Embed Size (px)
Citation preview

Jour
nal o
f Cel
l Sci
ence
RESEARCH ARTICLE
Subnuclear domain proteins in cancer cells support the functionsof RUNX2 in the DNA damage response
Seungchan Yang1, Alexandre J. C. Quaresma1,2, Jeffrey A. Nickerson1, Karin M. Green3, Scott A. Shaffer3,Anthony N. Imbalzano1, Lori A. Martin-Buley4, Jane B. Lian1,4, Janet L. Stein1,4, Andre J. van Wijnen1,5 andGary S. Stein1,4,*
ABSTRACT
Cancer cells exhibit modifications in nuclear architecture and
transcriptional control. Tumor growth and metastasis are
supported by RUNX family transcriptional scaffolding proteins,
which mediate the assembly of nuclear-matrix-associated gene-
regulatory hubs. We used proteomic analysis to identify RUNX2-
dependent protein–protein interactions associated with the nuclear
matrix in bone, breast and prostate tumor cell types and found that
RUNX2 interacts with three distinct proteins that respond to DNA
damage – RUVBL2, INTS3 and BAZ1B. Subnuclear foci containing
these proteins change in intensity or number following UV
irradiation. Furthermore, RUNX2, INTS3 and BAZ1B form UV-
responsive complexes with the serine-139-phosphorylated isoform
of H2AX (cH2AX). UV irradiation increases the interaction of BAZ1B
with cH2AX and decreases histone H3 lysine 9 acetylation levels,
which mark accessible chromatin. RUNX2 depletion prevents the
BAZ1B–cH2AX interaction and attenuates loss of H3K9 and H3K56
acetylation. Our data are consistent with a model in which RUNX2
forms functional complexes with BAZ1B, RUVBL2 and INTS3 to
mount an integrated response to DNA damage. This proposed
cytoprotective function for RUNX2 in cancer cells might clarify its
expression in chemotherapy-resistant and/or metastatic tumors.
KEY WORDS: RUNX2, DNA damage response, Nuclear matrix,
Proteomics, Cancer, Breast, Prostate, Osteosarcoma, INTS3,
BAZ1B
INTRODUCTIONNuclei are highly structured, and the spatial organization that
supports nuclear metabolism is increasingly well characterized.
This nuclear architecture encompasses two interconnected
structures – chromatin and a nuclear matrix (Berezney et al.,
1995; van Driel et al., 1995; Stenoien et al., 1998; Zink et al.,
2004; Kubben et al., 2010; Markaki et al., 2010; Meldi and
Brickner, 2011; Simon and Wilson, 2011). Molecular complexes
that perform and regulate transcription, RNA processing, DNA
replication, DNA repair and apoptosis, and that organize
chromatin structure are localized within distinct nuclear
domains. The ultrastructure of the nuclear matrix is well
characterized and, in addition to protein and DNA, requires
RNA to maintain its integrity (Nickerson et al., 1997). The
protein composition of the isolated nuclear matrix was initially
analyzed using two-dimensional gel electrophoresis (Fey and
Penman, 1988; Dworetzky et al., 1990; Getzenberg et al., 1991).
This approach indicated that the nuclear matrix is comprised of
§200 proteins and that its composition is partially cell-type
specific. Several dozen nuclear matrix proteins have been
identified, including Numa, matrin, HnRNPs, NMP1 (also
known as YY1) and NMP2 (also known as RUNX2) (Fey and
Penman, 1988; Bidwell et al., 2001; Merriman et al., 1995; Javed
et al., 2005), revealing that the nuclear matrix contains both
structural and regulatory components.
Molecular characterization of the nuclear matrix protein NMP2
(Bidwell et al., 1993; Merriman et al., 1995) established that this
protein is a member of the RUNX (AML/PEBP2alpha/CBFA)
family of lineage-specific transcription factors, which control
tissue development and have pathological roles in cancer (Zaidi
et al., 2007a). RUNX proteins are localized at specific subnuclear
domains through peptide-targeting sequences (Zeng et al., 1997;
Zeng et al., 1998). They participate in the scaffolding of
macromolecular protein–protein complexes that control gene
transcription by supporting chromatin-related epigenetic
mechanisms (Zaidi et al., 2001). These mechanisms involve
formation of multiple complexes with proteins that mediate
histone modifications or chromatin remodeling (Delcuve et al.,
2009), including protein histone acetyl transferases (HATs, such
as p300), histone deacetylases (e.g. HDACs), co-regulators (e.g.
TLE-1, a homolog of groucho), YAP and SMADs (Zaidi et al.,
2002; Zaidi et al., 2004) Disruption of RUNX protein subnuclear
targeting alters transcriptional programs and compromises cell
growth and differentiation (Zaidi et al., 2006), reflecting its
pathological linkage to acute myelogenous leukemia and breast
and prostate cancers (Zaidi et al., 2007b).
RUNX2 (NMP2) is a master regulator of skeletal development
(Lian et al., 2004; Lian et al., 2006; Kuo et al., 2009), and has
been linked to bone cancer and metastases from other cancers
(Pratap et al., 2011). Because RUNX2 is a rate-limiting
scaffolding protein that is crucial for the molecular organization
of both transcriptional and epigenetic complexes at multiple
target genes within matrix-associated subnuclear domains,
establishing which nuclear matrix proteins are linked to the
diverse biological functions of RUNX2 is a key objective. In this
1Department of Cell Biology, University of Massachusetts Medical School,Worcester, MA 01655, USA. 2Institute of Biomedicine, Department ofBiochemistry and Developmental Biology, FI-00014 University of Helsinki,Finland. 3Department of Biochemistry and Molecular Pharmacology andProteomics and Mass Spectrometry Facility, University of Massachusetts MedicalSchool, Worcester, MA 01605, USA. 4Department of Biochemistry & VermontCancer Center, University of Vermont Medical School, Burlington, VT 05405, USA.5Departments of Orthopedic Surgery & Biochemistry and Molecular Biology,Mayo Clinic, 200 First Street S.W., MSB 3-69, Rochester, MN 55905, USA.
*Author for correspondence ([email protected])
Received 21 July 2014; Accepted 11 December 2014
� 2015. Published by The Company of Biologists Ltd | Journal of Cell Science (2015) 128, 728–740 doi:10.1242/jcs.160051
728

Jour
nal o
f Cel
l Sci
ence
study, we used advanced protein mass spectrometry andproteomic analysis to identify proteins that are recruited to the
nuclear matrix in a RUNX2-dependent manner and that associatewith RUNX2 in bone, breast and prostate cancer cells. Strikingly,we found that three proteins with previously reported separateand distinct functions might associate with RUNX2 to form a
novel, multifunctional integrated protein complex involved in theUV-induced DNA damage response. Our data indicate thatRUNX2 supports the scaffolding of these proteins into an
integrated complex that is coupled to the DNA damageresponse. This finding suggests that RUNX2 is a directparticipant and regulator of the DNA damage response, thus
providing a new molecular dimension in our understanding ofDNA repair and its dysregulation in cancer.
