Embed Size (px)
Citation preview

474
STUDIES IN REPTILIAN COLOUR CHANGESII. THE PITUITARY AND ADRENAL GLANDS IN THE REGULA-
TION OF THE MELANOPHORES OF ANOLIS CAROLINENSIS
BY L. H. KLEINHOLZ, PH.D.
Biological Laboratories, Harvard University
(Received 12 January 1938)
(With Four Plates and Three Text-figures)
I. INTRODUCTIONTHE early literature of reptilian colour changes deals almost exclusively with theAfrican chameleon. In recent years the investigation of metachrosis in the chame-leontid reptiles has been leading to a comprehensive knowledge of the physiologicalmechanisms involved in this activity, chiefly through the studies of Hogben &Mirvish (1928) and of Zoond & Eyre (1934). Investigations of the pigmentaryactivity of lizards native to North America began at the beginning of the presentcentury. Our knowledge of the chromatic function in these forms has progressedthrough the publications of Parker and his associates. The early reports of Carlton(1903) and of Parker & Starratt (1904) on Anolis and of Parker (1906) and of Red-field (1918) on Phrynosoma served to define the bionomic aspects of colour changefor these forms.
Studies of pigmentary activity in the New World lizards were greatly influencedby the views of those nineteenth-century investigators who had engaged in theanalysis of colour mutability. Briicke (1852), Krukenberg (1880) and Keller (1895)found that the chromatic responses of African chameleons were under the controlof the nervous system. Later investigators of chromatophoral activity were soconvinced of this mode of control that for a long time no other regulating mechanismwas suspected. Redfield (1918), investigating the effects of adrenalin on the melano-phores of Phrynosoma, found this hormone to be a potent agent in inducingchromatophoral changes. Redfield's explanation of colour changes was a novel onebut soon gained apparent support by the demonstration that the chromatic functionin amphibians was under endocrinal control (Smith, 1916; Allen, 1917; Atwell,1919, 1921; Hogben & Winton, 1922a, 19226, 1923).
These recent developments in the physiology of chromatophoral behaviourwere a distinct advance in establishing a hormonal basis for the activity of sucheffectors. Little, however, is known of the role played by blood-borne substancesin reptilian colour changes. Redfield (1918) believed that in Phrynosoma "themelanophores are co-ordinated by two distinct mechanisms, the adrenal secretionand the direct action of nerves", either mechanism alone being capable of effecting

Studies in Reptilian Colour Changes 475
melanophore concentration (p. 309). But Hogben & Mirvish (1928) and Zoond& Eyre (1934), working with African chameleons, disputed Redfield's results.Carlton (1903) and May (1924) believed that chromatophoral responses in Anoliswere regulated by nerves. Noble & Bradley (1933), incident to their study of themoulting process in lizards, observed that hypophysectomized specimens of Herm-dactyhs were pale in colour.
While examining the hypophysis of Anolis carolinensis, I was prompted by thehighly secretory appearance of the intermediate lobe (unpublished results) toinvestigate the part played by the pituitary gland in the colour changes of thislizard. Results from preliminary experiments (Kleinholz, 1935, 1936), coupledwith the need of revising several concepts that were extant in the literature, ledto an extension of this work to include the various factors, nervous as well as endo-crine, that might be involved in the chromatic responses of this lizard. This studyis a report of the results obtained from experiments designed for that purpose.
It is a great pleasure to acknowledge my most appreciative thanks to Prof. AldenB. Dawson for his interest and helpful criticism and to Prof. G. H. Parker for hisfriendly encouragement during the course of this study.
II. MATERIALS AND METHODS
A plentiful supply of Anolis carolinensis was obtained from collectors in thesouth-eastern part of the United States, which is one of the natural habitats of thislizard. In the laboratory the animals were fed with meal-worm larvae and blow-flies. Water was supplied by sprinkling the cages daily.
Use was made of a heavy cardboard carton in studying adaptations to back-grounds. One hah0 of the carton was painted white and was separated by a partitionfrom the second half, which was painted black. Lizards were kept in individualglass jars which could be moved from one coloured background to the other withoutactual handling of the animals. The backgrounds were illuminated by a 100 W.electric lamp at a distance of 30 in. from the bottom of the box.
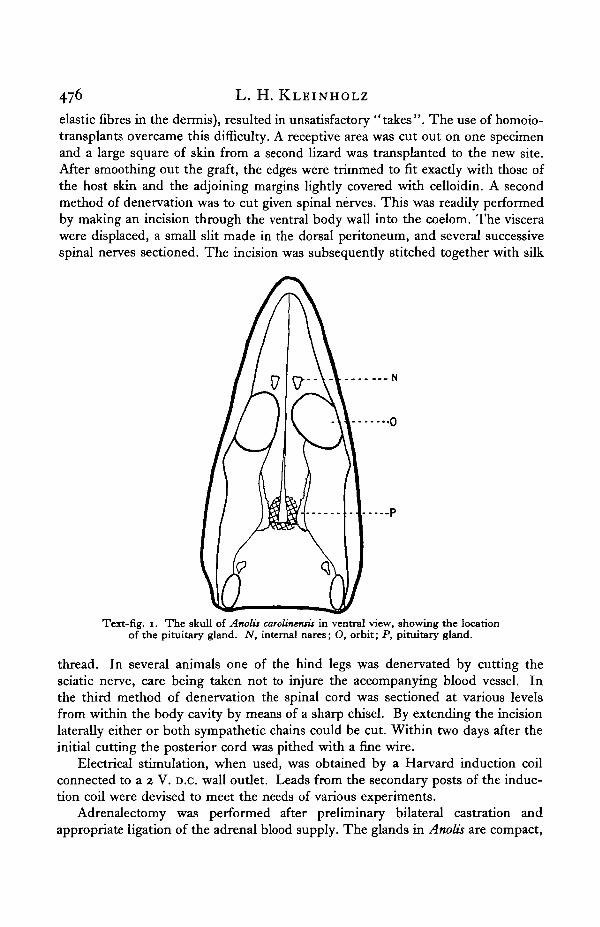
Surgical operations were performed under light ether anaesthesia, with noparticular regard for sterile conditions. Hypophysectomy was accomplishedthrough the oral route, the pituitary being visible through the floor of the skull asa pinkish-white mass, located about midway between the level of the orbits and theposterior margin of the skull (Text-fig. 1). After displacing the bone in this region,the membranous investment of the gland was slit and the entire pituitary removedwith the aid of a suction pipette. The slight amount of haemorrhage which occursis negligible and soon stops; recovery from the effects of the operation is prompt.
Lizards were blinded by cauterizing both eyes. Optic enucleation provedimpractical because of a large blood sinus situated close to each eye, so that anysuch attempts resulted in fatal haemorrhage.
Denervation of given regions of skin was accomplished in several ways, one ofwhich was by making transplants. In autotransplantations the areas of the graftsand those of their new sites, changing in shape and size (presumably because of
476 L. H. KLEINHOLZ
elastic fibres in the dermis), resulted in unsatisfactory "takes". The use of homoio-transplants overcame this difficulty. A receptive area was cut out on one specimenand a large square of skin from a second lizard was transplanted to the new site.After smoothing out the graft, the edges were trimmed to fit exactly with those ofthe host skin and the adjoining margins lightly covered with celloidin. A secondmethod of denervation was to cut given spinal nerves. This was readily performedby making an incision through the ventral body wall into the coelom. The viscerawere displaced, a small slit made in the dorsal peritoneum, and several successivespinal nerves sectioned. The incision was subsequently stitched together with silk
N
Text-fig. i. The skull of Anolii carotineniit in ventral view, showing the locationof the pituitary gland. N, internal nares; O, orbit; P, pituitary gland.
thread. In several animals one of the hind legs was denervated by cutting thesciatic nerve, care being taken not to injure the accompanying blood vessel. Inthe third method of denervation the spinal cord was sectioned at various levelsfrom within the body cavity by means of a sharp chisel. By extending the incisionlaterally either or both sympathetic chains could be cut. Within two days after theinitial cutting the posterior cord was pithed with a fine wire.
Electrical stimulation, when used, was obtained by a Harvard induction coilconnected to a 2 V. D.C. wall outlet. Leads from the secondary posts of the induc-tion coil were devised to meet the needs of various experiments.
Adrenalectomy was performed after preliminary bilateral castration andappropriate ligation of the adrenal blood supply. The glands in Anolis are compact,

