Embed Size (px)
Citation preview

32 Journal of Basic Microbiology 2009, 49, 32–41
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
Review Article
Sub-proteome analysis in the green flagellate alga Chlamydomonas reinhardtii
Volker Wagner*, Jens Boesger* and Maria Mittag
Institute of General Botany and Plant Physiology, Friedrich-Schiller-University Jena, 07743 Jena, Germany * Both authors have contributed equally
In the past years, research on the flagellate unicellular alga Chlamydomonas reinhardtii has
entered a new era based on the availability of its complete genome. Since this green alga can be
grown relatively easy in a short time-range, sufficient biological material is available to
efficiently establish biochemical purification procedures of sub-cellular fractions. Combined
with the available genome sequences, this paved the way to perform analysis of specific
sub-proteomes by mass spectrometry. In this review, several approaches that provided
comprehensive lists of components of certain sub-cellular compartments and their biological
relevance will be described. These include proteins of chloroplast ribosomes, of flagella, of the
eyespot as well as posttranslational and environmentally modified sub-proteomes. The power
of such proteome approaches lies in the identification of novel components and modifications
of a given sub-proteome that have not been discovered before. Information is usually gained at
a large scale and is very valuable to further understand biological processes of a given cellular
sub-compartment. But clearly the arduous task has then to be performed to further analyze
the function of specific proteins/genes by RNA interference technology, mutant analyses or
methods for identifying the protein interaction network within a sub-proteome.
Keywords: Chlamydomonas reinhardtii / Posttranslational Modifications / Sub-proteomes
Received: September 11, 2008; accepted: October 14, 2008
DOI 10.1002/jobm.200800292
Introduction*
The green unicellular alga Chlamydomonas reinhardtii,
whose nuclear, chloroplast and mitochondrial genomes
have been fully sequenced [1–3], has served for many
years as a model to study certain cellular processes and
organelles in depth. These include the biogenesis and
function of the chloroplast, its photosynthetic machin-
ery, the composition of flagella and the basal body for
their relevance to human diseases, metabolism, nutri-
ent acquisition as well as components of the light sig-
naling pathway and of its circadian clock [1, 4, 5]. A
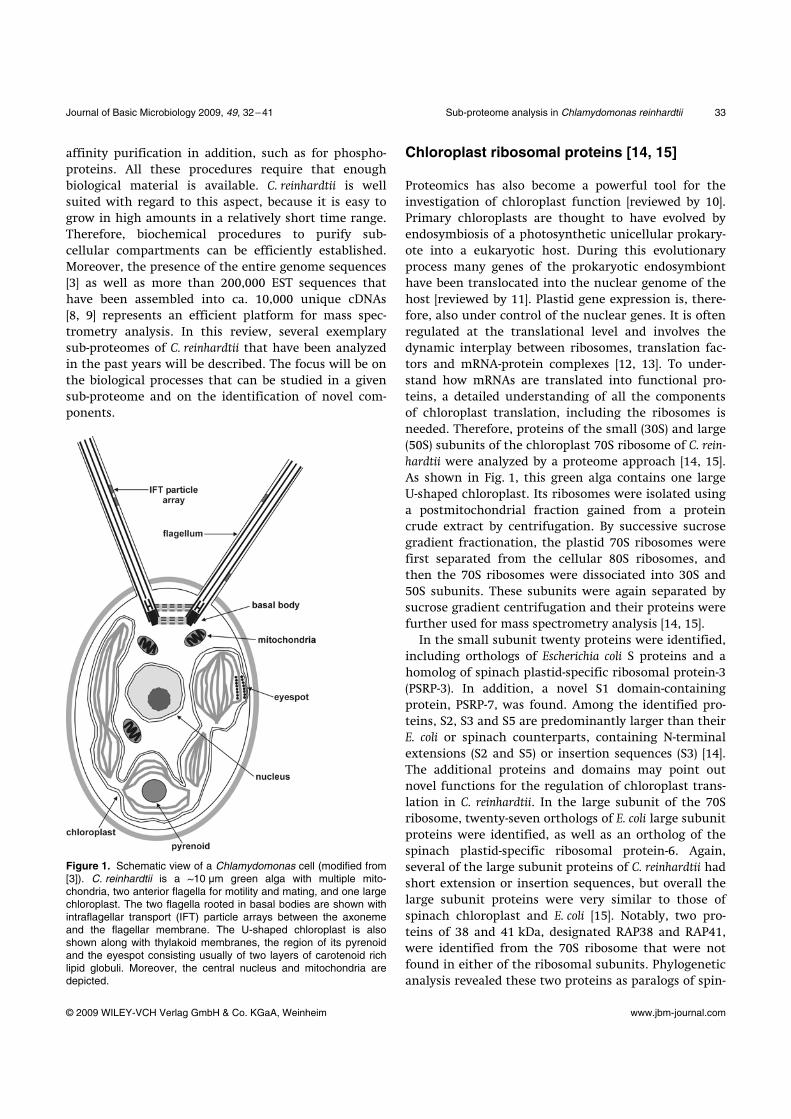
schematic view of a C. reinhardtii cell is presented in
Fig. 1. C. reinhardtii offers experimental advantages that
come close to yeast while yeast cannot be used for
many of the processes in focus. It is a haplont, but plus
Correspondence: Maria Mittag, Institute of General Botany and Plant Physiology, Friedrich-Schiller-University Jena, 07743 Jena, Germany E-mail: [email protected] Phone: 0049-(0)3641-949-201
and minus gametes can be induced by starvation in the
light. Genetic crosses and tetrad analysis are possible.
The alga can be grown photo-, mixo- and heterotrophi-
cally (on acetate) allowing analysis of mutants that
would be lethal, if obligatory photoautotroph. Numer-
ous molecular biology methods have been developed
over the past years for this alga and several reporter
genes (e.g. green fluorescent protein, luciferase) as well
as strong promoters are available [4–6].
During the last few years, proteomics has been estab-
lished as a powerful tool for understanding various
biological problems in several organisms [7]. Although
proteins can be identified down to the femtomol and
even the attomol range with modern mass spectrome-
try, many regulatory proteins within a crude extract
are not abundant enough to be unambiguously identi-
fied by mass spectrometry. For this reason, enrichment
of such proteins is often a prerequisite for efficient
proteome analysis. This can be achieved, for example,
by isolating cellular sub-compartments or conducting
Journal of Basic Microbiology 2009, 49, 32–41 Sub-proteome analysis in Chlamydomonas reinhardtii 33
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
affinity purification in addition, such as for phospho-
proteins. All these procedures require that enough
biological material is available. C. reinhardtii is well
suited with regard to this aspect, because it is easy to
grow in high amounts in a relatively short time range.