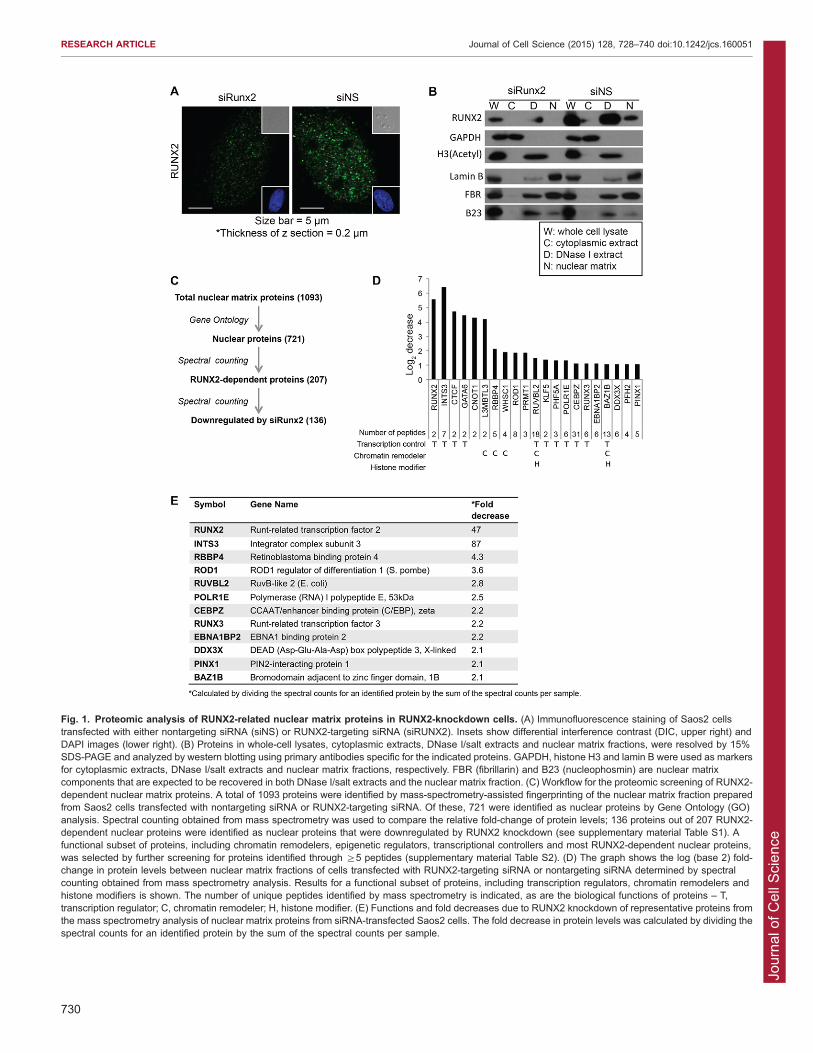
RESULTSDefinition of a RUNX2-dependent proteome associated withthe nuclear matrixRUNX2 localizes in subnuclear domains (Fig. 1A) that are
associated with the nuclear matrix (Zaidi et al., 2001). Toinvestigate RUNX2-related protein complexes in this nuclease-resistant chromatin compartment by proteomic analysis and analyze
the extent to which the protein profile of this compartment isRUNX2 dependent, we used siRNA-mediated knockdown ofRUNX2 in Saos2 osteosarcoma cells. We transiently transfected
cells with siRNA targeting RUNX2 and examined the subcellularlocalization of RUNX protein by confocal immunofluorescencemicroscopy. The number of RUNX2-containing nuclear foci was
diminished in cells transfected with RUNX2-targeting siRNA(siRUNX2), but gross nuclear morphology was maintained(Fig. 1A). Cells were then fractionated and proteins fromdifferent subcellular compartments were analyzed by SDS-PAGE
and western blotting, using markers to validate the fractionation ofcytoplasmic, chromatin and nuclear matrix compartments (e.g.GAPDH was used as a cytoplasmic marker, acetylated histone H3
as a chromatin marker and lamin B as a nuclear matrix marker)(Fig. 1B). siRNA-mediated depletion of RUNX2 greatly reducedRUNX2 protein levels in whole-cell lysates and chromatin
fractions, and rendered it undetectable in the nuclear matrixcompartment. RUNX2 knockdown did not, however, alter theoverall composition of the most abundant nuclear proteins residingin the nuclear matrix fraction, as visualized by staining the gel with
Coomassie Blue (data not shown).
