-acetyl-glucosamine, whereas Cytophaga/Flavobacteria
(now Sphingobacteria/Flavobacteria) of the Bacteroidetes phylum prefer polymers such as
chitin and protein, and gamma-Proteobacteria amino acids and proteins (Cottrell & Kirchman
2000). Various mesocosm studies have shown that distinct DOM components via direct
supply or the experimental induction of phytoplankton blooms select for specific bacterial
subcommunities or populations (LeBaron et al. 1999, Pinhassi et al. 2004, Riemann et al.
2000, Schäfer et al. 2001). The specific organic matter profile of various algae appears also to
be an important selection factor for distinct bacterial communities and populations evolving in
the phycosphere of algae (Grossart 1999, Grossart et al. 2005, Schäfer et al. 2002). In fact,
alpha-Proteobacteria, in particular the Roseobacter clade, and the Bacteroidetes appear to be
most responsive to inputs of phytoplankton-born DOM (Fandino et al. 2001, Grossart et al.
2005, Pinhassi et al. 2004, Riemann et al. 2000, Schäfer et al. 2001).
It is also well established that the community composition of particle-associated (PA)
bacteria differs from that of free-living (FL) bacteria. Several studies have shown that
Sphingobacteria and Flavobacteria preferentially colonize particles whereas alpha- and
gamma-Proteobacteria mainly dwell in free-living marine bacterial communities (Fandino et
al. 2001, Grossart et al. 2005, Simon et al. 2002). Our knowledge on the development and
succession of specific subcommunities and populations within PA bacterial communities
during phytoplankton blooms, however, is still fragmentary.
Experimental studies are important to elucidate single factors affecting the composition
of bacterial communities. As the aim of such studies is to better understand how the
composition of bacterial communities is controlled at ambient, but much more complex
conditions it is important to complement these studies by appropriate field observations. Such

Kapitel II Effects of a phytoplankton bloom on bacterial communities
24
studies have been carried out in various ecosystems and shown that the composition of
bacterial communities undergoes temporal changes during phytoplankton blooms (Fandino et
al. 2001, Larsen et al. 2004, Yager et al. 2001). These changes often reflect the changing
environmental conditions and DOM supply and also indicate which bacteria are mainly
involved in the biogeochemical cycling of elements and flux of energy. Denaturing gradient
gel electrophoresis (DGGE) of PCR-amplified 16S rRNA gene fragments using Bacteria-
specific primers (Muyzer et al. 1993) has been proven to be a powerful tool to assess the
composition and temporal changes of bacterial communities. Using Bacteria-specific primers
for this approach appears to be selective against the Bacteroidetes group (Cottrell &
Kirchman 2000, Selje et al. 2005, but see Castle & Kirchman 2004). Therefore, and to obtain
a more detailed insight into the composition of bacterial communities and their major players,
it is desirable to apply primers targeting specifically important groups such as Bacteroidetes
and alpha-Proteobacteria.
The aim of our study was to investigate the composition of free-living and aggregate-
associated bacterial communities during the course of the phytoplankton succession in spring
and early summer in the Wadden Sea, a tidal flat ecosystem of the southern North Sea. Based
on previous studies, we hypothesized that the expected bacterial response to the
phytoplankton succession would be reflected most pronounced by alpha-Proteobacteria and
the Sphingobacteria/Flavobacteria group. Therefore, we applied the DGGE approach and, in
addition to Bacteria-specific primers, primers specific for alpha-Proteobacteria, the
Roseobacter clade, and the Bacteroidetes group.
MATERIALS AND METHODS
Sample collection and processing. Surface water samples were collected weekly by
bucket from shipboard at high tide from 12 April to 29 June 2000 in the Backbarrier tidal flat
ecosystem of the German Wadden Sea near Spiekeroog Island (53° 44.4 N, 7° 41 E). This is a
mesotidal ecosystem characterized by high loads of suspended particulate matter (SPM). For
further details see Stevens et al. (2005a) and Lunau et al. (2006). For analysis of SPM and the
particulate carbon fractions 0.5-1 L of seawater was filtered onto pre-combusted (2 h at
550°C) and pre-weighed glass fiber filters (GF/F, Whatman, USA) and stored at –20°C in the
dark until further processing. For DGGE analysis, 250 ml of seawater were pre-filtered onto
5.0 µm polycarbonate-filters (Nuclepore) to obtain the fraction of aggregate-associated and

Kapitel II Effects of a phytoplankton bloom on bacterial communities
25
subsequently onto 0.2 µm polycarbonate-filters to obtain that of free-living bacteria. Filters
were stored at –20°C in the dark until further processing. For enumeration of bacterial and
phytoplankton cells 100 ml of water sample were fixed with formaldehyde (final
concentration 2% vol/vol) or Lugol and stored at 4°C. Hydrographic data (temperature,
salinity, pH, and oxygen) were measured by probes (LF 196, pH192, OXI 196, WTW,
Weilheim, Germany).
SPM dry weight, particulate carbon fractions. Filters were dried for 1 hour at 110°C
and weighed on a micro-balance (Sartorius, Germany). Total particulate carbon (TC) and
particulate inorganic carbon (PIC) were determined after high temperature combustion and
titration of the CO2 produced against Ba(ClO4)2. Particulate organic carbon (POC) was
calculated as the difference of TC and PIC. For further details see Stevens et al. (2005a).
Bacterial and algal cell counts. Abundance of free-living and aggregate-associated
bacteria was enumerated after DAPI (4´-6-diamidino-2-phenylindole) staining by
epifluorescence microscopy at 1000x magnification according to Crump et al. (1998). To
distinguish particle-attached and free-living bacteria, seawater was fractionated by filtration
onto 5.0 µm and subsequently onto 0.2 µm polycarbonate-filters. To reduce the background
fluorescence by inorganic matter filters were counter-stained with an acridine orange solution
(0.1%). Lugol-fixed phytoplankton samples were enumerated by inverted microscopy.
Phytoplankton was identified on the species level when possible. For estimating
phytoplankton biomass cell numbers were multiplied by cell carbon. The latter was estimated
from measured cell sizes of individual cells converted to carbon according to empirical
carbon/cell volume conversion factors from the Biologische Anstalt Helgoland (J. Berg,
unpubl. results).
Nucleic acid extraction. The isolation of genomic DNA was performed by phenol-
chloroform extraction after bead beating as described earlier with slight modifications (Selje
& Simon, 2003). The precipitation was done overnight at –20°C using isopropanol. The DNA
was resuspended in molecular grade water (Eppendorf, Germany) and stored at –20°C until
further processing.
Primer sets. PCR amplification of 16S rRNA gene fragments was performed with primer
pairs specific for Bacteria (GC 341F and 907RM), the Bacteroidetes phylum (GC-CF319f
and 907RM), alpha-Proteobacteria (GC 341F and ALF968r), and the Roseobacter clade
within alpha-Proteobacteria (GC ROSEO536Rf and GRb735r). Primer sequences and
references are given in Table 1. ’GC’ indicates that a GC clamp was added to the primer
(Muyzer et al. 1993). For the primer GC ROSEO536Rf the following GC clamp was used: 5’-

Kapitel II Effects of a phytoplankton bloom on bacterial communities
26
CGCCCGCCGCGCCCCGCGCCCGTCCCGCCGCCCCCGCCCG-3’. For the sequences of
the other GC clamps used in this study see the references cited in Table 1. Specificity of the
primers used for Bacteroidetes was described earlier (Jaspers et al. 2001, Kirchman 2002).
The oligonucleotide probe ALF968r (Neef, 1997), used as reverse primer for alpha-
Proteobacteria, was tested theoretically using the BLAST function of the NCBI server
(http://www.ncbi.nlm.nih.gov). Search results for this primer sequence revealed up to 10%
matches to other phylogenetic groups with 100% sequence similarity for the first one hundred
matches. The primer set used for the Roseobacter-group was tested theoretically with the
whole database of the ARB software package (Ludwig et al. 2004) and recently published
sequences present in GenBank (www.ncbi.nlm.nih.gov) of cultivated and uncultivated
organisms affiliated with the Roseobacter clade. In total 183 sequences affiliated with this
group were considered. Specificity was also tested in PCR assays using several described
species as positive and negative controls (Table 2), and 25 isolates affiliated with the
Roseobacter group, taken from our culture collection.
PCR amplification of 16S rRNA gene fragments. PCR amplifications were performed
with an Eppendorf Mastercycler (Eppendorf, Hamburg, Germany) as follows: One µl of
template was added to 49 µl of PCR mixture containing 1 U of Sigma RedTaqTM polymerase
and 5 µl 10 x RedTaqTM PCR buffer (Sigma, Deisenhofen, Germany), bovine serum albumin
(10 mg ml-1), 250 µM of each deoxynucleotide triphosphate, 2.1 µM MgCl2, and 20 pmol of
each primer. The PCR protocol for the Bacteria-specific primer set was performed as
described previously (Brinkhoff & Muyzer, 1997). Amplification of the 16S rRNA gene
fragments of alpha-Proteobacteria was performed under the same conditions with an
annealing temperature of 65°C for 10 cycles and subsequently 55°C for 20 cycles.
Roseobacter-specific PCR conditions were 5 cycles at 65°C and 25 cycles with an annealing
temperature of 63°C. For highest specificity, a maximum of 30 cycles is recommendable at
this step. PCR with the primer set specific for Bacteroidetes was performed as described
previously (Jaspers et al. 2001). Four µl of the amplification products were analyzed by
electrophoresis in 2% (w/v) agarose gels and stained with ethidium bromide (1 µg ml-1)
(Sambrook et al. 1989). For subsequent sequencing analysis PCR products were purified by
using the Qiaquick PCR purification kit (Qiagen Inc., Chatsworth, Calif.).
DGGE analysis of PCR products. DGGE was performed with the D-Code system (Bio-
Rad Laboratories, Inc.). For gene fragments of Bacteria and alpha-Proteobacteria, the
protocol described by Brinkhoff & Muyzer (1997) was used. For 16S rRNA gene fragments
obtained with the primer pair GC-CF319f and 907 RM the gradient was modified to 15 to

Kapitel II Effects of a phytoplankton bloom on bacterial communities
27
85% denaturant. DGGE analysis of Roseobacter 16S rRNA gene fragments was performed
with 20 to 70% denaturant and 9% (wt/vol) polyacrylamide content. After electrophoresis, the
gels were stained with SYBR Gold (Molecular Probes, Inc.) and photographed using a
BioDoc Analyze Transilluminator (Biometra, Göttingen, Germany). Bands were excised with
a scalpel sterilized with ethanol and transferred to sterile Eppendorf caps. Fifty µl of water
(molecular grade, Eppendorf, Germany) were added and the samples were stored at –20°C.
Cloning. Twenty four DGGE bands (GWS-e1-FL to GWS-e13-PA, GWS-c3-FL, GWS-
c16-PA, GWS-c9-PA, GWS-c10-PA, GWS-c18-PA and GWS-a10-PA to GWS-a13-PA,
GWS-a4-FL, GWS-a8-FL) were cloned using the pGEM®-T Vector System II (Promega,
Madison, USA) following the instruction manual. Clones with inserts were picked,
resuspended in molecular grade water (Eppendorf, Germany) and screened by DGGE to
check if the insert position matches the position of the corresponding DGGE band. Adequate
clones were amplified and subsequently sequenced using the primers pUC/M13f and
pUC/M13r (Messing, 1983) with an annealing temperature of 48°C.
Sequencing and phylogenetic analysis. PCR products were sequenced using the
DYEnamic Direct cycle sequencing kit (Amersham Life Science, Inc.) and a Model 4200
Automated DNA Sequencer (LI-COR, Inc.). Sequencing primers were 341F and 907RM for
direct sequencing of DGGE bands, or M13 primers as described above for cloned bands
labeled with IRDyeTM800. For all sequences, at least 400 bp were determined. Phylogenetic
affiliation of the sequences was compared to those in GenBank using the BLAST function of
the NCBI server (http://www.ncbi.nlm.nih.gov/BLAST/). Phylogenetic trees were constructed
using the ARB software package (Ludwig et al. 2004, http://www.arb-home.de). The
backbone tree was calculated with the maximum likelihood method using sequences with a
minimum length of 1300 bp including type strains of the selected phylogenetic groups. For
tree calculation, positions were excluded at which less than 50% of all sequences showed the
same residues to avoid uncertain alignments. Sequences with less than 1300 bp were added to
the backbone tree with the maximum parsimony method using the same filter. As an
outgroup, 16S rRNA gene sequences of seven type strains belonging to Cyanobacteria were
used.
The sequences obtained in this study are available from GenBank under accession no.
DQ080919 to DQ080962.
Statistics. Cluster analyses of DGGE banding patterns were performed using Gel
Compar II, version 2.5 (Applied maths, Kortrijk, Belgium). Calculations were done curve
based using Pearson correlation and UPGMA. A correspondence analysis of the DGGE

Kapitel II Effects of a phytoplankton bloom on bacterial communities
28
banding patterns and the phytoplankton composition was performed using ADE-4
(Thioulouse et al. 1997). To analyze the bacterial community structure, we exported the raw
data of the cluster analysis and generated a matrix based on the specific band heights. For
phytoplankton, we used relative species abundance. A modified correspondence analysis was
performed row weighted on a biplot scale. After calculation of the COA for each community
a Coinertia analysis was performed to connect the data. A permutation test based on the
Monte Carlo method was calculated using the Coinertia test (– Fixed D; number of random
matching: 1000).
RESULTS
Environmental conditions and SPM properties
From the start of the study period in mid-April until 10 May 2000 the water temperature
continuously increased from 8-17 °C (Fig. 1A). Thereafter it fluctuated between 17 and 13
°C. Salinity ranged between 29 and 32‰ (Fig. 1A) and SPM dry weight from 80-120 mg l-1
in April and May, but increased to 160 mg L-1 on 14 June (data not shown). POC
concentrations varied between 0.8 mg L-1 on 3 May and 4.7 mg L-1 on 26 April (Fig. 1B).
They steadily increased from 3 to 17 May and from 24 May to 14 June.
Phytoplankton and bacterial dynamics
The phytoplankton consisted exclusively of diatoms and few dinoflagellates. From 12
April to 3 May diatom cell numbers strongly decreased from 6.5x103-1.2x103 L-1 but
thereafter continuously increased until 24 May (Fig. 1C). After the decline of this bloom in
late May only low numbers remained. Whereas the initial bloom on 12 April exhibited a high
diversity and evenness, the bloom in May became more and more dominated by Guinardia
delicatula, constituting 70% of algal cell numbers and biomass on 24 May (Fig. 1C). One
week later, when diatom cell numbers declined to ~30% of the previous week, abundance of
Guinardia delicatula had strongly decreased while Pseudonitzschia pungens constituted 50%
of the cell numbers. At the onset of the study phytoplankton constituted 50% of POC, but on
26 April only 4%. Thereafter during the Guinardia bloom, phytoplankton carbon
continuously increased to 13% on 24 May (Fig. 1B).

Kapitel II Effects of a phytoplankton bloom on bacterial communities
29
Cell numbers of FL bacteria increased from 12 April until 3 May from 2.4x106-4.0x106
ml-1 and fluctuated thereafter around this value (Fig. 1D). Cell numbers of PA bacteria were
lower and ranged from 0.5-1.1x106 ml-1, accounting for 11 to 22% of total bacterial numbers.
Specificity of the Roseobacter primer set
Comparison of 16S rRNA gene fragments present in our ARB database revealed that the
forward primer GC ROSEO536Rf matched 131 of a total of 183 target sequences affiliated to
the Roseobacter clade. Forty three Roseobacter sequences had no or incomplete information
at the target site of the primer, and 8 sequences of uncultured Roseobacter-affiliated
organisms showed up to three mismatches to the primer sequence. Sulfitobacter pontiacus
(Acc. no. Y13155) had one mismatch at position 17 of the primer sequence. Reverse primer
GRb735r targeted 133 sequences after insertion of a wobble (G/A) at Escherichia coli
position 752. Forty four Roseobacter sequences had no or incomplete information at the target
site of the 16S rRNA gene. Six sequences of target bacteria had up to three mismatches:
Roseobacter sp. J8W (AF026462; two mismatches), Roseobacter sp. J2W (AF026462, three
mismatches), Roseobacter sp. KT11117 (AF173971, one mismatch), Adriatic 72 (AF030780,
one mismatch, two non-defined bases), Sulfitobacter pontiacus (Y13155, one mismatch) and
GWS-BW-H66M (AY515422, one mismatch). The non-target sequences of Rhodovulum
iodosum and clone SAR102 (Acc. no. L35460) had no mismatches to the primer sequence.
Considering all respective sequences in the ARB data base, the use of both primers paired
resulted in at least one mismatch to all other phylogenetic groups.
PCR results showed that the specificity and sensitivity of the Roseobacter primer set was
very high. With one step down from 65°C -63°C and 1 U of Taq polymerase, 0.2 ng genomic
DNA µl-1 of Roseobacter gallaeciensis was detectable. DNA of the non-target organism
Paracoccus aminophilus (1 mismatch to the target sequence) was detected down to 2 ng µl-1.
Specificity was higher but less sensitive under the same conditions with 0.5 U of polymerase,
detecting 0.1-1 ng DNA µl-1 of R. gallaeciensis and 20 ng DNA µl-1 of P. aminophilus. To
determine a possible sequence preference of the primer set, a DNA mixture of both organisms
with equal DNA amounts was amplified and the PCR products were analyzed using DGGE.
By this approach only amplicons of R. gallaeciensis could be detected (data not shown). This
notion suggests that the amplification of non-target organisms is suppressed under the chosen
PCR and DGGE conditions.

Kapitel II Effects of a phytoplankton bloom on bacterial communities
30
DGGE banding patterns
The DGGE analyses with the various primer sets showed distinctly different banding
patterns of the FL as well as PA bacterial communities (Fig. 2A-D). The Bacteria-specific
primer set yielded 12 to 15 bands per lane in the PA bacterial fraction and 12 to 18 bands in
the FL bacterial fraction (Fig. 2A). Changes in the banding patterns occurred mainly during
the Guinardia bloom in May, showing a slight increase of band numbers in the FL fraction on
17 May and the appearance of a strong band in the PA fraction (GWS-e11-PA). The cluster
analysis yielded distinct clusters for the FL and PA bacterial communities (Fig. 3A). Only
during the Guinardia bloom between 10 and 24 May the PA bacteria clustered separately and
closer with the FL bacterial community. Correspondence analysis did not yield a significant
correlation with the phytoplankton composition. The aggregate sample of 31 May was
reamplified from a former PCR product and showed reduced band numbers compared to the
other samples. Hence, the fingerprint of this sample appeared as an outgroup in the cluster
analysis and was not regarded for the further discussion.
The Bacteroidetes–specific banding patterns revealed 7-12 and 9-18 amplicons per lane
in the FL and PA bacterial fractions, respectively (Fig. 2B). Low numbers of 7-9 bands
occurred in the FL bacterial fraction before and after the Guinardia bloom and higher
numbers of 10-12 bands during the bloom. In contrast, the number of bands in the PA
bacterial community was high before and after and decreased during the bloom. The cluster
analysis showed a distinct cluster of the FL bacterial community, excluding the dates towards
the end of the Guinardia bloom, when banding patterns clustered together with those of the
PA bacterial community during the bloom (Fig. 3B). Furthermore, the latter fraction exhibited
clearly different patterns before and after the bloom. A correspondence analysis revealed a
moderate correlation of the banding patterns with the composition of the phytoplankton
(P=0.067).
DGGE banding patterns specific for alpha-Proteobacteria showed 7-8 and 8-13 bands per
lane in the FL and PA bacterial communities, respectively (Fig. 2C). Most of the bands were
permanently present but a few bands in both fractions occurred in the course of the bloom
(GWS-a11-PA, GWS-a6-FL, and GWS-a5-FL). The cluster analysis showed generally rather
complex patterns and that PA bacteria during the Guinardia bloom clustered together with FL
bacteria (Fig. 3C). The correspondence analysis did not yield a significant correlation of the
banding patterns with the phytoplankton composition.

Kapitel II Effects of a phytoplankton bloom on bacterial communities
31
The Roseobacter-specific DGGE banding patterns showed 5-8 bands per lane in the FL
bacterial fraction and 8-18 bands in the PA bacterial fraction (Fig. 2D). Quite a few bands
were permanently present in both fractions, but additional bands occurred during the decline
of the bloom in April and the Guinardia bloom in May, mainly in the PA bacterial fraction.
The cluster analysis yielded complex patterns with several subclusters both of FL and PA
associated bacterial fractions. A distinct subcluster comprised the banding patterns of both
fractions during the Guinardia bloom (Fig. 3D). The correspondence analysis showed a
highly significant correlation of the banding patterns with the composition of the
phytoplankton (P=0.03).
Phylogenetic affiliation
The sequence analysis of excised bands revealed a high diversity of the obtained
phylotypes for the 16S rRNA gene fragments of the Bacteria- and Bacteroidetes-specific
approaches (sequences obtained with Bacteria-specific primers were designated GWS-e and
sequences obtained with Bacteroidetes-specific primers as GWS-c; Figs. 4A, B). The
bacterial community detected by the alpha-Proteobacteria-specific primer set (sequences
GWS-a) was mainly composed of bacteria belonging to the Roseobacter clade. Most
phylotypes of this group, detected by the Bacteria- and alpha-Proteobacteria-specific primer
set, clustered within the recently described WAC I cluster (Stevens et al. 2005b) or RCA
cluster (Selje et al. 2004). The primer set used for alpha-Proteobacteria turned out to be not
specific, as sequencing results revealed that two sequences affiliated to delta-Proteobacteria
(GWS-a12-PA, GWS-a13-PA) and one to Bacteroidetes (GWS-a8-FL). In contrast, although
the primer GC-CF319f used for specific amplification of 16S rRNA gene sequences of
bacteria belonging to Bacteroidetes is known to be unspecific (Kirchman et al. 2003), all our
phylotypes of the sequenced bands fell into this phylum.
During the Guinardia bloom, DGGE derived phylotypes belonging to the WAC I cluster
dominated the FL bacterial fraction. DGGE band GWS-e7-FL was present during the bloom.
This phylotype was closely related to GWS-a6-FL and GWS-a5-FL (sequence differences
<0.8%, Fig. 4A) which were also present only during the Guinardia bloom. While these
organisms seem to be highly responsive to the phytoplankton composition, other members of
the WAC I cluster were present during the whole investigation period, e.g. GWS-e6-FL (Fig.
4A). This phylotype is closely related to DGGE band GWS-FL-3, which was persistently
detected throughout the year in the Wadden Sea, indicating that this organism is well adapted
to highly variable biotic and environmental conditions in this habitat (Stevens et al. 2005a). In

Kapitel II Effects of a phytoplankton bloom on bacterial communities
32
the PA bacterial fraction chloroplast DNA (GWS-e11-PA) represented the most significant
change within the community detected by the Bacteria-specific primer set. Sequencing of
other conspicuous bands was not possible, as the diffuse bands in the upper part of the gel
could not be reamplified.
One of the Bacteroidetes–specific phylotypes appeared during 24 May to 14 June at the
end of the Guinardia bloom both in the FL and PA bacterial fractions (GWS-c6-FL, GWS-c5-
FL and GWS-c15-PA). This phylotype is closely related to GWS-e9-FL in the FL bacterial
fraction (Fig. 4B). BLAST results revealed that the closest related sequence of these bands is
DGGE band GWS-AG-8, which was detected on aggregates in June 2000 in the same area
(Stevens et al. 2005a). Other phylotypes affiliated to the Bacteroidetes were present during
the whole investigation period. DGGE band GWS-c8-PA was detected in the PA bacterial
fraction from April to June and is closely related to strain T15 (AY177723, 99% similarity,
502/505 bp), isolated from the same habitat in October 1999 (Brinkhoff et al. 2004). The
phylotype was also detected in a seasonal sampling campaign in this habitat from 2000 to
2002 using GC-CF319f and 907RM for DGGE analysis (S. Seeberger, unpublished results).
DISCUSSION
Our results indicate that the composition of the bacterial communities in the Wadden Sea
underwent changes during the phytoplankton succession in spring and early summer. These
changes were detected as the disappearance of DGGE bands and the appearance of new ones,
were most pronounced during the Guinardia bloom and its decline in May, and occurred in
the FL as well as in the PA bacterial communities. Even though the application of Bacteria-
and alpha-Proteobacteria-specific primers in the DGGE approach detected some of these
changes, they were detected most clearly with the Roseobacter- and Bacteroidetes-specific
primer sets and supported by a correspondence analysis. Whereas the number of bands of the
FL bacterial fraction within the Bacteroidetes increased during the Guinardia bloom it
decreased in the PA bacterial fraction. Within the Roseobacter clade, the number of bands of
the PA bacterial fraction increased during the decline of both blooms, in late April and late
May. Hence our results show that the bacterial communities respond to the changing
phytoplankton community and organic matter field on a rather specific phylogenetic level and
call for applying class- and subclass-specific primer sets in the DGGE approach for such
investigations.

Kapitel II Effects of a phytoplankton bloom on bacterial communities
33
Our investigation complements mesocosm experiments which obtained similar findings
(LeBaron et al. 1999, Pinhassi et al. 2004, Riemann et al. 2000, Schäfer et al. 2001) and
demonstrate that changes in FL as well as PA bacterial communities during the development
of phytoplankton blooms also occur and can be detected at ambient conditions in a natural
ecosystem. Our results, however, go beyond the mentioned experimental studies by showing
in a much more detailed way the different responses of bacteria affiliated to the Roseobacter
clade and the Bacteroidetes.
To achieve these results we applied sets of published primers for all bacterial target
groups and modified specific probes for the Roseobacter clade from earlier studies to
optimize its specificity (Table 1). In addition, we developed a PCR to achieve highest
specifity for this primer set. As expected from the BLAST search, the results revealed that the
alpha-Proteobacteria-specific primer set was not specific. We detected two sequences
affiliated to delta-Proteobacteria and one to the Bacteroidetes phylum, indicating that
sequencing of bands is essential when applying this primer set. However, the great majority of
the bands sequenced affiliated to alpha-Proteobacteria and exclusively to the Roseobacter
clade, underscoring the significance to apply a primer set specific for this clade. In this study,
the Bacteroidetes-specific primer set, in fact, was specific, as all sequenced bands affiliated to
the respective target group. The fact that most changes within the bacterial communities were
only detected by applying primer sets for a lower phylogenetic level indicates that these
changes do not affect the most abundant or most readily amplifiable phylotypes.
Although various prominent bands were visible using the group-specific primer set, only
two Bacteroidetes affiliated phylotypes could be detected with the Bacteria-specific primer
set. Only one of these two phylotypes was also detected with the group-specific primer set
(GWS-e9-FL, Fig. 4B) suggesting that the Bacteria-specific primer set discriminates the
Bacteroidetes affiliated bacteria, as has been reported previously (Cottrell & Kirchman,
2000). In contrast, we had no indication of a biased amplification of phylotypes affiliated to
alpha-Proteobacteria by the Bacteria-specific primer set. Seven of the 17 sequences of this
subclass were amplified by the Bacteria-specific primer set and all except one sequence
(GWS-e10-FL) were very closely related or similar to those amplified by the alpha-
Proteobacteria-specific primer set (Fig. 4A). However, the latter and the Roseobacter-
specific primer set yielded a much better resolution and detected substantially more
phylotypes with a presumably lower abundance.
The decreasing number of bands of Bacteroidetes in the PA bacterial fraction detected
during the Guinardia bloom and its decline indicates the formation of a more specialized

Kapitel II Effects of a phytoplankton bloom on bacterial communities
34
environment on suspended particles and aggregates to which fewer bacteria of this
phylogenetic group were able to adapt. The number of bands of the Roseobacter clade,
however, remained rather unchanged and even increased at the decline of the bloom on 31
May, indicating that the niche diversity for this bacterial group did not decrease.
In contrast to aggregates, the number of bands of FL bacterial phylotypes belonging to
the Bacteroidetes increased during the bloom, indicating that the DOM supply became more
diverse, presumably including a variety of polymers released from growing and decaying
diatoms and solubilizing phytodetrital aggregates. Two of the newly occurring phylotypes
cluster together (GWS-c6-FL, GWS-c5-FL) and also together with other phylotypes which
occurred on 14 June (GWS-e9-FL) in the FL bacterial fraction and in the PA bacterial fraction
(GWS-c15-PA, Fig. 4B). These phylotypes are closely related to phylotypes which were
retrieved from the associated bacterial communities of two diatoms [SB-42-DB (Schäfer et al.
2001), Flo-21 (Grossart et al. 2005)], suggesting that they are particularly adapted to the
organic matter profile of diatoms.
Our results show that organisms of the Roseobacter clade and the Bacteroidetes are most
responsive to the changing organic matter field during the phytoplankton blooms. This is in
line with other studies (Dang & Lovell, 2002, Lebaron et al. 1999, Riemann et al. 2000,
Fandino et al. 2001, Pinhassi et al. 2004, Grossart et al. 2005) and thus indicates that members
of these two bacterial groups appear to be particularly adapted to such conditions, at least in
temperate waters. The significance of the Bacteroidetes affiliated bacteria in consuming
complex and polymeric DOM in marine systems is well known and fairly well understood,
mainly because of their specific properties to hydrolyze polymers (Cottrell & Kirchman,
2000; Kirchman, 2002). The significance of the Roseobacter clade is much less understood.
Some members of this clade exhibit aerobic anoxygenic photosynthesis but the significance of
this metabolic pathway at ambient conditions and varying trophic state is still unclear
(Schwalbach & Fuhrman, 2005). Other members of this clade appear to be involved in the
decomposition of DMS (Moran et al. 2003). Roseobacter strains have been isolated from FL
as well as PA bacterial communities and quite a few of them exhibit antibiotic and quorum
sensing properties (Long & Azam 2001, Gram et al. 2002; Grossart et al. 2004). Hence, it
appears conceivable that varying adaptive properties make this clade well suitable to dwell
successfully in marine systems. More work, however, is needed to better understand the
success of this clade at a physiological and genetic level.
The combined application of Bacteria- and group-specific primer sets revealed that a
hierarchical structure exists in the bacterial communities, both in the FL as well as the PA

Kapitel II Effects of a phytoplankton bloom on bacterial communities
35
fractions. The Bacteria-specific primer set detected mainly those phylotypes which constitute
the main and often dominant components of the bacterial communities, persisting throughout
most of the time and thus comprising bacteria able to adapt to rather variable environmental
conditions and exhibiting a rather generalistic life style. These phylotypes include members of
the RCA-cluster (Selje et al. 2004), and the WAC I cluster (Stevens et al. 2005b) of the
Roseobacter clade. In contrast, the group- and clade-specific primer sets detect, besides some
of these generalistic phylotypes, others which are probably less abundant but appear at
distinct environmental and biotic conditions, such as during certain periods of phytoplankton
blooms. The phylotypes detected by these primer sets reflect in a much more sensitive way
these changing conditions and thus allow a more detailed analysis of bacterial communities at
varying environmental conditions. The application of Bacteria-specific primer sets appears to
be appropriate to study the main components of bacterial communities and their variability at
greatly varying environmental conditions such as in salinity gradients (Selje & Simon, 2003;
Troussellier et al. 2002), PA vs. FL bacterial communities (Stevens et al. 2005a), or in
manipulated mesocosms (Lebaron et al. 1999, Pinhassi et al. 2004, Riemann et al. 2000). In
other cases when more subtle variations or discrimination against specific target groups may
occur, this approach appears not sensitive enough to comprehensively detect these changes.
Then the application of more specific primer sets is a valuable tool to detect these changes
which are an important indication of distinct responses of the bacterial communities to their
changing environment.
Acknowledgements. We appreciate the hospitality and assistance of the RV Senckenberg
crew. We thank B. Kuerzel and R. Weinert for dry weight analyses and H.-P. Grossart for
valuable discussions. This work was supported by the Deutsche Forschungsgemeinschaft
(DFG) within the research group “BioGeoChemistry of Tidal Flats” (FG 432 TP5).

Kapitel II Effects of a phytoplankton bloom on bacterial communities
36
LITERATURE CITED
Brinkhoff T, Muyzer G (1997) Increased species diversity and extended habitat range of sulfur-oxidizing Thiomicrospira spp. Appl Environ Microbiol 63: 3789-3796
Brinkhoff T, Bach G, Heidorn T, Liang L, Schlingloff A, Simon M (2004) Antibiotic production by a Roseobacter clade-affiliated species from the German Wadden Sea and its antagonistic effects on indigenous isolates. Appl Environ Microbiol 70: 2560-2565
Brinkmeyer R, Rappé M, Gallacher S, Medlin L (2000) Development of clade- (Roseobacter and Alteromonas) and taxon- specific oligonucleotide probes to study interactions between toxic dinoflagellates and their associated bacteria. Eur J Phycol 35: 315-329
Castle D, Kirchman DL (2004) Composition of estuarine bacterial communities assessed by denaturing gradient gel electrophoresis and fluorescence in situ hybridization. Limnol Oceanogr Methods 2: 303-314
Cottrell MT, Kirchman DL (2000) Natural assemblages of marine proteobacteria and members of the Cytophaga-Flavobacter cluster consuming low- and high-molecular-weight dissolved organic matter. Appl Environ Microbiol 66: 1692-1697
Crump BC, Baross JA, Simenstad CA (1998) Dominance of particle-attached bacteria in the Columbia River estuary, USA. Aquat Microb Ecol 14: 7-18
Dang H, Lovell CR (2002) Seasonal dynamics of particle-associated and free-living marine Proteobacteria in a salt marsh tidal creek as determined using fluorescence in situ hybridization. Environ Microbiol 4:287-295
Fandino LB, Riemann L, Steward GF, Long RA, Azam F (2001) Variations in bacterial community structure during a dinoflagellate bloom analyzed by DGGE and 16S rDNA sequencing. Aquat Microb Ecol 23:119-130
Giuliano L, De Domenico M, De Domenico E, Höfle MG, Yakimov MM (1999) Identification of culturable oligotrophic bacteria within naturally occurring bacterioplankton communities of the Ligurian Sea by 16S rRNA sequencing and probing. Microb Ecol 37:77-85
Gram L, Grossart HP, Schlingloff A, Kiorboe T (2002) Possible quorum sensing in marine snow bacteria: production of acylated homoserine lactones by Roseobacter strains isolated from marine snow. Appl Environ Microbiol 68:4111-4116
Grossart HP (1999) Interactions between marine bacteria and axenic diatoms (Cylindrotheca
fusiformis, Nitzschia laevis, and Thalassiosira weissflogii) incubated under various conditions in the lab. Aquat Microb Ecol 19:1-11
Grossart HP, Schlingloff A, Bernhard M, Simon M, Brinkhoff T (2004a) Antagonistic activity of bacteria isolated from organic aggregates of the German Wadden Sea. FEMS Microbiol Ecol 47:387-396
Grossart HP, Levold F, Allgaier M, Simon M, Brinkhoff T (2005) Marine diatom species harbour distinct bacterial communities. Environ Microbiol 7:860-873
Jaspers E, Nauhaus K, Cypionka H, Overmann J (2001) Multitude and temporal variability of ecological niches as indicated by the diversity of cultivated bacterioplankton. FEMS Microbiol Ecol 36:153-164
Kirchman DL (2002) The ecology of Cytophaga-Flavobacteria in aquatic environments. FEMS Microbiol Ecol 39:91-100

Kapitel II Effects of a phytoplankton bloom on bacterial communities
37
Kirchman DL, Yu L, Cottrell MT (2003) Diversity and abundance of uncultured Cytophaga-like bacteria in the Delaware estuary. Appl Environ Microbiol 69:6587-6596
Larsen A, Flaten GAF, Sandaa RA, Castberg T, Thyrhaug R, Erga SR, Jacquet S, Bratbak G (2004) Spring phytoplankton bloom dynamics in Norwegian coastal waters: Microbial community succession and diversity. Limnol Oceanogr 49:180-190
Lebaron P, Servais P, Troussellier M, Courties C, Vives-Rego J, Muyzer G, Bernard L, Guindulain T, Schäfer H, Stackebrandt E (1999) Changes in bacterial community structure in seawater mesocosms differing in their nutrient status. Aquat Microb Ecol 19:255-267
Long RA, Azam F (2001) Antagonistic interactions among marine pelagic bacteria. Appl Environ Microbiol 67:4975-4983
Ludwig W, Strunk O, Westram R et al. (2004) ARB: a software environment for sequence data. Nucl Acids Res 32:1363-1371
Lunau M, Lemke A, Dellwig O, Simon M (2006) Physical and biogeochemical controls of microaggregate dynamics in a tidally affected coastal ecosystem. Limnol Oceanogr 51:847-859
Manz W, Amann R, Ludwig W, Vancanneyt M, Schleifer KH (1996) Application of a suite of 16S rRNA-specific oligonucleotide probes designed to investigate bacteria of the phylum cytophaga-flavobacter-bacteroides in the natural environment. Microbiology 142:1097-1106
Messing J (1983) New M13 vectors for cloning. Meth Enzymol 101:20-78
Moran MA, Gonzalez JM, Kiene RP (2003) Linking a bacterial taxon to sulfur cycling in the sea: Studies of the marine Roseobacter group. Geomicrobiol J 20:375-388
Muyzer G, de Waal EC, Uitterlinden AG (1993) Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl Environ Microbiol 59:695-700
Neef A (1997) Anwendung der in-situ-Einzell-Identifizierung von Bakterien zur Populationsanalyse in komplexen mikrobiellen Biozönosen. PhD Thesis, Technische Universität München.
Pinhassi J, Sala MM, Havskum H, Peters F, Guadayol O, Malits A, Marrase CL (2004) Changes in bacterioplankton composition under different phytoplankton regimens. Appl Environ Microbiol 70:6753-6766
Riemann L, Steward GF, Azam F (2000) Dynamics of bacterial community composition and activity during a mesocosm diatom bloom. Appl Environ Microbiol 66:578-587
Sambrook J, Frisch EF, Maniatis T (1989) Northern Hybridisation. In Molecular Cloning: a laboratory manual, 2nd edn., Cold Spring Harbour Laboratory Press, New York
Schäfer H, Bernard L, Courties C, et al. (2001) Microbial community dynamics in Mediterranean nutrient-enriched seawater mesocosms: changes in the genetic diversity of bacterial populations. FEMS Microbiol Ecol 34:243-253
Schäfer H, Abbas B, Witte H, Muyzer G (2002) Genetic diversity of 'satellite' bacteria present in cultures of marine diatoms. FEMS Microbiol Ecol 42:25-35
Schwalbach MS & Fuhrman JA (2005) Wide-ranging abundances of aerobic anoxygenic phototrophic bacteria in the world ocean revealed by epifluorescence microscopy and quantitative PCR. Limnol Oceanogr 50:620-628