Studies in Reptilian Colour Changes 477
no scattered chromaffine tissue being macroscopically evident. Recovery from theoperation took place within 2 hr., and individuals survived from 10 to 24 hr.
III. THE HISTOLOGY OF THE MELANOPHORES IN COLOUR CHANGES
In its colour play Anotis ranges from dark brown to bright green, intermediateshades including light brown, yellow, yellowish green and emerald-green.
Individual lizards are not uniformly coloured but show considerable variationin certain regions of the body. A dewlap, or throat fan, is present, being mosthighly developed in males. When erected in combat or in courtship it is brilliantred, due, in part, to granules of pigment in the skin of that region. In some animalsa mid-dorsal longitudinal band is present, extending from the cervical region forvarying distances toward or even along the tail (Text-fig. 3); in many lizards it iscompletely absent.
When an animal is in the dark condition, a regular darker pattern is discernibleagainst the brown colour of the rest of the body; such markings vary among speci-mens but are constant in form for a particular individual. The darker areas consistof groups of from two to a score or more of scales scattered over the dorsal andlateral aspect of the body and appendages. In addition to this body pattern alllizards in the dark state have a darker quadrangular patch immediately posteriorto each orbit. In some individuals a similar area is located over each scapula. Theselocalized markings will be considered again below and will be referred to as thepost-orbital and the scapular regions.
In the light state the dorsal and lateral surfaces of the body are green. Theventral surface ranges from cream colour to brownish white in appearance and isfrequently peppered over with black spots.
A brief description of the important histological features of the skin will bereported here in order to provide a better understanding of the processes involvedin the mechanics of metachrosis. Von Geldern's (1921) paper gives a good descrip-tion of the finer details. Skin from lizards in various colour states was fixed byimmersion in hot water (80° C.) for 10 sec. to fix the position of the pigment.Embedding in paraffin was effected by the rapid dioxane method. Some sectionswere stained with Mallory's azan while others were mounted unstained.
The epidermis is a transparent layer, typically divisible into the stratumcorneum and the stratum germinativum. Just beneath the epidermis is a layer ofyellow oil droplets and xanthophores. These are not present in ordinary histologicalpreparations, having been dissolved by the fat solvents usually employed in suchtechniques, but can be seen in sections cut by the freezing method. Internal tothe oil droplet layer lies a much thicker region known as the leucophore layer;this is composed of a number of irregularly spaced blocks or plates, the long axesof which are parallel to each other. In unstained sections the leucophore regionappears brownish by transmitted light but is bluish white when examined byreflected light.
Below the leucophore layer are situated the melanophores, which are the cells