Mass spectrometry analysis of the nuclear matrix proteomein bone cancer cellsNanoliquid chromatography tandem mass spectrometry (nanoLC-MS/MS) was used to examine the molecular consequences of
RUNX2 depletion on the proteomic profile of nuclear matrixproteins. Proteins in the nuclear matrix fraction from control andsiRNA-transfected cells were separated by SDS-PAGE and
processed for nanoLC-MS/MS (Dzieciatkowska et al., 2014). Intotal, .1000 proteins were identified from the product ion spectrausing the Swissprot database (supplementary material Table S1),and .700 nuclear proteins were selected based on Gene Ontology
(GO) analysis (Fig. 1C). To assess differences in the relativeprotein abundance in the nuclear matrix fraction between cellstransfected with nontargeting control or RUNX2-targeting siRNA,
we used semi-quantitative spectral counting (Zybailov et al., 2005;Lundgren et al., 2010). With this approach, we found that .100proteins in the nuclear matrix fraction are downregulated .2-fold
in cells transfected with siRNA targeting RUNX2 (Fig. 1C). Of
this subset, we selected proteins that exhibited the greatest relativechange when RUNX2 was depleted by siRNA-mediated
knockdown or that are known to have a role in transcriptionalregulation, chromatin remodeling and/or histone modification(Fig. 1D,E; supplementary material Table S2). For example, incells with a 40-fold reduction in RUNX2 levels, INTS3 levels
exhibit a .80-fold decrease.Because the transcription factor RUNX2 might modulate the
levels of distinct proteins in the nuclear matrix by regulating
transcription or protein–protein interactions, we examined themRNA expression of selected proteins by reverse transcription-quantitative polymerase chain reaction (RT-qPCR) analysis. As
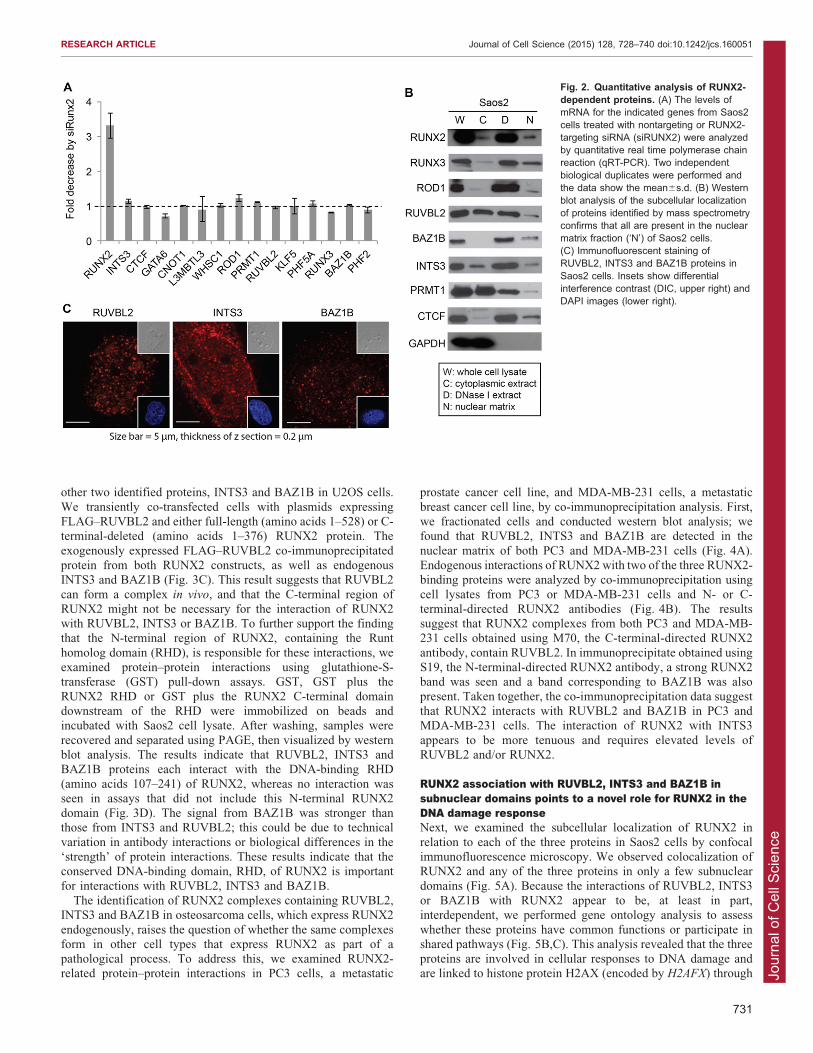
expected, RUNX2 mRNA levels in cells transfected with RUNX2siRNA were decreased 3–4-fold compared to levels in cellstransfected with nontargeting siRNA (Fig. 2A). However, mRNA
expression levels corresponding to the proteins identified in ourproteomic screen did not display significant changes due totransfection with siRNA targeting RUNX2 (Fig. 2A). Thissuggests that the differences in protein abundance observed by
proteomic analysis (see Fig. 1D) are not due to RUNX2-dependent transcriptional regulation. RUNX2-dependent proteinsthat were detected by at least five unique peptides in nanoLC-MS/
MS-based protein profiling and showed promise as regulatorycontributors by preliminary bioinformatic analysis were confirmedto be localized in the nuclear matrix compartment by western blot
analysis (Fig. 2B). Proteins of interest, PRMT1 and CTCF, werealso included in the analysis. Collectively, these data indicate thatchanges in the nuclear matrix association of proteins upon RUNX2
depletion are mediated by post-transcriptional mechanisms (e.g.protein–protein interactions, subcellular compartmentalization).
We selected three RUNX2-interacting proteins, RUVBL2,INTS3 and BAZ1B, for further studies, because their functions
are broadly related to chromatin organization, DNA repair and/orformation of protein–protein complexes that might be relevant tothe molecular pathology of cancer cells. To support our proteomic
identification, we examined the endogenous cellular localizationof these proteins in Saos2 cells using immunofluorescenceconfocal microscopy (Fig. 2C). Nuclear foci were seen using
primary antibodies that target RUVBL2, INTS3 and BAZ1B(Fig. 2C), consistent with the possibility that these proteinslocalize in the nuclear matrix in association with RUNX2.
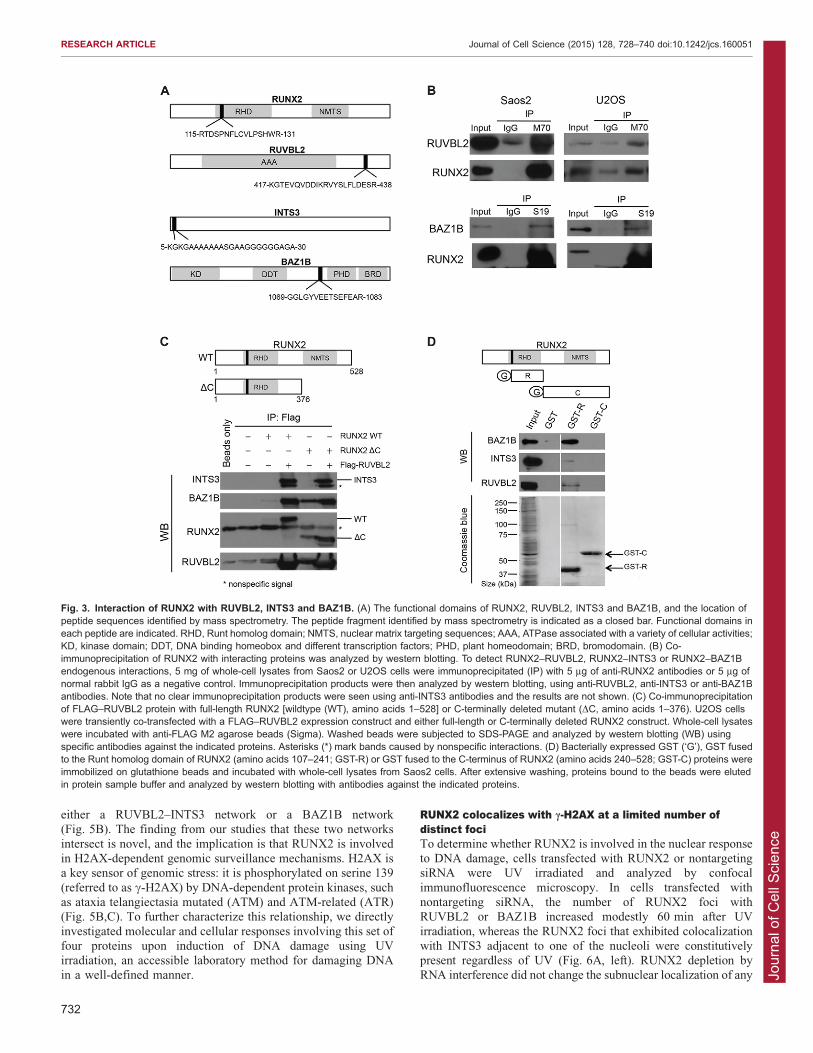
Selective interactions of RUNX2 with RUVBL2, INTS3and BAZ1BWe examined whether RUNX2 changes the nuclear matrix
association of RUVBL2, INTS3 and BAZ1B proteins by theirrecruitment through protein–protein interactions (Fig. 3A). Co-immunoprecipitation experiments revealed that endogenous
RUVBL2 and BAZ1B each exhibit interactions with RUNX2 inosteosarcoma cell lines, Saos2 and U2OS (Fig. 3B). EndogenousRUVBL2 protein present in lysates of Saos2 and U2OS cells
was effectively co-immunoprecipitated with M70 antibody,which recognizes a C-terminal domain in RUNX2. BAZ1B wasalso co-immunoprecipitated using S19 antibody, which interactswith the N-terminal region of RUNX2 (Fig. 3B). We did not
detect a clear interaction between endogenous RUNX2 andINTS3 using either the C- or N-terminally directed RUNX2antibodies, but were able to readily detect RUNX2–INTS3-
complexed proteins in RUNX2 and RUVBL2 co-expressionanalysis (see below).