Kapitel II Effects of a phytoplankton bloom on bacterial communities
38
Selje N, Simon M (2003) Composition and dynamics of particle-associated and free-living bacterial communities in the Weser estuary, Germany. Aquat Microb Ecol 30:221-237
Selje N, Simon M, Brinkhoff T (2004) A newly discovered Roseobacter cluster in temperate and polar oceans. Nature 427:445-448
Selje N, Brinkhoff T, Simon M (2005) Detection of abundant bacteria in the Weser estuary using culture-dependent and culture-independent approaches. Aquat Microb Ecol 39:17-34
Simon M, Grossart HP, Schweitzer B, Ploug H (2002) Microbial ecology of organic aggregates in aquatic ecosystems. Aquat Microb Ecol 28:175-211
Stevens H, Brinkhoff T, Simon M (2005a) Composition of free-living, aggregate-associated and sediment surface-associated bacterial communities in the German Wadden Sea. Aquat Microb Ecol 38:15-30
Stevens H, Stübner M, Simon M, Brinkhoff T (2005b) Phylogeny of Proteobacteria and Bacteroidetes from oxic habitats of a tidal flat ecosystem. FEMS Microbiol Ecol 54:351-365
Thioulouse J, Chessel D, Dolédec S, Olivier JM (1997) ADE-4: a multivariate analysis and graphical display software. Statistics and Computing 7:75-83
Troussellier M, Schäfer H, Batailler N, Bernard L, Courties C, Lebaron P, Muyzer G, Servais P, Vives-Rego J (2002) Bacterial activity and genetic richness along an estuarine gradient (Rhone River plume, France). Aquat Microb Ecol 28:13-24
Yager PL, Connelly TL, Mortazavi B, Wommack KE, Bano N, Bauer JE, Opsahl S, Hollibaugh JT (2001) Dynamic bacterial and viral response to an algal bloom at subzero temperatures. Limnol Oceanogr 46:790-801
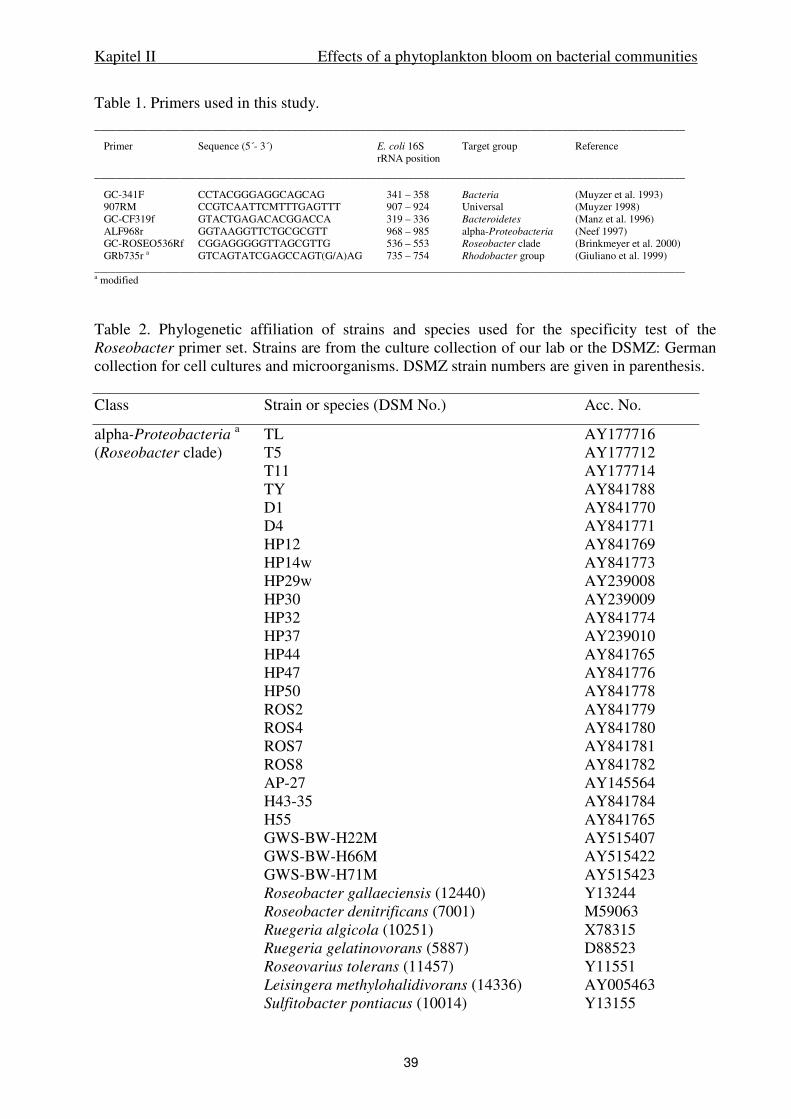
Kapitel II Effects of a phytoplankton bloom on bacterial communities
39
Table 1. Primers used in this study. _______________________________________________________________________________________________________________
Primer Sequence (5´- 3´) E. coli 16S Target group Reference rRNA position
_______________________________________________________________________________________________________________
GC-341F CCTACGGGAGGCAGCAG 341 – 358 Bacteria (Muyzer et al. 1993) 907RM CCGTCAATTCMTTTGAGTTT 907 – 924 Universal (Muyzer 1998) GC-CF319f GTACTGAGACACGGACCA 319 – 336 Bacteroidetes (Manz et al. 1996) ALF968r GGTAAGGTTCTGCGCGTT 968 – 985 alpha-Proteobacteria (Neef 1997) GC-ROSEO536Rf CGGAGGGGGTTAGCGTTG 536 – 553 Roseobacter clade (Brinkmeyer et al. 2000) GRb735r a GTCAGTATCGAGCCAGT(G/A)AG 735 – 754 Rhodobacter group (Giuliano et al. 1999)
_______________________________________________________________________________________________________________ a modified
Table 2. Phylogenetic affiliation of strains and species used for the specificity test of the Roseobacter primer set. Strains are from the culture collection of our lab or the DSMZ: German collection for cell cultures and microorganisms. DSMZ strain numbers are given in parenthesis. Class Strain or species (DSM No.) Acc. No.
alpha-Proteobacteria a TL AY177716
(Roseobacter clade) T5 AY177712 T11 AY177714 TY AY841788 D1 AY841770 D4 AY841771 HP12 AY841769 HP14w AY841773 HP29w AY239008 HP30 AY239009 HP32 AY841774 HP37 AY239010 HP44 AY841765 HP47 AY841776 HP50 AY841778 ROS2 AY841779 ROS4 AY841780 ROS7 AY841781 ROS8 AY841782 AP-27 AY145564 H43-35 AY841784 H55 AY841765 GWS-BW-H22M AY515407 GWS-BW-H66M AY515422 GWS-BW-H71M AY515423 Roseobacter gallaeciensis (12440) Y13244 Roseobacter denitrificans (7001) M59063 Ruegeria algicola (10251) X78315 Ruegeria gelatinovorans (5887) D88523 Roseovarius tolerans (11457) Y11551 Leisingera methylohalidivorans (14336) AY005463 Sulfitobacter pontiacus (10014) Y13155

Kapitel II Effects of a phytoplankton bloom on bacterial communities
40
Table 2 cont. Class Strain or species (DSM No.) Acc. No.
alpha-Proteobacteria b Paracoccus aminophilus (8538) Y16929
beta-Proteobacteria b Aquaspirillum delicatum (11558) AF078756
Burkholderia pyrrocinia (10685) AB021369 Sphaerotilus natans (6575) L33980 gamma-Proteobacteria
b Pseudomonas putida (548) AF094741
Pseudeoalteromonas atlantica (6839) X82134 Fundibacter jadensis (12178) AJ001150 delta-Proteobacteria
b Desulfococcus multivorans (2059) AF418173
Desulfobulbus mediterraneus (13871) AF354663 Pelobacter venetianus (2394) U41562 Flavobacteria
b Muricauda ruestringensis (13258) AF218782
Bacilli b Bacillus marinus (1297) AJ237708
Bacillus subtilis (7, 10) AJ276351 Lactobacillus plantarum (20205) n. a. Actinobacteria
b Streptomyces violaceoruber (40701) n. a.
Streptomyces glaucescens (40155) D44092 Streptomyces antibioticus (40715) n. a. Arthrobacter nicotinovorans (420) X80743 a positive control b negative control n. a. : not available

Kapitel II Effects of a phytoplankton bloom on bacterial communities
41
Figure legends
Fig. 1. Temperature and salinity (A), total particulate carbon, particulate organic carbon
(POC) and phytoplankton carbon as % of POC (B), phytoplankton cell counts and species
composition (C), and abundance of particle-attached and free-living bacteria (D) in the
German Wadden Sea from 12 April to 29 June 2000.
Fig. 2. DGGE fingerprints of the free-living (FL) and particle-attached (PA) bacterial
communities of the German Wadden Sea from 12 April to 29 June 2000 using primer sets for
16S rRNA genes of Bacteria (EUB; A), Bacteroidetes (CFB; B), alpha-Proteobacteria (ALF;
C) and the Roseobacter clade (ROS; D). The numbered arrows mark excised and sequenced
bands. Because of the small fragment size of the Roseobacter amplicons the DGGE bands
were not excised for sequencing.
Fig. 3. Cluster analyses of the DGGE banding patterns of particle-attached (PA) and free-
living (FL) Bacteria (EUB; A), Bacteroidetes (CFB; B), alpha-Proteobacteria (ALF; C) and
the Roseobacter clade (ROS; D) using UPGMA. The similarity matrix was calculated using
Pearson correlation.
Fig. 4. Phylogenetic trees of Proteobacteria (A) and the Bacteroidetes phylum (B) calculated
with Maximum-Likelihood based on 16S rRNA gene fragments. Sequences obtained in this
study are highlighted in bold.
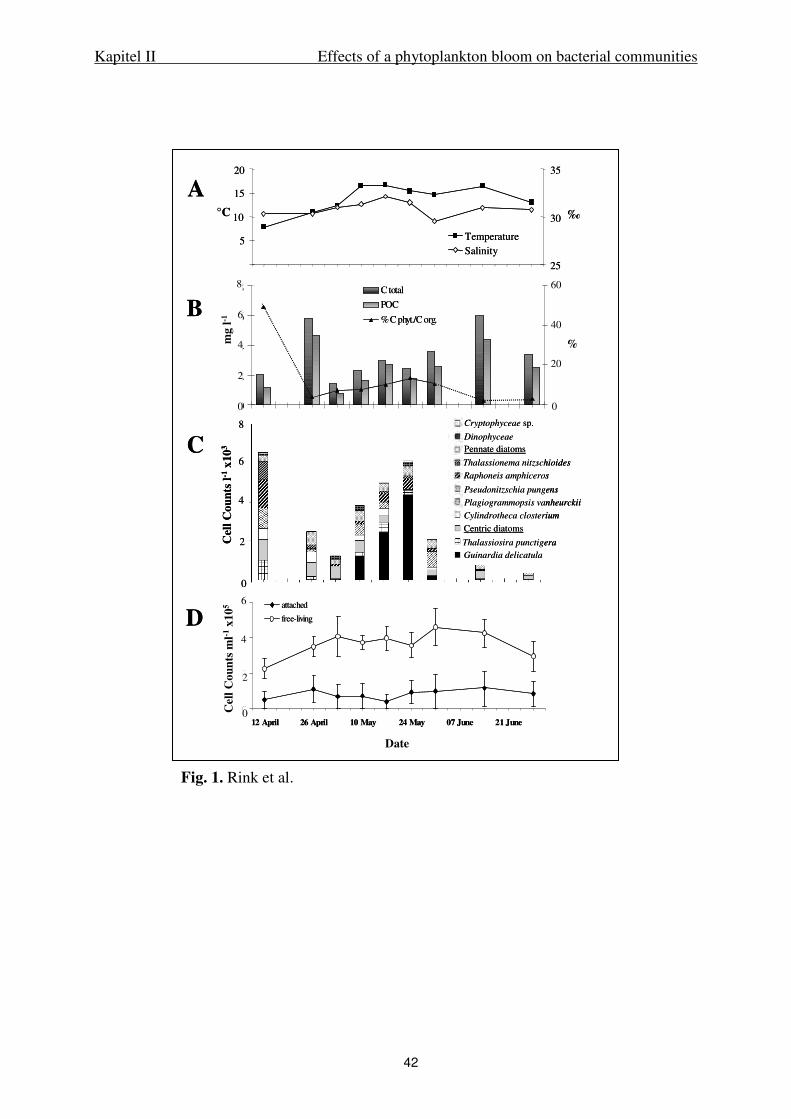
Kapitel II Effects of a phytoplankton bloom on bacterial communities
42
0
2
4
6
12 April 26 April 10 May 24 May 07 June 21 June
Date
Cel
l C
oun
ts m
l-1
*106
attached
free-living
0
2
4
6
8
12 April 26 April 10 May 24 May 07 June 21 June
mg
l-1
0
20
40
60
%
C total
POC
% C phyt./C org.
A
B
C
D
Cel
lCou
nts
l-1x1
03
5
10
15
20
12 April 26 April 10 May 24 May 07 June 21 June
°C
25
30
35
‰
TemperatureSalinity
Cel
lCou
nts
ml-1
x105
Date
mg
l-1
8
6
4
2
%
60
40
20
0
Cryptophyceae sp.
Dinophyceae
Pennate diatoms
Thalassionema nitzschioides
Raphoneis amphiceros
Pseudonitzschia pungens
Plagiogrammopsis vanheurckii
Cylindrotheca closterium
Centric diatoms
Thalassiosira punctigera
Guinardia delicatula
8
6
4
2
6
4
2
0
0
00
2
4
6
12 April 26 April 10 May 24 May 07 June 21 June
Date
Cel
l C
oun
ts m
l-1
*106
attached
free-living
0
2
4
6
8
12 April 26 April 10 May 24 May 07 June 21 June
mg
l-1
0
20
40
60
%
C total
POC
% C phyt./C org.
A
B
C
D
Cel
lCou
nts
l-1x1
03
5
10
15
20
12 April 26 April 10 May 24 May 07 June 21 June
°C
25
30
35
‰
TemperatureSalinity
Cel
lCou
nts
ml-1
x105
Date
mg
l-1
8
6
4
2
%
60
40
20
0
Cryptophyceae sp.
Dinophyceae
Pennate diatoms
Thalassionema nitzschioides
Raphoneis amphiceros
Pseudonitzschia pungens
Plagiogrammopsis vanheurckii
Cylindrotheca closterium
Centric diatoms
Thalassiosira punctigera
Guinardia delicatula
Cryptophyceae sp.
Dinophyceae
Pennate diatoms
Thalassionema nitzschioides
Raphoneis amphiceros
Pseudonitzschia pungens
Plagiogrammopsis vanheurckii
Cylindrotheca closterium
Centric diatoms
Thalassiosira punctigera
Guinardia delicatula
8
6
4
2
6
4
2
0
0
0
Fig. 1. Rink et al.
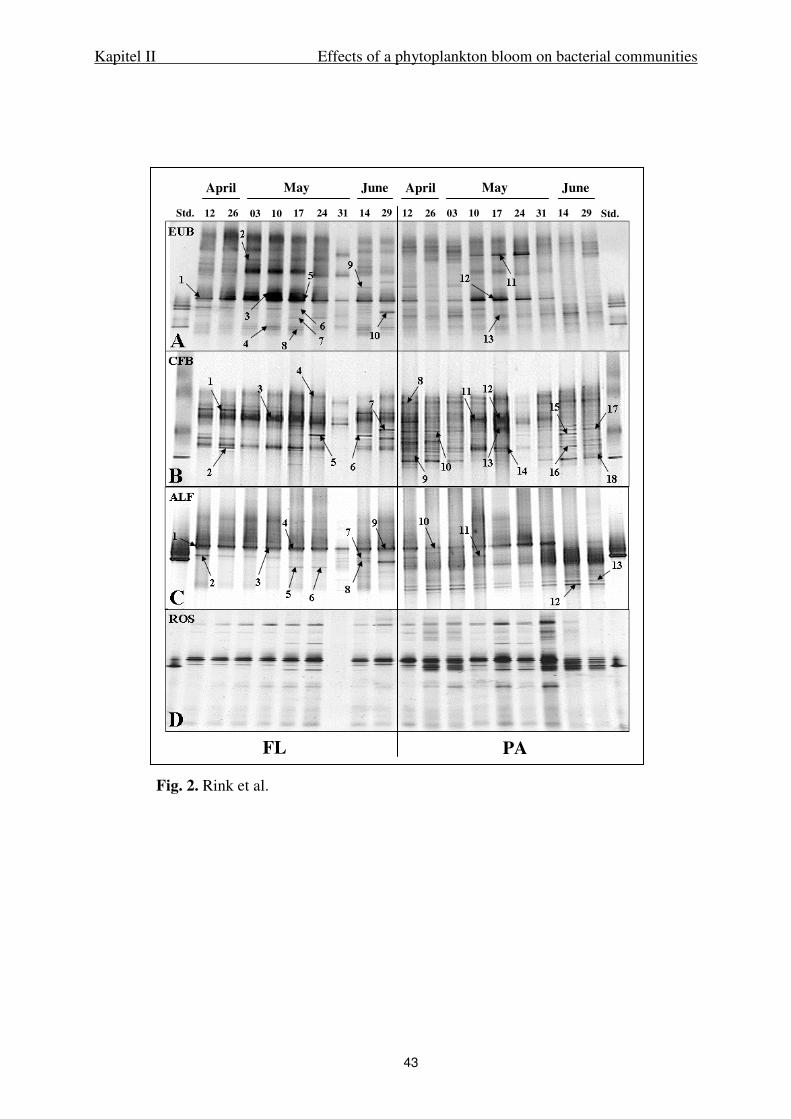
Kapitel II Effects of a phytoplankton bloom on bacterial communities
43
April May June April May June
12 26 03 10 17 24 31 14 29 12 26 03 10 17 24 31 14 29Std. Std.
EUB
CFB
ALF
ROS
A
B
C
D
FL PA
Fig. 2. Rink et al.
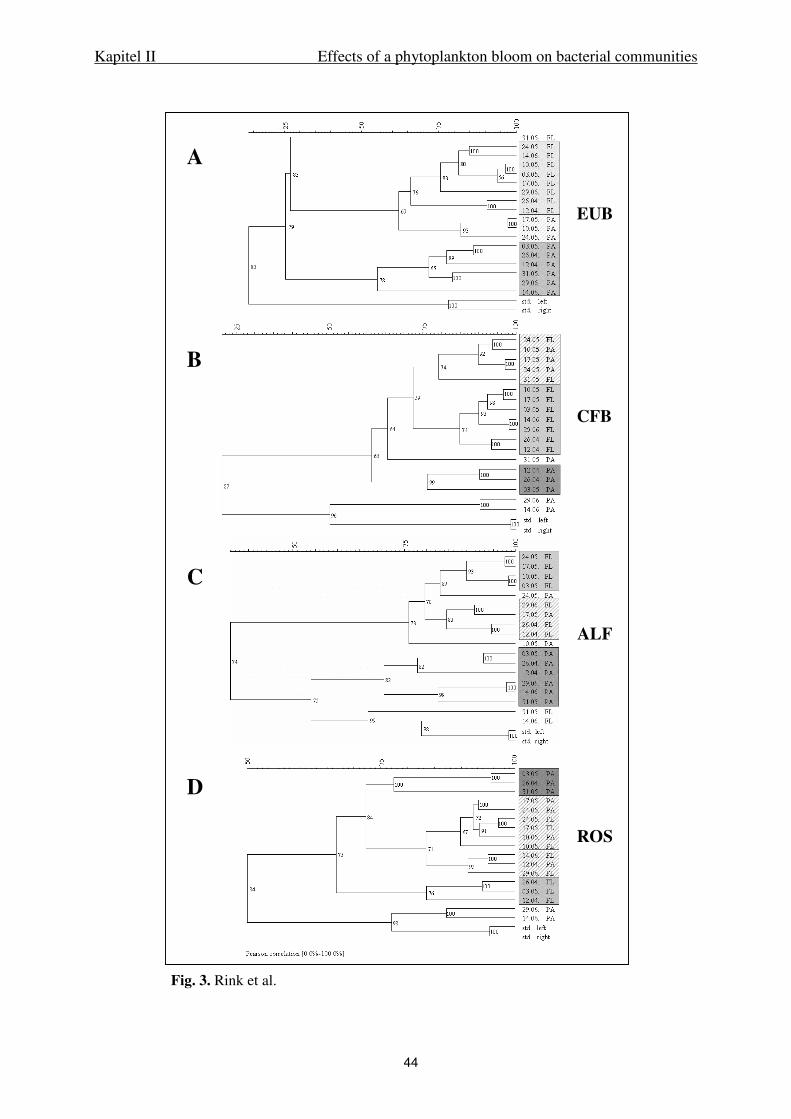
Kapitel II Effects of a phytoplankton bloom on bacterial communities
44
Fig. 3. Rink et al.
ROS
ALF
CFB
EUB
A
B
C
D

Kapitel II Effects of a phytoplankton bloom on bacterial communities
45
Fig. 4A. Rink et al.
A
gamma
beta
RCA
WAC I
alpha
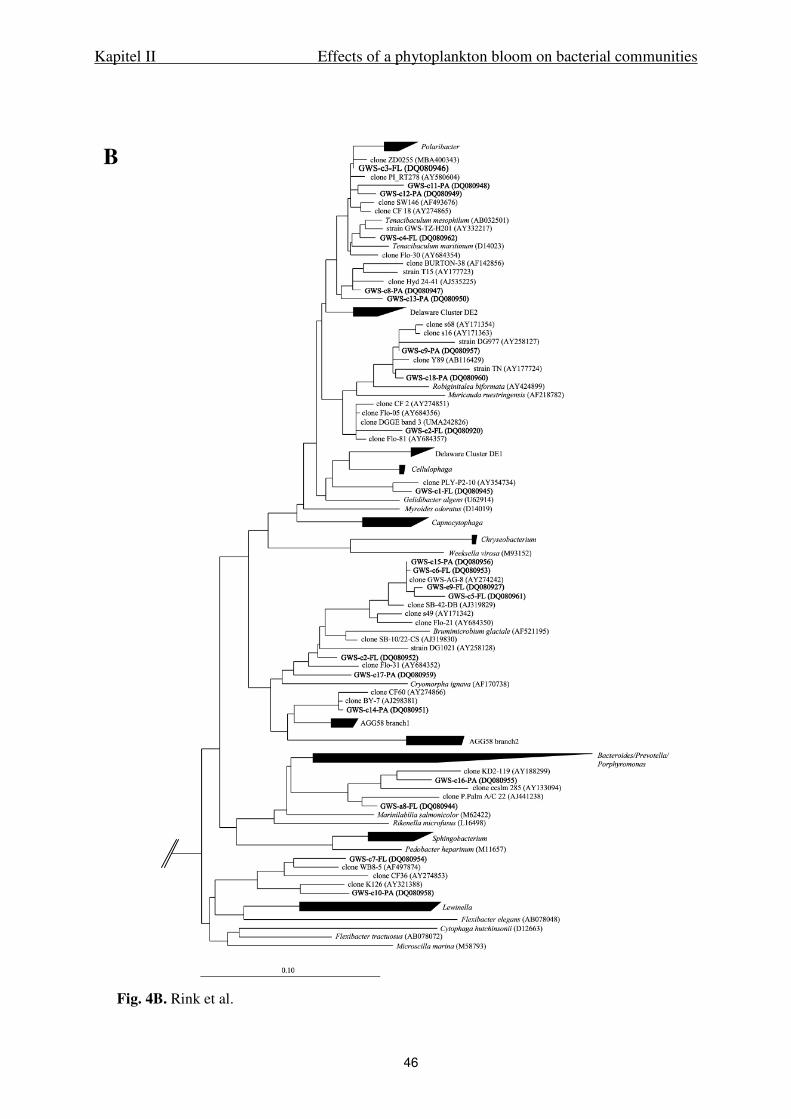
Kapitel II Effects of a phytoplankton bloom on bacterial communities
46
B
Fig. 4B. Rink et al.

47
III.
Tidal effects on coastal bacterioplankton

Kapitel III Tidal effects on coastal bacterioplankton
48
Tidal effects on coastal bacterioplankton
Beate Rink, Torben Martens, Doreen Fischer, Andreas Lemke, Hans-Peter Grossart, 1
Meinhard Simon, and Thorsten Brinkhoff *
Institute for Chemistry and Biology of the Marine Environment (ICBM),
University of Oldenburg, P.O. Box 2503, D-26111 Oldenburg, Germany
1 Present address: Institute of Freshwater Ecology and Inland Fisheries, Department of
Limnology of Stratified Lakes, Alte Fischerhuette 2, D-16775 Stechlin, Germany
Running head: Tidal effects on coastal bacterioplankton
Key words: Aggregates, bacteria, CARD-FISH, DGGE, tidal flat, Wadden Sea
__________________________________________________________________________
* To whom correspondence should be addressed ([email protected]).

Kapitel III Tidal effects on coastal bacterioplankton
49
Acknowledgements
We appreciate the hospitality and assistance of the RV Senckenberg crew. We gratefully
acknowledge K. Ishii and R. Amann for the introduction of the CARD-FISH method and M.
Mußmann for valuable methodical discussions. We thank B. Kuerzel and R. Weinert for Chl
a and dry weight analyses and R. Reuter and T. Badewien for the supply of temperature and
salinity data for July 2005. This work was supported by the Deutsche
Forschungsgemeinschaft (DFG) within the research group “BioGeoChemistry of Tidal Flats”
(FG 432-5).
Abstract
Tidal flats are highly dynamic and productive ecosystems, strongly influenced by
hydrodynamic forces and tidal events. We examined the impact of the tide on the composition
of free-living (FL) and particle-attached (PA) bacterioplankton in the Wadden Sea, southern
North Sea, in November 1999, May 2000 and July 2005 complementing the study of Grossart
et al. (2004) which focused mainly on the dynamics of suspended particulate matter (SPM)
and bacterial abundance. FISH, CARD-FISH and DGGE fingerprints with primer sets for
Bacteria, Bacteroidetes, alpha-Proteobacteria and the Roseobacter Clade based on 16S rRNA
genes and 16S rRNA were applied. In addition, suspended particulate matter (SPM),
particulate organic carbon (POC), chlorophyll a (Chl a), bacterial cell counts and bacterial
protein production rates (BPP) were measured. The introduction of North Sea water in the
Wadden Sea system was shown by changes of the water temperature, salinity values and
oxygen saturation during high tides (HT). In spring and summer, particulate organic carbon
(POC) ratio and cell specific bacterial production was increased and altered with the tide.
Surprisingly, these strong variations had only slight effect on the composition of the bacterial
communities. FISH counts revealed abundance of Bacteroidetes, alpha- and gamma-
Proteobacteria. On particles, also beta-Proteobacteria were detected with 15.29% DAPI in
May 2000 and 8.19% DAPI in July 2005. The Roseobacter Clade constituted almost all FL
but only one third of the PA alpha-Proteobacteria. In spring and summer, FISH counts
showed higher variations within the bacterial groups compared to autumn and increased
abundances of PA alpha-Proteobacteria. A clear relationship between these variations and the
tide was not found. DGGE banding patterns of 16S rRNA gene fragments were highly stable
even on the group-specific level. At higher resolution based on cDNA amplicons, higher
richness was detected but only few variations related to the tide appeared. Phylogenetic
analysis revealed that prominent bands were affiliated to the RCA, the WAC I and the

Kapitel III Tidal effects on coastal bacterioplankton
50
SAMMIC cluster. These clusters were found in previous studies of the Wadden Sea and are
distributed worldwide in temperate and polar marine regions. Our results suggest that the
stability of the abundant bacterial groups is a consequence of the strong changes within the
Wadden Sea ecosystem selecting highly adapted species which persist even on a long-term
scale.
Introduction
Tidal flats are coastal ecosystems appearing in temperate and tropical regions and are
composed of sandy or muddy sediment. They are strongly exposed to the tide and thus form a
very stressful environment for the indwelling organisms. Furthermore, tidal flats provide
complex substrates due to high loads of suspended particulate matter (SPM) deriving from the
land and the sea, and belong to the most productive ecosystems in the world (Dittmann,
1999). The German Wadden Sea, a tidal flat system in the southern part of the North Sea, is
the largest tidal flat system worldwide comprising an area of 7500 km2. Several monitoring
studies examined the status of this ecosystem on spatial, diurnal and seasonal scales. It is a
well studied area in terms of suspended particulate matter flux and particle size distribution
(Eisma and Li 1993; van Leussen 1996; Behrends und Liebezeit 1999; Mikkelsen 1998;
Fugate and Friedrichs 2003; Grossart et al. 2004; Lunau et al. 2006), primary production (van
Duyl et al. 1999; Tillmann et al. 2000; Niesel and Günther 1999) and several planktonic and
benthic organisms (Günther, 1999; Dittmann, 1999), but the main mediators for organic
matter decomposition, the bacterial communities, remained disregarded.
Correlation of SPM composition and -variation and microbial activities in the Wadden Sea
were investigated by Grossart et al. (2004) during two tidal cycles in November 1999 and
May 2000. Strong changes in several parameters were detected due to differences in tidal
currents and subsequent introduction of North Sea water masses into the Wadden Sea
ecosystem at high tides (HT). In November 1999, water temperatures and salinity were higher
during HT. SPM and PA bacteria were correlated and fluctuated during the tidal cycle
dependent from current velocities. In May 2000, water temperatures were higher but oxygen
saturation was increased during HT due to the North Sea water influence. Cell specific
bacterial production increased with rising tide indicating response of the bacterial community
to tidal changes. Also seasonal influences were detected particularly for the POC ratio and FL
cell counts which were increased in May 2000. These observations substantiated the
assumption that the tide may strongly affect the composition of pelagic bacterial
communities. So far, only few studies focused on short-term influences on bacterioplankton.

Kapitel III Tidal effects on coastal bacterioplankton
51
Pernthaler and Pernthaler (2005) described variations in the proliferation of North Sea
bacteria during a tidal cycle. Diurnal changes in pigment concentrations derived from bacteria
in Baltic Sea water were examined by Koblizek et al. (2005). In dilution cultures of North Sea
bacteria, the impact of UV on bacterial communities during a 24 hours period was
investigated using DGGE profiles (Winter et al., 2001).
In previous studies, the main interest of Wadden Sea investigations focused on the
composition and abundance of sediment bacteria. Llobet-Brossa et al. (1998) showed
dominance of Bacteroidetes and sulfate-reducing bacteria in the sediment surface layer. At
two different sampling sites of Wadden Sea sediment, Mussmann et al. (2005) isolated
several delta-Proteobacteria. In the water column, the Wadden Sea bacterioplankton was
recently examined on a qualitative level by Stevens et al. (2005a, b) and Rink et al. (2006)
using DGGE. It was demonstrated that many bands persisted over long-term periods, but also
variations in the banding patterns were detected in spring and summer months. Rink et al.
(2006) showed correlation of phytoplankton composition changes and alteration in the
bacterial community composition using group-specific primer sets. As these studies were
performed on a monthly or weekly scale, the influences of tidal currents and the fast exchange
of different water bodies in the Wadden Sea on the bacterioplankton remained unknown.
Thus, the aim of this study was to examine the impact of the tide on the composition and
abundance of the bacterial communities in the Wadden Sea with respect to seasonal aspects.
With this study, we complete the results by Grossart et al. (2004) with detailed bacterial
community composition analysis based on the same sampling campaigns and an additional
sampling in July 2005.

Kapitel III Tidal effects on coastal bacterioplankton
52
Materials and methods
Sample collection and processing– Samples were taken in November 1999, May 2000 and
July 2005 from shipboard in the major channel of the backbarrier tidal flat system of the
German Wadden Sea (53° 44.9´N, 07° 40.0´E). The sampling period was 19 hours in
November 1999, 22 hours in May 2000 and 12 hours in July 2005. In November 1999 and
May 2000, surface water was collected every hour and filtrated for suspended particulate
matter (SPM), chlorophyll a (Chl a), particulate organic carbon (POC) and bacterial cell
counts according to Grossart et al. (2004). In July 2005, samples for SPM, Chl a, POC and
bacterial cell counts were taken and processed according to Lunau et al. (2006). Hydrographic
data of July 2005 were measured by a CTD probe (SeaCat 19plus, Seabird, Washington,
USA).
For DGGE analyses, one hundred ml of seawater were filtered at high tides, mean tides
and low tides, fractionated onto polycarbonate-filters (diameter 47 mm, Nuclepore) and stored
at -20°C in the dark. A pore size of 5.0 µm was used to obtain particle-attached (PA) and 0.2
µm to obtain free-living (FL) bacteria. For Fluorescent In Situ Hybridization (FISH) and
Catalyzed Reporter Deposition (CARD)- FISH analysis, 2 - 4 ml were filtered onto 5.0 µm
polycarbonate-filters (diameter 25 mm, Nuclepore) and 2 ml of the filtrate were subsequently
filtered onto 0.2 µm polycarbonate-filters. After fixation with paraformaldehyde (4% w/v) for
one hour filters were stored at -20°C in the dark until further processing. Sediment cores in
November 1999 and May 2000 were taken with Plexiglas tubes (36 mm diameter) at low tide
on an intertidal mud flat about 200 m away from the ship. Two mm of the upper surface layer
were sliced, transferred into sterile caps and kept frozen at -20°C.
SPM dry weight, particulate organic carbon and chlorophyll a– Filters for SPM dry
weight were dried for 12 h at 60°C, adapted to room temperature and weighed on a
microbalance. Dry weight was calculated as the difference of filter weight before and after
filtration (500 ml filtration volume, glass fiber filters, GF/F, Whatman). For particulate
organic carbon, filters were exposed to hydrochloric acid fume for 12 h and subsequently
analyzed by a FlashEA 1112 CHN-analyzer (Thermo Finnigan). Chlorophyll a was
determined photometrically after extraction in hot ethanol according to Von Tuempling and
Friedrich (1999). For further details concerning SPM, POC and Chl a analyses see Lunau et
al. (2006).

Kapitel III Tidal effects on coastal bacterioplankton
53
Bacterial cell counts– Bacterial abundance was determined with SybrGreen I as described
by Lunau et al. (2006) with slight modifications. The detachment of particulate bacteria was
performed using methanol (30% v/v) and ultrasonic treatment for 15 min at 35°C. For the
enumeration of bacterial cells, subsamples of 1 ml were filtered onto black polycarbonate
filters (0.2 µm pore size, Poretics). Abundance of particulate bacteria was calculated as the
difference between total and free-living cell counts.
Bacterial Production –The bacterial production rate was estimated by the incorporation of 14C-Leucin (Simon and Azam 1989). Samples were incubated in triplicates with 14C-Leucin
(306 mCi/mmol, Hartmann Analytic, Germany) at a final concentration of 70 nmol l-1 to
ensure saturation of uptake systems. Formalin fixed water samples (2% v/v) were used as
controls. The samples were incubated in 5 ml plastic tubes in the dark at in situ temperature
for up to 1 hour. To avoid sedimentation, incubation was performed on a plankton wheel. The
incubation was linear for at least 1 hour and terminated by the addition of formaldehyde (2%
v/v). After fixation, the samples were filtered onto 0.45 µm nitrocellulose filters (Sartorius,
Germany) and extracted with ice-cold 5% trichloracetic acid (TCA) for 5 min. Subsequently,
filters were rinsed twice with 3 ml ice-cold TCA (5% v/v). To dissolve the filters, 4.5 ml of
scintillations cocktail was added. Vials were shaken vigorously and radioactivity was
determined afterwards in a scintillation counter. Standard deviation of triplicate measurement
was usually <15%.
FISH and CARD-FISH– FISH filters in November 1999 and May 2000 were rinsed with 1
ml phosphate buffered saline (PBS 1x) and dehydrated with 50%, 80% and 100% ethanol.
Subsequently, bacterial cells were hybridized for 5 h and washed 20 min at 46°C following
the protocol of Glöckner et al. (1996). Oligonucleotide probes for various phylogenetic
groups were used in November 1999: EUB 338 (Amann et al. 1990), ALF968, GAM42a
(Manz et al. 1992), CF319a (Cytophaga/Flavobacteria, Manz et al. 1996), ARCH915 (Stahl &
Amann 1991) and SRB385 (Amann et al. 1990). In May 2000, the additional probes BET42a
(Manz et al. 1992) and SRB385db (Rabus et al. 1996) were used. All filters were
counterstained with DAPI (4´,6´-Diamidino-2-phenylindol, 1µg/ml). In July 2005,
hybridizations were performed using the CARD-FISH method following the protocol of
Sekar et al. (2003). After fixation, samples were embedded in agarose (0.2%) and treated with
lysozyme (10mg/ml). Hybridization conditions were 3 h of hybridization at 35°C, 10 min
washing at 37°C and 30 min. amplification at 37°C. The following oligonucleotide probes

Kapitel III Tidal effects on coastal bacterioplankton
54
labeled with horseradish-peroxidase (HRP) were used for CARD-FISH: EUB338, ALF968,
BET42a, GAM42a, ROS536 (Brinkmeyer et al. 2000) and NON338 (Wallner et al. 1993).
Tyramine-HCl was labeled with Fluorescein-5-isothiocyanate (FITC) as described by
Pernthaler et al. (2002). To avoid unspecific accumulation of dye in the cells, the last washing
step in PBS (1x) amended with TritonX-100 (0.05%) was extended to 30 min.
Counterstaining was performed with Vectashield-mounting medium with DAPI (1.5µg/ml;
Vector Laboratories, Peterborough, England).
Nucleic acid extraction– For samples taken in November 1999 and May 2000, genomic
DNA was isolated by phenol-chloroform extraction after bead beating as described by Selje
and Simon (2003). In July 2005, DNA and RNA were co-extracted from the same filter using
phenol-chloroform calibrated with sodium acetate RNA-buffer (50 mM, pH 4.2) containing
EDTA (10 mM) and Polyvinylpolypyrrolidone (PVPP, 2 g l-1). All steps were done under
sterile conditions using Diethylpyrocarbonate (DEPC, 0,1%)-treated ingredients. After
precipitation in isopropanol at -20°C overnight nucleic acids were resuspended in
DNase/RNase-free molecular grade water (Eppendorf, Germany). The RNA was incubated
with DNase I (5 U ml-1) for 1 h at 37°C and precipitated as described above. DNase digestion
was repeated until no DNA contamination could be detected by PCR with primers specific for
bacterial 16S rRNA genes. Samples were stored at -20°C until further processing.
PCR amplification – 16S rRNA gene fragments for subsequent DGGE analyses were
amplified using the primer pair GC 341F and 907RM (Muyzer et al. 1998) in an Eppendorf
Mastercycler (Eppendorf, Hamburg, Germany). In July 2005, the additional primer pairs GC
CF319aF and 907RM, GC 341F and ALF968R, and GC ROS 536F and GRB 735R were used
to investigate the bacterial community composition on a group-specific level. Primer
sequences and PCR conditions were described earlier by Rink et al. (2006). Four µl of the
amplification products were analyzed by electrophoresis in 2% (w/v) agarose gels and stained
with ethidium bromide (1 µg ml-1). For sequencing analysis, PCR products were purified by
using the E.Z.N.A. Microspin Cycle-Pure Kit (Peqlab Biotechnologie GmbH, Erlangen,
Germany) following the instruction manual. To amplify bacterial RNA, the Qiagen RT-PCR
kit (Qiagen, Hilden, Germany) was used following the instruction manual for reverse
transcription of the RNA. Reverse transcription was done directly with bacteria- or group-
specific primer sets as described above. Subsequent PCR was performed in the same reagent
mix directly after transcription using specific conditions for the applied primer set.

Kapitel III Tidal effects on coastal bacterioplankton
55
DGGE analysis of PCR products– DGGE was performed with the INGENY phorU System
(INGENY International BV, Goes, Netherlands) using specific conditions for the applied
primer sets according to Rink et al. (2006). Gels were stained with SYBR Gold (Molecular
Probes, Inc.) after electrophoresis and documented digitally using a BioDoc Analyze
Transilluminator (Biometra, Göttingen, Germany). Bands were excised with a sterile scalpel,
suspended in 50 µl of molecular grade water (Eppendorf, Germany) and stored at –20°C until
reamplification.
Cloning– DGGE bands GWS-TC-a2-PA, GWS-TC-a6-FL, GWS-TC-c4-PA, GWS-TC-e9-
FL, GWS-TC-e11-FL, GWS-TC-e3-SE, GWS-TC-e4-PA, GWS-TC-e1-SE and GWS-TC-e2-
SE were cloned using the pGEM®-T Vector System II (Promega, Madison, USA) following
the instruction manual. Clones with inserts were picked, resuspended in molecular grade
water (Eppendorf, Germany) and screened by DGGE to check if the insert position matches
the position of the corresponding DGGE band. Adequate clones were amplified and
subsequently sequenced using the primers pUC/M13f and pUC/M13r (Messing 1983).
Sequencing and phylogenetic analysis – PCR products were sequenced using the
DYEnamic Direct cycle sequencing kit (Amersham Life Science, Inc.) and a Model 4200
Automated DNA Sequencer (LI-COR, Inc.) using GM8R (5´-TGGGTATCTAATCCT-3´) as
sequencing primer labeled with IRDyeTM800. In addition, DGGE bands were sequenced by
GeneArt (Regensburg, Germany) using the primer 907RM to enhance the sequence quality by
repeat determination. For all sequences, at least 400 bp were determined.
Construction of the phylogenetic trees was performed using the ARB software package
(http:/www.arb-home.de). Calculation of the backbone trees was done with the maximum
likelihood method using sequences of type strains of the selected phylogenetic groups with a
minimum sequence length of 1300 bp. To avoid uncertain alignments, positions at which less
than 50% of all sequences showed the same residues were excluded. Sequences with less than
1300 bp were added to the backbone tree using the maximum parsimony method and the
same filter. Five type strains belonging to Cyanobacteria were used as outgroup.
Nucleotide sequence accession number –The sequences obtained in this study are available
from GenBank under accession no. DQ911822 to DQ911842.

Kapitel III Tidal effects on coastal bacterioplankton
56
Statistics – Cluster analyses of DGGE banding patterns were performed using Gel Compar
II, version 2.5 (Applied maths, Kortrijk, Belgium). Calculations were done curve based using
Pearson correlation and UPGMA.
Results
Hydrographic data–In July 2005 water temperature ranged between 19.1 and 20.1°C
showing slight decrease at HT with incoming North Sea water. Salinity was almost constant
around 32 psu (Fig. 1a).
For hydrographic data as well as results concerning SPM dry weight, POC, Chl a, bacterial
abundance and bacterial production from November 1999 and May 2000 see Grossart et al.
(2004).
SPM dry weight, POC and Chl a–Dry weight of suspended particulate matter in July 2005
ranged from 7.5 to 8.2 mg l-1 at HT and reached the highest value of 36.8 mg l-1 at low tide
(Fig. 1b). Despite highest current values at mean tide, SPM dry weight was average (21 to
29.2 mg l-1). POC values (see supplementary data) were low around LT (0.39 mg l-1) and
increased towards HT (up to 2.15 mg l-1) and constituted 1.1% (LT) to 28.7% (HT) of the
SPM dry weight. The average value was 7.6% dry weight. Chlorophyll a increased from 3.13
µg l-1 at HT to 6.7 µg l-1 around LT and decreased again to 3.76 µg l-1 at high tide (Fig. 1c).
Phaeopigments showed the same tidal dynamic and ranged from 0.30 to 5.12 µg l-1 with the
highest ratio phaeopigments/chlorophyll a of 1.18 at mean tide.
Bacterial abundance and bacterial production– In July 2005, total bacterial cell counts
varied between 1.28 to 3.56 x 106 cells ml-1 (Fig 1d). Free-living (FL) bacteria showed a
minimum of 0.97 x 106 cells ml-1 one hour after the first high tide and increased slightly to a
maximum of 2.30 x 106 cells ml-1 at one hour after mean tide 1 (MT 1). Afterwards, FL cell
numbers decreased again until next mean tide (MT2) and then ranged between 1.46 and 1.75
x 106 cells ml-1 until second high tide. Particle-attached (PA) bacteria were lowest around
high tide with 0.09 to 0.45 x 106 cells ml-1 and highest between MT1 to MT2 (0.96 to 1.33 x
106 cells ml-1) with a slight decrease around LT. In average, PA bacteria accounted for 28% of
total bacterial cell counts. To compare cell densities of PA bacteria, cell counts per mg dry
weight were calculated (see supplementary data). Density of PA cells was highest one hour
after MT and two hours after LT due to lower dry weight values and equal cell counts.

