Embed Size (px)
Citation preview

i
STUDIES ON THERAPEUTIC POTENTIAL OF
MEDICINAL PLANTS AGAINST HEPATITIS C VIRUS
TARIQ JAVED
NATIONAL CENTRE OF EXCELLENCE IN MOLECULAR BIOLOGY
UNIVERSITY OF THE PUNJAB
LAHORE, PAKISTAN
(2014)

ii
Studies on Theraputic Potential of Medicinal Plants
against Hepatitis C Virus
A DISSERTATION
Submitted to
UNIVERSITY OF THE PUNJAB
In fulfillment
of the requirement for the degree
DOCTOR OF PHILOSOPHY
IN
MOLECULAR BIOLOGY
BY
TARIQ JAVED
Supervisor:
Dr. Tayyab Husnain
Professor & Acting Director
National Centre of Excellence in Molecular Biology
University of the Punjab, Lahore Pakistan.
(2014)

iii
In The Name Of Allah, the Beneficent, Merciful.

iv
CERTIFICATE
This is to certify that the research work described in this thesis is the original
work of the author Mr Tariq Javed and has been carried out under my direct
supervision. I have personally gone through the raw data and certify the
correctness/authenticity of all results reported herein. I further certify that these data
have not been used in part or full, manuscript already submitted or in the process of
submission in partial/complete fulfillment of the award of any other degree from any
other institution at home or abroad. It is also certified that the enclosed manuscript,
has been prepared under my supervision according to the prescribed format and I
endorse its evaluation for the award of PhD degree through the official procedures of
the University.
In accordance with the rules of the Centre, data book No. M-126, 904 and 985
are declared as unexpandable document that will be kept in the registry of the Centre
for a minimum of three years from the date of the thesis defense examination.
Signature of the Supervisor: __________________
Name: Dr. TAYYAB HUSNAIN
Designation: Professor & Acting Director

v
DEDICATION
“I dedicate my dissertation work to whom, who taught me even a single word, being my
teachers, I am indebted to them.
Mentor of the Universe

vi
SUMMARY
Medicinal plants are the natural reservoir of many antiviral, antimicrobial and anticancer agents.
Globally most of the population still relies on traditional medicinal plants for their primary health care.
Medicinal plants are considered to be less toxic, more effective and economical. Pakistan has a diverse flora
which potentially offers many unique phytochemicals against number of human diseases. Although there is
need for scientific analysis and research to be conducted on these medicinal plants used by the indigenous
people from centuries. Based on traditional knowledge, present study has identified some of the native
medicinal plants which possess activity against hepatitis C virus.
Hepatitis C virus infection is a serious health problem which causes liver damage, hepatocellular
carcinoma and ultimately leads to death. So far, it has affected more than 170 million individual worldwide and
10 million people in Pakistan are living with Hepatitis C virus. HCV genotype identification is most important
for prediction of treatment response and to determine the duration of antiviral therapy. The present HCV
regimen is administration of peglated interferon (PEG-IFN) and ribavirin, has limited efficacy, severe adverse
effects, and high cost. Moreover, HCV genotype 1 and 4 are more resistant to peg-interferon and rabavirin
therapy than other genotypes.
The present study was designed to search for phytochemicals from traditional medicinal plants against
Hepatitis C Virus (HCV) and study synergistic effect of purified fractions with interferon alpha which will
provide potential for future HCV drug development. Therefore, an in-vitro bioassay was developed for studying
the activity of plant extracts by infecting HCV inoculums of genotype 1a and 3a into Huh-7 cell line to screen
out potential phytochemicals against Hepatitis C virus.

vii
Twenty four medicinal plants were collected and extracted for toxicological studies on liver (Huh-7)
and fibroblast (CHO) cells lines by trypan blue dye explosive method and methylthiazol phenyltetrazolium
bromide (MTT) cell proliferation assay. Three plant extracts designated as NJ, PN and VJ showed toxic effect
in hepatoma cells, so there were excluded for further screening against Hepatitis C virus. For further antiviral
screening, HCV infected liver cells were treated with plant extracts at non toxic doses. It was found that five out
of twentyfour medicinal plant extracts designated as SN, GA, SC, AM and FC showed antiviral effect against
HCV 1a and 3a genotype in our in vitro assay. The HCV viral titer was analyzed through Quantitative Real
Time PCR and was further screened against HCV-NS3 proteases of genotype 1a and 3a. In order to identify the
active compound, corresponding plant extracts were separated into different fractions by thin layer
chromatography (TLC) and column chromatography. Purified effective fractions were then tested to find 50%
Effective concentration (EC50), and synergistic effect if any between purified fraction and interferon alpha
against Hepatitis C virus. Three fractions designated as SN8, GA15 and SC14 were active against HCV in a
dose dependent manner, and had synergistic effect when combined with interferon. The EC50 values of SN8,
GA15 and SC14 for HCV genotype 3a were 24.94 µg/ml, 9.46 µg/ml and 31.75 µg/ml and 1a were 47.68
µg/ml, 10.13 µg/ml and 71.96 µg/ml respectively.
In these in vitro studies three active fractions were identified that showed potential against HCV.
Therefore, these finding suggest that medicinal plants contain potential antiviral agents against HCV and
combination of these antiviral agents with interferon (IFN) will be better option for future HCV therapy.

viii
Acknowledgment
Praise be to Allah the most Merciful and Beneficent. Who created everything from atom to
universe and has shown us light in the darkness. Whose perpetuate patronage is treasure of my life.
Secondly, praise worthy His last Prophet (P.B.U.H) who is the torch of knowledge for humanity. It is
he, who showed us the way to success in this life and life hereafter.
I am grateful to my kind and worthy supervisor, Professor Dr. Tayyab Husnain, Director,
National Centre of Excellence in Molecular Biology, University of the Punjab, who provided me with
all possible research facilities in this institution.
I am highly gratified to Professor Dr. Sheikh Riazuddin (Ex- Director) National Centre of
Excellence in Molecular Biology, University of the Punjab, for his encouragement and scholarly
guidance during the course of my research. My work may not have seen the light of day without the
skilled advices and encouragement of Professor Dr. Shaheen N.Khan.
I would like to thank all people who have helped and inspired me during my doctoral study.
My cordial thanks go to Dr. Usman Ali Ashfaq, at National Centre of Excellence in Molecular
Biology, for his skillful support, guidance and cooperation all the time.
I am lucky enough to have prerogative to express my deep and sincere gratitude to my
seniors Dr. Tanveer Qasir, Dr. Rana Amjad, Dr. Muhammad Sharif Masoud, Dr. Muhammad Qasim
and Dr. Tahir Sarwar for helping me to get through the difficult times, and for all the emotional
support, camaraderie, entertainment, and caring; they provided, invaluable suggestions and their
sincere attitude throughout the course of my research. For me, they have been a source of great help
and inspiration.
I owe a special dept of cordial gratitude to my chums Muhammad Sohail Anjum, Muhammad
Ali Rana, Sultan Asad, Mureed Hussain, Muhammad Afzal, and Abdul Hafeez who are kind hearted,
compassionate and always there to help me whenever I needed. They always kept my spirits high.
particularly, their support, guidance, encouragement and prayers kept me to the road of optimism
and success. I am grateful I wish to extend my thanks to members of Molecular Medicine, Functional
Genomics and Stem Cell Lab, all the Scientific, Para scientific and Administrative staff of CEMB,
those had been directly and indirectly instrumental in my research work.
Special feeling of gratitude is towards my great parents, brothers Muhammad Tahir,
Muhammad Sajid, Muhammad Rashad, Muhammad Majid, Muhammad Adil and sisters, they are
outstanding their love and assistance is an asset of my life.
Finally, I bow my head before Almighty Allah, for giving me reverence for being the part of
this, most privileged and prestigious research institute CEMB, Which opens new horizons for being
explored.
I am obliged to acknowledge the Higher Education Commission (HEC) of Pakistan for
granting me a fully funded merit scholarship to meet my finances during PhD studies.
Tariq Javed
Dated: 10 -02- 2014

ix
ABBREVIATIONS AND SYMBOLS
°C Degrees Celsius
% Percentage
ABI Applied Biosystems
bp Base pair
cDNA Complementary DNA
CHO Chineese hamster ovary cells
dH2O Distilled water
DMEM DULBECCO'S MODIFIED EAGLE'S MEDIUM
DMSO Dimethyl sulfoxide
DNA Deoxyribonucleic acid
RNA Ribonucleic acid
dNTPs Deoxyribonucleotide triphosphate
EDTA ETHYLINEDIAMINE TETRAACETIC ACID
FBS Fetal Bovine Serum
GAPDH Glyceraldehyde-3-phosphate dehydrogenase
HAV Hepatitis A virus
HBV Hepatitis B virus
HCV Hepatitis C virus

x
HEV Hepatitis E virus
HIV Human Immunodeficiency virus
Huh-7 Human Hepatoma cell line
IFN Interferon
IRES Internal Ribosomal Entry Site
Kb Kilo base
kDa Kilo Dalton
L Liter
LDL Low density lipoprotein
LDL-R Low density lipoprotein recepter
MDBK Madin-Darby bovine kidney
Mg Magnesium
MgCl2 Magnesium Chloride
min Minute
ml Milliliter
mM Millimolar
mRNA Messenger ribonucleic acid
MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide)
NANBH Non-A, non B hepatitis
NC Non-coding
NCBI National Commission on Biotechonolgy
ng Nanogram

xi
NS Non-structural
ORF Open Reading Frame
PBS Phosphate buffer Saline
PCR Polymerase Chain Reaction
RNA Ribonucleic acid
RNA Ribonucleic acid
RNAase Ribonuclease enzyme
rpm Revolution per minute
RT-PCR Revesre Transcription PCR
SDS Sodium Dodecyl Sulphate
SVR Sustained viral response
Taq Thermus aquaticus
TNF-α Tumor necrosis factor alpha
UTR Untranslated region
UV Ultraviolet
μg Microgram
μl Microlitre

xii
TABLE OF CONTENTS
Certificate IV
Summary VI-VIII
Acknowledgements IX
Abbreviations ans Symbols X-X11
List of Tables XVI
List of Figures XVII
1 INTRODUCTION 1-4
2 LITERATURE REVIEW 5-21
2.1 History of Traditional Medicinal Plants 5
2.2 Ethnobotanic Flora of Pakistan 6
2.3 Cytotoxicity of Antiviral Phytochemicals 7
2.4 Antiviral Activity of Medicinal Plants 7
2.5 Medicinal Plants against Hepatitis C Virus 8
2.6 Future of Medicinal Plants 9
2.7 Hepatitis C virus (HCV) 11
2.8 HCV Molecular Evolution 11
2.9 Genotype and Ethnic Origion 12
2.10 Genetic Organization of HCV 14
2.10.1 Structural Proteins 15

xiii
2.10.2 Nonstructural Proteins 15
2.11 Model Systems for Investigating Life Cycle of HCV 15
2.11.1 Cell Lines and Primary Cell Culture 15
2.11.2 The Replicon System 16
2.11.3 Animal Models 18
2.12 Hepatitis C Virus Drug Development 19
2.12.1 NS3 Serine Protease as a Drug Target 19
2.13 HCV NS3 Protease Inhibitors 19
2.14 Disease Management 20
3 MATERIALS AND METHODS 22–34
3.1 Medicinal Plants Collection and Solvent extraction 22
3.2 Serum Samples Collection 23
3.3 Cell Lines 25
3.4 Plasmids 25
3.5 Chemicals 25
3.6 Primers Designing 26
3.7 Trypan Blue Dye Explosive Method for Cellular Toxicity 26
3.8 MTT Cell Proliferation Assay 27
3.9 Antiviral Analysis of Compounds in Liver Cells 28
3.10 Transfection of Huh-7 cells with pCR3.1/Flag TAG/HCV Nonstructural Gene 29
3.11 Co-transfection of Huh-7 cells with pCR3.1/FlagTAG/HCV Nonstructural 29

xiv
Gene and Plant Extracts
3.12 Pharmacological Analysis of isolated Fractions 31
3.13 Antiviral Analysis of Effective Fractions Along with Interferon 31
3.14 Protein Isolation and Estimation 32
3.15 Western Blotting 32
3.16 Separation and Purification Techniques 33
3.16.1 Thin Layer Chromatography 33
3.16.2 Column Chromatography 34
3.16.3 High Pressure Liquid Chromatography (HPLC) 34
3.17 Statistical Analysis 34
4 RESULTS 35-75
4.1 Medicinal Plants Collection and Solvent extraction 35
4.2 Cytotoxicity Study of Plant Extracts 35
4.3 Cellular Toxicity Through MTT Assay 37
4.4 Antiviral Effect of Plant Extracts against HCV of Genotype 3a and 1a 40
4.5 Antiviral Effect of Solanum nigrum against HCV Genotype 3a and 1a 41
4.5.1 Antiviral effect of Solanum nigrum against HCV-NS3 Proteases of Genotype 3a and 1a 42
4.5.2 Separation & Purification of Solanum nigrum Fractions by Chromatography 45
4.5.3 Antiviral Effect of Solanum nigrum Fractions 46
4.5.4 Dose Response Assay of Active Fraction of Solanum nigrum 49

xv
4.5.5 Synergistic Effect of Solanum nigrum Active Fraction (SN8) along with interferon (IFN) 51
4.6 Antiviral Effect of Grewia asiatica against HCV Genotype 3a and 1a 52
4.6.1 Antiviral Effect of Grewia asiatica against HCV-NS3 Proteases of Genotype 3a and 1a 55
4.6.2 Separation & Purification of Grewia asiatica Fractions by Chromatography 56
4.6.3 Antiviral Effect of Grewia asiatica Fractions 57
4.6.4 Dose Response Assay of Active Fraction of Grewia asiatica 60
614.6.5 Synergistic effect of Grewia asiatica Active Fraction (GA15) with Interferon (IFN) 62
4.7 Antiviral Effect of Syzgium cumine against HCV Genotype 3a and 1a 63
4.7.1 Antiviral Effect of Syzgium cumine against HCV-NS3 Proteases of Genotype 3a and 1a 64
4.7.2 Separation & Purification of Syzgium cumine Fractions by Chromatography 67
4.7.3 Antiviral Effect of Syzgium cumine Fractions 68
4.7.4 Dose Response Assay of Active Fraction of Syzgium cumine 71
4.7.5 Synergistic Effect of Syzgium cumine Active Fraction (SC14) with Interferon (IFN) 73
4.8 HPLC Analysis of Active Fractions 74
5 DISCUSSION 76-87
7 REFERENCES 88-99
8 APPENDICES
AppendixI 100-103
Appendix II (Publications) 104

xvi
LIST OF TABLE
Table 3.1 List of selected medicinal plants used for anti HCV activity 22
Table 3.2 Patients selected for medicinal plants screening, their viral loads and infecting
Genotype of HCV 24
Table 3.3.1 Primers of HCV Non-structure (NS3) gene of genotype 3a 26
Table 3.3.2 Primers of HCV Non-structure (NS3) gene of genotype 1a 26
Table 3.3.3 Primers for Glyceraldehyde-3-Phosphate Dehydrogenase gene (GAPDH) 26

xvii
LIST OF FIGURES
Figure 2.1 Hepatitis C virus (HCV): model structure 13
Figure 2.2 The HCV genome and expressed polyprotein 14
Figure 2.3 Hepatitis C virus (HCV) genome and potential drug discovery targets 20
Figure 4.1 Toxicological analysis of Plant Extracts in Huh-7 and CHO cells 36
Figure 4.2 Toxicological study of plant extracts in Huh-7 through MTT cell proliferation assay 38
Figure 4.3 Toxicological studies of SNSM, GALM and SCLM extracts in Liver (Huh-7) cells
through MTT cell proliferation assay 39
Figure 4.4 Antiviral effect of Plant extracts against HCV 3a and 1a genotype in liver cells 40
Figure 4.5 Antiviral effect of Solanum nigrum against HCV 3a and 1a genotype in liver cells 41
Figure 4.5.1.1 Antiviral effect of Solanum nigrum against HCV NS3 gene of genotype 3a and 1a
in liver cells 43
Figure 4.5.1.2 Real Time PCR analysis of Solanum nigrum against HCV NS3 gene of genotype
3a and 1a in Huh-7 cells 43
Figure 4.5.1.3 Antiviral effect of Solanum nigrum in different solvents against HCV NS3 gene
of genotype 3a and 1a in liver cells 44
Figure 4.5.1.4 Real Time PCR analysis of Solanum nigrum in different solvents against HCV
NS3 gene of genotype 3a and 1a in liver cells 44
Figure 4.5.2.1 TLC Chromatogram of Solanum nigrum (SN) 45

xviii
Figure 4.5.3.1 Antiviral effect of purified fraction of Solanum nigrum (SN8) against HCV
3a and 1a genotype in liver cells 46
Figure 4.5.3.2 Antiviral effect of column fractions (1-13) from Solanum nigrum against HCV
NS3 gene of genotype 3a and 1a in liver cells 47
Figure 4.5.3.3 Real Time PCR analysis of Solanum nigrum active fraction (SN8) against HCV
NS3 gene of genotype 3a and 1a in liver cells 47
Figure 4.5.3.4 HCV NS3 gene inhibition by S.nigrum at protein level 48
Figure 4.5.4.1 Dose dependant inhibition of active fraction (SN8) of Solanum nigrum against
HCV genotype 3a and 1a 49
Figure 4.5.4.2 Antiviral effect of active fraction (8th
) from Solanum nigrum in different
concentrations against HCV NS3 gene of genotype 3a and 1a in liver cells 50
Figure 4.5.5 Synergy in the antiviral activity of Solanum nigrum active fraction (SN8)
with interferon 51
Figure 4.6 Antiviral effect of Grewia asiatica against HCV 3a and 1a genotype in liver cells 52
Figure 4.6.1.1 Antiviral effect of Grewia asiatica against HCV NS3 gene of genotype 3a and 1a
in liver cells 54
Figure 4.6.1.2 Real Time PCR analysis of Grewia asiatica against HCV NS3 gene of genotype 3a and 1a in
liver cells 54
Figure 4.6.1.3 Antiviral effect of Grewia asiatica in different solvents against HCV NS3 gene of genotype
3a and 1a in Liver cells 55

xix
Figure 4.6.1.4 Real Time PCR analysis of Grewia asiatica in different solvents against HCV NS3 gene of
genotype 3a and 1a in liver cells 55
Figure 4.6.2.1 TLC Chromatogram of Grewia asiatica (GA) 56
Figure 4.6.3.1 Antiviral effect of purified fraction of Grewia asiatica (GA15) against HCV 3a and 1a
genotype in liver cells 57
Figure 4.6.3.2 Antiviral effect of column fractions (1-16) from Grewia asiatica against HCV NS3 gene of
genotype 3a and 1a in liver cells 58
Figure 4.6.3.3 Real Time PCR analysis of Grewia asiatica active fraction (GA15) against HCV NS3 gene of
genotype 3a and 1a in liver cells 58
Figure 4.6.3.4 HCV NS3 gene inhibition by G. asiatica at protein level 59
Figure 4.6.4.1 Dose dependent inhibition of active fraction of Grewia asiatica (GA15) against HCV of 3a
and 1a genotype 60
Figure 4.6.4.2 Antiviral effect of active fraction (15th
) from Grewia asiatica in different concentrations
against HCV NS3 gene of genotype 3a and 1a in liver cells 61
Figure 4.6.5 Synergy in the antiviral activity of Grewia asiatica active fraction (GA15) with
interferon (IFN) 62
Figure 4.7 Antiviral effect of Syzgium cumine against HCV 3a and 1a genotype in liver cells 63
Figure 4.7.1.1 Antiviral effect of Syzgium cumine against HCV NS3 gene of genotype 3a and 1a 65
Figure 4.7.1.2 Real Time PCR analysis of Syzgium cumine against HCV NS3 gene of genotype
3a and 1a in liver cells 65
Figure 4.7.1.3 Antiviral effect of Syzgium cumine in different solvents against HCV NS3 gene of
genotype 3a and 1a in liver cells 66

xx
Figure 4.7.1.4 Real Time PCR analysis of Syzgium cumine in different solvents against HCV
NS3 gene of genotype 3a and 1a in liver cells 66
Figure 4.7.2.1 TLC Chromatogram of Syzgium cumine (SC) 67
Figure 4.7.3.1 Antiviral effect of purified fraction of Syzgium cumine against HCV 3a and 1a
genotype in liver cells 68
Figure 4.7.3.2 Antiviral effect of column fractions (1-15) from Syzgium cumine against HCV
NS3 gene of genotype 3a and 1a in liver cells 69
Figure 4.7.3.3 Real Time PCR analysis of Syzgium cumine active fraction (SC14) against HCV
NS3 gene of genotype 3a and 1a in liver cells 69
Figure 4.7.3.4 HCV NS3 gene inhibition by S. cumine at protein level 70
Figure 4.7.4.1 Dose dependent inhibition of Active fraction of Syzgium cumine against HCV
of 3a and 1a genotype 71
Figure 4.7.4.2 Antiviral effect of active column fraction (14th
) from Syzgium cumine in different
concentrations against HCV NS3 gene of genotype 3a and 1a in liver cells 72
Figure 4.7.3 Synergy in the activity of Syzgium cumine active fraction (SC14) with
interferon (IFN) 73
Figure 4.8 The HPLC Chromatogram of purified fraction (A) SN8 (B) GA16 and (C) SC14 75

1
INTRODUCTION Traditional use of medicinal plants implies in ethnobotanical literature. In developing
countries, the majority of the people rely on medicinal plants for health care. South East
Asian countries possess a rich, diverse and unique flora. Therefore, people in these countries
depend upon traditional medicinal plants against variety of ailments. According to an
estimate less than 10% of ethnic medicinal plants have been scientifically analyzed for any
bioactivity (Mohanty and Cock, 2012). Plant extracts, infusions and powders have been used
for treatment of numerous ailments and have shown potential against different human
diseases (Vijayan et al., 2004). About 25% of the frequently used medicines are derived
from plant source (Mukhtar et al., 2008) which includes aspirin isolated from Salix alba, and
quinine extracted from bark of cinchona tree (Newman et al., 2000).
Phytochemicals like tannins, flavonoides, glycosides and alkaloids present in these
plants are found to be active against herpes simplex virus (Cavallaro et al., 1995), influenza
virus, hepatitis B and C viruses (Hudson, 1989; Kitazato et al., 2007). These phytochemicals
either inhibit the viral DNA or RNA formation or hinder the viral reproduction (Hudson et
al., 1991). However, few of the endemic herbs are used for the treatment of various viral
diseases including Hepatitis C virus.
Several antiviral drugs have severe side effects which discontinue the standard
regimen therapy of interferon and ribavirin. So, there is an ever increasing need for the
search of phytochemicals possessing antiviral potential (De Clercq, 2002). Since, herbal
medication is perceived to be less toxic, and more efficacious (Pak et al., 2004) which makes
them more advantageous for the population of low income countries.

2
Hepatitis is the foremost issue to community health worldwide which may further
develop liver fibrosis, cirrhosis, hepatocellular carcinoma. According to an estimate about
170 million people worldwide (Ge et al., 2009) and approximately 10% of Pakistani
population is living with HCV (Idrees et al., 2008). There are seven major genotypes of
HCV (Nakano, T., 2012) and more than 80 subtypes (Kuiken et al., 2005). Some of them are
distributed worldwide, while others confined geographically. More than 90% of the
infections in America, Europe, Russia, Central Asia are caused by HCV genotype 1a, 1b, 2a,
2b, 2c and 3a (Simmonds, 2004). Genotype 3 and its other subtypes are highly prevalent in
South Asian countries. In Pakistan, the subtype 3a is the most frequent HCV genotype
(49.05%) and other genotypes have different distribution (Idrees et al., 2008) whereas
genotype 4, 5 and 6 are rare ones (Attaullah et al., 2011).
However, the preventive measures are limited and about 50% patients are unable to
show response for current therapies including peglated Interferon and Ribavirin (Bacon et al.,
2009). Interferon monotherapy or combination of interferon with ribavirin both are
successful treatment in some of the patients (Moradpour et al., 2005), while the modified
forms of Interferon including peglated Interferon-α have sustained virologic response (SVR)
of 41% (Manns et al., 2001). Similarly, the treatment with combination therapy of IFN α and
ribavirin in patients with chronic hepatitis C has sustained virologic response of about 10%
(Bacon et al., 2009).
The side effects of Interferon and Peglated Interferon treatment include depression,
fatigue, headache, myalgia and thrombocytopenia. Meanwhile, as a result of these adverse
effects 10% of HCV patients discontinue this therapy. HCV genotype 1 patients are the most

3
complicated to treat and their SVR rate is approximately 40% after 48 weeks of therapy
(McHutchison et al., 2009).
To reduce the HCV viral load, a number of recent therapies are at different stages of
clinical trials (Zeuzem et al., 2011). These drugs include direct acting antiviral (DAA)
agents, which acts on the target during the viral life cycle. Recently, NS3/4A inhibitors are
the most widely studied and are successful direct acting antiviral therapy (Asselah and
Marcellin, 2011). The ultimate target of HCV eradication is to develop a shorter, more
efficient treatment schedule comprising of oral dosage that have a few side effect profile than
peglated Interferon α and ribavirin. Preliminary studies in this area are focusing on the
combination therapy of a protease and polymerase inhibitors (Jazwinski and Muir, 2011).
The triple combination of peglated Interferon, ribavirin plus a protease inhibitor has
increased sustained virologic response to about 60% in HCV genotype 1 patients (Asselah et
al., 2009; Asselah and Marcellin, 2011).
This study was an attempt to find out medicinal plants with potential antiviral agents.
Therefore, the twenty four medicinal plants were selected and their methanolic extracts were
concentrated and dried. In vitro toxicity of extracts was checked in Huh-7 and CHO cell lines
at 100 µg/µl concentration. Subsequently, blood serum from HCV positive patients of
genotype 3a and 1a were collected and pooled to evaluate the antiviral effect of extracts in
liver cell line and level of HCV-RNA was detected by Quantitative Real Time PCR. It was
found that five out of twenty four medicinal plants designated as SN, GA, SC, AM and FC
showed antiviral effect against HCV genotypes 3a and 1a in in vitro assay. Plant extracts
with antiviral potential were further fractionated by various chromatography techniques and

4
purified fractions were tested for antiviral activity against Hepatitis C virus, alone and in
combination with Interferon.
Present study shows that Pakistan‘s diverse flora has many potent antiviral plants.
Moreover, after determination of their chemical species responsible for anti HCV activity
and in combination therapy with interferon will help to develop future HCV therapies.
Therefore, more research is required on medicinal plants which are used by indigenous
people for treatment of Hepatitis C.

5
LITERATURE REVIEW
2.1 History of Traditional Medicinal Plants
The history of medicine dates back to the origin of human race. The use of plants as a
remedy of different diseases has been inherited and is an important element of health care
scheme worldwide. From last two decades use of traditional medicinal plants against various
ailments has increased. These medicinal plants are considered too rich in photochemicals of
interest for drug development (Calland et al., 2012). Traditional medical practitioners used to
hide the formulation and identity of plants used for the cure and healing of different diseases.
The reason behind was that patients should not learn to treat themselves.
The use of folk medicines throughout the world commonly depends upon local flora
and traditional experiences. For example, most of the Chinese people even now use Chinese
herbs from ancient times. In China, almost 5000 medications are in use by traditional healers
and these medicines account for roughly one fifth of the whole Chinese pharmaceutical
market (Murphy et al., 2002). Likewise Ayurveda includes herbal remedies is a traditional
Indian medical system for disease prevention and treatment (Morgan, 1994).
To avoid the adverse effects of synthetic medication, most of the people rely on
traditional medicinal plants. The first effort for the development of an antiviral agent was
performed by Boots a Drug Company of England for developing drug against influenza
virus and they screened more than 280 plants (Mukhtar et al., 2008). In the latter part of the
20th
century plant based medicine or ‗Alternative Medicine‖ became very common in USA
and western countries because of its natural origin. Traditional medicine use centuries old
plant formulations while modern plant based medicine use extraction of active
phytochemicals provides as raw material for synthesis of different drugs.

6
2.2 Ethnobotanic Flora of Pakistan
In countries like Japan, India, China, Pakistan, Srilanka, Thailand and neumerous
African countries large number of people depend upon herbal drugs to cure different diseases
(Hoareau and DaSilva, 1999). Pakistan possesses a very rich and unique flora with variety of
medicinal plants due to topography and distinct four seasons. About 6,000 species of
flowering plants have been reported from Pakistan and Kashmir and nearly 372 plant species
are endemic (Qureshi et al., 2009). In Pakistan about 75% of the population resides in
villages and remote areas with lack of health facilities. Most of these poor and ignorant
people have no option except to practice traditional medicines (Azaizeh et al., 2003). A vast
knowledge about use of local medicinal plants is expected to be present in such areas (Diallo
et al., 1999).
For the treatment of various diseases, traditional practitioners (Hakeems) utilize
remedies based on different parts of the plant for medicinal purposes. For instance, for the
cure of wounds the powdered leaves and bark of Caryopteris odorata is used. Likewise, the
bark and leaves of Daphne papyracea used as poultice for tumors and whole plant of
Cichorium intybus is used for jaundice and hepatitis while aerial parts of this plant are used
for asthma, typhoid, and ulcer (Abbasi et al., 2009; Haq and Rehman, 1990; Zafar and Ali,
1998). So, there is a need of scientific research to identify potential phytochemicals from
these traditional medicinal plants (Nisar et al., 2011).

7
2.3 Cytotoxicity of Antiviral Phytochemicals
The potential phytochemicals or bioactive compounds have to be confirmed as non
toxic to life for the production of new antiviral drug. For this reason, different cytotoxic
assays are performed by utilizing cell culture system. Cells may behave differently as a
result of phytochemical treatment. While evaluation, cells may found as lysed, reduced
growth or even apoptosis (programmed cell death). Some of the integral cytotoxic assays
includes, cell viability and proliferation assay which are based on evaluation of different
parameters.
Moreover, colorimeteric and luminescence based assay use microliter plate reader or
ELISA plate reader to measure the toxicity of bioactive compounds. To determine membrane
integrity, vital dyes assay can be performed such as trypan blue, intracellular components of
cell dyed with trypan blue dye, if cell membrane is ruptured (Jauregui et al., 1981). MTT is
another colorimetric assay for the measurement of reducing potential of metabolically active
cells. Tetrazolium salt (3-[4, 5-dimethylthiazol-2-y1]-2, 5- diphenyltetrazolium bromide)
(MTT) will reduce to blue colored formazan by viable cells (Dhawan, 2012).
2.4 Antiviral Activity of Medicinal Plants
World Health Organization (WHO) estimates that globally about 80% the people
fulfills their health care issues by utilizing phytochemicals (Grossmann et al., 2010;
Nascimento et al., 2000). Antiviral potential of several phytochemicals has been reported by
several research groups. Taking into account the vast number of plants and diversity of their

8
chemical constituents, there is a need to screen plants for their antiviral domain. Therefore,
utilizing these potent phytochemicals can be the better option for the treatment of viral
diseases in future.
Studies have also suggested the antiviral effect of medicinal herbs extract on various
viruses which includes herpes simplex virus type 2 (HSV-2), (Vermani and Garg, 2002) HIV
and Hepatitis B Virus (HBV) (Kapusta et al., 1999). Syzygium aromaticum (clove) is used in
traditional medical practice for its main compound, eugenol has many therapeutic benefits
including antiseptic, antibacterial, analgesic, antifungal, anticancer, antioxidant, anti-
inflammatory (Hussain et al., 2009) antimutagenic and as pesticidal agent against several
pests. The essential oil of S. aromaticum has number of antimicrobial agents for aquaculture
(Kumar et al.). Recently, Syzygium aromaticum has shown to possess antiviral potential, in
combination with acyclovir against HSV-1. Furthermore, it also limited the replication of
cytomegalovirus (CMV). Another cosmopolitan medicinal herb Solanum nigrum is
beneficial for the treatment of ulcers, nervous system disorders, liver disorders (Khattak et
al.; Saleem et al., 2010).
2.5 Medicinal Plants against Hepatitis C Virus
Recently, natural products are enormously employed for anti HCV activity. Studies
and clinical trials have shown that Glycyrrhiza uralensis (licorice root), glycyrrhizin sulphate
is involved in inhibition of HIV replication and induces IFN activity (Jatav et al., 2011).
Silybum marianum (milk thistle) possesses antioxidant, anti inflammatory,
immunomodulating and liver regeneration capacity with therapeutic effects against fatty
liver, cirrhosis and viral hepatitis (Ashfaq et al., 2011). Viscum album extract stabilizes liver

9
function and cyclosporine-A has shown effect in limiting the HCV RNA below the detection
level (Li et al., 2012; Wang et al., 2013).
Beneficial extracts obtained from plants such as Piper cubeba, Trachyspermum ammi,
Embelia schimperi, Boswellia carterii, Quercus infectoria, and Syzygium aromaticum were
examined for in vitro antiviral activity against HCV proteases (Kitazato et al., 2007).
Therapeutic phytochemicals includes terpenoids, triterpenoids, fatty component, thiophenes,
flavonoids and steroids which exhibited promising antiviral effect against different viral
infections. There is enormous potential of useful phytochemicals to expose, evaluate and
exploit for therapies against diverse viral family like HCV (Jassim and Naji, 2003). The
different species of genus Grewia are used for their medicinal importance throughout the
world. The roots of G.abutilifolia are helpful against abscesses, and G. asiatica leaves are
utilized in pustular eruptions (Ahaskar et al., 2007; Zia-Ul-Haq et al., 2012).
Hepatoprotective effect of Ayurvedic herb for antituberculosis treatment has been
studied to assess the hepatopprotective effect of some Ayurvedic herbs. Hydroalcoholic root
extract of Berberis aristata (Daruha ridra), Aloe vera (Ghritakumari) ariel parts and Solanum
nigrum (Kakmachi) whole plant, herb Phyllanthus fraternus (Bhumayamalaki) exhibited
hepatoprotective efficiency (Sharma et al., 2004).
2.6 Future of Medicinal Plants
Medicinal plants have a promising future, as more than 500,000 uninvestigated
medicinal plants throughout the world (Hassan, 2012). These potential therapeutic plants
needs to be explored, some of them possess even more potential than expected. On the other

10
hand, many food contents contain non-nutritive phytochemicals such as flavonoids, phenolic
acids and carotenoids, which are considered to provide protective effect against chronic
diseases (Boyer and Liu, 2004). These antioxidants reduce the threat of free radicals and
improve the immune response. According to research of Block and coworkers, people who
eat more fruits and vegetables have limited risks of cancers (Block et al., 1992).
So far, the research on medicinal plant based drugs has shown some exceptional
characteristics when used for treatment. They can act as synergic medicine to interact
simultaneously, thus their possible adverse effects are neutralized. Some plant base drugs can
be used for the support of approved medicine during the treatment of complex diseases like
cancers, so play an effective role and even reduces the side effect of synthetic medicines
(Hassan, 2012). Another important character of medicinal plants is their behavior as
preventive medicine, possess ability to prevent diseases; despite of synthetic medicines
which can only be used when the disease occurs.
Therefore, a systematic scientific study is required to identify and isolate bioactive
compounds from traditional medicinal plants. This should include cytotoxic effect followed
by invitro and invivo animal models and finally clinical trials. After passing through this
route promising novel bioactive compounds can be optimized and new medicinal plants
derived drugs can be introduced in the market for the improvement of individual‘s health all
over the world.

