Embed Size (px)
Citation preview

PROTEIN SEO UENCE MOTIFS oi= ........ 1 . : ' " ' . ' - - - , , - ;
I YE~H2 g~l'T IT f l qOTT~AT glNPLE, 7"AK~. 'zlTf E 1 N~.~£CIALAR ~loLe&Y O'F TtIE ~£LL~(AL~ERT£ Pt'.,~L~ ~
1
6to t.O~tG, q N/LLiON A~,~CI.E~
WOI(# L,IF~ IH TH~ "it TLE : TI~E ¢¢t~,E4¢ , ~lFg INY~ACt£E ¢0~ / ~ ,
To 1~£ y~ t/¢'c~,.tte oF u~e :=(¢)~ :;;t) tV 4 ~ ; l (
LL IH^V~ ~t ,Sc~i~o~lN,~g~ m ~ L~1YLE E,eo~, ee Ar~(~ Th~ i - " ~
~fglN~ T~ T~b( f f ]'Pig b i Vt'N[~ ,,,,,
10
Pete Jeffs is a freelancer working in Paris, France.
TIBS 2 0 - JANUARY 1 9 9 5
20 BreltenbacbSchmitt, I., Schmitt, H. O., Heinlsch, J. and Zimmermann, F. K. (1984) MoL Gen. Genet. 195, 536-540
21 Hohmann, S. et al. (1992) J. Bacteriol. 174, 4183-4188
22 Van Aelst, L. et al. (1993) Mol. Microbiol. 8, 927-943
23 Thevelein, J. M. (1992) Ant. van Leeuwenhoek I Microbiol. 62,109-130
24 Gonzalez, M. I. et al. (1992) Yeast 8, 183-192
25 Cannon, J. F., Pringle, J. R., Fiechter, A. and Kha]il, M. (1994) Genetics 136, 485-503
26 Bell, W. et al. (1992) Eur. J. Biochem. 209, 951-959
27 Vuorio, O. E., Kalkkinen, N. and Londesborough, J. (1993) Eur. Jo Biochem. 216, 849-861
28 McDougall, J., Kaasen, i. and Strcm, A. R. (1993) FEMS MicrobioL Lett. 107, 25-30
29 Vandercammen, A., Fran(~ois, J. and Hers, H-G. (1989) Eur. J. Biochem. 182, 613-620
30 De Virgilio, C. et aL (1993) Eur. 1
Biochem. 212, 315-323 31 Sur, L P., Loboo Z. and Maitra, P. F,. (1994)
Yeast 10, 199-209 32 Hohmann, 5. et al. (1993) Cuff. Genet. 23,
281-289 33 Luyten, K. et aL (1993) Eur. J. Biochem. 217,
701-713 34 Londesborough, J. and Vuono, O. E. (1993)
Eur. J. Biochem. 216, 841-848 35 Hohmann, S., Van Dijck, P., Luyten, K. and
Thevelein, J. M. (1994) Curt. Genet. 26, 295-301
36 Toda, T. et al. (1985) Ceil 40, 27-36 37 Bl~zquez, M. A., Lagunas, R., Gancedo, C. and
Gancedo, J. M. (1993) FEBS L~ % 329, 51-54