Because RUVBL2 is a molecular chaperone (Izumi et al.,
2010), we tested whether RUVBL2 can recruit RUNX2 and the
RESEARCH ARTICLE Journal of Cell Science (2015) 128, 728–740 doi:10.1242/jcs.160051
729
Jour
nal o
f Cel
l Sci
ence
Fig. 1. Proteomic analysis of RUNX2-related nuclear matrix proteins in RUNX2-knockdown cells. (A) Immunofluorescence staining of Saos2 cellstransfected with either nontargeting siRNA (siNS) or RUNX2-targeting siRNA (siRUNX2). Insets show differential interference contrast (DIC, upper right) andDAPI images (lower right). (B) Proteins in whole-cell lysates, cytoplasmic extracts, DNase I/salt extracts and nuclear matrix fractions, were resolved by 15%SDS-PAGE and analyzed by western blotting using primary antibodies specific for the indicated proteins. GAPDH, histone H3 and lamin B were used as markersfor cytoplasmic extracts, DNase I/salt extracts and nuclear matrix fractions, respectively. FBR (fibrillarin) and B23 (nucleophosmin) are nuclear matrixcomponents that are expected to be recovered in both DNase I/salt extracts and the nuclear matrix fraction. (C) Workflow for the proteomic screening of RUNX2-dependent nuclear matrix proteins. A total of 1093 proteins were identified by mass-spectrometry-assisted fingerprinting of the nuclear matrix fraction preparedfrom Saos2 cells transfected with nontargeting siRNA or RUNX2-targeting siRNA. Of these, 721 were identified as nuclear proteins by Gene Ontology (GO)analysis. Spectral counting obtained from mass spectrometry was used to compare the relative fold-change of protein levels; 136 proteins out of 207 RUNX2-dependent nuclear proteins were identified as nuclear proteins that were downregulated by RUNX2 knockdown (see supplementary material Table S1). Afunctional subset of proteins, including chromatin remodelers, epigenetic regulators, transcriptional controllers and most RUNX2-dependent nuclear proteins,was selected by further screening for proteins identified through §5 peptides (supplementary material Table S2). (D) The graph shows the log (base 2) fold-change in protein levels between nuclear matrix fractions of cells transfected with RUNX2-targeting siRNA or nontargeting siRNA determined by spectralcounting obtained from mass spectrometry analysis. Results for a functional subset of proteins, including transcription regulators, chromatin remodelers andhistone modifiers is shown. The number of unique peptides identified by mass spectrometry is indicated, as are the biological functions of proteins – T,transcription regulator; C, chromatin remodeler; H, histone modifier. (E) Functions and fold decreases due to RUNX2 knockdown of representative proteins fromthe mass spectrometry analysis of nuclear matrix proteins from siRNA-transfected Saos2 cells. The fold decrease in protein levels was calculated by dividing thespectral counts for an identified protein by the sum of the spectral counts per sample.
RESEARCH ARTICLE Journal of Cell Science (2015) 128, 728–740 doi:10.1242/jcs.160051
730
Jour
nal o
f Cel
l Sci
ence
other two identified proteins, INTS3 and BAZ1B in U2OS cells.
We transiently co-transfected cells with plasmids expressingFLAG–RUVBL2 and either full-length (amino acids 1–528) or C-terminal-deleted (amino acids 1–376) RUNX2 protein. The
exogenously expressed FLAG–RUVBL2 co-immunoprecipitatedprotein from both RUNX2 constructs, as well as endogenousINTS3 and BAZ1B (Fig. 3C). This result suggests that RUVBL2
can form a complex in vivo, and that the C-terminal region ofRUNX2 might not be necessary for the interaction of RUNX2with RUVBL2, INTS3 or BAZ1B. To further support the findingthat the N-terminal region of RUNX2, containing the Runt
homolog domain (RHD), is responsible for these interactions, weexamined protein–protein interactions using glutathione-S-transferase (GST) pull-down assays. GST, GST plus the
RUNX2 RHD or GST plus the RUNX2 C-terminal domaindownstream of the RHD were immobilized on beads andincubated with Saos2 cell lysate. After washing, samples were
recovered and separated using PAGE, then visualized by westernblot analysis. The results indicate that RUVBL2, INTS3 andBAZ1B proteins each interact with the DNA-binding RHD
(amino acids 107–241) of RUNX2, whereas no interaction wasseen in assays that did not include this N-terminal RUNX2domain (Fig. 3D). The signal from BAZ1B was stronger thanthose from INTS3 and RUVBL2; this could be due to technical
variation in antibody interactions or biological differences in the‘strength’ of protein interactions. These results indicate that theconserved DNA-binding domain, RHD, of RUNX2 is important
for interactions with RUVBL2, INTS3 and BAZ1B.The identification of RUNX2 complexes containing RUVBL2,
INTS3 and BAZ1B in osteosarcoma cells, which express RUNX2
endogenously, raises the question of whether the same complexesform in other cell types that express RUNX2 as part of apathological process. To address this, we examined RUNX2-related protein–protein interactions in PC3 cells, a metastatic
prostate cancer cell line, and MDA-MB-231 cells, a metastatic
breast cancer cell line, by co-immunoprecipitation analysis. First,we fractionated cells and conducted western blot analysis; wefound that RUVBL2, INTS3 and BAZ1B are detected in the
nuclear matrix of both PC3 and MDA-MB-231 cells (Fig. 4A).Endogenous interactions of RUNX2 with two of the three RUNX2-binding proteins were analyzed by co-immunoprecipitation using
cell lysates from PC3 or MDA-MB-231 cells and N- or C-terminal-directed RUNX2 antibodies (Fig. 4B). The resultssuggest that RUNX2 complexes from both PC3 and MDA-MB-231 cells obtained using M70, the C-terminal-directed RUNX2
antibody, contain RUVBL2. In immunoprecipitate obtained usingS19, the N-terminal-directed RUNX2 antibody, a strong RUNX2band was seen and a band corresponding to BAZ1B was also
present. Taken together, the co-immunoprecipitation data suggestthat RUNX2 interacts with RUVBL2 and BAZ1B in PC3 andMDA-MB-231 cells. The interaction of RUNX2 with INTS3
appears to be more tenuous and requires elevated levels ofRUVBL2 and/or RUNX2.