11
2.7 Hepatitis C Virus (HCV)
Hepatitis C virus (HCV) is a foremost health dilemma globally. World Health
Organization (WHO) appraised about 170 million infected people though out the world with
Hepatitis C Virus (Ghany et al., 2011). HCV virus is an envelope RNA virus, first identified
by Choo et al in 1989. Formerly, HCV was named as ―non A, non B (NANB) Hepatitis‖. It
is among seven different hepatotrapic viruses identified today, the other viruses include
Hepatitis A, B, D, E and G Virus (Bostan and Mahmood, 2010) (Lanford et al., 1994).
HCV was identified and characterized by molecular cloning techniques using serum
from a NANB hepatitis virus from infected chimpanzee and based on the similarity of the
genomic organization and hydropathy profiles of several precursor proteins. It is classified as
a member of Flaviviridae family (Collett et al., 1988) which also includes Dengue virus.
Hepatitis C Virus encodes for single polyprotein of about 3010 amino acid. However, the low
sequence homology compared to other flaviviruses ultimately lead to its classification into a
hepacivirus which is different from other flavivirus members (Bollati et al., 2010).
2.8 HCV Molecular Evolution
To estimate the origin of HCV, when it was introduced into human population,
remains hard to know because its inability to identify HCV or HCV like variants in Ape
species (Simmonds, 2004). However, in theory, it may be possible to calculate the
divergence time of the main clades and splitting of subtypes by using the constant nucleotide
substitution rate over time. The rate of HCV sequence change in whole genome is 1.44 x 10-3
nucleotide changeover per site per year. An evolutionary rate of 7.4 x 10 -4
nucleotide
replacement per site per year for E1 gene and 4.1x 10-4
or the NS5B gene was calculated by

12
Smith in 1997. The subtypes deviate around 300 years ago, and the variance of different
genotypes occurred around 500-2000 years ago (Smith et al., 1997).
2.9 Genotype and Ethnic Origin
Identification of Hepatitis C virus genotype is critically important and responsible for
the response and time period required for treatment (Noppornpanth et al., 2006). According
to facts, genotype 1 and 4 are more resistant to peg-IFN and ribavirin, standard therapy than
genotype 2 and 3. Furthermore, severe liver disease is reported in case of patients with
chronic HCV genotype 1b. (Trinks et al., 2012). In epidemiological studies HCV genotyping
is an easy method utilize genotype specific HCV antibodies.
Presently, three broad patterns of Hepatitis C virus genotype distribution exists (Fretz
et al., 1995). Genetically most diverse genotype pattern are 1 and 2 prevalent in West
African regions (Ndjomou et al., 2003). On the other hand, in developing countries and under
developed counties, there is lack of authentic data about the disease burden. One fifth of the
world‘s population resides in China, where HCV seroprevalence is about 4.9%. In South
Asian countries including India, Indonesia and Pakistan, seroprevalence ranges from 0.9 % to
6.5%. The most prevalent HCV genotype in Pakistan is genotype 3, which is also most
frequent genotype in northern and northeastern, and central India, with high infection rates of
more than 70% acute and more than 80% chronic hepatitis patients (Chaudhuri et al., 2005;
Hissar et al., 2006).
In China and Japan most HCV infections are with genotype 1b, whereas genotype 4
is frequently found in Middle East and North Africa and genotype 5a in South Africa, while
genotype 6 is frequent in Hong Kong (Dusheiko et al., 1994; Simmonds, 2004). Asian and

13
African countries are reported to have highest prevalence of HCV as compared to rest of the
world. Moreover, the rate of seroprevalence is 20% in Central Africa and Egypt, 4% in Asian
and Mediterranean countries (ANNEMARIE WASLEY and Alter, 2000).
On the contrary, the developed nations of North America, Western Europe and
Australia have low HCV seroprevalence rate i.e. in Germany 0.6%, Canada 0.08%, France
and Australia have 1.1% (Shepard et al., 2005). Slightly higher seroprevalence is reported in
some other developed countries including USA 1.8%, Japan 1.5-2.3% and Italy 2.2%
(Shepard et al., 2005).
Figure 2.1: Hepatitis C virus (HCV): Model structure Image taken from Louis E. Henderson (Frederick
Cancer Research Center)

14
Figure 2.2: The HCV genome and expressed polyprotein: Translation depends on an internal ribosome entry
site (IRES) within the 5‘ non translated region (NTR). The polyprotein precursor is posttranslationally
processed by host and viral proteases and the HCV structural and nonstructural proteins are localized within
the endoplasmic reticulum membrane.
2.10 Genetic Organization of HCV
HCV genome is comprised of open reading frame (ORF) and at 5‘ and 3‘ ends very
stable non translated regions (NTR) which further forms secondary and tertiary structures.
During translation, the polyprotein is processed from 5‘ to 3‘ reigon to produce core (C),
envelope E1, E2, p7, NS2, NS3, NS4A, NS5A and NS5S (Bartenschlager and Lohmann,
2000). All the substructures have different tasks in HCV life cycle.

15
2.10.1 Structural Proteins
The structural protein which form viral particles, have following order; core,
enveloped E1, E2 and p7. According to different studies, core protein affects the cellular
functions of host including gene transcription, apoptosis and signaling pathways
(Tellinghuisen and Rice, 2002). The envelope proteins are supposed to involve in cell entry
(Bartosch et al., 2003a). P7 has vital role in assembly and release of virus particles
depending upon the viral genotype (Steinmann et al., 2007).
2.10.2 Nonstructural Proteins
The second part of HCV genome encodes numerous NS proteins such as: NS2, NS3,
NS4A, NS4B and NS5A. The function of NS2 protein is not elucidated so far (Duvet et al.,
1998) whereas, NS3 serine protease influences the cellular host defense has emerged as
antiviral target (Foy et al., 2005; Meylan et al., 2005). For the development of anti HCV oral
drugs, NS3-4A serine protease and central component of HCV replicase NS5B are
considered to be most attractive targets (De Francesco and Migliaccio, 2005; Kolykhalov et
al., 1997).
2.11 Model Systems for Investigating Life Cycle of HCV
2.11.1 Cell Lines and Primary Cell Culture
Since HCV was identified in 1989, serological and epidemiological studies seemed to
be unsusceptible to HCV infection. The major limitation for hepatitis C virus research was
the lack of cell culture system. Moreover, Flaviviruses including HCV like other positive

16
strand viruses replicate by means of a negative strand RNA intermediate. Therefore, in in
vitro studies to detect negative sense molecule strand specific RT-PCR is used (Yan et al.,
2000).
Significant increase in HCV positive strand RNA has also been reported within the
first four days of infection in chimpanze by using strand specific RT-PCR. On contrary, the
negative strand RNA signal become detectable on day four and increases afterwards.
Furthermore, Lanford and colleagues noted that from baboons, primary liver cells isolated
are not vulnerable to infection sustaining the idea that HCV has only limited hosts. Several
studies on HepG2 and Huh-7 liver cell lines remained unsuccessful although there were
comprehensive variations in experimental conditions (Lanford et al., 1994). A large number
of potent targets for direct acting antiviral (DAA) agents has been identified through
replicative cell culture system and resolution of three dimentional structures of HCV protein
(Sarrazin and Zeuzem, 2010).
2.11.2 The Replicon System
However, the primary cell culture provided some insight into the basic principles of
HCV infection. The heterogeneity of the inoculums and the low RNA replication rate has
made it complicated to analyze all the features of viral life cycle. Later on, neomycin
selectable HCV mini genome (replicon) based on the Con1 consensus genome cloned from
liver derived viral RNA was created (Krieger et al., 2001). At first, a full-length genome was
used to transfect various cell lines and primary human hepatocytes.

17
Therefore, a full length genome was used to transfect various cell lines. The full
length RNA failed to replicate to generate a 16 number of bicistronic construct, with 5‘HCV
internal ribosomal entry sites (IRES) neomycin phosphotransferase gene, the genotype 1b
non-structural genes NS2 or NS3 to NS5B under the control of encephalomyocarditis virus
(EMCV) the HCV 3‘ nontranslated region (NTR). Distinctive to the full length counterparts
the subgenomic replicons replicated to a high level, was credited to cell culture based
mutations in the NS3, NS5A and NS5B region of HCV genome (Blight et al., 2003; Krieger
et al., 2001).
On these findings, several groups successfully generated full length replicons with
single amino acid substitutions in the genes encoding the non structural viral proteins (Blight
et al., 2000; Pietschmann et al., 2001). As expected these cell culture adaptive mutations
clearly improved RNA levels as well as the incidence of cells supporting replication.
Nevertheless, Huh-7 cells harbouring full length replicons of the prototypic viral strains
Con1 and HCV-H, still failed to produce infectious particles although the Con1 strain was
infectious in vivo (Bukh et al., 2002). Furthermore, the number of HCV-RNA replication
competent cells within the total population remained low even for adapted replicons,
suggesting that the cellular background was major determinant of replication efficiency
(Blight et al., 2003).
To enhance permissiveness of the Huh-7 cell line, cells were transfected with
subgenomic replicons with either the wild type amino acid sequence a serine to leucin
substitution (S2204I) in the NS5A region, or a 47 amino acid NS5A deletion (5AD47). Cells
supporting viral replication were chosen and cured of HCV-RNA by extended treatment with
interferon (IFN) α. The resulting clonal cell lines were then tested for their ability to support

18
HCV replication following transfection with subgenomic and full length replicons (Wakita et
al., 2005). However, mutation in RIG-I eliminates pathogen associated molecular patterns
(PAMP) signalling to IRF3, thus inhibiting the cellular antiviral response and presenting an
increased permissiveness for HCV RNA replication in Huh-7.5 cells (Sumpter et al., 2005).
The replicon system provides a important tool to study HCV replication. However, it
does not allow studies of virus attachment and entry (Baumert et al., 1998). Development of
infectious HCV pseudoparticles (HCVpp) by several research groups expressing the E1/E2
structural proteins in 293T cells with a packaging construct encoding the HIV genome minus
the envelop gene, and the gag and pol genes of murine leukaemia virus (MLV) (Bartosch et
al., 2003b). Huh-7 cells can be infected by; 293T cells secreted virus pseudoparticle which
can further be evaluated by luciferase or GFP assays (Cai et al., 2005; Hsu et al., 2003). The
Coexpression of these constructs led to the assembly of infectious replication deficient HCV
pseudoparticles, which makes it possible to study details of virus attachment (Yi et al., 2006).
2.11.3 Animal Models
The chimpanzees were the only animal model available since last two decades for the
study of HCV infection. Similar to humans, after few days of infection, chimpanzees also
have 105 and 10
7 RNA genome copies/ml in serum and raised aminotransferase (ALT) levels
(Tan, 2006).
Finally, the chimpanzee model was considered to be successful model system for the
establishment of molecular clones of HCV (Kolykhalov et al., 2000). Human beings are
mostly asymptomatic, which makes it difficult to study the acute phase of infection.
Researchers have reported the creation of chimeric (xenograft) mice harbouring human

19
hepatocytes (Mercer et al., 2001) are immunodeficient and suffer from severe, chronic liver
disease caused by over expression of the noxious protein urokinase. Chimeric mice, such as
the SCID/uPA mouse, are successfully infected with HCV derived from 21 human sera and
have shown to support viral replication at relevant titers (Meuleman et al., 2005).
2.12 Hepatitis C Virus Drug Development
2.12.1 NS3 Serine Protease as a Drug Target
So for the protease inhibitors are considered to be promising target for treatment of
severe viral diseases including HIV. Viral RNA replication can be inhibit by the NS3 protein
(Locarnini and Bartholomeusz, 2002). An additional task is to develop a molecule that mimic
the natural peptide ligand (peptidomimetic), where the cleavable amide bond is substituted
with a non cleavable isostere (Leung et al., 2000).
2.13 HCV NS3 Protease
NS3 protease is a small protein which belongs to sub-class of small chymotrypsin like
protease (Bazan and Fletterick, 1988). There are several compounds that influence the
activity of the NS3 protease and the drugs that inhibit NS5B polymerase are presently in
clinical trials, and will likely to become the next generation of anti HCV drugs. The segment
of HCV which acts as a helicase has attracted the attention of researchers interested in
developing novel antiviral drugs and interaction of proteins with nucleic acid.

20
Figure 2.3: Hepatitis C virus (HCV) genome and potent drug discovery targets (Asselah and Marcellin,
2011)
2.14 Disease Management
Timely clearance of HCV to prevent the risk of hepatocellular carcinoma (HCC) and
to decrease mortality is the aim of antiviral remedy in patients with HCV infection. In past,
several studies have reported that using 3 million International units (MIU) of IFN, thrice a
week for 6 to 24 weeks or similar dose of IFN intravenously for 4 to 7 weeks are beneficial
in clearing virus and normalizing ALT levels. Use of IFN (3 MIU, 3 times in a week)
intramuscularly for 4 weeks has shown similar results (Dhiman and Chawla, 2005).
Since 2001, recombinant interferon is replaced by pegylated interferon α (Peg-IFN α)
with Ribavirin (RBV) and more sustained virologic response has been observed in genotype
1 patients of chronic Hepatitis C (Ghany et al., 2011). The Peg-IFN and RBV therapy has
shown an efficacy limit to 30% with severe side effect. This combination therapy shown

21
better results in patients with HCV genotype 2 and 3 although only small numbers of patients
have shown complete clearance of virus by this method (Sánchez–Tapias et al., 2006).
Furthermore, the disease management in patients with HCV genotype 1 has low response to
combination therapy of pegylated interferon and rebavirin. (Biselli et al., 2006).
Recently, number of new HCV inhibitors has reached clinical trials at different
stages. However, only few NS3/4A protease inhibitors have studied on patients with HCV
genotype other than 1. Similarly, telaprevir has shown response against genotype 3 and a
limited effect on genotype 4, boceprevir has shown modest antiviral effect is shown in
genotype 2 and 3 (Tong et al., 2012). Adverse effects of telaprevir includes anemia, rash,
pruritus and these effects are sometimes more severe with limited treatment options
(Cunningham and Foster, 2012). This emphasizes the need for more efficient and less toxic
antiviral therapy against Hepatitis C Infection.
22
MATERIALS AND METHODS
3.1 Medicinal Plants Collection and Solvent Extraction
Medicinal plants were collected from different climat zones of Pakistan based on
indigenous knowledge (undocumented reports from Hakeems) and then shade dried.
Extraction of different parts of plants was carried out by simple maceration process with n-
hexane, chloroform, acetone and methanol successively at room temperature (25C0) for 24
hours. Extracts were concentrated by Rotavapor-R200 (Buchi) at 35ºC and stored at -70C0.
Stock solutions of each extract were prepared in DMSO (Dimethyl sulfoxide). The working
stock solution of plant extracts was prepared from main stock by diluting these in the culture
medium.
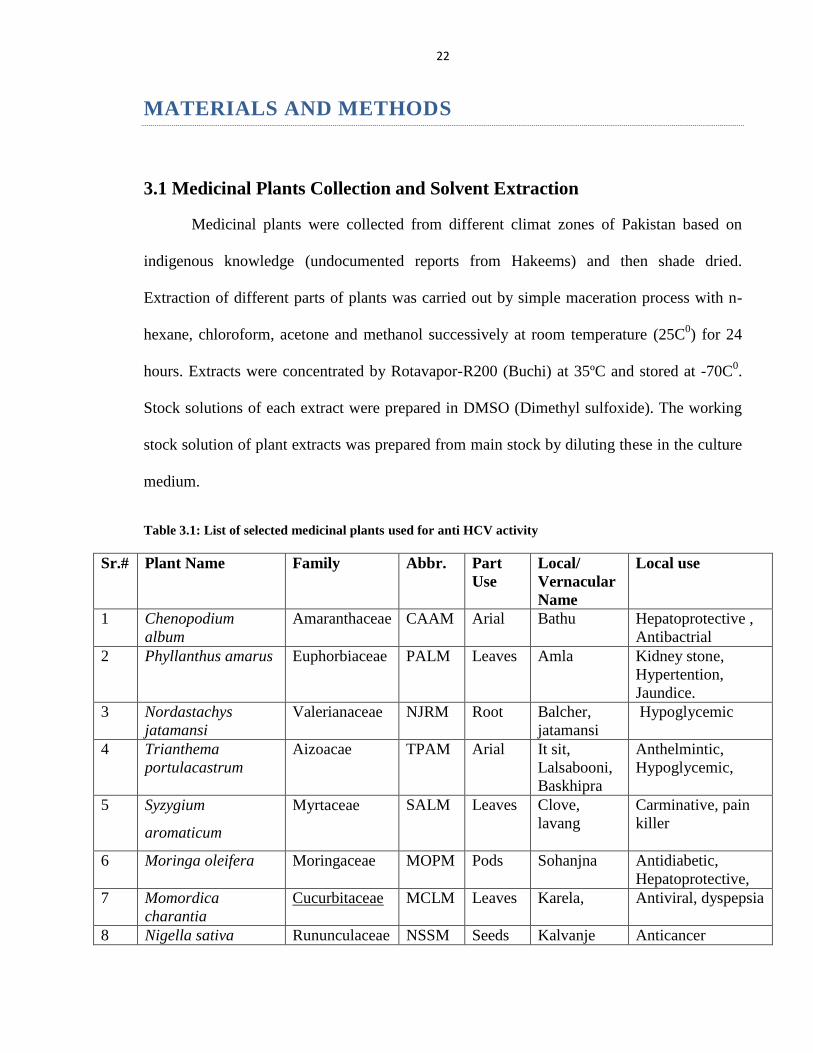
Table 3.1: List of selected medicinal plants used for anti HCV activity
Sr.# Plant Name Family Abbr. Part
Use
Local/
Vernacular
Name
Local use
1 Chenopodium
album
Amaranthaceae CAAM Arial Bathu Hepatoprotective ,
Antibactrial
2 Phyllanthus amarus Euphorbiaceae PALM Leaves Amla Kidney stone,
Hypertention,
Jaundice.
3 Nordastachys
jatamansi
Valerianaceae NJRM Root Balcher,
jatamansi
Hypoglycemic
4 Trianthema
portulacastrum
Aizoacae TPAM Arial It sit,
Lalsabooni,
Baskhipra
Anthelmintic,
Hypoglycemic,
5 Syzygium
aromaticum
Myrtaceae SALM Leaves Clove,
lavang
Carminative, pain
killer
6 Moringa oleifera Moringaceae MOPM Pods Sohanjna Antidiabetic,
Hepatoprotective,
7 Momordica
charantia
Cucurbitaceae MCLM Leaves Karela, Antiviral, dyspepsia
8 Nigella sativa Rununculaceae NSSM Seeds Kalvanje Anticancer
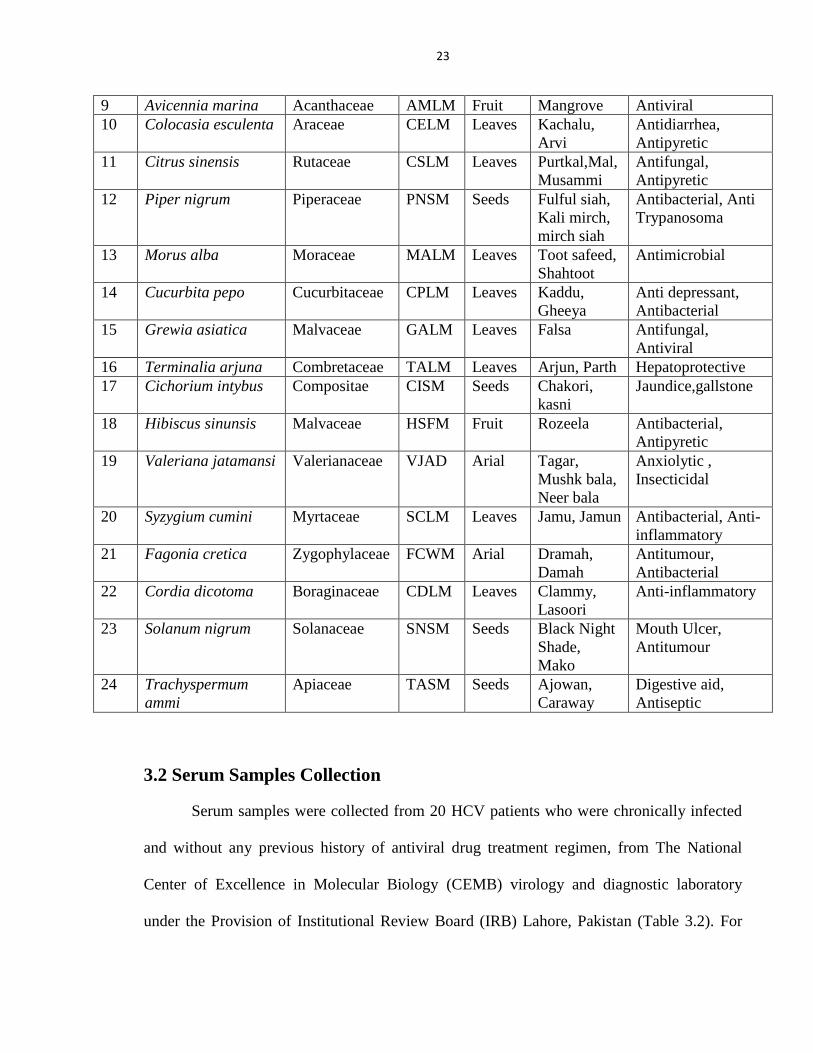
23
9 Avicennia marina Acanthaceae AMLM Fruit Mangrove Antiviral
10 Colocasia esculenta Araceae CELM Leaves Kachalu,
Arvi
Antidiarrhea,
Antipyretic
11 Citrus sinensis Rutaceae CSLM Leaves Purtkal,Mal,
Musammi
Antifungal,
Antipyretic
12 Piper nigrum Piperaceae PNSM Seeds Fulful siah,
Kali mirch,
mirch siah
Antibacterial, Anti
Trypanosoma
13 Morus alba Moraceae MALM Leaves Toot safeed,
Shahtoot
Antimicrobial
14 Cucurbita pepo Cucurbitaceae CPLM Leaves Kaddu,
Gheeya
Anti depressant,
Antibacterial
15 Grewia asiatica Malvaceae GALM Leaves Falsa Antifungal,
Antiviral
16 Terminalia arjuna Combretaceae TALM Leaves Arjun, Parth Hepatoprotective
17 Cichorium intybus Compositae CISM Seeds Chakori,
kasni
Jaundice,gallstone
18 Hibiscus sinunsis Malvaceae HSFM Fruit Rozeela Antibacterial,
Antipyretic
19 Valeriana jatamansi Valerianaceae VJAD Arial Tagar,
Mushk bala,
Neer bala
Anxiolytic ,
Insecticidal
20 Syzygium cumini Myrtaceae SCLM Leaves Jamu, Jamun Antibacterial, Anti-
inflammatory
21 Fagonia cretica Zygophylaceae FCWM Arial Dramah,
Damah
Antitumour,
Antibacterial
22 Cordia dicotoma Boraginaceae CDLM Leaves Clammy,
Lasoori
Anti-inflammatory
23 Solanum nigrum Solanaceae SNSM Seeds Black Night
Shade,
Mako
Mouth Ulcer,
Antitumour
24 Trachyspermum
ammi
Apiaceae TASM Seeds Ajowan,
Caraway
Digestive aid,
Antiseptic
3.2 Serum Samples Collection
Serum samples were collected from 20 HCV patients who were chronically infected
and without any previous history of antiviral drug treatment regimen, from The National
Center of Excellence in Molecular Biology (CEMB) virology and diagnostic laboratory
under the Provision of Institutional Review Board (IRB) Lahore, Pakistan (Table 3.2). For
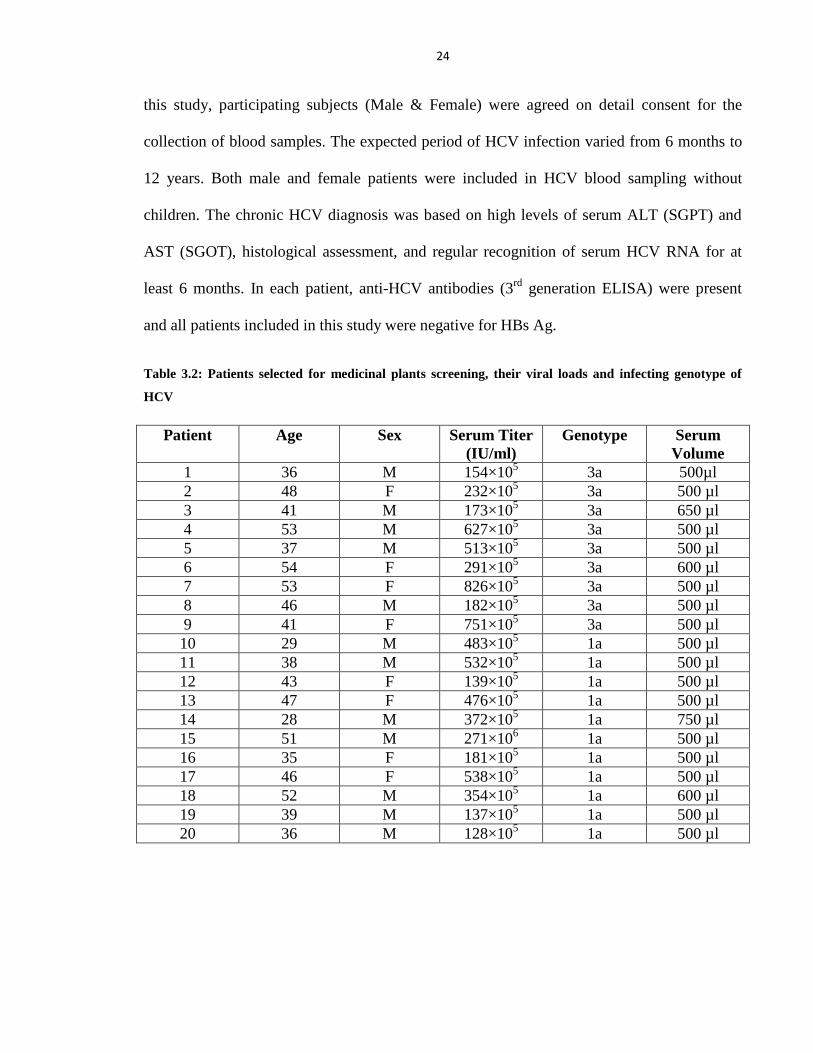
24
this study, participating subjects (Male & Female) were agreed on detail consent for the
collection of blood samples. The expected period of HCV infection varied from 6 months to
12 years. Both male and female patients were included in HCV blood sampling without
children. The chronic HCV diagnosis was based on high levels of serum ALT (SGPT) and
AST (SGOT), histological assessment, and regular recognition of serum HCV RNA for at
least 6 months. In each patient, anti-HCV antibodies (3rd
generation ELISA) were present
and all patients included in this study were negative for HBs Ag.
Table 3.2: Patients selected for medicinal plants screening, their viral loads and infecting genotype of
HCV
Patient Age Sex Serum Titer
(IU/ml)
Genotype Serum
Volume
1 36 M 154×105 3a 500µl
2 48 F 232×105 3a 500 µl
3 41 M 173×105 3a 650 µl
4 53 M 627×105 3a 500 µl
5 37 M 513×105 3a 500 µl
6 54 F 291×105 3a 600 µl
7 53 F 826×105 3a 500 µl
8 46 M 182×105 3a 500 µl
9 41 F 751×105 3a 500 µl
10 29 M 483×105 1a 500 µl
11 38 M 532×105 1a 500 µl
12 43 F 139×105 1a 500 µl
13 47 F 476×105 1a 500 µl
14 28 M 372×105 1a 750 µl
15 51 M 271×106 1a 500 µl
16 35 F 181×105 1a 500 µl
17 46 F 538×105 1a 500 µl
18 52 M 354×105 1a 600 µl
19 39 M 137×105 1a 500 µl
20 36 M 128×105 1a 500 µl

25
3.3 Cell Lines
Huh-7 and MDBK cells were propagated in Dulbecco‘s modified Eagle medium
(DMEM) with addition of cell culture tested 10% fetal bovine serum, 100IU/ml penicillin
and 100µg/ml streptomycin, in 37°C incubator of an atmosphere of 5% CO2. The cells of
Chinese Hamster Ovary (CHO) were cultured in DMEM Hams F12, adding 100IU/ml
penicillin and 100µg/ml streptomycin and 5% fetal bovine serum. The cells of Huh-7 cell
line were generously presented by Dr. Zafar Nawaz (Biochemistry and Molecular Biology
Department, University of Miami, USA). CHO cell line was a kind gift of Dr. Ahmad Usman
Zafar (Biopharmaceutical Lab CEMB, Lahore).
3.4 Plasmids
PCR 3.1 containing NS3 gene of genotype 3a and 1a was taken from functional
Genomics Lab, CEMB Lahore.
3.5 Chemicals
HCV-NS3 specific monoclonal antibody (Sc-52806) was purchased from Santa
Cruiz Biotechnology. Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) and secondary
gout antimouse monoclonal antibodies from Sigma Aldrich (St. Louis, MO, USA) were
purchased. TLC 60 F254 plates were obtained from Merck (Germany) and silica gel with
binder and fluorescent indicator (Cat# 34644-6) from Sigma Aldrich (St. Louis, MO, USA).

26
3.6 Primers Designing
For primers designing, sequences of HCV specific nonstructural gene (NS3) of 3a
and 1a genotype were retrieved from NCBI. The NCBI sequence was altered into FASTA
format and primers were designed through primer 3 software.
Table 3.3.1: Primers of HCV Non-structure (NS3) gene of genotype 3a
S. No Primer Sequence (5‘-3‘) Length
1 NS3F 3a GACCATTGTGACCAGCTTGA 20
2 NS3R 3a GCGGGTGACCAAGTACAAGT 20
Table 3.3.2: Primers of HCV Non-structure (NS3) gene of genotype 1a
S. No Primer Sequence (5‘-3‘) Length
1 NS3F 1a GGACGACGATGACAAGGACT 20 2 NS3R 1a CCTCGTGACCAGGTAAAGGT 20
Table 3.3.3: Primers for Glyceraldehyde-3-Phosphate Dehydrogenase gene (GAPDH)
S. No Primer Sequence (5‘-3‘) Length
1 GAPDH F ACCACAGTCCATGCCATCAC 20
2 GAPDH R TCCACCACCCTGTTGCTGTA 20
3.7 Trypan Blue Dye Explosive Method for Cellular Toxicity
For verification of Huh-7 and CHO cells viability, trypan blue dye explosive method
was employed before antiviral screening of plant extracts. Liver cells (Huh-7) were cultured
in concentration of 8×104 in twelve well plates for toxicological analysis of plant extracts.
First two wells of plate were considered as control and in the remaining wells plant extracts
from lower to higher concentrations were added. After 24 hours cells were trypsinized with
Trypsin EDTA solution, a suspension in 1:1ratio of trypan blue dye and cell suspension was
made and 10µl of this mixture was dispensed on a glass slide and the viable cells were
counted by using hemocytometer under the microscope.

27
3.8 MTT Cell Proliferation Assay
MTT (3-[4, 5-dimethylthiazol-2-y1]-2, 5- diphenyltetrazolium bromide) is a quick,
precise and sensitive in-vitro method to evaluate the toxicity of plant extracts in cell cultures.
The MTT is a yellow colour dye which is reduced to succinic dehydrogenases in living
(attached to bottom of the flask) cells to purple colour formazan crystals which are
precipitated and become insoluble in aqueous medium. The spectrophotometric absorption
wavelength of dissolved purple colour formazan in the visible range shows a direct
relationship with the quantity of cells attached to the bottom (viable cells) (Mosmann, 1983a)
In order to study the toxicity of Huh-7 and CHO cells, 4×104
cells/well of these cells was
cultured in 96 well cell culture plates. The test plant extracts of different concentrations were
added in culture plates after 24 hrs, and kept in an atmosphere of 5% CO2 at 37°C for 24 h.
After that the media having test plant extracts was removed and 100µl fresh media and 30µl
of MTT mixture (5mg/ml in PBS) were dispensed to all wells (1-12) of the 96 well plate. The
plate was wrapped with aluminium foil sheet and placed in 37°C incubator for 3-4 hrs. The
media was removed cautiously and the formazan crystals in all 1-12 columns were dissolved
in 100µl of Dimethyl sulfo-oxide (DMSO). The formazan end product was deduced by
measuring spectrophotometric absorbance at 570 nm (test wavelength) and 620 nm
(reference wavelength) by Enzyme Linked Immunosorbent Assay (ELISA) plate reader.
The viability of cells was determined by following equation.
Cell Viability (Percent) = (Test 570nm-620nm) / (Control 570nm-620nm) × 100

28
3.9 Antiviral Analysis of Compounds in Liver Cells
Liver cells (Huh-7) were propagated in 6 well plates at a concentration of 3×105 cells
per plate. After an incubation period of 24 hrs, 1xPBS was used to wash the cells thrice.
Then, HCV viral inoculations at concentrations of 105 IU of genotype 3a and 1a were added
in each well. In 6 well plate, the first well was chosen as control (only HCV contaminated
serum and the solvent) and dispensed a dose of test plant extract (minimum number of cell
death) in the remaining wells of culture plate for studying the antiviral effect of plant extracts
on the same day. Total RNA was extracted by Gentra RNA isolation kit (Gentra System
Pennsylvania, USA) in accordance with manufacturer‘s protocol. Briefly, cells attached to
bottom of the plate were scratched with cell lysis solution and the pallet of RNA was mixed
homogeneously in 1% Diethyl pyrocarbonate (DEPC) treated water. Then 5µl internal
control (Sacace Biotechnologies Caserta, Italy) was added in each tube. The absolute
quantification of HCV-RNA samples was performed by Real Time PCR Smart Cycler II
system (Cepheid Sunnyvale, USA) by using the Sacace HCV quantitative analysis kit
(Sacace Biotechnologies Caserta, Italy) in accordance with manufacturer‘s protocols.
Calculation of HCV RNA Concentrations
In order to determine the quantity of HCV-RNA, following formula was used.
Where IC represents the internal control for a particular lot prepared.