Kapitel III Tidal effects on coastal bacterioplankton
57
Bacterial production (BPP, Fig. 1e) in July 2005 ranged between 0.55 µg l-1 h-1 at both
high tides to 3.33 to 3.64 µg l-1 h-1 during LT and MT2.
FISH counts – FISH counts detected with probe EUB338 were lowest in May 2000 with a
mean value of 56.77% DAPI on particles and 48.42% DAPI in the FL fraction (Table 1, Fig.
2). Highest FISH counts were measured in November 1999 with 73.8% DAPI on particles and
61.24% DAPI in the FL fraction. In July 2005, FISH counts were similar to November with
70.42% DAPI on particles and 59.44% DAPI for free-living bacteria. Highest variation within
the cell counts were detected in May 2000 during a spring phytoplankton bloom, while lowest
variation was observed in November 1999 when hydrodynamic forcing controlled the
processes in the water column (Fig. 2). Generally, a higher percentage (% DAPI) of cells
could be detected on particles compared to the free-living fraction.
Specific probes revealed high numbers of Bacteroidetes, alpha-, beta- and gamma-
Proteobacteria in the Wadden Sea (Table 1, Fig. 2). In November 1999 and May 2000, also
low cell counts of Archaea and sulphate-reducing bacteria were detected (below 5% each).
In May 2000, cell counts of Bacteroidetes were 27.59% DAPI on particles and 13.55%
DAPI in the FL fraction. In November 1999, Bacteroidetes showed lower FISH counts on
particles 20.93% DAPI, but higher numbers (19.16% DAPI) in the FL fraction (mean values).
In July 2005, lowest cell counts of Bacteroidetes were detected with 15.02% DAPI in the PA
fraction and 11.52% DAPI in the FL fraction.
Abundances of alpha-Proteobacteria on particles were lowest in November 1999 with
9.53% DAPI and highest in May 2000 with 30.59% DAPI. In the free-living fraction, alpha-
Proteobacteria contributed 13.76% of the total cell counts in November 1999 but only 7.21 to
7.87% DAPI in spring and summer. In July 2005, the Roseobacter Clade, as part of the alpha-
Proteobacteria, represented 4.34% DAPI on particles and 5.09% DAPI in the FL fraction
contributing nearly 25% of the PA and 71% of the FL alpha-Proteobacteria.
Abundances of gamma-Proteobacteria ranged between 16.49 and 28.83% DAPI on
particles and between 9.83 and 17.12% DAPI in the FL fraction. Hybridization with probe
BET42a revealed 8.19 to 15.29% DAPI counts on particles in spring and summer showing a
certain abundance of PA beta-Proteobacteria in this coastal environment.
DGGE banding patterns and Cluster analysis–DGGE analysis of the tidal cycles in
November 1999, May 2000, and July 2005 revealed distinct bacterial populations for the
sediment surface, the PA and the FL bacterial communities (Fig. 3, 4, 5). In November 1999

Kapitel III Tidal effects on coastal bacterioplankton
58
and May 2000, fingerprints within the different fractions showed almost identical banding
patterns during the tidal cycles (data not shown). Highest band numbers in all seasons were
determined in the PA fraction (24 bands in Nov 1999, 13 bands in May 2000, 25 bands in July
2005) and a direct comparison of November 1999 and May 2000 showed only few variations
in the banding patterns at all (Fig. 3). The banding patterns of the PA bacterial communities
indicate an overlap with those resulted from sediment and the FL bacteria, as both fractions
show bands on the same height as in the PA fraction.
In July 2005, the FL and PA bacterial communities were also distinct as shown by the
DGGE fingerprints and cluster analysis. In addition, high stability of the communities was
observed on the DNA level, however, some bands appeared only in one or two samples, i. e.
at certain times of the tidal cycle (Fig. 4). Analysis with the bacteria specific primers revealed
for the PA fraction, e. g., that samples obtained during the first mean tide (MT1) and LT
showed one additional band (GWS-TC-e6-PA), which probably led to higher similarity of
these two samples in the cluster analysis (Fig. 5). In the FL fraction, one band appeared
exclusively at the first HT (GWS-TC-e8-FL).
The cDNA fingerprints of Bacteria revealed a slightly higher richness for the FL fraction
with up to 23 bands at HT compared to a maximum of 18 bands in the DNA banding patterns.
At HT, two additional bands appeared in the FL fraction which were neither detected in the
DNA fingerprints nor in the other cDNA samples (GWS-TC-e10-FL and GWS-TC-e11-FL).
On the group-specific level, DGGE analysis of Bacteroidetes showed stable DNA based
banding patterns in the PA fraction (Fig. 4, CFB). In the FL fraction, one prominent band
appeared exclusively at MT1 and LT (GWS-TC-c1-FL). The cDNA banding patterns showed
few differences compared to the DNA fingerprints and during the tidal cycle as well. In the
PA fraction, bands no. 2, 3, and 4 (GWS-TC-c2-PA, GWS-TC-c3-PA and GWS-TC-c4-PA)
occurred individually at different times and did not appear in the DNA based banding
patterns. Cluster analysis of the Bacteroidetes fingerprints revealed distinct fractions of PA
und FL fractions except for the second mean tide (MT2) of the cDNA FL fraction, which
showed higher similarity to the PA cluster (Fig. 5). The FL cluster was divided in two
subclusters of DNA and cDNA samples showing high similarity within DNA samples (>90%
Pearson correlation). The PA cluster was also subdivided into DNA and cDNA clusters
except the DNA sample at LT which fell into the cDNA group.
DGGE analysis of the alpha-Proteobacteria showed low richness within the DNA
fingerprints with 14 bands in the PA fraction and 8 bands in the FL fraction (Fig. 4). Slight
differences during the tidal cycle were detected on particles at LT with decreased band

Kapitel III Tidal effects on coastal bacterioplankton
59
intensity of bands at the standard height, also reflected by cluster analysis. The cDNA
banding patterns showed higher richness in both fractions compared to the DNA level. A
maximum of 25 bands was counted at LT on particles and 11 bands were visible at HT in the
FL fraction. These differences between DNA and cDNA resulted in distinct clusters as shown
by cluster analysis for both, PA and FL fraction (Fig. 5). Tidal differences traced back to
alpha-Proteobacteria were detected in the cDNA fingerprints during MT1 and LT (Fig. 4,
GWS-TC-a2-PA, and GWS-TC-a5-PA) or generated by chloroplast phylotypes (GWS-TC-
a3-PA, GWS-TC-a4-PA).
For the Roseobacter Clade, DNA based fingerprints revealed a maximum of 14 bands in
the PA fraction and 11 bands in the FL fraction. The cDNA banding patterns showed
significantly higher richness in the PA fraction (20 bands at LT) compared to DNA
fingerprints, forming distinct clusters (Fig. 5). Slight differences during the tidal cycle were
also found for this group (Fig. 4). While the cluster analysis demonstrated high similarity of
the DNA and cDNA patterns of the FL bacterial communites, the PA samples of the
Roseobacter Clade clustered distinct in DNA and cDNA (Fig. 5).
Phylogenetic affiliation – Sequence analysis of DGGE bands revealed that most of the
prominent bands were affiliated to recently described clusters of Wadden Sea bacteria (WAC
I, Stevens et al. 2005b) or of worldwide distributed bacteria (RCA, Selje et al. 2004;
SAMMIC, Stevens et al. 2005b) as shown in Fig 6A. From the DGGE banding patterns of
Bacteria, phylotypes belonging to the alpha-, gamma- and beta-Proteobacteria were obtained
reflecting the FISH and CARD-FISH results. One phylotype of delta-Proteobacteria and two
sequences of Actinobacteria were detected in the sediment fractions of November and May
(Fig. 3 and 6, GWS-TC-e1-SE, GWS-TC-e2-SE, GWS-TC-e3-SE). In total, four bands were
identified as chloroplasts (Fig. 3: GWS-TC-e5-PA; Fig. 4: GWS-TC-e7-FL, GWS-TC-a3-PA,
GWS-TC-a4-PA). GWS-TC-e11-FL was distantly related to Acidocella aminolytica (Fig. 6A)
and was not related to any sequence found in the Wadden Sea or North Sea before. Two
sequences fell in the WAC I cluster (GWS-e6-FL, GWS-e7-FL), GWS-TC-e9-FL clustered
with the Wadden Sea clone GWS-FL-5 (Stevens et al. 2005a), three sequences belonged to
the RCA cluster (GWS-TC-e6-PA, GWS-e12-PA, GWS-e5-FL) and GWS-e3-FL was related
to clone NAC11-7 (Fig. 6 A).
Two phylotypes of DNA and cDNA fingerprints clustered together within the gamma-
Proteobacteria (GWS-TC-e8-FL, GWS-TC-e10-FL). They were affiliated to
Stenotrophomonas maltophilia and appeared solely during HT in the FL fraction. Two of the

Kapitel III Tidal effects on coastal bacterioplankton
60
sediment derived sequences (GWS-TC-e3-SE, delta-Proteobacteria; GWS-TC-e2-SE,
Actinobacteria) were affiliated to clones which were found in the North Frisian Wadden Sea
sediments (Sylt 19, clones Sylt 21, Fig. 6 A).
Bacteroidetes affiliated phylotypes were exclusively obtained with the specific primer set
(GWS-TC-c1-FL to GWS-TC-c4-PA) confirming the bias against this phylum by the use of
the Bacteria-specific primer set (Kirchman 2002). Within the Bacteroidetes phylum, GWS-
TC-c1-FL represented the most pronounced diurnal change in the FL fraction of the DNA
fingerprints. This phylotype was related to GWS-c14-PA which was detected in May 2000
with specific primer sets for Bacteroidetes (Rink et al. 2006). Both were affiliated to clone
CF60 (AY274866) derived from the Delaware estuary. GWS-TC-c2-PA was detected during
HT in the PA fraction of the cDNA fingerprints and the phylogenetic affiliation showed no
relationship to known Wadden Sea or North Sea organisms. Another cDNA phylotype, GWS-
TC-c3-PA, was related to Lutibacter litoralis, isolated from a tidal flat system in Korea (Choi
and Cho 2006). GWS-TC-c4-PA was related to two novel species, Krokinobacter genikus
(Khan et al. 2006), isolated from marine sediment in Japan, and Dokdonia donghaensis (Yoon
et al. 2005), isolated from seawater of the Korean East Sea.
The application of the ALF968r primer revealed two chloroplast sequences (GWS-TC-a3-
PA, GWS-TC-a4-PA), and three phylotypes of alpha-Proteobacteria. Within the alpha-
Proteobacteria, the cDNA derived phylotypes clustered with Silicibacter lacuscaerulensis
(GWS-TC-a2-PA) and Sphingomonas paucimobilis (GWS-TC-a5-PA).
Discussion
Grossart et al. (2004) demonstrated that bacterial dynamics in the Wadden Sea are
controlled by two major factors: resuspension of sediment and phytoplankton growth.
Another strong influence is the introduction of North Sea water in the tidal basin at high tides
(HT). In November 1999, sediment resuspension was the dominating process in the water
column (Grossart et al. 2004). SPM flux and changes in particulate carbon directly reflected
the tidal dynamics. In May 2000, a strong influence of a phytoplankton bloom was observed
resulting in higher Chl a concentrations and an increase of the POC ratio and free-living
bacterial cells. In July 2005 the Chl a concentrations were lower than during the
phytoplankton bloom in May but typical for summer months in the Wadden Sea (Lunau et al.,
2006). SPM values in July were much lower compared to November and May, probably due
to a lower resuspension rate and lower abundance of phytoplankton, respectively (see

Kapitel III Tidal effects on coastal bacterioplankton
61
Grossart et al. 2004). Bacterial abundance on particles was similar in all seasons following
SPM dynamics. Cell counts of free-living bacteria were lower in July than in May but similar
to November suggesting lower dissolved organic carbon concentrations in the water column
(Grossart et al. 2004). The bacterial production was lowest around HT contrary to May and
other investigations (Lunau et al. 2006). Bacterial production values of May showed not a
tidal, but a diurnal, pattern, with high rates during the day and reduced values at night
(Grossart et al. 2004).
Grossart et al. (2004) showed that free-living bacteria were influenced by SPM
concentrations, total carbon (TC) and particulate organic carbon in November 1999 and May
2000. The abundance of particulate bacteria correlated with SPM, Chl a, total carbon and
particulate organic carbon. Thus, we investigated the composition of the bacterial
communities in the Wadden Sea by FISH and DGGE expecting strong changes of the
populations following the tidal dynamics of these parameters. By contrast, the fluctuation of
particulate carbon, SPM, Chl a and bacterial cell counts was not reflected by changes within
the phylogenetic groups during tidal cycles in all investigated seasons as shown by the FISH
results. Even the application of the highly sensitive CARD-FISH method in July 2005 did not
show any systematic impact of the tide on the bacterial community composition. In all
seasons, alpha- and gamma-Proteobacteria as well as Bacteroidetes were most abundant in
both fractions. This is in line with other studies from the North Sea (Eilers et al. 2000) and the
Weser estuary (Selje et al. 2003). The seasonal comparison of the FISH results revealed very
low variation within the bacterial groups during the tidal cycle in November in contrast to
May and July (Fig. 2). Largest variations and highest group-specific FISH counts were
obtained in May 2000 on particles. The exceeding of the group-specific over the EUB338
counts indicated high activity of the PA bacteria, as the signal strength of directly
fluorochrome-labeled oligonucleotide probes depends on the ribosomal RNA content of the
cells (Schut 1994). The SPM in May was enriched with phytoplankton derived POC which
intensely stimulated bacterial degradation processes of the organic matter as suggested by the
decoupling of activity parameters and tidal dynamics (Grossart et al. 2004). This may be one
reason for the fact that despite large variations within the bacterial groups, no clear
relationship was found between the FISH counts and the tide.
In spring and summer, beta-Proteobacteria were also investigated and very high
abundances were detected on particles. In May 2000, they constituted even 15.29% of the
total PA bacterial community (mean value, Table 1). This is in line with FISH counts of a
study showing that this group constituted 6% of DAPI cell counts in the marine section of the

Kapitel III Tidal effects on coastal bacterioplankton
62
nearby Weser estuary (Selje et al. 2003) and findings of several other authors (Rappé et al.
2000; Beja et al. 2002). High abundance of beta- Proteobacteria in this habitat was supported
by detection of phylotype GWS-e8-FL in May 2000, which clustered with other phylotypes
derived from the Wadden Sea (GWS-e4-FL, Rink et al. 2006; GWS-FL-6, Stevens et al.
2005) and the Weser estuary (DC11-51-11, Selje et al. 2005). The latter phylotype was
obtained from a 10-5 dilution step on marine medium even suggesting abundance of beta-
Proteobacteria in this saline environment.
DGGE results demonstrated that the composition of the bacterial communities in
November, May and July showed almost no changes during the tidal cycles. Sequencing of
prominent bands revealed that most of these phylotypes were affiliated to previously
described phylogenetic clusters, e. g. the RCA and the WAC I cluster within the alpha-
Proteobacteria (Selje et al. 2004; Stevens et al. 2005b) and the SAMMIC cluster within the
gamma-Proteobacteria (Stevens et al. 2005b). Organisms affiliated with the RCA cluster are
globally distributed in temperate and polar regions. They live exclusively in marine
environments freely suspended in the water column. The SAMMIC cluster comprises
organisms living on surfaces in marine environments. Bacteria of the SAMMIC cluster were
permanently detected on particles and on the sediment surface in the Wadden Sea during a
seasonal study (Stevens et al. 2005a). In parallel, Bowman et al. (2005) described the same
cluster with phylotypes from polar and temperate marine sediments detecting abundances of
up to 4% of the total bacterial community by quantitative RealTime-PCR. The distribution
and abundance of WAC I affiliated organisms is not yet clarified, but phylotypes belonging to
this cluster were also present in the Wadden Sea during all seasons (Stevens et al. 2005a).
Thus, bacteria belonging to these phylogenetic groups are not affected by short-term changes
generated by the tide as confirmed by our results.
In contrast to these permanently present organisms, some phylotypes were not detected
during the whole tidal cycles but in single samples (Fig. 4). The application of group-specific
primer sets revealed additional bands present at specific points of time. Although the use of
specific primer sets enhances the resolution of microbial studies (Abell & Bowman 2005;
Gich et al. 2005; Rink et al. 2006) the bacterial communities were remarkably stable even on
the specific level indicating low influence of the tide on the investigated groups.
Most tidal changes were detected by the RNA approach showing significantly higher
richness compared to the DNA fingerprints. The rRNA content of metabolically active
bacteria is higher than in dormant cells (Poulsen et al. 1993) and correlates with bacterial
growth rates (Delong et al. 1989). Thus, rRNA is suggested as activity indicator and reflects

Kapitel III Tidal effects on coastal bacterioplankton
63
responses of bacterial communities to environmental changes more pronounced than the
rRNA gene. Higher sensitivity concerning species diversity and appearance of community
shifts have been reported before, using rRNA based fingerprinting methods for marine
bacterioplankton (Schäfer et al. 2001; Moeseneder et al. 1999), drinking water supply systems
(Eichler et al. 2006) and bacterial assemblages of mariculture biofilter systems (Cytryn et al.
2005). Our study provides the first RNA based insights into bacterial communities of the
Wadden Sea tidal flat system, showing that most of the cDNA derived phylotypes, which
occured exclusively at specific points of the sampling, were not closely related to previously
described Wadden Sea organisms. Within the alpha-Proteobacteria, two of these phylotypes
were affiliated to described species (GWS-TC-a5-PA and GWS-TC-a2-PA, Fig. 6A).
Phylotype GWS-TC-a6-FL, which appeared at rising tide (MT2) was related to clone ZD0117
derived from the North Sea during a phytoplankton bloom (Zubkov et al. 2002). Largest
differences between DNA and cDNA fingerprints were reflected by the Roseobacter specific
fingerprints in the PA fraction. The richness within the cDNA banding patterns showed
almost two-fold increase compared to DNA. These findings were reflected by distinct clusters
of DNA and cDNA as revealed by cluster analysis. Hence, our results suggest that
significantly more particle-attached organisms are active than previously indicated by DNA
based methods and that the additional application of RNA based fingerprinting methods on a
group-specific level is essential to detect small-scale processes within microbial communities.
Relationships between additionally appearing bands and tidally generated processes were
hardly detected. In July 2005, bands GWS-TC-e8-FL and GWS-TC-e10-FL were pronounced
at HT in the FL fraction of DNA and cDNA samples. At HT, increased salinity, %POC values
and lower water temperature indicated influence of water from the open North Sea. The two
above mentioned phylotypes were closely related to Stenotrophomonas maltophilia (Fig. 6A,
96-97% sequence similarity). The genus Stenotrophomonas is widespread in terrestrial and
limnic habitats, but was also found in different marine samples, several times associated with
algae, sponges or dinoflagellates (e. g. Hagstrom et al. 2000; Seibold et al. 2001; Sfanos et al.
2005). Seibold et al. (2001) found a phylotype of Stenotrophomonas associated with the
dinoflagellate Noctiluca scintillans obtained from plankton hauls taken at Helgoland Roads
(German Bight, North Sea). Thus it is possible that the additional DGGE bands we observed
in our study derived from a Stenotrophomonas sp. introduced to the Wadden Sea with the
incoming tide. The same probably holds true for band GWS-TC-e11-FL (Fig. 4), which
belongs to the SAR116 cluster (Giovannoni et al. 1990) and band GWS-TC-a6-FL, which
appeared at rising tide (MT2) and was related to a clone obtained from the open North Sea

Kapitel III Tidal effects on coastal bacterioplankton
64
during a phytoplankton bloom (Zubkov et al. 2002). Appearance of these phylotypes only at
HT or rising tide, however, indicates low significance of the organisms within the Wadden
Sea ecosystem.
Cunha et al. (2001) showed that the activity of marine bacteria exposed to brackish water
increased significantly while bacteria of the estuary were less active when exposed to marine
water. Thus, the detection of differences at HT or rising tide may also be a single response of
marine bacteria to high concentrations of suspended substrates in the Wadden Sea.
Band GWS-TC-e6-PA (RCA cluster) disappeared in July 2005 at HT and MT2 (rising
tide) in parallel to the incoming North Sea water, but only in the particle fraction. This effect
was also detected in May 2000, but not in November 1999 (data not shown). In spring and
summer, SPM concentrations were lower compared to November with inversed tidal
dynamics (Grossart et al., 2004; this study). In May and July the inflow of North Sea water
resulted in lower amounts of SPM at HT suggesting that the disappearance of this band may
be due to a dilution effect. This assumption is supported by data showing much lower
numbers of RCA related phylotypes in the open North Sea compared to the Wadden Sea (H.
Giebel, University of Oldenburg, unpublished results). As no corresponding bands were
visible in the cDNA pattern of the particle fraction (Fig. 4), the significance of this organism
for SPM degradation in the Wadden Sea is also questionable.
On the group-specific level, GWS-TC-c1-FL appeared at MT1 and LT in the FL fraction
of the Bacteroidetes in the DNA and cDNA banding patterns. Closest relative was clone
GWS-c14-PA found in the Wadden Sea during a phytoplankton bloom in May 2000 (Rink et
al. 2006). This indicates that the organism is resident and active in the Wadden Sea. The
ecological function, however, remains unknown as this organism was found on particles
(Rink et al. 2006) and free-living (this study) as well.
Several other bands appeared on the group specific level at MT1 and LT (Fig. 4: GWS-TC-
c3-PA, GWS-TC-c4-PA, GWS-TC-a2-PA, GWS-TC-a5-PA) what might be explained by
resuspension of material from the sediment surface but this remains speculation.
The fingerprints of the Roseobacter Clade, which constituted a high fraction within the
alpha-Proteobacteria (approx. 25% on particles and 71% in the FL fraction as shown by
CARD-FISH results), were most stable even on the RNA level. Despite significantly higher
richness on particles (see above), almost no changes in the community composition were
observed for this group during the tidal cycle. Thus, our results indicate that the combined
application of group-specific primer sets and DNA/RNA based fingerprinting is sufficient for
the reliable examination of bacterial communities, i. e. to detect small changes within the

Kapitel III Tidal effects on coastal bacterioplankton
65
bacterial communities. In the tidal flat ecosystem of the Wadden Sea, however, only very few
changes could be observed and thus connection of these data with processes generated by the
tide were only sparely visible.
Overall our results demonstrate that even application of very sensitive investigation
methods like CARD-FISH and rRNA based DGGE analyses with specific primer sets
revealed highly stable FL and PA bacterial communities, almost not influenced by strong tidal
effects of this dynamic ecosystem. Seasonal changes and phytoplankton blooms may result in
larger tidal variations of group-specific abundances and the appearance of additional
organisms on a short time-scale but do not seem to influence the composition of the bacterial
communities significantly (Stevens et al. 2005a; Rink et al. 2006). Thus, we conclude that the
recurrent short- and long-term changes in the Wadden Sea resulted in the selection of highly
adapted organisms and lead to an exceptional stability of the bacterial communities in this
ecosystem. Further investigations are now required to clarify how the abundant bacteria are
involved in the microbial degradation processes and what makes them superior to other
organisms.

Kapitel III Tidal effects on coastal bacterioplankton
66
References
ABELL, G. C., AND J. P. BOWMAN. 2005. Ecological and biogeographic relationships of class Flavobacteria in the Southern ocean. FEMS Microbiol. Ecol. 51: 265-77.
AMANN, R. I., B. J. BINDER, R. J. OLSON, S. W. CHISHOLM, R. DEVEREUX, AND D. A. STAHL. 1990. Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. Appl. Environ. Microbiol. 56: 1919-1925.
BEHRENDS, B. AND G. LIEBEZEIT. 1999. Particulate amino acids in Wadden Sea waters – seasonal and tidal variations. J. Sea. Res. 41: 141-148.
BEJA, O., AND OTHERS. 2002. Unsuspected diversity among marine aerobic anoxygenic phototrophs. Nature 415: 630-633
BOWMAN, J. P., S. A. MCCAMMON, AND A. L. DANN. 2005. Biogeographic and quantitative analyses of abundant uncultivated γ-proteobacterial clades from marine sediment. Microb. Ecol. 49:451-460.
BRINKMEYER, R., M. RAPPE, S. GALLACHER, AND L. MEDLIN. 2000. Development of clade- (Roseobacter and Alteromonas) and taxon- specific oligonucleotide probes to study interactions between toxic dinoflagellates and their associated bacteria. Eur. J. Phycol. 35: 315-329.
CHOI, D.H, AND B.C. CHO. 2006. Lutibacter litoralis gen. nov., sp. nov., a marine bacterium of the family Flavobacteriaceae isolated from tidal flat sediment. Int. J. Syst. Evol. Microbiol. 56: 771-776.
CORZO-DELGADO, J.E., AND J.M.GOMEZ-MATEOS. 2006. Stenotrophomonas maltophilia, an increasingly important nosocomial pathogen. Enferm. Infecc. Microbiol. Clin. 24: 1-3.
CUNHA, M. A., M. A. ALMEIDA, AND F. ALCANTARA. 2001. Short-term responses of natural planktonic bacterial community to the changing water properties in an estuarine environment: Ectoenzymatic activity, glucose incorporation, and biomass production. Microb Ecol 42:69-79.
CYTRYN, E., J. VAN RIJN, A. SCHRAMM, A. GIESEKE, AND D. MINZ. 2005. Identification of bacteria potentially responsible for oxic and anoxic sulfide oxidation in biofilters of a recirculating mariculture system. Appl. Environ. Microbiol.71:6134-41.
DELONG, E.F., G.S. WICKHAM, AND N.R. PACE. 1989. Phylogenetic strains: ribosomal RNA-based probes for the identification of single cells. Science 243:1360-1363.
DITTMAN, S. 1999. The Wadden Sea ecosystem: Stability properties and mechanisms. New York: Springer-Verlag.
EICHLER, S., R. CHRISTEN, C. HOLTJE, P. WESTPHAL, J. BOTEL, I. BRETTAR, A. MEHLING, AND
M.G. HÖFLE. 2006. Composition and dynamics of a drinking water supply system as assessed by RNA- and DNA-based 16S rRNA gene fingerpinting. Appl. Environ. Microbiol. 72:1858-1872.
EILERS, H., J. PERNTHALER, F. O. GLÖCKNER, AND R. AMANN. 2000. Culturability and in situ abundance of pelagic bacteria from the North Sea. Appl. Environ. Microbiol. 66: 3044-3051.
EISMA, D., AND A. LI. 1993. Changes in suspended-matter floc size during the tidal cycle in the Dollard Estuary. Neth. J. Sea Res. 31: 107-117.
FUGATE, D.C., AND C.T. FRIEDRICHS. 2003. Controls on suspended aggregate size in partially mixed estuaries. Estuar. Coast. Shelf Sci. 58: 389-404.

Kapitel III Tidal effects on coastal bacterioplankton
67
GICH, F., K. SCHUBERT, A. BRUNS, H. HOFFELNER, AND J. OVERMANN. 2005. Specific detection, isolation, and characterization of selected, previously uncultured members of the freshwater bacterioplankton community. Appl. Environ. Microbiol. 71: 5908-19.
GLÖCKNER, F.O., R. AMANN, A. ALFREIDER, J. PERNTHALER, R. PSENNER, K. TREBESIUS, AND
K. H. SCHLEIFER. 1996. An in situ hybridization protocol for detection and identification of planktonic bacteria. Syst. Appl. Microbiol. 19: 403-406.
GROSSART, H. P., T. BRINKHOFF, T. MARTENS, C. DUERSELEN, G. LIEBEZEIT, AND M. SIMON. 2004. Tidal dynamics of dissolved and particulate matter and bacteria in a tidal flat ecosystem in spring and fall. Limnol. Oceanogr. 49: 2212-2222.
GÜNTHER, C.-P. 1999. Settlement, secondary dispersal and turnover rate of benthic macrofauna, p. 133-146. In S. Dittmann [ed.] The Wadden Sea Ecosystem. New York: Springer Verlag.
HAGSTROM, A., J. PINHASSI, AND U. L. ZWEIFEL. 2000. Biogeographical diversity among marine bacterioplankton. Aquat. Microb. Ecol. 21: 231-244.
KARAKURT, A., O. ABDIK, A. SENGUN, R. KARADAG, S. SARICAOGLU, H. Y. SARIKATIPOGLU, AND H. HASIRIPI. 2006. Stenotrophomonas maltophilia Endophthalmitis after cataract extraction. Ocul. Immunol. Inflamm. 14: 41-6.
KHAN, S. T., Y. NAKAGAWA, AND S. HARAYAMA. 2006. Krokinobacter gen. nov. , with three novel species, in the family Flavobacteriaceae. Int. J. Syst. Evol. Microbiol. 56: 323-328.
KIRCHMAN, D. L. 2002. The ecology of Cytophaga-Flavobacteria in aquatic environments. FEMS Microbiol. Ecol. 39: 91-100.
KOBLIZEK, M., J. STON-EGIERT, S. SAGAN, AND Z.S. KOLBER. 2005. Diel changes in bacteriochlorophyll a concentration suggest rapid bacterioplankton cycling in the Baltic Sea. FEMS Microbiol. Ecol. 51: 353-361.
LLOBET-BROSSA, E., R. ROSSELLO-MORA, AND R. AMANN. 1998. Microbial community composition of Wadden Sea sediments as revealed by fluorescence-in-situ-hybridization. Appl. Environ. Microbiol. 64: 2691-2696.
LUDWIG, W., AND OTHERS. 2004. ARB: a software environment for sequence data. Nucl. Acids Res. 32: 1363-1371.
LUNAU, M., A. LEMKE, O. DELLWIG, AND M. SIMON. 2006. Physical and biogeochemical controls of microaggregate dynamics in a tidally affected coastal ecosystem. Limnol. Oceanogr. 51: 847-859.
MANZ, W., R. AMANN, W. LUDWIG, M. WAGNER, AND K. H. SCHLEIFER. 1992. Phylogenetic oligodeoxynucleotide probes for the major subclasses of Proteobacteria – Problems and solutions. Syst. Appl. Microbiol. 15: 593-600.
MANZ, W., R. AMANN, W. LUDWIG, M. VANCANNEYT, AND K.H. SCHLEIFER. 1996. Application of a suite of 16S rRNA-specific oligonucleotide probes designed to investigate bacteria of the phylum Cytophaga-Flavobacter-Bacteroides in the natural environment. Microbiology 142: 1097-1106.
MESSING, J. 1983. New M13 vectors for cloning. Meth. Enzymol. 101: 20-78.
MIKKELSEN, O., AND A. PEJRUP. 1998. Comparison of flocculated and dispersed suspended sediment in the Dollard estuary, p. 199-209. In K. Black, D. Paterson and A. Cramp. [eds.] Sedimentary Processes in the Intertidal Zone. London, Geological Society.

Kapitel III Tidal effects on coastal bacterioplankton
68
MOESENEDER, M. M., J. M. ARRIETA, G. MUYZER, C. WINTER, AND G. J. HERNDL. 1999. Optimization of Terminal-Restriction Fragment Length Polymorphism Analysis for complex marine bacterioplankton communities and comparison with Denaturing Gradient Gel Electrophoresis. Appl. Environ. Microbiol. 65: 3518-3525.
MUSSMANN, M., K. ISHII, R. RABUS, AND R. AMANN. 2005. Diversity and vertical distribution of cultured and uncultured Deltaproteobacteria in an intertidal mudflat of the Wadden Sea. Environ. Microbiol. 7: 405-18.
MUYZER, G., T. BRINKHOFF, U. NÜBEL, C. SANTEGOEDS, H. SCHÄFER AND C. WAWER. 1998. Denaturing gradient gel electrophoresis (DGGE) in microbial ecology. In: Akkermans ADL. J.D. van Elsas, F. J. Bruijn (eds) Molecular microbial ecology manual. Kluwer Academic Publishers, Dordrecht, 1-27.
NEEF, A. 1997. Anwendung der in-situ-Einzell-Identifizierung von Bakterien zur Populationsanalyse in komplexen mikrobiellen Biozönosen. Ph.D. thesis. Technische Universität München.
NIESEL, V., AND C. P. GÜNTHER. 1999. Distribution of nutrients, algae and zooplankton in the Spiekeroog backbarrier system, p. 77-94. In S. Dittmann [ed.] The Wadden Sea Ecosystem. Stability properties and mechanisms. Springer Verlag.
PERNTHALER, A., J. PERNTHALER, AND R. AMANN. 2002. Fluorescence in situ hybridization and catalyzed reporter deposition for the identification of marine bacteria. Appl. Environ. Microbiol. 68: 3094-3101.
--------, AND J. PERNTHALER. 2005. Diurnal variation of cell proliferation in three bacterial taxa from coastal North Sea waters. Appl. Environ. Microbiol. 71:4638-4644.
POULSEN, L. K., G. BALLARD, AND D. A. STAHL. 1993. Use of rRNA fluorescence in situ hybridization for measuring the activity of single cells in young and established biofilms. Appl. Environ. Microbiol. 59:1354-1360.
RABUS R., M. FUKUI, H. WILKES, AND WIDDEL F. (1996). Degradative capacities and 16S rRNA-targeted whole-cell hybridization of sulfate-reducing bacteria in an anaerobic enrichment culture utilizing alkylbenzenes from crude oil. Appl. Environ. Microbiol. 62: 3605-3613
RAPPÉ, M. S., K. VERGIN, AND S. J. GIOVANNONI. 2000. Phylogenetic comparison of a coastal bacterioplankton community with its counterparts in open ocean and freshwater systems. Fems Microbiol Ecol 33: 219-232
RINK, B., S. SEEBERGER, T. MARTENS, C. DUERSELEN, M. SIMON, AND T. BRINKHOFF. 2006. Effects of a phytoplankton bloom in a coastal ecosystem on the composition of bacterial communities. Aquat. Microb. Ecol., submitted.
SAMBROOK, J., E. F. FRISCH, AND T. MANIATIS. 1989. Northern Hybridisation. In Molecular Cloning: a laboratory manual, 2nd edn., Cold Spring Harbour Laboratory Press, New York.
SCHÄFER, H., AND OTHERS. 2001. Microbial community dynamics in Mediterranean nutrient-enriched seawater mesocosms: changes in the genetic diversity of bacterial populations. FEMS Microbiol. Ecol. 34: 243-253.
SCHUT, F. (1994) Ph.D. thesis. University of Groningen, Groningen, The Netherlands.
SEIBOLD, A., A. WICHELS, AND C. SCHÜTT. 2001. Diversity of endocytic bacteria in the dinoflagellate Noctiluca scintillans. Aquat. Microb. Ecol. 25: 229-235.

Kapitel III Tidal effects on coastal bacterioplankton
69
SEKAR, R., A. PERNTHALER, J. PERNTHALER, F. WARNECKE, T. POSCH, AND R. AMANN. 2003. An improved protocol for quantification of freshwater Actinobacteria by fluorescence in situ hybridization. Appl. Environ. Microbiol. 69: 2928-2935.
SELJE N., AND M. SIMON. 2003. Composition and dynamics of particle-associated and free-living bacterial communities in the Weser estuary, Germany. Aquat. Microb. Ecol. 30:221-237.
SELJE, N., M. SIMON, AND T. BRINKHOFF. 2004. A newly discovered Roseobacter cluster in temperate and polar oceans. Nature 427: 445-448.
SFANOS, K., D. HARMODY, P. DANG, A. LEDGER, S. POMPONI, P. MCCARTHY, AND J. LOPEZ. 2005. A molecular systematic survey of cultured microbial associates of deep-water marine invertebrates. Syst. Appl. Microbiol. 28: 242-264.
SIMON, M., AND F. AZAM. 1989. Protein content and protein synthesis rates of planktonic marine bacteria. Mar. Ecol. Prog. Ser. 51: 201-213.
STAHL, D. A., AND R. AMANN. 1991. Development and application of nucleic acid probes. 205-248. In E. Stackebrandt and M. Goodfellow [eds.], Nucleic acid techniques in bacterial systematics. John Wiley & Sons Ltd., Chichester, England.
STEVENS, H., T. BRINKHOFF, AND M. SIMON. 2005a. Composition of free-living, aggregate-associated and sediment surface-associated bacterial communities in the German Wadden Sea. Aquat. Microb. Ecol. 38: 15-30.
--------, M. STÜBNER, M. SIMON, AND T. BRINKHOFF. 2005b. Phylogeny of Proteobacteria and Bacteroidetes from oxic habitats of a tidal flat ecosystem. FEMS Microbiol. Ecol. 54: 351-365.
TILLMANN, U., K.-J. HESSE AND F. COLIJN. 2000. Planktonic primary production in the German Wadden Sea. J. Plankton Res. 22: 1253-1276.
VAN DUYL, F. C., B. DE WINDER, A. J. KOP, AND U. WOLLENZIEN. 1999. Tidal coupling between carbohydrate concentrations and bacterial activities in diatom-inhabited intertidal mudflats. Mar. Ecol. Prog. Ser. 191: 19-32.
VAN LEUSSEN, W. 1996. Erosion/sedimentation cycles in the Ems estuary. Arch. Hydrobiol. Spec. Issues Advanc. Limnol. 47: 179-193.
WALLNER G., R. AMANN, AND W. BEISKER. 1993. Optimizing fluorescent in situ hybridization with rRNA-targeted oligonucleotide probes for flow cytometric identification of microorganisms. Cytometry. 14: 136-143.
WINTER, C., M. M. MOESENEDER, AND G. J. HERNDL. 2001. Impact of UV radiation on bacterioplankton community composition. Appl. Environ. Microbiol. 67:665-672.
YOON, J. H., S. J. KANG, C. H. LEE, AND T. K. OH. 2005. Dokdonia donghaensis gen. nov., sp. nov., isolated from sea water. Int. J. Syst. Evol. Microbiol. 55: 2323-2328.
ZUBKOV, M. V., B. M. FUCHS, S. D. ARCHER, R. P. KIENE, R. AMANN, AND P. H. BURKILL. 2002. A population of the alpha-Proteobacteria dominates the bacterioplankton and dimethylsulphoniopropionate uptake after an algal bloom in the North Sea. Deep-sea Res. II, Top. Stud. Oceanogr. 49: 3017-3038.