38 Sols, A. (1976) in Reflections i.,~ Biocflemistfy (Kornberg, A., ed.), pp. 199.-2od, Pergamon
39 Neves, M. J. and Fran(~ois, J. (1~92) Biochem. J. 288, 859-864
40 Bn~zquez, M. A., Stucka, R., Feldmann, H. and Gancedo, C. (1992) H/oF1~shop on con!rot of gene expression in yeast, The Center ,or International Meetings on Biology Vol. 9, p. 60
41 Neves, M. J. et al. Curt. Genet. (in press)
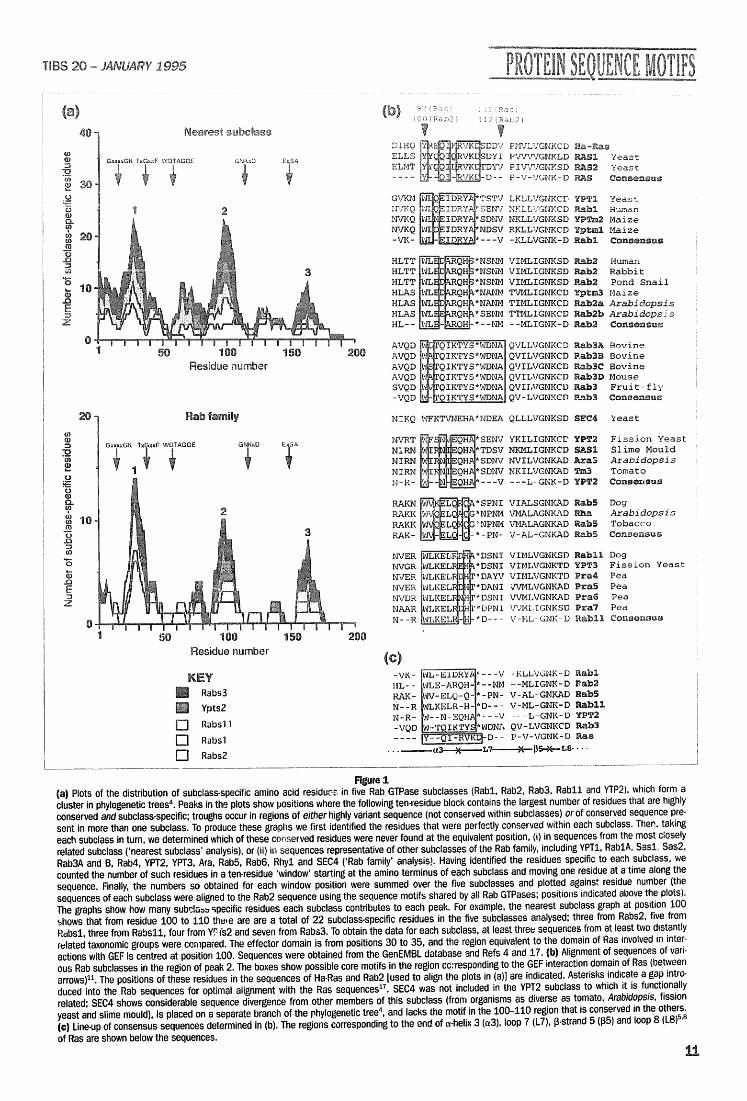
Sub©lass-specific sequen©e m0t[fs identified in Rab GTPases We have identified sequence motifs that may be involved in determining the functional specificity of small Ras-ltke GTPases of the Rub family. Members of the Rub family are critical [or the proper movement of transport vesicles between different compartments o| eukaryotic cells l,'~. The Rub family is divided into subclasses (numbered Rabl to Rub24) based on sequence similarities, in several cases these subclasses are known to reflect functional similarity or equivalence among members from different species. All members of the Rub family appear to be able to bind and hydrolyse GTP; however, members of individual subclasses appear to act specifically at one of several different steps in the endocytic and exocytic pathways. Thus, Rub GTPases have both individual and shared functions that are presumably reflected in their sequences.
The most highly conserved residues of Rub GTPases are involved in nucleotide binding and hydrolysis, functions common to all Ras-like GTPases. Other regions presumably contain the information that specifies individual functions. One such
region is the 'effector region', which interacts with subclass-specific GTPase- activating proteins (GAPs) 2,3. Its sequence is highly conserved within each Rab subclass, but diverges considerably between functionally distinct subclasses 2,3. We have now identified a second subclass-specific region in Rab GTPases.
