Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, JUlY 1990, p. 2352-23600019-9567/90/072352-09$02.00/0Copyright C) 1990, American Society for Microbiology
Study of Epitopes of Cholera Enterotoxin-Related Enterotoxins byCheckerboard Immunoblotting
MOHAMMAD KAZEMI AND RICHARD A. FINKELSTEIN*Department of Molecular Microbiology and Immunology, School of Medicine,
University of Missouri-Columbia, Columbia, Missouri 65212
Received 23 January 1990/Accepted 11 April 1990
Checkerboard immunoblotting, a versatile new technique for examining multiple antigen and antibodyinteractions simultaneously, was applied in studies of epitopes in the cholera enterotoxin (CT)-relatedheat-labile enterotoxin (LT) family. The purified antigens used included the following: the B-subunit proteinsfrom two CTs (CT-B-1 and CT-B-2), from classical and El Tor biotype strains of Vibrio cholerae, respectively;human LT-B-1 (H-LT-B-1) and porcine LT-B (P-LT-B) derived from LTs produced by Escherichia coli strainsof human (H) and porcine (P) origins, respectively; and genetically engineered chimeric P-LT-Bs with aminoacid substitutions from H-LT-B-1. The antigens were used in native, partially denatured, and CNBr-fragmented forms. The antisera included a variety of mouse monoclonal antibodies against these proteins as
well as polyclonal hyperimmune sera and sera from adult American volunteer vaccinees or convalescents frominduced cholera. Rabbit antisera against synthetic peptides of the CT-B-1 subunit were also used. In some
instances, the effect of GM, ganglioside on antibody binding was evaluated. The reactivity of the monoclonalantibodies was directed primarily against conformational epitopes: some were specific for homologous antigen;some were promiscuously reactive; and some recognized particular related proteins. Individual amino acids(most notably amino acid 46) exerted a dominant effect on epitope formation-in some instances, in a
complementary fashion. Epitope expression was also affected by distant amino acid residues (polar effects).Some reactions were blocked by GM, treatment of the immobilized antigen, indicating that the epitope was
involved in or affected by GM, binding. Polyclonal antibody responses varied within and among animal species.Human serum antitoxic responses were higher in convalescents from induced cholera than in recipients of a
genetically engineered live vaccine, and the convalescent sera (from El Tor biotype cholera patients) generallypreferred CT-B-2 to CT-B-1. The results demonstrate the potential significance of the differences among theseimmunologically related enterotoxins and may help provide direction to further vaccine development.
Despite intensive efforts over the past decades, an effec-tive and economical vaccine against cholera remains to bedeveloped and deployed (for reviews, see references 3, 4,and 10). With the recognition that cholera is a toxin-medi-ated disease, hopes were raised that, as with tetanus anddiphtheria, a toxoid vaccine would be an effective prophy-lactic. Indeed, numerous laboratory studies supported theconcept that antitoxic immunity could protect against exper-imental cholera. However, results of field trials of varioustoxoid vaccines were disappointing, at best. Because of (i)the demonstrated failure of parenterally administered killedwhole-cell bacterins to provide significant and lasting pro-tection, especially in children-the target population inendemic or epidemic areas; (ii) the failure of parenterallyadministered toxoids to provide effective and enduring im-munity; and (iii) the recognition that the disease itself is an
immunizing process, attention has shifted almost entirely toorally administered antigens to stimulate mucosal immuneresponses. These have been either living attenuated mutantsof Vibrio cholerae or cocktails consisting of large amounts ofkilled cells mixed with the B-subunit protein of the choleraenterotoxin (CT).CT was first isolated to homogeneity a little over 20 years
ago (6) from supernatants of cultures of classical biotypeInaba serotype V. cholerae 569B. Because of its hypertoxi-genicity in comparison with other strains of V. cholerae,
* Corresponding author.
strain 569B Inaba has been used exclusively for the produc-tion of the cholera toxoids which have been used in the fieldstudies alluded to above. In 1974, it was recognized that CTproduced by El Tor biotype Ogawa serotype V. cholerae3083 differed immunologically from the CT which had beenisolated earlier from strain 569B Inaba (7). That the immu-nologic differences between 569B CT (CT-1) and CT fromstrain 3083 (CT-2) could be meaningful was later demon-strated by Marchlewicz and Finkelstein (13), who showedthat various polyclonal antisera raised against CT-1 antigensneutralized CT-2 considerably less effectively than theyneutralized the homologous toxin. In that study, whichincluded CT-related heat-labile enterotoxins (LTs) fromEscherichia coli strains of human (H) and porcine (P) origin,it was also pointed out that "enterotoxin-type specific anti-bodies, in some instances, account(ed) for a major portion ofthe neutralizing activity (of the hyperimmune sera used)"(13).We suggest that part of the reason for the failure of cholera
toxoids in previous field trials may reside in the fact thatCT-1 antigen was used against the prevailing epitype of CT,which was primarily CT-2 (R. A. Finkelstein, K. L. Mackey,R. S. Newsham, and M. F. Burks, Abstr. Annu. Meet. Am.Soc. Microbiol. 1988, B103, p. 46). In fact, in the recent fieldstudy of orally administered cholera vaccines in Bangladesh(1), "while neither vaccine induced sufficient protection tojustify its being recommended for public health use in itscurrent form" (18), protection against El Tor biotype V.
2352
Vol. 58, No. 7
on June 9, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

CHOLERA TOXIN-RELATED ENTEROTOXINS
Pro Gln Asn lie5 Thr Asp Lou Cys Alal0 Glu
- - Ser - - Glu - - Ser -- - Thr - - - - - - -
Tyr His
- Arg
Asn Thr15 Gin lle His Thr Leu20- Tyr - -
- - - Ile
Tyr Thr Glu Ser30 Leu
- - - - Met
Ala Gly Lys Arg35 Glu
Phe Lys Asn Gly45 Ala
- - Ser - -
- - - - Glu
Lys Lys Ala 11965 Glu
Thr Phe Gin Val50 Glulie - - - -
Thr - - - -
Arg Met Lys (Asn) Thr- - - Mp -
Val Pro Gly Ser55 Gin- - Ser - -
- - Gly - -
Lou Arg Ile Ala75 Tyr
- - - Thr -
His lie Asp Ser60
Lou Thr Glu Ala80
- - - Thr
Val Glu Lys Leu85 Cys
lie Asp - - -
Val Trp Asn Asn90 Lys Thr Pro His Ala95 lie Ala Ala lie Ser10°
- - Asn Ser - - - -
CHIMERIC PLTBpDL-2 Glu °ZpDL-3 Ala46/Giu102pDL-5 Ala46pDL-7 Ser4/His13
SYNTHETIC PEPTIDESCT-P1 8-20 CT-P4 69-85CT-P2 30-42 CT-P5 75-85CT-P3 50-64 CT-P6 83-97CT-P7 45-64
FIG. 1. Amino acid sequences of B subunits of two CTs (CT-1 and CT-2), H-LT-1, P-LT, chimeras (pDL-2, pDL-3, pDL-5, and pDL-7)and synthetic peptides (CT-P1 through CT-P7). *, The validity of Asn-22 and Asn-70, originally defined by amino acid sequencing, has beenquestioned (16).
cholerae was less than that attained with classical biotypestrains which were present simultaneously.