29
3.10 Transfection of Huh-7 Cells with pCR3.1/Flag TAG/HCV
Nonstructural Gene
Huh-7 (liver) cells were rapidly transfected by HCV non structural gene of 3a and 1a
genotype with constructed plasmids in a dose dependant mode, using LipofectamineTM
2000
(Invitrogen) in accordance with manufacturer‘s instructions. Briefly, 300µl of DMEM
medium and 18µl of transfection reagent were mixed in an eppendroff and allowed to
incubate for 5 minutes at room temperature. DMEM medium of 300µl concentration and
mammalian expression vector of construct pCR3.1/ FlagTAG/HCV non structural gene were
mixed in separate eppendroff and allowed to incubate for 5 min at room temperature and
then, both mixtures were combined in a tube and incubated for 30 minutes at a temperature
of 37°C. The 1XPBS was used to wash the 6 well plate with 24 h incubated cells and 2ml
DMEM medium without any antibiotic and transfection reagent with pCR3.1/FlagTAG/HCV
non structural gene mammalian expression vector mixture in 500µl of media were dispensed
to all wells of the plate in drops and incubated in an atmosphere of 5% CO2 for 24 hrs, at
37°C.
To authenticate the results after transfection, cells were propagated from 24 h to 48 h
of post transfection for nonstructural gene transcription and expression studies.
3.11 Co-transfection of Huh-7 Cells with pCR3.1/FlagTAG/HCV
Nonstructural Gene and Plant Extracts
Huh-7 cells were propageted in DMEM medium, supported with fetal bovine serum
(10%) and antibiotics (1% penicillin/streptomycin) in an atmosphere of 5% CO2 at 37°C. For
transfection analysis, liver cells at 3×105 concentration were seeded in six well culture plates

30
for a period of 24 hrs. The medium was removed gently and cells were rinsed with 1X PBS
twice. Then the cells were momentarily transfected with expression plasmids of HCV
nonstructure gene with test plant extracts by LipofectamineTM
2000 (Invitrogen life
technologies, Carlsbad, CA) in accordance with the manufacturer‘s instructions. The trizol
reagent (Invitrogen life technologies, Carlsbad, CA) was used to extract the total RNA from
each sample according to the manufacturer‘s guidelines. To study the antiviral potential of
test plant extracts against HCV NS3 gene of 3a and 1a genotype, complementary DNA
(cDNA) library was formed with RNA concentration of 1µg, by using Revert Aid TM
First.Strand cDNA Synthesis.Kit (Fermentas, St. Leon-Rot/Germany). HCV-NS3 gene
expression study was performed by Polymerase Chain reaction (PCR) (Applied.Biosystems
Inc, USA) using 2X PCR Master Mix (Fermentas). For amplification of NS3 3a genotype,
following primers were used: forward primer:.GACCATTGTGACCAGCTTGA;.reverse
primer: GCGGGTGACCAAGTACAAGT;. 1a genotype: Forward
primer:.GGACGACGATGACAAGGACT;. Reverse primer:
.CCTCGTGACCAGGTAAAGGT;. while. GAPDH .Forward: primer
CCACAGTCCATGCCATCAC;. and GAPDH. Reverse;. TCCACCACCCTGTTGCTGTA.
Polymerase Chain Reaction was carried out by starting the initial denaturation step at a
temperature of 95°C for 5 minutes following 35 cycles. During PCR each step of
denaturation at 94°C for 1 min, annealing temperature was 58°C for 45sec and extension
time for 10 minutes at a temperature of 72°C. The final amplified DNA products were run on
2% agarose gel. Ultra Violet (UV) light was used to visualize the DNA bands and the gel
photographs were taken with gel documentation system (UVP).

31
3.12 Pharmacological Analysis of Isolated Fractions
After performing the antiviral screening of test plant extracts, the toxicological
studies of the effective extracts was done at higher doses. In order to study the effect of
active fractions at different dosed, liver cells at a concentration of 3×105 cell/well were
propagated in 6 well culturing plates. After 24 hrs of incubation period, cells were treated
with HCV virus copies (2×105) of 3a and 1a genotype in the absence and presence of
different concentrations of three antiviral fractions. For an additional 24 hours of incubation,
Huh-7 cells were kept in CO2 incubator at temperature of 37°C. After incubation period, cell
lysis solution was used to scratch the attached cells and total RNA (cells and serum) was
extracted. The absolute quantification of HCV-RNA was performed by Real Time PCR by
using the Sacace HCV quantitative analysis kit (Sacace.Biotechnologies Caserta,.Italy) in
accordance with the manufacturer‘s guidelines.
3.13 Antiviral Analysis of Effective Fractions along with Interferon (IFN)
After performing the pharmacological studies of isolated fractions, effective fractions
were examined in combination with interferon. Due to the presence of interferon receptors on
MDBK cell line, it is used as a model cell line for the assay of interferon (IFN) (Yanai et al.,
2001), For this purpose, MDBK cells were harvested in six well culturing plates at cell
density of 3×105 per well in DMEM rich medium supported with FBS (10% ) and placed in
incubator of 37C0 for 24hrs. The cells were then tested with active fractions singly and/or in
combination with interferon (IFN) and allowed to incubate for 6 hrs. After incubation time,
cells were treated with inoculums of 2× 105
IU of HCV genotype 3a and 1a per well and

32
again placed in incubator for another 18 hrs. After 24 hrs of incubation time, total RNA
(serum and cells) was take out by RNA isolation kit, and the Real Time Quantitative RT-
PCR was used to find the concentrations of HCV-RNA remaining.
3.14 Protein Isolation and Estimation
After 24 hrs of transfection period, the cells were propagated and protein extraction
was performed for expression studies of NS3 protease. 1X PBS (twice) was used to wash the
transfected cells. Cells were dislocated by adding 500µl of TEN buffer to the cells in
culturing plate and then peeled off after 20 sec. The scratched cells were taken in eppendorf
and centrifuged the cells for 10 min at 13000 rpm (Max. speed) at 4°C to pellet them down.
The cell lysis buffer (50mM Tris-Cl, pH 8.0, 150mM NaCl, 0.02% sodium azide, 1% Triton
X-100, 1µg/ml protease inhibitors, and 100 µg /ml PMSF) 100µl was added to pellet of cells
to homogenize them, allowed to incubate on ice cubes for 15 minutes and placed in
centrifuge at 13000 rpm (Max. speed) at temperature of 4°C for 30 minutes. In eppendorf,
supernatant liquid was taken out by pipette containing protein and freezed at -20°C for
further process. The spectrophotometric method was used to quantify the extracted protein.
Briefly, protein sample of 1µl was added in 800µl volume of 1x PBS and 200µl of Bio Red
dye. Then absorption of sample solutions was noted at 595nm of wavelength.
3.15 Western Blotting
Western blotting gives informative data about the presence of a protein, molecular
weight, and/or quantity of a specific antigen by protein separation in discrete bands via gel
electrophoresis, with specific recognition antigenic sites by antibodies. In order to investigate
protein expression analysis of HCV genes and potential HCV inhibitory potential of plant

33
extracts and their fractions, total protein samples of 100µg were loaded on 10% Sodium
Dodisyl Sulphate (SDS) Polyacrylamide Gel Electrophoresis (PAGE) and allowed to blot on
nitrocellulose membrane (Bio-Rad) electrophoretically. A solution of phosphate buffer saline
was mixed with 5% skim milk to block the membranes at room temperature for 1 h and
treated with HCV specific primary antibody. Then 1x PBS was used to wash the membrane
keeping Tween 20 in 0.1% of concentration. After washing, the nitrocellulose membranes
were placed on smooth surface and treated these membranes with monoclonal antibodies of
HCV-NS3 gene and GAPDH (Santa.Cruiz Biotechnology) specifically. Now the membranes
were washed thrice with 1X TBST and then treated with anti-mouse secondary antibody for a
period of 1 hour. Protein expression analysis was performed by using chemiluminescence
detection kit (Sigma) after washing with TBST thrice. The potential inhibitory effect of
different fractions was analyzed by the intensity of bands on photographic film after ECL.
3.16 Separation and Purification Techniques
3.16.1 Thin Layer Chromatography
Separation of plant extracts were made on precoated silica gel 60 F254 plastic sheets of
thin layer chromatography (TLC) by Merk, Germany. Briefly, 5ml solvent was taken to
prepare 1% sample solution and passed through 0.22µm filter. TLC plate (20×20cm2) was
cut in 10 cm length and 3cm width small plates. 3mm diameter of test sample was spotted on
plates. The spots on plates were air dried and put in chromatography tank, having a
homogeneous mixture of suitable mobile phase. The mobile phase moved on TLC plate for
half an hour. Chromatogram was taken out of tank and solvent front was marked. The

34
chromatogram was air dried and observed under UV light of 254nm and 366nm wavelength.
Marked the spots and Rf value of each spot was calculated.
3.16.2 Column Chromatography
Separation and isolation of compounds from active plant extracts on large scale was
achieved by column chromatography, with silica gel mesh size (70-230µ) as adsorbent with
suitable mobile phase. Flow rate of the eluents on column were 1ml/min at room
temperature.
3.16.3 High Pressure Liquid Chromatography (HPLC)
The active fractions of medicinal plants were analyzed by High Pressure Liquid
Chromatography (HPLC) of Shimadzu LC-10A system. A model LC 10 AT pump along
with wave length detector SPD-10A and CBM-10A were equipped with HPLC. An interface
module class LC-10 HPLC software and a Rheodyne injection valve with a 20 μL loop were
used. By using a Merck C-18 column 250×4.6, i.d., 5 μm particle size, chromatographic
separation was performed. The mobile phase was double distilled methanol and injection
volume was 20µl and with constant flow rate.
3.17 Statistical Analysis
All statistical data analysis was carried out by using GraphPad Prism 4.0 software
(GraphPad.Software, San.Diego, CA, USA). Data is presented as mean ± SE. Numerical data
was analyzed using 1 way ANOVA. *P value ˂ 0.05 and **P value ˂ 0.005 were considered
statistically significant.

35
RESULTS
4.1 Medicinal Plants Collection and Solvent Extraction
Medicinal plants were drawn together from different regions of Pakistan, based on
their local use and undocumented reports against viruses. Different parts (fruit, leaves, bark,
pods or aerial parts) were kept under shade for drying and ground in a grinding mill.
Extraction from these plants was carried out by simple maceration process. List of medicinal
plants with their local use is given in Table 3.1.
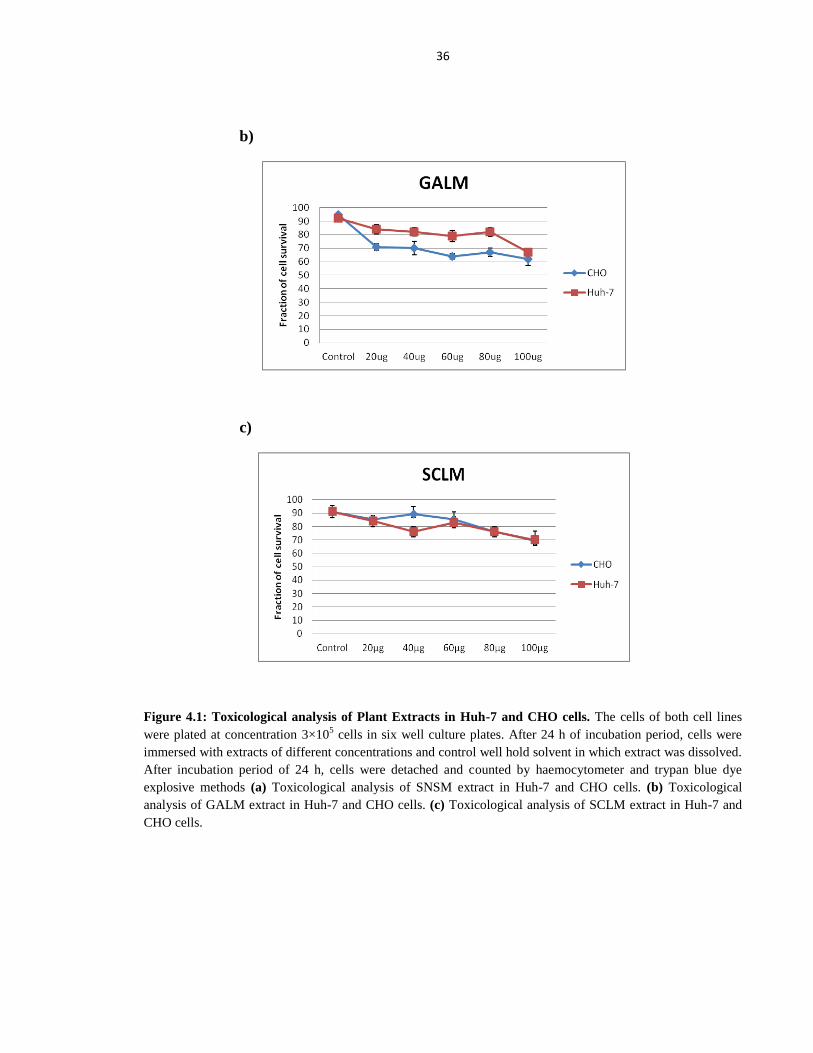
4.2 Cytotoxicity Study of Plant Extracts
Before the antiviral screening of plant extracts against Hepatitis C virus, toxicological
study of twenty four plant extracts were found out by treating cells of Huh-7 and CHO cell
line with different concentrations. In order to study the toxicological effect of plant extracts
to other cells of the body, Chinese Hamster Ovary (CHO) cells were utilized as a control cell
line. Figure 4.1 a, b and c show cytotoxicity analysis of three plants extract and demonstrates
the liver and CHO cells viability at a concentration of 100µg is not affected. For remaining
twenty one plant extracts similar results were observed at concentrations ranging from 10 to
100 µg.
a)
36
b)
c)
Figure 4.1: Toxicological analysis of Plant Extracts in Huh-7 and CHO cells. The cells of both cell lines
were plated at concentration 3×105 cells in six well culture plates. After 24 h of incubation period, cells were
immersed with extracts of different concentrations and control well hold solvent in which extract was dissolved.
After incubation period of 24 h, cells were detached and counted by haemocytometer and trypan blue dye
explosive methods (a) Toxicological analysis of SNSM extract in Huh-7 and CHO cells. (b) Toxicological
analysis of GALM extract in Huh-7 and CHO cells. (c) Toxicological analysis of SCLM extract in Huh-7 and
CHO cells.
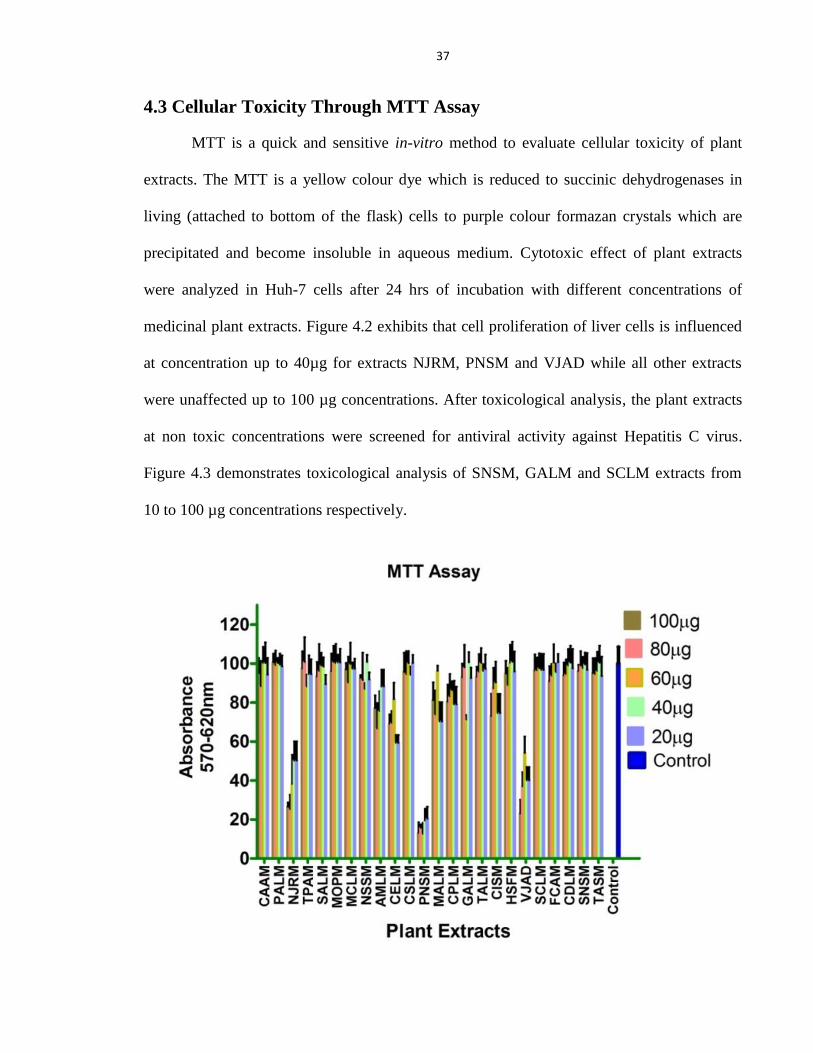
37
4.3 Cellular Toxicity Through MTT Assay
MTT is a quick and sensitive in-vitro method to evaluate cellular toxicity of plant
extracts. The MTT is a yellow colour dye which is reduced to succinic dehydrogenases in
living (attached to bottom of the flask) cells to purple colour formazan crystals which are
precipitated and become insoluble in aqueous medium. Cytotoxic effect of plant extracts
were analyzed in Huh-7 cells after 24 hrs of incubation with different concentrations of
medicinal plant extracts. Figure 4.2 exhibits that cell proliferation of liver cells is influenced
at concentration up to 40µg for extracts NJRM, PNSM and VJAD while all other extracts
were unaffected up to 100 µg concentrations. After toxicological analysis, the plant extracts
at non toxic concentrations were screened for antiviral activity against Hepatitis C virus.
Figure 4.3 demonstrates toxicological analysis of SNSM, GALM and SCLM extracts from
10 to 100 µg concentrations respectively.
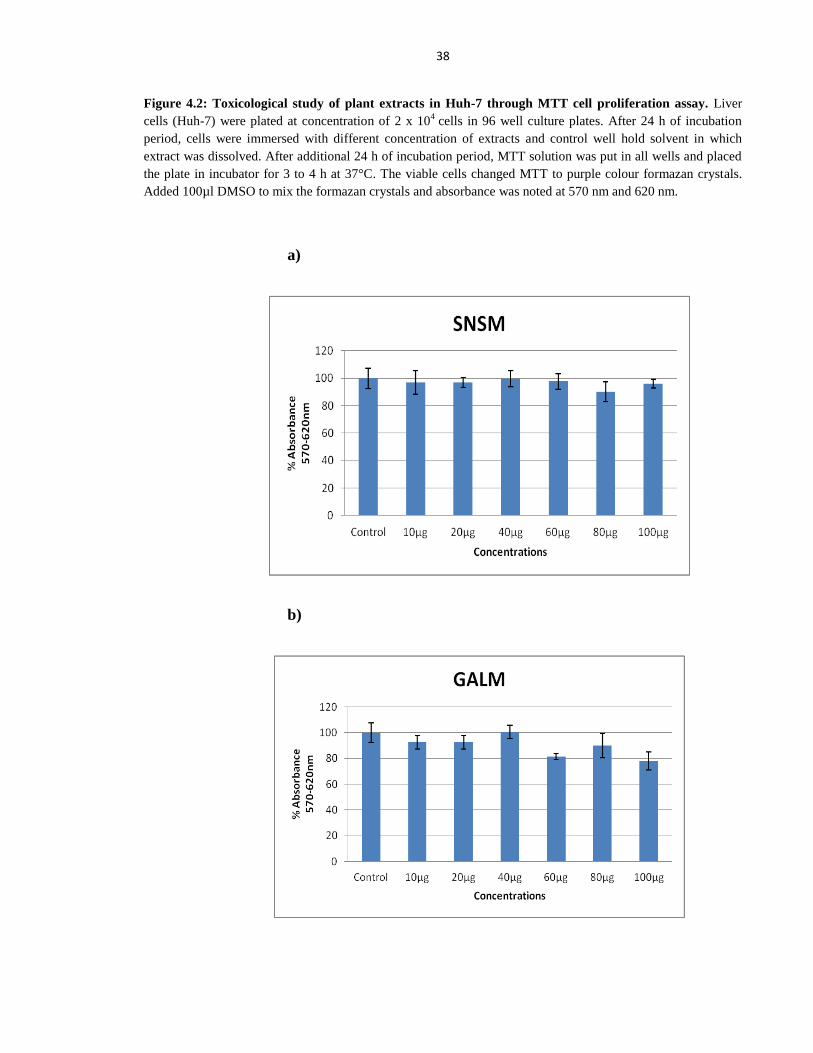
38
Figure 4.2: Toxicological study of plant extracts in Huh-7 through MTT cell proliferation assay. Liver
cells (Huh-7) were plated at concentration of 2 x 104
cells in 96 well culture plates. After 24 h of incubation
period, cells were immersed with different concentration of extracts and control well hold solvent in which
extract was dissolved. After additional 24 h of incubation period, MTT solution was put in all wells and placed
the plate in incubator for 3 to 4 h at 37°C. The viable cells changed MTT to purple colour formazan crystals.
Added 100µl DMSO to mix the formazan crystals and absorbance was noted at 570 nm and 620 nm.
a)
b)
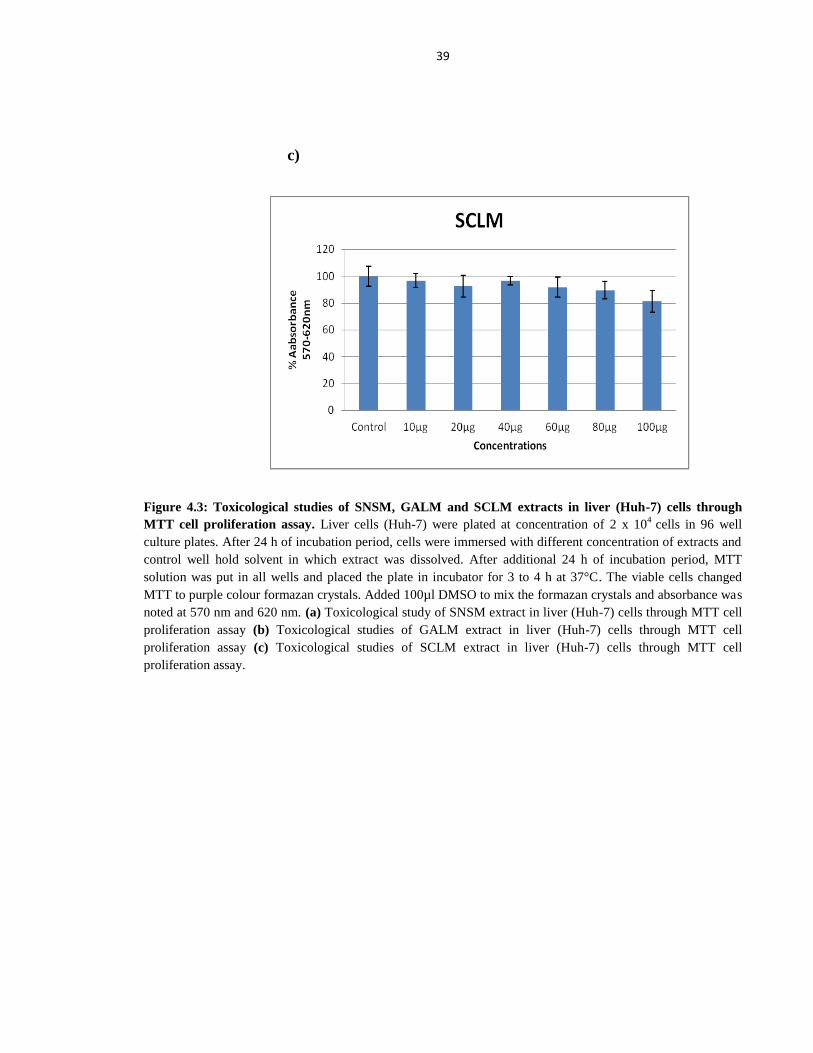
39
c)
Figure 4.3: Toxicological studies of SNSM, GALM and SCLM extracts in liver (Huh-7) cells through
MTT cell proliferation assay. Liver cells (Huh-7) were plated at concentration of 2 x 104
cells in 96 well
culture plates. After 24 h of incubation period, cells were immersed with different concentration of extracts and
control well hold solvent in which extract was dissolved. After additional 24 h of incubation period, MTT
solution was put in all wells and placed the plate in incubator for 3 to 4 h at 37°C. The viable cells changed
MTT to purple colour formazan crystals. Added 100µl DMSO to mix the formazan crystals and absorbance was
noted at 570 nm and 620 nm. (a) Toxicological study of SNSM extract in liver (Huh-7) cells through MTT cell
proliferation assay (b) Toxicological studies of GALM extract in liver (Huh-7) cells through MTT cell
proliferation assay (c) Toxicological studies of SCLM extract in liver (Huh-7) cells through MTT cell
proliferation assay.
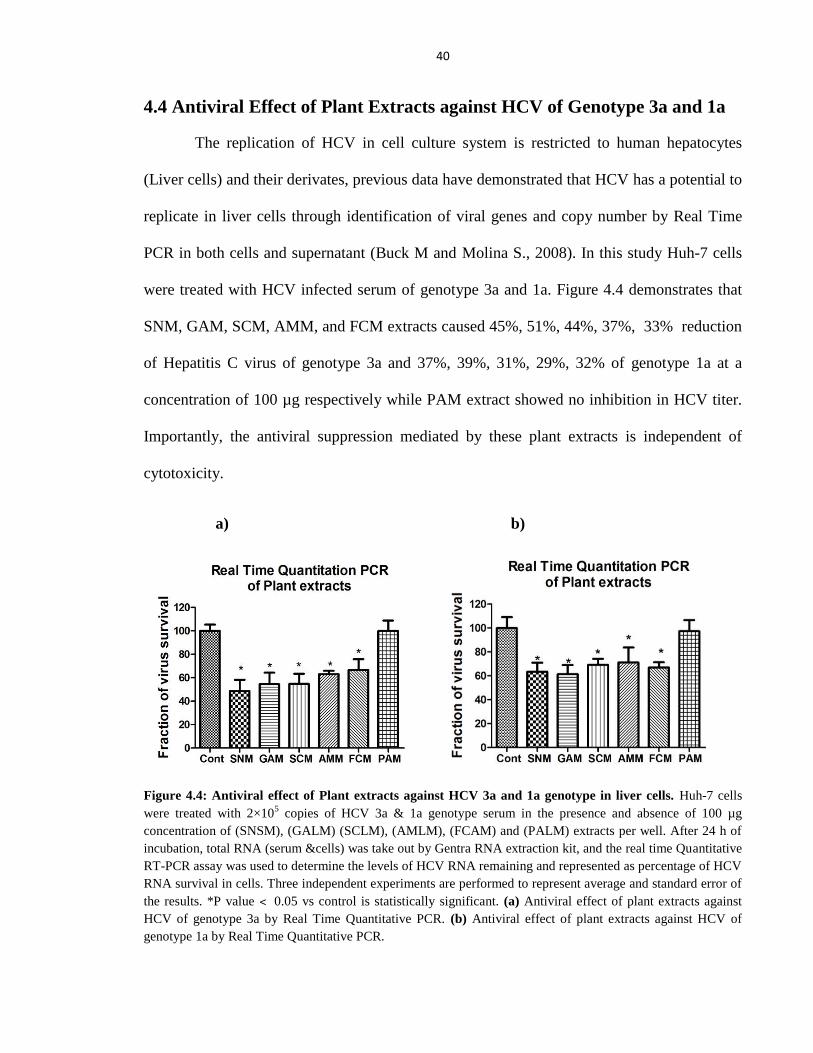
40
4.4 Antiviral Effect of Plant Extracts against HCV of Genotype 3a and 1a
The replication of HCV in cell culture system is restricted to human hepatocytes
(Liver cells) and their derivates, previous data have demonstrated that HCV has a potential to
replicate in liver cells through identification of viral genes and copy number by Real Time
PCR in both cells and supernatant (Buck M and Molina S., 2008). In this study Huh-7 cells
were treated with HCV infected serum of genotype 3a and 1a. Figure 4.4 demonstrates that
SNM, GAM, SCM, AMM, and FCM extracts caused 45%, 51%, 44%, 37%, 33% reduction
of Hepatitis C virus of genotype 3a and 37%, 39%, 31%, 29%, 32% of genotype 1a at a
concentration of 100 µg respectively while PAM extract showed no inhibition in HCV titer.
Importantly, the antiviral suppression mediated by these plant extracts is independent of
cytotoxicity.
a) b)
Figure 4.4: Antiviral effect of Plant extracts against HCV 3a and 1a genotype in liver cells. Huh-7 cells
were treated with 2×105 copies of HCV 3a & 1a genotype serum in the presence and absence of 100 µg
concentration of (SNSM), (GALM) (SCLM), (AMLM), (FCAM) and (PALM) extracts per well. After 24 h of
incubation, total RNA (serum &cells) was take out by Gentra RNA extraction kit, and the real time Quantitative
RT-PCR assay was used to determine the levels of HCV RNA remaining and represented as percentage of HCV
RNA survival in cells. Three independent experiments are performed to represent average and standard error of
the results. *P value ˂ 0.05 vs control is statistically significant. (a) Antiviral effect of plant extracts against
HCV of genotype 3a by Real Time Quantitative PCR. (b) Antiviral effect of plant extracts against HCV of
genotype 1a by Real Time Quantitative PCR.
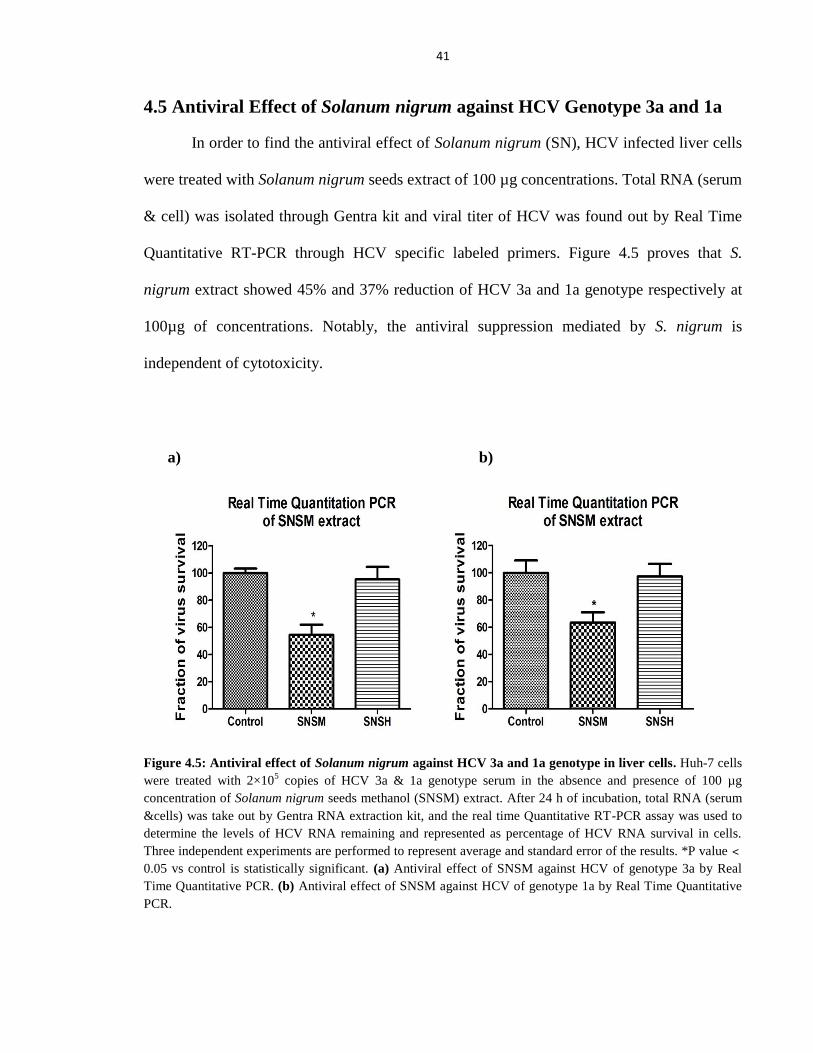
41
4.5 Antiviral Effect of Solanum nigrum against HCV Genotype 3a and 1a
In order to find the antiviral effect of Solanum nigrum (SN), HCV infected liver cells
were treated with Solanum nigrum seeds extract of 100 µg concentrations. Total RNA (serum
& cell) was isolated through Gentra kit and viral titer of HCV was found out by Real Time
Quantitative RT-PCR through HCV specific labeled primers. Figure 4.5 proves that S.
nigrum extract showed 45% and 37% reduction of HCV 3a and 1a genotype respectively at
100µg of concentrations. Notably, the antiviral suppression mediated by S. nigrum is
independent of cytotoxicity.
a) b)
Figure 4.5: Antiviral effect of Solanum nigrum against HCV 3a and 1a genotype in liver cells. Huh-7 cells
were treated with 2×105 copies of HCV 3a & 1a genotype serum in the absence and presence of 100 µg
concentration of Solanum nigrum seeds methanol (SNSM) extract. After 24 h of incubation, total RNA (serum
&cells) was take out by Gentra RNA extraction kit, and the real time Quantitative RT-PCR assay was used to
determine the levels of HCV RNA remaining and represented as percentage of HCV RNA survival in cells.
Three independent experiments are performed to represent average and standard error of the results. *P value ˂
0.05 vs control is statistically significant. (a) Antiviral effect of SNSM against HCV of genotype 3a by Real
Time Quantitative PCR. (b) Antiviral effect of SNSM against HCV of genotype 1a by Real Time Quantitative
PCR.