Therefore, biochemical procedures to purify sub-
cellular compartments can be efficiently established.
Moreover, the presence of the entire genome sequences
[3] as well as more than 200,000 EST sequences that
have been assembled into ca. 10,000 unique cDNAs
[8, 9] represents an efficient platform for mass spec-
trometry analysis. In this review, several exemplary
sub-proteomes of C. reinhardtii that have been analyzed
in the past years will be described. The focus will be on
the biological processes that can be studied in a given
sub-proteome and on the identification of novel com-
ponents.
Figure 1. Schematic view of a Chlamydomonas cell (modified from [3]). C. reinhardtii is a ~10 µm green alga with multiple mito-chondria, two anterior flagella for motility and mating, and one large chloroplast. The two flagella rooted in basal bodies are shown with intraflagellar transport (IFT) particle arrays between the axoneme and the flagellar membrane. The U-shaped chloroplast is also shown along with thylakoid membranes, the region of its pyrenoid and the eyespot consisting usually of two layers of carotenoid rich lipid globuli. Moreover, the central nucleus and mitochondria are depicted.
Chloroplast ribosomal proteins [14, 15]
Proteomics has also become a powerful tool for the
investigation of chloroplast function [reviewed by 10].
Primary chloroplasts are thought to have evolved by
endosymbiosis of a photosynthetic unicellular prokary-
ote into a eukaryotic host. During this evolutionary
process many genes of the prokaryotic endosymbiont
have been translocated into the nuclear genome of the
host [reviewed by 11]. Plastid gene expression is, there-
fore, also under control of the nuclear genes. It is often
regulated at the translational level and involves the
dynamic interplay between ribosomes, translation fac-
tors and mRNA-protein complexes [12, 13]. To under-
stand how mRNAs are translated into functional pro-
teins, a detailed understanding of all the components
of chloroplast translation, including the ribosomes is
needed. Therefore, proteins of the small (30S) and large
(50S) subunits of the chloroplast 70S ribosome of C. rein-
hardtii were analyzed by a proteome approach [14, 15].
As shown in Fig. 1, this green alga contains one large
U-shaped chloroplast. Its ribosomes were isolated using
a postmitochondrial fraction gained from a protein
crude extract by centrifugation. By successive sucrose
gradient fractionation, the plastid 70S ribosomes were
first separated from the cellular 80S ribosomes, and
then the 70S ribosomes were dissociated into 30S and
50S subunits. These subunits were again separated by
sucrose gradient centrifugation and their proteins were
further used for mass spectrometry analysis [14, 15].
In the small subunit twenty proteins were identified,
including orthologs of Escherichia coli S proteins and a
homolog of spinach plastid-specific ribosomal protein-3
(PSRP-3). In addition, a novel S1 domain-containing
protein, PSRP-7, was found. Among the identified pro-
teins, S2, S3 and S5 are predominantly larger than their
E. coli or spinach counterparts, containing N-terminal
extensions (S2 and S5) or insertion sequences (S3) [14].
The additional proteins and domains may point out
novel functions for the regulation of chloroplast trans-
lation in C. reinhardtii. In the large subunit of the 70S
ribosome, twenty-seven orthologs of E. coli large subunit
proteins were identified, as well as an ortholog of the
spinach plastid-specific ribosomal protein-6. Again,
several of the large subunit proteins of C. reinhardtii had
short extension or insertion sequences, but overall the
large subunit proteins were very similar to those of
spinach chloroplast and E. coli [15]. Notably, two pro-
teins of 38 and 41 kDa, designated RAP38 and RAP41,
were identified from the 70S ribosome that were not
found in either of the ribosomal subunits. Phylogenetic
analysis revealed these two proteins as paralogs of spin-

34 V. Wagner et al. Journal of Basic Microbiology 2009, 49, 32–41
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
ach CSP41, a chloroplast RNA-binding protein with
endoribonuclease activity [15].
Thus, proteomic analyses of the chloroplast ri-
bosomes identified a significant number of chloroplast-
unique additional domains of ribosomal proteins that
expand upon a basic bacterial 70S-like composition. In
a recent study [74], cryo-electron microscopy and single-
particle reconstruction revealed that these novel struc-
tural additions are located at optimal positions on the
chloroplast ribosome for interaction with mRNAs dur-
ing translation initiation. Visualization of these chloro-
plast-unique structures on the ribosome, combined
with mRNA cross-linking, allowed to propose a model
for translation initiation in chloroplasts in which
chloroplast-unique ribosomal proteins interact with
plastid-specific translation factors and RNA elements to
facilitate regulated translation of chloroplast mRNAs
[74].
The composition of the flagellum [16]
Cilia and flagella, which are essentially identical, are
widespread cell organelles that have been highly con-
served throughout evolution and play important roles
in motility, sensory perception and the life cycles of
eukaryotes ranging from protists to humans. Defects in
cilia in humans can cause severe diseases such as poly-
cystic kidney disease (PKD), retinal degeneration, hy-
drocephalus, or changes in the left-right symmetry of
organs, collectively known as ciliopathies [17]. Al-
though C. reinhardtii and mammals are separated by
more than 109 years of evolution, C. reinhardtii flagella
are amazingly similar in structure and function to
mammalian motile cilia [18]. They are composed of
nine microtubular doublets surrounding two central
microtubular singlets. The axoneme of motile flagella
includes substructures such as dynein arms and radial
spokes that generate and control axoneme bending [19].
The flagellum also contains matrix proteins that are
not tightly associated with the flagellar membrane or
the axoneme. They serve diverse functions and can be
involved in intraflagellar transport (IFT) [20].
Proteome analysis of flagellar proteins from C. rein-
hardtii was undertaken with different sub-fractions of
this organelle [16]. Therefore, algal cells were first
deflagellated. The flagella were then sub-fractionated in
different parts, including membrane and axonemes,
membrane and matrix as well as a KCl extract of
axonemes and the remaining axonemes. By this way,
a comprehensive list of flagellar proteins along with
sub-cellular localization within the flagellum was
provided. All identified peptides along with the
corresponding proteins can be accessed via the web
site http://labs.umassmed.edu/chlamyfp/index.php. 360
proteins were identified by five or more peptides, and
additional 292 proteins by two to five peptides [16].