In this study, we extended our previous observations (5)on the similarities and differences in the CT-related family ofenterotoxins by using a new technique, which we callcheckerboard immunoblotting (CBIB) (9a). CBIB facilitatesthe examination of multiple antigen and antibody interac-tions conveniently and reproducibly with minimal amountsof reactants. Reactions of numerous combinations of CT-related antigens and antibodies were examined to identifythe type, possible location, and role of selected amino acidresidues in the formation of different epitopes.
MATERIALS AND METHODS
Chemicals and reagents. Urea, nitrocellulose membranes,2-mercaptoethanol (2-ME), and other reagents for the immu-noassay were from Bio-Rad Laboratories (Richmond, Cal-if.). Horseradish peroxidase-conjugated immunoglobulinswere purchased from Cooper Biomedical, Inc. (WestChester, Pa.). Reagents for buffers, CNBr, hydrogen perox-ide, and o-dianisidine were obtained from Sigma ChemicalCo. (St. Louis, Mo.). lodoacetic acid was purchased fromICN Biochemicals (Cleveland, Ohio). The Miniblotter 45apparatus was purchased from Immunetics (Cambridge,Mass.). The GM, ganglioside was from Supelco (Bellefont,Pa.).
Antigens. The antigens used in this study were, primarilyas described previously (5, 12), the purified B-subunit pro-teins of CT-1 and CT-2, human LT-1 (H-LT-1), porcine LT(P-LT), and genetically engineered chimeric proteins, pDL-2(P-LT-B in which its Lys-102 was substituted with H-LT-BGlu-102), pDL-3 (P-LT-B with H-LT-B and CT-B Ala-46 and
H-LT-B Glu-102), pDL-5 (P-LT-B with H-LT-B and CT-BAla-46), and pDL-7 (P-LT-B with H-LT-B Ser-4 and H-LT-B-1 and CT-B-1 His-13) (5, 12). (By using the single letteramino acid code, pDL-2, pDL-3, pDL-5, and pDL-7 could bedesignated P-LT-B/Ki02E, P-LT-B/KiO2E and E46A, P-LT-B/E46A, and P-LT-B/T4S and R13H, respectively.)Amino acid sequences of the proteins used are summarizedin Fig. 1. Antigens were applied in the native state, or as
denatured (8 M urea), CNBr-digested, or heated and/orreduced (5% 2-ME) forms. CNBr digestion of antigens was
performed as follows: proteins (0.5 ,umol) were solubilized in1 ml of 0.18 M Tris buffer (pH 8.5) containing 8 M urea and5 mM EDTA, reduced (70 mM 2-ME), carboxymethylated(80 mM iodoacetic acid), precipitated by 1.5 volume of coldacetone, and digested by overnight incubation in a 50-foldmolar excess of CNBr over the protein concentration in 70%formic acid.
Antibodies. Several classes of monoclonal antibodies(MAbs) to CT-1, H-LT-1, pDL-2, pDL-3, pDL-5, and pDL-7used in this study were previously described (5). AdditionalMAbs directed against CT-2 (Finkelstein et al., Abstr. Annu.Meet. Am. Soc. Microbiol. 1988) and S-LT (LT from Sal-monella typhimurium, strain SSU 2635, isolated from poly-myxin extracts of fermentor-grown bacteria by chromatog-raphy with Detoxigel [Pierce Chemical Co., Rockford, Ill.],followed by immunoaffinity chromatography with immobi-lized immunopurified goat anti-H-LT [R. S. Newsham,M. F. Burks, K. L. Mackey, W. C. Katt, S. A. White, andR. A. Finkelstein, Abstr. Annu. Meet. Am. Soc. Microbiol.1988, B102, 46]) were prepared similarly. Polyclonal hyper-immune antisera raised against various CT-related B-subunitproteins in different animal species (e.g., rabbit, mouse,
goat, and horse) were also used. Hyperimmune rabbit anti-
CT-1 ThrCT-2 -
H-LT-1 AlaP-LT -
(Asn) LysAsp -
CT-1 AsnCT-2 -
H-LT-1 -P-LT -
lle Phe25 Ser
Leu -
CT-1 ThrCT-2 -
H-LT-1 -P-LT -
CT-1 GlnCT-2 -
H-LT-1 -P-LT -
Met Ala
- Val
Ile lie40
CT-1 LysCT-2 -
H-LT-1 -P-LT -
CT-1 MetCT-2 -
H-LT-1 -P-LT -
Ala Asn
Glu -Lys -
2353VOL. 58, 1990
on June 9, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

2354 KAZEMI AND FINKELSTEIN
NATIVE CmlICr14214.T4PLY-Sp01.-2p01.4p01.4pDL.7
+UREA CTB2HLT-SPLYSP.-2
p01.
Z oL-7+CNBr c
PLT-SpDL-
pOLUpOL.-6
+2ME t7'HLT4
p014pOL25pOL-
tHEAT ""CM-2
pOXLpDA-4
P01-?
MONOCLONAL ANTIBODYN 1iJ Ja a
0.iaCT-1 aCT-2 aHLT d apDL-3 d apDL.7REVta I n t, "^IV* Y MUAI A 1 I AN I VEVI MmlV V
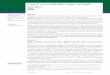
FIG. 2. CBIB reactions of various classes (defined by their patterns of reactivity [5]) of mouse MAbs (left to right) raised against CT-1,CT-2, H-LT-1, pDL-2, pDL-3, pDL-5, pDL-7, and S-LT with B-subunit antigens (100 ng per spot) CT-B-1, CT-B-2, H-LT-B-1, P-LT-B,pDL-2, pDL-3, pDL-5, and pDL-7 (top to bottom) in native, urea-denatured, CNBr-fragmented, 2-ME-treated, and heated forms. Positivecontrols for reactivity of CNBr fragments are given in Fig. 4.