We grouped Rub sequences into subclasses on tile basis of overall similarity '~ and lde~tical effector regions. We identified those residues that are hwa~ font h~ proteins of one subclass, but that are not found at the equivalent position in proteins of related subclasses. The analysis was limited to subclasses [or which members are known from distantly related taxonomic groups, as this allowed the identification of residues that are most highly conserved within the subclass. The results, plotted in Fig. la, show a peak of subclass-specific residues around the effector region as expected, but a second major peak occurs around position 100 (representing a 'window' from residues 100 to 110 and corresponding to ~-helix 3 and loop 7 of Ha-Ras4.S; see legend to Fig. 1). It is noteworthy that domain-swapping experiments betweeu two yeast Rub proteins showed that this region acts cooperatively with the effector region and the carboxy-terminal membrane-targeting signal to confer functional specificity to each Rub ~ . in fact, there is notable coincidence between the Rub specificity domains thus identified and the peaks of subclass-specific residues in Fig.la, testifying to the functional significance of our analysis. Stenmark e t ai. (1994) have also identified ~-heiix 3--loop 7 as a specificity domain in mammalian Rab5, and shown its functional importance 9.
© 1995, Elsevier Science Ltd 0968-0004/95/$09.50
T~8S 20 - JANUARY 1995
40 N e a r e s t s ~ b e ~ s ~
30
g
..Q
if3
0
ID
Gx:~xGK T~G~xF V~DTAGOE GNK~D
2
1 50 ~OO Residue number
o 0~
0"l
c~
E Z
20
lO
Rab family
Gx~GK Tx~xxF WDTAGQE GNK.D
PROTEIN SEQUENCE MOTIFS . . . . . . . . . . .
Za-P~s a L~L~S! Yeast
~A~2 "feast D~AS Consensus
YBTI Yeast Rabl Human
NVKQ YP%~m2 Maize Y~ml Maize
-VK- Rabl Consensus
HLTT Rab2 Human Rab2 Rabbit Rab2 Pond Snail Y p ~ m 3 Maize Rab2a Arabidopsis ~2b Arabidopsis Rab2 Consensus
15~ 2~0
GVKM ~ * T S T ~ J LKLLVG~,ZKCD
~L~-[E I DRYA~ ~ - - - V - KL LVGNK - D
~ *NShrM VIMLIGNKSD
m T~ Fm~pp~Q~*Ns~ vz~zG~s~ HLAS I~.,E~}~R.QH~*NAlkll~. 'I~v'MI.,IGNKCD HLAS IWL~ARQH~*NANM TINT. IGNKCD HLAS IWLE~ARQH~*SENM TTMLIGNKCD HL-- ~L~-~J .... NM --MLIGNK-D
~VQD F~FQ~_KTVS*~,mNAi QVLLVGm<C~ ~veD I~,~FQZKTYS*~I ewr.wm<c~ AVQD I%~QIKTYS*WDNA I QVILVGNKCD AVQD I%'~FQIKTYS*WDNA~ QVILVGNKCD SVQD ~QIKTYS*WDNA I QVII,VGNKCD -VQD k~-EQIKTYS*WDN ~ QV-LVGNKCD
NIKQ WFKTVNEHA*NDEA QLLLVGNKSD
FF~FFOH~* s~'v
,~- ~ - ~-~H~ . . . . v
RAKN ~ * S P N I VIALSGNKAD
RAKK I~L~G*NPNM \rMALAGNKAD RAK- L_~-~..~-L~-*-PN- V-AL-GNKAD
NVGR IWLKELP~*DSNI VIMLVGNKTD NVER IWLKELR~HF~ DAYV VIMLVGNKTD NVER ~LI<EL~F*DANI VVMLVGNKAD NVDR I~LKELF~*DSNI VVMLVGNKAD ~ R ~ , ~ " ~ m ,~v~,m~Ks~ N--R - * --- V-ML--GNK-D
Rab3A Bovine Rab3B Bovine Rab3C Bovine Rab3D Mouse Rab3 Fruit-fly P~b3 Consensus
SEC4 Neast
E A h[VRT YKILIGNKCD YPT2 NiRN NKMLIGNKCD SASI NIRN NVILVGNKADAra3 NIRN NKILVGNKAD %~m3
---L--Gh~-D YP~2
Fission Yeast Slime Mould Arabidopsis Tomato Consensus
Rab5 Dog Rha Arabidopsis Rab5 Tobacco Rab5 Consensus
Rabll Dog ~PT3 Fission Yeast Pra4 Pea Pra5 Pea Pra6 Pea Pray Pea Rabll Consensus
1 50 100 150 200 Residue number ( C )
KEY -vK- ~ . . . . v -m, LV~NK=D ~abl HL-- *--NM --MLIGNK-D P~b2
M Rabs3 *-PN- V-AL-GNKAD R a b 5 K- ~D--- V -ML-G~-D ~ b Z l
N-R- * .... V .... L-GNK-D YPT2 [] Rabs11 -VQD ~ ~ * W D N A QV-LVGNKCD Rab3
[ ] Rabs| ~ - D - - P - V - V G N K - D R a s
[ ] Rabs2 . . . . (~ 3 " " 'W" -=~ T ' 7 " - " - - - X - - ~ 5=X-- I* e . . . .