478 L. H. KLEINHOLZ
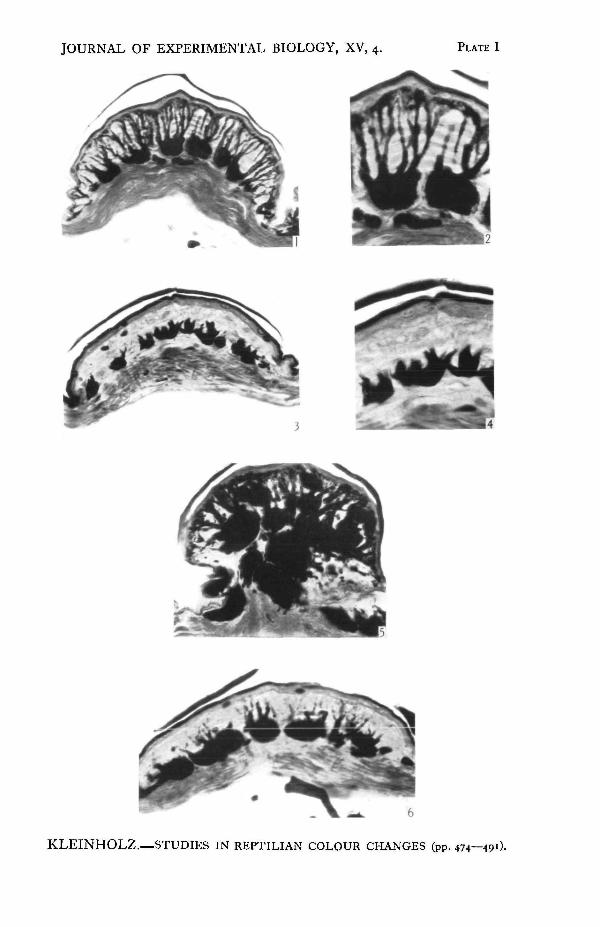
actively concerned in the colour changes of Anolis. The cell bodies of the melano-phores lie embedded partly in the leucophore layer and partly in the deeper con-nective tissue of the dermis. A varying number of branches, extending verticallyupward from the cell bodies of the melanophores (PL I, fig. 1), pass through theinterstices of the leucophore layer and are subdivided into many finer brancheswhich terminate beneath the inner surface of the epidermis.
When the skin is in the brown state the pigment granules are dispersed throughthe fine terminal branches of the melanophores, resulting in a layer of pigment justbeneath the epidermis (PL I, figs. 1 and 2). In the green phase of skin colour thepigment is present only in the proximal portions of the branches, having been with-drawn from beneath the epidermis (PL I, figs. 3 and 4). The finer distal branchesare practically transparent except for a few scattered granules which may havefailed to migrate proximally with the main mass of the pigment.
It is evident that the melanophores are the cells actively concerned in thevarious colour phases of Anolis. The other components of the skin, namely the oildroplets, the xanthophores and the leucophore layer, play a passive or at most avery restricted part in metachrosis, acting as filters or reflectors.
IV. THE EFFECT OF LIGHT ON THE COLOUR CHANGES OF ANOLIS
A. Responses to changes in backgrounds
It is now almost universally recognized that conditions of illumination and oftemperature are the principal environmental factors concerned in reptilian colourchanges. Normal specimens of Anolis carolinensis are brown at room temperature(18-280 C.) in the light and are green in darkness. These observations, first reportedby Carlton (1903), have been amply confirmed by Parker & Starratt (1904) and byvon Geldern (1921). Hadley (1929, 1931) extended the observations to includeAnolis porcatus and A. todurus. Similar behaviour of the melanophores of thehorned toad was reported by Parker (1906) for Phrynosoma blainvillei and by Red-field (1918) for P. comutum.
Colour responses of Anolis to changes in background have not been extensivelyinvestigated. Hadley (1931) found that Anolis todurus underwent adaptive changesto white and to black backgrounds, but failed to state the time required for theseprocesses. The species of Anolis used in this study showed active responses to suchenvironmental changes. Lizards in the dark state, when placed upon a brightlyilluminated white background, become green; conversely, green lizards, whenplaced upon an illuminated black background, rapidly become brown. The timecurves for such chromatic adaptations (Text-fig. 2) were plotted by assigning anumerical value (Table I) to each of several stages in the progressive changes ofcolour from the light to the dark condition (Hogben & Slome, 1931).
The time relations shown in Text-fig. 2 demonstrate that the onset of the darkstate (curve A) is rapid, approaching a maximum after the lizard has been on ablack background for approximately 6 min. The rate of change in colour of a lizardin the dark condition when it is placed on an illuminated white background (curve
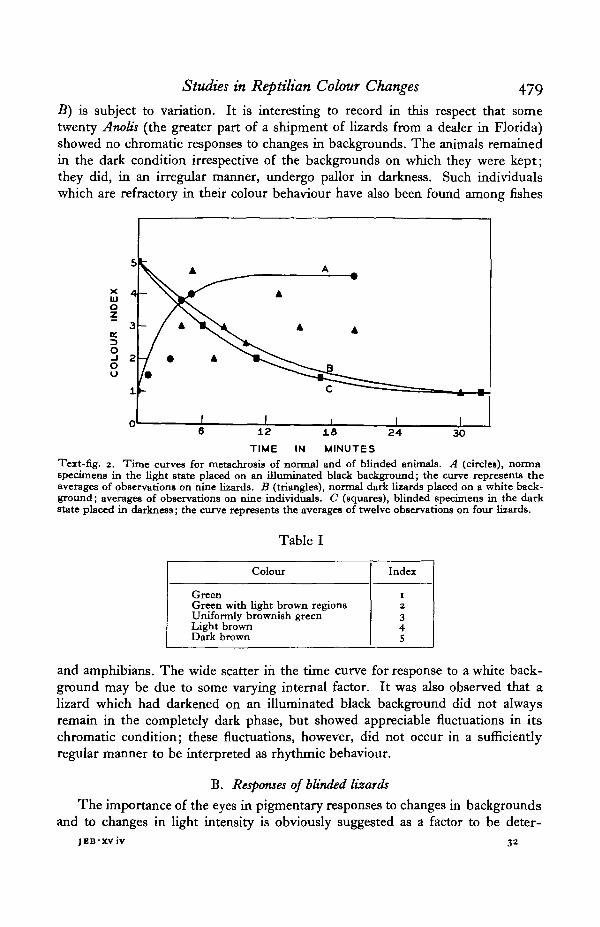
Studies in Reptilian Colour Changes 479
B) is subject to variation. It is interesting to record in this respect that sometwenty Anolis (the greater part of a shipment of lizards from a dealer in Florida)showed no chromatic responses to changes in backgrounds. The animals remainedin the dark condition irrespective of the backgrounds on which they were kept;they did, in an irregular manner, undergo pallor in darkness. Such individualswhich are refractory in their colour behaviour have also been found among fishes
TIME IN MINUTESText-fig. 2. Time curves for metachrosis of normal and of blinded animals. A (circles), nonnaspecimens in the light state placed on an illuminated black background; the curve represents theaverages of observations on nine lizards. B (triangles), normal dark lizards placed on a white back-ground; averages of observations on nine individuals. C (squares), blinded specimens in the darkstate placed in darkness; the curve represents the averages of twelve observations on four lizards.
Table I
Colour
GreenGreen with light brown regionsUniformly brownish greenLight brownDark brown
Index
1234S
and amphibians. The wide scatter in the time curve for response to a white back-ground may be due to some varying internal factor. It was also observed that alizard which had darkened on an illuminated black background did not alwaysremain in the completely dark phase, but showed appreciable fluctuations in itschromatic condition; these fluctuations, however, did not occur in a sufficientlyregular manner to be interpreted as rhythmic behaviour.
B. Responses of blinded lizards
The importance of the eyes in pigmentary responses to changes in backgroundsand to changes in light intensity is obviously suggested as a factor to be deter-
JEB-XViv 32