Among the identified proteins, many motor proteins
were present. Moreover, more than 90 putative signal
transduction proteins were found. These include, for
example, 21 protein kinases and 11 protein phosphata-
ses. These findings are consistent with the fact that a
large number of flagellar proteins are phosphorylated
[21], and with the roles of reversible protein phosphory-
lation in the control of flagellar motility [22], signaling
[23], and probably assembly [20]. Another functional
group of this category contained 27 EF hand-containing
proteins that are potential Ca2+-binding proteins, con-
sistent with the role of Ca2+ in the control of flagellar
behavior [24] and signaling [25, 26]. In addition to signal
transduction proteins, also proteins for nucleotide pro-
duction and metabolism were found, including en-
zymes of the late glycolytic pathway. Of note are also
homologues of vertebrate disease proteins that are
involved in PKD (e.g. homologues of polycystin2, fibro-
cystin, scorpion, quilin and three NIMA kinases), hy-
drocephalus (homologues of hydin and napa) or pro-
teins associated with male sterility in mouse (e.g.
homologues of “Parkin-coregulated gene” protein or of
a protein encoding a phosphatase1 catalytic subunit)
[16].
Proteins of the centriole [31]
Centrioles are composed of nine triplet microtubules
arranged in a cylinder [27]. During interphase, centri-
oles act as basal bodies by templating the formation of
cilia and flagella. Some human diseases such as the
Bardet-Biedl syndrome are known to result from de-
fects in proteins that are localized in the basal body
[28–30]. Biochemical purification of centrioles is subtle.
For this purpose, algal cells were first deflagellated and
then lysed in detergent. Basal bodies were enriched by
sucrose step gradients, followed by equilibrium cen-
trifugation in a continuous gradient of Nycodenz [31].
Proteomic analysis of this organelle revealed some pro-
teins already known to be localized in the basal body,
such as centrin, epsilon tubulin, and the cartwheel
protein BLD10p [31]. In addition, 27 so-called BUG
(basal-body proteins with up-regulated genes) proteins
were identified. Genes that are up-regulated during
flagellar assembly encode these. They are likely to be
directly involved in ciliogenesis. Moreover, 18 strong

Journal of Basic Microbiology 2009, 49, 32–41 Sub-proteome analysis in Chlamydomonas reinhardtii 35
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
candidate centriole proteins were found, designated
POC (proteome of centriole), which are not encoded by
genes up-regulated during flagellar regeneration [31]
and conserved in species with centrioles and/or in the
human centrosome. They are presumably core struc-
tural components of the centriole.
Analysis of the flagellar and basal body sub-
proteomes have provided an efficient basis to further
understand the biological structure and processes
within these cell organelles and notably, the knowledge
can be used to further gain insight into human
ciliopathies.
The eyespot and its components [33]
The eyespot apparatus of flagellate green algae is a
primordial visual system, which can detect both, light
direction and intensity and is thus important for the
phototactic orientation of these algae [reviewed in 32].
In C. reinhardtii, it is a singular structure usually com-
posed of two layers of highly ordered carotenoid-rich
lipid globuli that are situated at the periphery of the
chloroplast. Thylakoid membranes subtend these glob-
ule layers. Moreover, the outermost globule layer is
attached to specialized areas of the chloroplast enve-
lope membranes and the adjacent plasma membrane,
in which the photoreceptors are considered to be local-
ized. For purification of this complex ultrastructure,
flotation on successive sucrose gradients was applied
[33]. Thereby, a fraction enriched in the eyespot lipid
globuli and the associated parts of chloroplast and
plasma membranes was gained and used for its pro-
teomic analysis. 202 proteins of the eyespot apparatus
that were covered by at least two peptides per protein
were identified. These proteins included the already
known six proteins of the eyespot, EYE2 and MIN1, two
proteins that are relevant for eyespot assembly [re-
viewed in 34], two splicing variants of the retinal bind-
ing protein COP (Chlamydomonas opsin), and two unique
seven-transmembrane domain (TMD) photoreceptors,
COP3 and 4, which are better known as channelrho-
dopsins ChR-1 and ChR-2 [reviewed in 35]. The other
proteins include different functional categories. Some
are involved in carotenoid and retinal biosynthesis. One
group involving eight candidates contains a PAP fi-
brillin domain. Fibrillins were known as proteins of
carotenoid-rich fibrils and plastoglubules in chromo-
plasts and chloroplasts and play an important role in
carotenoid sequestration [36–38]. Yet another category
contains members of potential signal transduction
cascades such as Ca2+-binding proteins, kinases and
phosphatases and even potential chemotaxis-related
proteins [33]. The eyespot apparatus acquires light in-
formation via photoreceptors and forwards it through
signaling pathways to the flagella. In these signaling
cascades, Ca2+ is known to be involved [24, 39, 40]. In
the eyespot proteome, a significant number of thyla-
koid and chloroplast envelope related proteins were
also found. This can be expected since the carotenoid
lipid globule layers are subtented by thylakoid mem-
branes and overlaid by the chloroplast envelope mem-
branes. But one has to be careful to conclude that
known proteins of the chloroplast thylakoid have the
same localization and function in the eyespot. The al-
pha, beta and gamma subunits of the chloroplast ATP
synthase seem to have a specialized localization and
possibly function within the eyespot [41]. C. reinhardtii
proteome data from the eyespot have been deposited
on a protein network site (http://www2.uni-jena.de/
biologie/chlamy/index.php?page=search&cmd=search)
of C. reinhardtii.
Proteomic analysis of mitochondria from C. reinhardtii [43]
Like chloroplasts, mitochondria are of prokaryotic ori-
gin and have evolved by endosymbiosis. In C. reinhardtii,
mitochondria occupy about 1–3% of the cell volume of
photosynthetically grown algae, while the chloroplast
occupies about 40% [42]. A proteomic approach of mi-
tochondrial proteins from C. reinhardtii resulted in the
identification of proteins that are common among
strict anaerobic and microaerophilic prokaryotes, but
that are very rare in eukaryotes [43]. Thus, pyruvate
formate-lyase (PFL) that catalyzes the non-oxidative
conversion of pyruvate to formate and acetyl-CoA along
with its activating enzyme PFL-AE were found, showing
the existence of a PFL system in this photosynthetic
alga. Moreover, peptides from phosphotransacetylase
and acetate kinase were identified. The phosphotrans-
acetylase-acetate kinase pathway is a common route of
ATP synthesis or acetate assimilation among prokaryo-
tes, but not in eukaryotes. It is also remarkable that an
oxygen producing eukaryotic alga has a broad spectrum
of pyruvate-, ethanol- and acetate-metabolizing en-
zymes that were previously considered as specific to
anaerobic eukaryotic lineages [43]. However, this may
be necessary for the alga to adjust its metabolism to the
different environmental conditions to which it can be
exposed, including anaerobiosis, especially in darkness.