sera against synthetic peptides (CTPs) representing differentportions of CT-B-1 (8, 9) were kindly provided by Chaim 0.Jacob (Stanford University). Several pre- and postimmuni-zation or pre- and postchallenge sera from adult volunteercholera vaccinees or volunteer patients with induced cholera(generously provided by James B. Kaper and Myron M.Levine of the Center for Vaccine Development, Universityof Maryland School of Medicine, Baltimore) were alsoexamined for their reactivities.CBIB assay procedure. The checkerboard immunoblotting
(CBIB) procedure used in this study was described in detailelsewhere (9a). Briefly, antigens, diluted in coating buffer (15mM sodium carbonate, 35 mM sodium bicarbonate [pH9.6]), were immobilized on a solid-phase membrane in par-allel lanes by using a Miniblotter apparatus (Miniblotter 45;Immunetics). Coating buffer contained 8 M urea when urea-denatured antigens were used. Nonspecific binding siteswere saturated, normally at 4°C overnight, with 5% nonfatdry milk in phosphate-buffered saline (PBS) containing0.05% Tween 20 (PBST). Primary antibodies were thenapplied in lanes perpendicular to the antigens for 1 to 2 h atroom temperature, and the reactions were developed withappropriately labeled secondary antibody and substrate (us-ing the same conditions). After washing, substrate (200 puM)in 10 mM Tris hydrochloride buffer (pH 7.4) and H202 (0.1%[vol/vol]) were added. Reactions were stopped by rinsingwith H20. Positive reactions appear as colored squares atthe intersection of antibody and antigen, giving a checker-
board appearance to the developed blot. All antibodies werediluted appropriately in PBST containing 0.5% nonfat drymilk. In some assays, the effect of GM, ganglioside (the hostcell membrane receptor for the CT-related toxins) treatmentof the antigens on their reactivity with antibodies was tested.In those GM,' assays, after the nonspecific binding siteswere blocked, the blot was incubated for 2 h at roomtemperature in a GM, ganglioside solution (20 ,ug/ml inPBST) and then washed with PBST 3 to 4 times before beingreacted with primary antibodies.
RESULTS
Reactions of MAbs. Reactions of selected MAbs withvarious B-subunit proteins, in various forms, in our libraryare shown in Fig. 2. It should be noted that the MAbs were
generally directed against conformational epitopes; the re-
actions were very much weakened or eliminated when theantigens were denatured by urea or heat, and none of theMAbs reacted with CNBr cleavage products. Treatment ofnative antigens with 2-ME alone, under these conditions, didnot significantly alter their reactions. It may be seen thatsome of the MAbs (aCT-1 class I, aCT-2 class I, apDL-5class II, and apDL-7 class V) exhibited exquisite specificityand reacted with their homologous B-subunit proteins exclu-sively. Class II aCT-i and aCT-2 MAbs reacted only withCT epitypes, while aH-LT class III, as well as apDL-7 classII, reacted only with the LT-related antigens. The four aCT
aSLT%A"^O "-g* EU UUS U 11 U1:.5 Rivgi
-| - 111 KIME X a lXIl
- 3X Ue S 33 U-SE RUII iq
U. SE U U3.-3
aUa'
55533 mU *EW £ aUWtlS~~~~tlu urns ,M
-1 1*31 e,4 5
INFECT. IMMUN.
on June 9, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
~~~~~~CHOLERA TOXIN-RELATED ENTEROTOXINS 2355
MONOCLONAL ANTIBODY
C*4 LO~II I
j j~~.0 ~0
a
(xCT-2 alILT oxpDL-3 %PDL.7toaK MAVVIVRVIISAV1 I U1NA IN V IA I N ovW vjI
oxSLTI MUA I VVvI
ft
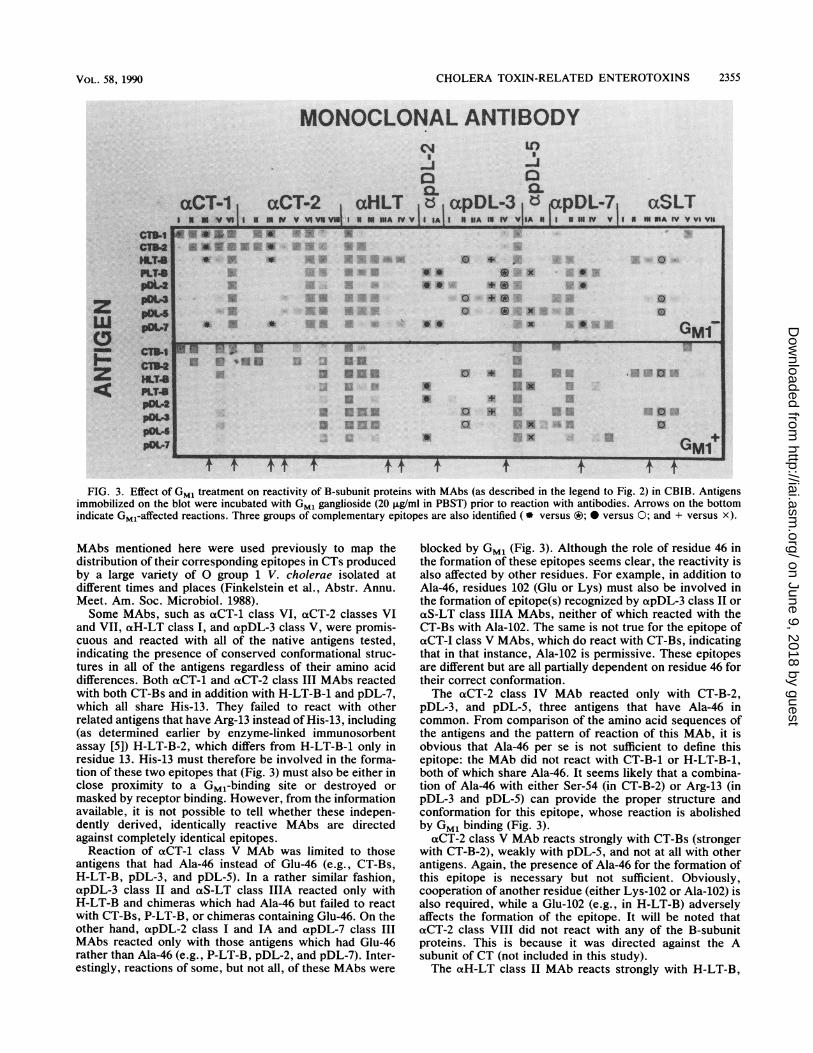
FIG. 3. Effect of Gm, treatment on reactivity of B-subunit proteins with MAbs (as described in the legend to Fig. 2) in CBIB. Antigensimmobilized on the blot were incubated with Gm, ganglioside (20 ~Lg/ml in PBST) prior to reaction with antibodies. Arrows on the bottom
indicate Gm,-affected reactions. Three groups of complementary epitopes are also identified (9 versus ®; versus 0; and + versus x).
MAbs mentioned here were used previously to map the
distribution of their corresponding epitopes in CTs produced
by a large variety of 0 group 1 V. cholerae isolated at
different times and places (Finkelstein et al., Abstr. Annu.
Meet. Am. Soc. Microbiol. 1988).