42
4.5.1 Antiviral Effect of Solanum nigrum against HCV-NS3 Proteases of
Genotype 3a and 1a
To evaluate the antiviral effect of Solanum nigrum (SN) against HCV-NS3 proteases,
liver (Huh-7) cells were seeded in six well plates. After the incubation period, cells were
propagated, RNA was isolated and complementary DNA (cDNA) was produced by ologo dT
priming. PCR was used to amplify cDNA using NS3-3a and NS3-1a gene specific primers of
HCV. The amplification of GAPDH mRNA served as internal control. The result of this
study reveals that methanolic extract of S. nigrum (SNSM) decreases the expression of HCV
RNA significantly at a concentration of 100 µg, while the expression of GAPDH mRNA is
not affected by the addition of extract (Figure 4.5.1.1). Figure 4.5.1.3 showed that S. nigrum
seeds chloroform (SNSC) and methanol (SNSM) extracts reduced the expression of HCV-
NS3 proteases while n-Hexane (SNSH) extract had no effect on HCV-NS3 proteases in Huh-
7 cells at a concentration of 100 µg. Similarly, Real Time PCR results exhibits that SNSM
extract resulted in 52% and 43% inhibition of HCV NS3 proteases level of genotype 3a and
1a respectively at a non toxic concentrations (Figure 4.5.1.2). Figure 4.5.1.4 demonstrates
that SNSH, SNSC and SNSM extracts inhibits 6%, 50%, 61% of NS3 protease level of
genotype 3a and 0%, 41%, 68% of genotype 1a respectively. Collectively, the data
demonstrate that phytochemicals present in medicinal plants significantly inhibit HCV-NS3
Protease expression in Huh-7 cells. Importantly, the antiviral suppression mediated by these
extracts is independent of cytotoxicity.
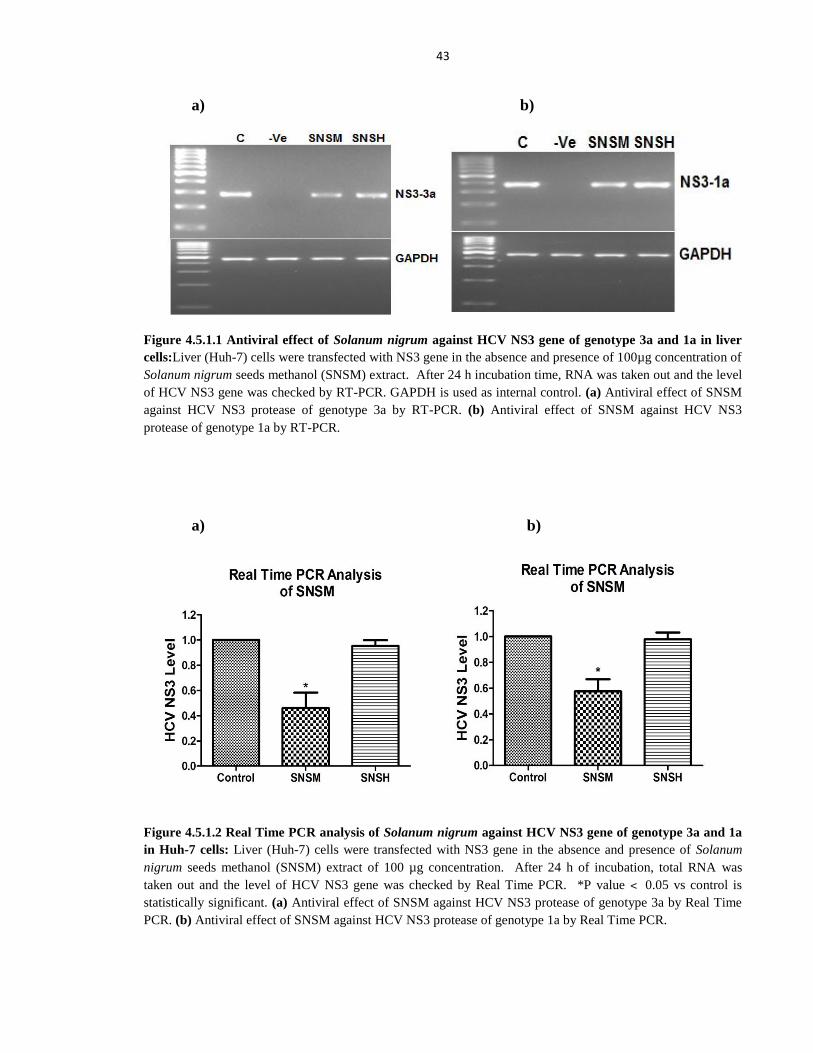
43
a) b)
Figure 4.5.1.1 Antiviral effect of Solanum nigrum against HCV NS3 gene of genotype 3a and 1a in liver
cells:Liver (Huh-7) cells were transfected with NS3 gene in the absence and presence of 100µg concentration of
Solanum nigrum seeds methanol (SNSM) extract. After 24 h incubation time, RNA was taken out and the level
of HCV NS3 gene was checked by RT-PCR. GAPDH is used as internal control. (a) Antiviral effect of SNSM
against HCV NS3 protease of genotype 3a by RT-PCR. (b) Antiviral effect of SNSM against HCV NS3
protease of genotype 1a by RT-PCR.
a) b)
Figure 4.5.1.2 Real Time PCR analysis of Solanum nigrum against HCV NS3 gene of genotype 3a and 1a
in Huh-7 cells: Liver (Huh-7) cells were transfected with NS3 gene in the absence and presence of Solanum
nigrum seeds methanol (SNSM) extract of 100 µg concentration. After 24 h of incubation, total RNA was
taken out and the level of HCV NS3 gene was checked by Real Time PCR. *P value ˂ 0.05 vs control is
statistically significant. (a) Antiviral effect of SNSM against HCV NS3 protease of genotype 3a by Real Time
PCR. (b) Antiviral effect of SNSM against HCV NS3 protease of genotype 1a by Real Time PCR.
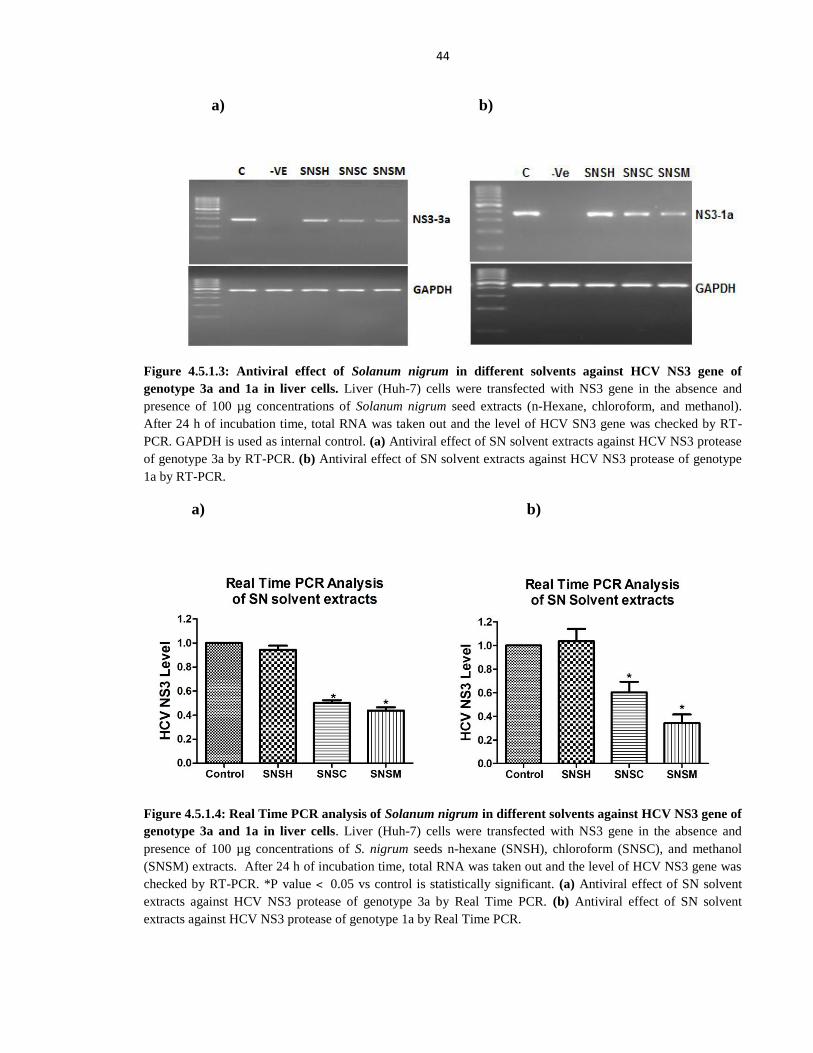
44
a) b)
Figure 4.5.1.3: Antiviral effect of Solanum nigrum in different solvents against HCV NS3 gene of
genotype 3a and 1a in liver cells. Liver (Huh-7) cells were transfected with NS3 gene in the absence and
presence of 100 µg concentrations of Solanum nigrum seed extracts (n-Hexane, chloroform, and methanol).
After 24 h of incubation time, total RNA was taken out and the level of HCV SN3 gene was checked by RT-
PCR. GAPDH is used as internal control. (a) Antiviral effect of SN solvent extracts against HCV NS3 protease
of genotype 3a by RT-PCR. (b) Antiviral effect of SN solvent extracts against HCV NS3 protease of genotype
1a by RT-PCR.
a) b)
Figure 4.5.1.4: Real Time PCR analysis of Solanum nigrum in different solvents against HCV NS3 gene of
genotype 3a and 1a in liver cells. Liver (Huh-7) cells were transfected with NS3 gene in the absence and
presence of 100 µg concentrations of S. nigrum seeds n-hexane (SNSH), chloroform (SNSC), and methanol
(SNSM) extracts. After 24 h of incubation time, total RNA was taken out and the level of HCV NS3 gene was
checked by RT-PCR. *P value ˂ 0.05 vs control is statistically significant. (a) Antiviral effect of SN solvent
extracts against HCV NS3 protease of genotype 3a by Real Time PCR. (b) Antiviral effect of SN solvent
extracts against HCV NS3 protease of genotype 1a by Real Time PCR.

45
4.5.2 Separation & Purification of Solanum nigrum Fractions by
Chromatography
Crude methanol extract of S. nigrum seeds (SNSM) was fractioned by thin layer
chromatography (TLC). The seeds extract separates into thirteen components on TLC plate
in (C: M: W 7:2:1) solvent system with Rf value 0.86, 0.80, 0.77, 0.64, 0.51, 0.44, 0.38, 0.29,
0.15, 0.13, 0.10, 0.08, and 0.05 respectively (Figure 4.5.2.1). For large scale purification
column chromatography was performed and more than 60 fractions were collected. Each
fraction was run on TLC plate and then combined fractions on the basis of their Rf values.
The isolated fractions were then dissolved in DMSO and assessed for antiviral screening
against HCV.
Figure 4.5.2.1: TLC Chromatogram of Solanum nigrum (SN). S. nigrum seeds methanol (SNSM) extract
was run on TLC plate. The crude extract (SNSM) and pure fraction (SN8) was visualized under UV light of
254nm wavelength. In first lane, SNSM extract separates into thirteen components in solvent system
(chloroform: methanol: water 70:20:10) with Rf values 0.86, 0.80, 0.77, 0.64, 0.51, 0.44, 0.38, 0.29, 0.15, 0.13,
0.10, 0.08 and 0.05 respectively. Second lane shows pure SN8 fraction (Rf 0.29).
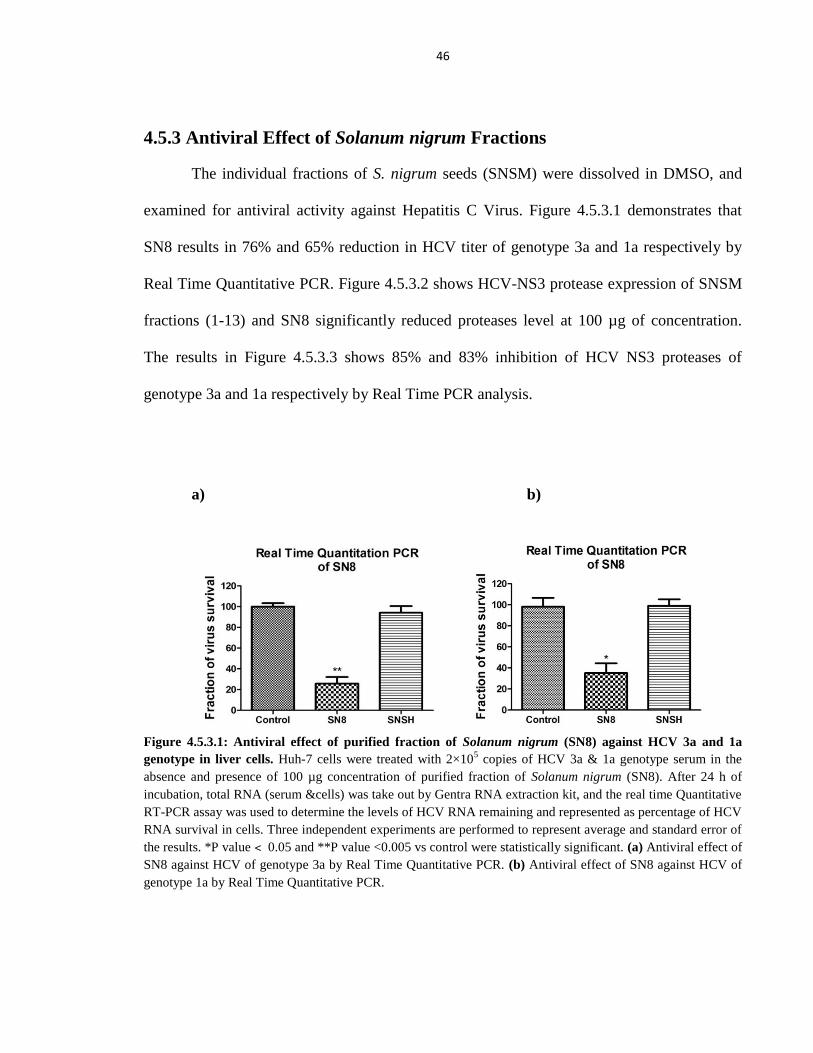
46
4.5.3 Antiviral Effect of Solanum nigrum Fractions
The individual fractions of S. nigrum seeds (SNSM) were dissolved in DMSO, and
examined for antiviral activity against Hepatitis C Virus. Figure 4.5.3.1 demonstrates that
SN8 results in 76% and 65% reduction in HCV titer of genotype 3a and 1a respectively by
Real Time Quantitative PCR. Figure 4.5.3.2 shows HCV-NS3 protease expression of SNSM
fractions (1-13) and SN8 significantly reduced proteases level at 100 µg of concentration.
The results in Figure 4.5.3.3 shows 85% and 83% inhibition of HCV NS3 proteases of
genotype 3a and 1a respectively by Real Time PCR analysis.
a) b)
Figure 4.5.3.1: Antiviral effect of purified fraction of Solanum nigrum (SN8) against HCV 3a and 1a
genotype in liver cells. Huh-7 cells were treated with 2×105 copies of HCV 3a & 1a genotype serum in the
absence and presence of 100 µg concentration of purified fraction of Solanum nigrum (SN8). After 24 h of
incubation, total RNA (serum &cells) was take out by Gentra RNA extraction kit, and the real time Quantitative
RT-PCR assay was used to determine the levels of HCV RNA remaining and represented as percentage of HCV
RNA survival in cells. Three independent experiments are performed to represent average and standard error of
the results. *P value ˂ 0.05 and **P value <0.005 vs control were statistically significant. (a) Antiviral effect of
SN8 against HCV of genotype 3a by Real Time Quantitative PCR. (b) Antiviral effect of SN8 against HCV of
genotype 1a by Real Time Quantitative PCR.
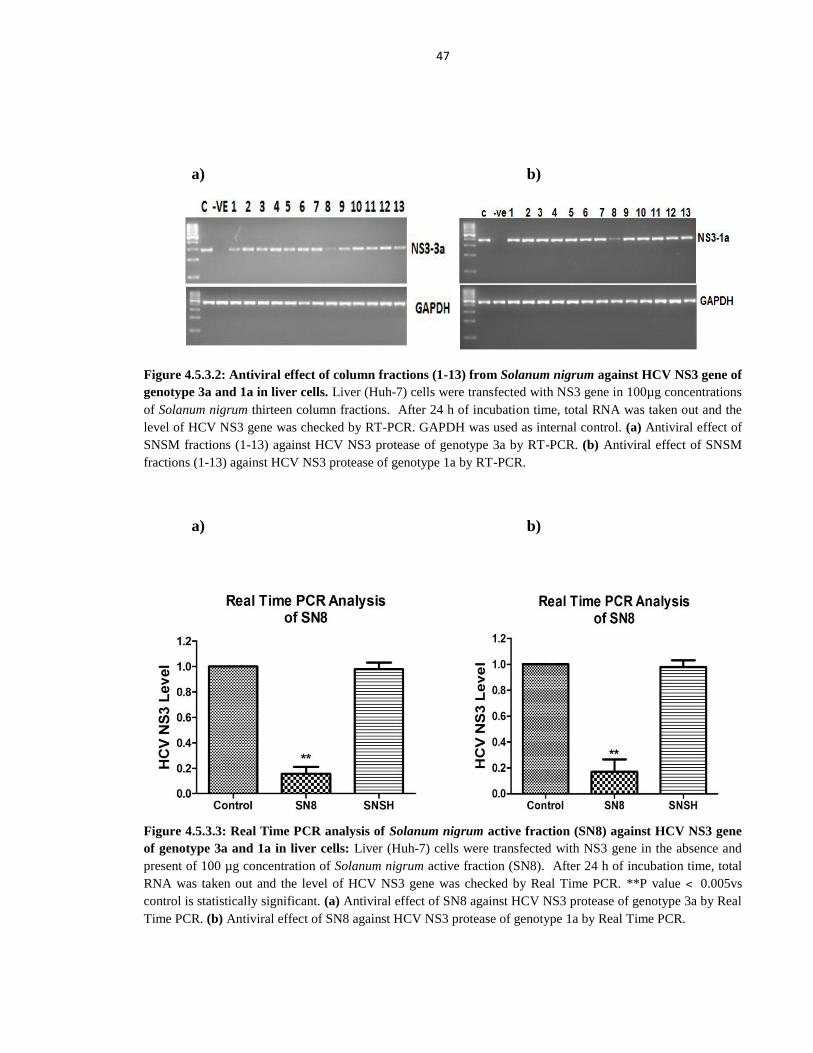
47
a) b)
Figure 4.5.3.2: Antiviral effect of column fractions (1-13) from Solanum nigrum against HCV NS3 gene of
genotype 3a and 1a in liver cells. Liver (Huh-7) cells were transfected with NS3 gene in 100µg concentrations
of Solanum nigrum thirteen column fractions. After 24 h of incubation time, total RNA was taken out and the
level of HCV NS3 gene was checked by RT-PCR. GAPDH was used as internal control. (a) Antiviral effect of
SNSM fractions (1-13) against HCV NS3 protease of genotype 3a by RT-PCR. (b) Antiviral effect of SNSM
fractions (1-13) against HCV NS3 protease of genotype 1a by RT-PCR.
a) b)
Figure 4.5.3.3: Real Time PCR analysis of Solanum nigrum active fraction (SN8) against HCV NS3 gene
of genotype 3a and 1a in liver cells: Liver (Huh-7) cells were transfected with NS3 gene in the absence and
present of 100 µg concentration of Solanum nigrum active fraction (SN8). After 24 h of incubation time, total
RNA was taken out and the level of HCV NS3 gene was checked by Real Time PCR. **P value ˂ 0.005vs
control is statistically significant. (a) Antiviral effect of SN8 against HCV NS3 protease of genotype 3a by Real
Time PCR. (b) Antiviral effect of SN8 against HCV NS3 protease of genotype 1a by Real Time PCR.

48
Additionally, the antiviral effect of S. nigrum active fraction (SN8) was also
evaluated against nonstructural gene by transfection of NS3 gene in liver cells. The lysates
from the Huh-7 cells transfected with HCV NS3 gene were studied by western blot, using
HCV NS3 specific antibodies and GAPDH served as internal control. There was a
remarkable decrease in HCV NS3 protein level in cells treated with S. nigrum active fraction
(SN8) where as the GAPDH protein expression level remained same in control as compare to
treated cells (Figure 4.5.3.4).
Figure 4.5.3.4: HCV NS3 gene inhibition by S.nigrum at protein level: Protein expression level was
determined by transfection of Huh-7 cells with NS3 gene in presence of S.nigrum seeds n-hexane (SNSH)
extract and active fraction (SN8). After 48 h of incubation time, protein was extracted and analyzed by western
blot with HCV NS3 monoclonal antibody and GAPDH monoclonal antibody serves as internal control.
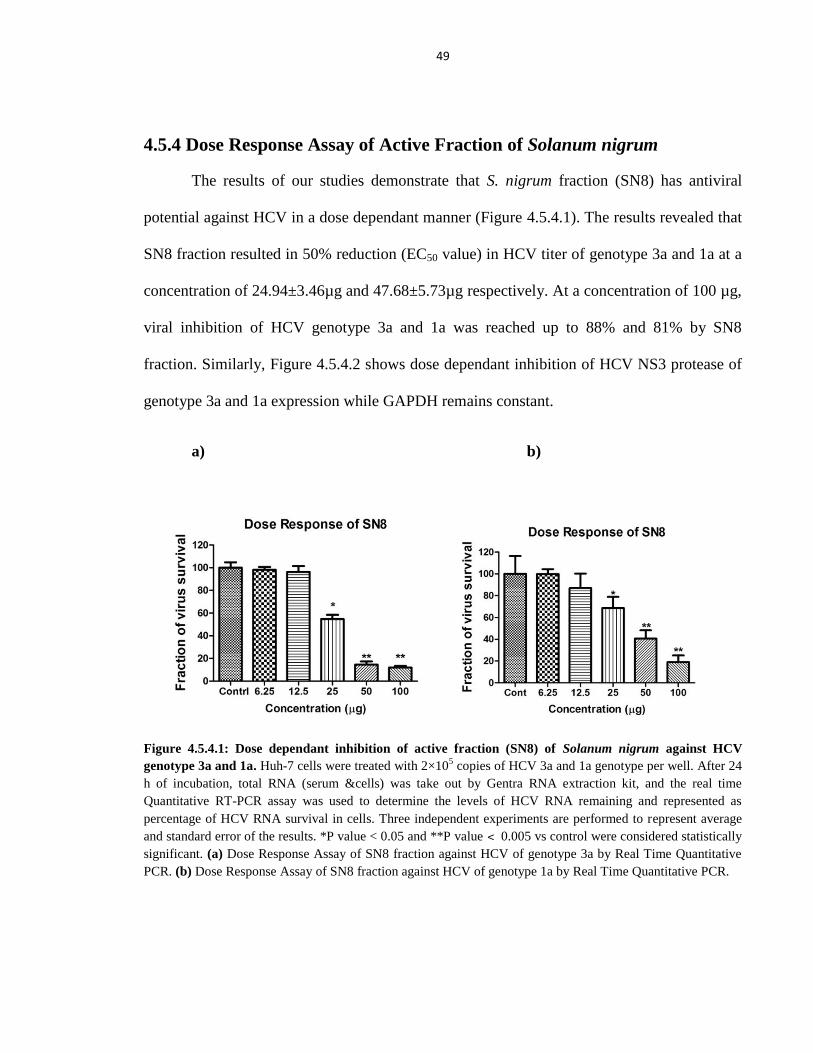
49
4.5.4 Dose Response Assay of Active Fraction of Solanum nigrum
The results of our studies demonstrate that S. nigrum fraction (SN8) has antiviral
potential against HCV in a dose dependant manner (Figure 4.5.4.1). The results revealed that
SN8 fraction resulted in 50% reduction (EC50 value) in HCV titer of genotype 3a and 1a at a
concentration of 24.94±3.46µg and 47.68±5.73µg respectively. At a concentration of 100 µg,
viral inhibition of HCV genotype 3a and 1a was reached up to 88% and 81% by SN8
fraction. Similarly, Figure 4.5.4.2 shows dose dependant inhibition of HCV NS3 protease of
genotype 3a and 1a expression while GAPDH remains constant.
a) b)
Figure 4.5.4.1: Dose dependant inhibition of active fraction (SN8) of Solanum nigrum against HCV
genotype 3a and 1a. Huh-7 cells were treated with 2×105 copies of HCV 3a and 1a genotype per well. After 24
h of incubation, total RNA (serum &cells) was take out by Gentra RNA extraction kit, and the real time
Quantitative RT-PCR assay was used to determine the levels of HCV RNA remaining and represented as
percentage of HCV RNA survival in cells. Three independent experiments are performed to represent average
and standard error of the results. *P value < 0.05 and **P value ˂ 0.005 vs control were considered statistically
significant. (a) Dose Response Assay of SN8 fraction against HCV of genotype 3a by Real Time Quantitative
PCR. (b) Dose Response Assay of SN8 fraction against HCV of genotype 1a by Real Time Quantitative PCR.

50
a) b)
Figure 4.5.4.2: Antiviral effect of active fraction (8th
) from Solanum nigrum in different concentrations
against HCV NS3 gene of genotype 3a and 1a in liver cells: Liver (Huh-7) cells were transfected with NS3
gene in different concentrations of Solanum nigrum 8th
column fraction. After 24 h of incubation time, total
RNA was taken out and the level of HCV NS3 gene was determined by RT-PCR. GAPDH serves as internal
control. (a) Dose Response Assay of SN8 fraction against HCV NS3 protease of genotype 3a by RT-PCR. (b)
Dose Response Assay of SN8 fraction against HCV NS3 protease of genotype 1a by RT-PCR.
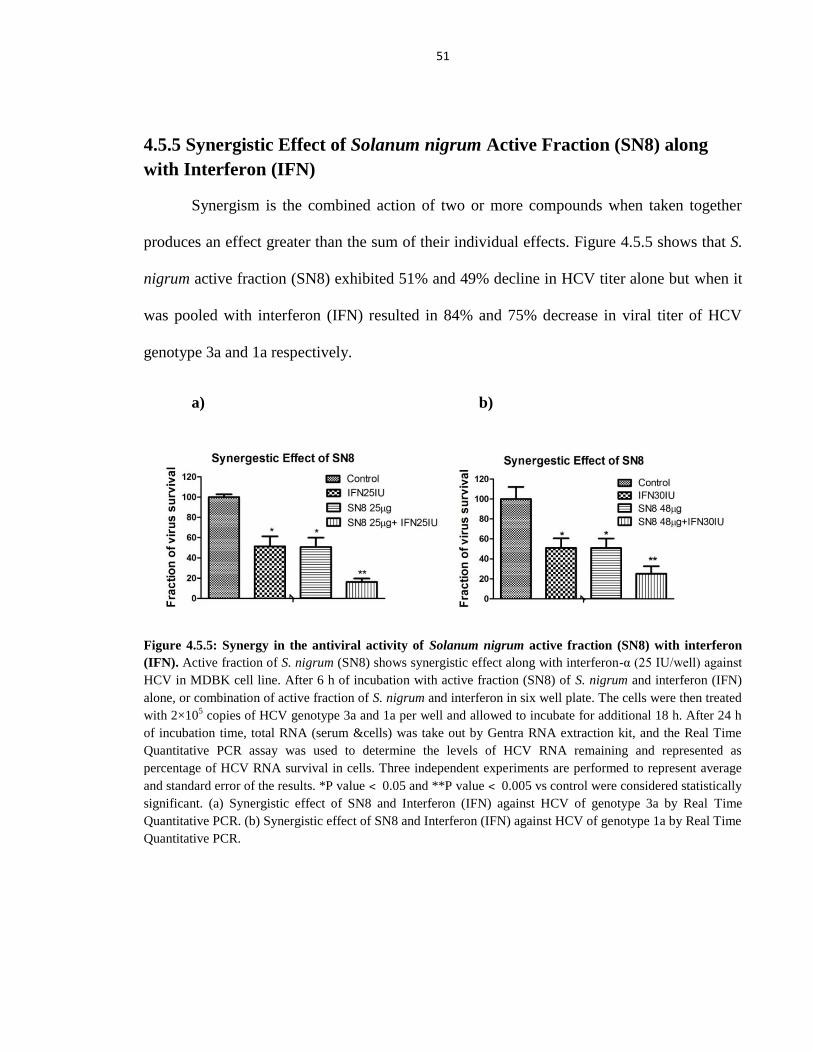
51
4.5.5 Synergistic Effect of Solanum nigrum Active Fraction (SN8) along
with Interferon (IFN)
Synergism is the combined action of two or more compounds when taken together
produces an effect greater than the sum of their individual effects. Figure 4.5.5 shows that S.
nigrum active fraction (SN8) exhibited 51% and 49% decline in HCV titer alone but when it
was pooled with interferon (IFN) resulted in 84% and 75% decrease in viral titer of HCV
genotype 3a and 1a respectively.
a) b)
Figure 4.5.5: Synergy in the antiviral activity of Solanum nigrum active fraction (SN8) with interferon
(IFN). Active fraction of S. nigrum (SN8) shows synergistic effect along with interferon-α (25 IU/well) against
HCV in MDBK cell line. After 6 h of incubation with active fraction (SN8) of S. nigrum and interferon (IFN)
alone, or combination of active fraction of S. nigrum and interferon in six well plate. The cells were then treated
with 2×105 copies of HCV genotype 3a and 1a per well and allowed to incubate for additional 18 h. After 24 h
of incubation time, total RNA (serum &cells) was take out by Gentra RNA extraction kit, and the Real Time
Quantitative PCR assay was used to determine the levels of HCV RNA remaining and represented as
percentage of HCV RNA survival in cells. Three independent experiments are performed to represent average
and standard error of the results. *P value ˂ 0.05 and **P value ˂ 0.005 vs control were considered statistically
significant. (a) Synergistic effect of SN8 and Interferon (IFN) against HCV of genotype 3a by Real Time
Quantitative PCR. (b) Synergistic effect of SN8 and Interferon (IFN) against HCV of genotype 1a by Real Time
Quantitative PCR.
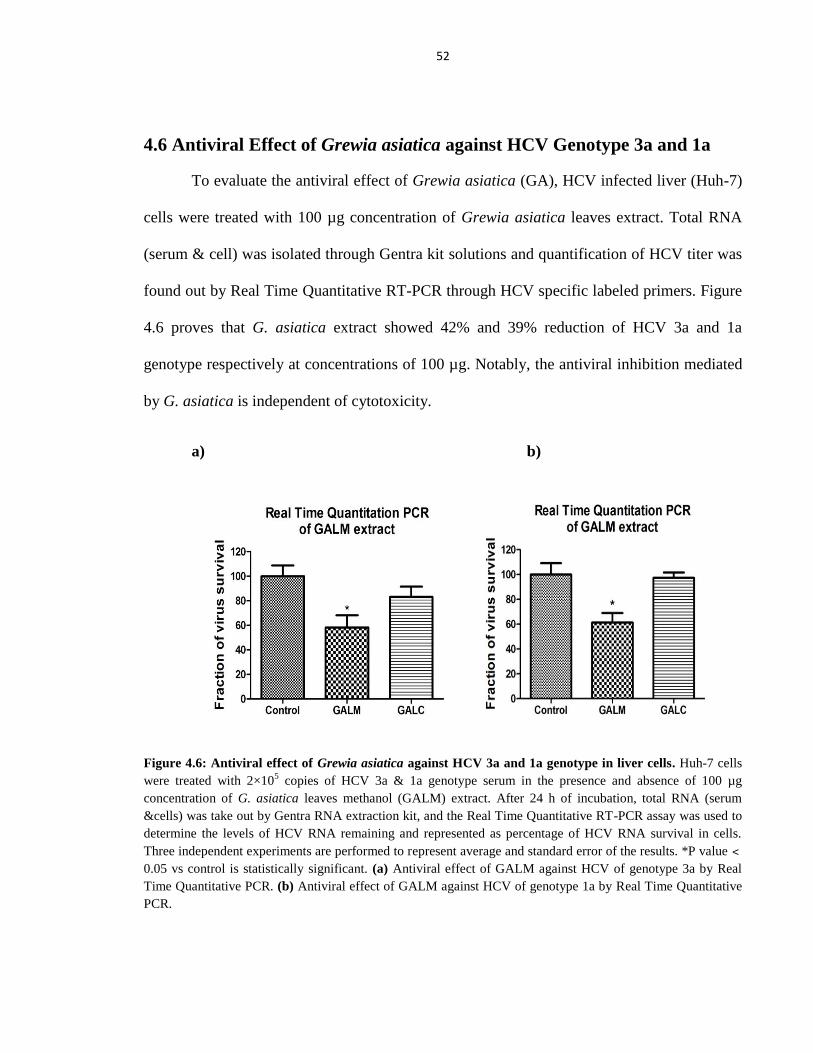
52
4.6 Antiviral Effect of Grewia asiatica against HCV Genotype 3a and 1a
To evaluate the antiviral effect of Grewia asiatica (GA), HCV infected liver (Huh-7)
cells were treated with 100 µg concentration of Grewia asiatica leaves extract. Total RNA
(serum & cell) was isolated through Gentra kit solutions and quantification of HCV titer was
found out by Real Time Quantitative RT-PCR through HCV specific labeled primers. Figure
4.6 proves that G. asiatica extract showed 42% and 39% reduction of HCV 3a and 1a
genotype respectively at concentrations of 100 µg. Notably, the antiviral inhibition mediated
by G. asiatica is independent of cytotoxicity.
a) b)
Figure 4.6: Antiviral effect of Grewia asiatica against HCV 3a and 1a genotype in liver cells. Huh-7 cells
were treated with 2×105 copies of HCV 3a & 1a genotype serum in the presence and absence of 100 µg
concentration of G. asiatica leaves methanol (GALM) extract. After 24 h of incubation, total RNA (serum
&cells) was take out by Gentra RNA extraction kit, and the Real Time Quantitative RT-PCR assay was used to
determine the levels of HCV RNA remaining and represented as percentage of HCV RNA survival in cells.
Three independent experiments are performed to represent average and standard error of the results. *P value ˂
0.05 vs control is statistically significant. (a) Antiviral effect of GALM against HCV of genotype 3a by Real
Time Quantitative PCR. (b) Antiviral effect of GALM against HCV of genotype 1a by Real Time Quantitative
PCR.