480 L. H. KLEINHOLZ
mined experimentally. So far as I know, the effects of blinding upon colour muta-bility have not been thoroughly studied in any of the American lizards.
The generalized statements of some investigators on the colour mutabilityresulting from blinding are confusing, because in most cases no distinctions aredrawn between responses to backgrounds and responses to light and to darknessafter such operations. Von Buddenbrock (1928) says (p. 397): "Bei vielen Krebsen,den Fischen und den Amphibienlarven ist der Erfolg dieses Eingriffs stets einestarke Verdunklung. Anderseits hat Blendung keinen Einfluss auf die Farbung derTintenfische, und Reptilien."
Parker (1936, p. 2) says: "The eyes of animals are essential to their colourchanges, for when these organs are removed or effectually covered all signs of suchchanges disappeared, and the given animal so far as an alteration of its tint wasconcerned was largely incapacitated"; although in the same publication (p. 23) adistinction is made between background responses and the responses to change inlight intensity: "Another feature which should be noted in passing is that aFundulus from which the eyes have been removed is pale in darkness, as normalindividuals are, but moderately dark in bright illumination."
Four specimens of Anolis, after they had been tested and found to be normalin their colour responses to background changes, were blinded by cauterization ofthe eyes. Twenty-four hours were allowed for recovery from the shock of theoperation before the animals were used in the following experiments.
When these otherwise normal lizards were tested, it was found that the colourresponses to backgrounds had disappeared, but that the typical chromatic reactionsto light and to darkness persisted apparently unimpaired. The animals were placedin the dark room while they were brown. At the end of 7 min. one individual wasentirely green and the remaining three were perceptibly lighter in colour. A secondlizard was green at the end of 17 min., while the other two required 38 min. toattain the stage of complete pallor. When the animals were kept on either black orwhite backgrounds which were illuminated from above, they attained the darkstate within 6 min. The average time required for darkening of blinded individualswhen illuminated, computed from eighteen observations on these four lizards, was47 min. The average time for attaining a condition of complete pallor in darkness(Text-fig. 2, curve C) was 22-4 min., calculated from twelve readings on the sameanimals. These time periods agree with those reported by Carlton (1903) and byParker & Starratt (1904) for normal animals (Table II).
The fact that blinded lizards, although they have lost the ability to adapt theircolours to backgrounds, are still able to undergo metachrosis in response to changesin light intensity implies either that the melanophores in the skin are directlyresponsive to light and to darkness, or that some photoreceptive mechanism, inaddition to the eyes, is involved in the system responsible for colour changes.Such possibilities will be considered more critically later.

Studies in Reptilian Colour Changes 484
V. THE DARK PHASE
A. The effects of hypophysectomy
Within an hour after hypophysectomy such successfully operated animals turngreen and remain permanently in the light condition (Text-fig. 3); colour responsesto background changes disappear. In several cases where removal of the pituitary
Text-fig. 3. On the left is a normal Anolit in the dark condition; the white mid-dorsal stripe extendsfrom the cervical region on to the tail. The animal on the right has been hypophysectomized and isconsequently in the light state.
was apparently incomplete the lizards remained in the light state for 10 to 20 days,after which they darkened slightly when kept on an illuminated black background;post-mortem examination of the pituitary region of such individuals showed thatportions of the intermediate lobe were still present, protected by the ridge of bonethat forms the posterior wall of the sella turcica. The pale condition of hypo-
is-*

482 L. H. KLEINHOLZ
physectomized specimens of Anolis is not a result of mechanical manipulation orinjury undergone during the operation because lizards in which the bone sur-rounding the pituitary had been chipped away, and animals from which the greaterpart of the anterior lobe had been removed, underwent normal colour changes.Lizards from which the pituitary gland has been removed, and which have beenkept in darkness, do not respond to illumination by darkening but remain in acondition of pallor.
It was shown above that blinded individuals, although having lost the abilityto respond to changes in backgrounds, undergo chromatic changes in response t6changes in light intensity, becoming dark when illuminated and pale when kept indarkness. Such behaviour implies either a direct reactivity of the melanophores
Table II
Average times to change
From brownto green
2 5 0 min.IQ-622-4
From greento brown
40 min.4-247
Temperature
?20° C.22°
Investigator
CarltonParker-StarrattThis paper
to light or the presence of photoreceptors other than the eyes. If blinded lizardsare hypophysectomized, however, they become green and remain so, notwith-standing that they are maintained in an illuminated environment. Equally note-worthy is the fact that specimens of Anolis which have been successfully hypo-physectomized do not darken, either in direct or in diffuse light, after they havebeen blinded. These experiments preclude the possibility that melanophores inthe intact skin of Anolis carolinensis are directly responsive to light. Such resultsdiffer from those reported by Hadley (1931, p. 325), who studied the behaviour ofisolated skin.
If the circulation is excluded from any part of the skin of Anolis, that regionbecomes green and remains in this light condition regardless of the fact that theanimal is kept on an illuminated black background. This can be readily demon-strated by ligating one of the legs, the skin of which becomes green within 15 min.Removal of the ligature while the lizard is still on its black background soon resultsin a darkening of the hitherto green skin. Similar results were obtained after thefemoral artery of a green lizard was exposed and ligated. It follows from theseresults that a portion of skin entirely removed from the body should, since thechromatophorotropic hormone can no longer act on the melanophores through theblood stream, turn green. Such is actually the case.
B. Injection of pituitary extracts
To supplement the experiments involving hypophysectomy, extracts of thepituitary glands of various animals were injected into lizards that were in the lightstate, either as a result of hypophysectomy, or of having been kept in darkness or