The natural habitats of C. reinhardtii are fresh water
ponds and soil. Notably, the first strain was isolated in

36 V. Wagner et al. Journal of Basic Microbiology 2009, 49, 32–41
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
a potato field in Amherst, Massachusetts (USA) in 1945
and designated by G.M. Smith as isolate 137c [42].
Phosphoproteomes in C. reinhardtii
a. General considerations Many processes are regulated at the posttranslational
level. One of the key modifications of cellular signaling
processes represents phosphorylation. It can affect
proteins functions, activities, localization and interac-
tions. About one-third of all proteins are thought to be
phosphorylated [44]. The analysis of phosphorylated
proteins and the characterization of in vivo phosphory-
lation sites along with their relevant kinases under
different biological conditions are still considered to be
one of the most challenging tasks in current proteomic
research [45, 46]. This has several reasons. First, the
stoichiometry of phosphorylation is generally relatively
low; only a small fraction of the available intracellular
pool of a protein is phosphorylated at any given time as
a result of a stimulus. Second, the phosphorylated sites
on proteins might vary implying that any given phos-
phoprotein is heterogeneous (i.e. exists in several dif-
ferent phosphorylated forms). Third, many of the sig-
naling molecules are present at low abundance within
cells [47]. Connecting specific kinases with relevant
targets remains a further challenge [46].
Specific enrichment of phosphorylated proteins or
peptides is a prerequisite for efficient phosphoprotein
analysis. Immobilized metal-affinity chromatography
(IMAC) is a frequently applied technique for phos-
phopeptide and -protein enrichments [45]. Thereby,
phosphorylated peptides and proteins are bound to the
stationary phase with positively charged metal-ions
that are on their part bound to the column material.
Various metal-ions such as Fe3+, Ga3+, Al3+ or Zr3+ have
been used for selectivity and phosphopeptide recovery.
Especially Ga3+ ions have proven well in different stud-
ies [48–49]. Moreover, metal oxide affinity chromatog-
raphy (MOAC) using, for example, titanium dioxide has
been used to enrich phosphopeptides. Like the above-
mentioned metal ions, metal oxides preferentially in-
teract with phosphopeptides in comparison to unmodi-
fied peptides. In C. reinhardtii, both IMAC and MOAC
have been established [49–53].
b. Environmentally modulated phosphoproteomes of photosynthetic membranes [52, 53, 75, 76] The thylakoid membranes in chloroplasts of plants
and green algae represent the location for photosynthe-
sis. Several proteins of the thylakoid membrane that
make up photosystem II complex (PSII) and its light-
harvesting chlorophyll a/b binding proteins (LHCII)
antennae undergo light- and redox-dependent phos-
phorylation [54, 55]. Phosphorylation of LHCII controls
a process designated photosynthetic State transitions.
In State 1 more energy is transferred to PSII, whereas in
State 2 a proportion of the excitation energy is redis-
tributed to PSI [56–58]. To map in vivo phosphorylation
sites in photosynthetic membranes of C. reinhardtii un-
der different environmental conditions, the photosyn-
thetic membranes were isolated from the algal cells
and exposed to different conditions that affect photo-
synthesis [52]. These included (i) dark aerobic, corre-
sponding to photosynthetic State 1; (ii) dark under ni-
trogen atmosphere, corresponding to photosynthetic
State 2; (iii) moderate light and (iv) high light. The sur-
face-exposed phosphorylated peptides were cleaved
from the membrane by trypsin and finally enriched by
IMAC and analyzed by mass spectrometry. By this way,
19 in vivo phosphorylation sites belonging to 15 proteins
in C. reinhardtii were found [52]. The State 1- to State
2-transition induced phosphorylation of the PSII core
components D2 and PsbR and quadruple phosphoryla-
tion of a minor LHCII antennae subunit, CP29, as well
as phosphorylation of constituents of a major LHCII
complex, Lhcbm1 and Lhcbm10. Exposure of the cells
to either moderate or high light caused additional
phosphorylation of the D1 and CP43 proteins of the
PSII core. The high light treatment led to specific phos-
phorylation of CP29 at seven distinct residues, phos-
phorylation of another minor LHCII constituent, CP26
at a single threonine, and double phosphorylation of
additional subunits of a major LHC complex.
Mass spectrometry analysis of C. reinhardtii in the
State 1 and State 2 also revealed that the LHCII-like
CP29 protein was phosphorylated at Thr6, Thr16,
Thr32, and Ser102 in State 2, whereas in State
1-exposed cells only phosphorylation of Thr6 and Thr32
was detected. The LHCI-PSI supercomplex isolated from
the alga in State 2 was found to contain strongly associ-
ated CP29 in phosphorylated form. Electron microscopy
suggested the binding site for this highly phosphory-
lated CP29 in PSI and it was postulated that redox-
dependent multiple phosphorylation of CP29 in green
algae is an integral part of the State transition process
in which the structural changes of CP29, induced by
reversible phosphorylation, determine the affinity of
LHCII for either of the two photosystems [75].
Besides the phosphorylation sites detection, mass
spectrometry analyses of the surface-exposed peptides
cleaved by trypsin from the thylakoid membranes of
the green alga uncovered the N-termini of nuclear-

Journal of Basic Microbiology 2009, 49, 32–41 Sub-proteome analysis in Chlamydomonas reinhardtii 37
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
encoded chlorophyll a/b-binding proteins and their
N-terminal acetylation. The N-termini of mature chlo-
rophyll a/b-binding proteins differed from those earlier
predicted by bioinformatics [52, 76].
In another approach, C. reinhardtii was acclimated to
limiting environmental CO2 conditions. Under these
conditions, a CO2-concentrating mechanism (CCM) is
induced that is associated with transcriptional regula-
tion of many genes [59]. Thereby, specific protein phos-
phorylation at the surface of thylakoid membranes was
investigated and found to be redox-dependent [53]. One
phosphopeptide belonged to an unknown expressed
protein (UEP). Moreover, three phosphorylated peptides
of the Lci5 protein that is encoded by the low-CO2-
inducible gene 5 were identified.