Figure 1 (a) Plots of the distribution of subclass-specific amino acid residues, in five Rab GTPase subclasses (Rabl, Rab2, Rab3, Rabll and YTP2), which form a cluster in phylogenetic trees 4. Peaks in the plots show positions where the following ten-residue block contains the largest number of residues ttlat are highly conserved and subclass-specific; troughs occur in regions of either highly variant sequence (not conserved within subclasses) or of conserved sequence pre- sent in more than one subclass. To produce these graphs we first identified the residues that were perfectly conserved within each subclass. Then, taking each subclass in turn, we determined which of these conserved residues were never found at the equivalent position, (i) in sequences from the most closely related subclass ('nearest subclass' analysis), or (ii) in sequences representative of other subclasses of the Rab family, including YPT1, RablA, Sasl, Sas2, Rab3A and B, Rab4, YPT2, YPT3, Ara, Rab5, Rab6, Rhyl and SEC4 ('Rab family' analysis). Having identified the residues specific to each subclass, we counted the number of such residues in a ten-residue 'window' starting at the amino terminus of each subclass and moving one residue at a time along the sequence. Finally, the numbers so obtained for each window position were summed over the five subclasses and plotted against residue number (the sequences of each subclass were aligned to the Rab2 sequence using the sequence motifs shared by all Rab GTPases; positions indicated above the plots). The graphs show how many subda=~.,~pecific residues each subclass contributes to each peak. For example, the nearest subclass graph at position 100 shows that from residue 100 to 110 the, e are are a total of 22 subclass-specific residues in the five subclasses analysed; three from Rabs2, five from Rabsl, three from Rabs11, four from YF fs2 and seven from Rabs3. To obtain the data for each subclass, at least three sequences from at least two distantly related taxonomic groups were compared. The effector domain is from positions 30 to 35, and the region equivalent to the domain of Ras involved in inter- actions with GEF is centred at position 100. Sequences were obtained from the GenEMBL database and Refs 4 and 17. {b) Alignment of sequences of vari- ous Rab subclasses in the region of peak 2. The boxes show possible core motifs in the region cc:responding to tile GEF interaction domain of Ras (between arrows)l< The positions of these residues in the sequences of Ha-Ras and Rab2 [used to align the plots in (a)] are indicated. Asterisks indicate a gap intro- duced into the Rab sequences for optimal alignment with the Ras sequenceslL SEC4 was not included in the YPT2 subclass to which it is functionally related; SEC4 shows considerable sequence divergence from other members of this subclass (from organisms as diverse as tomato, Arabidopsis, fission yeast and slime mould), is placed on a separate branch of the phylogenetic tree 4, and lacks the motif in the 100-110 region that is conserved in the others. (¢} Line-up of consensus sequences determined in (b). The regions corresponding to the end of a-helix 3 (e3), loop 7 (L7), 13-strand 5 (135) and loop 8 (L8) 5'8 of Ras are shown below the sequences.