Some MAbs, such as aCT-1 class VI, aCT-2 classes VI
and VII, aHl-LT class I, and apDL-3 class V, were promis-cuous and reacted with all of the native antigens tested,
indicating the presence of conserved conformational struc-
tures in all of the antigens regardless of their amino acid
differences. Both aCT-i and aCT-2 class III MAbs reacted
with both CT-Bs and in addition with H-LT-B-1 and pDL-7,which all share His-13. They failed to react with other
related antigens that have Arg-13 instead of His-13, including
(as determined earlier by enzyme-linked immunosorbent
assay [5]) H-LT-B-2, which differs from H-LT-B-1 only in
residue 13. His-13 must therefore be involved in the forma-
tion of these two epitopes that (Fig. 3) must also be either in
close proximity to a Gm,-binding site or destroyed or
masked by receptor binding. However, from the information
available, it is not possible to tell whether these indepen-
dently derived, identically reactive MAbs are directed
against completely identical epitopes.Reaction of aCT-i class V MAb was limited to those
antigens that had Ala-46 instead of Glu-46 (e.g., CT-Bs,
H-LT-B, pDL-3, and pDL-5). In a rather similar fashion,
apDL-3 class and cxS-LT class IIIA reacted only with
H-LT-B and chimeras which had Ala-46 but failed to react
with CT-Bs, P-LT-B, or chimeras containing Glu-46. On the
other hand, oapDL-2 class I and IA and apDL-7 class III
MAbs reacted only with those antigens which had Glu-46
rather than Ala-46 (e.g., P-LT-B, pDL-2, and pDL-7). Inter-
estingly, reactions of some, but not all, of these MAbs were
blocked by Gm, (Fig. 3). Although the role of residue 46 in
the formation of these epitopes seems clear, the reactivity is
also affected by other residues. For example, in addition to
Ala-46, residues 102 (Glu or Lys) must also be involved in
the formation of epitope(s) recognized by oapDL-3 class II or
otS-LT class IIIA MAbs, neither of which reacted with the
CT-Bs with Ala-102. The same is not true for the epitope of
aCT-I class V MAbs, which do react with CT-Bs, indicatingthat in that instance, Ala-102 is permissive. These epitopesare different but are all partially dependent on residue 46 for
their correct conformation.
The aCT-2 class IV MAb reacted only with CT-B-2,
pDL-3, and pDL-5, three antigens that have Ala-46 in
common. From comparison of the amino acid sequences of
the antigens and the pattern of reaction of this MAb, it is
obvious that Ala-46 per se is not sufficient to define this
epitope: the MAb did not react with CT-B-i or H-LT-B-1,
both of which share Ala-46. It seems likely that a combina-
tion of Ala-46 with either Ser-54 (in CT-B-2) or Arg-13 (in
pDL-3 and pDL-5) can provide the proper structure and
conformation for this epitope, whose reaction is abolished
by Gm, binding (Fig. 3).
oaCT-2 class V MAb reacts strongly with CT-Bs (strongerwith CT-B-2), weakly with pDL-5, and not at all with other
antigens. Again, the presence of Ala-46 for the formation of
this epitope is necessary but not sufficient. Obviously,
cooperation of another residue (either Lys-102 or Ala-102) is
also required, while a Glu-102 (e.g., in H-LT-B) adverselyaffects the formation of the epitope. It will be noted that
aCT-2 class VIII did not react with any of the B-subunit
proteins. This is because it was directed against the A
subunit of CT (not included in this study).The axH-LT class II MAb reacts strongly with H-LT-B,
aCT-I1I a lvy I,
CT".NLT4PLT41
pm74'ow4
PO"4
z
wU
z
N 'U 0 9,'~~~~~~~~~~~~~~~~~0f
*U9p ** x ~~~~~~GMf+It maA[J~
VOL. 58, 1990
t
on June 9, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

2356 KAZEMI AND FINKELSTEIN
CT-B-2 (but not CT-B-1), pDL-3, and pDL-5, all of whichpossess Ala-46. Once again, residue 46 and one or moreother residue(s), that in this case are only present in CT-B-2and LT-related antigens (Tyr-18, Asp-22, or Asp-70), arerequired for the epitope. Since the validity of Asp-22 andAsp-70 has been questioned (16), it is likely that Tyr-18 is theother residue involved in this epitope. Class IIIA otH-LTMAb, like class III, reacts only with LT-related antigens, butunlike class III reacts much more strongly to the homolo-gous protein. In contrast to class III, reaction of thisantibody is blocked by GM, (Fig. 3). Class IV otH-LT MAbreacted with H-LT-B-1 and pDL-7 but not with other anti-gens. (This MAb was previously shown not to react withH-LT-B-2 [5].) The two reactive antigens differ from otherantigens by having both Ser-4 and His-13 residues. Thesetwo residues can therefore be implied to be necessary for theformation of this epitope, which is affected by GM, binding.Interestingly, however, cxpDL-7 class V MAb, which isspecific for the homologous chimera with Ser-4 and His-13,does not react with H-LT-B-1 and is not blocked by GM,;other residues in the P-LT-B backbone of pDL-7 are influ-encing its reactivity.Among the MAbs to chimeric antigens not mentioned
earlier is apDL-3 class I, which is weakly reactive withpDL-2 and pDL-3 only. Both of these antigens are distinctfrom others by having Thr-4 (from its P-LT-B backbone) andGlu-102 residues, which together appear to define thisepitope. The reactions of apDL-3 class IIA with H-LT-B,pDL-3, and pDL-5 are similar to, but weaker than, those ofclass II. Peculiarly, the reactions ofaS-LT classes III and IVare stronger in the GM,' assay (Fig. 3), which means that theepitope is more exposed upon receptor binding. The reac-tions of opDL-3 class III MAb are interesting in that itprimarily reacts with those antigens that have Glu-102 (H-LT-B, pDL-2, and pDL-3). In contrast to these reactions,apDL-5 class IA reacts only with Lys-102-containing anti-gens. This phenomenon, within the LT-related antigens, isinteresting as changing residue 102 from P-LT-B Lys toH-LT-B Glu results in the formation of some epitopes (e.g.,the one recognized by apDL-3 class III) and the disappear-ance of others (e.g., the ones recognized by apDL-5 classIA). As antibodies to these epitopes react with antigens in acomplementary manner, we call them complementaryepitopes. Other complementary-type epitopes can also beseen (Fig. 3), such as those recognized by apDL-2 classes Iand IA, as well as apDL-7 class III versus apDL-3 class IIand oaS-LT class IIIA.No purified S-LT antigen was available to include in this
study; therefore, reactivities of aS-LT MAbs are defined interms of their reactions with other related antigens. TheoS-LT class II reacted only with H-LT-B, while the class Vreaction was only to CT-B-1. This indicates that S-LTprobably contains distinct structural elements from bothCT-B-1 and H-LT-B. Both class III and class IV reactedonly with H-LT-B and pDL-3, with their Ala-46 and Glu-102distinguishing them from other related antigens. The uniqueconformation resulting from the presence of these tworesidues presumably defines the epitopes and may predictresidues in S-LT.