53
4.6.1 Antiviral effect of Grewia asiatica against HCV-NS3 Proteases of
Genotype 3a and 1a
To evaluate the antiviral effect of Grewia asiatica (GA) against HCV-NS3 proteases,
liver (Huh-7) cells were propagated in six well culture plates. After the incubation period,
cells were propagated, RNA was isolated and complementary DNA (cDNA) was produced
by ologo dT priming. PCR was used to amplify cDNA using NS3-3a and NS3-1a gene
specific primers of HCV. The amplification of GAPDH messenger RNA is used as internal
control. The result of this study demonstrates that methanolic extract of G. asiatica (GALM)
decreases HCV RNA level significantly at a concentration of 100 µg, while the expression of
GAPDH messenger RNA is not affected by the addition of plant extract (Figure 4.6.1.1).
Figure 4.6.1.3 showed that G. asiatica methanol (GALM) extract reduced the expression of
HCV-NS3 proteases while chloroform (GALC) and acetone (GALA) extracts had no effect
on HCV-NS3 proteases in Huh-7 cells at a concentration of 100 µg. Similarly, Real Time
PCR results exhibits that GALM extract resulted in 59% and 35% inhibition of HCV NS3
proteases level of genotype 3a and 1a respectively at a non toxic concentrations (Figure
4.6.1.2). Figure 4.6.1.4 demonstrates that GALC, GALA and GALM extracts inhibits 3%,
4%, 72% of NS3 protease level of genotype 3a and 0%, 3%, 91% of genotype 1a
respectively. Collectively, the data demonstrate that phytochemicals present in medicinal
plants significantly inhibit HCV-NS3 Protease expression in Huh-7 cells. Importantly, the
antiviral suppression mediated by these extracts is independent of cytotoxicity.
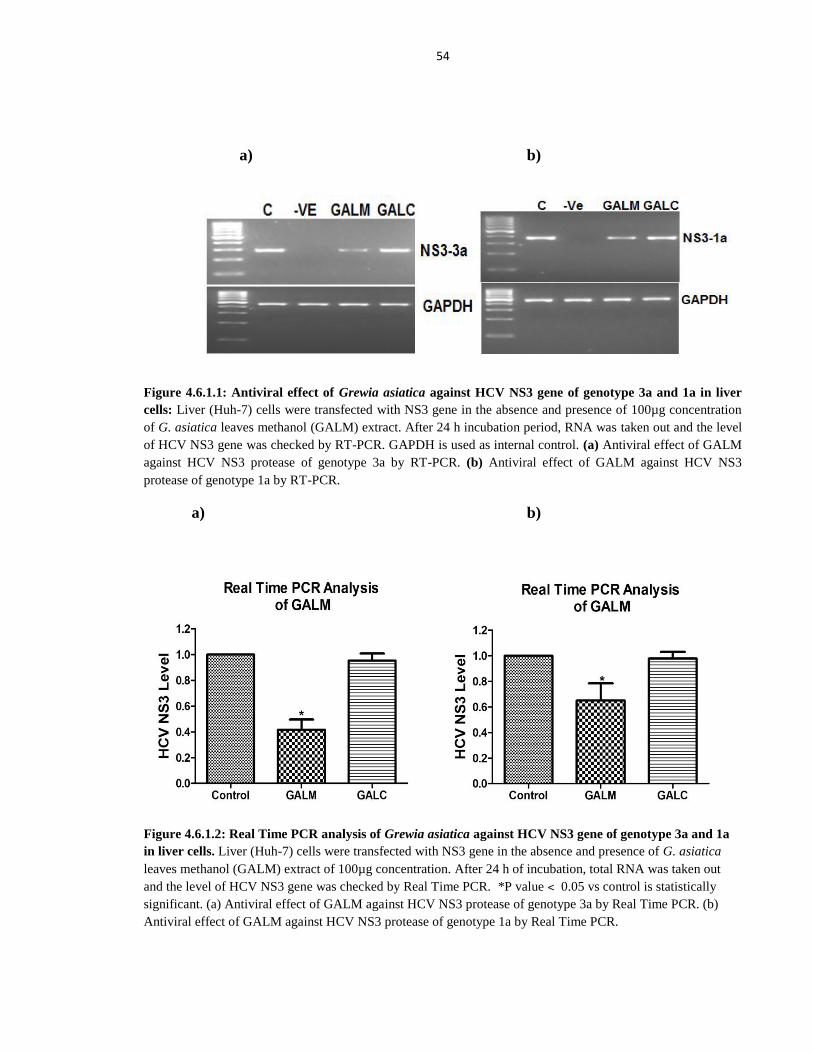
54
a) b)
Figure 4.6.1.1: Antiviral effect of Grewia asiatica against HCV NS3 gene of genotype 3a and 1a in liver
cells: Liver (Huh-7) cells were transfected with NS3 gene in the absence and presence of 100µg concentration
of G. asiatica leaves methanol (GALM) extract. After 24 h incubation period, RNA was taken out and the level
of HCV NS3 gene was checked by RT-PCR. GAPDH is used as internal control. (a) Antiviral effect of GALM
against HCV NS3 protease of genotype 3a by RT-PCR. (b) Antiviral effect of GALM against HCV NS3
protease of genotype 1a by RT-PCR.
a) b)
Figure 4.6.1.2: Real Time PCR analysis of Grewia asiatica against HCV NS3 gene of genotype 3a and 1a
in liver cells. Liver (Huh-7) cells were transfected with NS3 gene in the absence and presence of G. asiatica
leaves methanol (GALM) extract of 100µg concentration. After 24 h of incubation, total RNA was taken out
and the level of HCV NS3 gene was checked by Real Time PCR. *P value ˂ 0.05 vs control is statistically
significant. (a) Antiviral effect of GALM against HCV NS3 protease of genotype 3a by Real Time PCR. (b)
Antiviral effect of GALM against HCV NS3 protease of genotype 1a by Real Time PCR.
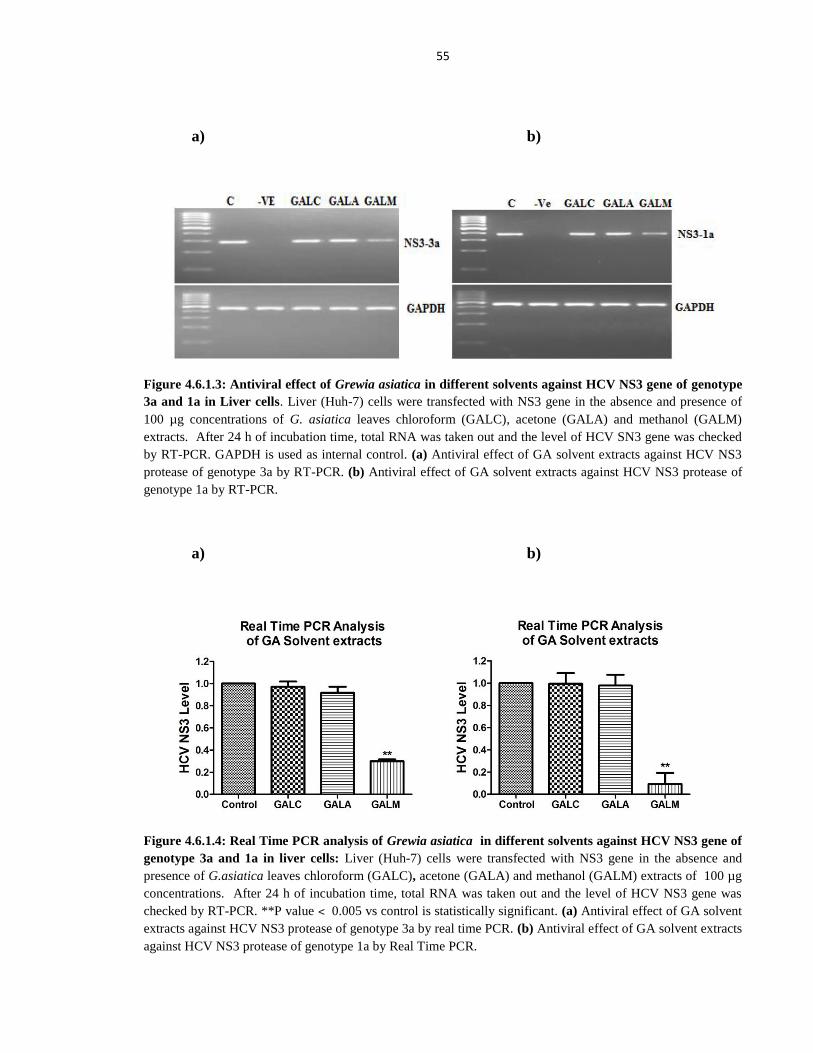
55
a) b)
Figure 4.6.1.3: Antiviral effect of Grewia asiatica in different solvents against HCV NS3 gene of genotype
3a and 1a in Liver cells. Liver (Huh-7) cells were transfected with NS3 gene in the absence and presence of
100 µg concentrations of G. asiatica leaves chloroform (GALC), acetone (GALA) and methanol (GALM)
extracts. After 24 h of incubation time, total RNA was taken out and the level of HCV SN3 gene was checked
by RT-PCR. GAPDH is used as internal control. (a) Antiviral effect of GA solvent extracts against HCV NS3
protease of genotype 3a by RT-PCR. (b) Antiviral effect of GA solvent extracts against HCV NS3 protease of
genotype 1a by RT-PCR.
a) b)
Figure 4.6.1.4: Real Time PCR analysis of Grewia asiatica in different solvents against HCV NS3 gene of
genotype 3a and 1a in liver cells: Liver (Huh-7) cells were transfected with NS3 gene in the absence and
presence of G.asiatica leaves chloroform (GALC), acetone (GALA) and methanol (GALM) extracts of 100 µg
concentrations. After 24 h of incubation time, total RNA was taken out and the level of HCV NS3 gene was
checked by RT-PCR. **P value ˂ 0.005 vs control is statistically significant. (a) Antiviral effect of GA solvent
extracts against HCV NS3 protease of genotype 3a by real time PCR. (b) Antiviral effect of GA solvent extracts
against HCV NS3 protease of genotype 1a by Real Time PCR.

56
4.6.2 Separation & Purification of Grewia asiatica Fractions by
Chromatography
Crude methanol extract of G. asiatica leaves (GALM) was fractioned by thin layer
chromatography (TLC). The leaves extract separates into twelve components on TLC plate in
(C: M: EA: W 5:3:1:1) solvent system with Rf value 0.93, 0.89, 0.84, 0.80, 0.70, 0.66, 0.61,
0.55, 0.50, 0.47, 0.44 and 0.34 respectively. Furthermore, the spot on base line was again run
in (C: EA: AA: W 4: 3: 2: 1) solvent system and It separates into four components with Rf
value 0.93, 0.89, 0.72, and 0.21 respectively. On large scale purification column
chromatography was performed and more than 75 fractions were collected. Each fraction was
run on TLC plate and then combined fractions on the basis of their Rf values. The isolated
fractions were then dissolved in DMSO and tested for antiviral screening against HCV.
Figure 4.6.2.1: TLC Chromatogram of Grewia asiatica (GA). G. asiatica leaves methanol (GALM) extract
was run on TLC plate. the crude extract (GALM) and pure fraction (GA15) were visualized under UV light of
254nm wavelength. In first lane, GALM extract separates into four components in solvent system (C: EA: AA:
W 4: 3: 2: 1) with Rf values 0.93, 0.89, 0.72, and 0.21 respectively. Second lane shows pure GA15 fraction with
Rf value 0.72.
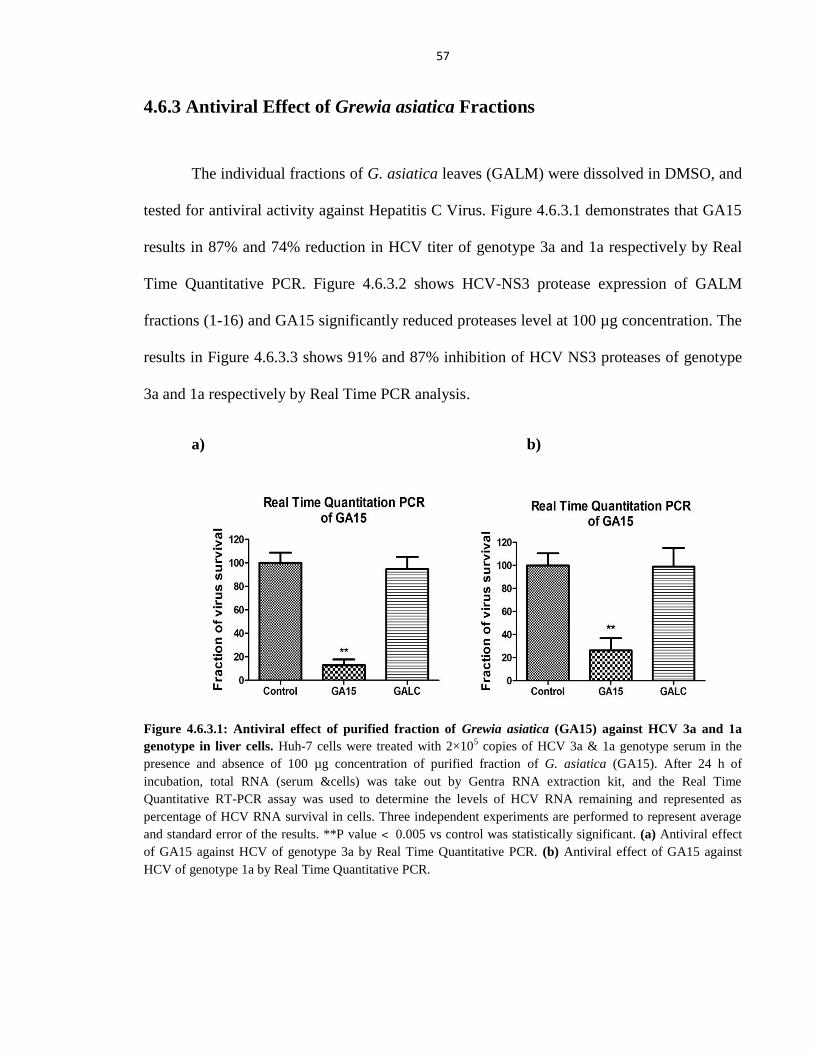
57
4.6.3 Antiviral Effect of Grewia asiatica Fractions
The individual fractions of G. asiatica leaves (GALM) were dissolved in DMSO, and
tested for antiviral activity against Hepatitis C Virus. Figure 4.6.3.1 demonstrates that GA15
results in 87% and 74% reduction in HCV titer of genotype 3a and 1a respectively by Real
Time Quantitative PCR. Figure 4.6.3.2 shows HCV-NS3 protease expression of GALM
fractions (1-16) and GA15 significantly reduced proteases level at 100 µg concentration. The
results in Figure 4.6.3.3 shows 91% and 87% inhibition of HCV NS3 proteases of genotype
3a and 1a respectively by Real Time PCR analysis.
a) b)
Figure 4.6.3.1: Antiviral effect of purified fraction of Grewia asiatica (GA15) against HCV 3a and 1a
genotype in liver cells. Huh-7 cells were treated with 2×105 copies of HCV 3a & 1a genotype serum in the
presence and absence of 100 µg concentration of purified fraction of G. asiatica (GA15). After 24 h of
incubation, total RNA (serum &cells) was take out by Gentra RNA extraction kit, and the Real Time
Quantitative RT-PCR assay was used to determine the levels of HCV RNA remaining and represented as
percentage of HCV RNA survival in cells. Three independent experiments are performed to represent average
and standard error of the results. **P value ˂ 0.005 vs control was statistically significant. (a) Antiviral effect
of GA15 against HCV of genotype 3a by Real Time Quantitative PCR. (b) Antiviral effect of GA15 against
HCV of genotype 1a by Real Time Quantitative PCR.
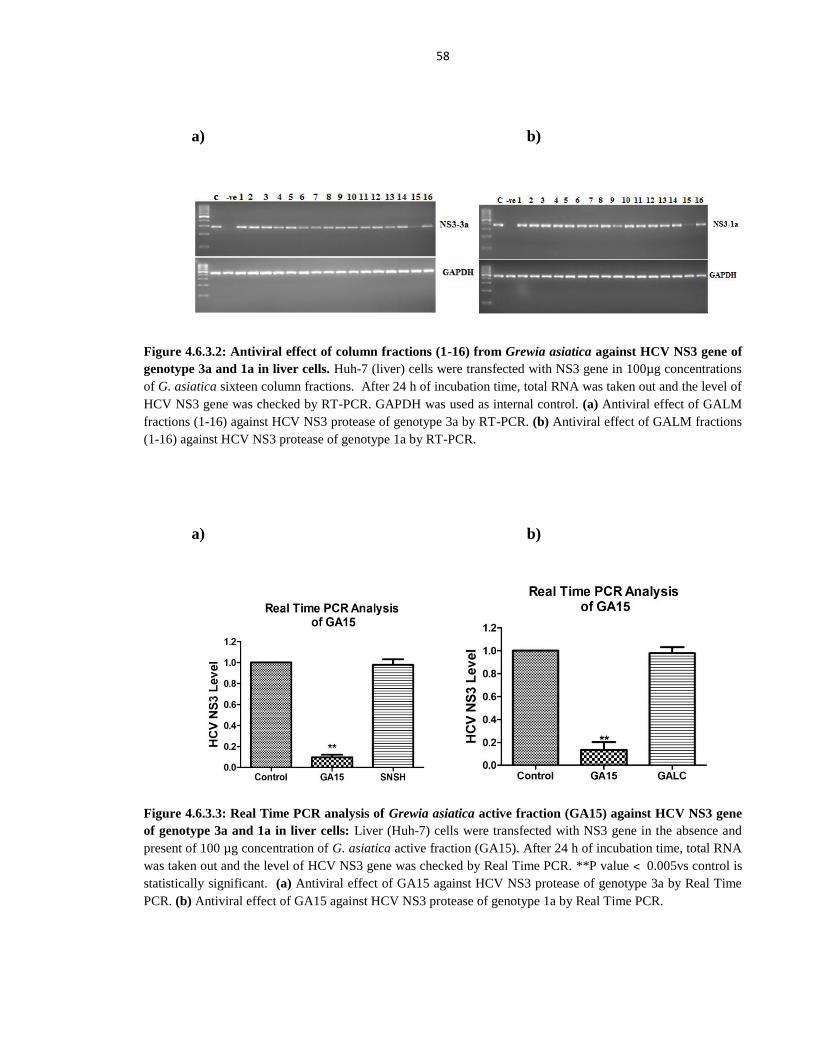
58
a) b)
Figure 4.6.3.2: Antiviral effect of column fractions (1-16) from Grewia asiatica against HCV NS3 gene of
genotype 3a and 1a in liver cells. Huh-7 (liver) cells were transfected with NS3 gene in 100µg concentrations
of G. asiatica sixteen column fractions. After 24 h of incubation time, total RNA was taken out and the level of
HCV NS3 gene was checked by RT-PCR. GAPDH was used as internal control. (a) Antiviral effect of GALM
fractions (1-16) against HCV NS3 protease of genotype 3a by RT-PCR. (b) Antiviral effect of GALM fractions
(1-16) against HCV NS3 protease of genotype 1a by RT-PCR.
a) b)
Figure 4.6.3.3: Real Time PCR analysis of Grewia asiatica active fraction (GA15) against HCV NS3 gene
of genotype 3a and 1a in liver cells: Liver (Huh-7) cells were transfected with NS3 gene in the absence and
present of 100 µg concentration of G. asiatica active fraction (GA15). After 24 h of incubation time, total RNA
was taken out and the level of HCV NS3 gene was checked by Real Time PCR. **P value ˂ 0.005vs control is
statistically significant. (a) Antiviral effect of GA15 against HCV NS3 protease of genotype 3a by Real Time
PCR. (b) Antiviral effect of GA15 against HCV NS3 protease of genotype 1a by Real Time PCR.

59
Additionally, the antiviral effect of G. asiatica active fraction (GA15) was examined
against nonstructural gene by transfection of NS3 gene in liver cells. The lysates from the
Huh-7 cells transfected with HCV-NS3 gene were studied by western blot, by using HCV
NS3 specific antibodies and GAPDH served as internal control. There was a remarkable
decrease in HCV NS3 protein level in cells treated with G. asiatica active fraction (GA15)
where as the GAPDH protein expression level remained same in control as compare to
treated cells (Figure 4.6.3.4).
Figure 4.6.3.4: HCV NS3 gene inhibition by G. asiatica at protein level. Protein expression level was
determined by transfection of Huh-7 cells with NS3 gene in the presence of G. asiatica leaves chloroform
(GALC) extract and active fraction (GA15). After 48 h of incubation time, protein was extracted and analyzed
by western blot with HCV NS3 monoclonal antibody and GAPDH monoclonal antibody served as internal
control.
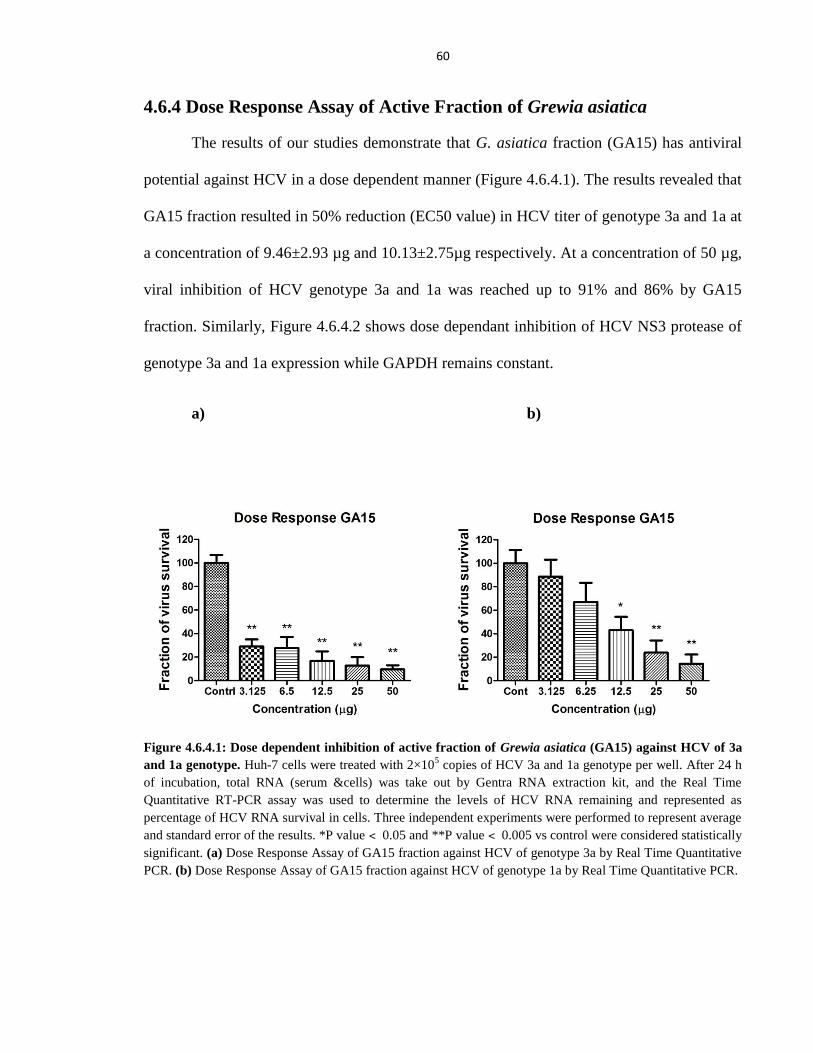
60
4.6.4 Dose Response Assay of Active Fraction of Grewia asiatica
The results of our studies demonstrate that G. asiatica fraction (GA15) has antiviral
potential against HCV in a dose dependent manner (Figure 4.6.4.1). The results revealed that
GA15 fraction resulted in 50% reduction (EC50 value) in HCV titer of genotype 3a and 1a at
a concentration of 9.46±2.93 µg and 10.13±2.75µg respectively. At a concentration of 50 µg,
viral inhibition of HCV genotype 3a and 1a was reached up to 91% and 86% by GA15
fraction. Similarly, Figure 4.6.4.2 shows dose dependant inhibition of HCV NS3 protease of
genotype 3a and 1a expression while GAPDH remains constant.
a) b)
Figure 4.6.4.1: Dose dependent inhibition of active fraction of Grewia asiatica (GA15) against HCV of 3a
and 1a genotype. Huh-7 cells were treated with 2×105 copies of HCV 3a and 1a genotype per well. After 24 h
of incubation, total RNA (serum &cells) was take out by Gentra RNA extraction kit, and the Real Time
Quantitative RT-PCR assay was used to determine the levels of HCV RNA remaining and represented as
percentage of HCV RNA survival in cells. Three independent experiments were performed to represent average
and standard error of the results. *P value ˂ 0.05 and **P value ˂ 0.005 vs control were considered statistically
significant. (a) Dose Response Assay of GA15 fraction against HCV of genotype 3a by Real Time Quantitative
PCR. (b) Dose Response Assay of GA15 fraction against HCV of genotype 1a by Real Time Quantitative PCR.
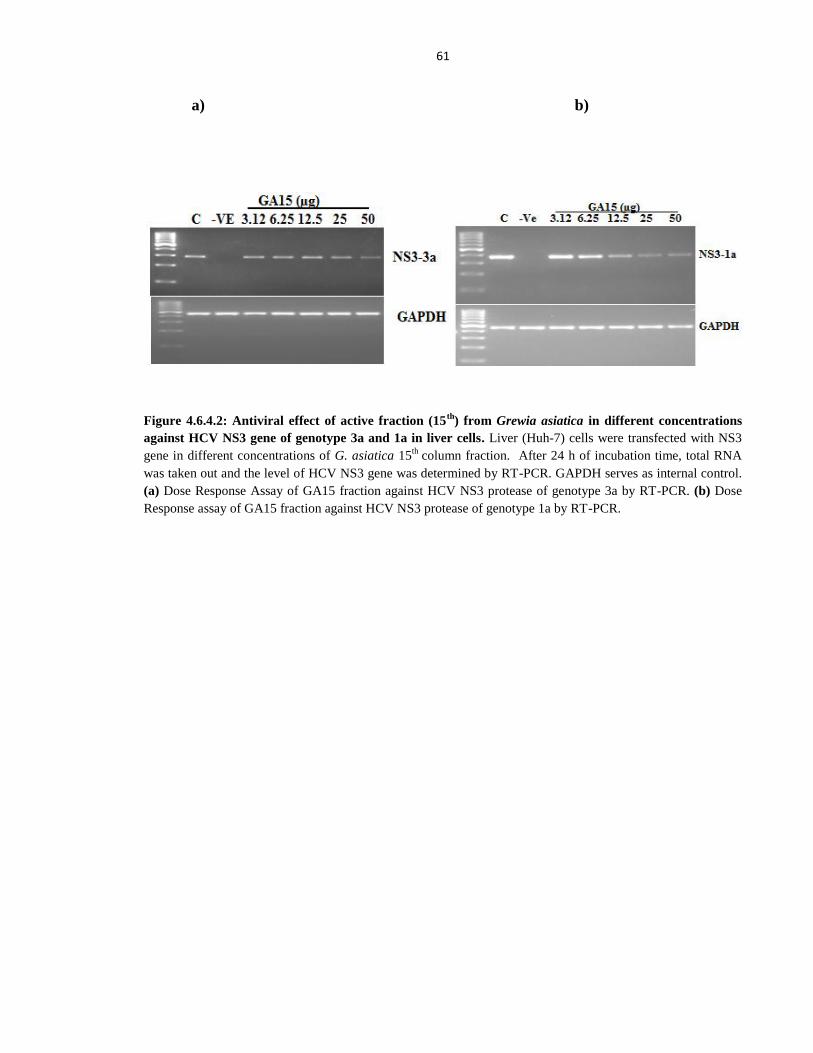
61
a) b)
Figure 4.6.4.2: Antiviral effect of active fraction (15th
) from Grewia asiatica in different concentrations
against HCV NS3 gene of genotype 3a and 1a in liver cells. Liver (Huh-7) cells were transfected with NS3
gene in different concentrations of G. asiatica 15th
column fraction. After 24 h of incubation time, total RNA
was taken out and the level of HCV NS3 gene was determined by RT-PCR. GAPDH serves as internal control.
(a) Dose Response Assay of GA15 fraction against HCV NS3 protease of genotype 3a by RT-PCR. (b) Dose
Response assay of GA15 fraction against HCV NS3 protease of genotype 1a by RT-PCR.
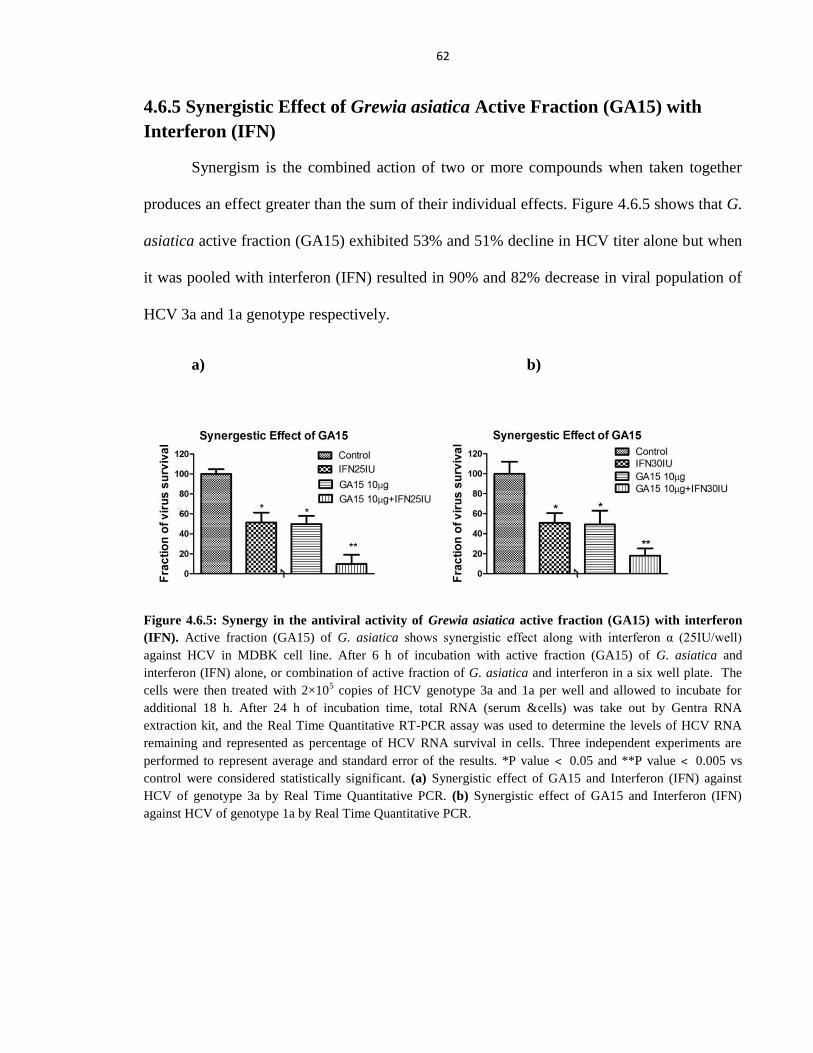
62
4.6.5 Synergistic Effect of Grewia asiatica Active Fraction (GA15) with
Interferon (IFN)
Synergism is the combined action of two or more compounds when taken together
produces an effect greater than the sum of their individual effects. Figure 4.6.5 shows that G.
asiatica active fraction (GA15) exhibited 53% and 51% decline in HCV titer alone but when
it was pooled with interferon (IFN) resulted in 90% and 82% decrease in viral population of
HCV 3a and 1a genotype respectively.
a) b)
Figure 4.6.5: Synergy in the antiviral activity of Grewia asiatica active fraction (GA15) with interferon
(IFN). Active fraction (GA15) of G. asiatica shows synergistic effect along with interferon α (25IU/well)
against HCV in MDBK cell line. After 6 h of incubation with active fraction (GA15) of G. asiatica and
interferon (IFN) alone, or combination of active fraction of G. asiatica and interferon in a six well plate. The
cells were then treated with 2×105 copies of HCV genotype 3a and 1a per well and allowed to incubate for
additional 18 h. After 24 h of incubation time, total RNA (serum &cells) was take out by Gentra RNA
extraction kit, and the Real Time Quantitative RT-PCR assay was used to determine the levels of HCV RNA
remaining and represented as percentage of HCV RNA survival in cells. Three independent experiments are
performed to represent average and standard error of the results. *P value ˂ 0.05 and **P value ˂ 0.005 vs
control were considered statistically significant. (a) Synergistic effect of GA15 and Interferon (IFN) against
HCV of genotype 3a by Real Time Quantitative PCR. (b) Synergistic effect of GA15 and Interferon (IFN)
against HCV of genotype 1a by Real Time Quantitative PCR.
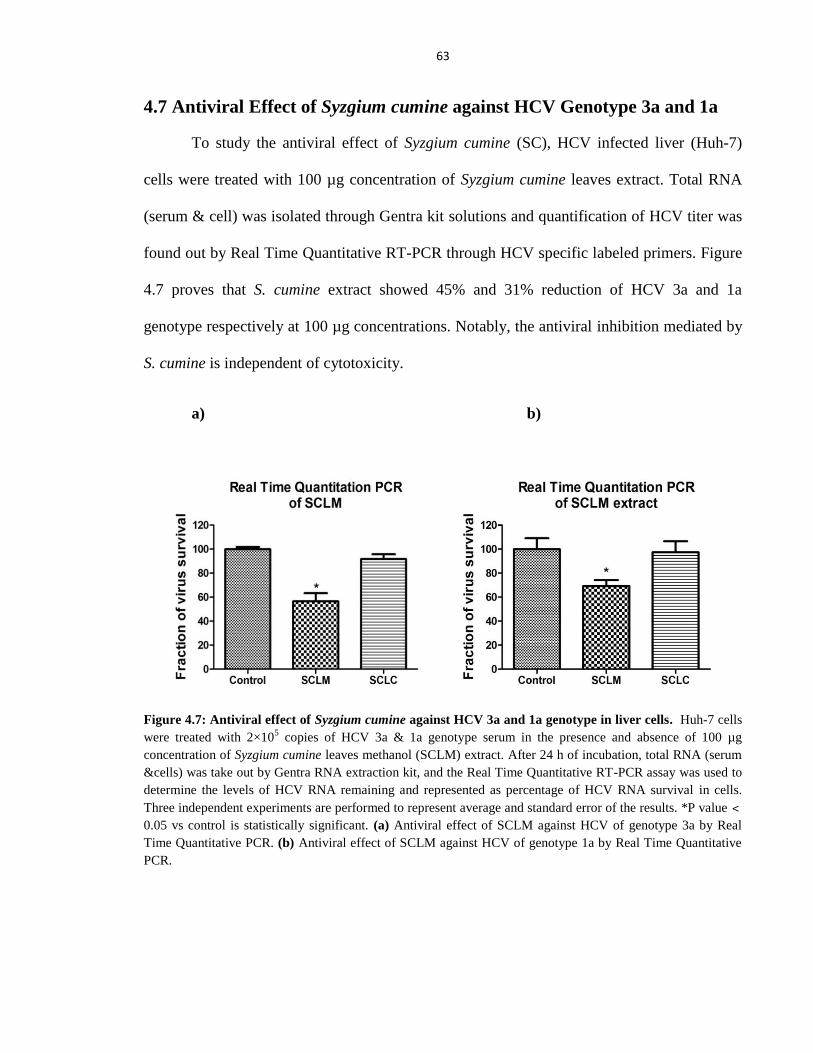
63
4.7 Antiviral Effect of Syzgium cumine against HCV Genotype 3a and 1a
To study the antiviral effect of Syzgium cumine (SC), HCV infected liver (Huh-7)
cells were treated with 100 µg concentration of Syzgium cumine leaves extract. Total RNA
(serum & cell) was isolated through Gentra kit solutions and quantification of HCV titer was
found out by Real Time Quantitative RT-PCR through HCV specific labeled primers. Figure
4.7 proves that S. cumine extract showed 45% and 31% reduction of HCV 3a and 1a
genotype respectively at 100 µg concentrations. Notably, the antiviral inhibition mediated by
S. cumine is independent of cytotoxicity.
a) b)
Figure 4.7: Antiviral effect of Syzgium cumine against HCV 3a and 1a genotype in liver cells. Huh-7 cells
were treated with 2×105 copies of HCV 3a & 1a genotype serum in the presence and absence of 100 µg
concentration of Syzgium cumine leaves methanol (SCLM) extract. After 24 h of incubation, total RNA (serum
&cells) was take out by Gentra RNA extraction kit, and the Real Time Quantitative RT-PCR assay was used to
determine the levels of HCV RNA remaining and represented as percentage of HCV RNA survival in cells.
Three independent experiments are performed to represent average and standard error of the results. *P value ˂
0.05 vs control is statistically significant. (a) Antiviral effect of SCLM against HCV of genotype 3a by Real
Time Quantitative PCR. (b) Antiviral effect of SCLM against HCV of genotype 1a by Real Time Quantitative
PCR.