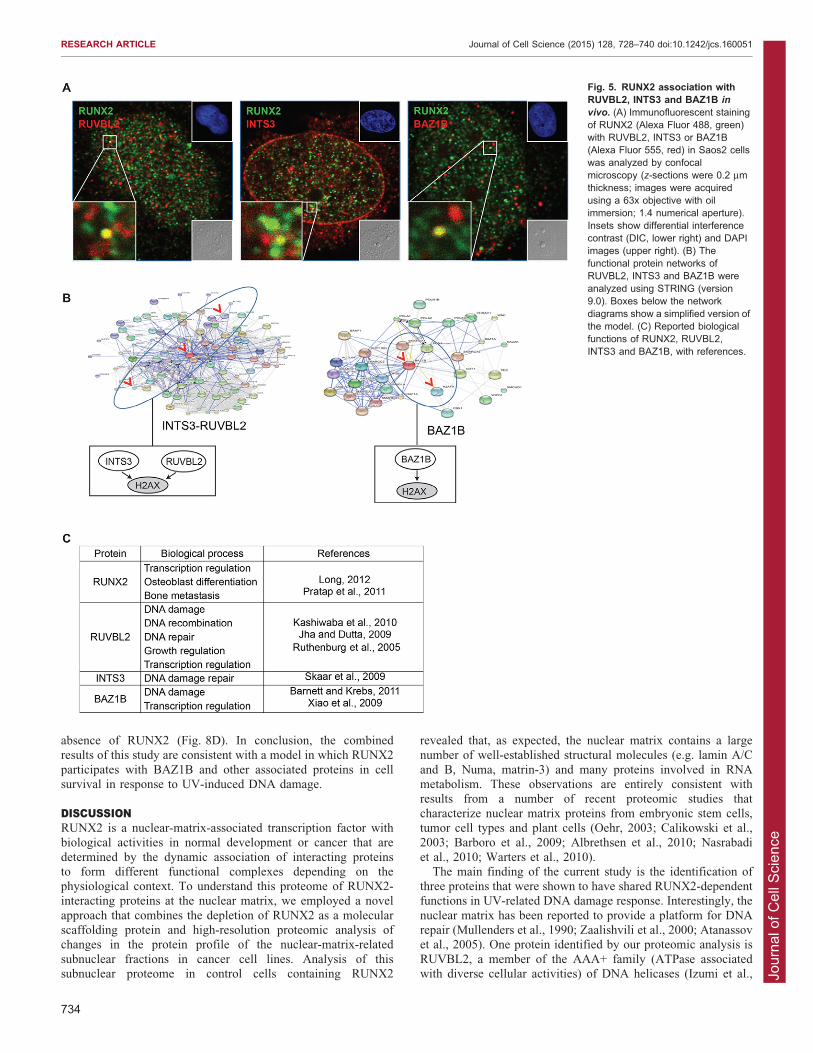
RUNX2 association with RUVBL2, INTS3 and BAZ1B insubnuclear domains points to a novel role for RUNX2 in theDNA damage responseNext, we examined the subcellular localization of RUNX2 in
relation to each of the three proteins in Saos2 cells by confocalimmunofluorescence microscopy. We observed colocalization ofRUNX2 and any of the three proteins in only a few subnuclear
domains (Fig. 5A). Because the interactions of RUVBL2, INTS3or BAZ1B with RUNX2 appear to be, at least in part,interdependent, we performed gene ontology analysis to assess
whether these proteins have common functions or participate inshared pathways (Fig. 5B,C). This analysis revealed that the threeproteins are involved in cellular responses to DNA damage andare linked to histone protein H2AX (encoded by H2AFX) through
Fig. 2. Quantitative analysis of RUNX2-dependent proteins. (A) The levels ofmRNA for the indicated genes from Saos2cells treated with nontargeting or RUNX2-targeting siRNA (siRUNX2) were analyzedby quantitative real time polymerase chainreaction (qRT-PCR). Two independentbiological duplicates were performed andthe data show the mean6s.d. (B) Westernblot analysis of the subcellular localizationof proteins identified by mass spectrometryconfirms that all are present in the nuclearmatrix fraction (‘N’) of Saos2 cells.(C) Immunofluorescent staining ofRUVBL2, INTS3 and BAZ1B proteins inSaos2 cells. Insets show differentialinterference contrast (DIC, upper right) andDAPI images (lower right).
RESEARCH ARTICLE Journal of Cell Science (2015) 128, 728–740 doi:10.1242/jcs.160051
731
Jour
nal o
f Cel
l Sci
ence
either a RUVBL2–INTS3 network or a BAZ1B network(Fig. 5B). The finding from our studies that these two networks
intersect is novel, and the implication is that RUNX2 is involvedin H2AX-dependent genomic surveillance mechanisms. H2AX isa key sensor of genomic stress: it is phosphorylated on serine 139
(referred to as c-H2AX) by DNA-dependent protein kinases, suchas ataxia telangiectasia mutated (ATM) and ATM-related (ATR)(Fig. 5B,C). To further characterize this relationship, we directly
investigated molecular and cellular responses involving this set offour proteins upon induction of DNA damage using UVirradiation, an accessible laboratory method for damaging DNAin a well-defined manner.