Figure 3 illustrates the effects of adding GM, ganglioside tothe immobilized antigens prior to their reaction with theprimary antibodies. As mentioned above, some of the reac-tions were weakened or obliterated by treatment with GM1,thus implying either that the epitopes are at or near a sitewhich contributes to GM, binding or that receptor bindingresults in conformational modifications of protein that de-
stroy the epitope. This is particularly noticeable with aCT-1class III and aCT-2 class III as well as aCT-2 class VIMAbs, oxH-LT class IIIA and IV MAbs, and apDL-2 classIA, which otherwise strikingly resembles apDL-2 class I anda-pDL-7 class III. From these results and others, it is likelythat several different regions of the B-subunit structurecontribute cooperatively to GM, binding and that the sitesinvolved in, or affected by, GM, binding may be only verysubtly different from those which are not.
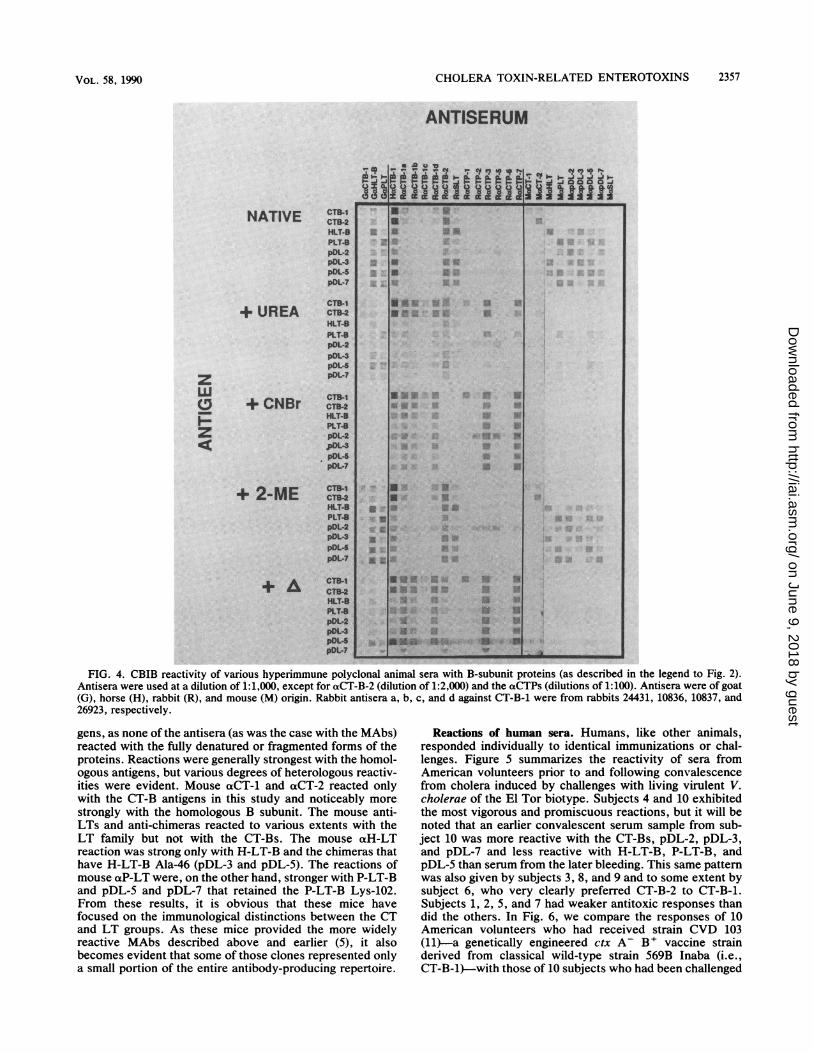
Reactions of hyperimmune polyclonal antisera. In contrastto the observations with MAbs summarized above, poly-clonal antisera frequently reacted with denatured forms ofthe antigen. Shown in Fig. 4 are the reactions of a variety ofpolyclonal hyperimmune sera against eight B-subunit pro-teins in various forms. A number of statements can be madeabout these reactions, and we will cover only a selective fewhere. First, despite the immunological relatedness of all ofthe proteins studied, even hyperimmune sera reacted withsome degree of specificity and, generally, reactions werestronger with homologous antigen. It will also be noted thatindividual animals gave different responses to the sameantigens. Thus, for example, goat aP-LT reacted veryweakly with the CT-Bs and H-LT-B, while goat aH-LT-Breacted better with H-LT-B, pDL-3, pDL-5, and pDL-7.Intact protein conformation was essential for the strongestreactions of these two antisera. The horse aCT-B-1 (equineanti-choleragenoid [2]) was promiscuous in its activity butreacted most strongly with the CT-Bs and with chimeraswith Ala-46 (pDL-3 and pDL-5). Although homologousreactions were strong even with denatured and fragmentedantigens, heterologous reactions were weaker with the al-tered forms of the proteins. Serum samples from four rabbitsimmunized with CT-B-1, interestingly, appeared to preferparticular denatured forms to the native form of the antigensbut with various degrees of reactivity. This was not the case,however, for sera from rabbits immunized with CT-B-2(which reacted promiscuously with native antigens) or S-LT(which, like goat aH-LT-B, reacted better with H-LT-B,pDL-3, pDL-5, and pDL-7). Neither of these two antiserareacted with CNBr-fragmented antigens.The rabbit xCTP sera (Fig. 4) (provided earlier by Chaim
0. Jacob [5, 8, 9]) were raised against synthetic peptidesrepresenting portions of CT-B-1. Anti-CTP-1 (raised againstresidues 8 through 20 of CT-B-1) was very specific forCT-B-1. This suggests that a CT-B-1-specific residue in thatregion must be definitive for reactivity with this antiserum.As the serum does not react with CT-B-2, and the onlydifference between CT-B-1 and CT-B-2 in that region is Hisversus Tyr at position 18, the definitive residue must beHis-18. Apparently, this epitope, in the native antigen, is nottotally exposed as the native antigen is not reactive. Anti-CTP-3 and otCTP-7, raised against a conserved region of theB-subunit protein (consisting of residues 50 through 64 and45 through 64, respectively), reacted vigorously with all ofthe CNBr-fragmented and heat-denatured antigens tested.However, little reactivity was seen with the native antigensin this study. Although this sequence contains, or is part of,a surface epitope, as shown by previous neutralization andprotection studies (8), the antigen(s) is apparently moreexposed when the protein is unfolded or, possibly, when itsurrounds the A subunit in the holotoxin (unpublished ob-servations).The mouse antisera used for reactions shown in Fig. 4
were from the same mice as the MAbs described earlier (5).Apparently, the immune responses in these mice wereprimarily directed toward the native conformation of anti-
INFECT. IMMUN.