Kapitel III Tidal effects on coastal bacterioplankton
70
Figure legends
Fig. 1. Temperature and salinity (a), dry weight (DW) and percent particulate organic carbon
(%POC) (b), chlorophyll a (chl a), phaeopigments and phaeopigment/chl a ratio (c),
abundance of total, particle-attached (PA) and free-living (FL) bacteria (d), and bacterial
protein production (BPP) (e) in the Wadden Sea during a tidal cycle in July 2005. X-axis:
Time of high tide (HT), mean tide (MT1/2) and low tide (LT).
Fig. 2. FISH counts for samples taken in November 1999, May 2000 and July 2005 during
tidal cycles at high tide, mean tide, and low tide. Results of July 2005 were obtained using
CARD-FISH. Calculation includes all values of single counting grids to display outliners
(dots). Box-Whisker-Plots show the 25th/75th percentile (box), the mean (dashed line), the
median (solid line) and the 5th/95th percentile (error bars). Abbreviations: EUB (EUB338),
CFB (CFB319a), ALF (ALF968), GAM (GAM42a), BET (BET42a), ROS (ROS536), ARCH
(ARCH915), SRB (SRB385), SRBdb (SRB385db), NON (NON338).
Fig. 3. DGGE fingerprints of sediment surface (SE), particle-attached (PA), and free-living
(FL) bacterial communities of the Wadden Sea during tidal cycles in November 1999 (Nov)
and May 2000 using primer sets for 16S rRNA genes of Bacteria. The arrows mark excised
and sequenced bands. Bands sequenced in this study include “TC” for tidal cycle, bands
named GWS-eX- PA/FL refer to sequences of Rink et al. (2006). Std. = standard.
Fig. 4. DGGE fingerprints of free-living (FL) and particle-attached (PA) bacterial
communities during a tidal cycle in July 2005, using primer sets for 16S rRNA genes of
Bacteria (EUB), Bacteroidetes (CFB), alpha-Proteobacteria (ALF) and the Roseobacter
clade (ROS). “DNA” marks banding patterns obtained from 16S rRNA gene fragments and
“cDNA” refers to banding patterns derived from 16S rRNA after reverse transcription. HT =
high tide, MT1/2 = mean tide, LT = low tide. The numbered arrows mark excised and
sequenced bands. Because of the small fragment size of the Roseobacter amplicons (approx.
200 bp) the DGGE bands were not excised for sequencing. Std. = standard.
Fig. 5. Cluster analyses of DGGE banding patterns (July 2005, Fig. 4) of particle-attached
(PA) and free-living (FL) Bacteria (EUB), Bacteroidetes (CFB), alpha-Proteobacteria (ALF)
and the Roseobacter clade (ROS). “DNA” marks banding patterns obtained from 16S rRNA

Kapitel III Tidal effects on coastal bacterioplankton
71
gene fragments and “cDNA” refers to banding patterns derived from 16S rRNA after reverse
transcription. Samples were taken during a tidal cycle at high tide (HT), mean tide (MT1/2)
and low tide (LT). The similarity matrix was calculated using UPGMA and Pearson
correlation. Std. = standard.
Fig. 6. Phylogenetic trees of Proteobacteria and Actinobacteria (A) and Bacteroidetes (B)
calculated with Maximum-Likelihood based on 16S rRNA gene fragments. Sequences
obtained in this and one earlier study partially including the same samples [Rink et al. (2006),
compare Fig. 3] are highlighted in bold. Alpha, delta, gamma and beta = different
phylogenetic groups of Proteobacteria, WAC I = Wadden Sea Alpha Cluster (Stevens et al.
2005), RCA = Roseobacter Clade Affiliated (Selje et al. 2004), SAMMIC = Surface Attached
Marine MICrobes (Stevens et al. 2005). “GWS” (German Wadden Sea) indicates clones
obtained from the same habitat in this and earlier studies. The percentage of sequence
divergence is indicated by the scale bars.
Supplementary Data
Fig. I. Particulate organic carbon (POC), ratio of POC per dry weight (DW) and density of
particle-attached bacteria at high tide (HT), mean tide (MT) and low tide (LT) in July 2005.
Fig. II. DGGE fingerprints of sediment surface, particle-attached, and free-living bacterial
communities of the Wadden Sea during tidal cycles in November 1999 and May 2000 using
primer sets for 16S rRNA genes of Bacteria.
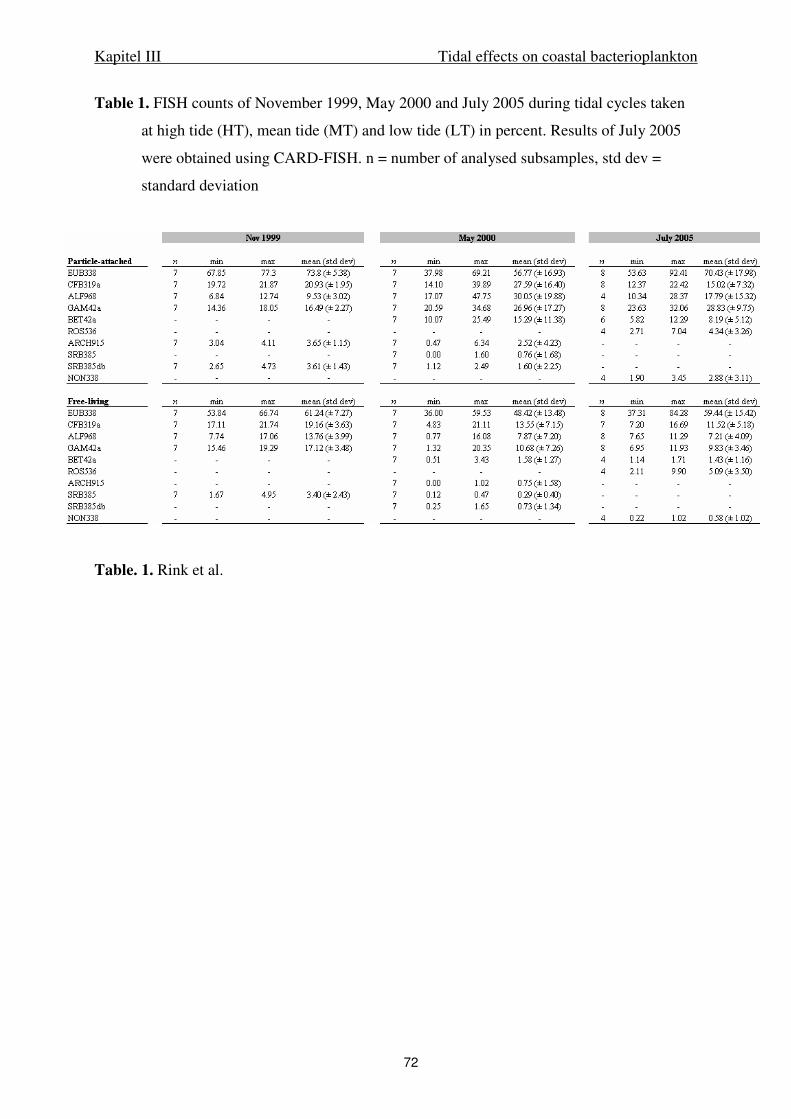
Kapitel III Tidal effects on coastal bacterioplankton
72
Table 1. FISH counts of November 1999, May 2000 and July 2005 during tidal cycles taken
at high tide (HT), mean tide (MT) and low tide (LT) in percent. Results of July 2005
were obtained using CARD-FISH. n = number of analysed subsamples, std dev =
standard deviation
Table. 1. Rink et al.
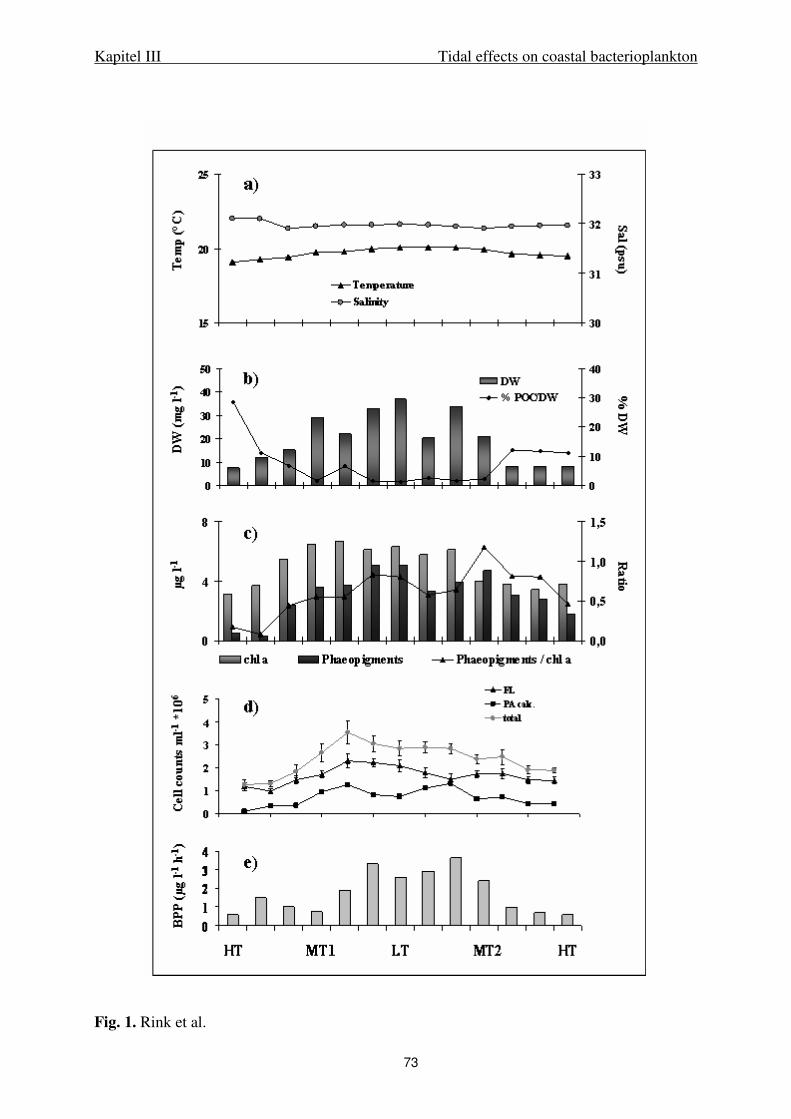
Kapitel III Tidal effects on coastal bacterioplankton
73
Fig. 1. Rink et al.
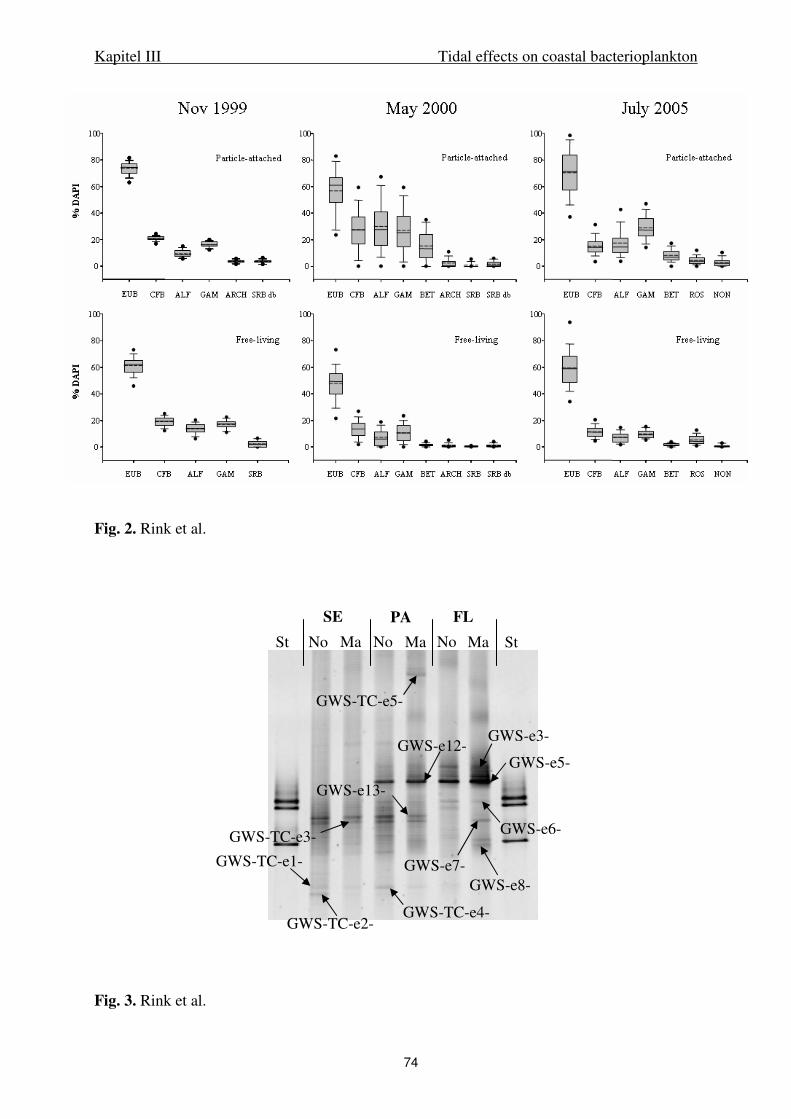
Kapitel III Tidal effects on coastal bacterioplankton
74
Fig. 2. Rink et al.
Fig. 3. Rink et al.
GWS-TC-e5-
GWS-e3-
GWS-e6-
GWS-e7-
GWS-e5-
GWS-e8-
GWS-e13-
GWS-TC-e4-
GWS-e12-
GWS-TC-e3-
GWS-TC-e1-
GWS-TC-e2-
No Ma No Ma No MaSt St
SE PA FL

Kapitel III Tidal effects on coastal bacterioplankton
75
EUB
H MT
L MT
H MT
L MT
H MT
L MT
H MT
L MT
PA FL PA FL
DNA cDNA
CFB
ALF
ROS
Std. Std.
GWS-TC-e6-GWS-TC-e7-
GWS-TC-e8-GWS-TC-e9-
GWS-TC-e10-
GWS-TC-e11-
GWS-TC-c1-
GWS-TC-c2-GWS-TC-c3- GWS-TC-c4-
GWS-TC-a1-
GWS-TC-a2-GWS-TC-a3-
GWS-TC-a4-
GWS-TC-a5-
GWS-TC-a6-
Fig. 4. Rink et al.

Kapitel III Tidal effects on coastal bacterioplankton
76
Fig. 5. Rink et al.
EUB
CFB
ALF
ROS
Pearson correlation [0.0%-100.0%]
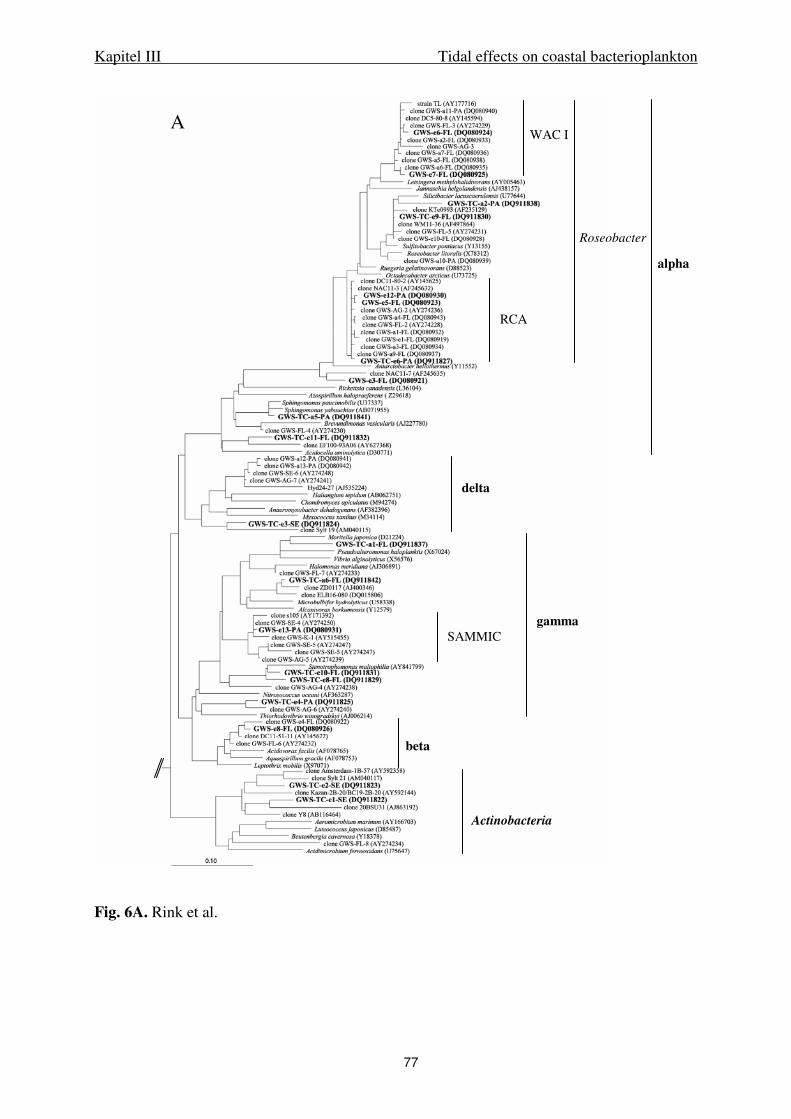
Kapitel III Tidal effects on coastal bacterioplankton
77
Fig. 6A. Rink et al.
alpha
delta
gamma
Actinobacteria
WAC I
RCA
SAMMIC
beta
A
Roseobacter

Kapitel III Tidal effects on coastal bacterioplankton
78
Fig. 6B. Rink et al.
B
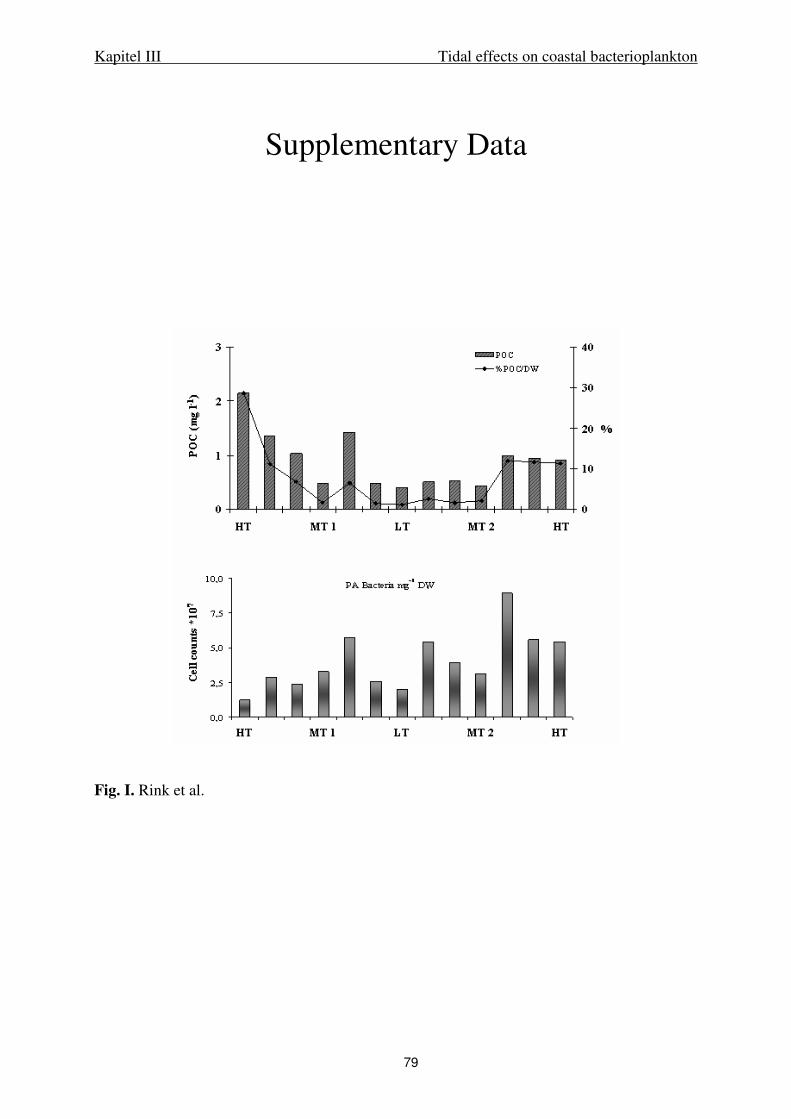
Kapitel III Tidal effects on coastal bacterioplankton
79
Fig. I. Rink et al.
Supplementary Data
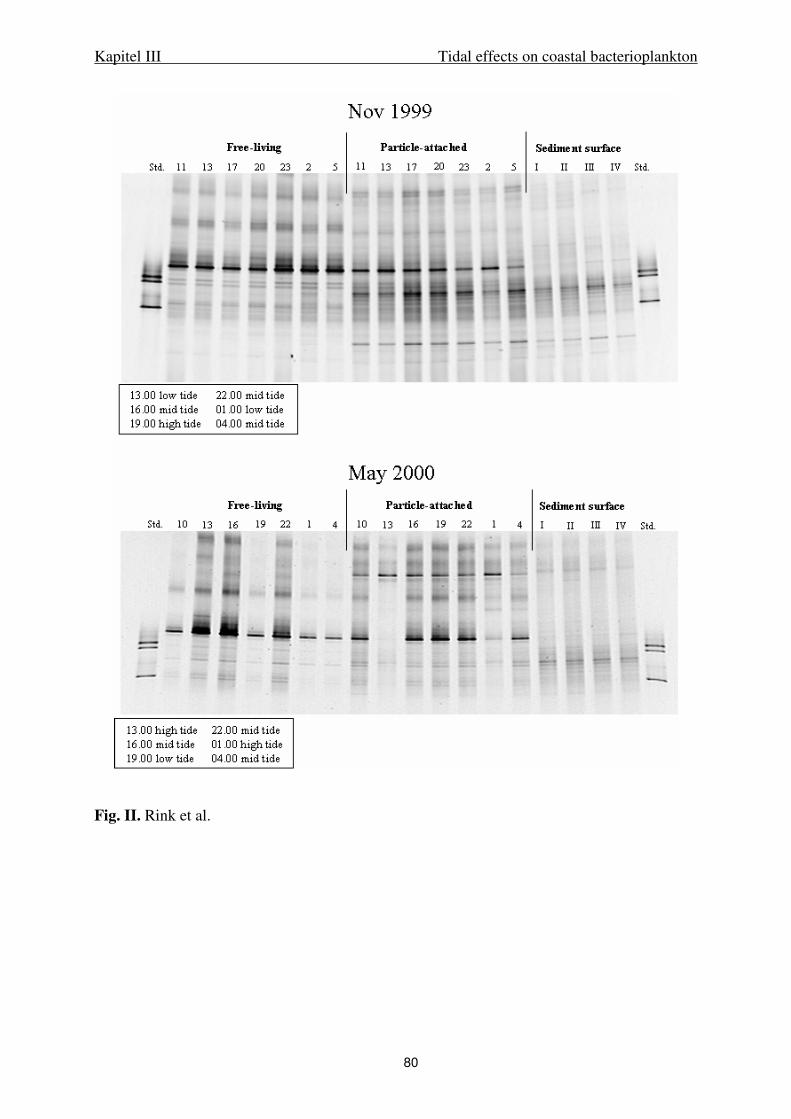
Kapitel III Tidal effects on coastal bacterioplankton
80
Fig. II. Rink et al.

81
IV.
Diversity and abundance of Gram-positive bacteria
in a tidal flat ecosystem

Kapitel IV Diversity and abundance of Gram-positive bacteria
82
Diversity and abundance of Gram-positive bacteria in a tidal flat ecosystem
Heike Stevens, Thorsten Brinkhoff, Beate Rink, John Vollmers, and Meinhard Simon*
Institute for Chemistry and Biology of the Marine Environment (ICBM),
University of Oldenburg, Germany
Running title: Gram-positive bacteria in tidal flats
Key words: Gram-positive bacteria, Actinobacteria, Firmicutes, tidal flats, dilution cultures,
fluorescence in situ hybridization, DGGE
___________________________________________________________________________
*Corresponding author
Mailing address: ICBM, University of Oldenburg, PO Box 2503, D-26111 Oldenburg,
Germany. Phone: +49 (0) 441 / 798-5361. Fax: +49 (0) 441 / 798-3438. E-mail:

Kapitel IV Diversity and abundance of Gram-positive bacteria
83
Abstract.
Gram-positive (Gram+) bacteria recently have been identified as important components of
freshwater ecosystems and are also present in marine environments. However, their
quantitative significance and possible role in the latter systems is still little studied, in
particular in coastal regions. Therefore, we investigated the abundance and composition of
Gram+ bacteria in the Wadden Sea, a tidal flat ecosystem in the German Bight of the North
Sea. Applying fluorescence in situ hybridization we found that Actinobacteria constitute 4-7%
of total bacteria in the Wadden Sea and slightly higher proportions in a freshwater drainage
channel connected to the sea by a sluice. The application of DGGE of 16S rRNA gene
fragments after amplification by an Actinobacteria-specific primer set and subsequent
sequencing showed that the composition of the actinobacterial community in the Wadden Sea
was distinctly different from that in the freshwater system. A clone library of 103 clones
yielded 8 Gram+ phylotypes which are related closely to other marine phylotypes including the
Marine Actinobacteria Clade but also to freshwater phylotypes. We applied dilution cultures,
enriched with various biopolymers for isolating bacteria from the bulk water, suspended
aggregates, the oxic surface and oxic/anoxic transition zone of the sediment, Marine Broth and
Fucus vesiculosus extracts. Fifty three isolates affiliated to seven families of the order
Actinomycetales and 9 isolates to the family Bacillaceae. The salinity range (1 to 45‰ NaCl)
and growth optimum of fourteen strains from various families showed that all except one strain
exhibited a rather broad range of sustained growth from 1 to >20‰ NaCl and several strains
exhibited an optimum of >10‰ NaCl. The results indicate that the Gram+ bacterial community
in the Wadden Sea is surprisingly diverse and consists mainly of indigenous species which
appear to be well adapted to the environmental conditions of this coastal ecosystem.

Kapitel IV Diversity and abundance of Gram-positive bacteria
84
Introduction
In the recent past, more and more evidence accumulated which indicated that Gram-positive
(Gram+) bacteria, in particular Actinobacteria, are of hitherto unknown significance in aquatic
ecosystems (Bull et al., 2005). Studies in freshwater systems showed a surprising diversity and
abundance of Actinobacteria even though the specific ecological role remains to be unveiled
(Glöckner et al., 2000; Hahn et al., 2003; Stepanauskas et al., 2003; Warnecke et al., 2004;
Haukka et al., 2005; Allgaier and Grossart, 2006). It has been known for more than 60 years
that Gram+ bacteria also exist in marine environments, but they were thought not to be
indigenous but introduced from terrestrial habitats (Zobell and Upham., 1944; Goodfellow and
Haynes, 1984). Since the mid-1990s, studies using culture-independent but also refined
culture-dependent methods indicated that Gram+ bacteria reveal an unexpected diversity in
marine bacterioplankton communities (Fuhrman et al., 1993; Rappé et al., 1997; Suzuki et al.,
1997; Rappé et al., 1999; Fuchs et al., 2005) as well as in marine sediments and a surprisingly
high abundance of up to 13% (Jensen and Fenical, 1995; Moran et al., 1995; Urakawa et al.,
1999; Mincer et al., 2002; Maldonado et al., 2005; Pathom-aree et al., 2006). Phylogenetic
analyses of the phylotypes and isolates obtained in these studies implied that, in fact,
indigenous marine Gram+ bacteria exist. Some of the Gram+ bacteria obtained from marine
habitats fall into distinct “marine” clusters, only distantly related to clusters comprising also
Gram+ bacteria from freshwater and soil. The Marine Actinobacteria Clade, (Rappé et al.,
1999), which includes the “BD1-5 cluster” (Fuhrman et al., 1993), a deeply branching cluster
within the Actinobacteria, comprises exclusively marine bacterioplankton phylotypes. The
MAR 1 cluster (Mincer et al., 2002) consists of Actinobacteria isolates from tropical and
subtropical marine sediments, but is branching not as deeply as the Marine Actinobacteria
Clade.
Quantitative studies on marine Gram+ bacteria are still scarce. A biomarker study on the basis
of the composition of phospholipid ester-linked fatty acids (PLFA) indicated that Gram+
bacteria are major components of bacterial communities in sediments of a eutrophic bay
(Rajendran et al., 1994). Actinobacteria constituted up to 5% of total bacteria in shallow
marine sediments and <1.4% in an Arctic deep-sea sediment as determined by dot blot and
fluorescence in situ hybridization (Moran et al., 1995; Llobet-Brossa et al., 1998; Ravenschlag
et al., 1999). In the Sargasso Sea and the Arabian Sea, Actinobacteria have been identified as
substantial components of the bacterioplankton (Fuchs et al., 2005; Morris et al., 2005). In the
Delaware estuary, USA, Actinobacteria constitute a decreasing fraction of total
bacterioplankton numbers and of the proportions assimilating glucose and polysaccharide with

Kapitel IV Diversity and abundance of Gram-positive bacteria
85
increasing salinity (Elifantz et al., 2005; Kirchman et al., 2005). It is of particular interest to
reveal the significance, composition and abundance of Gram+ bacterial communities in coastal
tidally affected ecosystems and whether in these systems indigenous Gram+ bacteria do exist
These systems are severely understudied with respect to Gram+ bacteria as compared to other
marine systems (Bull et al., 2005).
The Wadden Sea is a shallow and nutrient-rich tidally affected coastal ecosystem of the
southern North Sea stretching from the Netherlands (Den Helder) to Denmark (Esbjerg). Due
to the pronounced tidal dynamics and inputs of organic and inorganic nutrients from land,
freshwaters, and the North Sea it can be considered as a melting pot in which microbial
processes are of major significance (Dittmann, 1999; Poremba et al., 1999). In the recent past
extensive work on microbial processes and on the composition of bacterial communities in the
bulk water, on suspended aggregates and in the sediment of the Wadden Sea has been carried
out (e.g. Llobet-Brossa et al., 1998; Köpke et al., 2005; Stevens et al., 2005a, 2005b; Lunau et
al., 2006; Wilms et al., 2006).
This study investigated the composition and abundance of the Gram+ bacterial community in
the Wadden Sea. We applied cultivation-based approaches, using enrichment and dilution
cultures amended with a variety of biopolymers, as well as cultivation-independent approaches
using Gram+-specific CARD-FISH (CAtalyzed Reporter Deposition Fluorescence In Situ
Hybridization, Sekar et al., 2003), DGGE (Denaturing Gradient Gel Electrophoresis, Muyzer
et al., 1993) and clone library construction. The salt requirements for growth of representative
isolates were examined by determining their growth adaptation to the ambient salinity range.
RESULTS
Isolation
Seventeen of 63 bulk water isolates (27%) obtained from dilution cultures of the May sample
were Gram+ strains. Isolates affiliating to α- and γ-Proteobacteria constituted proportions of
44.4% and 20.6%, respectively, and those affiliating to Bacteroidetes 7.9%. In October, the
majority of strains affiliated to α -Proteobacteria (31.0%), Actinobacteria (30.2%) and γ -
Proteobacteria (24.0%). Including the Firmicutes isolates (n=6) 45 of the 129 October strains
(34.9%) were Gram+ bacteria, of which 35.6% originated from the bulk water, 20% from
aggregates and the transition zone each and 24.4% from the oxic layer of the sediment. In
other studies of which Gram+ isolates were included in the phylogenetic analysis, only the

Kapitel IV Diversity and abundance of Gram-positive bacteria
86
bulk water (Bruns et al., 2003; Selje et al., 2005) or aggregates (Grossart et al., 2004) were
investigated.
Gram+ isolates were obtained with all substrates used. The only isolate obtained with
laminarin also affiliated to Gram+ bacteria. The highest MPN dilution steps which yielded
isolates, was 10-8. These isolates were derived from bulk water in May with casein and from
the sediment transition zone in October with MB as substrate and affiliated to Actinobacteria
and Firmicutes, respectively. Overall most Gram+ isolates obtained in May and October were
isolated with alginate (n=11), followed by agar (n=9), MB and starch (n=8). Only few isolates
were retrieved from dilution cultures amended with casein (n=2) and palmitate (n=4).
Dilution steps 10–1, 10–2 and 10–4 yielded >10 and the others not more than 7 Gram+ isolates
(Fig. 2).
Salinity-dependent growth
All strains except one exhibited a rather broad salinity range of sustained growth and 9 strains
grew equally well from 1 to >20‰ NaCl (Table 1) and thus appeared well adapted to the
salinities occurring in the Wadden Sea. Two strains grew significantly better at salinity ranges
>10‰ (GWS-BW-H260, GWS-BW-H252). One strain, affiliating to Bacilli, grew
significantly better at salinities <5‰.
CARD-FISH
Hybridization efficiencies varied from 48.9 to 67.7% as indicated by the ratio of numbers of
EUB338 positive over DAPI cell counts and subtracting the non-EUB338 numbers which
remained below 1.4% (Table 2). Gram+ bacteria were detected at all five locations and
proportions ranged from 4.2 to 8.0% of DAPI cell counts (Table 2). The highest proportion
occurred in the freshwater location behind the sluice.
DGGE analysis
The banding patterns of the Actinobacteria–specific DGGE revealed distinct communities of
the freshwater and the marine locations (Fig. 3). The marine samples were rather similar with
one prominent band occurring in all samples. However, the particle-associated and free-living
bacterial fractions exhibited also different bands. Band numbers ranged between 4 in the
particle-associated bacterial fraction of station 4 and 9 in the free-living bacterial fraction of

Kapitel IV Diversity and abundance of Gram-positive bacteria
87
station 2. Sequencing of the excised bands yielded only actinobacterial 16S rRNA phylotypes,
indicating the specificity of the primers.
Clone library
Altogether 103 different clones were sequenced . Two of them were identified as chimera.
Eight of the remaining clones (7.8%) affiliated to Gram+ bacteria and the majority to γ- and
δ-Proteobacteria.
Phylogenetic affiliation
A total of 94 Gram+ 16S rRNA gene sequences was phylogenetically analyzed (Fig. 4A and
B, Tab. 3), 82 from isolates, 8 from the clone library and 4 from DGGE bands. To avoid an
overestimation of diversity, sequences with similarities of ≥99% were merged into 15
“sequence-groups” (I – XV) resulting in a total of 58 different sequence types (Tab. 3, Fig. 4).
Sixty-three sequences from isolates, 7 from clones and 4 from DGGE bands affiliated to
Actinobacteria. The sequences mainly clustered into seven families of the order
Actinomycetales, the Microbacteriaceae, Micrococcaceae, Mycobacteriaceae, Nocardiaceae,
Nocardioidaceae, Promicromonosporaceae, Pseudonocardiaceae, and the
Sanguibacteraceae. Most sequences group with the Microbacteriaceae (18 isolates, 1 clone),
with the Micrococcaceae (16 isolates) and with the Nocardioidaceae (15 isolates). Because of
their low sequence similarity to the next relative, Streptomyces cinnabarus and candidatus
“Microthrix parvicella”, clone K39 and DGGE bands GWS-DG2, GWS-DG1, GWS-FL-8,
GWS-DG3 could only be classified on the class-level (Table 3). The latter three phylotypes
clustered together with various phylotypes of uncultured marine Actinobacteria. Clone GWS-
K46 and isolates GP-5 and GP-6 belong to the order Actinobacteria with next related species
of the subclass Frankinae. Clones GWS-K72, GWS-K105 and GWS-K112 clustered within
the Marine Actinobacteria Clade.
Sixteen of the 58 Gram+ sequence types (28%) obtained from the Wadden Sea exhibit
a closest relative of marine origin, 37 of other environments, such as freshwater, soil, plants,
or endophytic habitats. For 5 sequence types no information is available on the source of
isolation of the next related sequence (Table 3). Our strains affiliating with the
Micrococcaceae were mainly isolated from bulk water, isolates affiliating with the
Nocardioidaceae from bulk water and aggregates. For all other sequences and isolates no

Kapitel IV Diversity and abundance of Gram-positive bacteria
88
relationship exists between phylogeny and habitat or substrate from which they were
obtained.
Eight of our strains, 10 from other studies of the same habitat and one clone (K48, not
shown in Fig. 4B) affiliate to the phylum Firmicutes. Fifteen of the isolates were merged into
five sequence groups. All isolates affiliate to the order Bacillales and, except sequence group
XIV (GP-13, GP-14) and isolate GWS-TZ-H232, fall within the family Bacillaceae.
Sequence group XIV affiliates to the Planococcaceae. The clone K48 affiliates within the
class Clostridia to the “Peptostreptococcaceae” (family name not validly published), with the
next relative Fusibacter paucivorans (Table 3). Isolates affiliating to the Firmicutes were
never obtained from aggregates, but from the sediment layers and the bulk water. They were
obtained from assays performed with MB, agar, alginate, and stearine.
Discussion
Our results show that Gram+ bacteria constitute a substantial fraction of the bacterial
community in the Wadden Sea and that they are indigenous to this environment. They
comprise around 5% of total bacterial numbers and are a major component of its so far
cultivated fraction. However, the isolated strains were only distantly related to most
phylotypes retrieved from the clone library and from the Actinobacteria-specific DGGE
bands. Most of the Gram+ isolates tested grew at a rather wide salinity range including 20 and
30‰ NaCl, thus indicating that they are well adapted to growing in the Wadden Sea , in
which salinities of 26 to 33 psu occur (http://las.physik.uni-oldenburg.de/wattstation). Even
though it seems possible that Gram+ bacteria are also washed in from terrestrial run off and
by releasing drained fresh water through the dike sluices these bacteria do not appear to
constitute prominent members of the marine Gram+ bacterial community. The DGGE bands
of the freshwater sample were distinctly different from the marine ones and only one isolated
strain exhibited a distinct growth optimum at salinities <5‰. Further, several clones of the
clone library affiliated to the Marine Actinobacteria Clade (Rappé et al., 1999), and the
sequence of one prominent DGGE band detected at all marine stations (GWS-DG3) affiliated
to marine Actinobacteria phylotypes and most closely to a phylotype detected in the
northwestern coastal Pacific (Fig. 4; Table 3; Morris et al., 2006).
Gram+ bacteria comprised 27 and 35% of all isolates retrieved in May and October
1999, respectively, and the majority affiliated to Actinobacteria. Thus, Gram+ bacteria
constitute one of the two major cultivable phylogenetic classes of isolates in the Wadden Sea.

Kapitel IV Diversity and abundance of Gram-positive bacteria
89
Because the Gram+ bacteria were obtained with various substrates we assume that the
frequent isolation of these organisms was not due to a cultivation bias but rather a result of the
various biopolymers used as enrichment substrates. Even though most of the isolates were
retrieved from low dilution steps, several isolates from various habitats were obtained from
10-6 to 10-8 dilutions, suggesting that some of them are significant constituents of the bacterial
community in this ecosystem. This is particularly true for sequence groups I
(Microbacteriaceae) and X (Nocardioidaceae). Three of the strains within group I were
obtained from 10-6 or higher, and 6 of the 7 strains in group X from 10-4 or higher (Fig. 4).
This suggestion is supported by the finding that the recently described Aeromicrobium
marinum, isolated from a 10-4 dilution culture retrieved from the Wadden Sea and the first
described marine species within the family Nocardioidaceae, constitutes up to 1% of total
bacteria in the water column (Bruns et al., 2003).
Several other recent studies also isolated Actinobacteria from various marine
environments, but in most cases they constituted lower fractions of all isolates as compared
with our results. In studies from various regions including the German Bight of the North Sea,
the Baltic Sea and the Oregon coast of the Pacific, which examined the diversity of isolates
from bacterioplankton samples and enrichments with various substrates, between <1 and 15%
of all isolates affiliated to Actinobacteria (Suzuki et al., 1997; Eilers et al., 2000; Hagström et
al., 2000; Uphoff et al., 2001). Du et al. (2006) reported that Gram+ bacteria were a
prominent fraction of the pigmented cultivable bacterial community in Chinese estuaries and
were also present in the coastal sea, even though to much lower proportions. In various
marine sediments, ranging from shallow coastal regions to the deepest ocean, Gram+ bacteria
of a rather wide diversity have been isolated in recent studies (Mincer et al. 2002; Köpke et
al., 2005 ; Maldonado et al., 2005; Pathom-aree et al., 2006). These notions indicate that
Gram+ bacteria constitute a greater fraction of the so far cultivable proportion of the bacterial
community in coastal environments than previously assumed.
The fraction of Actinobacteria we assessed by CARD-FISH is in the same range as
that reported from the Delaware Bay, USA (Kirchman et al., 2005). These authors found that
Actinobacteria constitute decreasing proportions of 20-30% of total bacterial numbers to 5%
from the freshwater end of the estuary to the marine Bay. This study did not examine the
phylogenetic composition of the Gram+ bacterial community in the Bay. According to our
DGGE and sequencing results, however, it appears most likely that the composition of the
Gram+ bacterial community in the Bay was different from the freshwater section and that the
decreasing proportion of this phylogenetic class was not only a dilution effect. Our results

Kapitel IV Diversity and abundance of Gram-positive bacteria
90
further imply that an indigenous marine actinobacterial community exists, also in such near
shore environments with intense water exchange with estuarine and freshwater habitats. The
proportions of Actinobacteria we and Kirchman et al. (2005) found in the water column of the
Wadden Sea and the Delaware Bay are somewhat higher than proportions reported from the
upper 4 cm of Wadden Sea sediments, <1 to 3.6% (Llobet-Brossa et al., 1998) and also as
those of the genus Streptomyces detected in a shallow marine sediment by dot blot
hybridization with a genus-specific probe (2.0 to 5.1% of total extracted rRNA; Moran et al.,
1995). Studies based on PLFA indicate that Gram+ bacteria are significant components of the
bacterial communities in eutrophic bays in Japan (Rajendran et al., 1994; Rajendran and
Nagatomo, 1999). In open ocean environments the significance of Actinobacteria appears to
be variable. Whereas in the Sargasso Sea, Actinobacteria constitute <2% of total bacterial
numbers (Morris et al., 2005), in the Arabian Sea variable proportions of 2 and 13% of total
bacteria were found in oligotrophic waters and in the oxygen minimum zone, respectively
(Fuchs et al., 2005). Hence, these data indicate that Actinobacteria are a prominent
component of the bacterial community in shallow coastal ecosystems, in the water column as
well as in the sediment, even though its quantitative proportion is lower than that of other
important classes such as - and -Proteobacteria and Bacteroidetes (Kirchman et al., 2005;
Llobet-Brossa et al., 1998) and lower than that of Actinobacteria in freshwater ecosystems
(Glöckner et al., 2000; Allgaier and Grossart, 2006).
Our phylogenetic analysis of sequences from the isolates, the DGGE bands and clone
library shows a surprisingly high diversity of the Gram+ bacteria. The sequences from the
DGGE bands and the clone library affiliated to distinctly different groups than the isolates of
this phylogenetic lineage. The diversity within Actinobacteria we detected was greater than
that described in other studies using either culture-dependent (Jensen and Fenical, 1995;
Suzuki et al., 1997; Köpke et al., 2005; Maldonado et al., 2005; Du et al., 2006; Pathom-aree
et al., 2006) or culture–independent approaches (Fuhrman et al., 1993; Gray and Herwig,
1996; Suzuki et al., 1997; Rappé et al., 1999; Urakawa et al., 1999; Wilms et al., 2006). This
may be due to the various isolation procedures we applied, such as dilution series and
different substrates and to the various habitats we sampled, but may also reflect the specific
signature of the Wadden Sea ecosystem as a melting pot with marine as well as terrestrial
impacts.
The fact that we successfully enriched and isolated Actinobacteria from various
habitats of the Wadden Sea with various biopolymers shows that these strains are capable of
degrading a variety of polymeric substances. Some of these substances are typical for coastal

Kapitel IV Diversity and abundance of Gram-positive bacteria
91
marine environments such as F. vesiculosus, cellulose, starch, chitin, and laminarin.
Actinobacteria are well known to be capable to degrade various polymeric substances such as
cellulose and lignin, but also rubber and polyester (Haider et al., 1978; Godden and
Penninckx, 1984; Jendrossek, 1997; Pranamuda et al., 1999; Linos et al., 2002). Their
hydrolytic potential appears comparable or even greater than that of the Sphingobacteria and
Flavobacteria group of the Bacteroidetes phylum (Reichenbach, 1992; Kirchman, 2002). It
may explain why Actinobacteria prosper in the bulk water and sediment of the Wadden Sea
and other coastal environments which are characterized by high concentrations of various
biopolymers (Harvey and Mannino, 2001). In fact, Piza et al. (2004) found a surprising
diversity of Actinobacteria in a Brazilian estuary subjected to high pollution during the last
fifty years, and in particular at its brackish end. This high diversity obviously reflects the high
potential of Actinobacteria to degrade complex organic substances including recalcitrant
compounds and pollutants.
In contrast to sequences affiliating to Actinobacteria, our sequences affiliating to
Firmicutes were much less divers. Sequences of all bacterial isolates of this group clustered
within the class Bacilli. The only clone obtained from the Firmicutes belongs to the class
Clostridia. Firmicutes appear to be more prominent members of the subsurface sediment,
possibly because of their ability to produce endospores. In a recent study a surprisingly high
diversity of Firmicutes was found in subsurface sediments of the German Wadden Sea
(Köpke et al., 2005). Since the early 1970s various Bacilli were isolated from marine habitats
(e.g., Denis, 1971; Bonde, 1976; Stackebrandt et al., 1997; Urakawa et al., 1999; Siefert et al.,
2000), but only very few marine isolates affiliated to the classes Lactobacillales, Clostridia,
and Mollicutes (Finne and Matches, 1974; Timmis et al., 1974; Marty, 1986; Gray and
Herwig, 1996). Hence, Bacilli seem to be the most abundant marine Firmicutes.
A further feature, explaining the high diversity of Gram+ bacteria in coastal
ecosystems may be their ability to grow at wide salinity ranges. From the 14 strains whose
growth adaptation to salinity we tested, all except one were able to grow at salinities >5‰
NaCl and exhibited growth optima ranging to 20‰ or higher (Table 1). Four of the 14 isolates
had as closest relative a strain also isolated from a marine habitat (GWS-BW-H301M, GWS-
AG-H268, GWS-SE-H117, GWS-TZ-H232, Table 3) and two of the latter isolates grew only
at salinities >5‰ NaCl. The fact that the next relatives of the other isolates were not of marine
origin is no indication that our isolates growing at higher salinity ranges were not truly marine
strains. Aeromicrobium marinum, the recently described marine species within the family
Nocardioidaceae, clusters with terrestrial isolates but exhibits a requirement for seawater

Kapitel IV Diversity and abundance of Gram-positive bacteria
92
typical of marine bacteria (Bruns et al., 2003). Hence, these marine species clustering closely
together with other non-marine species may indicate that they were introduced into coastal
habitats from adjacent soil and freshwater habitats. This notion underlines the significance of
the close interactions of the land sea transition zone for evolutionary processes and may
further explain the great diversity of Gram+ bacteria occurring in estuarine and coastal
environments.
Four of the five phylotypes we obtained from the clone library and the DGGE-bands
of the marine samples clustered closely together with other phylotypes of exclusively marine
origin and only distantly related to phylotypes or isolates of freshwater or soil origin (Fig. 4).
Because none of the isolates appeared as a prominent DGGE band with the Actinobacteria-
specific PCR, the in situ dominant components of the Gram+ bacterial community
presumably constituted of these yet uncultured phylotypes to a great extent. Obviously the
community of Gram+ bacteria in coastal environments consists of two fractions, one
including the cultivated strains and one including only not-yet cultivated phylotypes, mainly
affiliated to the Marine Actinobacteria Clade (Rappé et al., 1999), and whose physiological
traits are basically unknown. Whereas the indigenous community appears to be most
important for the turnover of organic matter at ambient conditions, the cultivable fraction
appears as a valuable resource for isolates capable of interesting catalytic pathways and for
bioactive compounds (Grossart et al., 2004; Bull et al., 2005; Maldonado et al., 2005).
Our analysis of the community of Gram+ bacteria in the Wadden Sea shows that it
was surprisingly divers, mainly consisting of various groups of Actinobacteria, and to a much
lesser extent of Firmicutes. Phylotypes and isolates clustered to distinctly different groups.
The phylogenetic affiliation of the phylotypes and the broad salinity range of most of the
isolates indicates that the Gram+ bacterial community in the Wadden Sea is well adapted and
indigenous to the marine environment. On the basis of the CARD-FISH results we estimate
that Gram+ bacteria constitute around 5% of total bacteria in the water column of the Wadden
Sea. Hence, they appear to be prominent members of the bacterial communities and, because
of their high potential to degrade various biopolymers, are important in the turnover and
decomposition of organic matter in this ecosystem.