64
4.7.1 Antiviral Effect of Syzgium cumine against HCV-NS3 Proteases of
Genotype 3a and 1a
To evaluate the antiviral effect of Syzgium cumine (SC) against HCV NS3 proteases,
liver (Huh-7) cells were harvested in six well culture plates. After the incubation period, cells
were propagated, RNA was isolated and complementary DNA (cDNA) was produced by
ologo dT priming. PCR was used to amplify cDNA using NS3-3a and NS3-1a gene specific
primers of HCV. The amplification of GAPDH messenger RNA is used as internal control.
The result of this study demonstrates that methanolic extract of S. cumine (SCLM) reduces
HCV RNA expression level extensively at a concentration of 100 µg, while the expression of
GAPDH mRNA is not affected by the addition of the extract (Figure 4.7.1.1). Figure 4.7.1.3
showed that S. cumine methanol (SCLM) extract reduced the expression of HCV-NS3
proteases while chloroform (SCLC) and acetone (SCLA) extracts had no effect on HCV-NS3
proteases in Huh-7 cells at a concentration of 100 µg. Similarly, Real Time PCR results
exhibits that SCLM extract resulted in 48% and 34% inhibition of HCV NS3 proteases level
of genotype 3a and 1a respectively at non toxic concentrations (Figure 4.7.1.2). Figure
4.7.1.4 demonstrates that SCLC, SCLA and SCLM extracts inhibits 6%, 9%, 51% of NS3
protease level of genotype 3a and 3%, 0%, 79% of genotype 1a respectively. Collectively,
the results showed that phytochemicals present in medicinal plants significantly inhibited
HCV-NS3 protease expression in Huh-7 cells. Importantly, the antiviral inhibition mediated
by these plant extracts is independent of cytotoxicity.
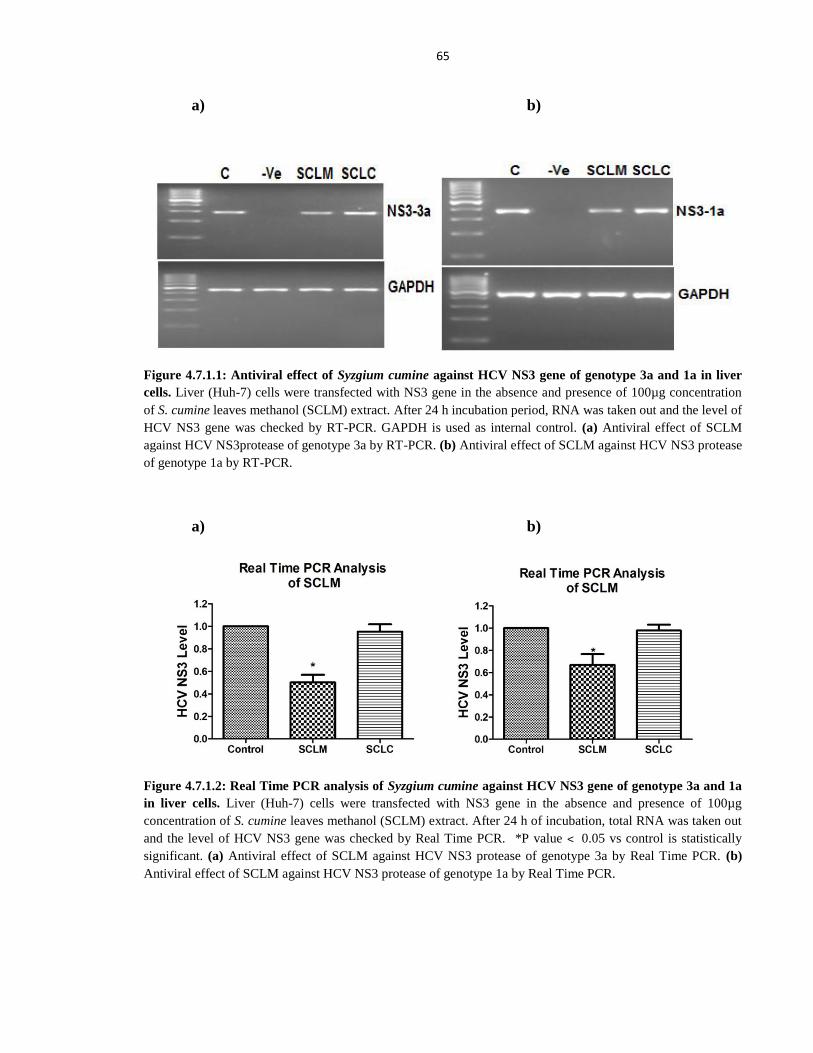
65
a) b)
Figure 4.7.1.1: Antiviral effect of Syzgium cumine against HCV NS3 gene of genotype 3a and 1a in liver
cells. Liver (Huh-7) cells were transfected with NS3 gene in the absence and presence of 100µg concentration
of S. cumine leaves methanol (SCLM) extract. After 24 h incubation period, RNA was taken out and the level of
HCV NS3 gene was checked by RT-PCR. GAPDH is used as internal control. (a) Antiviral effect of SCLM
against HCV NS3protease of genotype 3a by RT-PCR. (b) Antiviral effect of SCLM against HCV NS3 protease
of genotype 1a by RT-PCR.
a) b)
Figure 4.7.1.2: Real Time PCR analysis of Syzgium cumine against HCV NS3 gene of genotype 3a and 1a
in liver cells. Liver (Huh-7) cells were transfected with NS3 gene in the absence and presence of 100µg
concentration of S. cumine leaves methanol (SCLM) extract. After 24 h of incubation, total RNA was taken out
and the level of HCV NS3 gene was checked by Real Time PCR. *P value ˂ 0.05 vs control is statistically
significant. (a) Antiviral effect of SCLM against HCV NS3 protease of genotype 3a by Real Time PCR. (b)
Antiviral effect of SCLM against HCV NS3 protease of genotype 1a by Real Time PCR.
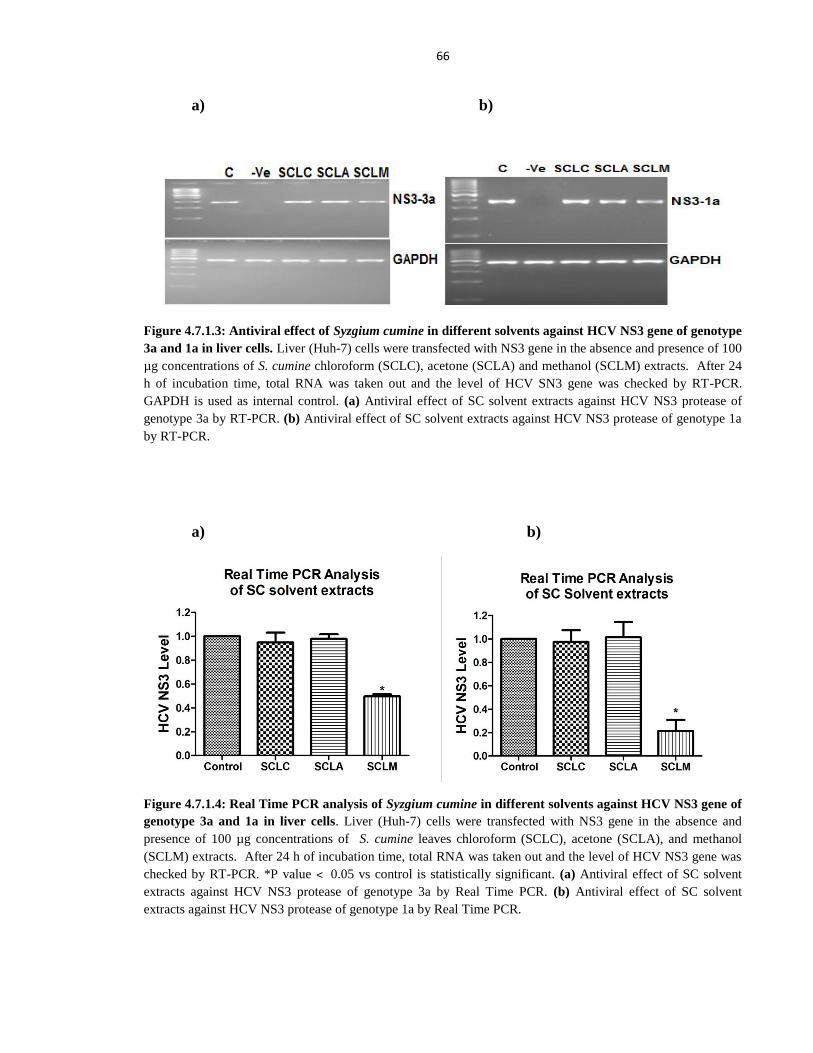
66
a) b)
Figure 4.7.1.3: Antiviral effect of Syzgium cumine in different solvents against HCV NS3 gene of genotype
3a and 1a in liver cells. Liver (Huh-7) cells were transfected with NS3 gene in the absence and presence of 100
µg concentrations of S. cumine chloroform (SCLC), acetone (SCLA) and methanol (SCLM) extracts. After 24
h of incubation time, total RNA was taken out and the level of HCV SN3 gene was checked by RT-PCR.
GAPDH is used as internal control. (a) Antiviral effect of SC solvent extracts against HCV NS3 protease of
genotype 3a by RT-PCR. (b) Antiviral effect of SC solvent extracts against HCV NS3 protease of genotype 1a
by RT-PCR.
a) b)
Figure 4.7.1.4: Real Time PCR analysis of Syzgium cumine in different solvents against HCV NS3 gene of
genotype 3a and 1a in liver cells. Liver (Huh-7) cells were transfected with NS3 gene in the absence and
presence of 100 µg concentrations of S. cumine leaves chloroform (SCLC), acetone (SCLA), and methanol
(SCLM) extracts. After 24 h of incubation time, total RNA was taken out and the level of HCV NS3 gene was
checked by RT-PCR. *P value ˂ 0.05 vs control is statistically significant. (a) Antiviral effect of SC solvent
extracts against HCV NS3 protease of genotype 3a by Real Time PCR. (b) Antiviral effect of SC solvent
extracts against HCV NS3 protease of genotype 1a by Real Time PCR.
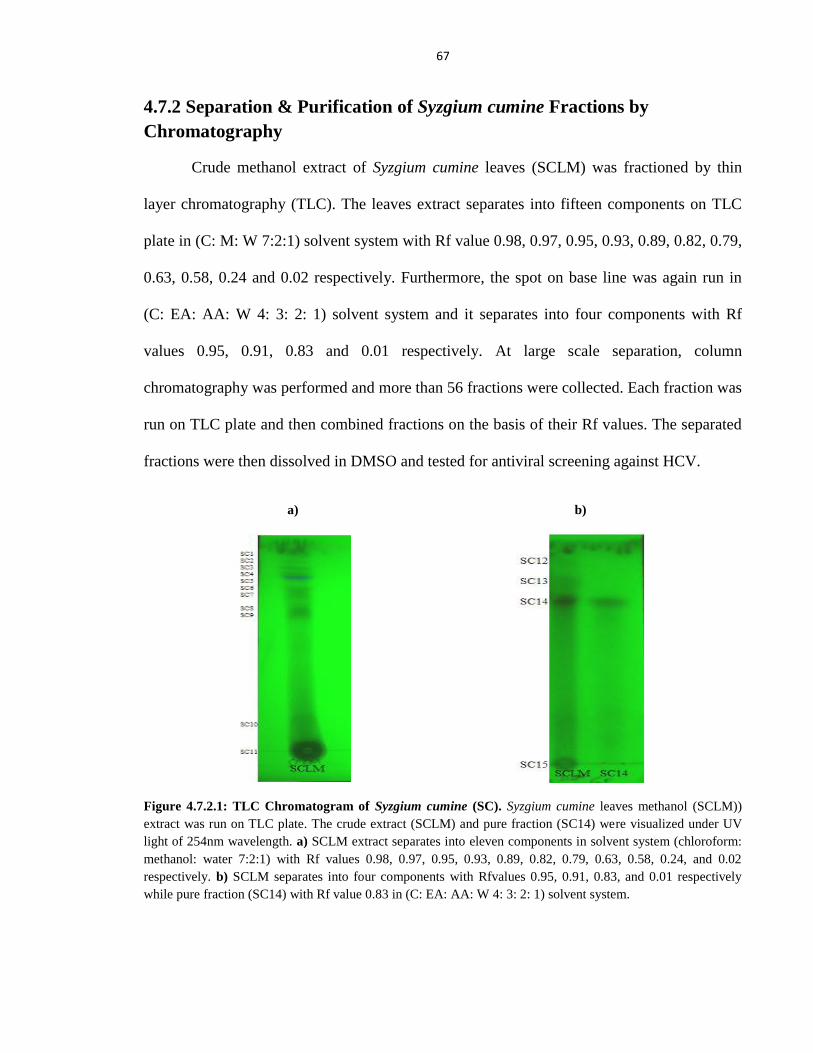
67
4.7.2 Separation & Purification of Syzgium cumine Fractions by
Chromatography
Crude methanol extract of Syzgium cumine leaves (SCLM) was fractioned by thin
layer chromatography (TLC). The leaves extract separates into fifteen components on TLC
plate in (C: M: W 7:2:1) solvent system with Rf value 0.98, 0.97, 0.95, 0.93, 0.89, 0.82, 0.79,
0.63, 0.58, 0.24 and 0.02 respectively. Furthermore, the spot on base line was again run in
(C: EA: AA: W 4: 3: 2: 1) solvent system and it separates into four components with Rf
values 0.95, 0.91, 0.83 and 0.01 respectively. At large scale separation, column
chromatography was performed and more than 56 fractions were collected. Each fraction was
run on TLC plate and then combined fractions on the basis of their Rf values. The separated
fractions were then dissolved in DMSO and tested for antiviral screening against HCV.
a) b)
Figure 4.7.2.1: TLC Chromatogram of Syzgium cumine (SC). Syzgium cumine leaves methanol (SCLM))
extract was run on TLC plate. The crude extract (SCLM) and pure fraction (SC14) were visualized under UV
light of 254nm wavelength. a) SCLM extract separates into eleven components in solvent system (chloroform:
methanol: water 7:2:1) with Rf values 0.98, 0.97, 0.95, 0.93, 0.89, 0.82, 0.79, 0.63, 0.58, 0.24, and 0.02
respectively. b) SCLM separates into four components with Rfvalues 0.95, 0.91, 0.83, and 0.01 respectively
while pure fraction (SC14) with Rf value 0.83 in (C: EA: AA: W 4: 3: 2: 1) solvent system.
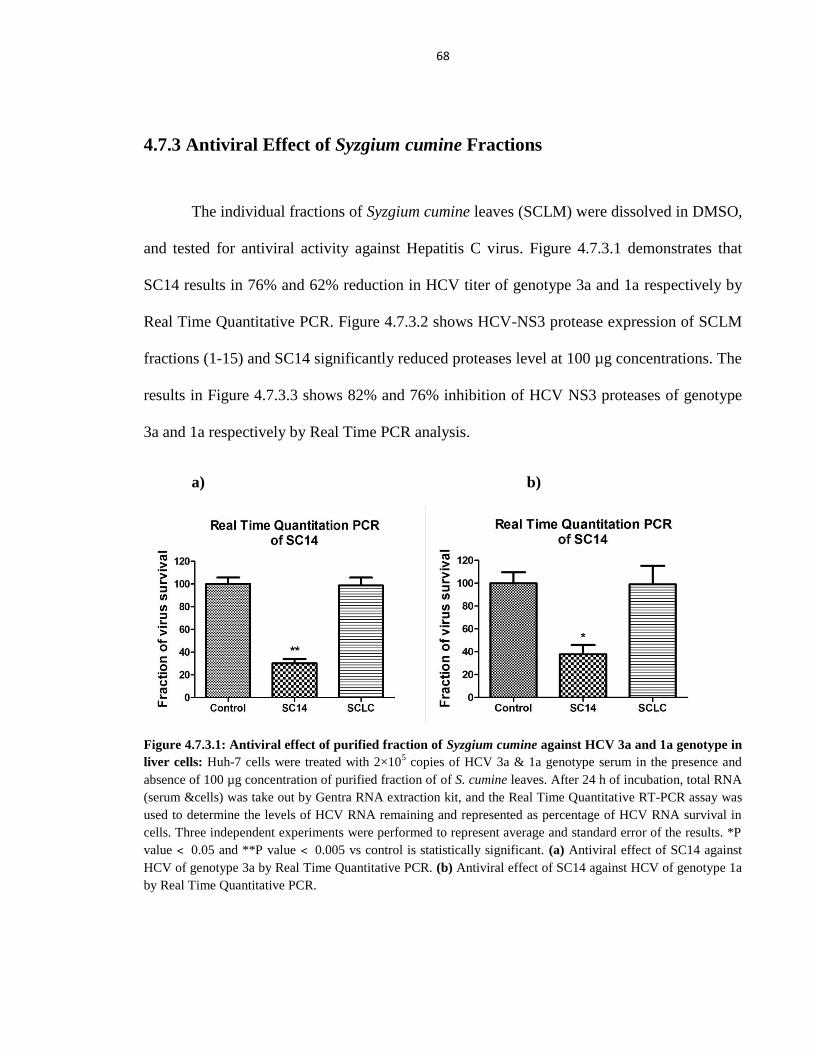
68
4.7.3 Antiviral Effect of Syzgium cumine Fractions
The individual fractions of Syzgium cumine leaves (SCLM) were dissolved in DMSO,
and tested for antiviral activity against Hepatitis C virus. Figure 4.7.3.1 demonstrates that
SC14 results in 76% and 62% reduction in HCV titer of genotype 3a and 1a respectively by
Real Time Quantitative PCR. Figure 4.7.3.2 shows HCV-NS3 protease expression of SCLM
fractions (1-15) and SC14 significantly reduced proteases level at 100 µg concentrations. The
results in Figure 4.7.3.3 shows 82% and 76% inhibition of HCV NS3 proteases of genotype
3a and 1a respectively by Real Time PCR analysis.
a) b)
Figure 4.7.3.1: Antiviral effect of purified fraction of Syzgium cumine against HCV 3a and 1a genotype in
liver cells: Huh-7 cells were treated with 2×105 copies of HCV 3a & 1a genotype serum in the presence and
absence of 100 µg concentration of purified fraction of of S. cumine leaves. After 24 h of incubation, total RNA
(serum &cells) was take out by Gentra RNA extraction kit, and the Real Time Quantitative RT-PCR assay was
used to determine the levels of HCV RNA remaining and represented as percentage of HCV RNA survival in
cells. Three independent experiments were performed to represent average and standard error of the results. *P
value ˂ 0.05 and **P value ˂ 0.005 vs control is statistically significant. (a) Antiviral effect of SC14 against
HCV of genotype 3a by Real Time Quantitative PCR. (b) Antiviral effect of SC14 against HCV of genotype 1a
by Real Time Quantitative PCR.
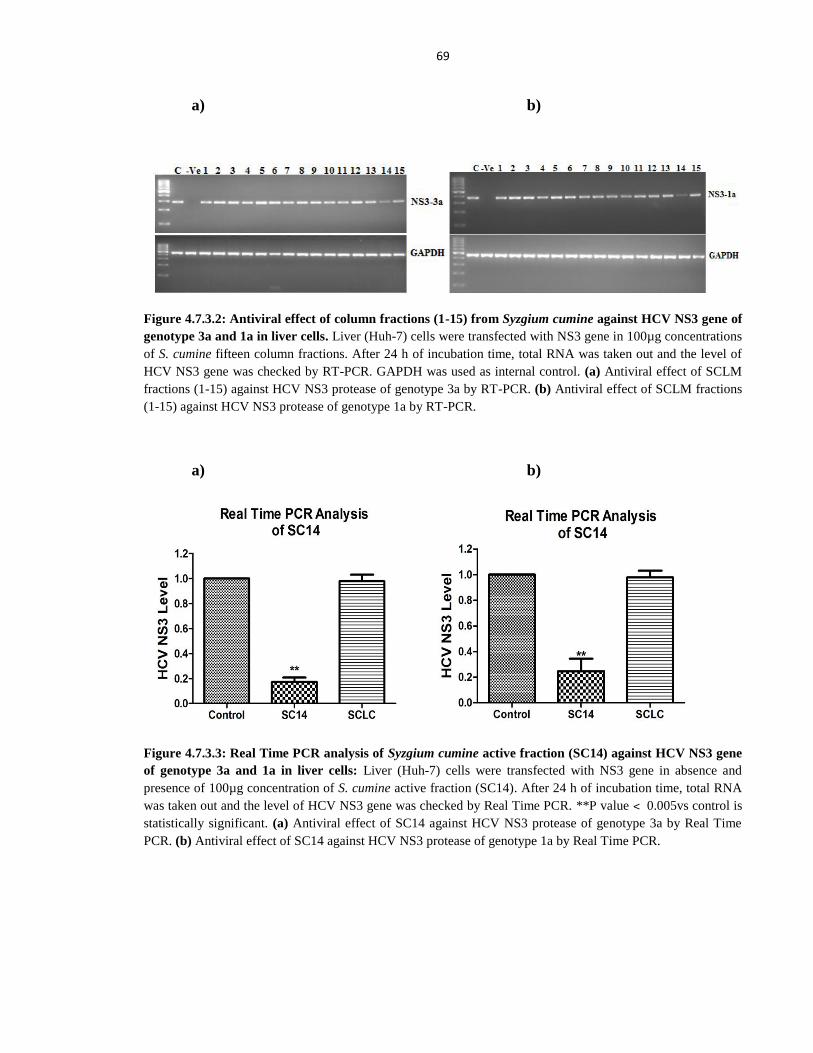
69
a) b)
Figure 4.7.3.2: Antiviral effect of column fractions (1-15) from Syzgium cumine against HCV NS3 gene of
genotype 3a and 1a in liver cells. Liver (Huh-7) cells were transfected with NS3 gene in 100µg concentrations
of S. cumine fifteen column fractions. After 24 h of incubation time, total RNA was taken out and the level of
HCV NS3 gene was checked by RT-PCR. GAPDH was used as internal control. (a) Antiviral effect of SCLM
fractions (1-15) against HCV NS3 protease of genotype 3a by RT-PCR. (b) Antiviral effect of SCLM fractions
(1-15) against HCV NS3 protease of genotype 1a by RT-PCR.
a) b)
Figure 4.7.3.3: Real Time PCR analysis of Syzgium cumine active fraction (SC14) against HCV NS3 gene
of genotype 3a and 1a in liver cells: Liver (Huh-7) cells were transfected with NS3 gene in absence and
presence of 100µg concentration of S. cumine active fraction (SC14). After 24 h of incubation time, total RNA
was taken out and the level of HCV NS3 gene was checked by Real Time PCR. **P value ˂ 0.005vs control is
statistically significant. (a) Antiviral effect of SC14 against HCV NS3 protease of genotype 3a by Real Time
PCR. (b) Antiviral effect of SC14 against HCV NS3 protease of genotype 1a by Real Time PCR.

70
Additionally, the antiviral effect of S. cumine active fraction (SC14) was analyzed
against nonstructural gene by transfection of HCV-NS3 gene in liver cells. The lysates from
the Huh-7 cells transfected with HCV NS3 gene were studied by western blot, by using
HCV-NS3 specific antibodies and GAPDH served as internal control. There was a
remarkable decrease in HCV NS3 protein level in cells treated with S. cumine active fraction
(SC14) where as the GAPDH protein expression level remained same in control as compare
to treated cells (Figure 4.7.3.4).
Figure 4.7.3.4: HCV NS3 gene inhibition by S. cumine at protein level. Protein expression level was
determined by transfection of Huh-7 cells with NS3 gene in presence of S. cumine leaves chloroform (SCLC)
extract and active fraction (SC14). After 48 h of incubation time, protein was extracted and analyzed by western
blot with HCV NS3 monoclonal antibody and GAPDH monoclonal antibody served as internal control.
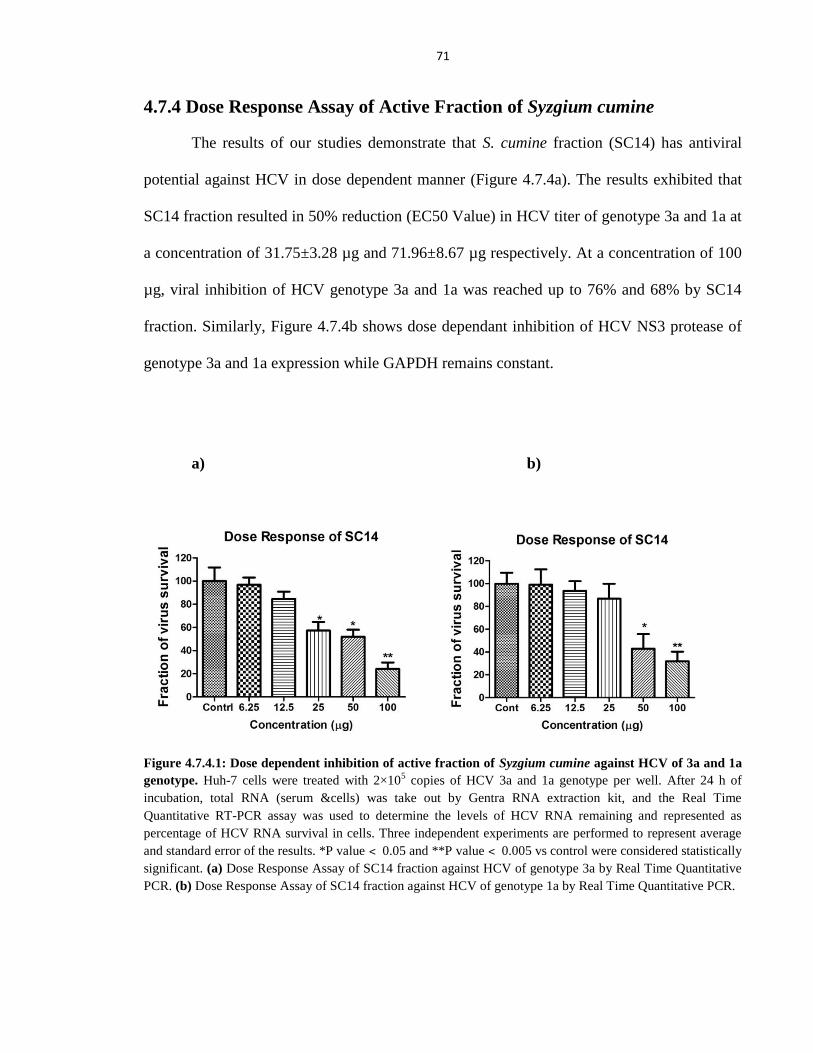
71
4.7.4 Dose Response Assay of Active Fraction of Syzgium cumine
The results of our studies demonstrate that S. cumine fraction (SC14) has antiviral
potential against HCV in dose dependent manner (Figure 4.7.4a). The results exhibited that
SC14 fraction resulted in 50% reduction (EC50 Value) in HCV titer of genotype 3a and 1a at
a concentration of 31.75±3.28 µg and 71.96±8.67 µg respectively. At a concentration of 100
µg, viral inhibition of HCV genotype 3a and 1a was reached up to 76% and 68% by SC14
fraction. Similarly, Figure 4.7.4b shows dose dependant inhibition of HCV NS3 protease of
genotype 3a and 1a expression while GAPDH remains constant.
a) b)
Figure 4.7.4.1: Dose dependent inhibition of active fraction of Syzgium cumine against HCV of 3a and 1a
genotype. Huh-7 cells were treated with 2×105 copies of HCV 3a and 1a genotype per well. After 24 h of
incubation, total RNA (serum &cells) was take out by Gentra RNA extraction kit, and the Real Time
Quantitative RT-PCR assay was used to determine the levels of HCV RNA remaining and represented as
percentage of HCV RNA survival in cells. Three independent experiments are performed to represent average
and standard error of the results. *P value ˂ 0.05 and **P value ˂ 0.005 vs control were considered statistically
significant. (a) Dose Response Assay of SC14 fraction against HCV of genotype 3a by Real Time Quantitative
PCR. (b) Dose Response Assay of SC14 fraction against HCV of genotype 1a by Real Time Quantitative PCR.