on June 9, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
CHOLERA TOXIN-RELATED ENTEROTOXINS 2357
ANTISERUM
NATIVE CTlHLT-BPLT4PIT-2p01-3p01.apOL-5pDL-7
+ UREA cm-2HLT-BPLT4pDL-2
pDL5
O + CNBr CTB-PT-BPLT4
,pDL4pDL-7
+ 2-ME CT8HLT4PLT4pDL,2
p04Lpow4pDL-7
CTB-ip014
CT -2HLT4PLT4
p0O4p014
p01-2pDL-2HIT-UPILTB
*aA X
FIG. 4. CBIB reactivity of various hyperimmune polyclonal animal sera with B-subunit proteins (as described in the legend to Fig. 2).Antisera were used at a dilution of 1:1,000, except for oaCT-B-2 (dilution of 1:2,000) and the aCTPs (dilutions of 1:100). Antisera were of goat(G), horse (H), rabbit (R), and mouse (M) origin. Rabbit antisera a, b, c, and d against CT-B-1 were from rabbits 24431, 10836, 10837, and26923, respectively.
gens, as none of the antisera (as was the case with the MAbs)reacted with the fully denatured or fragmented forms of theproteins. Reactions were generally strongest with the homol-ogous antigens, but various degrees of heterologous reactiv-ities were evident. Mouse aCT-1 and aCT-2 reacted onlywith the CT-B antigens in this study and noticeably more
strongly with the homologous B subunit. The mouse anti-LTs and anti-chimeras reacted to various extents with theLT family but not with the CT-Bs. The mouse acH-LTreaction was strong only with H-LT-B and the chimeras thathave H-LT-B Ala-46 (pDL-3 and pDL-5). The reactions ofmouse aP-LT were, on the other hand, stronger with P-LT-Band pDL-5 and pDL-7 that retained the P-LT-B Lys-102.From these results, it is obvious that these mice havefocused on the immunological distinctions between the CTand LT groups. As these mice provided the more widelyreactive MAbs described above and earlier (5), it alsobecomes evident that some of those clones represented onlya small portion of the entire antibody-producing repertoire.
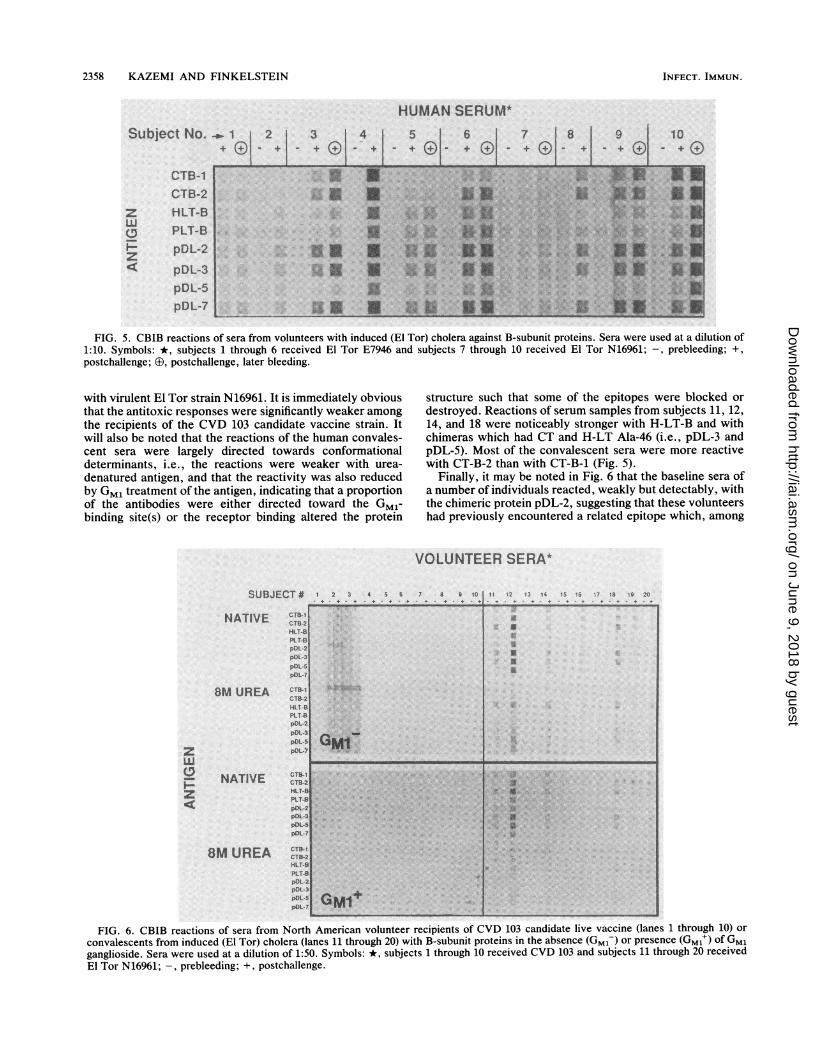
Reactions of human sera. Humans, like other animals,responded individually to identical immunizations or chal-lenges. Figure 5 summarizes the reactivity of sera fromAmerican volunteers prior to and following convalescencefrom cholera induced by challenges with living virulent V.cholerae of the El Tor biotype. Subjects 4 and 10 exhibitedthe most vigorous and promiscuous reactions, but it will benoted that an earlier convalescent serum sample from sub-ject 10 was more reactive with the CT-Bs, pDL-2, pDL-3,and pDL-7 and less reactive with H-LT-B, P-LT-B, andpDL-5 than serum from the later bleeding. This same patternwas also given by subjects 3, 8, and 9 and to some extent bysubject 6, who very clearly preferred CT-B-2 to CT-B-1.Subjects 1, 2, 5, and 7 had weaker antitoxic responses thandid the others. In Fig. 6, we compare the responses of 10American volunteers who had received strain CVD 103(11)-a genetically engineered ctx A- B+ vaccine strainderived from classical wild-type strain 569B Inaba (i.e.,CT-B-1)-with those of 10 subjects who had been challenged
* U5
~~~~~~~~~4.
2 U USE .:
UU Ui U
_. .. o. r.
S~~~ U
W
-U:
US.us ~ U * *
.5k:' 5k" *'W -
i 9 .-
VOL. 58, 1990
UX0 47 19 N.. 4y 0- ).- Nd dll-
I still
on June 9, 2018 by guesthttp://iai.asm
.org/D
ownloaded from
2358 KAZEMI AND FINKELSTEIN
HUMAN SERUM*Subject No. -_
CTB-1CTB-2
z HLT-Bw
PLT-B
pDL-2z
< pDL-3
pDL-5pDL-7
3 4 5 6 7 8 9 10+ D(3 + - G+G) - +(i) .E - + - + (i - + (i)
I -----I.........