c. The phosphoproteome of the eyespot [51] As mentioned before, proteomic analysis of the eyespot
of C. reinhardtii revealed the presence of a limited num-
ber of kinases and phosphatases [33], suggesting that
reversible protein phosphorylation might play a role in
the light-signaling cascade. To identify the targets of
the kinases and phosphatases, a functional phospho-
proteome approach was applied [51]. Since the proteins
of the eyespot have a rather hydrophobic character,
a specialized protocol involving digestion with the
endopeptidase LysC prior to trypsin was used for gener-
ating the phosphopeptides from the eyespot that were
enriched by IMAC. Proteins from the eyespot were dis-
solved in 4 M urea, in which LysC still has an activity of
86%. In total, 68 different phosphopeptides correspond-
ing to 32 known proteins of the eyespot along with 52
precise in vivo phosphorylation sites were identified by
mass spectrometry. Comparisons to other known phos-
phoproteins from C. reinhardtii obtained by phosphopro-
teomic approaches with whole cell extracts [49] or thy-
lakoid membranes [52, 53] revealed that only a few of
these phosphoproteins (6) were overlapping with the
eyespot phosphoproteome. Thus, a significant number
of phosphoproteins could only be identified when the
sub-cellular eyespot fraction was enriched prior to en-
richment of its phosphopeptides by IMAC. The identi-
fied phosphoproteins belong mainly to four functional
categories: carotenoid and fatty acid metabolism, (puta-
tive) light signaling pathway(s) and retina-related pro-
teins, thylakoid and chloroplast envelope-related pro-
teins and conserved and novel proteins of yet unknown
functions. Some phosphoproteins are clearly involved
in the light signaling pathway(s). These include the two
photoreceptors ChR-1 and ChR-2, which contain three
and one phosphorylation sites, respectively. These sites
are localized in a cytoplasmatic loop with close prox-
imity to the seven transmembrane domain channel-
forming regions. It is striking that the relative position
of the functional sites of phosphorylation is highly
conserved within the green algal and vertebrate rho-
dopsins, implying functional relevance for the regula-
tion of these unique directly light-gated channels [51].
In this context, it should be also pointed out that ChR-2
has been successfully expressed for light stimulation of
different systems including Caenorhabditis elegans and
mammalian neurons [60, 61]. It was even delivered to
retinal ganglion cells in a rodent model of inherited
blindness [62].
Two other examples of phosphoproteins that repre-
sent potential members of the light-signaling path-
way(s) within the eyespot include an EF hand contain-
ing Ca2+-binding protein and the SOUL3 heme-binding
protein [51]. It is known that extra-cellular calcium-
fluxes are intricately involved in the behavioral re-
sponses of C. reinhardtii to light [reviewed in 23] and
that both ChRs can conduct Ca2+ [63]. A SOUL heme-
binding protein was found in a screen for chicken
mRNAs specifically expressed in the retina and pineal
gland [64], indicating that certain proteins seem to be
indeed conserved with regard to their localization from
primitive visual algal systems to the highly sophisti-
cated visual system of animals.
Redox signaling in C. reinhardtii and the influence of Cadmium [65–67]
Pollution of the environment with heavy metals in soil
or water is often a result of industrial activity. Exposure
to Cadmium (Cd) can result in serious health problems.
Using proteomics along with comparative two-dimen-
sional gel electrophoresis, the effect of Cd on the so-
luble proteome of C. reinhardtii grown on acetate was
examined [65]. 20 proteins were found in a larger
amount compared to non-treated cells and 26 proteins
were decreased in the amount after Cd treatment. The
most drastic effect was the decrease in abundance of
both large and small subunits of the CO2 fixation en-
zyme, ribulose-1,5-bisphosphate carboxylase/oxygenase,
in correlation with several other enzymes involved in
photosynthesis, Calvin cycle and chlorophyll biosynthe-
sis. Other down regulated processes included fatty acid
biosynthesis as well as amino acid and protein biosyn-
thesis. On the other hand, proteins involved in glu-
tathione synthesis, ATP metabolism, response to oxida-
tive stress and protein folding were up-regulated in the
presence of Cd [65]. It was also found that many of the
Cd-sensitive proteins were regulated via two major

38 V. Wagner et al. Journal of Basic Microbiology 2009, 49, 32–41
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
cellular thiol redox systems, thioredoxin and glutare-
doxin. Notably, targets of thioredoxins in C. reinhardtii
are known from a proteome approach [66]. Thereby, a
thioredoxin affinity approach was taken and 55 targets
were identified by mass spectrometry. Some of these
targets were known from higher plants and cyanobacte-
ria, but 26 were novel, providing an efficient platform
for further studying their functions and importance
within redox signaling. Recently, 25 in vivo targets of
S-thiolation were identified in C. reinhardtii [67]. Gluta-
thionylation is the major form of S-thiolation in cells.
This reversible redox post-translational modification
consists of the formation of a mixed disulfide between a
free thiol on a protein and a molecule of the tripeptide
glutathione. Several of the identified targets were relat-
ed to photosynthesis, such as the Calvin cycle enzymes
phosphoglycerate kinase and ribose-5-phosphate isom-
erase. A number of targets were also related to stress
responses, such as chaperones and peroxiredoxins [67].
Further proteome approaches in C. reinhardtii
There have been also other functional proteomic ap-
proaches in C. reinhardtii [68, 69]. A few further strate-
gies are shortly described below. For example, func-
tional proteomics was applied to find proteins that are
regulated by the circadian clock. Two circadian ex-
pressed proteins, a protein disulfide isomerase and a
tetratricopeptide repeat protein, which were enriched
by heparin affinity chromatography, were identified by
this way [70]. Shotgun proteomic profiling technologies
were applied to C. reinhardtii that detected 1069 proteins
of this green alga. These data were used to validate the
in silico gene models [71]. Co-immunoprecipitations with
antibodies directed against a specific protein were car-
ried out to precipitate interaction partners that can
then be analyzed by mass spectrometry. Such an effort
was undertaken, for example, to characterize the “in-
teractome” of the vesicle-inducing protein in plastids,
designated VIPP1 in C. reinhardtii [72].
Comparative quantitative proteomics is of high in-
terest to study variations in the protein pool of biologi-
cal processes that are induced under certain conditions.
Thereby, stable isotope labeling by amino acids in cell
culture can be used. Such an approach was undertaken
to investigate the remodeling of bioenergetic pathways
under iron deficiency in C. reinhardtii [73]. Proteins that
show a decreased or increased expression under iron
deficiency were identified. The data suggested that
iron-deprivation induces a transition from photohet-
erotrophic to a primarily heterotrophic metabolism.
Conclusions
This review has focused mainly on large-scale proteome
approaches that have been undertaken in the past years
with C. reinhardtii. The proteomes described in this re-
view underline the power of functional proteome ap-
proaches, where cellular compartments are purified
and the proteins are identified by mass spectrometry.
By this way, hundreds of proteins have been character-
ized that belong to specific sub-cellular compartments
of C. reinhardtii and/or that are modified at the post-
translational level or modulated by the environment.
These data have provided new insights into a variety of
biological processes in C. reinhardtii. They create an
efficient basis for the further functional characteriza-
tion of involved proteins and provide a platform for
knowledge transfer to other organisms including not
only plant, but also prokaryotic and animal systems.