Kapitel IV Diversity and abundance of Gram-positive bacteria
93
Experimental procedures
Sampling
Samples were collected on 27 May and 25 October 1999 in the East Frisian Wadden Sea,
Germany (station A, 53° 37´ N, 07° 08´ E; station B 53° 42´ N, 07° 43´ E; Fig 1). Water
samples were taken at high tide with pre-rinsed 10 l-plastic jugs. Sediment cores from an
intertidal mixed sand/mud flat were taken only in October with Plexiglas tubes (36 mm
diameter) at low tide. Samples were brought to the lab on ice in cooling boxes and processed
further within 2 h. Water samples for CARD-FISH and DGGE analysis were taken on 17
August 2005 at five sampling points along a salinity gradient from 0.3 to 31 psu close to
station B (Fig. 1) and processed further within 3 hours. For DGGE analysis, 100 ml of water
were filtered onto 5.0 µm polycarbonate filters (Nuclepore, Whatman) to obtain particle-
associated bacteria and subsequently onto 0.2 µm polycarbonate filters to obtain free-living
bacteria. Filters were stored at -20°C until nucleic acid extraction.
Isolation of bacteria
For the October samples we applied the MPN (most probable number) technique (Trolldenier,
1993) with different substrates and subsequent isolation of bacteria (Stevens et al., 2005b).
For the May samples dilution series were used for isolation of bacteria. Therefore, 1 ml of
bulk water was used as inoculum for 10-fold dilution series. Mineral media amended with
various substrates and MB 2216 (Difco, Germany) were prepared as described previously
(Stevens et al., 2005b). The following substrates were added (0.1%): agar, alginate, casein,
cellulose, chitin, laminarin, dried and pestiled Fucus vesiculosus (a brown algae growing
copiously along the coast line of the Wadden Sea), palmitate, starch and stearate. Growth was
checked by turbidity and microscopically. Bacteria were isolated from various dilution steps
on agar plates containing the same media as in the MPN assays amended with 1.5% agar. For
further cultivation Marine Agar 2216 (Difco, Germany) was used. Additionally, three isolates
were obtained from 1 l-rolling tanks filled with natural seawater and amended with 0.1% agar
and alginate, respectively, and incubated for 100 days at 15 °C in the dark. Single colonies
were transferred at least five times until considered as pure. The purity of the isolated strains
was examined by DGGE analysis according to Brinkhoff and Muyzer (1997). Isolates from
the same habitat and from assays with the same substrate with sequence similarities of ≥99%
[as determined by a similarity matrix calculated with ARB (Ludwig et al., 2004)] were

Kapitel IV Diversity and abundance of Gram-positive bacteria
94
considered identical and only one sequence, either from the highest dilution step or from the
highest number of sequenced bp, was used in this study and submitted to GenBank.
Salinity dependent growth
In order to examine growth adaptation to the ambient salinity range 14 selected isolates from
the major families of Gram+ bacteria (Table 1) were grown on MB agar plates and transferred
to liquid MB medium of a salinity of 20‰ NaCl (w/v). These cultures were used to inoculate
5 ml liquid MB cultures in triplicate test tubes of salinities of 1, 5, 10, 20, 30 and 45‰ NaCl.
Test tubes were incubated in the dark at 20°C and growth was monitored over 4 to 10 days by
examining periodically the optical density at 660 nm spectrophotometrically directly in the
test tubes. Growth rates were determined as the slope of the exponential growth phase of the
log-plotted growth curves.
Fluorescence in situ hybridization (CARD-FISH)
CARD-FISH analyses were carried out basically following the protocol of Pernthaler et al.
(2004). Four replicates of one ml of each sample were filtered onto a 0.2 µm polycarbonate
filter (25 mm diameter, Nuclepore, Whatman) and fixed with 3 ml of paraformaldehyde (4%
w/v) for one hour. Subsequently, filters were embedded in low-gelling point agarose (0.2%
w/v, Metaphor) and incubated in lysozyme (10 mg ml-1) and achromopeptidase (2 U µl-1) as
described by Sekar et al. (2003). Cells were hybridized with the probes HRP-EUB338
(Amann et al., 1990), HRP-HGC69a (Roller et al., 1994) and HRP-NON338 (Wallner et al.,
1993) for 2 hours at 35°C. Amplification was performed at 37°C using FITC-labeled
Tyramide (fluorescein-5-isothiocyanate, Invitrogen) for 30 min in the dark. Samples were
incubated in 1x PBS (pH 7.3) amended with Triton X-100 (0.05%) for 30 min at room
temperature in the dark to remove residual dye. Cells were counterstained using Vectashield
with DAPI (4´,6´-diamidino-2-phenylindol, 1.5 µg ml-1; Vector Laboratories, Peterborough,
UK) and kept frozen at -20°C until further processing. Enumeration was performed by
epifluorescence microscopy (Axiolab, Zeiss, Germany) at 1,000 x magnification.
Nucleic acid extraction
Genomic DNA was extracted after bead-beating by phenol-chloroform as described earlier
(Stevens et al., 2005a). Precipitation of nucleic acids was done overnight at -20°C using
isopropanol. DNA pellets were resuspended in molecular grade water (Eppendorf, Hamburg,
Germany) and stored at -20°C until further processing.

Kapitel IV Diversity and abundance of Gram-positive bacteria
95
PCR amplification of 16S rRNA gene fragments
PCR amplifications were performed with an Eppendorf Mastercycler (Eppendorf) as
described previously (Brinkhoff and Muyzer, 1997). 16S rRNA gene fragments of
Actinobacteria were amplified using the primers S-C-Act-235-a-S-20
(CGCGGCCTATCAGCTTGTTG, forward) and S-C-Act-878-a-A-19
(CCGTACTCCCCAGGCGGGG, reverse, Stach et al., 2003). A GC-clamp was added to the
forward-primer for subsequent DGGE analysis (Muyzer et al., 1998). Amplification was
performed according to Stach et al. (2003) with the following modifications: Denaturing and
annealing were extended from 45 to 60 s and elongation from 1 to 3 min. Touchdown from
72°C to 67°C in 0.5°C steps was done with two instead of one cycle per step and annealing at
68°C was extended from 15 to 20 cycles.
Four µl of the amplification products were analyzed by electrophoresis in 2% (w/v)
agarose gels and stained with ethidium bromide (1 µg ml-1) (Sambrook et al., 1989). For
subsequent sequencing analysis PCR products were purified by using the Qiaquick PCR
purification kit (Qiagen Inc.).
DGGE analysis of PCR products
DGGE was performed with the D-Code system (Bio-Rad Laboratories, Inc.). For the 16S
rRNA gene fragments of Actinobacteria, a gradient of 35 to 85% denaturant was used. After
electrophoresis, the gels were stained with SYBR Gold (Molecular Probes, Inc.) and
photographed using a BioDoc Analyze Transilluminator (Biometra, Göttingen, Germany).
Bands were excised with a scalpel sterilized with ethanol and transferred to sterile Eppendorf
caps. Fifty µl of water (molecular grade, Eppendorf, Germany) were added and the samples
were stored at –20°C.
Clone library construction
A water sample of 250 ml collected in October at station B was filtered onto a 0.2 µm
Nuclepore filter (47 mm diameter). The filter was immediately frozen at –80°C until DNA-
extraction. Bacterial genomic DNA of the sample was isolated after bead beating, phenol-
chloroform extraction, and isopropanol precipitation as described previously (Stahl et al.,
1988; MacGregor et al., 1997), but slightly modified. Lysozyme treatment was not applied,
precipitation done at –20°C and molecular grade water (Eppendorf, Hamburg, Germany)
was used for resuspension at 4°C over night. PCR amplification of almost complete 16S
rRNA gene fragments was performed as previously described (Brinkhoff and Muyzer, 1997)

Kapitel IV Diversity and abundance of Gram-positive bacteria
96
with primers GM3F (8F) and GM4R (1492R) (Muyzer and Ramsing, 1995). Amplification
was done in triplicates and the products were pooled prior to cloning. For cloning the
pGEM®-T Vector System II (Promega, Madison, USA) was used according to the
manufacturer’s instructions. Sequencing of the clones was performed as described previously
(Stevens et al., 2005a). Clone sequences were checked for chimera formation with the
CHECK_CHIMERA software of the Ribosomal Database Project II (Maidak et al., 2001).
Sequencing and phylogenetic analysis
PCR amplification of 16S rRNA gene fragments of bacterial isolates and subsequent
sequencing was performed as described before (Brinkhoff and Muyzer, 1997). Sequences
were compared with similar sequences of reference organisms by BLAST search
(http://www.ncbi.nlm.nih.gov/blast (Altschul et al., 1998). Phylogenetic analysis was
performed with the ARB software package [http://www.arb-home.de (Ludwig et al., 2004)].
For tree calculation, only sequences with more than 1300 bp were considered using
maximum-likelihood analysis. Shorter sequences were added later to the final tree using the
maximum parsimony option of the ARB program. Alignment positions at which less than
50% of sequences of the entire set of data had the same residues were excluded from the
calculations to prevent uncertain alignments within highly variable positions of the 16S rRNA
gene fragments, which cause mistakes in tree topology (Ludwig et al., 2004). A phylogenetic
analysis of Gram+ bacteria of the Wadden Sea included sequences of the Gram+ isolates
obtained in May and October 1999 (this study), of a clone library and Actinobacteria-specific
DGGE bands (this study), and of previous studies of the water column of the Wadden Sea
using DGGE analysis or cultivation-based methods (Bruns et al., 2003; Brinkhoff et al., 2004;
Grossart et al., 2004; Selje et al., 2005; Stevens et al., 2005a). Sequences obtained from
isolates and clones in this study are available from GenBank under accession no. given in
Table 4.

Kapitel IV Diversity and abundance of Gram-positive bacteria
97
ACKNOWLEDGEMENTS
We are grateful to Andrea Schlingloff for the sequencing, and to H.P. Grossart for the
introduction into the rolling tank incubation method. This work was supported by grants from
the Volkswagen Foundation within the Lower Saxonian priority Program Marine
Biotechnology and by the Deutsche Forschungsgemeinschaft within the research group
BioGeoChemistry of the Wadden Sea (FG-432, TP 5).

Kapitel IV Diversity and abundance of Gram-positive bacteria
98
REFERENCES Allgaier, M., and Grossart, H.P. (2006) Diversity and seasonal dynamics of Actinobacteria in
four lakes of northeastern Germany. Appl Environ Microbiol 72: 3489-3497.
Amann R., Krumholz, L., Stahl, D.A. (1990) Fluorescent-oligonucleotide probing of whole cells for determinative, phylogenetic, and environmental studies in microbiology. J
Bacteriol 172: 762-770
Altschul, S., Madden, T., Schaffer, A., Zhang, J.H., Zhang, Z., Miller, W., and Lipman, D. (1998) Gapped BLAST and PSI-BLAST: A new generation of protein database search proGrams. Faseb Journal 12: A1326-A1326.
Bonde, G.J. (1976) Marine Bacillus. J Appl Bacteriol 41: R7-R8.
Brinkhoff, T., and Muyzer, G. (1997) Increased species diversity and extended habitat range of sulfur-oxidizing Thiomicrospira spp. Appl Environ Microbiol 63: 3789-3796.
Brinkhoff, T., Bach, G., Heidorn, T., Liang, L., Schlingloff, A., and Simon, M. (2004) Antibiotic production by a Roseobacter Clade-affiliated species from the German Wadden Sea and its antagonistic effects on indigenous isolates. Appl Environ Microbiol 70: 2560-2565.
Bruns, A., Phillip, H., Cypionka, H., and Brinkhoff, T. (2003) Aeromicrobium marinum sp. nov., an abundant pelagic bacterium isolated from the German Wadden Sea. Int J Syst
Evol Microbiol 53: 1917-1923.
Bull, A.T., Stach, J.E.M., Ward, A.C., and Goodfellow, M. (2005) Marine actinobacteria: perspectives, challenges, future directions. Antonie van Leeuwenhoek 87: 65-79.
Denis, F. (1971) Bacillus in marine environment - study of 120 strains. Cr Soc Biol 165: 2404-&.
Dittmann, S. (1999) The Wadden Sea ecosystem: stability, properties and mechanisms. Springer- New York, p. 336 pp.
Du, H.L., Jiao, N.Z., Hu, Y.H., and Zeng, Y.H. (2006) Diversity and distribution of pigmented heterotrophic bacteria in marine sediments. FEMS Microbiol Ecol 57: 92-105.
Eilers, H., Pernthaler, J., Glöckner, F.O., and Amann, R. (2000) Culturability and in-situ abundance of pelagic bacteria from the North Sea. Appl Environ Microbiol 66: 3044-3051.
Elifantz, H., Malmstrom, R.R., Cottrell, M.T., and Kirchman, D.L. (2005) Assimilation of polysaccharides and glucose by major bacterial groups in the Delaware estuary. Appl
Environ Microbiol 71: 7799-7805.
Finne, G., and Matches, J.R. (1974) Low-temperature-growing clostridia from marine sediments. Can J Microbiol 20: 1639-1645.
Fuhrman, J.A., McCallum, K., and Davis, A.A. (1993) Phylogenetic diversity of subsurface marine microbial communities from the Atlantic and Pacific Oceans. Appl Environ
Microbiol 59: 1294-1302.
Fuchs, B.M., Woebken, D., Zubkov, M.V., Burkill, P., and Amann, R. (2005) Molecular identification of picoplankton populations in contrasting waters of the Arabian Sea. Aquat
Microb Ecol 39: 145-157.

Kapitel IV Diversity and abundance of Gram-positive bacteria
99
Glöckner, F.O., Zaichikov, E., Belkova, N., Denissova, L., Pernthaler, J., Pernthaler, A., and Amann, R. (2000) Comparative 16S rRNA analysis of lake bacterioplankton reveals globally distributed phylogenetic clusters including an abundant group of Actinobacteria. Appl Environ Microbiol 66: 5053-5065.
Godden, B., and Penninckx, M.J. (1984) Identification and evolution of the cellulolytic microflora present during composition of cattle manure: on the role of Actinomycetes.sp. Ann. Microbiol. (Paris) 135B: 69-78.
Goodfellow, M, and Haynes, J.A. (1984) Actinomycetes in marine sediments. In: Ortiz-Ortiz, L. (ed.), Biological, biochemical and biomedical aspects of actinomycetes. Academic Press, New York, pp. 453-472.
Gray, J.P., and Herwig, R.P. (1996) Phylogenetic analysis of the bacterial communities in marine sediments. Appl Environ Microbiol 62: 4049-4059.
Grossart, H.P., Schlingloff, A., Bernhard, M., Simon, M., and Brinkhoff, T. (2004) Antagonistic activity of bacteria isolated from organic aggregates of the German Wadden Sea. FEMS Microbiol Ecol 47: 387-396.
Hagström, A., Pinhassi, J., and Zweifel, U.L. (2000) Biogeographical diversity among marine bacterioplankton. Aquat Microb Ecol 21: 231-244.
Hahn, M.W., Lunsdorf, H., Wu, Q.L., Schauer, M., Höfle, M.G., Boenigk, J., and Stadler, P. (2003) Isolation of novel ultramicrobacteria classified as actinobacteria from five freshwater habitats in Europe and Asia. Appl Environ Microbiol 69: 1442-1451.
Haider, K., Trojanowski, J., and Sundmann, V. (1978) Screening for lignin degrading bacteria by means of 14C-labelled ligninds. Arch Microbiol 119: 103-106.
Harvey, H.R., and Mannino, A. (2001) The chemical composition and cycling of particulate and macromolecular dissolved organic matter in temperate estuaries as revealed by molecular organic tracers. Org Geochem 32: 527-542.
Haukka, K., Heikkinen, E., Kairesalo, T., Karjalainen. H., and Sivonen, K. (2005) Effect of humic material on the bacterioplankton community composition in boreal lakes and mesocosms. Environ Microbiol 7: 620-630.
Jendrossek, ?? (1997) Bacterial degradation of natural rubber: a privilege of actinomycetes? FEMS Microbiol Lett 150: 179-188.
Jensen, P.R., and Fenical, W. (1995) The relative abundance and seawater requirements of Gram- positive bacteria in near-shore tropical marine samples. Microb Ecol 29: 249-257.
Kirchman, D.L. (2002) The ecology of Cytophaga-Flavobacteria in aquatic environments. FEMS Microbiol Ecol 39: 91-100.
Kirchman, D.L., Dittel, A.I., Malmstrom, R.R., and Cottrell, M.T. (2005) Biogeography of major bacterial groups in the Delaware estuary. Limnol Oceanogr 50: 1697-1706.
Köpke, B., Wilms, R., Engelen, B., Cypionka, H., and Sass, H. (2005) Microbial diversity in coastal subsurface sediments: a cultivation approach using various electron acceptors and substrate gradients. Appl Environ Microbiol 71: 7819-7830.
Linos, A., Berekaa, M.M., Steinbüchel, A., Kim, K.K., Sproer, C., and Kroppensted, R.M. (2002) Gordonia westfalica sp. nov., a new rubber-degrading actinomycete. Int J Syst
Evol Microbiol 52: 1133-1139.

Kapitel IV Diversity and abundance of Gram-positive bacteria
100
Llobet-Brossa, E., Rossello-Mora, R., and Amann, R. (1998) Microbial community composition of Wadden Sea sediments as revealed by fluorescence-in-situ-hybridization. Appl Environ Microbiol 64: 2691-2696.
Ludwig, W., Strunk, O., Westram, R., Richter, L., Meier, H., Yadhukumar et al. (2004) ARB: a software environment for sequence data. Nuc Acids Res 32: 1363-1371.
Lunau, M., Lemke, A., Dellwig, O., and Simon, M. (2006) Physical and biogeochemical controls of microaggregate dynamics in a tidally affected coastal ecosystem. Limnol
Oceanogr 51: 847-859.
MacGregor, B.J., Moser, D.P., Alm, E.W., Nealson, K.H., and Stahl, D. (1997) Crenarchaeota in Lake Michigan sediment. Appl Environ Microbiol 63: 1178-1181.
Maidak, B.L., Cole, J.R., Lilburn, T.G., Parker, C.T., Saxman, P.R., Farris, R.J. et al. (2001) The RDP-II (Ribosomal Database Project). Nuc Acids Res 29: 173-174.
Maldonado, L.A., Stach, J.E.M., Pathom-aree, W., Ward, A.C., Bull, A.T., and Goodfellow, M (2005) Diversity of cultivable actinobacteria in geographically widespread marine sediments. Antonie van Leeuwenhoek 87: 11-18.
Marty, D.G. (1986) Description of 23 cellulolytic and non-cellulolytic marine clostridia. Ann
Inst Pasteur Mic 137A: 33-43.
Mincer, T.J., Jensen, P.R., Kauffman, C.A., and Fenical, W. (2002) Widespread and persistent populations of a major new marine actinomycete taxon in ocean sediments. Appl Environ
Microbiol 68: 5005-5011.
Moran, M.A., Rutherford, L.T., and Hodson, R.E. (1995) Evidence for indigenous Streptomyces populations in a marine environment determined with a 16S ribosomal-RNA probe. Appl Environ Microbiol 61: 3695-3700.
Morris, R.M., Vergin, K.L., Cho, J.C., Rappé, M.S., Carlson, C.A. and Giovannoni, S.J. (2005) Temporal and spatial response of bacterioplankton lineages to annual convective overturn at the Bermudas Atlantic Time-series Study site. Limnol Oceanogr 50: 1687-1696.
Morris, R.M., Longnecker, K., and Giovannoni, S.J. (2006) Pirellula and OM43 are among the dominant lineages identified in an Oregon coast diatom bloom. Environ Microbiol 8: 1361-1370.
Muyzer, G., and Ramsing, N.B. (1995) Molecular methods to study the organization of microbial communities. Water Sci Technol 32: 1-9.
Muyzer, G., de Waal, E.C., and Uitterlinden, A.G. (1993) Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl Environ Microbiol 59: 695-700.
Muyzer, G., Brinkhoff, T., Nübel, U., Santegoeds, C., Schäfer, H., and Wawer, C. (1998) Denaturing gradient gel electrophoresis (DGGE) in microbial ecology. In: Akkermans ADL, van Elsas, J.D., Bruijn, F.J. (eds.) Molecular Microbial Ecology Manual. Kluwer Academic Publishers, Dordrecht, p 1-27.
Pathom-aree, W., Stach, J.E.M., Ward, A.C., Horikoshi, K., Bull, A.T., and Goodfellow, M. (2006) Diversity of actinomycetes isolated from Challenger Deep sediment (10,898 m) from the Mariana Trench. Extremophiles 10: 181-189.
Pernthaler, A., Pernthaler, J., and Amann, R. (2004) Sensitive multi-color fluorescence In situ hybridization for the identification of environmental microorganisms. Molecular Microbial Ecology Manual (2nd Edition) 3.11: 711-726.

Kapitel IV Diversity and abundance of Gram-positive bacteria
101
Piza, H.I., Prado, P.I., and Manfio, G.P. (2004). Investigation of bacterial diversity in Brazilian tropical estuarine sediments reveals high actinobacterial diversity. Antonie van
Leeuwenhoek 86: 317-328.
Poremba, K., Tillmann, U., and Hesse, K.J. (1999) Tidal impact on planktonic primary and bacterial production in the German Wadden Sea. Helgol Mar Res 53: 19-27.
Pranamuda, H., Chollakup, R., and Tokiwa, Y. (1999) Degradation of polycarbonate by a polyester-degrading strain, Amycolatopsis sp. strain HT-6. Appl Environ Microbiol 65: 4220-4222.
Rajendran, N., Matsuda, O., Urushigawa, Y., and Simidu, U. (1994) Characterization of microbial community structure in the surface sediment of Osaka Bay, Japan, by phospholipid fatty- acid analysis. Appl Environ Microbiol 60: 248-257.
Rajendran, N., and Nagatomo, Y. (1999) Seasonal changes in sedimentary microbial communities of two eutrophic bays as estimated by biomarkers. Hydrobiologia 393: 117-125.
Rappé, M., Kemp, P., and Giovannoni, S. (1997) Phylogenetic diversity of marine coastal picoplankton 16S rRNA genes cloned from the continental shelf off Cape Hatteras, North Carolina. Limnol Oceanogr 42: 811-826.
Rappé, M.S., Gordon, D.A., Vergin, K.L., and Giovannoni, S.J. (1999) Phylogeny of actinobacteria small subunit (SSU) rRNA gene clones recovered from marine bacterioplankton. Syst Appl Microbiol 22: 106-112.
Ravenschlag, K., Sahm, K., Pernthaler, J., and Amann, R. (1999) High bacterial diversity in permanently cold marine sediments. Appl Environ Microbiol 65: 3982-3989.
Reichenbach, H. (1992) The Order Cytophagales. In: Starr, M. P., H. Stolp, H. G. Trüper, A. Balows, H. G. Schlegel (eds.). The Prokaryotes. 2nd edition, Vol.1, Springer press, N. Y. pp. 3632-3666.
Roller, C., Wagner, M., Amann, R., Ludwig, W., Schleifer, K.-H. (1994) In situ probing of gram-positive bacteria with high DNA G+C content using 23S rRNA-targeted oligonucleotides. Microbiology 140: 2849-2859.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989) Northern Hybridisation. Molecular Cloning: a laboratory manual, 2nd ed, Cold Spring Harbour Laboratory Press; New York.
Sekar R., Pernthaler, A., Pernthaler, J., Warnecke, F., Posch, T., and Amann, R. (2003) An improved protocol for quantification of freshwater Actinobacteria by fluorescence in situ hybridization. Appl Environ Microbiol 69: 2928-2935.
Selje, N., Brinkhoff, T., and Simon, M. (2005) Detection of abundant bacteria in the Weser estuary by culture-dependent and culture-independent approaches. Aquat Microb Ecol 39: 17-34.
Siefert, J.L., Larios-Sanz, M., Nakamura, L.K., Slepecky, R.A., Paul, J.H., Moore, E.R.B. et al. (2000) Phylogeny of marine bacillus isolates from the Gulf of Mexico. Curr Microbiol 41: 84-88.
Stach, J. E. M., Maldonado, L. A., Ward, A. C., Goodfellow, M., and Bull, A. T. (2003) New primers specific for Actinobacteria: application to marine and terrestrial environments. Environ. Microbiol. 5:828-841.
Stackebrandt, E., Rainey, F.A., and WardRainey, N.L. (1997) Proposal for a new hierarchic classification system, Actinobacteria classis nov. Int J Syst Bacteriol 47: 479-491.

Kapitel IV Diversity and abundance of Gram-positive bacteria
102
Stahl, D.A., Flesher, B., Mansfield, H.R., and Montgomery, L. (1988) Use of phylogenetically based hybridization probes for studies of ruminal microbial ecology. Appl Environ Microbiol 54: 1079-1084.
Stepanauskas, R., Moran, M.A., Bergamaschi, B.A., and Hollibaugh, J.T. (2003) Covariance of bacterioplankton composition and environmental variables in a temperate delta system. Aquat Microb Ecol 31: 85-98.
Stevens, H., Brinkhoff, T., and Simon, M. (2005a) Composition of free-living, aggregate- and sediment surface-associated bacterial communities in the German Wadden Sea. Aquat
Microb Ecoly 38: 15-30.
Stevens, H., Stübner, M., Simon, M., Brinkhoff, T. (2005b) Phylogeny of Proteobacteria and Bacteroidetes from oxic habitats of a tidal flat ecosystem. FEMS Microbiol Ecol 54: 351-364.
Suzuki, M.T., Rappé, M.S., Haimberger, Z.W., Winfield, H., Adair, N., Strobel, J., and Giovannoni, S.J. (1997) Bacterial diversity among small-subunit rRNA gene clones and cellular isolates from the same seawater sample. Appl Environ Microbiol 63: 983-989.
Timmis, K., Hobbs, G., and Berkeley, R.C. (1974) Chitinolytic clostridia isolated from marine mud. Can J Microbiol 20: 1284-1285.
Trolldenier, G. (1993) Bestimmung physiologischer Gruppen nach der MPN-Methode. Schinner F., Öhlinger R., Kandeler E., and Margesin R. (eds.) Bodenbiologische Arbeitsmethoden, 2nd edition. Springer-Verlag, Berlin, Germany: 30-32.
Uphoff, H.U., Felske, A., Fehr, W., and Wagner-Döbler, I. (2001) The microbial diversity in picoplankton enrichment cultures: a molecular screening of marine isolates. FEMS
Microbiol Ecol 35: 249-258.
Urakawa, H., Kita-Tsukamoto, K., and Ohwada, K. (1999) Microbial diversity in marine sediments from Sagami Bay and Tokyo Bay, Japan, as determined by 16S rRNA gene analysis. Microbiology 145: 3305-3315.
Wallner, G., Amann, R., and Beisker, W. (1993) Optimizing fluorescent in situ hybridization with rRNA-targeted oligonucleotide probes for flow cytometric identification of microorganisms. Cytometry 14: 136–143.
Warnecke, F., Amann, R., and Pernthaler, J. (2004) Actinobacterial 16S rRNA genes from freshwater habitats cluster in four distinct lineages. Environ Microbiol 6: 242-253.
Wilms, R., Köpke, B., Sass, H., Chang, T.S., Cypionka, H., Engelen, B. (2006) Deep biosphere-related bacteria within the subsurface of tidal flat sediments. Environ
Microbiol 8: 709-719.
Zobell, C.E., and Upham., H.C. (1944) A list of marine bacteria including descriptions of sixty new species. Bull Scripps Inst Oceanogr Univ Calif 5: 239-292.
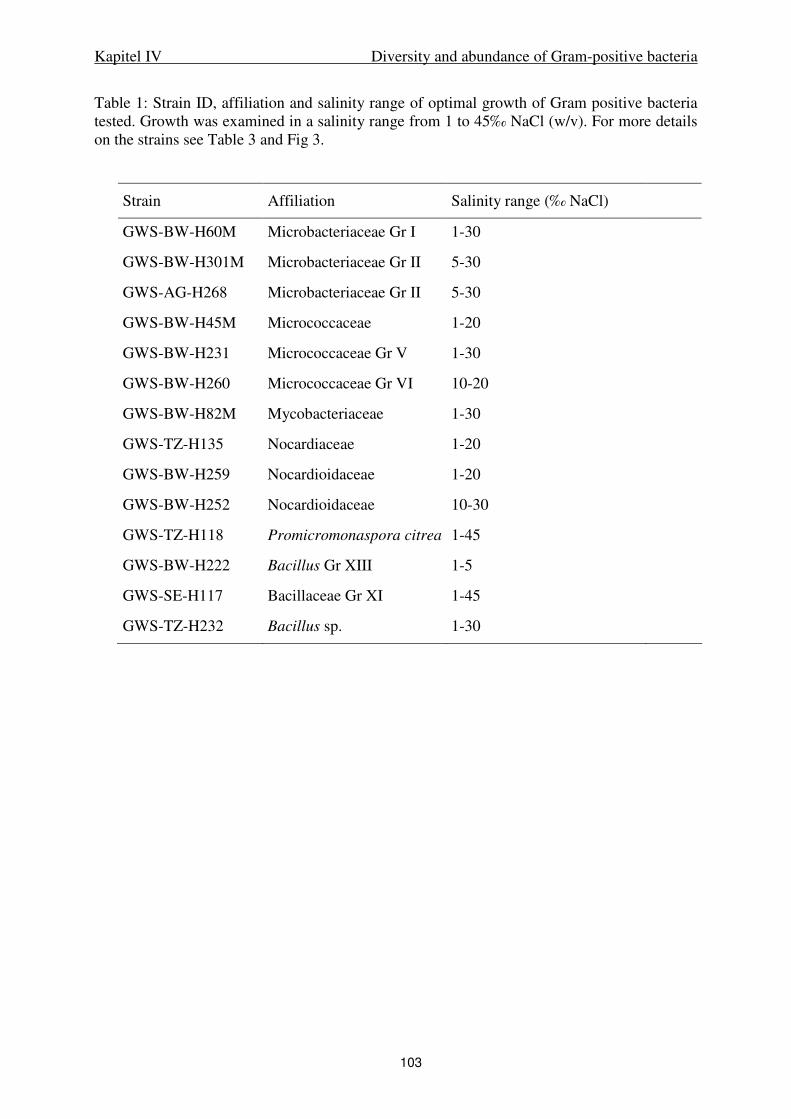
Kapitel IV Diversity and abundance of Gram-positive bacteria
103
Table 1: Strain ID, affiliation and salinity range of optimal growth of Gram positive bacteria tested. Growth was examined in a salinity range from 1 to 45‰ NaCl (w/v). For more details on the strains see Table 3 and Fig 3.
Strain Affiliation Salinity range (‰ NaCl)
GWS-BW-H60M Microbacteriaceae Gr I 1-30
GWS-BW-H301M Microbacteriaceae Gr II 5-30
GWS-AG-H268 Microbacteriaceae Gr II 5-30
GWS-BW-H45M Micrococcaceae 1-20
GWS-BW-H231 Micrococcaceae Gr V 1-30
GWS-BW-H260 Micrococcaceae Gr VI 10-20
GWS-BW-H82M Mycobacteriaceae 1-30
GWS-TZ-H135 Nocardiaceae 1-20
GWS-BW-H259 Nocardioidaceae 1-20
GWS-BW-H252 Nocardioidaceae 10-30
GWS-TZ-H118 Promicromonaspora citrea 1-45
GWS-BW-H222 Bacillus Gr XIII 1-5
GWS-SE-H117 Bacillaceae Gr XI 1-45
GWS-TZ-H232 Bacillus sp. 1-30
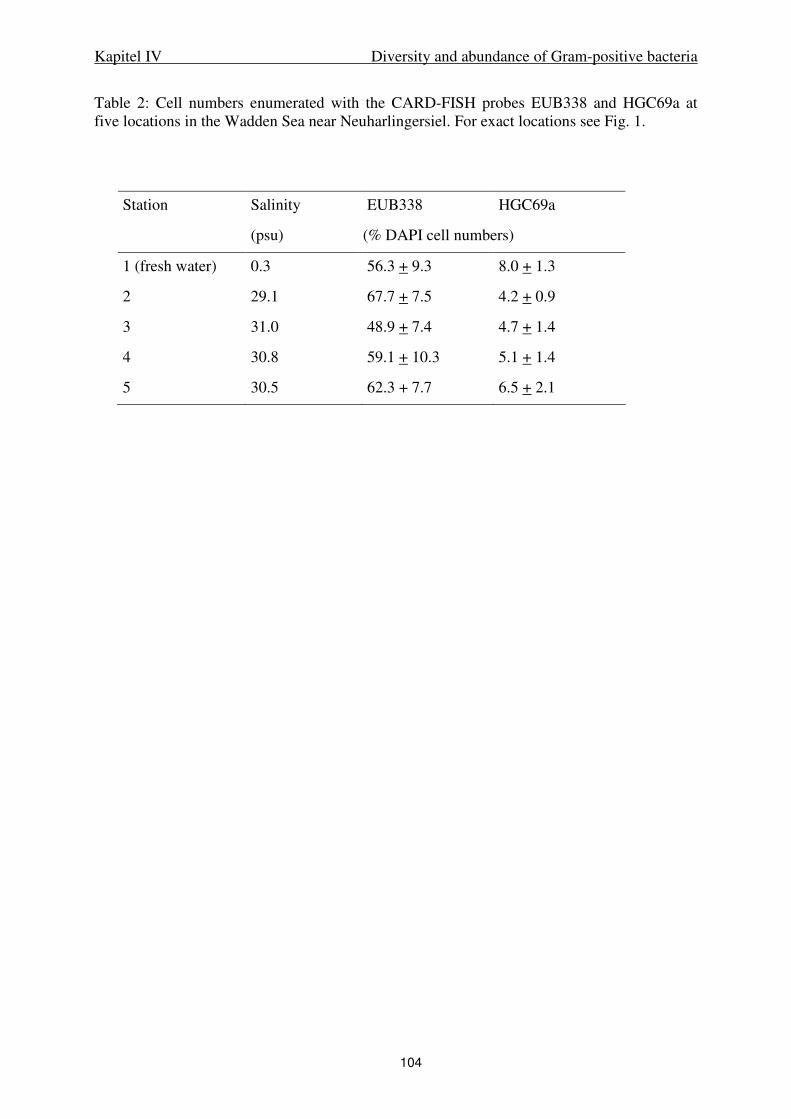
Kapitel IV Diversity and abundance of Gram-positive bacteria
104
Table 2: Cell numbers enumerated with the CARD-FISH probes EUB338 and HGC69a at five locations in the Wadden Sea near Neuharlingersiel. For exact locations see Fig. 1.
Station Salinity EUB338 HGC69a
(psu) (% DAPI cell numbers)
1 (fresh water) 0.3 56.3 + 9.3 8.0 + 1.3
2 29.1 67.7 + 7.5 4.2 + 0.9
3 31.0 48.9 + 7.4 4.7 + 1.4
4 30.8 59.1 + 10.3 5.1 + 1.4
5 30.5 62.3 + 7.7 6.5 + 2.1