FIG. 5. CBIB reactions of sera from volunteers with induced (El Tor) cholera against B-subunit proteins. Sera were used at a dilution of1:10. Symbols: *, subjects 1 through 6 received El Tor E7946 and subjects 7 through 10 received El Tor N16961; -, prebleeding; +,
postchallenge; G3, postchallenge, later bleeding.
with virulent El Tor strain N16961. It is immediately obviousthat the antitoxic responses were significantly weaker amongthe recipients of the CVD 103 candidate vaccine strain. Itwill also be noted that the reactions of the human convales-cent sera were largely directed towards conformationaldeterminants, i.e., the reactions were weaker with urea-
denatured antigen, and that the reactivity was also reducedby GM, treatment of the antigen, indicating that a proportionof the antibodies were either directed toward the GMl-binding site(s) or the receptor binding altered the protein
structure such that some of the epitopes were blocked or
destroyed. Reactions of serum samples from subjects 11, 12,14, and 18 were noticeably stronger with H-LT-B and withchimeras which had CT and H-LT Ala-46 (i.e., pDL-3 andpDL-5). Most of the convalescent sera were more reactivewith CT-B-2 than with CT-B-1 (Fig. 5).
Finally, it may be noted in Fig. 6 that the baseline sera ofa number of individuals reacted, weakly but detectably, withthe chimeric protein pDL-2, suggesting that these volunteershad previously encountered a related epitope which, among
VOLUNTEER SERA*
SUBJECT#
NATIVE CTs-1
HLT-BPILT-8p0L-2
pOL-5pDL- 7
8AW UREA CTWI
HLT-BPLT-8p0DL2pOL-3pOL-5 (pDL-7
CTB-I
NATIVE CTB-2HLT-UPlT-UpDL-2pDL-3pOL-5poL- 7
8M UREA CTW2HLT E
P1 T-8pOL-2pOL-3pOL-5pOL -X
2 3 4 5 6 a 910U 11 12 73 14 15 16 17 18 !9 2
FIG. 6. CBIB reactions of sera from North American volunteer recipients of CVD 103 candidate live vaccine (lanes 1 through 10) or
convalescents from induced (El Tor) cholera (lanes 11 through 20) with B-subunit proteins in the absence (GM,-) or presence (GM1+) of GM1ganglioside. Sera were used at a dilution of 1:50. Symbols: *, subjects 1 through 10 received CVD 103 and subjects 11 through 20 receivedEl Tor N16961; -, prebleeding; +, postchallenge.
------------*X ;
s ~~~~~~~~~~~~~~~......... ''.,
" "-
'~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.
zw
z
a
.m .......... =.............. . .i ... ... U. ::
aM4.
GMIt~~~~~~. .
INFECT. IMMUN.
1.
7
on June 9, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

CHOLERA TOXIN-RELATED ENTEROTOXINS 2359
the proteins examined, is unique to pDL-2 (which differsfrom the others in having Glu-102 on a P-LT-B backbone).
DISCUSSION
By using CBIB, we have demonstrated that mouse MAbsraised against the purified proteins of various members of theCT-related enterotoxin family, as well as against geneticallyengineered chimeric B-subunit proteins, exhibited multiplepatterns of reactivity primarily against conformational (dis-continuous) epitopes; some were highly specific and recog-nized only the homologous antigen, some were completelypromiscuous (directed against conserved epitopes), andsome recognized a few closely related proteins. Of these,some reacted with patterns which indicated that individualamino acids had a dominant role in the formation of partic-ular epitopes-in some instances in a complementary man-
ner, i.e., amino acid substitutions simultaneously created anepitope while eliminating the reactivity of another. Thisinvolvement of one or more residues in the formation of an
epitope does not necessarily mean that the residue is part ofthe epitope. In some cases, epitope expression was affectedby amino acid residues which were sequentially distant fromthe dominant residue (polar effects). The results, particularlythose derived from the use of the chimeric B-subunit pro-teins and their antibodies, emphasized the dominant role ofresidue 46 in the formation of a good number of epitopes.However, epitopes dominated by residue 46 were, in manyinstances, affected by residues which were sequentially farfrom it, upstream or down.Some of the reactions between MAbs and the B-subunit
proteins were reduced or eliminated by GM, treatment of theimmobilized antigen, indicating that either the epitope wasinvolved in GM, binding or that receptor binding exertsstructural modifications of the protein that result in altera-tion of the epitope region. Curiously, none of the mousepolyclonal serum samples or MAbs in our library recognizedcontinuous (sequence-related) epitopes, as evidenced bytheir failure to react with CNBr fragments.The reactions of the promiscuous MAbs indicated that
there are conserved conformational regions in these antigensthat are not altered even by other sequence differencesbetween them. Some of these epitopes are distinct, as GM,saturation of antigens differentially affected their reactions.In this regard, the reaction of aCT-1 class VI MAb (Fig. 3) isinteresting since GM, blocks its reaction with all antigens butCT-B-1. This suggests that the Gm1-binding sites may beslightly different in different antigens.
Polyclonal immune responses varied among and withinanimal species. Whereas the mice recognized primarilyconformational (discontinuous) epitopes, rabbits frequentlyrecognized sequence-determined epitopes which were un-masked by denaturation of the protein. It is not clearwhether these represent species differences or differences inthe immune responses to the immunizing regimens. Themouse MAbs were originally identified primarily by enzyme-linked immunosorbent assays with native proteins, but eventhe mouse polyclonal sera from the same mice did not reactwell with denatured antigens. Antisera against syntheticpeptides, as might be expected, recognized sequence-de-fined epitopes. The reactions of the polyclonal antisera, inmany instances, emphasized the specificity, rather than therelatedness, of the CT-related toxin proteins examined,although some sera were promiscuous in their reactivity.
Finally, the immune response of humans varies amongindividuals. People who were convalescent from El Tor
cholera tended to have a stronger response to CT-2 than toCT-1. Volunteers who had received the CVD 103 A- B+candidate vaccine strain clearly had weaker antitoxic im-mune responses than did volunteers who had recovered frominduced cholera. CVD 103 is a derivative of strain 569BInaba (11). In 1974, we (7) isolated a hypotoxinogenicmutant, M13, of strain 569B which was shortly thereafterfound to induce protection against induced cholera in volun-teers. The protective effect was of the same magnitude (17)as was reported recently for CVD 103 (11), which is substan-tially less than that observed following convalescence frominduced cholera in volunteers. In an earlier study in rabbits(15), we were unable to demonstrate significant adherenceby strain 569B to the small bowel. Migasena et al. (14) haverecently reported that CVD 103-HgR (a further derivative ofCVD 103) could be recovered from only 2 of 12 Thaivolunteer vaccinees. These observations imply that therelative inability of strain 569B and its derivatives to colo-nize the small bowel may be reflected in a reduced immuneresponse and suggest that other attenuated strains of V.cholerae would make better vaccines. In such further en-deavors, the immunologic differences between CT-1 andCT-2 should be considered.