Studies in Reptilian Colour Changes 483
on an illuminated white background. Injections were in all cases made intra-peritoneally. This is necessary because subcutaneous injections of distilled wateror of Ringer's solution are capable of producing a dark spot at the site of injection,possibly because of the pressure exerted on the melanophores by the wheal offluid. Carlton (1903) found that a piece of isolated skin turned green but could bemade to darken by tapping it gently with a blunt instrument. The results of suchinjections are shown in Table III.
In teleost fishes the hypophyses are not distinctly divided into definite lobes;consequently the entire gland was used in the case of Fundulus heteroclitus. In-jection of such extracts into normal specimens of Anolis which were in the lightstate (through having been kept in darkness) and into hypophysectomized lizards(Kleinholz, 1935) resulted in a pronounced darkening. In frogs and in Anolis onlythe neuro-intermediate lobes were used in preparing extracts. Experiments B andC illustrate in the first case the rapidity with which the darkening response wasevoked and in the latter that the induced dispersion of the melanophores maypersist for several hours. In experiment E a commercial preparation of mammalianposterior lobe in 0-5 % chloretone was used; control injections with chloretonesolution were not made.
C. Early evidence for nervous control of darkening
The results from hypophysectomy and from injecting pituitary extracts proveclearly that the dispersed condition of the melanophores in the skin of Anolis iseffected by a hormone from the hypophysis.
The evidence of earlier investigators that melanophore dispersion in Anoliswas under direct nervous control is fragmentary and inconclusive. Carlton (1903),describing the darkening that resulted when a green Anolis was brought from a darkbox into the light, offered two possible explanations for such behaviour; the changemight be due either to the direct action of light on the melanophores or to themediation of the nervous system which was stimulated by light. By proving thatlight which fell on one part of the animal's body induced darkening in the othernon-illuminated portions, he eliminated the possibility of the direct action of lighton the melanophores, and came to the alternative view because he knew "of noway of explaining the induced changes except on the assumption that nervesserved as intermediate organs". Cutting and destruction of the posterior cord hadno effect on the ability of a lizard to darken completely under environmental con-ditions which normally evoked dispersion of the melanophores. By a process ofelimination Carlton came to the conclusion that this phase of melanophore activitywas controlled by the sympathetic nervous system. His sole evidence for this viewwas the fact that brown lizards turned green after injection of nicotine. It has beencommonly observed, however, that ordinary handling of this lizard readily inducesthe green state. At that time the possibility of an endocrine factor in metachrosiswas practically unsuspected.
May's belief (1924) that the colour changes of Anolis were due to the trans-mission of stimuli over nerve fibres supplying the melanophores resulted from a
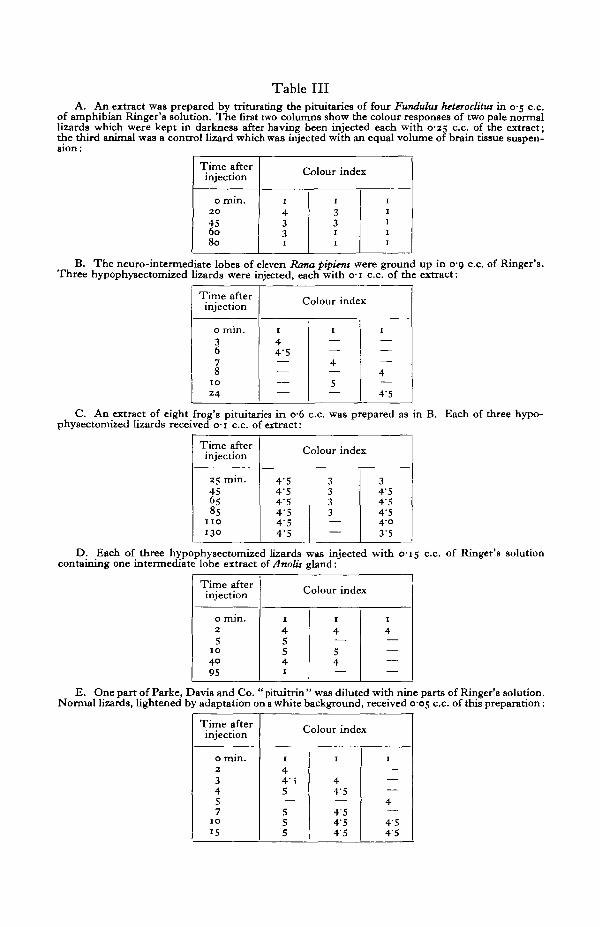
Table IIIA. An extract was prepared by triturating the pituitaries of four Funthdus heteroclitw in 0-5 c.c.
of amphibian Ringer's solution. The first two columns show the colour responses of two pale normallizards which were kept in darkness after having been injected each with 0-25 c.c. of the extract;the third animal was a control lizard which was injected with an equal volume of brain tissue suspen-
Time afterinjection
0 min.2 0
456080
1
4331
Colour index
1
331
1
1
1
1
1
1
B. The neuro-intermediate lobes of eleven Rana pipiens were ground up in 0-9 c.c. of Ringer's.Three hypophysectomized lizards were injected, each with o-i c.c. of the extract:
Time afterinjection
0 min.
6
810
24
Colour index
1
44-S
1
4
5
1
4
4'5
C. An extract of eight frog's pituitariea in o-6 c.c. was prepared as in B. Each of three hypo-physectomized lizards received o-i c.c. of extract:
Time afterinjection
25 min.456585
110
130
Colour index
4'54-54-54-54-54-5
3333
34'54-54-5403-5
D. Each of three hypophysectomized lizards was injected with C15 c.c. of Ringer's solutioncontaining one intermediate lobe extract of Anolis gland:
Time afterinjection
0 min.2
5I O
4095
Colour index
1
4554I
I
4—54—
1
4————
E. One part of Parke, Davis and Co. " pituitrin " was diluted with nine parts of Ringer's solution.Normal lizards, lightened by adaptation on a white background, received 0-05 c.c. of this preparation:
Time afterinjection
0 min.23457
10
15
Colour index
1
44i5
55
1
44'5
454'545
1
4
4'545