Kapitel IV Diversity and abundance of Gram-positive bacteria
105
Table 2: Clones and isolates obtained from the German Wadden Sea affiliating with Gram+ bacteria as well as their closest relatives determined by BLAST analysis (http://www.ncbi.nlm.nih.gov/blast). Sequences with a similarity ≥ 99% were grouped (sequence-groups I - XV), the given information pertains the longest obtained sequence of a sequence group. Given are phylogenetic affiliation, sequence / isolate ID and sequence group where applicable, summarized data on sequence or isolate, closest relative according to BLAST analysis and similarity of the 16S rRNA gene (%), and information concerning the closest relative. Remarks on the isolates give the MPN dilution steps (Dil. [10x]), habitats, substrates, and isolation dates. CAS= casein, CEL= cellulose; CHI= chitin, FUV= Fucus
vesiculosus; LAM= laminarin, PAL= palmitate; STA= starch, MB = Marine Broth 2216, MB* = Marine Broth 2216 prepared with natural sea water (Grossart et al., 2004). SW = autoclaved natural seawater amended with trace elements and vitamins (Selje et al., 2005), BW= bulk water, AG= aggregate, SE= oxic sediment; TZ= oxic/anoxic transition zone of the sediment; rt= rolling tank.
Phylum / family Sequence / isolate ID
(Representative of seq.- group)
Remarks on isolate / summary of sequence group
(Dil. [10x], habitat, substrate, date)
Closest relative (acc. number) [%] Habitat or environmental features of closest
relative
Actinobacteria
Microbacteriaceae GWS-SE-H242a -2, SE, PAL, Oct 99 Microbacterium sp. OS-6 (AJ296094)
99 coastal marsh (Galicia, Spain)
GWS-AG-H197 -3, AG, CHI, Oct 99 Microbacterium sp. V4.BP.11 (AJ244677)
98 marine bacterioplankton (Mediterranea)
GWS-TZ-H305 -2, TZ, FUV, Oct 99 Microbacterium esteraromaticum
(Y17231) 95 soil
GWS-TZ-H139 -1, ALG, TZ, Oct 99 Microbacterium testaceum SE034 (AF474327)
97 endophytic, agronomic crop
GWS-BW-H60M
(Sequence-group I)
-8, -7, -6, -1; BW, SE, TZ, AGA, CAS, CEL, STA; May, Oct 99
Microbacterium sp. VKM
Ac-2050 (AB042084)
99 plant nematode
GWS-SE-H300 -5, ALG, SE, Oct 99 Microbacterium sp. LB030 (AF474326)
99 endophytic, prairie plant
GWS-SE-H149 -2, CEL, SE, Oct 99 Gram+ bacterium strain 12-8 (AB008510)
99 copiotrophic, urban soil
GWS-BWrt-H97M
(Sequence-group II)
-1, rt; BW, AG, AGA, CAS, MB*, STA; May, Oct 99
Marine bacterium P_wp0234 (AY188942)
98 deep sea sediment/degrading PAH
GWS-SE-H243 -2, SE, PAL, Oct 99 Frigoribacterium faeni (Y18807) 98 psychrophilic, non-marine
Clone GWS- K13 From clone-library, BW, Oct 99
Actinobacterium MWH-Dar4 (AJ565416)
98 0.2 µm filtered freshwater
Sanguibacteraceae GWS-AG-H192 -3; AG, CHI, Oct 99 Cellulomonas fermentans (X79458) 94 municipal dumping site
Promicromonosporacea
e GWS-TZ-H118 -5, TZ, AGA, Oct 99 Cellulomonas sp. IFO16243
(AB023364) 96 no information available
Micrococcaceae GWS-BW-H45M
(Sequence-group III)
-5, -4, -1; BW, ALG, CEL, MB; May 99
Arthrobacter nicotianae SB42 (AJ315492)
97 starter culture (cheese)
GWS-SE-H161 -2, SE, CEL, Oct 99 Bacterium PS32 (AF200218) 99 psychrophilic, marine
GWS-BW-H126
(Sequence-group IV)
-4, -2; BW, SE; ALG, CEL, MB*; Oct 99
Bacterium isolate SI-12 (AJ252579)
99 agricultural soil
GWS-BW-H15M
(Sequence-group V)
-7, -2, -1; BW; CEL, FUV, MB; May, Oct 99
Micrococcus luteus strain Ballarat (AJ409096)
99 activated sludge
HP42 From aggregates Micrococcus sp. V4.MO.20 (AJ244665)
98 marine bacterioplankton (Mediterranea)
GWS-BWrt-H158
(Sequence-group VI)
-4, -2, rt; BW; CEL, STA; May, Oct 99
Kocuria rosea (Y11330) 99 soil and water.
Mycobacteriaceae GWS-BW-H82M -1, BW, MB, May 99 Mycobacterium sp. IP20010961 (AY163341)
99 water supplies
GWS-BW-H50M -1, BW, STA, May 99 Mycobacterium sp. TH-2003 (AY266138)
98 associated with sepsis
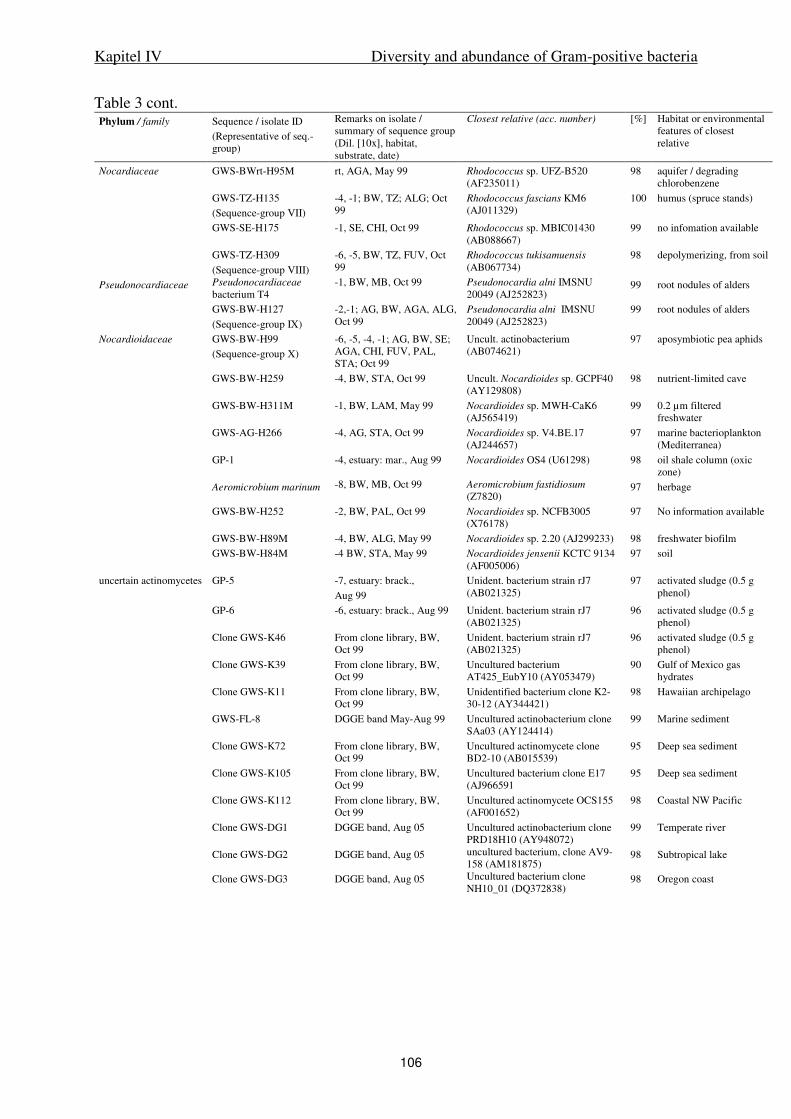
Kapitel IV Diversity and abundance of Gram-positive bacteria
106
Table 3 cont. Phylum / family Sequence / isolate ID
(Representative of seq.- group)
Remarks on isolate / summary of sequence group (Dil. [10x], habitat, substrate, date)
Closest relative (acc. number) [%] Habitat or environmental features of closest relative
Nocardiaceae GWS-BWrt-H95M rt, AGA, May 99 Rhodococcus sp. UFZ-B520 (AF235011)
98 aquifer / degrading chlorobenzene
GWS-TZ-H135
(Sequence-group VII)
-4, -1; BW, TZ; ALG; Oct 99
Rhodococcus fascians KM6 (AJ011329)
100 humus (spruce stands)
GWS-SE-H175 -1, SE, CHI, Oct 99 Rhodococcus sp. MBIC01430 (AB088667)
99 no infomation available
GWS-TZ-H309
(Sequence-group VIII)
-6, -5, BW, TZ, FUV, Oct 99
Rhodococcus tukisamuensis (AB067734)
98 depolymerizing, from soil
Pseudonocardiaceae Pseudonocardiaceae bacterium T4
-1, BW, MB, Oct 99 Pseudonocardia alni IMSNU 20049 (AJ252823)
99 root nodules of alders
GWS-BW-H127
(Sequence-group IX)
-2,-1; AG, BW, AGA, ALG, Oct 99
Pseudonocardia alni IMSNU 20049 (AJ252823)
99 root nodules of alders
Nocardioidaceae GWS-BW-H99
(Sequence-group X)
-6, -5, -4, -1; AG, BW, SE; AGA, CHI, FUV, PAL, STA; Oct 99
Uncult. actinobacterium (AB074621)
97 aposymbiotic pea aphids
GWS-BW-H259 -4, BW, STA, Oct 99 Uncult. Nocardioides sp. GCPF40 (AY129808)
98 nutrient-limited cave
GWS-BW-H311M -1, BW, LAM, May 99 Nocardioides sp. MWH-CaK6 (AJ565419)
99 0.2 µm filtered freshwater
GWS-AG-H266 -4, AG, STA, Oct 99 Nocardioides sp. V4.BE.17 (AJ244657)
97 marine bacterioplankton (Mediterranea)
GP-1 -4, estuary: mar., Aug 99 Nocardioides OS4 (U61298) 98 oil shale column (oxic zone)
Aeromicrobium marinum -8, BW, MB, Oct 99 Aeromicrobium fastidiosum (Z7820)
97 herbage
GWS-BW-H252 -2, BW, PAL, Oct 99 Nocardioides sp. NCFB3005 (X76178)
97 No information available
GWS-BW-H89M -4, BW, ALG, May 99 Nocardioides sp. 2.20 (AJ299233) 98 freshwater biofilm
GWS-BW-H84M -4 BW, STA, May 99 Nocardioides jensenii KCTC 9134 (AF005006)
97 soil
uncertain actinomycetes GP-5 -7, estuary: brack.,
Aug 99
Unident. bacterium strain rJ7 (AB021325)
97 activated sludge (0.5 g phenol)
GP-6 -6, estuary: brack., Aug 99 Unident. bacterium strain rJ7 (AB021325)
96 activated sludge (0.5 g phenol)
Clone GWS-K46 From clone library, BW, Oct 99
Unident. bacterium strain rJ7 (AB021325)
96 activated sludge (0.5 g phenol)
Clone GWS-K39 From clone library, BW, Oct 99
Uncultured bacterium AT425_EubY10 (AY053479)
90 Gulf of Mexico gas hydrates
Clone GWS-K11 From clone library, BW, Oct 99
Unidentified bacterium clone K2-30-12 (AY344421)
98 Hawaiian archipelago
GWS-FL-8 DGGE band May-Aug 99 Uncultured actinobacterium clone SAa03 (AY124414)
99 Marine sediment
Clone GWS-K72 From clone library, BW, Oct 99
Uncultured actinomycete clone BD2-10 (AB015539)
95 Deep sea sediment
Clone GWS-K105 From clone library, BW, Oct 99
Uncultured bacterium clone E17 (AJ966591
95 Deep sea sediment
Clone GWS-K112 From clone library, BW, Oct 99
Uncultured actinomycete OCS155 (AF001652)
98 Coastal NW Pacific
Clone GWS-DG1 DGGE band, Aug 05 Uncultured actinobacterium clone PRD18H10 (AY948072)
99 Temperate river
Clone GWS-DG2 DGGE band, Aug 05 uncultured bacterium, clone AV9-158 (AM181875)
98 Subtropical lake
Clone GWS-DG3 DGGE band, Aug 05 Uncultured bacterium clone NH10_01 (DQ372838)
98 Oregon coast
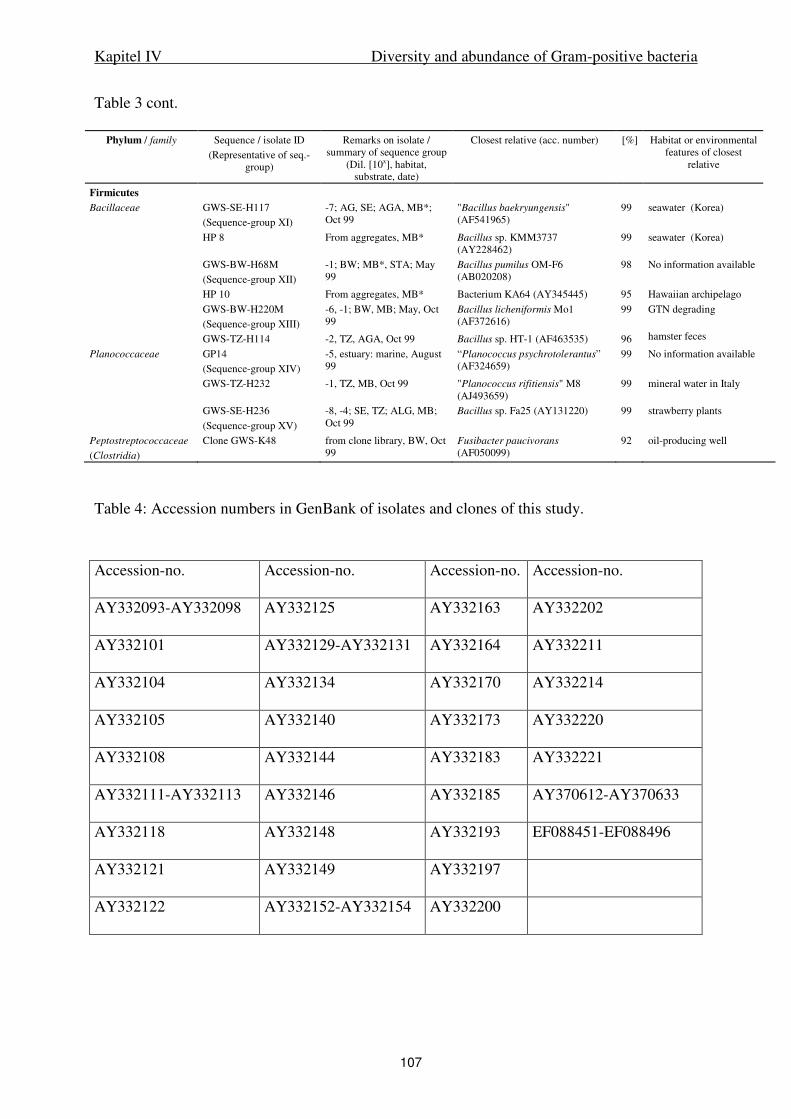
Kapitel IV Diversity and abundance of Gram-positive bacteria
107
Table 3 cont.
Phylum / family Sequence / isolate ID
(Representative of seq.- group)
Remarks on isolate / summary of sequence group
(Dil. [10x], habitat, substrate, date)
Closest relative (acc. number) [%] Habitat or environmental features of closest
relative
Firmicutes
Bacillaceae GWS-SE-H117
(Sequence-group XI)
-7; AG, SE; AGA, MB*; Oct 99
"Bacillus baekryungensis" (AF541965)
99 seawater (Korea)
HP 8 From aggregates, MB* Bacillus sp. KMM3737 (AY228462)
99 seawater (Korea)
GWS-BW-H68M
(Sequence-group XII)
-1; BW; MB*, STA; May 99
Bacillus pumilus OM-F6 (AB020208)
98 No information available
HP 10 From aggregates, MB* Bacterium KA64 (AY345445) 95 Hawaiian archipelago
GWS-BW-H220M
(Sequence-group XIII)
-6, -1; BW, MB; May, Oct 99
Bacillus licheniformis Mo1 (AF372616)
99 GTN degrading
GWS-TZ-H114 -2, TZ, AGA, Oct 99 Bacillus sp. HT-1 (AF463535) 96 hamster feces
Planococcaceae GP14
(Sequence-group XIV)
-5, estuary: marine, August 99
“Planococcus psychrotolerantus” (AF324659)
99 No information available
GWS-TZ-H232 -1, TZ, MB, Oct 99 "Planococcus rifitiensis" M8 (AJ493659)
99 mineral water in Italy
GWS-SE-H236
(Sequence-group XV)
-8, -4; SE, TZ; ALG, MB; Oct 99
Bacillus sp. Fa25 (AY131220) 99 strawberry plants
Peptostreptococcaceae
(Clostridia)
Clone GWS-K48 from clone library, BW, Oct 99
Fusibacter paucivorans (AF050099)
92 oil-producing well
Table 4: Accession numbers in GenBank of isolates and clones of this study.
Accession-no. Accession-no. Accession-no. Accession-no.
AY332093-AY332098 AY332125 AY332163 AY332202
AY332101 AY332129-AY332131 AY332164 AY332211
AY332104 AY332134 AY332170 AY332214
AY332105 AY332140 AY332173 AY332220
AY332108 AY332144 AY332183 AY332221
AY332111-AY332113 AY332146 AY332185 AY370612-AY370633
AY332118 AY332148 AY332193 EF088451-EF088496
AY332121 AY332149 AY332197
AY332122 AY332152-AY332154 AY332200

Kapitel IV Diversity and abundance of Gram-positive bacteria
108
Figure legends
Fig. 1:
Locations of sample collection for the isolates (A, B), the clone library (B), and the DGGE
and CARD-FISH analyses (1-5) in the Wadden Sea (lower panel). Station 1 is a fresh water
drainage channel and stations 2-5 are marine. Between stations 1 and 2 is a sluice.
Fig. 2:
Numbers of Gram+ strains obtained in May and October 1999 from various dilution steps of
dilution cultures of bulk water samples (May and October), of suspended aggregates
(October), the sediment surface (October), and the oxic-anoxic transition zone in the sediment
(October).
Fig. 3:
Banding patterns of an Actinobacteria-specific DGGE analysis of free-living (FL) and
particle-associated bacterial communities (PA) collected at stations 1 to 5 in the vicinity of
station B in the Wadden Sea. For location of station B see Fig. 1. Arrows indicate bands
excised for sequencing.
Fig. 4:
Maximum likelihood trees of all Gram+ isolates and clones obtained from the Wadden Sea
(bold) showing the affiliation within the Actinobacteria (A) and the Firmicutes (B).
Sequences <1300 bp were added with maximum parsimony. The scale bars indicate 10%
sequence divergence. The Marine Actinobacteria Clade was adopted from Rappé et al. (1999)
and the MAR 1 cluster from Mincer et al. ( 2002). Isolates from May are marked with an "M"
at the end of the name. If available, dilution step and substrate were added to the accession
number (CAS = casein, CEL = cellulose; CHI = chitin, FUV = Fucus vesiculosus; LAM =
laminarin, PAL = palmitate; STA = starch, MB = Marine Broth 2216, MB* = Marine Broth
2216 prepared with natural sea water (Grossart et al., 2004), SW = autoclaved natural
seawater amended with trace elements and vitamins (Selje et al., 2005). The sub-habitat can
be derived from the name of the May and October isolates (BW = bulk water, AG =
aggregates, SE = sediment surface, TZ = oxic /anoxic transition zone of the sediment).
Numbers on branches with pooled sequences indicate the number of sequences used to
calculate the cluster.
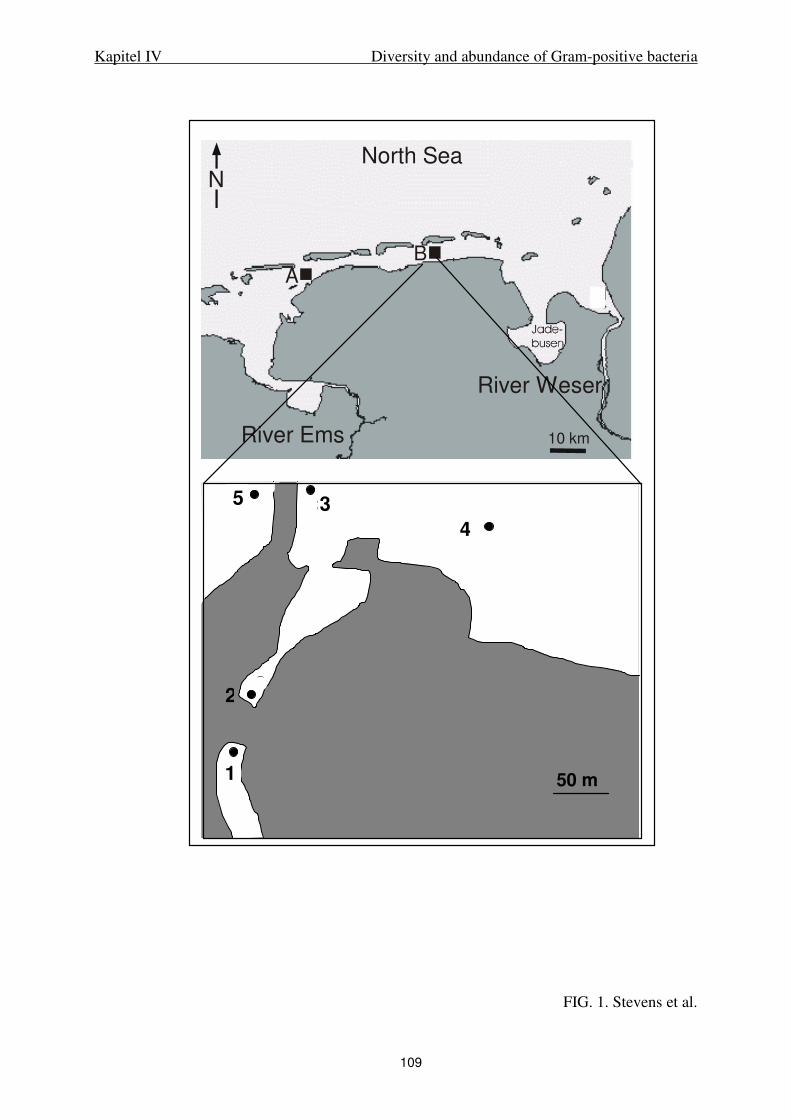
Kapitel IV Diversity and abundance of Gram-positive bacteria
109
FIG. 1. Stevens et al.
River Ems
River Weser
North Sea
N
10 km
A
B
2
53
4
1
2
53
4
1
50 m
5
2
3
4
1
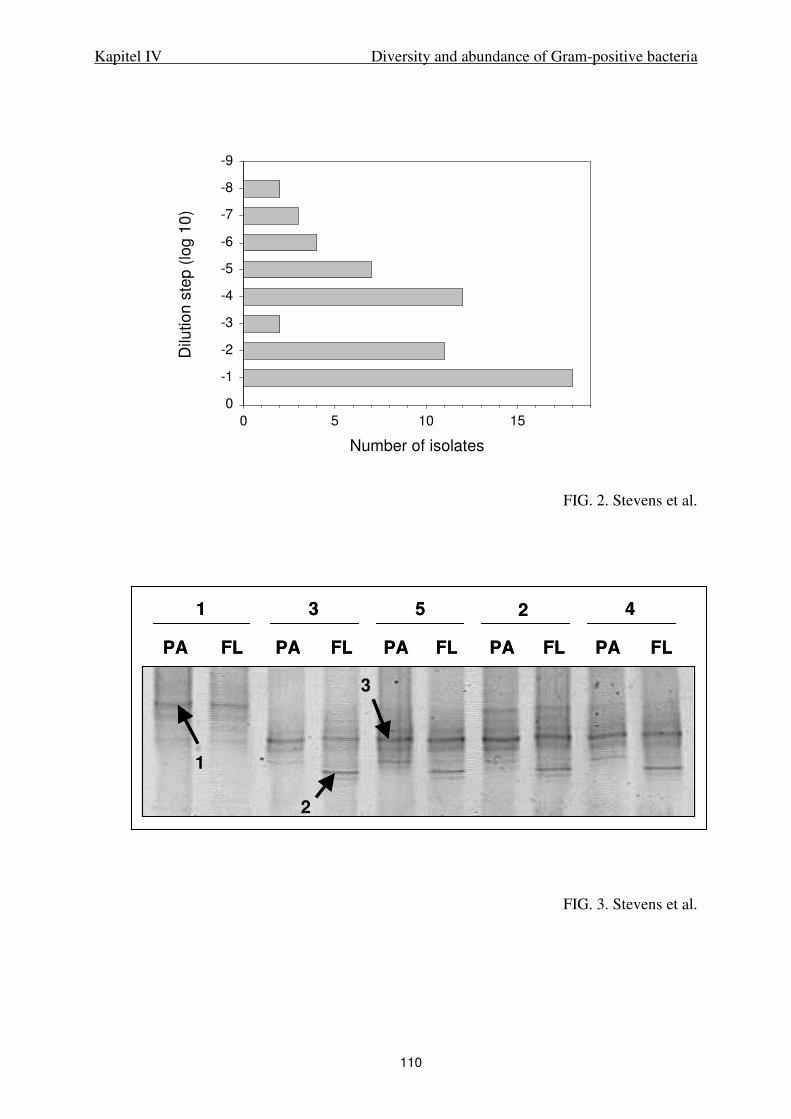
Kapitel IV Diversity and abundance of Gram-positive bacteria
110
FIG. 2. Stevens et al.
FIG. 3. Stevens et al.
Number of isolates
0 5 10 15
Dilu
tio
n s
tep
(lo
g 1
0)
-9
-8
-7
-6
-5
-4
-3
-2
-1
0
42531
PA FL PA FL PA FL PA FL PA FL
1
2
3
42531
PA FL PA FL PA FL PA FL PA FL
1
2
3

Kapitel IV Diversity and abundance of Gram-positive bacteria
111
FIG. 4A, Stevens et al.

Kapitel IV Diversity and abundance of Gram-positive bacteria
112
FIG. 4A cont., Stevens et al.
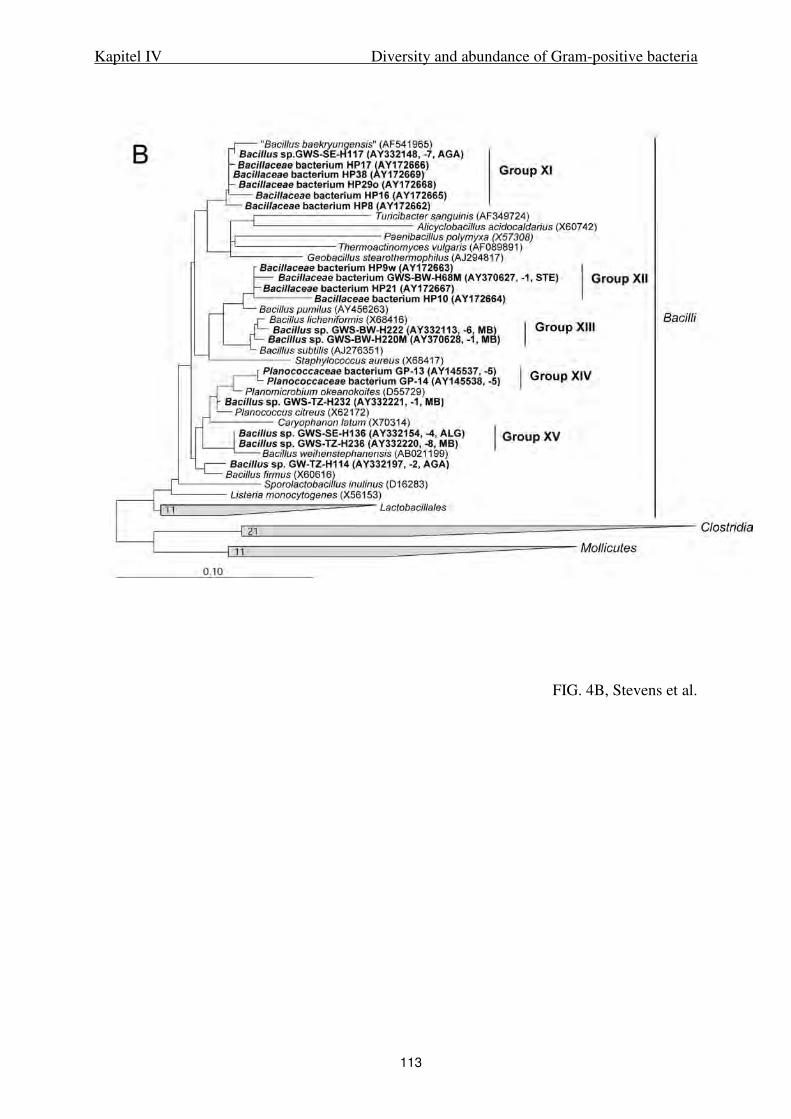
Kapitel IV Diversity and abundance of Gram-positive bacteria
113
FIG. 4B, Stevens et al.

114
V.
High regional variability of bacterial communities
in the German Bight, North Sea

Kapitel V Regional variability of bacterial communities in the German Bight
115
High regional variability of bacterial communities
in the German Bight, North Sea
Beate Rink, Thorsten Brinkhoff, Katja Ziegelmüller, Meinhard Simon*
Institute for Chemistry and Biology of the Marine Environment (ICBM),
University of Oldenburg, D-26111 Oldenburg, Germany
Running title: Bacteria in the North Sea
Key words: North Sea, free-living and attached bacteria, Roseobacter, DGGE, phytoplankton
___________________________________________________________________________
* Corresponding author. Institute for Chemistry and Biology of the Marine Environment
(ICBM), University of Oldenburg, PO Box 2503, D-26111 Oldenburg, Germany,
Phone: +49-441-798-5361. Fax: +49-441-798-3438. E-mail: [email protected]

Kapitel V Regional variability of bacterial communities in the German Bight
116
ABSTRACT
The German Bight of the North Sea is characterized by near shore tidal flat regions with high
loads of suspended matter and estuarine inputs of organic matter and pelagic off shore
regions. Due to tidal and wind-induced currents its hydrography is highly dynamic. In order to
examine how these highly dynamic properties affect the regional distribution and composition
of the bacterioplankton we conducted two surveys in June in two consecutive years during
which we assessed the composition of the free-living (FL, 0.2-5.0 µm fraction) and particle-
associated (PA, >5.0 µm fraction) bacterial communities on the background of hydrographic
(salinity, temperature) and biogeochemical properties (suspended matter, particulate organic
carbon, chlorophyll, phytoplankton composition). The composition of the bacterial
communities was determined by denaturing gradient gel electrophoretic (DGGE) analysis of
16S rRNA gene fragments PCR-amplified by Bacteria-, α-Proteobacteria and Bacteroidetes-
specific primer sets and subsequent sequencing of excised bands.
The results showed that the FL-bacterial community was rather evenly distributed in the
German Bight irrespective of the regional hydrographic and biogeochemical differences.
Several prominent bands, identified as phylotypes affiliated to the Roseobacter clade of α-
Proteobacteria, persisted throughout all 10 stations visited. The PA bacterial community
exhibited distinct differences among the various stations. These differences were not simply
attributed to properties of the near shore tidal flat regions and to the more homogeneous
hydrographic situation of the off shore region. They were rather site-specific, obviously
reflecting local conditions of the phytoplankton present and its growth phase and the
resuspended particles in the tidal flat regions. The results of the PA bacterial community
showed that unspecific PCR-amplifications were obtained by the Bacteria– (chloroplasts) and
α-Proteobacteria–specific primer sets (δ-Proteobacteria), biasing the results to a certain
extent. Because one primer applied for amplifying α-proteobacterial 16S rRNA gene
fragments (ALF968) is frequently used as a probe in fluorescence in situ hybridization (FISH)
analyses, its application leads to overestimates of α-Proteobacteria in samples containing δ-
Proteobacteria.

Kapitel V Regional variability of bacterial communities in the German Bight
117
INTRODUCTION
In aquatic environments, complex communities of free-living and particle-associated
heterotrophic bacteria are the main decomposers of dissolved (DOC) and particulate organic
carbon (POC) and play a key role in the global carbon cycle (Cotner & Biddanda 2002). It has
been shown that origin and composition of organic matter affect the composition and biomass
production of bacterial communities (Covert & Moran 2001, Crump et al. 2003, Lebaron et al.
1999) and that phylogenetic bacterioplankton groups exhibit distinct preferences for low and
high molecular weight carbon sources (Cottrell & Kirchman 2000). Marine ecosystems are
structured into the open sea and coastal environments including the polyhaline estuarine
regions, and the environmental conditions in these habitats comprise different physical and
biogeochemical properties which may affect the ambient microbial communities. In the
coastal regions of the southern North Sea, the German Bight, high loads of dissolved and
particulate inorganic and organic matter are introduced from the tidal flats and the rivers
Weser and Elbe, thus providing organic and inorganic nutrients as well as refractory organic
matter (Loewe et al. 2005). An easterly current follows the southern coastal line along the
East Frisian Islands and the Weser estuary and encounters the polyhaline plume of the Elbe
estuary. Both water masses circulate along the north Frisian coast, often building separate
layers of distinct salinities or, in the case of persisting strong winds, a salinity gradient.
Because of the shallow water depth (10 – 40 m), the variable river discharge and because of
often variable winds the extent and position of the different water masses may change
considerably even on a short-term scale.
Various aspects of the North Sea bacterioplankton have been the scope of quite a few
investigations, e.g. the culturability of pelagic bacteria (Eilers et al. 2001), seasonal and
interannual dynamics and abundance of specific phylogenetic groups (Eilers et al. 2000,
Gerdts et al. 2004) and the composition of bacterial communities as a function of bacterial
respiration and growth (Reinthaler et al. 2005). The spatial distribution of the bacterial
community composition, e.g. in and off shore gradients including the tidal flat areas and
pelagic regions has not been considered. It is not known how the diversity of the bacterial
communities varies within and among the various water bodies mentioned above, possibly as
a function of a patchy distribution of phytoplankton blooms and the strong tidal currents, or
whether the community composition remains unaffected. The strong currents within the
German Bight and the generally shallow water depth may also lead to a well mixed situation,
preventing the establishment of pronounced regional differences of the bacterial community

Kapitel V Regional variability of bacterial communities in the German Bight
118
composition. Obtaining insight into such regional distributions of the composition of bacterial
communities is also important for designing sampling strategies for future investigations,
linking the community composition to hydrographical and biogeochemical processes, not
only in the German Bight, but also in other shallow coastal marine regions exhibiting strong
currents, and estuarine and terrestrial inputs of dissolved and particulate matter.
We investigated the bacterial communities at various near shore and off shore stations
in the German Bight together with properties to characterize the suspended particulate matter
(SPM) and phytoplankton. We used denaturing gradient gel electrophoresis (DGGE) of 16S
rRNA gene fragments and applied primer sets specific for Bacteria, α-Proteobacteria and the
Bacteroidetes phylum.
MATERIALS AND METHODS
Study area and sampling. Surface water samples were collected at various locations in
the German Bight from 11 to 13 June 2002 and from 24 to 27 June 2003 (Fig. 1, Table 1) on
board RV Heincke with a 10 L Niskin bottle. For analysis of suspended matter (SPM) dry
weight (DW), particulate organic carbon (POC), and chlorophyll a (Chl a) 500 to 1000 ml of
sample water were filtered in duplicates on precombusted and preweighed glass fiber filters
(GF/F, Whatman) and stored at –20°C in the dark until further processing in the lab within
four weeks. For enumeration of bacteria 100 ml of seawater were fixed with formaldehyde
(2% v/v) and stored at 4°C until further processing within four weeks. Phytoplankton cells
were fixed with Lugol’s solution as described elsewhere (Utermöhl 1958). For DGGE
analysis, 250 ml of sample water were fractionated by filtration on polycarbonate-filters
(Nuclepore) with pore sizes of 5.0 µm (particle-associated bacteria) and subsequently of 0.2
µm (free-living bacteria) and stored at –20°C until further processing within four months.
Temperature and salinity were recorded by a built-in probe of RV Heincke.
Enumeration of bacteria and algae. Bacteria were enumerated by epifluorescence
microscopy after staining with DAPI (4´-6-diamidino-2-phenylindole) on black 0.2 µm
Nuclepore filters at 1000x magnification (Porter & Feig 1980). We did not differentiate
between free-living (FL) and particle-associated (PA) cells, mainly because it was rather
difficult to reliably enumerate particle-associated cells in the near shore stations with high
concentrations of SPM. When numbers were assessed the reliable desorption technique of PA
bacteria by Lunau et al. (2005) was not yet available. Phytoplankton cells were counted by

Kapitel V Regional variability of bacterial communities in the German Bight
119
inverted microscopy (Utermöhl 1958) and phytoplankton species were identified according to
Drebes (1974).
Phytoplankton pigments, SPM and POC. For chlorophyll analysis filters were
extracted at 75°C in 90% ethanol and concentrations of Chl a were determined by standard
procedures (Parsons et al. 1984). For phaeopigment determination, samples were acidified
with HCl (2N) prior to spectrophotometric analysis. For determination of DW, filters were
dried for 1 hour at 110°C and weighed on a micro-balance (Sartorius, Germany). In 2002,
DW was corrected for salt according to Lunau et al. (2006) and in 2003 filters were rinsed
with distilled H2O. POC was determined with a FlashEA 1112 CHN-analyzer (Thermo
Finnigan).
Nucleic acid extraction, primer sets and PCR amplification of 16S rRNA gene
fragments. Genomic DNA was extracted with phenol-chloroform as described in Rink et al.
(2006a) with slight modifications. DNA was precipitated at –20°C overnight using
isopropanol and resuspended in molecular grade water. Samples were stored at –20°C until
further processing. For the amplification of 16S rRNA gene fragments, primer sets were used
targeting eubacterial DNA (primer pair GC 341F, Muyzer et al. 1993; 907RM, Muyzer et al.
1998), the Bacteroidetes phylum (primer pair GC CF319aF, Jaspers et al. 2001, and 907RM)
and α-Proteobacteria (primer pair GC 341F and ALF 968R, Rink et al. 2006a). Specificity of
the primer sets and the applied PCR conditions are described by Rink et al. (2006a).
Amplification products were analyzed by electrophoresis in 1.5% (w/v) agarose gels and
stained with ethidium bromide (1 µg ml-1) (Sambrook et al. 1989). For subsequent sequence
analysis PCR products were purified by using the Qiaquick PCR purification kit (Qiagen Inc.,
Chatsworth, California).
DGGE analysis of PCR products and cluster analysis. DGGE was performed with an
INGENYphorU system (Ingeny International BV, Leiden, The Netherlands) following the
protocol of Rink et al. (2006a). After electrophoresis, the gels were stained with SYBR Gold
(Molecular Probes, Inc.) and documented using a BioDoc Analyze Transilluminator
(Biometra, Göttingen, Germany). Bands were excised, suspended in 50 µl of water (molecular
grade, Eppendorf, Germany) and centrifuged for 2 min. at 3,000 rpm. Samples were stored at
–20°C and 1 µl was used as template in subsequent PCR reactions. A cluster analysis of the
DGGE banding patterns was performed using the software GelCompare II, Version 2.5
(Applied Maths, St. Martens-Latem, Belgium). We applied 5 to 20% background subtraction
depending on the signal-to-noise ratio of the corresponding gel. Patterns were compared
curve-based using Pearson correlation as similarity coefficient and UPGMA (unpaired group

Kapitel V Regional variability of bacterial communities in the German Bight
120
method of analysis) to generate the dendrogram. We used the position tolerance optimization
option of the software to fit the curves to the best possible matching.
Cloning. 16 DGGE bands were cloned using the pGEM®-T Vector System II (Promega,
Madison, USA) following the instruction manual. At least five clones per DGGE band with
inserts were picked after blue-white-screening and amplified with the specific DGGE primers.
Fragment length of the inserts was screened by agarose gel electrophoresis and positive
inserts were examined for their specific height using DGGE. Adequate clones were amplified
for sequencing using the primers pUC/M13f and pUC/M13r (Sambrook et al. 1989).
Sequencing and phylogenetic analysis. PCR products were sequenced with an
Automated DNA Sequencer (Model 4200, LI-COR Inc.) using the primers 341F and 907RM,
labeled with IRDyeTM800, and the DYEnamic Direct cycle sequencing kit (Amersham Life
Science Inc.). Clones were sequenced by Geneart (Regensburg, Germany) using the primer
M13f. At least 400 bp were determined for all sequences and the phylogenetic affiliation was
compared to those in GenBank using the BLAST function of the NCBI server
(http://www.ncbi.nlm.nih.gov). The phylogenetic trees were constructed using the ARB
software package (http://www.arb-home.de, Ludwig et al. 2004). The backbone tree was
calculated with the maximum likelihood method using sequences with a minimum of 1300 bp
length including type strains of the selected phylogenetic groups. To avoid uncertain
alignments, positions were excluded at which less than 50% of all sequences showed the same
residues. Sequences with less than 1300 bp were added to the backbone tree with the
maximum parsimony method using the same filter. 16S rRNA gene sequences of seven type
strains belonging to Cyanobacteria were used as outgroup.
Nucleotide sequence accession number. The sequences obtained in this study are
available from GenBank under accession no. DQ911759 to DQ911821.

Kapitel V Regional variability of bacterial communities in the German Bight
121
RESULTS
Hydrography. In both years, the off shore stations with depths >17 m (sta 1, 8, 9, 10)
were characterized by lower water temperatures and higher salinities than the near shore
stations (sta 2-7, Fig. 2). At the latter stations water depths is only 8-11 m except at station 2
which exhibits more off shore than near shore properties (see below). In general, temperature
and salinity were higher in 2003 as compared to 2002, when the survey was carried out two
weeks earlier. In 2002, surface water temperatures ranged from 13.7°C (sta 1, 8) to 16.5°C
(sta 7) and in 2003 from 14.4°C (sta 10) to 17.6°C (sta 4). Salinity ranged from 29.6 (sta 7) to
33.2 psu (sta 9) in 2002 and from 31.8 (sta 6) to 35.5 psu (sta 9) in 2003.
Abundance and composition of SPM. In both years, SPM concentrations were higher at
the near shore than at the off shore stations (Fig. 3A, 3B). In 2002, SPM ranged from 4 to 5
mg DW l-1 at the off shore stations and station 2 to 7.6 to 16 mg DW l-1 at the near shore
stations with the highest value at station 5. In 2003, respective values at the off shore stations
(sta 1, 2, 8, 9, 10) were between 2 and 4 mg DW l-1 and at the near shore stations 6.5 to 15.4
mg DW l-1. POC varied from 0.10 mg l-1 at station 9 to 0.78 mg l-1 at station 6 with generally
higher values at the near shore than at the off shore stations (Fig. 3B). At stations 9, 10 and 2
POC constituted around 10% of DW but at the near shore stations generally <6%.
Phytoplankton. Chl a concentrations in 2002 were lower than in 2003 and varied from
0.8 to 4.5 µg l-1 (Fig. 3C, 3D). In 2002, concentrations at the near shore stations were higher
than at the offshore stations except at station 8. In 2003, highest concentrations occurred at
stations 3 and 8 with up to 6.3 µg Chl a l-1 without any clear-cut difference between the near
and off shore stations. Patterns of phaeopigment concentrations, which are only available for
2003, were quite different from that of Chl a (Fig. 3D), indicating that the growth phase and
thus the physiological status of the phytoplankton was quite variable among the stations.
Highest proportions of phaeopgments, close to or exceeding those of Chl a, occurred at
stations 6, 7 and 9. In 2002, Leptocylindricus danicus dominated the phytoplankton at the off
shore stations and Rhizosolenia imbricata at the near shore stations (data not shown). At the
North Frisian coast (sta 6, 7), Guinardia delicatula and Guinardia flaccida were also present
to substantial proportions. In 2003, the phytoplankton at all stations was highly dominated by
Rhizosolenia imbricata except at station 10 which showed a more diverse composition
including substantial proportions of Guinardia spp. (Fig. 3E). Also at station 8, Guinardia
spp. were the second most abundant taxa. Phytoplankton cell numbers generally reflected
concentrations of Chl a with highest values at stations 3 and 8. At station 6 and 7 the high

Kapitel V Regional variability of bacterial communities in the German Bight
122
sediment load in the samples and low phytoplankton cell numbers prevented a reliable
enumeration.
Bacterial abundance. In 2002, bacterial abundance varied between 1.8 x 106cells ml-1 at
station 1 and 3.5 x 106cells ml-1 at station 7. Numbers did not covary with Chl a or SPM. In
2003, the off shore stations exhibited low bacterial numbers and highest numbers were
recorded at stations 3 and 6 together with high concentrations of Chl a and SPM.
DGGE and cluster analysis. Pronounced differences were detected between DGGE
banding patterns of FL and PA bacterial communities. These differences were substantiated
by a cluster analysis (Fig. 4). The application of the Bacteria-specific primer set yielded 7 to
13 DGGE bands of the FL bacterial community in 2002 and 15 to 24 bands in 2003. In the
former year, the lowest and highest number of bands occurred at stations 2 and 1 and in the
latter year at stations 1 and 6. Two prominent bands persisted throughout all stations in both
years, identified as clones GB02-e8-FL, GB02-e9-FL, GB03-e16-FL and GB03-e17-FL
(Table 2). Other bands occurred only at a few or individual stations such as at stations 1 and
10 (GB03-e15-FL), 6 and 7 (GB03-e24-FL, GB03-e25-FL), and 5 (GB03-e23-FL). There was
no band detected only at the off shore or near shore stations. In 2003, stations 6 and 7,
however, exhibited distinctly different patterns than the other stations.
In the PA bacterial community in both years, the number of bands exceeded that of the
FL bacterial community. In 2002, between 10 and 17 bands were detected with lowest and
highest numbers at stations 10 and 1, respectively. In 2003, band numbers ranged between 12
and 28 with lowest and highest numbers at stations 1 and 6. The variability of the PA bacterial
community among the different stations was more pronounced than that of the FL bacterial
community. No single band was detected at all stations. The banding patterns of the PA
bacterial community were highly biased by chloroplast-derived 16S rRNA gene fragments.
Two of the 7 bands of the 2002 samples and 8 of the 14 bands of the 2003 samples sequenced
turned out as chloroplast-like phylotypes.
The cluster analysis substantiated the different banding patterns of the FL- and PA
bacterial communities detected by the Bacteria-specific primer set (Fig. 4). The banding
patterns of the PA bacterial communities exhibited a lower similarity (>55% Pearson
correlation) than those of the FL bacterial communities (>76% Pearson correlation). Even
though micro-clusters occurred, near shore and off shore stations did not exhibit distinct
clusters. Stations 6 and 7, however, formed a separate cluster in both bacterial communities in
2003.