In conclusion, we have analyzed a variety of immuneresponses to various forms of CT-related enterotoxin B-subunit proteins by using a new technique which we callCBIB. This rapid and reproducible technique permits thesimultaneous analysis of large numbers of antigen and anti-body combinations with minimal amounts of reagents. Theresults are available within 24 h and are in the form of apermanent record. The observations of this study, althoughadmittedly incomplete, should be considered, together withthe earlier results of Jacob et al. (8, 9) with syntheticpeptides, in future efforts to construct synthetic peptideswhich may be used as vaccines against cholera and theCT-related enterotoxic enteropathies. Studies in progresswith sequential overlapping synthetic hexapeptides of theB-subunit protein are delineating continuous (sequence-related) epitopes, which may constitute portions of confor-mational epitopes described in this work (Kazemi and Fin-kelstein, unpublished data).
ACKNOWLEDGMENTS
This work was supported in part by Public Health Service grantsAl 16776 and Al 17312 from the National Institute of Allergy andInfectious Diseases.We appreciate the contributions and assistance of Mary Boesman-
Finkelstein, Allen Maddy, Joe Flynn, and Farhad Gharib duringportions of the study.
LITERATURE CITED1. Clemens, J. D., J. R. Harris, D. A. Sack, J. Chakraborty, F.
Ahmed, B. F. Stanton, M. U. Khan, B. A. Kay, N. Hoda, M. R.Khan, M. Yunus, M. R. Rao, A.-M. Svenerholm, and J. Holm-gren. 1988. Field trial of oral cholera vaccines in Bangladesh:results of one year of follow-up. J. Infect. Dis. 158:60-69.
2. Finkelstein, R. A. 1970. Monospecific equine antiserum againstcholera exo-enterotoxin. Infect. Immun. 2:691-697.
3. Finkelstein, R. A. 1984. Cholera, p. 107-136. In R. Germanier(ed.), Bacterial vaccines. Academic Press, Inc., New York.
4. Finkelstein, R. A. 1988. Cholera, the cholera enterotoxins, andthe cholera enterotoxin-related enterotoxin family, p. 85-102. InP. Owen and T. J. Foster (ed.), Immunochemical and moleculargenetic analysis of bacterial pathogens. Elsevier Science Pub-lishers, Amsterdam, The Netherlands.
5. Finkelstein, R. A., M. F. Burks, A. Zupan, W. S. Dallas, C. 0.Jacob, and D. S. Ludwig. 1987. Epitopes of the cholera family ofenterotoxins. Rev. Infect. Dis. 9:544-561.
VOL. 58, 1990
on June 9, 2018 by guesthttp://iai.asm
.org/D
ownloaded from

2360 KAZEMI AND FINKELSTEIN
6. Finkelstein, R. A., and J. J. LoSpalluto. 1969. Pathogenesis ofexperimental cholera: preparation and isolation of choleragenand choleragenoid. J. Exp. Med. 130:185-202.
7. Finkelstein, R. A., M. L. Vasil, and R. K. Holmes. 1974. Studieson toxinogenesis in Vibrio cholerae. I. Isolation of mutants withaltered toxinogenicity. J. Infect. Dis. 129:117-123.
8. Jacob, C. O., R. Arnon, and R. A. Finkelstein. 1986. Immunitytowards heat-labile enterotoxins of porcine and human Esche-richia coli strains achieved with synthetic cholera toxin pep-tides. Infect. Immun. 52:562-567.
9. Jacob, C. O., M. Sela, and R. Arnon. 1983. Antibodies againstsynthetic peptides of the B-subunit of cholera toxin: cross-reaction and neutralization of the toxin. Proc. Natl. Acad. Sci.USA 80:7611-7615.
9a.Kazemi, M., and R. A. Finkelstein. 1990. Checkerboard immu-noblotting (CBIB): an efficient, rapid, and sensitive method ofassaying multiple antigen/antibody cross-reactivities. J. Immu-nol. Methods 128:143-146.
10. Levine, M. M., J. B. Kaper, R. E. Black, and M. L. Clements.1983. New knowledge on pathogenesis of bacterial entericinfections as applied to vaccine development. Microbiol. Rev.47:510-550.
11. Levine, M. M., D. Herrington, G. Losonsky, B. Tall, J. B. Kaper,J. Ketley, C. 0. Tacket, and S. Cryz. 1988. Safety, immunoge-nicity and efficacy of recombinant live oral cholera vaccines,CVD 103 and CVD 103-HgR. Lancet ii:467-470.
12. Ludwig, D. S., R. A. Finkelstein, A. E. Karu, W. S. Dallas, E. R.Ashby, and G. K. Schoolnik. 1987. Anti-idiotypic antibodies asprobes of protein active sites: application to cholera toxinB-subunit. Proc. Natl. Acad. Sci. USA 84:3673-3677.
13. Marchlewicz, B. A., and R. A. Finkelstein. 1983. Immunologicdifferences among the cholera/coli family of enterotoxins. Di-agn. Microbiol. Infect. Dis. 1:129-138.
14. Migasena, S., P. Pitisuttitham, B. Prayurahong, P. Suntharasa-mai, W. Supanaranond, V. Desakorn, U. Vongsthongsri, B. Tall,J. Ketley, G. Losonsky, S. Cryz, J. B. Kaper, and M. M. Levine.1989. Preliminary assessment of the safety and immunogenicityof live oral cholera vaccine strain CVD 103-HgR in healthy Thaiadults. Infect. Immun. 57:3261-3264.
15. Nelson, E. T., J. D. Clements, and R. A. Finkelstein. 1976. Vibriocholerae adherence and colonization in experimental cholera:electron microscopic studies. Infect. Immun. 14:527-547.
16. Takao, T., H. Watanabe, and Y. Shimonishi. 1985. Facileidentification of protein sequences by mass spectrometry. Eur.J. Biochem. 146:503-508.
17. Woodward, W. E., R. H. Gilman, R. B. Hornick, J. P. Libonati,and R. A. Cash. 1976. Efficacy of a live oral cholera vaccine inhuman volunteers. Dev. Biol. Stand. 33:108-112.
18. World Health Organization. 1989. Diarrhoeal diseases controlprogramme: report of tenth meeting of the technical advisorygroup (Geneva, 13-17 March 1989). World Health Organization/CDD/89.32:1-21.
INFECT. IMMUN.
on June 9, 2018 by guesthttp://iai.asm
.org/D
ownloaded from










![Phosphorylation-dependentepitopes antibodies Alzheimertau · ment antibodies SMI31, SMI34, SMI35, or SMI310 (with phosphorylated epitopes) and SM133 [unphosphorylated epitopes (3)]](https://img.pdfslide.net/doc/110x75/5e62d2f4d3d32f22a55ed9e3/phosphorylation-dependentepitopes-antibodies-alzheimertau-ment-antibodies-smi31.jpg)