Studies in Reptilian Colour Changes 485
study of the chromatic responses of skin in auto- and in homoio-transplantations.In his experiments, areas of skin which had been denervated (by the operation oftransplantation) lost their capacity for colour changes over a period of 2 to 3 weeks,although during this period the skin of the host was normal in its chromaticbehaviour. At the end of this time the engrafted skin resumed pigmentary activityin concurrence with the skin of the recipient. May attributed these results to theregeneration of nerve fibres into the hitherto denervated region.
D. Behaviour of denervated skin
I am able to confirm Carlton's results obtained from cutting nerves. Lizardswhose spinal cords had been sectioned at various levels were able to becomeuniformly brown if kept on black backgrounds; similarly, animals whose posteriorcords had been pithed did not differ in their colour changes from normal in-dividuals. Cutting the sciatic nerve of one leg had no effect on the pigmentarycondition, the skin of that leg behaving in all respects like that of the controlmember. Uni- or bi-lateral section of the sympathetic chain had no visible effecton the ability of the lizard to darken. It is also interesting to note that neither vonGeldern (1921) nor May (1924) were successful in their attempts to demonstrateinnervated melanophores in skin treated by the usual neuro-histological methods.
By making skin transplants and then testing for colour changes on black and onwhite backgrounds, it was found that, although considerable variation existed, thegrafts showed unmistakable melanophore activity. As can be seen from Table IV,the grafts in several cases of auto-transplantation evidenced metachrosis within24 to 48 hr. after the " denervating " operation of transplantation; in other instancesthe denervated patches did not change from the initial green colour until the graftwas 7 to 8 days old. In animal 6 (Table IV) when the reciprocal auto-transplantswere one day old the graft on one side required 75 min. longer than the other tobecome brown, while several days later the two skin patches darkened at approxi-mately the same time. These variations are undoubtedly due to differences in thevascularization of the grafts, since the transplants did not "take" in good contactwith the receptive areas in many cases. That this explanation is correct is seen fromthe fact that homoioplastically transplanted skin (which was fitted smoothly intothe region excised for it on the host) in all cases turned brown when the host waskept on an illuminated black background. Further support is given this view whenthe effects of injections of "pituitrin" on host and on graft skin colour are studied.In all cases there was an obvious temporal difference in the response, the trans-planted regions being appreciably slower than the skin of the host in their rates ofdarkening (Table IV and PI. II).
E. Effects of electrical stimulation
To determine whether the dark state might in addition be induced by a nervousagency, various methods of electrical stimulation were used. Leads from thesecondary posts of the induction coil were inserted one into the cloaca and thesecond into the animal's mouth; the passage of current caused the lizard to clamp
486 L. H. KLEINHOLZ
its jaws upon the mouth electrode, thereby maintaining a closed circuit. Thesource of current was a 2 V. D.c. wall outlet. Stimulation was kept constant in allcases, the secondary coil being at a distance of 6 cm. from the primary and thecurrent being allowed to pass through the animal for exactly 2 min.
Table IVA. Auto-transplants. The numbers in parentheses after the method of treatment indicate the
age of the graft in days. The time periods which are marked with a cross (+ ) are only approximate,the mean values having been taken of the two observations between which the colour change occurred.The "pituitrin" was diluted with four parts of distilled water.
Serialnumber
1225
6
66122
Method oftreatment
Background (7)Background (2)Background (9)Background (i)Background (7)Background (1)
Background (7)Background (10)Pituitrin inj. (5)Pituitrin inj. (2)Pituitrin inj. (8)
Time to change
From brown to green
Host
60 min.
25
Graft
95 min.
15
From green to brown
Host
15 min.
15
525
Graft
40 min.195240 +200210195270210
60251512
B. Homoio-transplants. The pituitrin was diluted with nine parts of distilled water.
Serialnumber
899
12131314121314
Method oftreatment
Background (1)Background (1)Background (1)Background (2)Background (1)Background (1)Background (3)Background (3)Pituitrin inj. (3)Pituitrin inj. (3)Pituitrin inj. (3)
Time to change
From brown to green
Host
5 min.10108
186
Graft
30 min.120 +90 +54
3232
From green to brown
Host
S min.
15
449
Graft
9hr.+
2iomin.+60
131014
Five normal animals were placed in darkness to attain the light state. Each wasthen stimulated in turn with a tetanizing current. At the end of the period ofstimulation, when examined for an instant by the rays of a flashlight, the lizardswere found to be darkening; several minutes later they were uniformly dark brownin colour. On the average, it required 4-2 min. after cessation of electrical stimula-tion for the five lizards to become maximally dark (in agreement with the time formelanophore dispersion shown in Text-fig. 2 and in Table II).

Studies in Reptilian Colour Changes 487
These results might be interpreted as being due to reflex stimulation of thepituitary gland with consequent liberation of the melanophore-dispersing hormoneinto the circulation. When, however, the method of stimulation was slightlychanged, the same results were not obtained. In the first modification both pointsof a bipolar pencil electrode were placed in the cloaca; with the second method oneelectrode was inserted in the cloaca and the other into the spinal cord. Although theintensity and duration of the stimulation were the same as those which had causedgeneralized darkening in the original experiments, in neither of these cases wasgeneralized darkening effected; the only observable change was a darkening of thepost-orbital regions and of scattered groups of scales over the otherwise green body.It appears unlikely that the generalized darkening was due to reflex stimulation ofthe pituitary; such melanophore dispersion resulting from electrical excitation wasprobably due to direct stimulation of the pituitary. If this be the case it is difficultto understand the darkening of scattered groups of scales which occurred with themodified methods of electrical stimulation. Such results could be explained on thebasis of a lower concentration threshold of these regions for the melanophore-dispersing principle.
To test this last possibility more critically, four hypophysectomized lizardswere stimulated electrically, one of the electrodes being placed in the cloaca andthe other in the mouth; the duration and the intensity of the stimulation were thesame as those used in the earlier trials. Within two minutes after cessation ofelectrical stimulation the post-orbital regions of these individuals became intenselyblack; in addition, clusters of black spots appeared in an irregular pattern againstthe generalized green colour of the skin (PI. III). The black spots consisted ofgroups of from two to twenty or more scales. This pigmentary condition of thelizard I shall call mottled.
The mottling pattern persists in an hypophysectomized animal for at least20 min. after cessation of electrical stimulation. Within an hour it has usuallydisappeared and the lizard is again uniformly green. This experiment provesdefinitely that the generalized darkening obtained originally by electrical stimulationis dependent upon the pituitary and not at all upon a direct nervous agency; it alsoshows that the mottling pattern may be mediated by an entirely different agency, sincethe response was evoked in hypophysectomized lizards. The nature of the controllingmechanism for the mottling response was next investigated.
VI. THE MOTTLING RESPONSE
A. Behaviour of denervated skin
Since the mottling pattern and darkening of the post-orbital areas are evokedunder the same conditions, the latter regions were selected as convenient ones forexperimental study. Denervation of the skin of this area on one side of an hypo-physectomized lizard by excision and subsequent replacement in its original positiondid not prevent darkening on appropriate stimulation. Two minutes after thecessation of stimulation the control side began to darken while the graft on the left

L. H. KLEINHOLZ
side was still green, but 4 min. later the post-orbital regions of both sides wereblack. This condition persisted for at least an hour; at the end of 2 hr. both sideswere again green. When the responses of denervated post-orbital patches toelectrical stimulation were observed in several lizards on the third, fifth, tenth andeleventh days after transplantation (ample time for degeneration of severed nervefibres) they were found not to differ from the control regions of the unoperatedsides. Darkening of the post-orbital regions is therefore not under direct nervouscontrol.
B. The role of the adrenals
In the absence of nervous mediation, it appeared that the mottling responsemight be regulated through the circulatory system. Darkening of the post-orbitalregions and mottling of the body also appear when the animal is excited by mechan-ical manipulation, such as prodding the head with a glass rod, or pinching the snoutwith forceps. Von Geldern (1921) has reported a similar colour change in the skinof males preparing for combat; Dr L. T. Evans, while studying territorial defenceand the fighting behaviour of Anolis carolinensis, has observed the same pigmentarycondition (personal communication).
This reaction is apparently associated with conditions of emotional excitementin these lizards. Redfield (1918) found that in Phrynosoma tetanization of the roofof the mouth caused generalized pallor, a condition which he called excitementpallor and which he attributed to the secretion of adrenalin. Sand (1935) found thatemotional excitement of the African chameleon, Lophosaura, provoked by pinchingthe feet of the animal with forceps, induced darkening of the skin within 2 min.
To determine whether the mottling response might be evoked by adrenalin,five hypophysectomized lizards were each injected with 0-05 c.c. of Parke, Davisand Co. solution of adrenalin chloride, in a concentration of 1:40,000. The mottlingpattern became evident with 1 min., and 2 min. later stood out strikingly againstthe otherwise green colour of the body. A Ringer-solution extract of both adrenalsfrom one Anolis was similarly effective in inducing the mottling pattern in a secondlizard. Because these results are exact duplications of the pigmentary conditionsresulting from' electrical and from mechanical stimulation of hypophysectomizedspecimens, they indicate that the hormone which appears in the circulation ofstimulated lizards may be adrenalin.
The assumption that the mottling reaction of Anolis is due to the secretion ofadrenalin is substantiated by the following series of experiments. A lizard washypophysectomized, and 6 hr. later, when recovery from the effects of the operationwas complete, the animal was stimulated electrically in the standardized manner.The mottling pattern appeared within 1 min. after cessation of electrical stimulation.On the following day the animal was anaesthetized and a bilateral adrenalectomyperformed. Three hours later, when the mottling colouration provoked by theoperation had worn off completely and the lizard was uniformly green, the animalwas again stimulated. No mottling appeared, even for as long as 15 min. followingstimulation. The animal was then injected with 0-05 c.c. of 1:40,000 adrenalin.