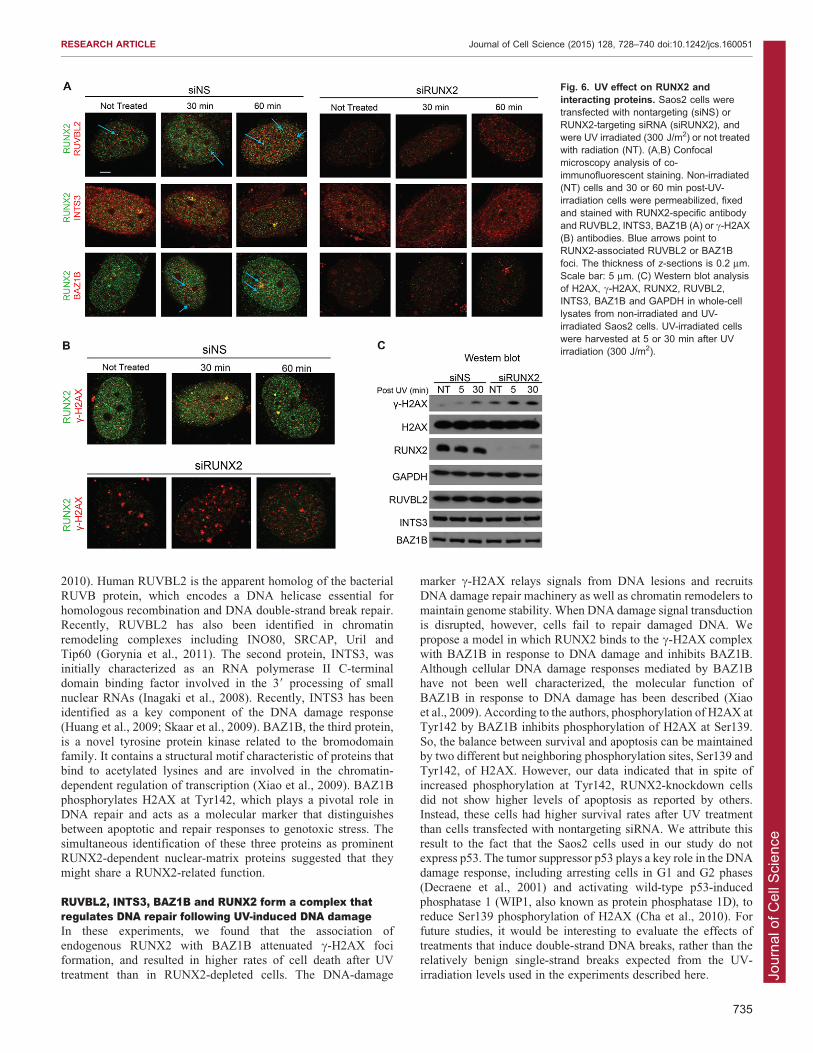
RUNX2 colocalizes with c-H2AX at a limited number ofdistinct fociTo determine whether RUNX2 is involved in the nuclear responseto DNA damage, cells transfected with RUNX2 or nontargetingsiRNA were UV irradiated and analyzed by confocal
immunofluorescence microscopy. In cells transfected withnontargeting siRNA, the number of RUNX2 foci withRUVBL2 or BAZ1B increased modestly 60 min after UV
irradiation, whereas the RUNX2 foci that exhibited colocalizationwith INTS3 adjacent to one of the nucleoli were constitutivelypresent regardless of UV (Fig. 6A, left). RUNX2 depletion byRNA interference did not change the subnuclear localization of any
Fig. 3. Interaction of RUNX2 with RUVBL2, INTS3 and BAZ1B. (A) The functional domains of RUNX2, RUVBL2, INTS3 and BAZ1B, and the location ofpeptide sequences identified by mass spectrometry. The peptide fragment identified by mass spectrometry is indicated as a closed bar. Functional domains ineach peptide are indicated. RHD, Runt homolog domain; NMTS, nuclear matrix targeting sequences; AAA, ATPase associated with a variety of cellular activities;KD, kinase domain; DDT, DNA binding homeobox and different transcription factors; PHD, plant homeodomain; BRD, bromodomain. (B) Co-immunoprecipitation of RUNX2 with interacting proteins was analyzed by western blotting. To detect RUNX2–RUVBL2, RUNX2–INTS3 or RUNX2–BAZ1Bendogenous interactions, 5 mg of whole-cell lysates from Saos2 or U2OS cells were immunoprecipitated (IP) with 5 mg of anti-RUNX2 antibodies or 5 mg ofnormal rabbit IgG as a negative control. Immunoprecipitation products were then analyzed by western blotting, using anti-RUVBL2, anti-INTS3 or anti-BAZ1Bantibodies. Note that no clear immunoprecipitation products were seen using anti-INTS3 antibodies and the results are not shown. (C) Co-immunoprecipitationof FLAG–RUVBL2 protein with full-length RUNX2 [wildtype (WT), amino acids 1–528] or C-terminally deleted mutant (DC, amino acids 1–376). U2OS cellswere transiently co-transfected with a FLAG–RUVBL2 expression construct and either full-length or C-terminally deleted RUNX2 construct. Whole-cell lysateswere incubated with anti-FLAG M2 agarose beads (Sigma). Washed beads were subjected to SDS-PAGE and analyzed by western blotting (WB) usingspecific antibodies against the indicated proteins. Asterisks (*) mark bands caused by nonspecific interactions. (D) Bacterially expressed GST (‘G’), GST fusedto the Runt homolog domain of RUNX2 (amino acids 107–241; GST-R) or GST fused to the C-terminus of RUNX2 (amino acids 240–528; GST-C) proteins wereimmobilized on glutathione beads and incubated with whole-cell lysates from Saos2 cells. After extensive washing, proteins bound to the beads were elutedin protein sample buffer and analyzed by western blotting with antibodies against the indicated proteins.
RESEARCH ARTICLE Journal of Cell Science (2015) 128, 728–740 doi:10.1242/jcs.160051
732

Jour
nal o
f Cel
l Sci
ence
of the three proteins after UV irradiation, albeit that thelocalization of INTS3 in the nucleus was transiently altered
30 min after UV (Fig. 6A, right). The total protein level of c-H2AX in irradiated cells increased after UV exposure; however,RUNX2, RUVBL2, INTS3 and BAZ1B levels did not change
(Fig. 6C). This suggests that foci for RUVBL2, INTS3 andBAZ1B change in number or appearance upon UV exposure, butcan form independently of RUNX2 (Fig. 6A). Because BAZ1B
binds to both c-H2AX (Barnes et al., 2010) and RUNX2 (Fig. 3),we examined whether RUNX2 associates with c-H2AX in situ.We found that both large and small RUNX2 foci containing c-H2AX were present at 30 min after UV irradiation (Fig. 6B). The
partial colocalization of RUNX2 with c-H2AX is consistent withour observation that the c-H2AX-binding protein BAZ1Binteracts with RUNX2.