Kapitel V Regional variability of bacterial communities in the German Bight
123
The DGGE analysis of the FL- and PA associated bacterial communities applying the
Bacteroidetes-specific primer set was only done in 2003. The results also revealed
pronounced differences between both communities with a higher diversity in the PA bacterial
community. In the FL bacterial community between 7 (sta 8) and 13 bands (sta 1) were
detected, and in the PA bacterial community between 9 (sta 7) and 17 bands (sta 5). There
was no band which was detected at all stations, neither in the FL- nor in the PA bacterial
community. However, several bands occurred at distinct stations, such as at stations 1-5 and 8
(GB03-c5-PA, Table 3), at stations 2-5 (GB03-c8-FL, Table 2), and at stations 1, 8 and 10
(GB03-c12-FL). In the PA bacterial community, stations 2-5 and 10 clustered together, as did
stations 6 and 7 (Fig. 4). The off shore stations 1, 8 and 9 branched deeply separated. In the
FL fraction, no specific sub-clusters were detected.
The α-Proteobacteria–specific DGGE banding patterns revealed lowest band numbers of
all target groups with 5 to 7 bands in the FL bacterial community and 7 to 16 bands in the PA
bacterial community. In the former community, two conspicuous bands (GB-a14-FL, GB-
a15-FL, Table 2) dominated the banding patterns at all stations and the similarity between the
banding patterns was very high, as confirmed by the cluster analysis (Fig. 4).The banding
patterns of the PA bacterial community were much more diverse. Only stations 6 and 7
clustered together.
Phylogenetic affiliation. Sequencing of the excised DGGE bands obtained from the
Bacteria-specific amplicons showed that 12 bands of a total of 38 contained chloroplast-
derived 16S rRNA gene fragments. Ten of them were detected in the PA fraction. In addition,
13 sequences affiliated to α-, 1 to β, 2 to γ-Proteobacteria and 10 to the Bacteroidetes-
phylum.
All sequences of α-Proteobacteria obtained from the Bacteria-specific DGGE gels
affiliated to the Roseobacter clade except GB02-e3-PA (station 4), which was related most
closely to Acidiphilium aminolytica (Tables 2, 3, Fig. 5A). The phylotypes occurring at all
stations in both years, GB02-e8-FL, GB03-e16-FL, GB02-e9-FL and GB03-e17-FL, affiliated
to the NAC11-7 cluster detected in the North Atlantic (Gonzalez et al. 2000) and the WM11-
36 cluster identified in the polyhaline section of the Weser estuary (Selje & Simon 2003).
The two clones which affiliated to γ-Proteobacteria (GB03-e7-PA, GB03-e10-PA) were
detected in 2003 in the PA bacterial community of station 6 and were closely related to clones
from the East Frisian Wadden Sea (GWS-AG-6, GWS-SE-4, Fig. 5C).
Sequences of DGGE bands obtained with the Bacteroidetes specific primer set revealed 2
unspecific amplifications. DGGE band GB03-c1-PA (sta 1, PA fraction) was related to a

Kapitel V Regional variability of bacterial communities in the German Bight
124
Firmicutes species (Table 3) and band GB03-c6-PA to chloroplast-derived 16S rRNA genes.
The sequences of two DGGE bands obtained from the PA fraction of station 6 (North Frisian
coast, GB03-c3-PA and GB03-c4-PA) were closely related to 16S rRNA gene fragments
detected as a diatom-associated bacterium and in coastal bacterioplankton (GWS-AG-8,
GWS-c2-FL, Fig. 5D), respectively. Clone GB03-c2-PA (sta 6) was closely related to strain
T15, isolated from a high dilution step of a dilution culture from the East Frisian Wadden Sea
(Brinkhoff et al. 2004).
Sequencing of 16S rRNA gene fragments obtained from the α-Proteobacteria specific
DGGE gel showed unspecific amplification in the PA bacterial community. Nine of the 13
sequences obtained were identified as δ-Proteobacteria predominantly originating from
sediments and only 4 as α-Proteobacteria (Fig. 5A and B, Table 3). Two of them affiliated to
the Roseobacter clade, to the RCA cluster (GB03-a5-PA) and to Sulfitobacter pontiacus (GB-
a4-PA), and the other two to Rhodobacterales (GB-a7-PA) and to the genus Sphingomonas
(GB03-a1-PA), respectively. The 4 sequenced bands of the FL bacterial community of the α-
Proteobacteria specific DGGE gel affiliated to the clusters RCA (GB03-a14-FL) and WM11-
36 (GB03-a15-FL, GB03-a16-FL) of the Roseobacter clade and one closely to a phylotype
retrieved from the German Wadden Sea and related to Acidiphilum aminolytica. (GB03-a17-
FL).
DISCUSSION
We found a surprisingly high variability of the DGGE banding patterns both of FL and
PA bacterial communities of either primer set applied in the German Bight of the North Sea
which, however, only partially reflected the clear differences of salinity and SPM
concentrations between the near shore and off shore stations. Similarly, banding patterns did
not reflect patterns in the distribution of Chl a or phytoplankton composition. We did find
distinct differences between banding patterns of PA and FL-bacterial communities,
substantiated by the cluster analysis. A number of phylotypes in both communities only
occurred at certain stations near shore or off shore, indicating that hydrographic and
biogeochemical differences did affect the composition of the bacterial communities to a
certain extent. The only stations which exhibited clearly different banding patterns and
formed a distinct subcluster in 2003 were stations 6 and 7 close to the North Frisian coast.
When samples were collected at these shallow stations rather strong wind (Beaufort scale 6-7)
prevailed leading to high sediment resuspension. The unspecific detection of sediment-

Kapitel V Regional variability of bacterial communities in the German Bight
125
associated δ-Proteobacteria in these samples is a further indication of the sediment
resuspension. Several prominent DGGE bands of the FL bacterial communities amplified
with the Bacteria- and α-Proteobacteria-specific primer sets, however, were present at all
stations, indicating that several populations of this community persisted in the German Bight,
irrespective of the given hydrographic and biogeochemical conditions. The variability within
the PA bacterial community among the various stations was greater than that of the FL
bacterial community, indicating that site-specific properties affected more the former than the
FL bacterial community. However, unspecific amplification of chloroplast-derived 16S rRNA
gene fragments by the Bacteria-specific primer set and of δ-proteobacterial 16S rRNA gene
fragments by the α-Proteobacteria-specific primer set contributed to this variability and
biased the banding patterns and cluster analysis of the PA bacterial community.
Our investigation was carried out during several days in June of two consecutive years,
thus covering only a short period of the annual development of the German Bight. There are
consistent reports of general annual patterns of the hydrography and biological development
at individual stations such as at Helgoland Roads, at Norderney (East Frisian Wadden Sea)
and Büsum and Sylt (North Frisian Wadden Sea) (BSH 2002 and 2003, Loewe et al. 2005).
Even though they exhibit a general seasonal trend, the patterns of each station do vary.
Further, the individual stations show short-term deviations from the seasonal trends, also for
June 2002 and 2003, presumably because of wind- and current-induced movements of water
masses with different physico-chemical and biological properties. Our surveys in both years
encountered two different biological situations, as shown by the different Chl a
concentrations and composition of the phytoplankton, and also reflected by the generally
higher number of DGGE bands detected both in the FL and the PA bacterial community in
2003. The physiological state of the phytoplankton among the near shore and off shore station
varied, as indicated by the variable proportions of phaeopigments relative to total chlorophyll.
Hence it seems not surprising that we did not find consistent patterns of the composition of
the bacterial communities in the near and off shore regions in a rather dynamic regional
coastal sea.
As mentioned, our DGGE and cluster analysis was biased by unspecific PCR
amplifications. Unspecific amplifications by the Bacteria-specific primer set applied has been
reported previously (e.g. Selje & Simon 2003, Stevens et al. 2005a) and amplification of
chloroplast-derived 16S rRNA gene fragments in the PA bacterial community with this
primer set appears a general problem for samples containing phytoplankton cells. Unspecific
amplification with primer ALF968r (α-Proteobacteria) has also been reported by Rink et al.

Kapitel V Regional variability of bacterial communities in the German Bight
126
(2006a) for samples from the East Frisian Wadden Sea and by Overmann et al. (2005). The
latter authors found amplification of Actinobacteria in freshwater samples and possible
detection of few γ-Proteobacteria. Unspecific amplification of δ-Proteobacteria by the
ALF968r primer predominantly occurred in the PA bacterial community at the shallow near
shore stations (3-7) with high concentrations of resuspended SPM. Marine sediments contain
high numbers of sulfate reducing bacteria affiliated to δ-Proteobacteria (Llobet-Brossa et al.
1998, Musat et al. 2006) and also myxobacteria which affiliate to this subclass of
Proteobacteria as well (Stevens et al. 2005a). Hence, when these bacteria are present in the
samples to detectable amounts, their 16S rRNA genes are amplified by this primer. In the FL
bacterial community, no unspecific amplification occurred, indicating that this community did
not include δ-Proteobacteria in proportions high enough to be amplified. In conclusion, a
reliable assessment of the composition of PA α-Proteobacteria by the subclass-specific
primer set was not possible. Our finding of unspecific amplification of the ALF968r primer
has important implications for the interpretation of data obtained by fluorescence in situ
hybridization (FISH) applying this oligonucleotide as a probe (ALF968). In habitats with high
proportions of δ-Proteobacteria they may be included in the detection of α-Proteobacteria
and thus lead to overestimating this subclass.
Also the Bacteroidetes–specific primer set (Jaspers et al. 2001) resulted in unspecific
amplification in three of 12 cases. Even though this is also a bias of the DGGE results, it
appears not as critical as that with the α-Proteobacteria–specific primer set, but emphasizes
the importance for sequencing of prominent bands applying group-specific primer sets in
DGGE analyses.
These biases of unspecific amplification predominantly affected the DGGE banding
pattern of the PA bacterial community obtained by the Bacteria–specific (Fig. 4B) and α-
Proteobacteria–specific primer sets in 2003 (Fig. 4D). The other banding patterns were only
marginally affected. Hence our general findings of greater differences of the composition of
the PA bacterial community among the various stations and a greater diversity as compared to
the FL bacterial community is unaffected by these biases. Our findings are consistent with
previous reports from single stations of the same region (Gerdts et al. 2004, Stevens et al.
2005a). However, similarly as these two reports, we did not enumerate the PA bacteria and
thus can not directly assess their significance relative to that of the FL bacteria. Particle-
associated bacteria have been enumerated in the SPM-rich East Frisian Wadden Sea by Lunau
et al. (2006). In June and July of various years, numbers of PA bacteria vary from 23 to 32%
of total bacteria, and we assume that these numbers are representative for the near shore

Kapitel V Regional variability of bacterial communities in the German Bight
127
stations in general. Comparable numbers are not available for the off shore region of the
German Bight. This region is characterized by low numbers of aggregates (Riebesell 1991),
and PA bacteria in such neritic seas usually constitute <10% of total bacterial biomass and
activity (Simon et al. 2002). Therefore, we assume that PA bacteria are of much lower
significance in the off shore than in the near shore region of the German Bight.
The phylogenetic lineages we identified have previously been shown to constitute the
bacterial communities in the German Bight to a great extent. Applying FISH, Eilers et al.
(2000 and 2001) found that at Helgoland Roads Bacteroidetes and α- and γ-Proteobacteria
constitute 18–30, 15–25 and 6–9% of DAPI-stainable bacteria, respectively, at various
seasonal situations including June. These groups also constitute the bacterial communities in
the Wadden Sea to a great extent, with rather equal proportions in the FL (each group 10–
20% of DAPI stainable bacteria) and PA fractions (each group 15–40%) (Rink et al. 2006b).
Our results, in agreement with previous studies (Eilers et al. 2000, Selje et al. 2004, Stevens et
al. 2005a, Zubkov et al. 2002), show that members of the Roseobacter clade and in particular
of distinct subclusters (RCA, WM11-36, NAC11-7) are important components of the FL
bacterial community (Fig. 5A). As indicated by the prominent DGGE bands persisting at all
stations (GB02-e8-FL, GB02-e9-FL, GB-a14-FL, GB-a15-FL, GB03-e16-FL, GB03-e17-FL)
and the high similarity of the banding patterns, the FL α-Proteobacteria sub-community was
most evenly distributed in the German Bight and least affected by hydrographic and
biogeochemical differences of the various regions. The occurrence of closely related
phylotypes in other neritic and oceanic regions (Gonzalez et al. 2000, Selje et al. 2004)
emphasizes that bacteria of this lineage are important components of the marine
bacterioplankton globally.
The FL sub–community of the Bacteroidetes phylum exhibited a greater variability and
diversity than that of α-Proteobacteria, as shown by the group-specific DGGE banding
patterns and cluster analysis and the occurrence of more phylotypes at distinct stations and of
none at all stations. The phylotypes were retrieved from the bands amplified by both the
Bacteria-specific and Bacteroidetes–specific primer sets. This notion indicates the
significance of site-specific factors for controlling the composition of this sub-community.
The phylotypes affiliated to rather different clusters of the Bacteroidetes phylum but in most
cases were closely related to other phylotypes previously detected in the Wadden Sea, the
German Bight or North Sea during phytoplankton blooms (Table 2, Fig. 5D, Rink et al.
2006a, Stevens et al. 2005a and 2005b, Zubkov et al. 2002, J. Pernthaler et al. unpubl.). Close
relationships between phylotypes and isolates of the Bacteroidetes phylum and distinct

Kapitel V Regional variability of bacterial communities in the German Bight
128
phytoplankton and algal populations have been reported (Schäfer et al. 2002, Grossart et al.
2005, Rooney-Varga et al. 2005). Further, bacteria of this phylum are known to degrade
complex organic polymers (Cottrell & Kirchman 2000, Kirchman 2002). Therefore, it appears
that the phylotypes affiliated to Bacteroidetes reflect more the specific substrate conditions
related to the phytoplankton community of different growth stages and composition and to
other substrate sources at the various stations.
Particles and suspended aggregates provide much more diverse micro-habitats than the
surrounding water. Depending on the given conditions and the water depth, they may include
phytoplankton-derived organic matter as well as organic and inorganic matter resuspended
from the sediment. Therefore, it appears not surprising that the PA bacterial community was
more diverse than the FL bacterial community and contained more site-specific phylotypes
reflecting the local environmental conditions. The PA bacterial community contained quite a
few phylotypes affiliated to various clusters of the Bacteroidetes phylum (Fig. 5D, Table 3),
but also various phylotypes affiliated to α- and γ-Proteobacteria and distinct from those in the
FL bacterial community. The PA bacterial phylotypes affiliated to α-Proteobacteria were
related mainly to various clusters of the Roseobacter clade which differ from those containing
the FL bacterial phylotypes. Others affiliated to Sphingomonas and Acidiphilum aminolyticum
(Fig. 5A, Table 2). The two γ-Proteobacteria phylotypes were detected in 2003 at stations 6
and 7 in the North Frisian Wadden Sea and closely related to phylotypes retrieved from the
East Frisian Wadden Sea (Fig. 5B, Stevens et al. 2005b). One phylotype fell into the
SAMMIC-cluster (Surface Attached Marine MICrobes, Stevens et al. 2005b). Members of
this cluster are uncultured, globally distributed and always associated to suspended aggregates
in coastal systems or to sediments. The unspecific amplification of the tentatively α-
Proteobacteria–specific primer set also retrieved exclusively phylotypes of sediment-
associated δ-Proteobacteria (Fig. 5B, Table 3).
In conclusion, our results show a variable diversity and distribution patterns of the
bacterial community in the German Bight of the North Sea. The FL bacterial community is
rather similar in this coastal sea and harbors several widely distributed members, affiliated to
the Roseobacter clade of α-Proteobacteria and obviously little affected by site-specific
environmental conditions. However, other members occurred only at specific locations,
obviously as the result of site-specific environmental conditions of the tidal flat areas and the
phytoplankton communities. The PA bacterial community was more divers and reflected
mainly local environmental features, also related to the specific environmental conditions of

Kapitel V Regional variability of bacterial communities in the German Bight
129
the tidal flat areas with high SPM concentrations and intense resuspension and of the
phytoplankton communities.
ACKNOWLEDGEMENTS
We appreciate the hospitality and cooperation of the captain and crew of RV Heincke. We
thank B. Kuerzel for dry weight and chlorophyll analyses and A. Luek for phytoplankton and
bacterial cell counts. This work was supported by the Deutsche Forschungsgemeinschaft
(DFG) within the research group “BioGeoChemistry of the Wadden Sea” (FG 432 TP5).

Kapitel V Regional variability of bacterial communities in the German Bight
130
REFERENCES
Brinkhoff T, Bach G, Heidorn T, Liang L, Schlingloff A, Simon M (2004) Antibiotic production by a Roseobacter clade affiliated species from the German Wadden Sea and its antagonistic effects on indigenous isolates. Appl Environ Microbiol 70:2560-2565
BSH (2002) MURSYS Reportsystem 2002. Bundesamt für Seeschifffahrt und Hydrographie, Hamburg (www.bsh.de)
BSH (2003) MURSYS Reportsystem 2003. Bundesamt für Seeschifffahrt und Hydrographie, Hamburg (www.bsh.de)
Cotner JB, Biddanda BA (2002) Small players, large role: Microbial influence on biogeochemical processes in pelagic aquatic ecosystems. Ecosystems 5:105-121
Cottrell MT, Kirchman DL (2000) Natural assemblages of marine Proteobacteria and members of the Cytophaga-Flavobacteria cluster consuming low- and high-molecular weight dissolved organic matter. Appl Environ Microbiol 66:1692-1697
Covert JS, Moran MA (2001) Molecular characterization of estuarine bacterial communities that use high- and low-molecular weight fractions of dissolved organic carbon. Aquat Microb Ecol 25:127-139
Crump BC, Kling GW, Bahr M, Hobbie JE (2003) Bacterioplankton community shifts in an arctic lake correlate with seasonal changes in organic matter. Appl Environ Microbiol 69:2253-2268
Drebes G (1974) Marines Phytoplankton. Georg-Thieme-Verlag, Stuttgart
Eilers H, Pernthaler J, Glöckner FO, Amann R (2000) Culturability and in situ abundance of pelagic bacteria from the North Sea. Appl Environ Microbiol 66:3044-3051
Eilers H, Pernthaler J, Peplies J, Glöckner FO, Gerdts G, Amann R (2001) Isolation of novel pelagic bacteria from the German Bight and their seasonal contributions to surface picoplankton. Appl Environ Microbiol 67:5134-5142
Gerdts G, Wichels A, Döpke H, Klings K-W, Gunkel W, Schütt C (2004) A long-term study of microbial parameters near Helgoland (German Bight, North Sea): Historical view and future perspectives. Helgoland Mar Res 58: 230-242
Gonzalez JM, Simo R, Massana R, Covert J, Casamayor E, Pedros-Alio C, Moran MA (2000) Bacterial community structure associated with a dimethylsulfonioproponiate-producing North Atlantic algal bloom. Appl Environ Microbiol 66:4237-4246
Grossart HP, Levold F, Allgaier M, Simon M, Brinkhoff T (2005) Composition of bacterial communities associated with marine diatoms. Environ Microbiol 7:960-973
Jaspers E, Nauhaus K, Cypionka H, Overmann J (2001) Multitude and temporal variability of ecological niches as indicated by the diversity of cultivated bacterioplankton. FEMS Microbiol Ecol 36:153-164
Kirchman DL (2002) The ecology of Cytophaga-Flavobacteria in aquatic environments. FEMS Microb Ecol 39:91-100
Lebaron P, Servais P, Trousselier M, Courties C, Vives-Rego J, Muyzer G, Bernard L, Guindulain T, Schäfer H, Stackebrandt E (1999) Changes in bacterial community structure in seawater mesocosms differing in their nutrient status. Aquat Microb Ecol 19:255-2267

Kapitel V Regional variability of bacterial communities in the German Bight
131
Loewe P, Schmolke S, Becker G, Brockmann U Dick S, Engelke C, Frohse A, Horn W, Klein H, Müller-Navarra S, Nies H, Schmelzer N, Schrader D, Schulz A, Theobald N, Weigelt S (2005) Berichte des Bundesamtes für Seeschiffahrt und Hydrographie. Nr. 38/2005, Hamburg (www.bsh.de)
Ludwig W, Strunk O, Westram R, Richter L, Meier H, Yadhukumar, Buchner A, Lai T, Steppi S, Jobb G, Forster W, Brettske I, Gerber S, Ginhart AW, Gross O, Grumann S, Hermann S, Jost R, Konig A, Liss T, Lussmann R, May M, Nonhoff B, Reichel B, Strehlow R, Stamatakis A, Stuckmann N, Vilbig A, Lenke M, Ludwig T, Bode A, Schleifer KH (2004) ARB: a software environment for sequence data. Nucleic Acid Res 32:1363-1371
Lunau M, Lemke A, Walther K, Martens-Habbena W, Simon M (2005) An improved method for counting bacteria in samples with high proportions of particle-associated cells by epifluorescence microscopy. Environ Microbiol 7:961-968
Lunau M, Lemke A, Dellwig O, Simon M (2006) Physical and biogeochemical controls of microaggregate dynamics in a tidally affected coastal ecosystem. Limnol Oceanogr 51:847-859
Musat N, Werner U, Knittel K, Dodenhof T, van Beusekom JE, De Beer D, Dubilier N, Amann R (2006) Microbial community structure of sandy intertidal sediments in the North Sea, Sylt-Romø Basin, Wadden Sea. Syst Appl Microbiol 4:333-348
Muyzer G, de Waal EC, Uitterlinden AG (1993) Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl Environ Microbiol 59:695-700
Muyzer G, Brinkhoff T, Nübel U, Santegods C, Schäfer H, Wawer C (1998) Denaturing gradient gel electrophoresis (DGGE) in microbial ecology. Molecular Microbial Ecology Manual, p 1-27. Kluwer Academic Publishers, Dordrecht
Parsons TR, Maita Y, Lalli CM (1984) A manual of chemical and biological methods for seawater analysis, p101-112. Pergamon Press, New York
Porter K, Feig Y (1980) The use of DAPI for identifying and counting aquatic microflora. Limnol Oceanogr 25:943-948
Reinthaler T, Winter C, Herndl GJ (2005). Relationship between bacterioplankton richness, respiration, and production in the southern North Sea. Appl Environ Microbiol 71:2260-2266
Riebesell U (1991) Particle aggregation during a diatom bloom. II. Biological aspects. Mar Ecol Prog Ser 69:281-291
Rink B, Seeberger S, Martens T, Duerselen CD, Simon M, Brinkhoff T (2006a) A phytoplankton bloom in a coastal ecosystem affects the composition of bacterial communities. Aquat Microb Ecol, submitted
Rink B, Martens T, Fischer D, Lemke A, Grossart HP, Simon M, Brinkhoff T (2006b) Tidal effects on coastal bacterioplankton. Manuscript to be submitted to Aquat Microb Ecol
Rooney-Varga JN, Giewat MW, Savin MC, Sood S, LeGresley M, Martin JL (2005) Links between phytoplankton and bacterial community dynamics in a coastal marine environment. Microb Ecol 49:163-175
Sambrook J, Frisch EF, Maniatis T (1989) Northern hybridisation. In : Molecular Cloning: a laboratory manual, 2nd edn., Cold Spring Harbour Laboratory Press, New York

Kapitel V Regional variability of bacterial communities in the German Bight
132
Schäfer H, Abbas B, Witte H, Muyzer G (2002) Genetic diversity of 'satellite' bacteria present in cultures of marine diatoms. FEMS Microbiol Ecol 42:25-35
Selje N, Simon M (2003) Composition and dynamics of particle-associated and free-living bacterial communities in the Weser estuary, Germany. Aquat Microb Ecol 30:221-236
Selje N, Simon M, Brinkhoff T (2004) A newly discovered Roseobacter cluster in temperate and polar oceans. Nature 427:445-448
Simon M, Grossart HP, Schweitzer B, Plough H (2002) Microbial ecology of organic aggregates in aquatic ecosystems. Aquat Microb Ecol 28:175-211
Stevens H, Brinkhoff T, Simon M (2005a) Composition and seasonal dynamics of free-living, aggregate- and sediment surface-associated bacterial communities in the German Wadden Sea. Aquat Microb Ecol 38:15-30
Stevens H, Stübner M, Simon M, Brinkhoff T (2005b). Phylogeny of Proteobacteria and Bacteroidetes from oxic habitats of a tidal flat ecosystem. FEMS Microbiol Ecol 54:351-365
Utermöhl H (1958) Zur Vervollkommnung der quantitativen Phytoplanktonmethodik. Mitt Int Verh Theor Angew Limnol 9:1-38
Zubkov MV, Fuchs BM, Archer SD, Kiene RP, Amann R, Burkill PH (2002) Rapid turnover of dissolved DMS and DMSP by defined bacterioplankton communities in the stratified euphotic zone of the North Sea. Deep-Sea Res II 49:3017-3038
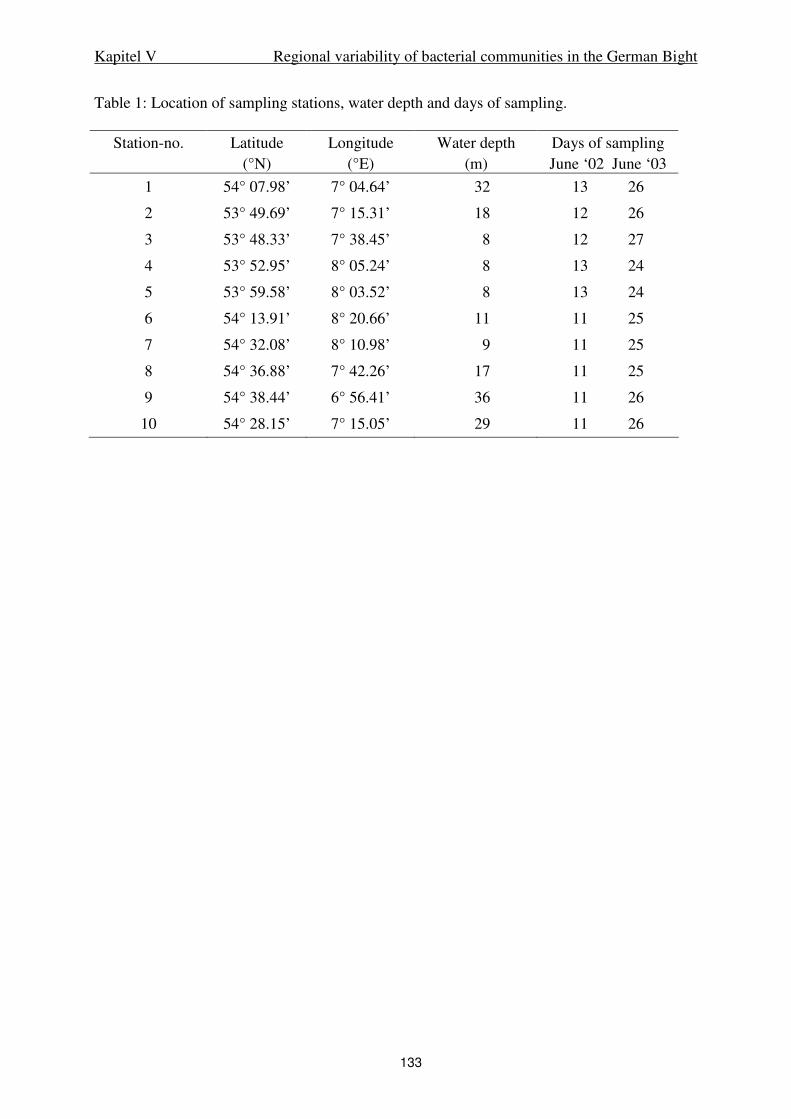
Kapitel V Regional variability of bacterial communities in the German Bight
133
Table 1: Location of sampling stations, water depth and days of sampling.
Station-no.
Latitude (°N)
Longitude (°E)
Water depth (m)
Days of sampling June ‘02 June ‘03
1 54° 07.98’ 7° 04.64’ 32 13 26
2 53° 49.69’ 7° 15.31’ 18 12 26
3 53° 48.33’ 7° 38.45’ 8 12 27
4 53° 52.95’ 8° 05.24’ 8 13 24
5 53° 59.58’ 8° 03.52’ 8 13 24
6 54° 13.91’ 8° 20.66’ 11 11 25
7 54° 32.08’ 8° 10.98’ 9 11 25
8 54° 36.88’ 7° 42.26’ 17 11 25
9 54° 38.44’ 6° 56.41’ 36 11 26
10 54° 28.15’ 7° 15.05’ 29 11 26
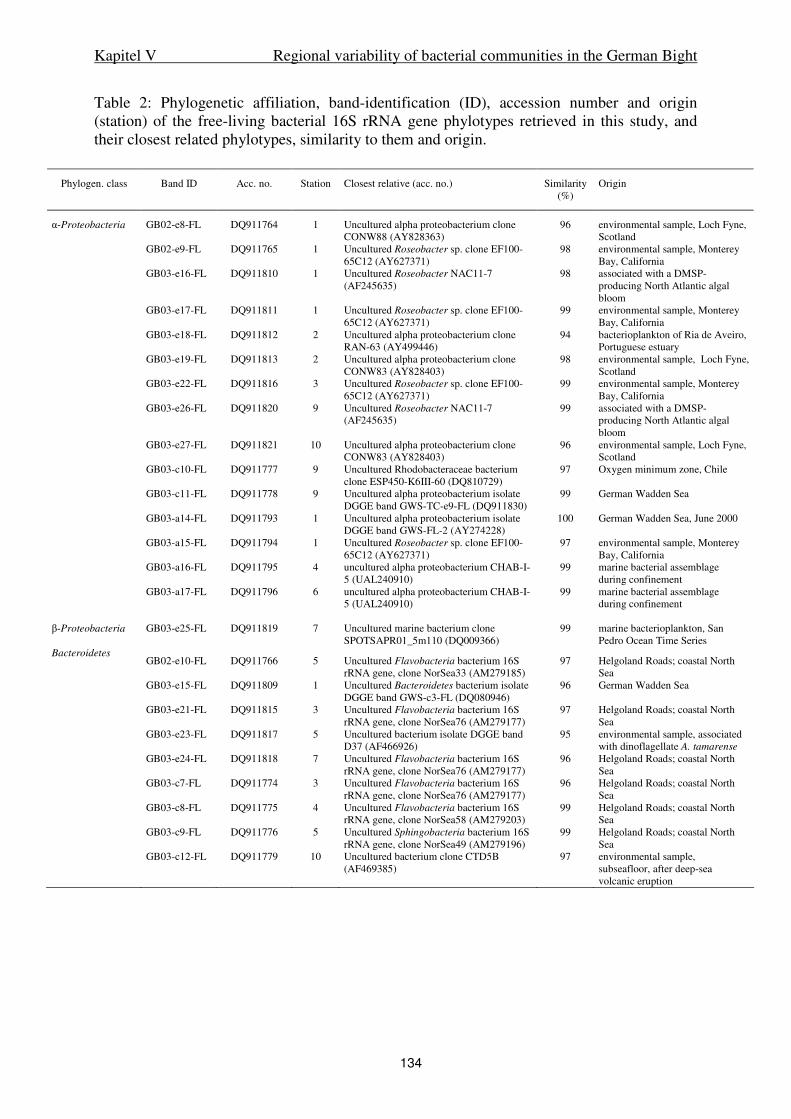
Kapitel V Regional variability of bacterial communities in the German Bight
134
Table 2: Phylogenetic affiliation, band-identification (ID), accession number and origin (station) of the free-living bacterial 16S rRNA gene phylotypes retrieved in this study, and their closest related phylotypes, similarity to them and origin.
Phylogen. class Band ID Acc. no. Station Closest relative (acc. no.) Similarity (%)
Origin
α-Proteobacteria GB02-e8-FL DQ911764 1 Uncultured alpha proteobacterium clone CONW88 (AY828363)
96 environmental sample, Loch Fyne, Scotland
GB02-e9-FL DQ911765 1 Uncultured Roseobacter sp. clone EF100-65C12 (AY627371)
98 environmental sample, Monterey Bay, California
GB03-e16-FL DQ911810 1 Uncultured Roseobacter NAC11-7 (AF245635)
98 associated with a DMSP-producing North Atlantic algal bloom
GB03-e17-FL DQ911811 1 Uncultured Roseobacter sp. clone EF100-65C12 (AY627371)
99 environmental sample, Monterey Bay, California
GB03-e18-FL DQ911812 2 Uncultured alpha proteobacterium clone RAN-63 (AY499446)
94 bacterioplankton of Ria de Aveiro, Portuguese estuary
GB03-e19-FL DQ911813 2 Uncultured alpha proteobacterium clone CONW83 (AY828403)
98 environmental sample, Loch Fyne, Scotland
GB03-e22-FL DQ911816 3 Uncultured Roseobacter sp. clone EF100-65C12 (AY627371)
99 environmental sample, Monterey Bay, California
GB03-e26-FL DQ911820 9 Uncultured Roseobacter NAC11-7 (AF245635)
99 associated with a DMSP-producing North Atlantic algal bloom
GB03-e27-FL DQ911821 10 Uncultured alpha proteobacterium clone CONW83 (AY828403)
96 environmental sample, Loch Fyne, Scotland
GB03-c10-FL DQ911777 9 Uncultured Rhodobacteraceae bacterium clone ESP450-K6III-60 (DQ810729)
97 Oxygen minimum zone, Chile
GB03-c11-FL DQ911778 9 Uncultured alpha proteobacterium isolate DGGE band GWS-TC-e9-FL (DQ911830)
99 German Wadden Sea
GB03-a14-FL DQ911793 1 Uncultured alpha proteobacterium isolate DGGE band GWS-FL-2 (AY274228)
100 German Wadden Sea, June 2000
GB03-a15-FL DQ911794 1 Uncultured Roseobacter sp. clone EF100-65C12 (AY627371)
97 environmental sample, Monterey Bay, California
GB03-a16-FL DQ911795 4 uncultured alpha proteobacterium CHAB-I-5 (UAL240910)
99 marine bacterial assemblage during confinement
GB03-a17-FL DQ911796 6 uncultured alpha proteobacterium CHAB-I-5 (UAL240910)
99 marine bacterial assemblage during confinement
β-Proteobacteria GB03-e25-FL DQ911819 7 Uncultured marine bacterium clone
SPOTSAPR01_5m110 (DQ009366) 99 marine bacterioplankton, San
Pedro Ocean Time Series Bacteroidetes
GB02-e10-FL DQ911766 5 Uncultured Flavobacteria bacterium 16S rRNA gene, clone NorSea33 (AM279185)
97 Helgoland Roads; coastal North Sea
GB03-e15-FL DQ911809 1 Uncultured Bacteroidetes bacterium isolate DGGE band GWS-c3-FL (DQ080946)
96 German Wadden Sea
GB03-e21-FL DQ911815 3 Uncultured Flavobacteria bacterium 16S rRNA gene, clone NorSea76 (AM279177)
97 Helgoland Roads; coastal North Sea
GB03-e23-FL DQ911817 5 Uncultured bacterium isolate DGGE band D37 (AF466926)
95 environmental sample, associated with dinoflagellate A. tamarense
GB03-e24-FL DQ911818 7 Uncultured Flavobacteria bacterium 16S rRNA gene, clone NorSea76 (AM279177)
96 Helgoland Roads; coastal North Sea
GB03-c7-FL DQ911774 3 Uncultured Flavobacteria bacterium 16S rRNA gene, clone NorSea76 (AM279177)
96 Helgoland Roads; coastal North Sea
GB03-c8-FL DQ911775 4 Uncultured Flavobacteria bacterium 16S rRNA gene, clone NorSea58 (AM279203)
99 Helgoland Roads; coastal North Sea
GB03-c9-FL DQ911776 5 Uncultured Sphingobacteria bacterium 16S rRNA gene, clone NorSea49 (AM279196)
99 Helgoland Roads; coastal North Sea
GB03-c12-FL DQ911779 10 Uncultured bacterium clone CTD5B (AF469385)
97 environmental sample, subseafloor, after deep-sea volcanic eruption
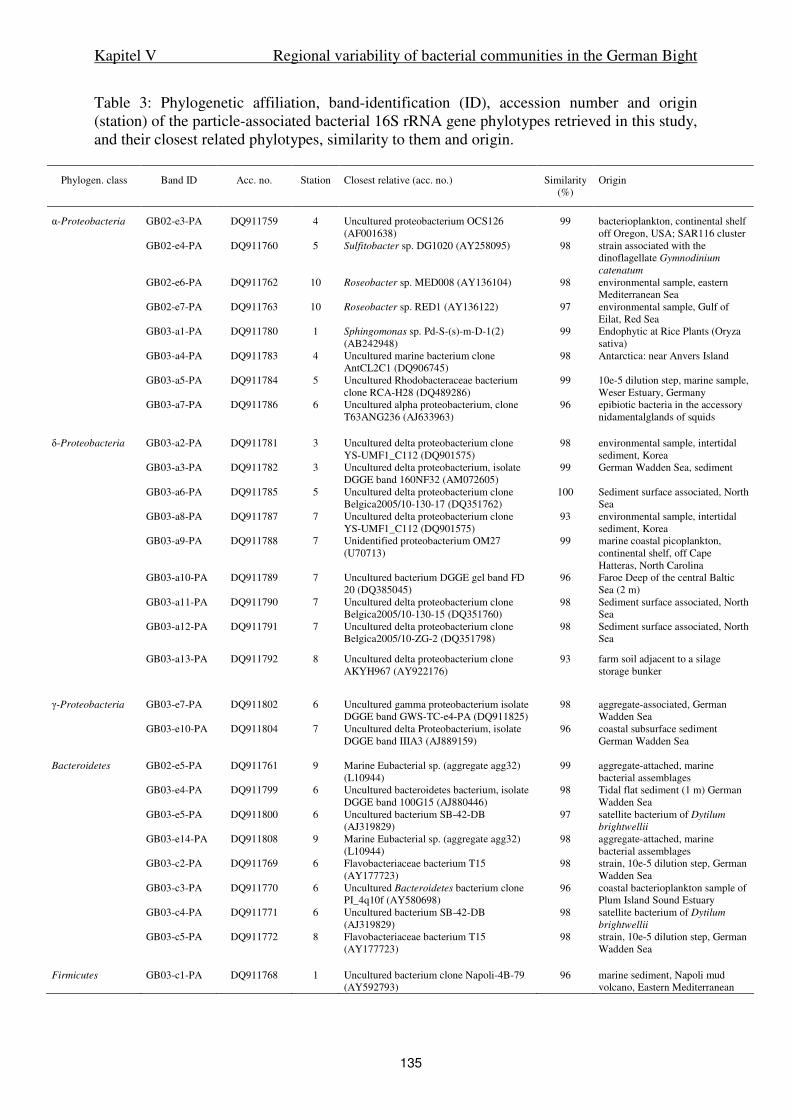
Kapitel V Regional variability of bacterial communities in the German Bight
135
Table 3: Phylogenetic affiliation, band-identification (ID), accession number and origin (station) of the particle-associated bacterial 16S rRNA gene phylotypes retrieved in this study, and their closest related phylotypes, similarity to them and origin.
Phylogen. class Band ID Acc. no. Station Closest relative (acc. no.) Similarity (%)
Origin
α-Proteobacteria GB02-e3-PA DQ911759 4 Uncultured proteobacterium OCS126 (AF001638)
99 bacterioplankton, continental shelf off Oregon, USA; SAR116 cluster
GB02-e4-PA DQ911760 5 Sulfitobacter sp. DG1020 (AY258095) 98 strain associated with the dinoflagellate Gymnodinium
catenatum GB02-e6-PA DQ911762 10 Roseobacter sp. MED008 (AY136104) 98 environmental sample, eastern
Mediterranean Sea GB02-e7-PA DQ911763 10 Roseobacter sp. RED1 (AY136122) 97 environmental sample, Gulf of
Eilat, Red Sea GB03-a1-PA DQ911780 1 Sphingomonas sp. Pd-S-(s)-m-D-1(2)
(AB242948) 99 Endophytic at Rice Plants (Oryza
sativa) GB03-a4-PA DQ911783 4 Uncultured marine bacterium clone
AntCL2C1 (DQ906745) 98 Antarctica: near Anvers Island
GB03-a5-PA DQ911784 5 Uncultured Rhodobacteraceae bacterium clone RCA-H28 (DQ489286)
99 10e-5 dilution step, marine sample, Weser Estuary, Germany
GB03-a7-PA DQ911786 6 Uncultured alpha proteobacterium, clone T63ANG236 (AJ633963)
96 epibiotic bacteria in the accessory nidamentalglands of squids
δ-Proteobacteria GB03-a2-PA DQ911781 3 Uncultured delta proteobacterium clone
YS-UMF1_C112 (DQ901575) 98 environmental sample, intertidal
sediment, Korea GB03-a3-PA DQ911782 3 Uncultured delta proteobacterium, isolate
DGGE band 160NF32 (AM072605) 99 German Wadden Sea, sediment
GB03-a6-PA DQ911785 5 Uncultured delta proteobacterium clone Belgica2005/10-130-17 (DQ351762)
100 Sediment surface associated, North Sea
GB03-a8-PA DQ911787 7 Uncultured delta proteobacterium clone YS-UMF1_C112 (DQ901575)
93 environmental sample, intertidal sediment, Korea
GB03-a9-PA DQ911788 7 Unidentified proteobacterium OM27 (U70713)
99 marine coastal picoplankton, continental shelf, off Cape Hatteras, North Carolina
GB03-a10-PA DQ911789 7 Uncultured bacterium DGGE gel band FD 20 (DQ385045)
96 Faroe Deep of the central Baltic Sea (2 m)
GB03-a11-PA DQ911790 7 Uncultured delta proteobacterium clone Belgica2005/10-130-15 (DQ351760)
98 Sediment surface associated, North Sea
GB03-a12-PA DQ911791 7 Uncultured delta proteobacterium clone Belgica2005/10-ZG-2 (DQ351798)
98 Sediment surface associated, North Sea
GB03-a13-PA DQ911792 8 Uncultured delta proteobacterium clone AKYH967 (AY922176)
93 farm soil adjacent to a silage storage bunker
γ-Proteobacteria GB03-e7-PA DQ911802 6 Uncultured gamma proteobacterium isolate DGGE band GWS-TC-e4-PA (DQ911825)
98 aggregate-associated, German Wadden Sea
GB03-e10-PA DQ911804 7 Uncultured delta Proteobacterium, isolate DGGE band IIIA3 (AJ889159)
96 coastal subsurface sediment German Wadden Sea
Bacteroidetes GB02-e5-PA DQ911761 9 Marine Eubacterial sp. (aggregate agg32)
(L10944) 99 aggregate-attached, marine
bacterial assemblages GB03-e4-PA DQ911799 6 Uncultured bacteroidetes bacterium, isolate
DGGE band 100G15 (AJ880446) 98 Tidal flat sediment (1 m) German
Wadden Sea GB03-e5-PA DQ911800 6 Uncultured bacterium SB-42-DB
(AJ319829) 97 satellite bacterium of Dytilum
brightwellii GB03-e14-PA DQ911808 9 Marine Eubacterial sp. (aggregate agg32)
(L10944) 98 aggregate-attached, marine
bacterial assemblages GB03-c2-PA DQ911769 6 Flavobacteriaceae bacterium T15
(AY177723) 98 strain, 10e-5 dilution step, German
Wadden Sea GB03-c3-PA DQ911770 6 Uncultured Bacteroidetes bacterium clone
PI_4q10f (AY580698) 96 coastal bacterioplankton sample of
Plum Island Sound Estuary GB03-c4-PA DQ911771 6 Uncultured bacterium SB-42-DB
(AJ319829) 98 satellite bacterium of Dytilum
brightwellii GB03-c5-PA DQ911772 8 Flavobacteriaceae bacterium T15
(AY177723) 98 strain, 10e-5 dilution step, German
Wadden Sea Firmicutes GB03-c1-PA DQ911768 1 Uncultured bacterium clone Napoli-4B-79
(AY592793) 96 marine sediment, Napoli mud
volcano, Eastern Mediterranean