Studies in Reptilian Colour Changes 489
Four minutes later the mottling pattern stood out in strong contrast against thegeneralized green colour of the skin (PI. IV). Repetition of the same procedure onsimilarly prepared lizards consistently yielded the same results: electrical stimula-tion of individuals which had been both hypophysectomized and adrenalectomizedfailed to evoke any pigmentary change, while injection of adrenalin, in all cases,called forth the mottling response.
In the third section of this paper, describing the colour variations in differentregions of the body, it was stated that an animal in the dark state shows a darkquadrangular patch posterior to the orbit and a scattering of dark spots againstthe brown skin of the body. It is now evident that these darker regions constitutethe mottling pattern. (Except for being slightly larger and more densely pigmented,the melanophores in the scales of the mottling pattern do not appear much differentfrom those in non-mottling skin; see PI. I, figs. 5 and 6.) The above experimentsdemonstrated adrenal control of mottling, but since there is no reason to believethat adrenalin is being secreted when the lizard is adapting itself to a black back-ground, it follows that the pituitary hormone which causes dispersion of themelanophore pigment of the body has a similar effect on the melanophores of thescales composing the mottling pattern. This conclusion is supported by the factthat the mottling pattern is distinctly recognizable in adrenalectomized lizardswhich darken in adaptation to an illuminated black background.
VII. SUMMARY
1. Anolis becomes dark brown on a light-absorbing background and brightgreen on a light-dispersing background, the changes being accomplished by a dis-persion or concentration of pigment within melanophores.
2. Blinded lizards lose the ability to respond to changes in backgrounds, butbecome brown when in light and green when kept in darkness.
3. Hypophysectomy results in permanent pallor. The brown colour can betemporarily elicited in such lizards by injection of appropriate extracts of thepituitary gland.
4. Denervated regions of skin undergo normal colour changes.5. Electrical stimulation of hypophysectomized lizards evokes a mottled
pattern.6. Mottling is under hormonal and not under nervous regulation.
REFERENCESALLEN, B. M. (1917). Biol. Bull. Wood's Hole, 32, 117.ATWBLL, W. J. (1919). Science, 49, 48.
(1921). Endocrinology, 6, 221.BROCKE, E. (1852). Denkickr. Akad. Wiss. Wien, 4, 179.VON BUDDENBROCK, W. (1928). Grundriss der vergleichenden Pkytiologie. Berlin.CARLTON, F. C. (1903). Proc. Amer. Acad. Arts Sci. 39, 259.VON GELDERN, C. E. (1921). Proc. Calif. Acad. Sci. 10, 77.HADLEY, C. E. (1929). Bull. Mus. comp. Zool. Harv. 69, 107.