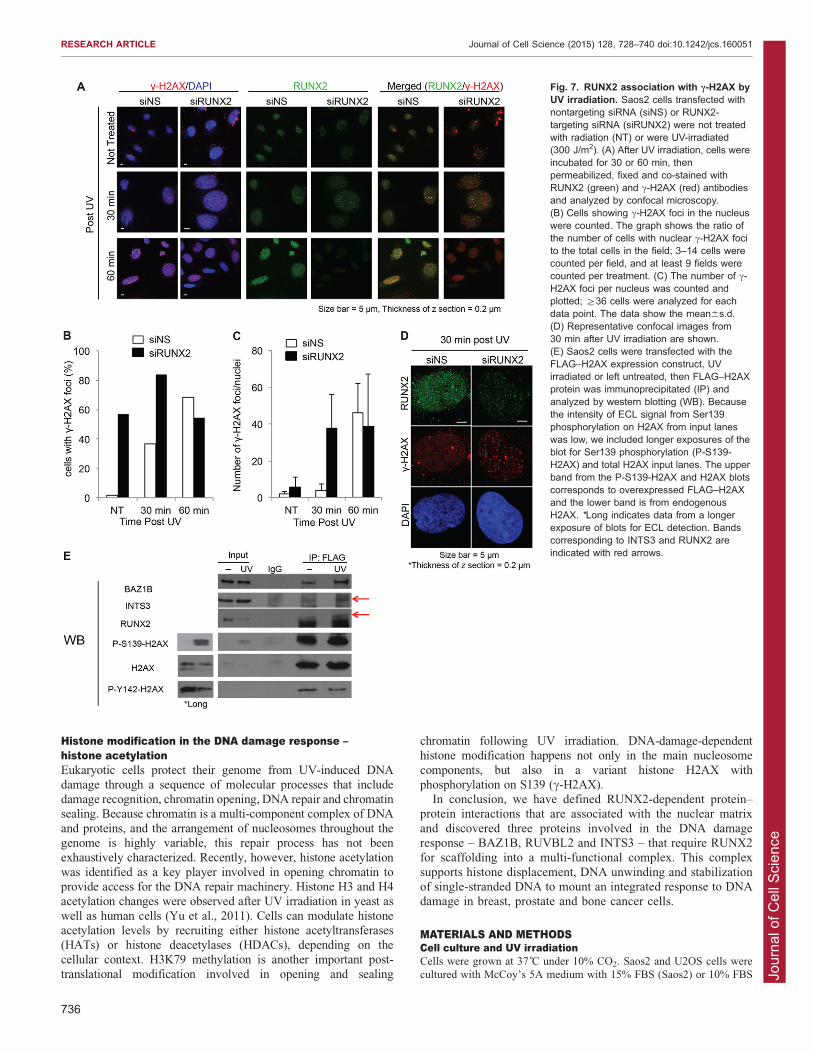
To investigate whether RUNX2 plays a role in the formation ofc-H2AX foci, we performed RUNX2 knockdown in Saos-2 cells(Figs 6, 7). Importantly, the total levels of c-H2AX were
increased in siRUNX2-transfected cells compared to the levelsin cells transfected with negative-control siRNA (siNS) (Fig. 6C).This finding is consistent with similar observations in RUNX2-null cells (Zaidi et al., 2007b). As expected, UV irradiation
increased the percentage of cells that exhibited nuclear c-H2AXfoci (Fig. 7B), as well as the number of foci per cell innontargeting-siRNA-treated control cells (Fig. 7C). However, in
RUNX2-depleted cells, there was a basal level of c-H2AX foci inthe majority of the cells (Fig. 7A,B), which increased after UVtreatment (Fig. 7C,D). Taken together, our results suggest that
RUNX2 depletion enhances the formation of c-H2AX foci in thenucleus.
Because RUNX2 interacts with the c-H2AX-binding protein
BAZ1B, we examined whether RUNX2 and c-H2AX participatein protein–protein interactions within a larger complex. Saos2cells were transfected with FLAG–H2AX expression construct,then FLAG–H2AX was immunoprecipitated from cell lysates. As
expected, BAZ1B did co-immunoprecipitate with FLAG–H2AX(Fig. 7E). The immunoprecipitates also contained INTS3 andRUNX2, but signal above background was only seen in lysates
from UV-irradiated cells. BAZ1B is a protein kinase thatphosphorylates H2AX on tyrosine 142 (Y142) (Xiao et al.,
2009). Indeed, phosphorylation of Y142 is observed in H2AXimmunoprecipitates (Fig. 7E). These results suggest that RUNX2
binds to BAZ1B and c-H2AX in response to UV.