72
a) b)
Figure 4.7.4.2: Antiviral effect of active column fraction (14th
) from Syzgium cumine in different
concentrations against HCV NS3 gene of genotype 3a and 1a in liver cells. Liver (Huh-7) cells were
transfected with NS3 gene in different concentrations of Syzgium cumine 14th
column fraction. After 24 h of
incubation time, total RNA was taken out and the level of HCV NS3 gene was determined by RT-PCR.
GAPDH serves as internal control. (a) Dose Response Assay of SC14 fraction against HCV NS3 protease of
genotype 3a by RT-PCR. (b) Dose Response Assay of SC14 fraction against HCV NS3 protease of genotype 1a
by RT-PCR.
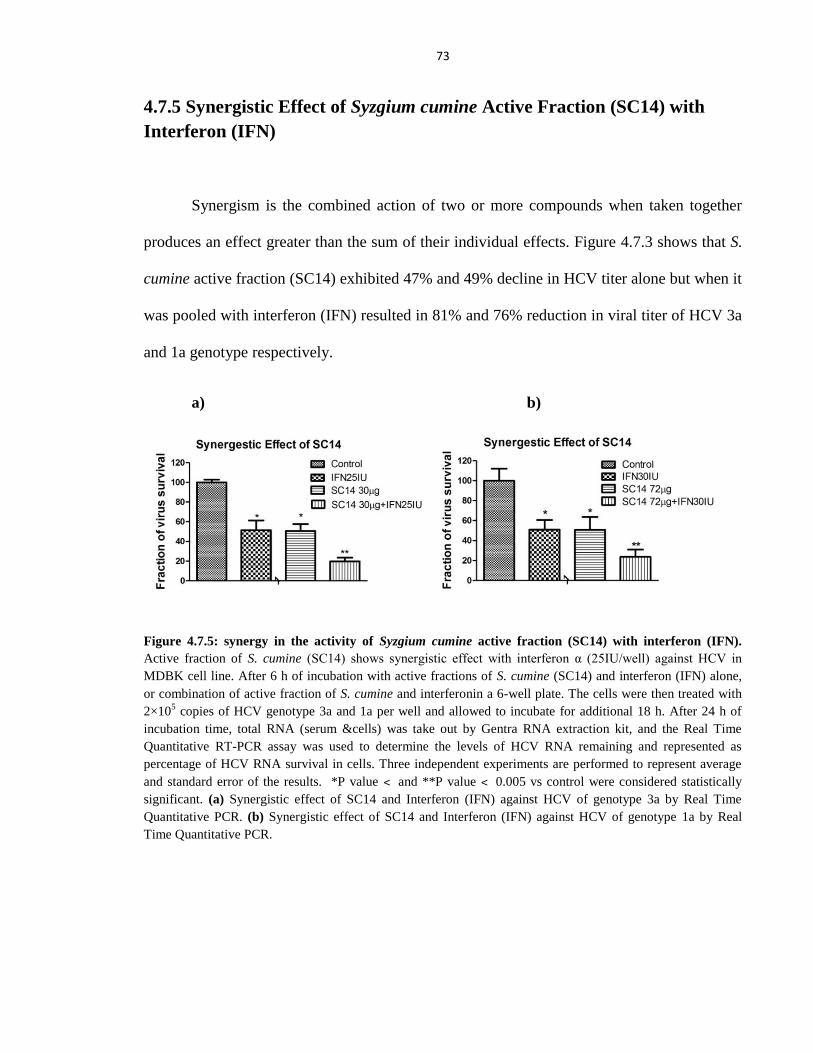
73
4.7.5 Synergistic Effect of Syzgium cumine Active Fraction (SC14) with
Interferon (IFN)
Synergism is the combined action of two or more compounds when taken together
produces an effect greater than the sum of their individual effects. Figure 4.7.3 shows that S.
cumine active fraction (SC14) exhibited 47% and 49% decline in HCV titer alone but when it
was pooled with interferon (IFN) resulted in 81% and 76% reduction in viral titer of HCV 3a
and 1a genotype respectively.
a) b)
Figure 4.7.5: synergy in the activity of Syzgium cumine active fraction (SC14) with interferon (IFN).
Active fraction of S. cumine (SC14) shows synergistic effect with interferon α (25IU/well) against HCV in
MDBK cell line. After 6 h of incubation with active fractions of S. cumine (SC14) and interferon (IFN) alone,
or combination of active fraction of S. cumine and interferonin a 6-well plate. The cells were then treated with
2×105 copies of HCV genotype 3a and 1a per well and allowed to incubate for additional 18 h. After 24 h of
incubation time, total RNA (serum &cells) was take out by Gentra RNA extraction kit, and the Real Time
Quantitative RT-PCR assay was used to determine the levels of HCV RNA remaining and represented as
percentage of HCV RNA survival in cells. Three independent experiments are performed to represent average
and standard error of the results. *P value ˂ and **P value ˂ 0.005 vs control were considered statistically
significant. (a) Synergistic effect of SC14 and Interferon (IFN) against HCV of genotype 3a by Real Time
Quantitative PCR. (b) Synergistic effect of SC14 and Interferon (IFN) against HCV of genotype 1a by Real
Time Quantitative PCR.
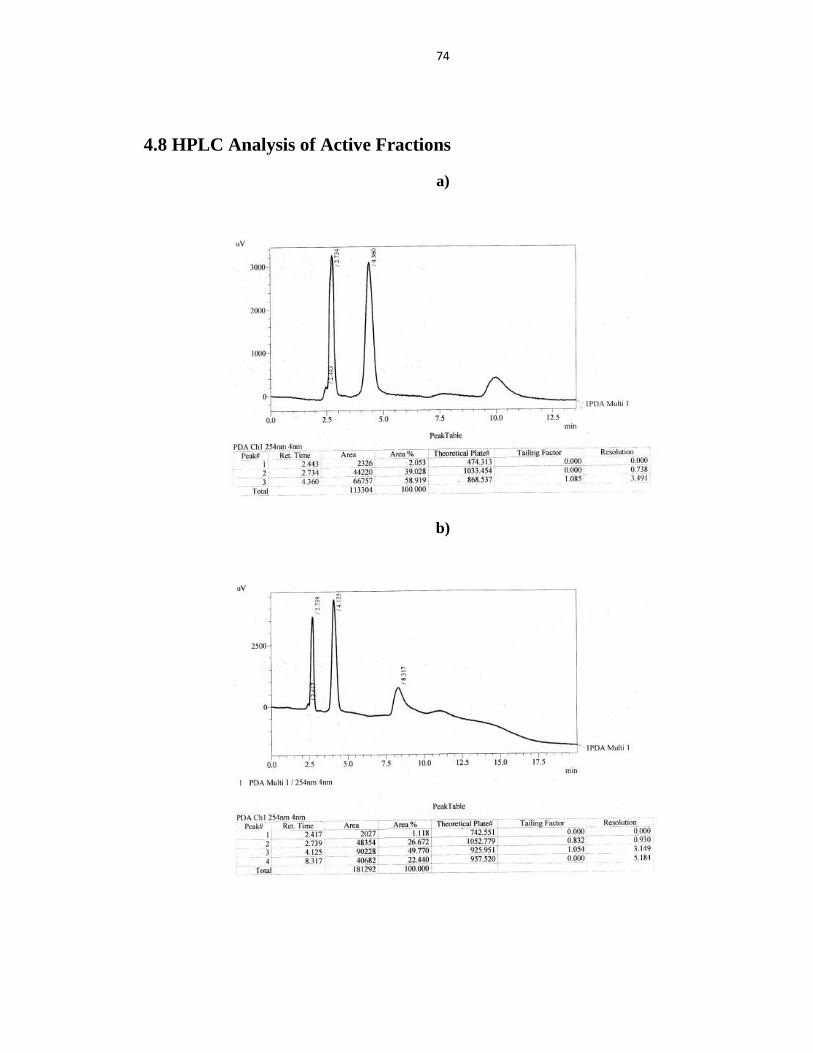
74
4.8 HPLC Analysis of Active Fractions
a)
b)
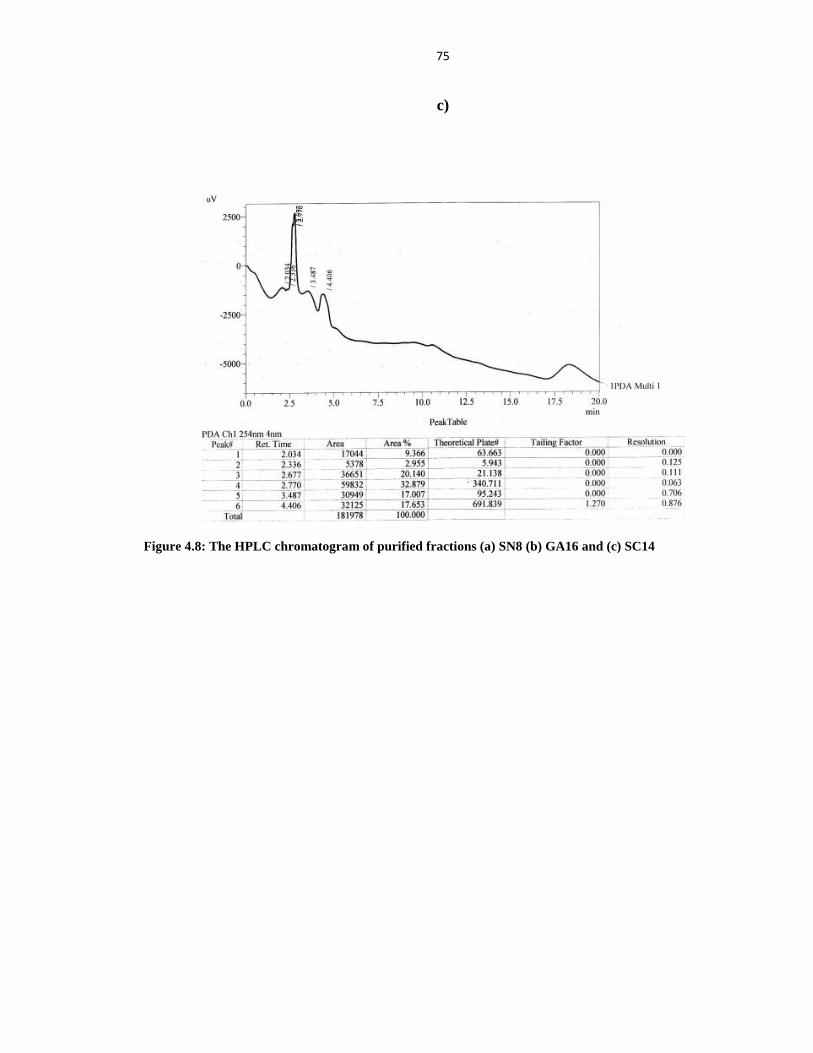
75
c)
Figure 4.8: The HPLC chromatogram of purified fractions (a) SN8 (b) GA16 and (c) SC14

76
DISCUSSION
Medicinal plants are used for different ailments throughout the world. In Pakistan a
large number of indigenous populations utilize medicinal plants to fulfill their health care
needs. As medicinal plants are cheaper and easily accessible, local people prefer herbs
against different diseases. In present study different plants extracts were examined through
trypan blue dye and MTT cell proliferation assay for cytotoxic studies in fibroblast and liver
cells. Previous data showed cytotoxic activity of plant extracts in two cancer cell lines and
all the extracts have cytotoxic effect on HeLa (Human cervix cancer) and T47D (human
breast cancer) cells (Vega-Avila et al., 2009). Some plant's methanolic extracts revealed low
or no cytotoxicity against the MCF7, HepG2 and MDBK cell lines, whereas F. szowitsiana
showed the most potent cytotoxicity against all of them (Sahranavard et al., 2009). In our
studies, extracts presented the best cytotoxic effect in Huh-7 cells correspond to a potential
for cancer treatment (Figure 4.2). The extracts that were nontoxic to Huh-7 cells were
included for screening against Hepatitis C virus.
In present study, Twenty four medicinal plants were screened against HCV. Before
antiviral screening, the extracts were examined for toxicological effects, if any, in fibroblast
and liver cells through trypan blue dye exclusion method and results indicated that extracts
NJRM, PNSM and VJAD were toxic to hepatoma cells up to 40µg concentration while the
remaining extracts were non toxic to cells at 100µg of concentrations (Data not shown).
These results were further verified by MTT cell proliferation assay (Mosmann, 1983b) which
showed that extracts NJRM, PNSM and VJAD were toxic to liver cells (Figure 4.2) so, did
not permit them for further screening against HCV. Trypan blue dye method showed that
SNSM, GALM and SCLM extracts are nontoxic to Huh-7 and CHO cells at 100 µg

77
concentrations (Figure 4.2.2). Similarly, MTT assay proved that SNSM, GALM and SCLM
extracts are nontoxic to Huh-7 cells at a concentration of 100µg (Figure 4.3).
Subsequently, antiviral activities of all nontoxic extracts were investigated against
HCV by infecting liver cells with infectious viral particles. The present study showed that
five out of twenty one plant extracts exhibited anti-HCV effect. These extracts were
designated as Solanum nigrum (SN), Grewia asiatica (GA), Syzygium cumini (SC),
Avicennia marina (AM) and Fagonia cretica (FC). Interestingly, SNSM, GALM, SCLM,
FCAM and AMLM inhibited HCV 3a and 1a genotype and did not show appreciable toxicity
against Huh-7 and CHO cells (Figure 4.4). Hence, these extracts used for further cell culture
studies against HCV.
Solanum nigrum traitionaly named as Makoi or black nightshade, is a weed that
grows in variety of habitats and it belongs to family solanaceae (Kiran et al., 2009). Mostly
S.nigrum is used conventionally to treat a range of diseases which includes inflammation,
(Acharya and Pokhrel, 2006) dressing of warts (Moshi et al., 2009), erysipelas (acute
streptococcus bacterial infection) (Leporatti and Ghedira, 2009), stomach ulcer (Sivaperumal
et al., 2010), liver tonic, indigestion (Kala, 2005), increase fertility in women (Singh et al.,
2010) and treating asthma and whooping cough (Sikdar and Dutta, 2008).
Both the crude extract of S. nigrum and purified components showed antiproliferative
action on different cancer cell lines. The antiproliferative activity of crude plant extract and
isolated components were examined on cancerous liver cell line (HepG2) and colon (HT29
and HCT-116) (Ji et al., 2008; Lee et al., 2004), breast cancer (MCF-7) (Li et al., 2009),
cervical cancer cell line (U14) (Joo et al., 2009; Li et al., 2008) and HeLa cells (Son et al.,

78
2003). The results of this study demonstrated that SNSM extract is non toxic to Huh-7 and
CHO cells in trypan blue dye exclusion method (Figure 4.1) and MTT cell viability assay
(Figure 4.3) at 100 µg concentrations.
Traditionally used many medicinal plants and herbal formulations like Glycyrrhiza
glabra which was used in traditional medicine across the globe, reported to have strong
antiviral activity against RNA and DNA viruses (Saxena, 2005). In addition, the S. nigrum
crude extract declined the eminent levels of plasma alanine aminotransferase (ALT) and
billirubin level to normal value (Hsieh et al., 2008). Present study clearly demonstrated that
at a concentration of 100 µg S. nigrum seeds extract showed 45% and 37% reduction of HCV
3a and 1a titer respectively (Figure 4.5). Clinical trials on polyherbal formulation like Liv52,
includes S. nigrum as one of the major components, explore its hepatoprotective action
(Debajyoti et al., 2012). Similarly, 50% of the tumor size in mice was reduced by aqueous
extract of S. nigrum plant (Wang et al., 2010).
In this study, we found that S. nigrum seeds chloroform (SNSC) and methanol
(SNSM) extracts reduced the expression of HCV NS3 proteases (Figure 4.5.1.3) and our Real
Time PCR results showed that SNSM extract demonstrated 52% and 43% inhibition of HCV
NS3 proteases level of genotype 3a and 1a respectively (Figure 4.5.1.2). Furthermore, S.
nigrum methanolic extract was dissolved in different solvents from lower to higher polarity
and found that SNSH, SNSC and SNSM extracts inhibits 6%, 50%, 61% of HCV NS3
protease level of genotype 3a and 0%, 41%, 68% of genotype 1a respectively at 100 µg
concentrations (Figure 4.5.1.4). So, S. nigrum seeds extract has a potential to decrease HCV
titer of genotype 3a and 1a, which is most prevalent in Pakistan and Africa respectively.

79
The previous literature showed the presence of carbohydrates, coumarins,
phytosterols and flavonoids in ethanolic and aqueous extracts of S. nigrum (Ravi et al.,
2009). Ayesha mohy-ud-din in 2009 studied thin layer chromatography (TLC) of SN extract
and found 6 components with Rf values 0.93, 0.83, 0.58, 0.55, 0.36 and 0.15 in n-
butanol:acetic acid:water, with ratio of 4:1:5 (UD-DIN et al., 2009). In another study, it was
found that SN leaf consists of 11 flavonoids and 8 phenolic acids (Huang et al., 2010). In our
thin layer chromatography data S. nigrum extract separates into thirteen components on TLC
plate in (Chloroform- Methanol- Water 7:2:1) mobile phase with Rf value 0.86, 0.80, 0.77,
0.64, 0.51, 0.44, 0.38, 0.29, 0.15, 0.13,0.10, 0.08, 0.05 respectively and pure fraction SN8
with Rf value 0.29 (Figure 4.5.2.1). Then separated fractions were dissolved in DMSO to
find out fraction active against HCV and resulted that HCV viral titer is decreased 76% and
65% of genotype 3a and 1a respectively (Figure 4.5.3.1).
NS3 protease is considered to be most attractive target for drug development
because of the fact that is essential for the replication of hepatitis C virus (Kolykhalov et al.,
2000; Reed and Rice, 2000). The Figure 4.5.3.2 proves that HCV NS3 level is significantly
decreased by SN8 fraction as compare to remaining S. nigrum fractions and Real Time PCR
analysis shows 85% and 83% inhibition of HCV NS3 level of genotype 3a and 1a
respectively (Figure 4.5.3.3). Previously, it was noted that a small molecule BILN 2061
belongs to a group of potent HCV NS3 protease inhibitors (Lamarre et al., 2003) which
support our results of SN8 fraction. Previously, the inhibition of polyprotein processing via
NS3 and reduction of protein expression by BLN 2061 was observed in replicon system

80
(Lamarre et al., 2003). Figure 4.5.3.4 clearly shows reduced NS3 expression by SN8 fraction
at protein level and GAPDH protein expression remains constant.
The results in Figure 4.5.4.1 shows that S. nigrum active fraction (SN8) has antiviral
effect against HCV of genotype 3a and 1a in a dose-dependent manner and at 100 µg
concentration inhibition of HCV titer upto 88% and 81% respectively. Furthermore, EC50
value was calculated by using non linear regression equation and demonstrated 50%
reduction in HCV titer of genotype 3a and 1a at 24.94±3.46µg and 47.68±5.73µg
respectively. In Figure 4.5.4.2 the results investigated that expression of HCV NS3 level
decreases as we increase the dose of SN8 while GAPDH expression remains same at all
doses. In this findings it is demonstrated that 51% and 49% decrease in viral titer was
observed by S. nigrum active fraction (SN8) alone but in combination therapy with
interferon (IFN) exhibited 84% and 75% decrease in HCV viral load of genotype 3a and 1a
respectively (Figure 4.5.5).
Grewia asiatica is locally called as Falsa, it is Southern Asian native shrub belongs to
species of Grewia. The fruit is of 6 to 12 mm diameter, with purple black color when ripped.
Falsa is used as traditional medicine in South Asian countries including Ayurvedic Indian
system. Its extract is considered very useful and an important ingredient of many herbal
formulations. (Poonam and Singh, 2009); (Gupta et al., 2010). In this study, our findings
showed that Grewia asiatica extract has antiviral potential against Hepatitis C virus (Figure
4.4).

81
The previous studies demonstrated that G. asiatica has antibacterial (Duraipandiyan
et al., 2006), anti-platelet (Kumar et al., 2012), antidiabetic, hypoglycemic, antifertility,
antipyretic and analgesic activity (Zia-Ul-Haq et al., 2013). The results from this study
proves that G. asiatica extract showed 42% and 39% reduction of HCV 3a and 1a genotype
respectively at non toxic concentration of 100 µg. On the other, (Bhanuprakash et al., 2008)
any antiviral drug active against virus should be non toxic to host cells. Importantly, the
decrease in HCV titer mediated by G. asiatica is independent of cytotoxicity (Figure 4.6).
Hepatitis C virus junction, NS3-4A is made of NS3 and cofactor NS4A which is a
non covalent complex. NS3 is a 70 kDa multifactorial protein, with a serine protease domain
positioned in the N-terminal one third and an NTPase/RNA helicase domain in the C-
terminal two third (Pietschmann et al., 2001; Raney et al., 2010). In our study we have
evaluated that G. asiatica methanol (GALM) extract reduced HCV NS3 protease expression
while chloroform (GALC) and acetone (GALA) extracts had no effect on NS3 protease at
non toxic concentrations (Figure 4.6.1.3). In addition, our Real Time PCR data proves that
GALM extract resulted in 59% and 35% inhibition of HCV NS3 protease level of genotype
3a and 1a respectively at non toxic doses (Figure 4.6.1.2). When G. asiatica was macerated
in increasing polarities of solvents, resulted in 3%, 4%, 72% inhibition of NS3 protease level
of genotype 3a and 0%, 3%, 91% of genotype 1a by GALC, GALA and GALM extracts
respectively (Figure 4.6.1.4). The recent investigation reports that methanol extract of G.
asiatica exhibits antiplatelet (Zia-Ul-Haq et al., 2012) antitumor, cytotoxic (Marya et al.,
2011) analgesic and antipyretic activities (Debajyoti et al., 2012).

82
In this study we also separated different fractions through thin layer and column
chromatography. Figure 4.6.2.1 shows TLC chromatogram of different components
separated from GALM extract and the pure fraction (GA15). The crude GALM extract
separated into sixteen components in (Chloroform: Methanol: Ethyl Acetate: Water 5:3:1:1)
followed by (Chloroform: Ethyl Acetate: Acetic Acid: Water 4: 3: 2: 1) solvent system. For
antiviral screening against HCV, each fraction was dissolved in DMSO and tested for
activity. In another study silymarin crude extract was observed to compromise HCV core
gene of genotype 3a in Huh-7 cells (Ashfaq et al., 2011).
In the reporter assay system hydroxyurea inhibited HCV RNA replication (Nozaki et
al., 2010). The results from this study demonstrate that each fraction was tested for antiviral
activity against HCV and found GA15 fraction reduced HCV titer 87% and 74% of genotype
3a and 1a respectively by Real Time Quantitative PCR analysis (Figure 4.6.3.1). Several
studies have shown that cellular targets of the NS3-4A protease are MAVS (blocking RIG-1
signaling), TRIF (blocking toll-like receptor 3 signaling), TC-PTP (enhance EGF signaling
and basal Akt activity) thereby inactivate mitochondrial host protein (Morikawa et al., 2011).
Similarly, the cleavage of IPS-1 adaptor molecule by HCV NS3/4A protease induced further
blocking of the interferon inducing signaling pathway (Foy et al., 2005). This study indicates
significant downstream blocking of HCV NS3 gene expression by addition of GA15 fraction
at nontoxic concentration of both 3a and 1a genotypes and GAPDH gene expression remains
constant (Figure 4.6.3.2). Our Real Time PCR data shows 91% and 87% inhibition of HCV
NS3 proteases level of genotype 3a and 1a respectively (Figure 4.6.3.3). HCV NS3 gene
expression was also examined at protein level and found significant decrease in NS3 protein
level by addition of GA15 fraction (Figure 4.6.3.4).

83
Dose dependent antiviral activity of active (GA15) fraction was found by serial
dilution method. The EC50 value is the amount of compound required to reduce the virus
50% of its control value (healthy cells). Previous literature indicates that EC50 values of
inhibition for L.radiata, P.lingua, A.annua and L. aggregata were 2.4±0.2, 34.5±2.6,
43.2±14.1 and 88.2±7.7 µg/ml respectively (Li et al., 2005). The results in this study show
that EC50 value of GA15 fraction is 9.46±2.93µg/ml and 10.13±2.75µg/ml for genotype 3a
and 1a respectively. Furthermore, GA15 fraction reduced viral titer of HCV genotype 3a and
1a upto 91% and 86% at 50µg concentration (Figure 4.6.4.1). Similarly, Figure 4.6.4.2 shows
downstream blocking of HCV NS3 protease of genotype 3a and 1a expression in a dose
dependent manner while GAPDH remains constant.
Previous results suggested that hydroxyurea would be an effective anti HCV agent
that can be used not only singly but also in combination with interferon to treat chronic
hepatitis C. Moreover, anti HCV activity of hydroxyurea was higher in combination with
IFN alpha than alone (Nozaki et al., 2010). In present study, the data indicates that GA15
fraction reduced HCV viral titer to 53% and 51% alone in combination with interferon,
resulted in 90% and 82% of genotype 3a and 1a respectively (Figure 4.6.5). These results
support the potential of anti HCV compound alone and also in combination with interferon to
treat chronic Hepatitis C.
Syzygium cumini extract decrease the growth and stimulate apoptosis in HeLa and
SiHa cancer cell lines (Barh and Viswanathan, 2008). In this research, S. cumini leaves
extract (SCLM) demonstrated the Huh-7 and CHO cells remained unaffected at 100 µg of

84
concentration in trypan blue dye explosive assay (Figure 4.1). The cytotoxic effect of
Syzygium cumini crude extract was determined by MTT assay on Huh-7 cells at different
concentrations (Figure 4.3). Recently, S. cumini aqueous extract was evaluated for cell
viability by MTT assay and found non toxic at 100 µg of concentration (Bellé et al., 2013).
Three major steps of Hepatitis C virus life cycle begins with: attachment and entry of
virus into its target cells, replication of genomic RNA, and assembly and release of viruses
from the infected cells (Calland et al., 2012). The mouse hepatocyte lines make it possible to
establish a HCV infection model with diverse applications (Aly et al., 2011). The present
study reports that S. cumine extract showed 45% and 31% inhibition of HCV 3a and 1a
genotype respectively at non toxic concentrations (Figure 4.7)
HCV NS3 protease is a vital component of viral replication and regarded as an
attractive target for patients infected with HCV (Gu and Rice, 2013). The present study
reveals that our methanol extract of S. cumine leaves down regulated the expression level of
HCV NS3 proteases while chloroform (SCLC) and acetone (SCLA) extracts showed no
effect in Huh-7 cells at a concentration of 100 µg and GAPDH expression remains constant
(Figure 4.7.1.3). Furthermore, our Real Time PCR analysis reveals 48% and 34% inhibition
of HCV NS3 proteases level of genotype 3a and 1a respectively at non toxic concentrations
(Figure 4.7.1.2). Similarly, Syzgium cumine leaves in different polarity solvents demonstrated
6%, 9%, 51% inhibition of NS3 protease level of genotype 3a and 3%, 0%, 79% inhibition of
genotype 1a respectively (Figure 4.7.1.4).

85
Phytochemical studies of Syzgium cumine extract contains flavonoids, cardiac
glycosides, terpenoids, phenols, saponins and tannins (Gowri and Vasantha, 2010). The
present study clearly reports that S. cumini leaves extract separates into fifteen components in
(C: M: W 7:2:1) solvent system and four more components in (C: EA: AA: W 4: 3: 2: 1)
mobile phase (Figure 4.7.2.1). All the fractions were dissolved in DMSO and tested against
HCV at nontoxic concentrations and found that fraction (SC14) results in 76% and 62%
decrease in HCV titer of genotype 3a and 1a respectively by Real Time Quantitative PCR
analysis (Figure 4.7.3.1). Previously, crude methanol extract of S. cumini led to the isolation
of 7-hydroxycalamenene, methyl-β-orsellinate , β-sitosterol and oleanolic acid (Sikder et al.,
2012).
The results of this study demonstrated that the expression level of HCV NS3 protease
is significantly reduced by SC14 fraction as compare to all other fractions isolated from S.
cumini leaves methanol (SCLM) extract while GAPDH expression remains constant for all
tested fractions (Figure 4.7.3.2). Furthermore, the results of SC14 fraction were verified by
Real Time PCR analysis and found the inhibition of NS3 protease level to 82% and 76% of
genotype 3a and 1a respectively (Figure 4.7.3.3). Previous literature of western blotting
revealed that the quantity of α-fetoprotein was reduced by 1.0 mg/ml of fucoidan in Huh-7
cells (Nagamine et al., 2009). Results in Figure 4.7.3.4 demonstrates that the protein level of
NS3 protease is decreased by SC14 fraction in Huh-7 cells and GAPDH protein expression
remains constant. So, SC14 fraction decreases HCV NS3 level at RNA as well as protein
level.

86
The outcome of this study exhibited that SC14 fraction reduced 50% (EC50 Value) of
HCV titer of genotype 3a and 1a at a concentration of 31.75±3.28 µg/ml and 71.96±8.67
µg/ml respectively (Figure 4.7.4a). Furthermore, Figure 4.7.4b shows dose dependant
inhibition of HCV NS3 protease of genotype 3a and 1a expression while GAPDH expression
remains constant.
It is well known that IFNs, importantly type I interferon plays a vital role in reducing
viral population through the motivation of interferon stimulating genes (ISGs) (O'Neill and
Bowie, 2010). Currently, peg-Interferon-α and ribavirin is the only standard remedy for all
genotypes of Hepatitis C Virus, but tolerability of this combination treatment is poor with
few side effects and an impaired quality of life (Asselah and Marcellin, 2011). In present
study, effective fraction SC14 was tested without or combination of interferon and viral titers
were quantified through Quantitative RT-PCR. S. cumini active fraction (SC14) exhibited
47% and 49% reduction in viral load, on the other hand its combination with interferon
shows 81% and 76% decline in HCV virus of 3a and 1a genotype respectively (Figure 4.7.5).
So we can suggest that the use of SC14 in combination with interferon would be encouraging
treatment for HCV patients.
The present study was designed to search for phytochemicals from traditional
medicinal plants against HCV. Therefore, an in-vitro bioassay was developed by using HCV
inoculums of genotype 3a and 1a into Huh-7 cell line for screening the active component
from selected plants. Twenty four medicinal plants were collected and their solvent
extraction was performed for further toxicological studies on liver (Huh-7) and fibroblast
(CHO) cells lines by colorimeteric methods, trypan blue dye and MTT cell proliferation

87
assay. Plant extracts designated as NJ, PN and VJ showed toxic effect on Huh 7 cell lines
cells, so excluded for further screening. For antiviral screening, HCV infected cells were
treated with plant extracts at non toxic doses. In in vitro assay on HCV 3a and 1a genotype
five medicinal plant extracts designated as SN, GA, SC, AM and FC showed antiviral
activity. Through Quantitative real time PCR, HCV titer was analyzed and further evaluated
against NS3 proteases of HCV genotype 1a and 3a. Potent plant extracts were fractionated
by thin layer chromatography (TLC) and column chromatography. These purified potent
fractions were then analyzed against Hepatitis C virus to find out EC50 (50% Effective
concentration) and synergistic effect with interferon alpha if exists. Finally, the active
fractions SN8, GA15 and SC14 were effective against HCV and had synergistic effect when
combined with IFN α.
Present in vitro study has identified three active fractions from traditional medicinal
plants as potential anti HCV candidates. Combination of these active ingredients with
interferon (IFN) will be a successful future therapy against Hepatitis C virus. Further studies
are necessary for the better understanding of mechanism to clarify their role in the treatment
of HCV.

88
REFERENCES
Abbasi, A.M., Khan, M.A., Ahmad, M., Zafar, M., Khan, H., Muhammad, N., and Sultana,
S. (2009). Medicinal plants used for the treatment of jaundice and hepatitis based on socio-
economic documentation. African journal of biotechnology 8.
Acharya, E., and Pokhrel, B. (2006). Ethno-medicinal plants used by Bantar of Bhaudaha,
Morang, Nepal. Our Nature 4, 96-103.
Ahaskar, M., Sharma, K., Singh, S., and Sisodia, R. (2007). Radioprotective effect of the
fruit extract of Grewia asiatica in Swiss albino mice against lethal dose of γ-irradiation.
Asian J Exp Sci 21, 295-308.
Aly, H.H., Oshiumi, H., Shime, H., Matsumoto, M., Wakita, T., Shimotohno, K., and Seya,
T. (2011). Development of mouse hepatocyte lines permissive for hepatitis C virus (HCV).
PLoS ONE 6, e21284.
ANNEMARIE WASLEY, S.D., and Alter, M.J. (2000). Epidemiology of hepatitis C:
geographic differences and temporal trends. Hepatitis C: State of the Art at the Millennium
20, 1.
Ashfaq, U.A., Javed, T., Rehman, S., Nawaz, Z., and Riazuddin, S. (2011). Inhibition of
HCV 3a core gene through Silymarin and its fractions. Virology journal 8, 1-7.
Asselah, T., Bieche, I., Sabbagh, A., Bedossa, P., Moreau, R., Valla, D., Vidaud, M., and
Marcellin, P. (2009). Gene expression and hepatitis C virus infection. Gut 58, 846-858.
Asselah, T., and Marcellin, P. (2011). New direct‐acting antivirals' combination for the
treatment of chronic hepatitis C. Liver International 31, 68-77.
Attaullah, S., Khan, S., and Ali, I. (2011). Hepatitis C virus genotypes in Pakistan: a systemic
review. Virol J 8, 433.
Azaizeh, H., Fulder, S., Khalil, K., and Said, O. (2003). Ethnobotanical knowledge of local
Arab practitioners in the Middle Eastern region. Fitoterapia 74, 98-108.
Bacon, B.R., Shiffman, M.L., Mendes, F., Ghalib, R., Hassanein, T., Morelli, G., Joshi, S.,
Rothstein, K., Kwo, P., and Gitlin, N. (2009). Retreating chronic hepatitis C with daily
interferon alfacon‐1/ribavirin after nonresponse to pegylated interferon/ribavirin: DIRECT
results. Hepatology 49, 1838-1846.
Barh, D., and Viswanathan, G. (2008). Syzygium cumini inhibits growth and induces
apoptosis in cervical cancer cell lines: a primary study. ecancermedicalscience 2.
Bartenschlager, R., and Lohmann, V. (2000). Replication of hepatitis C virus. Journal of
General Virology 81, 1631-1648.