1 t

. . . . . . . . . . . . . . . . . . . . . . . . w . . . . . . . . . .
PROTEIN SEQUENCE MOTIFS The role of the newly Identified motifs
is not yet clear, However, it Is noteworthy that residues 100 to 110 coincide wl~h a region of Ras that is critical for its interaction with guanine-nucleotide exchange factors (GEFs) l°'lt. GEFs catalyse the release of GDP following nucleotide hydrolysis, allowing a new GTP molecule to be bound. Rub GEF activities 12-t4 appear to act when Rab proteins are recruited onto budding vesicles ts,ts. Considering the similarities between Ras and Rab proteins 4, Rab GEFs might also interact with a-helix 3-loop 7 (Ref, 11). If so, we speculate that the subclass-specific a-helix 3-.loop 7 motifs might facilitate interaction with specific Rab GEFs.
References I Pfeffer, S. (1992) Trer~O's Cell Biol. 2, 41-46 2 Zerlal, M. and Stenmark, H. (1993) Curt. Opin.
Cell Biol. 5, 613-620 3 Haubruck, H., Engelke, U., Mertins, P. and
Galwitz, D. (1990) EML~)J. 9, 1957-1962 4 Valencia, A., Chardin, P., Wittinghoffer, A. and
Sander, C. (1991) Biochemistry 30, 4637-4648 5 Pal, E. F. etal. (1989) Nature 341, 209-214 6 Brennwald, P. and Novick, P. (1993) Nature 362,
560-563 7 Chavrier, P. etal. (1991) Nature 353, 769-772 8 Dunn, B., Stearns, T. and Botstein, D. (1993)
Nature 362, 563-565 9 Stenmark, H. et al. (1994) EMBO J. 13,
575-583 10 Novick, P. and Brennwald, P. (1993) Cell 75,
597-601 11 Segal, M., Willumsen, B. M. and Levitzki, A.
(1993) Proc. Nat/Acad. Sci. USA 90, 5564-5568
TIBS 2 0 - JANUARY 1 9 9 5
12 Burnstein, E. S. and Macara, I. G. (1992) Proc. Natl Acad. Sci. USA 89, 1154-1158
13 Burton, J. etal. (1993) Nature 361,464-467 14 Moya, M., Robe=ts, D. and Novick, P. (1.993)
Nature 361, 460~63 15 Soldati, T., Shapiro, A., Dirac Svejstrup, A. B.
and Pfeffer, S. R. (1994) Nature 369, 76-78 16 UIIrich, 0., Hoduchi, H., ~.ucci, C. and ZeriaL M.
(1994) Nature 368, 157-160 17 Palme, K. etal. (1992) Proc. Natf Acad. Sci.
USA 89, 787-791
IAN MOORE
Department of Plant Sciences, University of Oxford, South Parks Road, Oxford, UK OXl 3RB.
JEFF $CHELL AND KL~.U$ PALME
Max Plenck lnstitut f~r ZQchtungsforschung, Cafl-von-Linn6.Weg 10, D-50829, K61n, Germany.
A new family of peptidyl-prolym isomemses
Peptidyhprolyl [somerases (PPlases) are ubiquitous and are thought to assist in protein folding by catalysing the cis/trans isomerlzatlon of the peptidyl-prolyl bonds in peptides and proteins ~-7, They are also called Immunophilins, since they bind immuuosuppresslve drugs such as cyclosporln, FKS06 and rapamycm. The PPlases described so far fall Into two families, the cyclophllins (cyclosporln- binding proteins) and the FKS06~bindlng proteins (FKBPs), which also bind rapamyctn, The amino add sequences are highly conserved within each faintly but there Is no detectable conservation between the f~milles ~ . Here we delineate a new, third family of PHases, based on the Identification of the gone for the prototype member of the group, namely the Escherichia colt PplC protein. Inclusion In this family allows us to predict that the bacterial proteins PrsA, SurA and NIfM, for which no enzymatic activity has been reported so far, are PPhses. The known properties of these proteins support the hypothesis that they are involved in protein folding.