Acknowledgements
We thank Carsten Milkowski for helpful comments on
the manuscript. Proteome analysis of C. reinhardtii
would not have reached such a level without the free
delivery of EST and genome sequences from the ge-
nome projects of C. reinhardtii in the USA (Department
of Energy, Joint Genome Institute) and Japan (Kazusa
DNA Research Institute). Our work was supported by
grants of the Deutsche Forschungsgemeinschaft.
References
[1] Grossman, A.R., 2005. Paths toward algal genomics. Plant Physiol., 137, 410–427.
[2] Grossman, A.R., 2007. In the grip of algal genomics. Adv. Exp. Med. Biol., 616, 54–76.
[3] Merchant, S.S., Prochnik, S.E., Vallon, O., Harris, E.H., Karpowicz, S.J., Witman, G.B. et al., 2007. The evolution of key animal and plant functions is revealed by analysis of the Chlamydomonas genome. Science, 318, 245–250.
[4] Harris, E.H., 2001. Chlamydomonas as model organism. Ann. Rev. Plant Physiol. Plant Mol. Biol., 52, 363–406.
[5] Mittag, M. and Wagner, V., 2003. The circadian clock of the unicellular eukaryotic model organism Chlamydomo-nas reinhardtii. Biol. Chem., 384, 689–695.
[6] Heitzer, M., Eckert, A., Fuhrmann, M. and Griesbeck, C., 2007. Influence of codon bias on the expression of foreign genes in microalgae. Adv. Exp. Med. Biol., 616, 46–53.
[7] Reinders, J., Lewandrowski, U., Moebius, J., Wagner, Y. and Sickmann, A., 2004. Challenges in mass spectrome-try-based proteomics. Proteomics, 4, 3686–3703.

Journal of Basic Microbiology 2009, 49, 32–41 Sub-proteome analysis in Chlamydomonas reinhardtii 39
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
[8] Asamizu, E., Nakamura, Y., Sato, S., Fukuzawa, H. and Tabata, S., 1999. A large scale structural analysis of cDNAs in a unicellular green alga, Chlamydomonas reinhardtii. I. Generation of 3433 non-redundant expressed sequence tags. DNA Res., 6, 369–373.
[9] Grossman, A.R., Harris, E.E., Hauser, C., Lefebvre, P.A., Martinez, D., Rokhsar, D., Shrager, J., Silflow, C.D., Stern, D., Vallon, O. and Zhang, Z., 2003. Chlamydomonas rein-hardtii at the crossroads of genomics. Eukaryot. Cell, 2, 1137–1150.
[10] van Wijk, K.J., 2001. Challenges and prospects of plant proteomics. Plant Physiol., 126, 501–508.
[11] Subramanian, A.R., 1993. Molecular genetics of chloro-plast ribosomal proteins. Trends Biochem. Sci., 18, 177–181.
[12] Mayfield, S.P., Yohn, C.B., Choen, A. and Danon, A., 1995. Regulation of chloroplast gene expression. Annu. Rev. Plant Physiol. Plant Mol. Biol., 46, 147–166.
[13] Barkan, A. and Goldschmidt-Clermont, M., 2000. Partici-pation of nuclear genes in chloroplast gene expression. Biochimie, 82, 559–572.
[14] Yamaguchi, K., Prieto, S., Beligni, M.V., Haynes, P.A., McDonald, W.H., Yates III, J.R. and Mayfield, S.P., 2002. Proteomic characterization of the small subunit of Chla-mydomonas reinhardtii chloroplast ribosome: Identification of a novel S1 domain-containing protein and unusually large orthologs of bacterial S2, S3, and S5. Plant Cell, 14, 2957–2974.
[15] Yamaguchi, K., Beligni, M.V., Prieto, S., Haynes, P.A., McDonald, W.H., Yates III, J.R. and Mayfield, S.P., 2003. Proteomic characterization of the Chlamydomonas reinhard-tii chloroplast ribosome. J. Biol. Chem., 278, 33774–33785.
[16] Pazour, G.J., Agrin, N., Leszyk, J. and Witman, G.B., 2005. Proteomic analysis of a eukaryotic cilium. J. Cell. Biol., 170, 103–113.
[17] Marshall, W.F., 2008. The cell biological basis of ciliary disease. J. Cell Biol., 180, 17–21.
[18] Silflow, C.D. and Lefebvre, P.A., 2001. Assembly and moti-lity of eukaryotic cilia and flagella. Lessons from Chlamy-domonas reinhardtii. Plant Physiol., 127, 1500–1507.
[19] Pazour, G.J. and Witman, G.B., 2003. The vertebrate pri-mary cilium is a sensory organelle. Curr. Opin. Cell Biol., 15, 105–110.
[20] Rosenbaum, J.L. and Witman, G.B., 2002. Intraflagellar transport. Nat. Rev. Mol. Cell Biol., 3, 813–825.
[21] Piperno, G., Huang, B., Ramanis, Z. and Luck, D., 1981. Radial spokes of Chlamydomonas flagella: polypeptide com-positon and phosphorylation of stalk components. J. Cell Biol., 88, 73–79.
[22] Porter, M.E. and Sale, W.S., 2000. The 9 + 2 axoneme anchors multiple inner arm dyneins and a network of ki-nases and phosphatases that control motility. J. Cell Biol., 151, F37–42.
[23] Pan, J. and Snell, W.J., 2000. Regulated targeting of a protein kinase into an intact flagellum. An aurora/Ip11p-like protein kinase translocates from the cell body into the flagella during gamete activation in Chlamydomonas. J. Biol. Chem., 275, 24106–24114.
[24] Witman, G.B., 1993. Chlamydomonas phototaxis. Trends Cell Biol., 3, 403–408.
[25] Bloodgood, R.A., 1992. Calcium-regulated phosphoryla- tion of proteins in the membrane-matrix compartment of the Chlamydomonas flagellum. Exp. Cell Res., 198, 228–236.
[26] Praetorius, H.A. and Spring, K.R., 2001. Bending the MDCK cell primary cilium increases intracellular calcium. J. Membr. Biol., 184, 71–79.
[27] Beisson, J. and Wright, M., 2003. Basal body/centriole assembly and continuity. Curr. Opin. Cell Biol., 15, 96–104.
[28] Li, J.B., Gerdes, J.M., Haycraft, C.J., Fan, Y., Teslovich, T.M., May-Simera, H., Li, H., Blacque, O.E., Li, L., Leitch, C.C. et al., 2004. Comparative genomics identifies a flagel-lar and basal body proteome that includes the BBS5 hu-man disease gene. Cell, 117, 541–552.