Kapitel V Regional variability of bacterial communities in the German Bight
136
Figure Legends
Fig. 1.
Study area and locations of the sampling stations in the German Bight, Southern North Sea.
Fig. 2.
Scatter plot of temperature versus salinity of the sampling stations in the German Bight in
June 2002 and 2003. For exact locations and dates see Table 1.
Fig. 3.
Suspended matter dry weight (SPM) in 2002 (A) and 2003 (B), POC in 2003 (B), Chlorophyll
a in 2002 (C) and 2003 (D), phaeopigments in 2003 (D), phytoplankton cell numbers in 2003
(E) bacterial numbers in 2002 (F) and 2003 (G) at various stations in the German Bight.
Samples were collected in June of both years. Stations 8-10 and 1 are off shore, stations 2-7
near shore. For exact locations and dates see Table 1. Missing bars: data not available.
Fig. 4.
Cluster analyses of the DGGE banding patterns of particle-attached and free-living Bacteria
(EUB02, A; EUB03, B), Bacteroidetes (CFB03, C) and α-Proteobacteria (ALF03, D)
retrieved from samples at various stations in the German Bight in June 2002 (02) and 2003
(03). PA: Particle associated bacterial community.
Fig. 5.
Phylogenetic trees of α-Proteobacteria (A), δ-Proteobacteria (B), β- and γ-Proteobacteria
(C) and the Bacteroidetes phylum (D) calculated with Maximum-Likelihood based on 16S
rRNA gene fragments. Sequences obtained in this study are highlighted in bold.
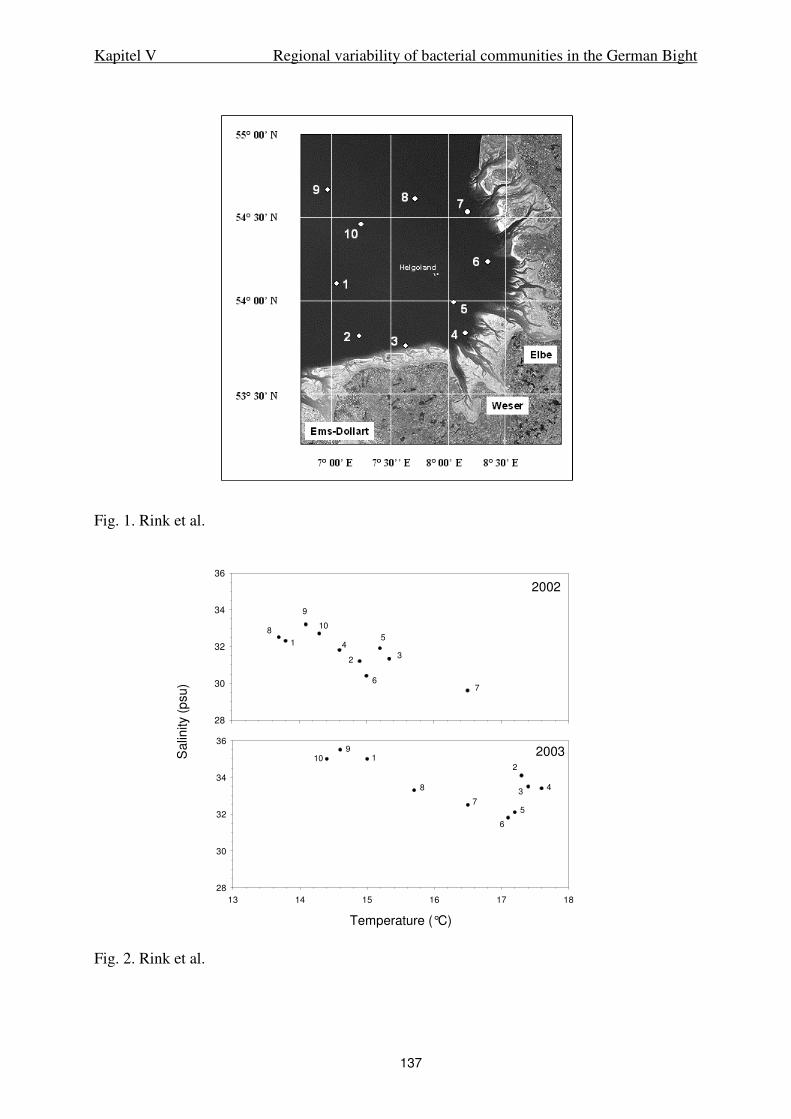
Kapitel V Regional variability of bacterial communities in the German Bight
137
Fig. 1. Rink et al.
Fig. 2. Rink et al.
Salin
ity
(psu)
28
30
32
34
36
Temperature (°C)
13 14 15 16 17 18
28
30
32
34
36
2002
2003
7
54
2
6
9
10
1
8
3
8
7
19
102
34
5
6
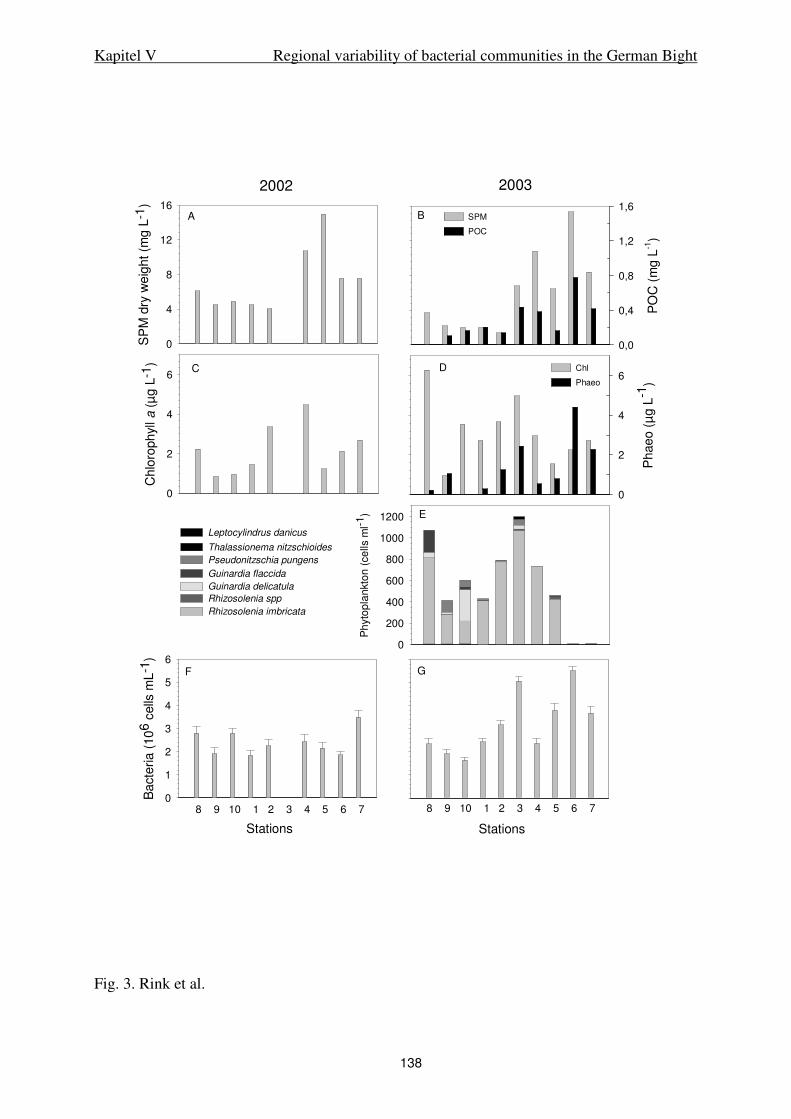
Kapitel V Regional variability of bacterial communities in the German Bight
138
Fig. 3. Rink et al.
SP
M d
ry w
eig
ht
(mg
L-1
)
0
4
8
12
16
PO
C (
mg L
-1)
0,0
0,4
0,8
1,2
1,6
Ch
loro
ph
yll
a (
µg
L-1
)
0
2
4
6
SPM
POC
Ph
ae
o (
µg L
-1)
0
2
4
6Chl
Phaeo
2002 2003
Ph
yto
pla
nkto
n (
ce
lls m
l-1)
0
200
400
600
800
1000
1200
8 9 10 1 2 3 4 5 6 78 9 10 1 2 3 4 5 6 7
Stations
A B
C D
E
F G
Stations
Ba
cte
ria
(10
6 c
ells
mL
-1)
0
1
2
3
4
5
6
Rhizosolenia imbricata
Rhizosolenia spp
Guinardia delicatula
Guinardia flaccida
Pseudonitzschia pungens
Thalassionema nitzschioides
Leptocylindrus danicus
F
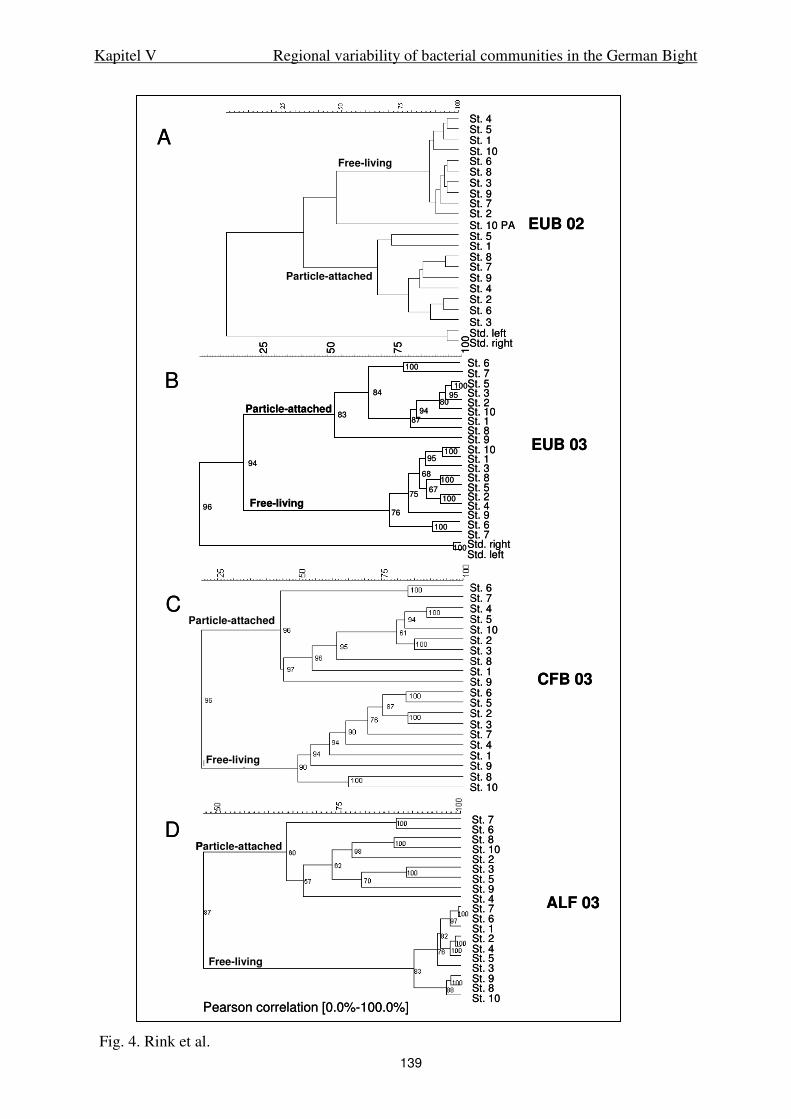
Kapitel V Regional variability of bacterial communities in the German Bight
139
Fig. 4. Rink et al.
St. 6 St. 7
St. 3 St. 8
St. 6
100
75
50
25
100
10095
8094
87
84
83
10095
100
10067
68
75
100
76
94
100
96
St. 7 St. 5 St. 3 St. 2 St. 10 St. 1 St. 8 St. 9 St. 10 St. 1
St. 5 St. 2 St. 4 St. 9
Std. rightStd. left
Particle-attached
Free-living
St. 6 St. 7 St. 4 St. 5 St. 10 St. 2 St. 3 St. 8 St. 1 St. 9 St. 6 St. 5 St. 2 St. 3St. 7 St. 4St. 1St. 9St. 8St. 10
Pearson correlation [0.0%-100.0%]
St. 7St. 6 St. 8St. 10St. 2St. 3St. 5St. 9St. 4St. 7St. 6St. 1St. 2St. 4St. 5St. 3St. 9 St. 8 St. 10
Particle-attached
Free-living
D
Free-living
Particle-attached
St. 4St. 5St. 1St. 10St. 6St. 8St. 3St. 9St. 7St. 2St. 10 PASt. 5St. 1St. 8St. 7St. 9St. 4St. 2St. 6St. 3Std. leftStd. right
EUB 02
A
EUB 03
CFB 03
ALF 03
B
CParticle-attached
Free-living
St. 6 St. 7
St. 3 St. 8
St. 6
100
75
50
25
100
10095
8094
87
84
83
10095
100
10067
68
75
100
76
94
100
96
St. 7 St. 5 St. 3 St. 2 St. 10 St. 1 St. 8 St. 9 St. 10 St. 1
St. 5 St. 2 St. 4 St. 9
Std. rightStd. left
Particle-attached
Free-living
St. 6 St. 7 St. 4 St. 5 St. 10 St. 2 St. 3 St. 8 St. 1 St. 9 St. 6 St. 5 St. 2 St. 3St. 7 St. 4St. 1St. 9St. 8St. 10
Pearson correlation [0.0%-100.0%]
St. 7St. 6 St. 8St. 10St. 2St. 3St. 5St. 9St. 4St. 7St. 6St. 1St. 2St. 4St. 5St. 3St. 9 St. 8 St. 10
Particle-attached
Free-living
D
Free-living
Particle-attached
St. 4St. 5St. 1St. 10St. 6St. 8St. 3St. 9St. 7St. 2St. 10 PASt. 5St. 1St. 8St. 7St. 9St. 4St. 2St. 6St. 3Std. leftStd. right
EUB 02
A
EUB 03
CFB 03
ALF 03
B
CParticle-attached
Free-living
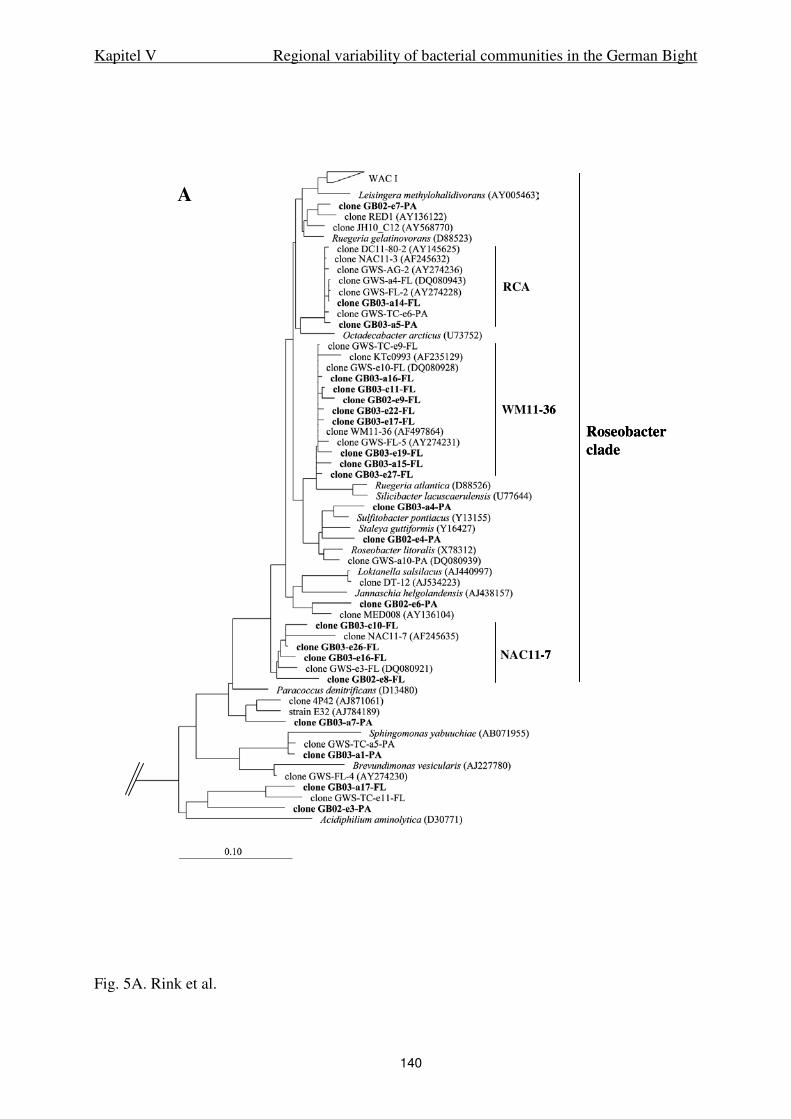
Kapitel V Regional variability of bacterial communities in the German Bight
140
Fig. 5A. Rink et al.
A
Roseobacterclade
RCA
WM11-36
NAC11-7
A
Roseobacterclade
RCA
WM11-36
NAC11-7
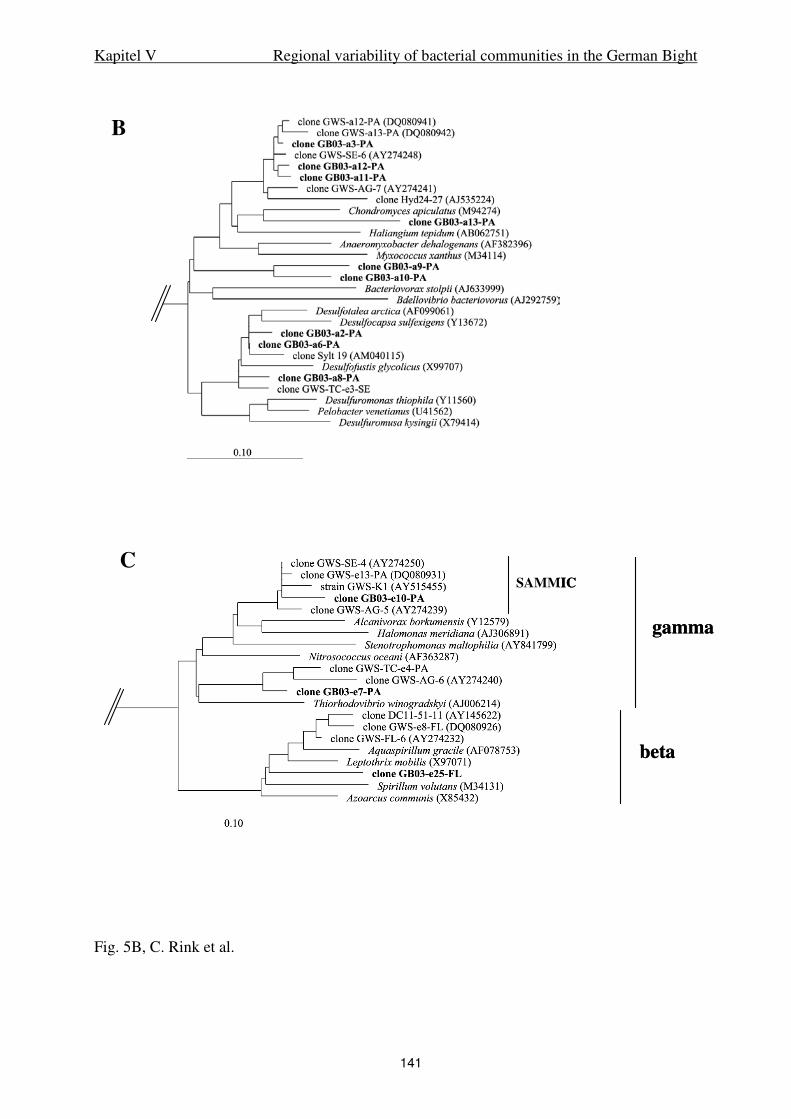
Kapitel V Regional variability of bacterial communities in the German Bight
141
Fig. 5B, C. Rink et al.
B
C
gamma
SAMMIC
beta
B
C
gamma
SAMMIC
beta

Kapitel V Regional variability of bacterial communities in the German Bight
142
D
Fig. 5D. Rink et al.
D
Fig. 5D. Rink et al.
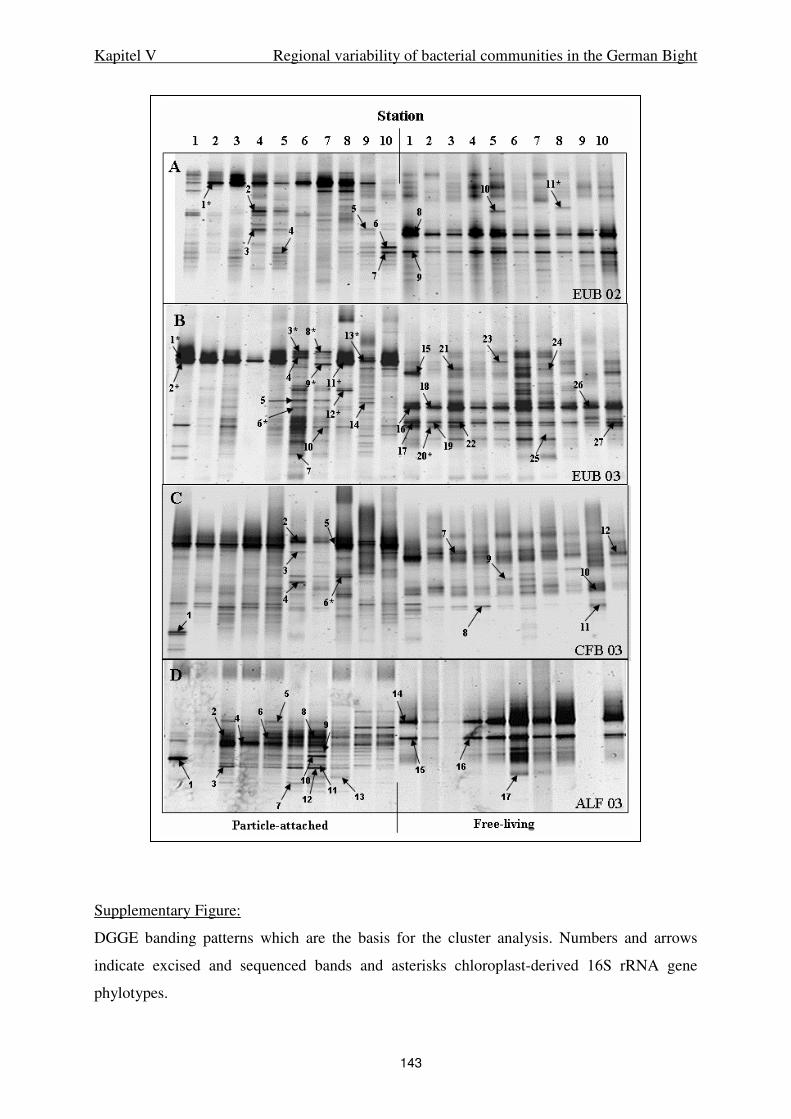
Kapitel V Regional variability of bacterial communities in the German Bight
143
Supplementary Figure:
DGGE banding patterns which are the basis for the cluster analysis. Numbers and arrows
indicate excised and sequenced bands and asterisks chloroplast-derived 16S rRNA gene
phylotypes.

144
VI.
Schlussbetrachtung und Ausblick

Kapitel VI Schlussbetrachtung und Ausblick
145
Die vorliegende Arbeit umfasst detaillierte Untersuchungen zur Struktur und Abundanz des
Bakterioplankton im ostfriesischen Wattenmeer sowie der Deutschen Bucht. Besondere
Berücksichtigung fand die Entwicklung und Anwendung spezifischer Nachweise für
Bakteriengruppen, die durch methodische Einschränkungen in der Vergangenheit als bisher
unterrepräsentiert angesehen werden mußten. Die Ergebnisse dieser Arbeit wurden im
Rahmen der DFG geförderten Forschergruppe „BioGeoChemie des Watts“ gewonnen und
stellen eine wesentliche Grundlage für zukünftige Untersuchungen bezüglich des
Stoffumsatzes abundanter Bakteriengruppen dar.
Das Bakterioplankton im Wattenmeer kann in die Kompartimente freilebend, Aggregat- und
Sediment- assoziiert unterteilt werden (vgl. Stevens et al. , 2005,). Die DGGE-Auswertung
von Probenahmen in monatlichen Abständen ergab eine relative Stabilität der
Bakteriengemeinschaften im Wattenmeer (Stevens et al., 2005). Diese veränderten sich nur
geringfügig zu extremen Ereignissen, z. B. während und nach Phytoplanktonblüten und bei
Sturm. Da diese Ereignisse fester Bestandteil der Jahreszeiten sind und regelmäßig
wiederkehren, wurden sie als stabile Merkmale des Ökosystems Wattenmeer gewertet und die
Schlussfolgerung war, dass das Bakterioplankton praktisch keinen Änderungen unterliegt. Die
Aussagen von H. Stevens sind, basierend auf den damals erhobenen Daten, durchaus
zutreffend, bedürfen allerdings durch die Ergebnisse der vorliegenden Arbeit nun der
Ergänzung.
In Kapitel II wurde deutlich, dass die große Stabilität der Bakteriengemeinschaft bei
genauerer Betrachtung über den Zeitraum einer Phytoplanktonblüte moduliert wird. Man kann
annehmen, dass durch Ausscheidungen der Diatomeen und auch deren Absterben bestimmte
Bakteriengruppen in einem kurzen Zeitintervall stark beeinflusst werden. Dies betrifft vor
allem Bakterien des Phylums Bacteroidetes und der Roseobacter-Gruppe innerhalb der alpha-
Proteobakterien, die mit Änderungen der Phytoplanktonzusammensetzung korreliert waren.
Es zeigte sich auch, dass die Bakteriengemeinschaften der unterschiedlichen Kompartimente
(freilebend und Aggregat-assoziiert) auf unterschiedliche Weise beeinflusst werden. Eine
überraschend hohe Artenvielfalt von Bakterien der Roseobacter-Gruppe war auf Aggregaten
zu beobachten, die annehmen lässt, dass diese Gruppe für die Zersetzung partikulären
organischen Materials (POM) bisher unterschätzt wurde. Ebenso deutlich wurde, dass frei
lebende Bacteroidetes sehr schnell von Änderungen in der Umgebung beeinflusst werden und
ihre Rolle im Abbau von gelöstem organischem Material (DOM) wahrscheinlich als
wesentlich größer eingeschätzt werden kann als bisher angenommen. Die Bedeutung der

Kapitel VI Schlussbetrachtung und Ausblick
146
vorliegenden Arbeit liegt daher in der Beschreibung des Potentials einzelner
Bakteriengruppen, die im Ökosystem Wattenmeer extremen Einflüssen unterliegen und
sowohl Stabilität als auch Flexibilität besitzen. Die Auswirkung der einzelnen
Bakteriengruppen auf den gesamten Stoffumsatz ist daher komplex zu betrachten und sollte in
weiteren Untersuchungen besonderen Stellenwert einnehmen.
Hinweise über Veränderungen bakterieller Aktivität wurden sogar während eines Tidenzyklus
beschrieben (Grossart et al. 2004). Diese Beobachtung ließ die Frage entstehen, auf welcher
Zeitskala Reaktionen der Bakteriengemeinschaft nachweisbar sind. In Kapitel II war
ersichtlich, dass Bakteriengemeinschaften in wöchentlichen Zeitabständen starken
Änderungen unterliegen können. Die Abhängigkeit der Änderungen in der
Bakteriengemeinschaft von saisonalen Variablen, z. B. Sturmereignisse im Herbst und die
Phytoplanktonblüte im Frühjahr und Sommer, war bereits beschrieben worden (Stevens et al.
2005).
Daher wurde in Kapitel III anhand von Probenahmen zu den Kenterpunkten bei Hoch- und
Niedrigwasser sowie zum Strömungsmaximum und hoch sensitiver Nachweismethoden der
Einfluss der Tide auf die Zusammensetzung der Bakteriengemeinschaften zu
unterschiedlichen Jahreszeiten untersucht. Die Ergebnisse zeigen, dass wenige Änderungen
innerhalb der Bakteriengemeinschaften sogar im Gezeitenwechsel zu den Kenterpunkten
stattfinden. Einige Phylotypen waren stets nachweisbar und wurden schon beschrieben, z. B.
das RCA-Cluster und das WAC I-Cluster (Selje et al., 2004, Stevens et al, 2005), andere
Phylotypen jedoch erschienen nur kurzzeitig zu bestimmten Ereignissen. Diese Änderungen
waren im Wesentlichen durch den kombinierten Einsatz von RNA-basierter PCR und
spezifischen Primern für Bacteroidetes, alpha-Proteobakterien und Roseobacter detektierbar,
wodurch die Bedeutung dieser Bakterien für den gesamten Stoffumsatz als eher gering
einzuschätzen ist. Die Frage nach ihrer Funktion bleibt dennoch interessant, da es sich
innerhalb der Bakteriengemeinschaft um funktionell wichtige Prozesse handeln könnte, die zu
bestimmten Zeitpunkten relevant sind. Darüber hinaus war auffällig, dass die Bandenmuster
der aktiven Bakterien (RNA-basiert) stark abwichen von den Bandenmustern der als häufig
anzunehmenden Bakterien (DNA-basiert). Dies lässt den Rückschluss zu, dass die
Anwendung RNA-basierter Methoden bei weiteren Untersuchungen im Wattenmeer dringend
notwendig ist, um Korrelationen zwischen Aktivität und Phylogenie herstellen zu können.
Große Bedeutung hat die Anwendung der FISH- und CARD-FISH Methode, durch die
erstmalig die Abundanz der verschiedenen Bakteriengruppen in der Wassersäule des

Kapitel VI Schlussbetrachtung und Ausblick
147
Wattenmeeres und damit auch der jeweilige potentielle Anteil der Gruppen am Stoffumsatz
gezeigt werden konnte. Die Verknüpfung zu spezifischen tidalen Ereignissen war trotz der
hochauflösenden CARD-FISH nicht erkennbar, jedoch zeigte sich, dass in den hoch
produktiven Frühjahrs- und Sommermonaten wesentlich höhere Varianz innerhalb der
Bakteriengruppen auftrat, was ebenfalls auf erhöhte Aktivität der Organismen schließen
hinweist. Dies bedarf ebenfalls weiterer Untersuchungen, z. B. in Kombination mit der Micro-
FISH Methode, um diese Aussagen stützen zu können.
Kapitel IV zeigte, dass die Actinobakterien im Wattenmeer unerwartet hohe Abundanz und
Artenvielfalt aufweisen. Die Frage nach der ökologischen Bedeutung dieser Organismen für
Küstengebiete rückt daher immer mehr in den Vordergrund und stellt eine interessante
Perspektive für weitere Forschungsarbeiten dar. Auch für diese Gruppe zeigte sich, dass die
Einteilung in die Kompartimente frei lebend und aggregat-assoziiert essentiell ist, um weitere
Rückschlüsse ziehen zu können. Beide Fraktionen zeigten Unterschiede in der
Zusammensetzung der Bakteriengemeinschaft und sind daher auch gesondert zu betrachten.
Besonders interessant ist die klare Unterscheidung von Actinobakterien, die im Süßwasser
leben, und den halotoleranten marinen Vertretern im Wattenmeer. Actinobakterien wurden
bisher durch Produktion von Sekundärstoffen in biotechnologischen Fragestellungen
fokussiert, aber auch im ökologischen Sinne ist diese Eigenschaft untersuchenswert.
Besonders auf Aggregaten können diese als Sekundärstoffe bezeichneten Botenstoffe
besondere Bedeutung für Kommunikation oder Abwehr innerhalb der
Bakteriengemeinschaften einnehmen. Als Hauptursache für die Artenvielfalt und Abundanz
der Actinobakterien in diesem schwebstoffreichen Grenzbereich zwischen Land und Meer
kann jedoch der Abbau komplexer organischer Verbindungen, z. B. Huminstoffe,
angenommen werden.
Der Austausch von Wasserkörpern zwischen dem Wattenmeer und der offenen Nordsee und
damit verbundene Stoffflüsse sind lange Zeit Mittelpunkt sedimentologischer und weiterer
geochemischer Untersuchungen gewesen. Inwiefern Aggregat-assoziierte oder auch frei
lebende Bakterien durch diese Prozesse beeinflusst sind und welchen Einfluss
standortgebundene Bedingungen auf die Bakteriengemeinschaften haben, zeigen die
Ergebnisse aus Kapitel V.

Kapitel VI Schlussbetrachtung und Ausblick
148
Trotz standortspezifischer Unterschiede, die durch die Bandenmuster der partikel-assoziierten
Bakteriengemeinschaften reflektiert wurden, war eine homogene Verteilung frei lebender
Bakteriengemeinschaften zu beobachten. Die phylogenetischen Stammbäume ergaben große
Ähnlichkeit der partikulären Bakteriengemeinschaften des ostfriesischen und nordfriesischen
Wattenmeeres. Es konnten Indikatororganismen ausgemacht werden, die ausschließlich
Aggregat-assoziiert leben und somit den Verlauf partikulären Materials aus den
Wattenmeeren und von resuspendiertem Sediment in der Wassersäule aufzeigen.
Überraschend waren auch hier die eindeutige Trennung der Kompartimente, und die große
Stabilität innerhalb der alpha-Proteobakterien bzw. der Roseobacter-Gruppe. Nicht
veröffentlichte Daten, die im Rahmen dieser Arbeit erhoben wurden, ergaben für die frei
lebenden Roseobacter nahezu identische DGGE-Bandenmuster, unabhängig von der
Jahreszeit und vom Standort. Die ökologische Bedeutung dieser Gruppe für den Umsatz von
gelöstem organischen Kohlenstoff (DOC) ist daher als essentiell anzunehmen und bedarf
dringend einer weiteren Erforschung.
Zusammenfassend ergibt sich daher aus dieser Arbeit ein komplexes Bild mit vertieften
Erkenntnissen über die Zusammensetzung der Bakteriengemeinschaften im ostfriesischen
Wattenmeer und der Deutschen Bucht. Die sensitiven Untersuchungsmethoden konnten
zeigen, welche Einflussfaktoren in diesem als persistent bezeichneten Ökosystem eine
wesentliche Rolle einnehmen und bilden daher die Grundlage für weitere Forschungsarbeiten.
Aktivitätsmessungen und Erhebung des genetischen Potentials für Stoffwechselwege stellen
daher eine geeignete Ergänzung dar, die durch weitere Arbeiten im Rahmen des
Forschergruppenverbundes erhoben werden können. Intensive Datenerhebung von
bakterieller Biomasseproduktion findet derzeit in stündlichen bis saisonalen Zeitskalen statt,
die gleichzeitige Etablierung der Micro-CARD-FISH stellt eine weitere, höchst effektive
Möglichkeit zur Aktivitätsmessung im Wattenmeer dar. Die Überprüfung der Ergebnisse über
experimentelle Ansätze ist derzeit ebenfalls in der Ausführung, und die Verknüpfung zum
natürlichen System wird über mathematische Modelle gelenkt. Die Metagenomik bleibt
vorerst ein zukunftsträchtiger Ausblick, wird aber im Zuge der Fortschritte in der molekularen
Ökologie ein logischer Schritt zur weiteren Erforschung der Bakteriengemeinschaften in der
Wassersäule des Wattenmeeres und der Deutschen Bucht darstellen.

Erklärung
Hiermit bestätige ich, dass ich die vorliegende Dissertation selbständig verfasst und nur die
angegebenen Quellen und Hilfsmittel verwendet habe.
Weiterhin erkläre ich, dass diese Dissertation weder in ihrer Gesamtheit noch in Teilen einer
anderen wissenschaftlichen Hochschule zur Begutachtung in einem Promotionsverfahren
vorliegt oder vorgelegen hat.
Oldenburg, den

Danksagung
Mein besonderer Dank gilt Prof. Meinhard Simon für das Vertrauen, die Unterstützung und
die große Flexibilität, vor allem zum Ende der Fertigstellung dieser Arbeit. Diese
Eigenschaften sind keineswegs selbstverständlich und ich bin dankbar, dass ich diese positive
Erfahrung machen durfte! Vielen Dank an dieser Stelle auch an Prof. Heribert Cypionka, der
unkompliziert die Aufgabe des Gutachters übernommen hat. Für ihn habe ich mich besonders
um Prägnanz bemüht - ich hoffe, es ist gelungen.
Für Zielstrebigkeit, fachliche Diskussion und einfach immer wohltuenden freundlichen Rat
möchte ich Dr. Thorsten Brinkhoff danken. Er hat mich mehr als einmal vor dem Verzetteln
bewahrt und ist mir ein großes Vorbild im Umgang mit Kollegen. Vielen Dank auch für die
große Flexibilität zum Ende meiner Arbeit!
Der gesamten Arbeitsgruppe, besonders Andrea Schlingloff und Birgit Kürzel, möchte ich
danken für die Geduld, Offenheit und Hilfsbereitschaft. Heike Stevens bin ich mehr als
dankbar für 2500000000 motivierende Gespräche und ihre erfrischende Art, das Leben positiv
zu sehen. Und für die Fahrt im Schäbi Pop durch Yucatan! Katja Walther und Beate Köpke
danke ich für Fachgespräche zu jeder Tages- und Nachtzeit und für die Freundschaft, die eine
sehr wichtige Begleitung für mich war und ist.
Die größte Unterstützung fand ich bei meiner Familie, die mir jederzeit zur Seite stand und
mir gezeigt hat, wie wichtig dieser Halt für mich ist. Ihr habt alle einen großen Teil zu dieser
Arbeit beigetragen! Das werde ich nie vergessen und daher widme ich Euch meine Arbeit aus
tiefstem Dank.
Meinem Freundeskreis, für den das Wort „Doktorarbeit“ mittlerweile fest mit Begrüßungs-
und Abschiedsformeln verknüpft ist, danke ich für unendliche Ausdauer und Geduld, für
seelischen Ausgleich, für tolle Gespräche, und fürs Dasein. Es ist geschafft!!! Ihr könnt
wieder ans Telefon gehen!

Lebenslauf
Beate Rink, geboren am 23.01.1974 in Bremerhaven
Schulausbildung
1980-1984 Grundschule
Friedrich-Ebert-Schule, Bremerhaven
1984-1986 Orientierungsstufe
Wilhelm-Raabe-Schule, Bremerhaven
1986-1990 Gymnasium
Wilhelm-Raabe-Schule, Bremerhaven
1990-1993 Gymnasiale Oberstufe
Geschwister-Scholl-Schule, Bremerhaven
Leistungskurse: Latein, Kunst
Abschluß: Abitur
Berufsausbildung
1993-1995 Ausbildung zur Industriekauffrau
Fa. Schottke GmbH & Co. KG, Bremerhaven
Abschluß: Industriekauffrau
1995-1997 Biologiestudium
Universität Regensburg
1997-2001 Biologiestudium
Carl-von-Ossietzky-Universität, Oldenburg
Hauptfächer: Mikrobiologie, Genetik, Biochemie
Abschluß: Diplom
Thema Diplomarbeit: „Besiedelung und Abbau von Fucus-detritus durch
heterotrophe marine Bakterien“
2001-2006 Wissenschaftliche Angestellte am Institut für Chemie und Biologie des Meeres
Carl-von-Ossietzky-Universität, Oldenburg
Arbeitsgruppe „Biologie geologischer Prozesse“, Prof. Meinhard Simon