In £'. colt, two cyclophllln genes s and two genes coding for FKBP-related proteins 9,|° have been identified. However, a novel PHase activity has recently been purified from E. coli and assigned a molecular weight of 10102 ± 2 Da by high-precision electrospray mass spectrometry u. The sequence of the 21 amino-terminal residues of this protein has been determined u. When this sequence was compared with those in the protein sequence databases, no significant similarity was revealed. However, when
12
we compared it with the conceptually translated nucleotide sequence databases using the TBLASTN program ~2, 100% identity with a previously uncharacterized sequence encoded in the 'gray hole' (chromosome region without genes) between the E. coil ilvC and rap genes in the ECOUW85U sequence t3 was observed. The open reading frame (ORF) coding for the new PHase was interrupted by two DNA sequencing frameshlft errors that we have now corrected by re-sequencing (see legend of F!g. ! for details). This new ORF encodes a proteln cons!sting o| 93 residues, and the mature protein (no fMet) has a calculated molecular weight of l0101 Do. This is remarkably close to the expe'Amental value. We therefore delh~e tMs protel~ as the new E. coli PHase, PplC (a different name, parvulin, has been given to this protein by its discoverers; G, Fischer, pars. commun.).
A database search with the PplC sequence detected highly significant similarity with the Bacillus subtilis iipoprotein PrsA (probability of matching by chance, P < 10 -l~- as computed using the BLASTP program), the E. coil protein SurA (e < I0"9), and NifM proteins from nitrogen-assimilating bacteria (P < 104); a limited similarity was also observed with Lactococcus protease maturation protein PrtM. Further computer analysis, combining searches with BLASTP and TBLa, STN with multiple alignment using the MACAW program ~, revealed three motifs that are conserved in all these proteins as well as in an uncharacterized B. subtilis ORF product, and two putative eukaryotlc proteins encoded by a previously unrecognized ORF from Dictyostelium and a human expressed- sequence tag or EST (Fig. 1). The putative human EST product showed significant similarity to PpiC (e < 10 -4) whereas the
Dictyostelium sequence is related to SurA (P < 10-4). Together, the conserved alignment blocks spanned almost the entire length of PpiC. The alignment in Fig. 1 defines a new protein family that is conserved in both bacteria and eukaryotes. By analogy with PpiC, it is likely that all the proteins in this family have PHase activity.
The new family of putative PPlases showed no detectable similarity to either cyclophilins or FKBP. Moreover, while the experimentally determined structures o| both cyclophilhls and FKBP are 9-barre!s L~,"~, the PpiC~related domains were predicted to consist predominantly o[ ~-helices, with only two 9ostrands near the termini off PpiC (Fig. l; Re|. 17). The proteiris in the new PPlase family are similar to the other two PPlase classes in that they are hydropMlic and do not contain any predicted transmembrane segments 4. However, PrsA and PrtM are associated with the cytoplasmic membrane via a conjugated lipid moietytS.~9; these proteins also have amino-terminal signal peptides that are cleaved by the signal peptidase concomitantly with their transport across the cytoplasmic membrane and into the periplasm m.tg. A similar signal is prescott at the amino terminus of SurA and is cleaved from the mature protein that has been isolated from the periplasm (S. Lazar and R. Kolter, unpublished). On the other hand, no signal peptide is predicted in PplC or NifM. PpiC, which is smaller than any other known PPIase, is likely to comprise the basic PHase domain. Interestingly, this domain is duplicated in SurA, while all the other proteins in the new family apparently contain additional domains (Fig. 1) r
Motifs il and Ill are each predicted to contain a long, presumably flexible loop with a conserved glycine (Fig. 1);





