[29] Kim, J.C., Badano, J.L., Sibold, S., Esmail, M.A., Hill, J., Hoskins, B.E., Leitch, C.C., Venner, K., Ansley, S.J., Ross, A.J. et al., 2004. The Bardet-Biedl protein BBS4 targets car-go to the pericentriolar region and is required for micro-tubule anchoring and cell cycle progression. Nat. Genet., 36, 462–470.
[30] Kim, J.C., Ou, Y.Y., Badano, J.L., Esmail, M.A., Leitch, C.C., Fiedrich, E., Beales, P.L., Archibald, J.M., Katsanis, N., Rattner, J.B. et al., 2005. MKKS/BBS6, a divergent chapero-nin-like protein linked to the obesity disorder Bardet-Biedl syndrome, is a novel centrosomal component requi-red for cytokinesis. J. Cell Sci., 118, 1007–1020.
[31] Keller, L.C., Romijn, E.P., Zamora, I., Yates III, J.R. and Marshall, W.F., 2005. Proteomic analysis of isolated Chla-mydomonas centrioles reveals orthologs of ciliary-disease genes. Curr. Biol., 15, 1090–1098.
[32] Kreimer, G., 2001. Light reception and signal modulation during photoorientation of flagellate algae. In: Compre-hensive Series in Photosciences (Lebert, M. and Häder, D.-P., eds), pp. 193–227. Elsevier Amsterdam, The Nether-lands.
[33] Schmidt, M., Geßner, G., Luff, M., Heiland, I., Wagner, V., Kaminski, M., Geimer, S., Eitzinger, N., Reißenweber, T., Voytsekh, O., Fiedler, M., Mittag, M. and Kreimer, G., 2006. Proteomic analysis of the eyespot of Chlamydomonas reinhardtii provides novel insights into its components and tactic movements. Plant Cell, 18, 1908–1930.
[34] Dieckmann, C.L., 2003. Eyespot placement and assembly in the green alga Chlamydomonas. Bioessays, 25, 410–416.
[35] Kateriya, S., Nagel, G., Bamberg, E. and Hegemann, P., 2004. “Vision” in single-celled algae. News Physiol. Sci., 19, 133–137.
[36] Deruère, J., Römer, S., D’Harlingue, A., Backhaus, R.A., Kuntz, M. and Camara, B., 1994. Fibril assembly and caro-tenoid overaccumulation in chromoplasts: A model for supramolecular lipoprotein structures. Plant Cell, 6, 119–133.
[37] Pozueta-Romero, J., Rafia, F., Houlné, G., Cheniclet, C., Carde, J.P., Schantz, M.-L. and Schantz, R., 1997. A ubiqui-tous plant housekeeping gene, PAP, encodes a major protein component of bell pepper chromoplasts. Plant Physiol., 115, 1185–1194.

40 V. Wagner et al. Journal of Basic Microbiology 2009, 49, 32–41
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
[38] Bréhélin, C., Kessler, F. and van Wijk, K.J., 2007. Plas-toglobules: versatile lipoprotein particles in plastids. Trends Plant Sci., 12, 260–266.
[39] Pazour, G., Sineshchekov, O.A. and Witman, G.B., 1995. Mutational analysis of the phototransduction pathway of Chlamydomonas reinhardtii. J. Cell Biol., 131, 427–440.
[40] Sineshchekov, O.A. and Govorunova, E.G., 1999. Rho-dopsin-mediated photosensing in green flagellated algae. Trends Plant Sci., 4, 58–63.
[41] Schmidt, M., Luff, M., Mollwo, A., Kaminski, M., Mit- tag, M. and Kreimer, G., 2007. Evidence for a specialized localization of the chloroplast ATP-synthase subunits α, β and γ in the eyespot apparatus of Chlamydomonas reinhard-tii (Chlorophyceae). J. Phycol., 43, 284–294.
[42] Harris, E.H., 1989. The Chlamydomonas Sourcebook: A Comprehensive Guide to Biology and Laboratory Use. Academic Press, San Diego.
[43] Atteia, A., van Lis, R., Gelius-Dietrich, G., Adrait, A., Ga-rin, J., Joyard, J., Rolland, N. and Martin, W., 2006. Pyru-vate formate-lyase and a novel route of eukaryotic ATP synthesis in Chlamydomonas mitochondria. J. Biol. Chem., 281, 9909–9918.
[44] Hubbard, M.J. and Cohen, P., 1993. On target with a new mechanism for the regulation of protein phosphoryla-tion. Trends Biochem. Sci., 18, 172–177.
[45] Reinders, J. and Sickmann, A., 2005. State-of-the-art in phosphoproteomics. Proteomics, 5, 4052–4061.
[46] Sopko, R. and Andrews, B.J., 2008. Linking the kinome and phosphorylome – a comprehensive review of ap-proaches to find kinase targets. Mol. Bio. Syst., 4, 920–933.
[47] Mann, M., Ong, S.E., Grønborg, M., Steen, H., Jensen, O.N. and Pandey, A., 2002. Analysis of protein phosphorylation using mass spectrometry: deciphering the phosphopro-teome. Trends Biotechnol., 20, 261–268.
[48] Shu, H., Chen, S., Bi, Q., Mumby, M. and Brekken, D.L., 2004. Identification of Phosphoproteins and their phos-phorylation sites in the WEHI-231 B lymphoma cell line. Mol. Cell Proteomics, 3, 279–286.
[49] Wagner, V., Geßner, G., Heiland, I., Kaminski, M., Hawat, S., Scheffler, K. and Mittag, M., 2006. Analysis of the phos-phoproteome of Chlamydomonas reinhardtii provides new insights into various cellular pathways. Eukaryot Cell, 5, 457–468.
[50] Wolschin, F. and Weckwerth, W., 2005. Combining metal oxide affinity chromatography (MOAC) and selective mass spectrometry for robust identification of in vivo protein phosphorylation sites. Plant Methods, 1, 9.
[51] Wagner, V., Ullmann, K., Mollwo, A., Kaminski, M., Mit-tag, M. and Kreimer, G., 2008. The phosphoproteome of a Chlamydomonas reinhardtii eyespot fraction includes key proteins of the light signaling pathway. Plant Physiol., 146, 772–788.
[52] Turkina, M.V., Kargul, J., Blanco-Rivero, A., Villarejo, A., Barber, J. and Vener, A.V., 2006a. Environmentally modu-lated phosphoproteome of photosynthetic membranes in the green alga Chlamydomonas reinhardtii. Mol. Cell. Pro- teom., 5, 1412–1425.