89
Bartosch, B., Bukh, J., Meunier, J.C., Granier, C., Engle, R.E., Blackwelder, W.C., Emerson,
S.U., Cosset, F.L., and Purcell, R.H. (2003a). In vitro assay for neutralizing antibody to
hepatitis C virus: evidence for broadly conserved neutralization epitopes. Proc Natl Acad Sci
U S A 100, 14199-14204.
Bartosch, B., Dubuisson, J., and Cosset, F.-L. (2003b). Infectious hepatitis C virus pseudo-
particles containing functional E1–E2 envelope protein complexes. The Journal of
experimental medicine 197, 633-642.
Baumert, T.F., Ito, S., Wong, D.T., and Liang, T.J. (1998). Hepatitis C virus structural
proteins assemble into viruslike particles in insect cells. Journal of virology 72, 3827-3836.
Bazan, J.F., and Fletterick, R.J. (1988). Viral cysteine proteases are homologous to the
trypsin-like family of serine proteases: structural and functional implications. Proceedings of
the National Academy of Sciences 85, 7872-7876.
Bellé, L.P., Bitencourt, P.E.R., Abdalla, F.H., de Bona, K.S., Peres, A., Maders, L.D.K., and
Moretto, M.B. (2013). Aqueous seed extract of Syzygium cumini inhibits the dipeptidyl
peptidase IV and adenosine deaminase activities, but it does not change the CD26 expression
in lymphocytes in vitro. Journal of physiology and biochemistry 69, 119-124.
Bhanuprakash, V., Hosamani, M., Balamurugan, V., Gandhale, P., Naresh, R., Swarup, D.,
and Singh, R. (2008). In vitro antiviral activity of plant extracts on goatpox virus replication.
Indian journal of experimental biology 46, 120.
Biselli, M., Andreone, P., Gramenzi, A., Lorenzini, S., Loggi, E., Bonvicini, F., Cursaro, C.,
and Bernardi, M. (2006). Pegylated interferon plus ribavirin for recurrent Hepatitis C
infection after liver transplantation in naive and non-responder patients on a stable
immunosuppressive regimen. Digestive and liver disease 38, 27-32.
Blight, K.J., Kolykhalov, A.A., and Rice, C.M. (2000). Efficient initiation of HCV RNA
replication in cell culture. Science 290, 1972-1974.
Blight, K.J., McKeating, J.A., Marcotrigiano, J., and Rice, C.M. (2003). Efficient replication
of hepatitis C virus genotype 1a RNAs in cell culture. Journal of virology 77, 3181-3190.
Block, G., Patterson, B., and Subar, A. (1992). Fruit, vegetables, and cancer prevention: a
review of the epidemiological evidence. Nutrition and cancer 18, 1-29.
Bollati, M., Alvarez, K., Assenberg, R., Baronti, C., Canard, B., Cook, S., Coutard, B.,
Decroly, E., de Lamballerie, X., and Gould, E.A. (2010). Structure and functionality in
flavivirus NS-proteins: Perspectives for drug design. Antiviral research 87, 125-148.
Bostan, N., and Mahmood, T. (2010). An overview about hepatitis C: a devastating virus.
Critical reviews in microbiology 36, 91-133.
Boyer, J., and Liu, R.H. (2004). Apple phytochemicals and their health benefits. Nutr J 3, 12.
Bukh, J., Pietschmann, T., Lohmann, V., Krieger, N., Faulk, K., Engle, R.E., Govindarajan,
S., Shapiro, M., Claire, M.S., and Bartenschlager, R. (2002). Mutations that permit efficient

90
replication of hepatitis C virus RNA in Huh-7 cells prevent productive replication in
chimpanzees. Proceedings of the National Academy of Sciences 99, 14416-14421.
Cai, Z., Zhang, C., Chang, K.S., Jiang, J., Ahn, B.C., Wakita, T., Liang, T.J., and Luo, G.
(2005). Robust production of infectious hepatitis C virus (HCV) from stably HCV cDNA-
transfected human hepatoma cells. J Virol 79, 13963-13973.
Calland, N., Dubuisson, J., Rouillé, Y., and Séron, K. (2012). Hepatitis C virus and natural
compounds: a new antiviral approach? Viruses 4, 2197-2217.
Cavallaro, L., Garcia, G., Broussalis, A., Ferraro, X., Martino, V., Coussio, J., De Torres, R.,
and Campos, R. (1995). Antiherpetic in vitro activity of Gamochaeta simplicicaulis extract.
Phytotherapy Research 9, 176-179.
Chaudhuri, S., Das, S., Chowdhury, A., Santra, A., Bhattacharya, S., and Naik, T. (2005).
Molecular epidemiology of HCV infection among acute and chronic liver disease patients in
Kolkata, India. Journal of clinical virology 32, 38-46.
Collett, M.S., Larson, R., Gold, C., Strick, D., Anderson, D.K., and Purchio, A. (1988).
Molecular cloning and nucleotide sequence of the pestivirus bovine viral diarrhea virus.
Virology 165, 191-199.
Cunningham, M., and Foster, G.R. (2012). Efficacy and safety of telaprevir in patients with
genotype 1 hepatitis C infection. Therapeutic advances in gastroenterology 5, 139-151.
De Clercq, E. (2002). Cidofovir in the treatment of poxvirus infections. Antiviral research 55,
1-13.
De Francesco, R., and Migliaccio, G. (2005). Challenges and successes in developing new
therapies for hepatitis C. Nature 436, 953-960.
Debajyoti, D., Achintya, M., Debdas, D., Achintya, S., and Jayram, H. (2012). Evaluation of
antipyretic and analgesic activity of parusaka (Grewia asiatica Linn.): An indigenous Indian
plant. Int J Res Ayurveda Pharm 3, 519-524.
Dhawan, B. (2012). Anti-Viral Activity of Indian Plants. Proceedings of the National
Academy of Sciences, India Section B: Biological Sciences 82, 209-224.
Dhiman, R.K., and Chawla, Y.K. (2005). Herbal medicines for liver diseases. Digestive
diseases and sciences 50, 1807-1812.
Diallo, D., Hveem, B., Mahmoud, M.A., Berge, G., Paulsen, B.S., and Maiga, A. (1999). An
ethnobotanical survey of herbal drugs of Gourma district, Mali. Pharmaceutical Biology 37,
80-91.
Duraipandiyan, V., Ayyanar, M., and Ignacimuthu, S. (2006). Antimicrobial activity of some
ethnomedicinal plants used by Paliyar tribe from Tamil Nadu, India. BMC complementary
and alternative medicine 6, 35.

91
Dusheiko, G., Schmilovitz‐Weiss, H., Brown, D., McOmish, F., Yap, P.L., Sherlock, S.,
McIntyre, N., and Simmonds, P. (1994). Hepatitis C virus genotypes: an investigation of
type‐specific differences in geographic origin and disease. Hepatology 19, 13-18.
Duvet, S., Cocquerel, L., Pillez, A., Cacan, R., Verbert, A., Moradpour, D., Wychowski, C.,
and Dubuisson, J. (1998). Hepatitis C virus glycoprotein complex localization in the
endoplasmic reticulum involves a determinant for retention and not retrieval. Journal of
Biological Chemistry 273, 32088-32095.
Foy, E., Li, K., Sumpter, R., Loo, Y.-M., Johnson, C.L., Wang, C., Fish, P.M., Yoneyama,
M., Fujita, T., and Lemon, S.M. (2005). Control of antiviral defenses through hepatitis C
virus disruption of retinoic acid-inducible gene-I signaling. Proceedings of the National
Academy of Sciences of the United States of America 102, 2986-2991.
Fretz, C., Jeannel, D., Stuyver, L., Herve, V., Lunel, F., Boudifa, A., Mathiot, C., and
Fournel, J. (1995). HCV infection in a rural population of the Central African Republic
(CAR): evidence for three additional subtypes of genotype 4. Journal of medical virology 47,
435-437.
Ge, D., Fellay, J., Thompson, A.J., Simon, J.S., Shianna, K.V., Urban, T.J., Heinzen, E.L.,
Qiu, P., Bertelsen, A.H., and Muir, A.J. (2009). Genetic variation in IL28B predicts hepatitis
C treatment-induced viral clearance. Nature 461, 399-401.
Ghany, M.G., Nelson, D.R., Strader, D.B., Thomas, D.L., and Seeff, L.B. (2011). An update
on treatment of genotype 1 chronic hepatitis C virus infection: 2011 practice guideline by the
American Association for the Study of Liver Diseases. Hepatology 54, 1433-1444.
Gowri, S.S., and Vasantha, K. (2010). Phytochemical screening and antibacterial activity of
Syzygium cumini (L.)(Myrtaceae) leaves extracts. Int J Pharm Tech Res 2, 1569-1573.
Grossmann, M.E., Mizuno, N.K., Schuster, T., and Cleary, M.P. (2010). Punicic acid is an ω-
5 fatty acid capable of inhibiting breast cancer proliferation. International journal of
oncology 36, 421.
Gu, M., and Rice, C.M. (2013). Structures of hepatitis C virus nonstructural proteins required
for replicase assembly and function. Current opinion in virology.
Gupta, M., Shaw, B., and Mukherjee, A. (2010). A new glycosidic flavonoid from Jwarhar
mahakashay (antipyretic) Ayurvedic preparation. International journal of Ayurveda research
1, 106.
Haq, I., and Rehman, M. (1990). Medicinal plants of upper Swat (NWFP) Pakistan. Hamdard
Med 33, 51-68.
Hassan, B.A.R. (2012). Medicinal Plants (Importance and Uses). Pharmaceutica Analytica
Acta.
Hissar, S.S., Goyal, A., Kumar, M., Pandey, C., Suneetha, P., Sood, A., Midha, V., Sakhuja,
P., Malhotra, V., and Sarin, S.K. (2006). Hepatitis C virus genotype 3 predominates in North

92
and Central India and is associated with significant histopathologic liver disease. Journal of
medical virology 78, 452-458.
Hoareau, L., and DaSilva, E.J. (1999). Medicinal plants: a re-emerging health aid. Electr J
Biotechnol 2, http://www.ejb.org/content/vol2/issue2/full2.
Hsieh, C.-C., Fang, H.-L., and Lina, W.-C. (2008). Inhibitory effect of< i> Solanum
nigrum</i> on thioacetamide-induced liver fibrosis in mice. Journal of Ethnopharmacology
119, 117-121.
Hsu, M., Zhang, J., Flint, M., Logvinoff, C., Cheng-Mayer, C., Rice, C.M., and McKeating,
J.A. (2003). Hepatitis C virus glycoproteins mediate pH-dependent cell entry of pseudotyped
retroviral particles. Proc Natl Acad Sci U S A 100, 7271-7276.
Huang, H.-C., Syu, K.-Y., and Lin, J.-K. (2010). Chemical composition of Solanum nigrum
linn extract and induction of autophagy by leaf water extract and its major flavonoids in
AU565 breast cancer cells. Journal of agricultural and food chemistry 58, 8699-8708.
Hudson, J. (1989). Plant photosensitizers with antiviral properties. Antiviral research 12, 55-
74.
Hudson, J., Lopez-Bazzocchi, I., and Towers, G. (1991). Antiviral activities of hypericin.
Antiviral research 15, 101-112.
Hussain, A., Sasidharan, S., Ahmed, T., Ahmed, M., and Sharma, C. (2009). Clove
(Syzygium aromaticum) extract potentiates gemcitabine cytotoxic effect on human cervical
cancer cell line. Int J Cancer Res 5, 95-104.
Idrees, M., Lal, A., Naseem, M., and Khalid, M. (2008). High prevalence of hepatitis C virus
infection in the largest province of Pakistan. Journal of digestive diseases 9, 95-103.
Jassim, S.A., and Naji, M.A. (2003). Novel antiviral agents: a medicinal plant perspective. J
Appl Microbiol 95, 412-427.
Jatav, V.S., Singh, S.K., Khatri, P., and Sharma, A.K. (2011). Recent pharmacological trends
of Glycyrrhiza glabra Linn. Unani Res 1, 1-4.
Jauregui, H.O., Hayner, N.T., Driscoll, J.L., Williams-Holland, R., Lipsky, M.H., and
Galletti, P.M. (1981). Trypan blue dye uptake and lactate dehydrogenase in adult rat
hepatocytes—freshly isolated cells, cell suspensions, and primary monolayer cultures. In
vitro 17, 1100-1110.
Jazwinski, A.B., and Muir, A.J. (2011). Direct-acting antiviral medications for chronic
hepatitis C virus infection. Gastroenterology & hepatology 7, 154.
Ji, Y., Gao, S., Ji, C., and Zou, X. (2008). Induction of apoptosis in HepG< sub> 2</sub>
cells by solanine and Bcl-2 protein. Journal of Ethnopharmacology 115, 194-202.

93
Joo, H.Y., Lim, K., and Lim, K.T. (2009). Phytoglycoprotein (150 kDa) isolated from
Solanum nigrum Linne has a preventive effect on dextran sodium sulfate‐induced colitis in
A/J mouse. Journal of Applied Toxicology 29, 207-213.
Kala, C.P. (2005). Ethnomedicinal botany of the Apatani in the Eastern Himalayan region of
India. Journal of ethnobiology and ethnomedicine 1, 1-8.
Kapusta, J., Modelska, A., Figlerowicz, M., Pniewski, T., Letellier, M., Lisowa, O., Yusibov,
V., Koprowski, H., Plucienniczak, A., and Legocki, A. (1999). A plant-derived edible
vaccine against hepatitis B virus. The FASEB journal 13, 1796-1799.
Khattak, J.Z.K., Anwar, Z., Aftab, S., Afzal, M., Islam, M., and Khan, A. Solenum nigrum as
Potent Therapy: A Review. British Journal of Pharmacology 3.
Kiran, B., Kudesia, R., Rani, M., and Pal, A. (2009). Reclaiming degraded land in India
through the cultivation of medicinal plants. Bot Res Int 2, 174-181.
Kitazato, K., Wang, Y., and Kobayashi, N. (2007). Viral infectious disease and natural
products with antiviral activity. Drug discoveries & therapeutics 1, 14.
Kolykhalov, A.A., Agapov, E.V., Blight, K.J., Mihalik, K., Feinstone, S.M., and Rice, C.M.
(1997). Transmission of hepatitis C by intrahepatic inoculation with transcribed RNA.
Science 277, 570-574.
Kolykhalov, A.A., Mihalik, K., Feinstone, S.M., and Rice, C.M. (2000). Hepatitis C virus-
encoded enzymatic activities and conserved RNA elements in the 3′ nontranslated region are
essential for virus replication in vivo. Journal of virology 74, 2046-2051.
Krieger, N., Lohmann, V., and Bartenschlager, R. (2001). Enhancement of hepatitis C virus
RNA replication by cell culture-adaptive mutations. Journal of virology 75, 4614-4624.
Kuiken, C., Yusim, K., Boykin, L., and Richardson, R. (2005). The Los Alamos hepatitis C
sequence database. Bioinformatics 21, 379-384.
Kumar, P., Jaiswal, P., Singh, V.K., Singh, D.K., and Singh, D. MEDICI AL,
THERAPEUTIC AD PHARMACOLOGICAL EFFECTS OF SYZYGIUM
AROMATICUM (LAU G).
Kumar, R.V., Venkatrajireddy, G., Bikshapathi, T., and Reddy, M.K. (2012). Antioxidant-the
maximum expressed activity among 63 medicinal plants. J Phyto Pharmacol 1, 1-13.
Lamarre, D., Anderson, P.C., Bailey, M., Beaulieu, P., Bolger, G., Bonneau, P., Bös, M.,
Cameron, D.R., Cartier, M., and Cordingley, M.G. (2003). An NS3 protease inhibitor with
antiviral effects in humans infected with hepatitis C virus. Nature 426, 186-189.
Lanford, R.E., Sureau, C., Jacob, J.R., White, R., and Fuerst, T.R. (1994). Demonstration of<
i> in Vitro</i> Infection of Chimpanzee Hepatocytes with Hepatitis C Virus Using Strand-
Specific RT/PCR. Virology 202, 606-614.

94
Lee, K.-R., Kozukue, N., Han, J.-S., Park, J.-H., Chang, E.-y., Baek, E.-J., Chang, J.-S., and
Friedman, M. (2004). Glycoalkaloids and metabolites inhibit the growth of human colon
(HT29) and liver (HepG2) cancer cells. Journal of agricultural and food chemistry 52, 2832-
2839.
Leporatti, M.L., and Ghedira, K. (2009). Journal of Ethnobiology and Ethnomedicine.
Journal of ethnobiology and ethnomedicine 5, 31.
Leung, D., Abbenante, G., and Fairlie, D.P. (2000). Protease inhibitors: current status and
future prospects. Journal of medicinal chemistry 43, 305-341.
Li, H.-M., Tang, Y.-L., Zhang, Z.-H., Liu, C.-J., Li, H.-Z., Li, R.-T., and Xia, X.-S. (2012).
Compounds from Arnebia euchroma and their related anti-HCV and antibacterial activities.
Planta medica 78, 39-45.
Li, J., Li, Q., Feng, T., and Li, K. (2008). Aqueous extract of< i> Solanum nigrum</i> inhibit
growth of cervical carcinoma (U14) via modulating immune response of tumor bearing mice
and inducing apoptosis of tumor cells. Fitoterapia 79, 548-556.
Li, J., Li, Q.W., Gao, D.W., Han, Z.S., and Lu, W.Z. (2009). Antitumor and
immunomodulating effects of polysaccharides isolated from Solanum nigrum Linne.
Phytotherapy Research 23, 1524-1530.
Li, S.-y., Chen, C., Zhang, H.-q., Guo, H.-y., Wang, H., Wang, L., Zhang, X., Hua, S.-n., Yu,
J., and Xiao, P.-g. (2005). Identification of natural compounds with antiviral activities against
SARS-associated coronavirus. Antiviral research 67, 18-23.
Locarnini, S.A., and Bartholomeusz, A. (2002). Advances in hepatitis C: What is coming in
the next 5 years? Journal of gastroenterology and hepatology 17, 442-447.
Manns, M.P., McHutchison, J.G., Gordon, S.C., Rustgi, V.K., Shiffman, M., Reindollar, R.,
Goodman, Z.D., Koury, K., Ling, M.-H., and Albrecht, J.K. (2001). Peginterferon alfa-2b
plus ribavirin compared with interferon alfa-2b plus ribavirin for initial treatment of chronic
hepatitis C: a randomised trial. The Lancet 358, 958-965.
Marya, B., Dattani, K.H., Patel, D.D., Patel, P.D., Patel, D., Suthar, M.P., Patel, V.P., and
Bothara, S.B. (2011). In vitro cytotoxicity evaluation of aqueous fruit and leaf extracts of
Grewia asiatica using MTT assay. Scholars Research Library Der Pharma Chemica 3, 282-
287.
McHutchison, J.G., Lawitz, E.J., Shiffman, M.L., Muir, A.J., Galler, G.W., McCone, J.,
Nyberg, L.M., Lee, W.M., Ghalib, R.H., and Schiff, E.R. (2009). Peginterferon alfa-2b or
alfa-2a with ribavirin for treatment of hepatitis C infection. New England Journal of
Medicine 361, 580-593.
Mercer, D.F., Schiller, D.E., Elliott, J.F., Douglas, D.N., Hao, C., Rinfret, A., Addison, W.R.,
Fischer, K.P., Churchill, T.A., and Lakey, J.R. (2001). Hepatitis C virus replication in mice
with chimeric human livers. Nature medicine 7, 927-933.

95
Meuleman, P., Libbrecht, L., De Vos, R., de Hemptinne, B., Gevaert, K., Vandekerckhove,
J., Roskams, T., and Leroux‐Roels, G. (2005). Morphological and biochemical
characterization of a human liver in a uPA‐SCID mouse chimera. Hepatology 41, 847-856.
Meylan, E., Curran, J., Hofmann, K., Moradpour, D., Binder, M., Bartenschlager, R., and
Tschopp, J. (2005). Cardif is an adaptor protein in the RIG-I antiviral pathway and is targeted
by hepatitis C virus. Nature 437, 1167-1172.
Mohanty, S., and Cock, I.E. (2012). The chemotherapeutic potential of Terminalia
ferdinandiana: Phytochemistry and bioactivity. Pharmacognosy reviews 6, 29.
Moradpour, D., Brass, V., and Penin, F. (2005). Function follows form: The structure of the
N‐terminal domain of HCV NS5A. Hepatology 42, 732-735.
Morgan, K. (1994). Medicine of the gods: Basic principles of Ayurvedic medicine (Charles
T. Banford Company).
Morikawa, K., Lange, C., Gouttenoire, J., Meylan, E., Brass, V., Penin, F., and Moradpour,
D. (2011). Nonstructural protein 3‐4A: the Swiss army knife of hepatitis C virus. Journal of
viral hepatitis 18, 305-315.
Moshi, M.J., Otieno, D.F., Mbabazi, P.K., and Weisheit, A. (2009). Journal of Ethnobiology
and Ethnomedicine. Journal of ethnobiology and ethnomedicine 5, 24.
Mosmann, T. (1983a). Rapid colorimetric assay for cellular growth and survival: application
to proliferation and cytotoxicity assays. Journal of immunological methods 65, 55-63.
Mosmann, T. (1983b). Rapid colorimetric assay for cellular growth and survival: application
to proliferation and cytotoxicity assays. J Immunol Methods 65, 55-63.
Mukhtar, M., Arshad, M., Ahmad, M., Pomerantz, R.J., Wigdahl, B., and Parveen, Z. (2008).
Antiviral potentials of medicinal plants. Virus Research 131, 111-120.
Murphy, W.L., Andersen, J.M., and Ebelin, R.M. (2002). Assessment of geology as it
pertains to modeling uplift in jointed rock :
a basis for inclusion of uncertainty in flow models (Vicksburg, Miss., US Army Corps of
Engineers, Engineer Research and Development Center, Geotechnical and Structures
Laboratory, Information Technology Laboratory).
Nagamine, T., Hayakawa, K., Kusakabe, T., Takada, H., Nakazato, K., Hisanaga, E., and Iha,
M. (2009). Inhibitory effect of fucoidan on Huh7 hepatoma cells through downregulation of
CXCL12. Nutrition and cancer 61, 340-347.
Nascimento, G.G., Locatelli, J., Freitas, P.C., and Silva, G.L. (2000). Antibacterial activity of
plant extracts and phytochemicals on antibiotic-resistant bacteria. Brazilian Journal of
Microbiology 31, 247-256.

96
Ndjomou, J., Pybus, O.G., and Matz, B. (2003). Phylogenetic analysis of hepatitis C virus
isolates indicates a unique pattern of endemic infection in Cameroon. Journal of General
Virology 84, 2333-2341.
Newman, D.J., Cragg, G.M., and Snader, K.M. (2000). The influence of natural products
upon drug discovery. Natural product reports 17, 215-234.
Nisar, M.F., Ismail, S., Arshad, M., Majeed, A., and Arfan, M. (2011). Ethnomedicinal Flora
of District Mandi Bahaudin, Pakistan. Middle-East Journal of Scientific Research 9, 233-238.
Noppornpanth, S., Sablon, E., De Nys, K., Lien, T.X., Brouwer, J., Van Brussel, M., Smits,
S.L., Poovorawan, Y., Osterhaus, A.D., and Haagmans, B.L. (2006). Genotyping hepatitis C
viruses from Southeast Asia by a novel line probe assay that simultaneously detects core and
5′ untranslated regions. Journal of clinical microbiology 44, 3969-3974.
Nozaki, A., Morimoto, M., Kondo, M., Oshima, T., Numata, K., Fujisawa, S., Kaneko, T.,
Miyajima, E., Morita, S., and Mori, K. (2010). Hydroxyurea as an inhibitor of hepatitis C
virus RNA replication. Archives of virology 155, 601-605.
O'Neill, L.A., and Bowie, A.G. (2010). Sensing and signaling in antiviral innate immunity.
Current Biology 20, R328-R333.
Pak, E., Esrason, K.T., and Wu, V.H. (2004). Hepatotoxicity of herbal remedies: an emerging
dilemma. Progress in Transplantation 14, 91-96.
Pietschmann, T., Lohmann, V., Rutter, G., Kurpanek, K., and Bartenschlager, R. (2001).
Characterization of cell lines carrying self-replicating hepatitis C virus RNAs. Journal of
virology 75, 1252-1264.
Poonam, K., and Singh, G.S. (2009). Ethnobotanical study of medicinal plants used by the
Taungya community in Terai Arc Landscape, India. Journal of Ethnopharmacology 123, 167-
176.
Qureshi, R., Waheed, A., Arshad, M., and Umbreen, T. (2009). Medico-ethnobotanical
inventory of Tehsil Chakwal, Pakistan. Pak J Bot 41, 529-538.
Raney, K.D., Sharma, S.D., Moustafa, I.M., and Cameron, C.E. (2010). Hepatitis C virus
non-structural protein 3 (HCV NS3): a multifunctional antiviral target. Journal of Biological
Chemistry 285, 22725-22731.
Ravi, V., Saleem, T., Maiti, P., Gauthaman, K., and Ramamurthy, J. (2009). Phytochemical
and pharmacological evaluation of Solanum nigrum Linn. African Journal of Pharmacy and
Pharmacology 3, 454-457.
Reed, K., and Rice, C. (2000). Overview of hepatitis C virus genome structure, polyprotein
processing, and protein properties. In The Hepatitis C Viruses (Springer), pp. 55-84.
Sahranavard, S., Naghibi, F., Mosaddegh, M., Esmaeili, S., Sarkhail, P., Taghvaei, M., and
Ghafari, S. (2009). Cytotoxic activities of selected medicinal plants from Iran and

97
phytochemical evaluation of the most potent extract. Research in pharmaceutical sciences 4,
133.
Saleem, T.M., Chetty, C.M., Ramkanth, S., Rajan, V., Kumar, K.M., and Gauthaman, K.
(2010). Hepatoprotective herbs—a review. International Journal of Research in
Pharmaceutical Sciences 1, 1-5.
Sánchez–Tapias, J.M., Diago, M., Escartín, P., Enríquez, J., Romero–Gómez, M., Bárcena,
R., Crespo, J., Andrade, R., Martínez–Bauer, E., and Pérez, R. (2006). Peginterferon-alfa2a
plus ribavirin for 48 versus 72 weeks in patients with detectable hepatitis C virus RNA at
week 4 of treatment. Gastroenterology 131, 451-460.
Sarrazin, C., and Zeuzem, S. (2010). Resistance to direct antiviral agents in patients with
hepatitis C virus infection. Gastroenterology 138, 447-462.
Saxena, S. (2005). Glycyrrhiza glabra: medicine over the millennium. Nat Prod Rad 4, 358-
367.
Sharma, Y., Singh, H., and Mehra, B. (2004). Hepatoprotective effect of few Ayurvedic
herbs in patients receiving antituberculus treatment. Indian Journal of Traditional Knowledge
3, 391-396.
Shepard, C.W., Finelli, L., and Alter, M.J. (2005). Global epidemiology of hepatitis C virus
infection. The Lancet infectious diseases 5, 558-567.
Sikdar, M., and Dutta, U. (2008). Traditional phytotherapy among the Nath people of Assam.
Ethno Med 2, 39-45.
Sikder, M.A.A., Kaisar, M.A., Rahman, M.S., Hasan, C.M., Al-Rehaily, A.J., and Rashid,
M.A. (2012). Secondary Metabolites from Seed Extracts of Syzygium Cumini (L.). Journal
of Physical Science 23, 83-87.
Simmonds, P. (2004). Genetic diversity and evolution of hepatitis C virus–15 years on.
Journal of General Virology 85, 3173-3188.
Singh, B., Gupta, V., Bansal, P., Singh, R., and Kumar, D. (2010). Pharmacological potential
of plant used as aphrodisiacs. International Journal of Pharmaceutical Sciences Review and
Research 5, 104-113.
Sivaperumal, R., Ramya, S., Ravi, A.V., Rajasekaran, C., and Jayakumararaj, R. (2010).
Ethnopharmacological studies on the medicinal plants used by tribal inhabitants of Kottur
Hills, Dharmapuri, Tamilnadu, India. Environ We Int J Sci Technol 5, 57-64.
Smith, D.B., Pathirana, S., Davidson, F., Lawlor, E., Power, J., Yap, P.L., and Simmonds, P.
(1997). The origin of hepatitis C virus genotypes. Journal of General Virology 78, 321-328.
Son, Y.-O., Kim, J., Lim, J.-C., Chung, Y., Chung, G.-H., and Lee, J.-C. (2003). Ripe fruits
of< i> Solanum nigrum</i> L. inhibits cell growth and induces apoptosis in MCF-7 cells.
Food and chemical toxicology 41, 1421-1428.

98
Steinmann, E., Penin, F., Kallis, S., Patel, A.H., Bartenschlager, R., and Pietschmann, T.
(2007). Hepatitis C virus p7 protein is crucial for assembly and release of infectious virions.
PLoS Pathog 3, e103.
Sumpter, R., Loo, Y.-M., Foy, E., Li, K., Yoneyama, M., Fujita, T., Lemon, S.M., and Gale,
M. (2005). Regulating intracellular antiviral defense and permissiveness to hepatitis C virus
RNA replication through a cellular RNA helicase, RIG-I. Journal of virology 79, 2689-2699.
Tan, S.-L. (2006). Hepatitis C viruses: genomes and molecular biology (The Publisher).
Tellinghuisen, T.L., and Rice, C.M. (2002). Interaction between hepatitis C virus proteins
and host cell factors. Current opinion in microbiology 5, 419-427.
Tong, J., Wang, Y.-w., and Lu, Y.-a. (2012). New developments in small molecular
compounds for anti-hepatitis C virus (HCV) therapy. Journal of Zhejiang University
SCIENCE B 13, 56-82.
Trinks, J., Gadano, A., and Argibay, P. (2012). Evolving trends in the hepatitis C virus
molecular epidemiology studies: from the viral sequences to the human genome.
Epidemiology Research International 2012.
UD-DIN, A.M., DIN KHAN, Z.-U., Ahmad, M., AKRAM KASHMIRI, M., Yasmin, S., and
Mazhar, H. (2009). Chemotaxonomic significance of flavonoids in the Solanum nigrum
complex. Journal of the Chilean Chemical Society 54, 486-490.
Vega-Avila, E., Espejo-Serna, A., Alarcón-Aguilar, F., and Velasco-Lezama, R. (2009).
Cytotoxic activity of four Mexican medicinal plants. Paper presented at: Proceedings of the
Western Pharmacology Society.
Vermani, K., and Garg, S. (2002). Herbal medicines for sexually transmitted diseases and
AIDS. Journal of Ethnopharmacology 80, 49-66.
Vijayan, P., Raghu, C., Ashok, G., Dhanaraj, S., and Suresh, B. (2004). Antiviral activity of
medicinal plants of Nilgiris. Indian Journal of Medical Research 120, 24-29.
Wakita, T., Pietschmann, T., Kato, T., Date, T., Miyamoto, M., Zhao, Z., Murthy, K.,
Habermann, A., Kräusslich, H.-G., and Mizokami, M. (2005). Production of infectious
hepatitis C virus in tissue culture from a cloned viral genome. Nature medicine 11, 791-796.
Wang, H.-C., Wu, D.-H., Chang, Y.-C., Li, Y.-J., and Wang, C.-J. (2010). Solanum nigrum
Linn. water extract inhibits metastasis in mouse melanoma cells in vitro and in vivo. Journal
of agricultural and food chemistry 58, 11913-11923.
Wang, X., Zhang, A., and Sun, H. (2013). Recent Advances in Natural Products from Plants
for Treatment of Liver Diseases. European journal of medicinal chemistry.
Yan, F.M., Chen, A.S., Hao, F., Zhao, X.P., Gu, C.H., Zhao, L.B., Yang, D.L., and Hao, L.J.
(2000). Hepatitis C virus may infect extrahepatic tissues in patients with hepatitis C. World
Journal of Gastroenterology 6, 805-811.

99
Yanai, Y., Sanou, O., Kayano, T., Ariyasu, H., Yamamoto, K., Yamauchi, H., Ikegami, H.,
and Kurimoto, M. (2001). Analysis of the antiviral activities of natural IFN-α preparations
and their subtype compositions. Journal of Interferon & Cytokine Research 21, 835-841.
Yi, M., Villanueva, R.A., Thomas, D.L., Wakita, T., and Lemon, S.M. (2006). Production of
infectious genotype 1a hepatitis C virus (Hutchinson strain) in cultured human hepatoma
cells. Proceedings of the National Academy of Sciences of the United States of America 103,
2310-2315.
Zafar, R., and Ali, S. (1998). Cichorium intybus Linn. A review. Hamdard Med 41, 98-109.
Zeuzem, S., Andreone, P., Pol, S., Lawitz, E., Diago, M., Roberts, S., Focaccia, R.,
Younossi, Z., Foster, G.R., and Horban, A. (2011). Telaprevir for retreatment of HCV
infection. New England Journal of Medicine 364, 2417-2428.
Zia-Ul-Haq, M., Shahid, S.A., Muhammed, S., Qayum, M., Khan, I., and Ahmad, S. (2012).
Antimalarial, antiemetic and antidiabetic potential of Grewia asiatica L. leaves. J Med Plants
Res 6, 3213-3216.
Zia-Ul-Haq, M., Stanković, M.S., Rizwan, K., and Feo, V.D. (2013). Grewia asiatica L., a
food plant with multiple uses. Molecules 18, 2663-2682.

100
7. APPENDICES
APPENDIX-I
5x TBE Buffer
Trizma base 54g
Boric acid 27.5g
0.5M EDTA 20mL
Adjust volume to 1 liter with distilled water
6x DNA Loading Dye
Ficoll 20 %
EDTA 0.1M
SDS 1 %
Bromophenol blue 0.25 %
Xylene Cyanol 0.25 %
Protein Extraction Buffer (5mL)
Glycerol 0.5mL
0.5M EDTA 0.4mL (pH 7.5)
5M NaCl 0.15mL
1M TrisCl 0.05mL (pH 7.5)
NH4Cl 26.7mg
DTT 15mg
PMSF 2mM

101
Phosphate Buffered Saline (PBS)
CO 2
NaCl 8 g
KCl 0.2 g
KH2PO40.24 g
Na2HPO4 1.44 g .
Dissolve in 1 liter of distilled water; adjust pH to 7.4 and autoclaved.
1X Tris buffered saline tween 20 TBST
From stock 10X TBS make one liter 1X TB
10X TBS 100 m
dH2O 900 m
Twin 20 1 ml
Phosphate Buffered Saline-Tween 20 (PBST)
Tween-20 500 µL (0.05 %)
Dissolved in 1 liter of 1x PBS
12% separating gel
Water 2.8mL
Acrylamide (30%) 3.2mL
4xTris-SDS 2.5mL (pH8.8)
APS (10 %) 26.7µL
TEMED 5.3µL

102
4% stacking gel
Water 2.5mL
Acrylamide (30%) 0.533mL
4xTris-SDS 1mL (pH6.8)
APS (10 %) 31.5µL
TEMED 6.3µL
10% APS (ammonium per sulphate)
APS 0.1g
Water 1mL
30% Acrylamide
Acrylamide 29gm
Bisacrylamide 1g
dH2O
To make volume 100 ml
10% Sodium dodidyl sulphate SDS
SDS 10g
dH2O 90 ml
5% Skimmed Milk (Blocking Solution)
Skimmed Milk 0.5 g
TBST 10 ml

103
10x Running Buffer
Trizma base 30.05g
Glycine 142.5g
SDS 10g
Dissolve in 1 liter of distilled water
Transfer Buffer
Trizma base 3.032g
Glycine 14.416g
Methanol 200mL
Adjust volume upto 1 liter.
Coomassie Stain
Coomassie blue 2.5g
Methanol 455mL
Glacial acetic acid 91mL
Adjust volume to 1 liter with distilled water.
Coomassie Destain
Methanol 250mL
Glacial acetic acid 70mL
Adjust volume to 1 liter with distilled water.
distilled water.

104
APPENDIX-II
Publications:
Javed, T., Ashfaq, U. A., Riaz, S., Rehman, S., & Riazuddin, S.
(2011). In-vitro antiviral activity of Solanum nigrum against
Hepatitis C Virus. Virol J, 8, 26.
Ashfaq, U. A., Javed, T., Rehman, S., Nawaz, Z., & Riazuddin,
S. (2011). An overview of HCV molecular biology, replication
and immune responses. Virol J,8, 161.

