490 L. H. KLEINHOLZ
HADLEY, C. E. (1931). J. exp. Zool. 58, 3*1.HOGBEN, L. T. & MraviSH, L. (1928). J. exp. Biol. 5, 295.HOGBEN, L. T. & SLOME, D. (1931). Proe. roy. Soc. B, 108, 10.HOGBEN, L. T. & WINTON, F. R. (1922 a). Proc. roy. Soc. B, 93, 318.
(19226). Proc. roy. Soc. B, 94, 151.(1923). Proc. roy. Soc. B, 96, 15.
KELLEK, R. (1895). PflUg. Arch. ges. Phytiol. 61, 121.KLEINHOLZ, L. H. (1935). Biol. Bull. Wood's Holt, 69, 379.
(1936). Proc. Nat. Acad. Set., Wash., 22, 454.KHUKENBERG, C. F. W. (1880). Vergl. physiol. Stud. Heidelberg, 1, 23.MAY, R. M. (1924). J. exp. Biol. 1, 539.NOBLE, G. K. & BRADLEY, H. T. (1933). Biol. Bull. Wood's Hole, 64, 289.PARKER, G. H. (1906). J. exp. Zool. 3, 401.
(1936). Color Changes of Animals in Relation to Nervous Activity. Philadelphia.PARKER, G. H. & STARRATT, S. A. (1904). Proc. Amer. Acad. Arts Sci. 40, 457.REDFIELD, A. C. (1918). J. exp. Zool. 26, 275.SAND, A. (1935). Biol. Rev. 10, 361.SMITH, P. E. (1916). Anat. Rec. 11, 57.ZOOND, A. & EYRE, J. (1934). Philos. Trans. B, 223, 27.
EXPLANATION OF PLATES I-IV
PLATE I
Vertical sections of scales from lizards in their colour phases. Sections stained with Mallory's azan.
Fig. 1. From a lizard in the brown state. The stratum corneum has been displaced in sectioning;the branches of the melanophores extend to the stratum germinativum.Fig." 2. In the brown phase the terminal processes of the melanophores form a thin line underneaththe stratum germinativum. The lamellae of the leucophore layer are visible.Fig. 3. In the green condition the pigment of the melanophores has become concentrated in thecell bodies.Fig. 4. The distal processes of the melanophores, even though devoid of pigment, can be seenextending toward the epidermis.Fig. 5. From the post-orbital region of a lizard in the mottled state.Fig. 6. A post-orbital scale in the green phase.
PLATE II
Colour changes of transplanted skin. All figures are of the same lizard (serial number 9, Table IV B),photographed within the first 48 hr. after transplantation.Fig. 1. Twelve hours after the operation, the animal is brown and the graft is green.Fig. 2. About 10 hr. later, after having been kept on an illuminated black background; host »kinand graft are brown.Fig. 3. The lizard was in the dark phase when placed on a white background; 8 min. later the lizardbecame green but the graft remained brown.Fig. 4. Still being kept on the illuminated white background, the lizard became uniformly greenless than an hour later.
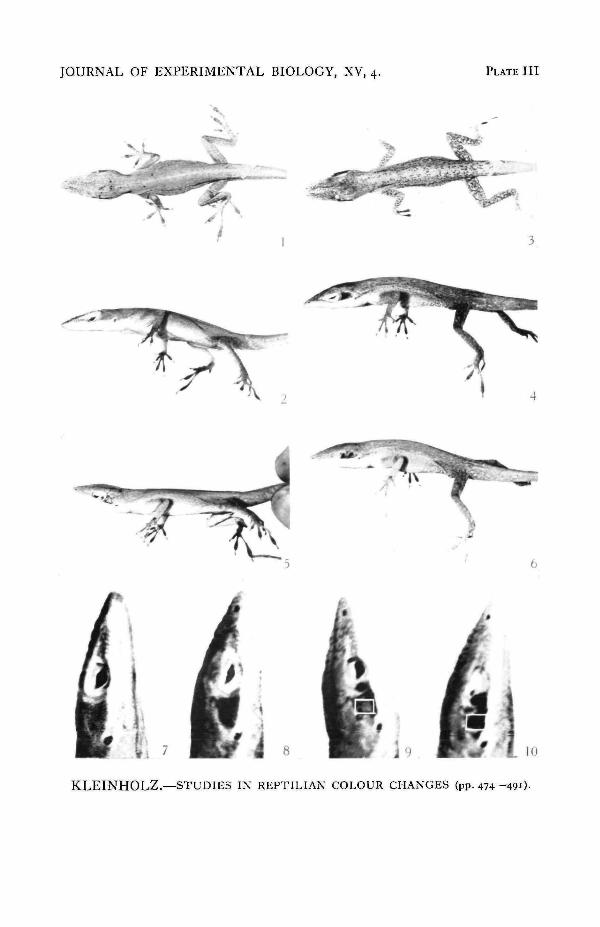
PLATE III
The mottling pattern. All figures are photographs of the same animal.Fig. 1. Dorsal view of an hypophysectomized lizard.Fig. 2. Lateral view.Fig. 3. Dorsal view of the lizard, photographed immediately after electrical stimulation for 2 min.The mottling pattern has appeared.Fig. 4. Lateral view showing the intense darkening of the post-orbital region.
JOURNAL OF EXPERIMENTAL BIOLOGY, XV, 4. PLATE I
KLEINHOLZ.—STUDIES IN REPTILIAN COLOUR CHANGES (pp. 474—491).


JOURNAL OF EXPERIMENTAL BIOLOGY, XV, 4. PLATE II
KLEINHOLZ.—STUDIES IN REPTILIAN COLOUR CHANGES (pp. 474—49!).

JOURNAL OF EXPERIMENTAL BIOLOGY, XV, 4. PLATE III
]9 10
KLEINHOLZ.—STUDIES IX REPTILIAN COLOUR CHANGES (pp. 474—49*1-

JOURNAL OF EXPERIMENTAL BIOLOGY, XV, 4. PLATE IV
#*'.
\
KLEINHOLZ.—STUDIES IN REPTILIAN COLOUR CHANGES (pp. 474—491).


Studies in Reptilian Colour Ctianges 491Fig. 5. Lateral view of the lizard, 3 days after the skin of the post-orbital region had been denervated.The transplant and the host skin are green.Fig. 6. Lateral view of the animal after 2 min. of electrical stimulation. The mottling pattern hasappeared and the denervated post-orbital patch is black.Fig. 7. The post-orbital region of the animal in fig. 2.Fig. 8. The darkened region after stimulation.Fig. 9. The denervated post-orbital patch in the green condition.Fig. 10. The same denervated post-orbital patch after electrical stimulation.
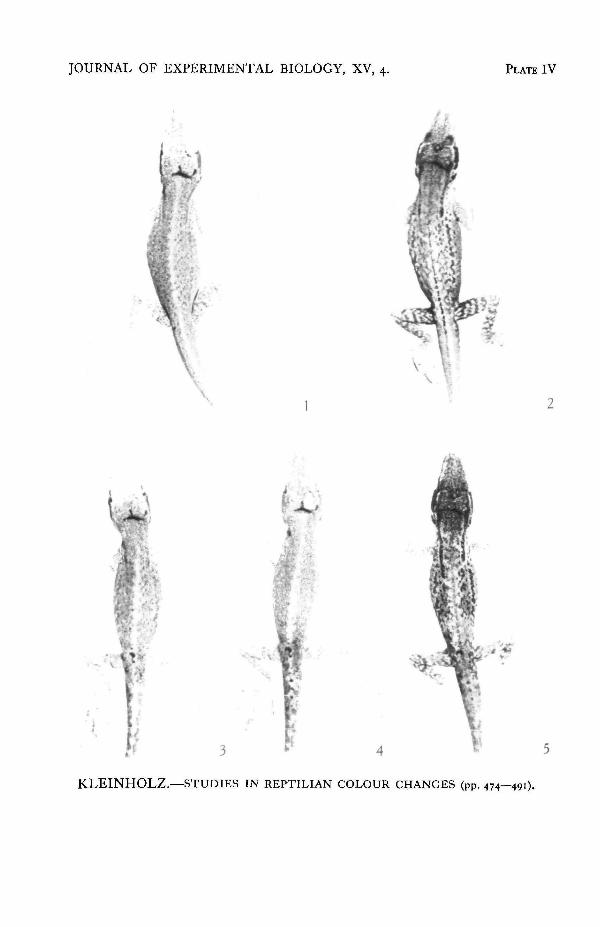
PLATE IV
Adrenal control of the mottling response. All figures are photographs of the same individual.Fig. 1. Hypophysectomized lizard in the green state.Fig. 2. The mottling response to electrical stimulation.Fig. 3. Photographed 3J hr. after the removal of both adrenals. The tail and pelvic region show aslight darkening, probably because of ligation and injury in that region.Fig. 4. The hvpophysectomized and adrenalectomized lizard after standard electrical stimulation.The mottling pattern has failed to appear.Fig. 5. Four minutes after the injection of 0-05 c.c. of 1:40,000 adrenalin. The mottling pattern hasappeared.





