[53] Turkina, M.V., Blanco-Rivero, A., Vainonen, J.P., Vener, A.V. and Villarejo, A., 2006b. CO2 limitation induces spe-
cific redox-dependent protein phosphorylation in Chlamy-domonas reinhardtii. Proteomics, 6, 2693–2704.
[54] Vener, A.V., Van Kan, P.J.M., Rich, P.R., Ohad, I. and Andersson, B., 1997. Plastoquinol at the quinol oxidation site of reduced cytochrome bf mediates signal transduc-tion between light and protein phosphorylation: thyla-koid protein kinase deactivation by a single-turnover flash. Proc. Natl. Acad. Sci. USA, 94, 1585–1590.
[55] Zito, F., Finazzi, G., Delosme, R., Nitschke, W., Picot, D. and Wollman, F.A., 1999. The Qo site of cytochrome b6f complexes controls the activation of the LHCII kinase. EMBO J., 18, 2961–2969.
[56] Allen, J.F., 2003. Botany. State transitions – a question of balance. Science, 299, 1530–1532.
[57] Depege, N., Bellafiore, S. and Rochaix, J.D., 2003. Role of chloroplast protein kinase Stt7 in LHCII phosphorylation and state transition in Chlamydomonas. Science, 299, 1572–1575.
[58] Bellafiore, S., Barneche, F., Peltier, G. and Rochaix, J.D., 2005. State transitions and light adaptation require chloroplast thylakoid protein kinase STN7. Nature, 433, 892–895.
[59] Yamano, T., Miura, K. and Fukuzawa, H., 2008. Expres-sion analysis of genes associated with the induction of the carbon-concentrating mechanism in Chlamydomonas rein-hardtii. Plant Physiol., 147, 340–354.
[60] Boyden, E.S., Zhang, F., Bamberg, E., Nagel, G. and Deisse-roth, K., 2005. Millisecond-timescale, genetically targeted optical control of neural activity. Nature Neurosci., 8, 1263–1268.
[61] Nagel, G., Brauner, M., Liewald, J.F., Adeishvili, N., Bam-berg, E. and Gottschalk, A., 2005. Light activation of chan-nel rhodopsin-2 in excitable cells of Caenorhabditis elegans triggers rapid behavioral responses. Curr. Biol., 15, 2279–2284.
[62] Bi, A., Cui, J., Ma, Y.P., Olshevskaya, E., Pu, M., Dizhoor, A.M. and Pan, Z.-H., 2006. Ectopic expression of a micro-bial-type rhodopsin restores visual responses in mice with photoreceptor degeneration. Neuron, 50, 23–33.
[63] Hegemann, P. and Tsunoda, S.P., 2007. Light tools for neuroscience: Channelrhodopsin and light-activated en-zymes. Cell Sci. Rev., 3, 108–123.
[64] Zylka, M.J. and Reppert, S.M., 1999. Discovery of a putati-ve hemebinding protein family (SOUL/HBP) by two-tissue suppression subtractive hybridization and database sear-ches. Brain Res. Mol. Brain Res., 74, 175–181.
[65] Gillet, S., Decottignies, P., Chardonnet, S. and Le Maré-chal, P., 2006. Cadmium response and redoxin targets in Chlamydomonas reinhardtii: a proteomic approach. Photo-synth. Res., 89, 201–211.
[66] Lemaire, S.D., Guillon, B., Le Maréchal, P., Keryer, E., Miginiac-Maslow, M. and Decottignies, P., 2004. New thio-redoxin targets in the unicellular photosynthetic eu-karyote Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA, 101, 7475–7480.
[67] Michelet, L., Zaffagnini, M., Vanacker, H., Le Maréchal, P., Marchand, Ch., Schroda, M., Lemaire S.D. and Decottig-nies, P., 2008. In vivo targets of S-thiolation in Chlamydo-monas reinhardtii. J. Biol. Chem., 283, 21571–21578.

Journal of Basic Microbiology 2009, 49, 32–41 Sub-proteome analysis in Chlamydomonas reinhardtii 41
© 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
[68] Stauber, E.J. and Hippler, M., 2004. Chlamydomonas reinhardtii proteomics. Plant Physiol. Biochem., 42, 989–1001.
[69] Wagner, V., Geßner, G. and Mittag, M., 2005. Functional proteomics: a promising approach to find novel compo-nents of the circadian system. Chronobiol. Internat., 22, 403–415.
[70] Wagner, V., Fiedler, M., Markert, Ch., Hippler, M. and Mittag, M., 2004. Functional proteomics of circadian ex-pressed proteins from Chlamydomonas reinhardtii. FEBS Let-ters, 559, 129–135.
[71] May, P., Wienkoop, S., Kempa, S., Usadel, B., Christian, N., Rupprecht, J., Weiss, J., Recuenco-Munoz, I., Ebenhöh, O., Weckwerth, W. and Walther, D., 2008. Metabolomics- and proteomics-assisted genome annotation and analysis of the draft metabolic network of Chlamydomonas reinhard-tii. Genetics, 179, 157–166.
[72] Liu, C., Willmund, F., Golecki, J.R., Cacace, S., Heß, H., Markert, C. and Schroda, M., 2007. The chloroplast
HSP70B-CDJ2-CGE1 chaperones catalyse assembly and dis-assembly of VIPP1 oligomers in Chlamydomonas. Plant J., 50, 265–277.
[73] Naumann, B., Busch, A., Allmer, J., Ostendorf, E., Zeller, M., Kirchhoff, H. and Hippler, M., 2007. Comparative quantitative proteomics to investigate the remodeling of bioenergetic pathways under iron deficiency in Chlamy-domonas reinhardtii. Proteomics, 7, 3964–3979.
[74] Manuell, A.L., Quispe, J. and Mayfield, S.P., 2007. Struc-ture of the chloroplast ribosome: Novel domains for translational regulation. PLOS Biology, 5, 1785–1797.
[75] Kargul, J., Turkina, M.V., Nield, J., Benson, S. Vener A.V. and Barber, J., 2005. Light-harvesting complex II protein CP29 binds to photosystem I of Chlamydomonas reinhardtii under State 2 conditions. FEBS Journal, 272, 4797–4806.
[76] Turkina, M.V., Villarejo, A. and Vener, A.V., 2004. The transit peptide of CP29 thylakoid protein in Chlamydomo-nas reinhardtii is not removed but undergoes acetylation and phosphorylation. FEBS Letters, 564, 104–108.
((Funded by:
• Deutsche Forschungsgemeinschaft))





