RUNX2 decreases histone H3 acetylation on lysine 9 inresponse to UV irradiationBecause of the interrelationships between RUNX2, BAZ1B, c-H2AX and the cellular response to UV, we further explored the
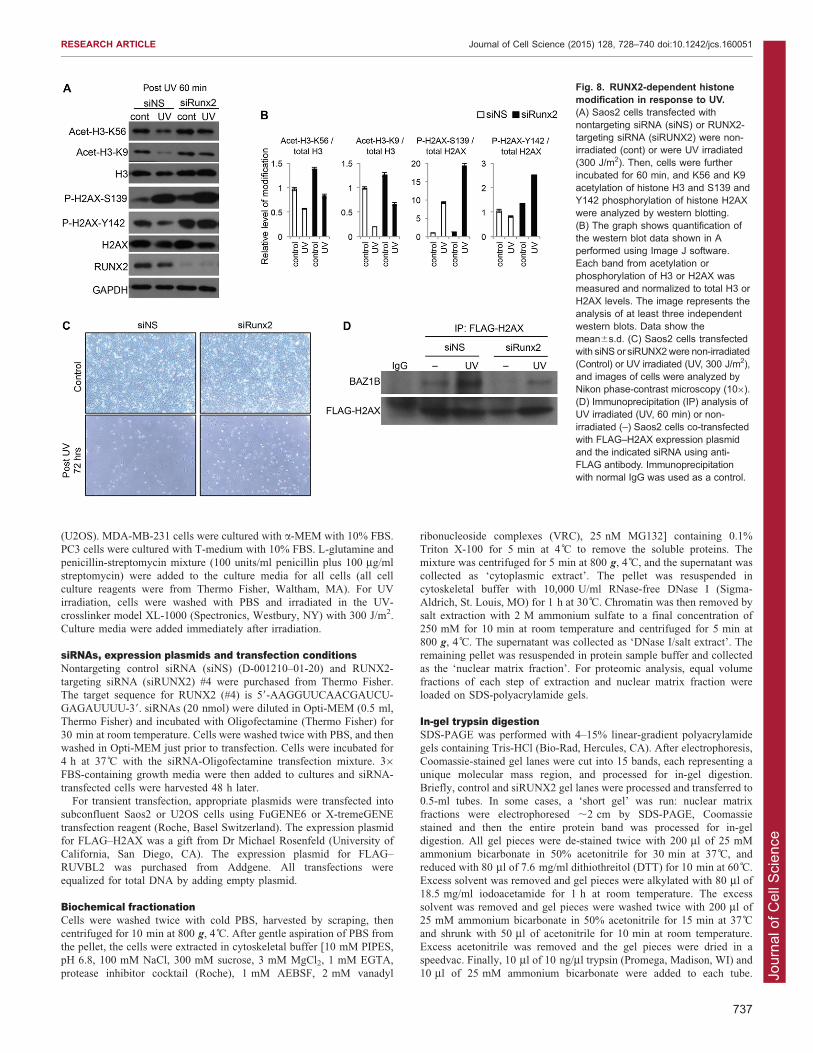
biological function of RUNX2 in the UV-induced DNA damageresponse. For example, other post-translational modifications ofhistones occur in addition to phosphorylation of H2AX inresponse to DNA damage, including decreased acetylation of
H3K9 and H3K56 by activation of HDACs or inhibition ofacetyltransferases (e.g. GCN5) (Yu et al., 2011). In osteosarcomacells transfected with nontargeting siRNA and exposed to UV
irradiation, acetylation of H3K9 and H3K56 decreased, whereasphosphorylation of H2AX S139 increased (Fig. 8A,B). H2AXY142 phosphorylation appeared to be slightly decreased when the
signal intensity was normalized relative to total H2AX recovery(Fig. 8B).
RUNX2 depletion by siRNA knockdown modestly attenuatedthe UV-dependent reductions in acetylation of both H3K9 and
H3K56. The results of RUNX2 knockdown on UV-dependentphosphorylation of H2AX were more dramatic. We saw anincrease in H2AX S139 phosphorylation after UV irradiation
and the abolishment of the UV-related reduction in Y142phosphorylation in cells transfected with RUNX2-specificsiRNA (Fig. 8A,B). The latter finding supports the concept that
RUNX2 modulates BAZ1B functions linked to H2AXphosphorylation and the response to UV irradiation. BecauseBAZ1B-mediated phosphorylation of Y142 in H2AX triggers
apoptosis (Xiao et al., 2009), we examined cell survival inresponse to UV. Cells with decreased levels of RUNX2 appearedto be protected from UV-induced cell loss compared tonontargeting-siRNA-transfected control cells (Fig. 8C). Finally,
immunoprecipitation analysis of Saos2 cells co-transfected withthe FLAG–H2AX expression construct and RUNX2-targeting ornontargeting siRNA, and treated with UV irradiation indicated
that the BAZ1B interaction with H2AX, which is normallyincreased by UV exposure, was actually diminished in the
Fig. 4. Interaction of RUNX2 with RUVBL2, INTS3 and BAZ1B in metastatic prostate PC3 or breast MDA-MB-231 cells. (A) Biochemical fractionation ofPC3 and MDA-MB-231 cells. Proteins from whole-cell lysates (‘W’), cytoplasmic extracts, (‘C’), DNase I/salt extracts (‘D’) and nuclear matrix fraction (‘N’) fromPC3 or MDA-MB-231 cells were resolved on 8% or 15% SDS-PAGE and analyzed by western blotting with the indicated primary antibodies. (B) Co-immunoprecipitation of RUNX2 with interacting proteins was analyzed by western blotting. To detect RUNX2–RUVBL2, RUNX2–INTS3 or RUNX2–BAZ1Bendogenous interactions, 5 mg of whole-cell lysates from Saos2 or U2OS cells were immunoprecipitated (IP) with 5 mg of anti-RUNX2 antibodies or 5 mg ofnormal rabbit IgG as a negative control. Proteins were separated by SDS-PAGE and blotted, and immunoblots were probed with anti-RUVBL2, anti-INTS3 oranti-BAZ1B antibodies. Antibodies against GAPDH, histone H3 and lamin B were used as markers for cytoplasmic extracts, DNase I/salt extracts and thenuclear matrix fraction, respectively.
RESEARCH ARTICLE Journal of Cell Science (2015) 128, 728–740 doi:10.1242/jcs.160051
733
Jour
nal o
f Cel
l Sci
ence
absence of RUNX2 (Fig. 8D). In conclusion, the combinedresults of this study are consistent with a model in which RUNX2
participates with BAZ1B and other associated proteins in cellsurvival in response to UV-induced DNA damage.
DISCUSSIONRUNX2 is a nuclear-matrix-associated transcription factor withbiological activities in normal development or cancer that are
determined by the dynamic association of interacting proteinsto form different functional complexes depending on thephysiological context. To understand this proteome of RUNX2-interacting proteins at the nuclear matrix, we employed a novel
approach that combines the depletion of RUNX2 as a molecularscaffolding protein and high-resolution proteomic analysis ofchanges in the protein profile of the nuclear-matrix-related
subnuclear fractions in cancer cell lines. Analysis of thissubnuclear proteome in control cells containing RUNX2
revealed that, as expected, the nuclear matrix contains a largenumber of well-established structural molecules (e.g. lamin A/C
and B, Numa, matrin-3) and many proteins involved in RNAmetabolism. These observations are entirely consistent withresults from a number of recent proteomic studies that
characterize nuclear matrix proteins from embryonic stem cells,tumor cell types and plant cells (Oehr, 2003; Calikowski et al.,2003; Barboro et al., 2009; Albrethsen et al., 2010; Nasrabadi
et al., 2010; Warters et al., 2010).The main finding of the current study is the identification of
three proteins that were shown to have shared RUNX2-dependentfunctions in UV-related DNA damage response. Interestingly, the
nuclear matrix has been reported to provide a platform for DNArepair (Mullenders et al., 1990; Zaalishvili et al., 2000; Atanassovet al., 2005). One protein identified by our proteomic analysis is
RUVBL2, a member of the AAA+ family (ATPase associatedwith diverse cellular activities) of DNA helicases (Izumi et al.,
Fig. 5. RUNX2 association withRUVBL2, INTS3 and BAZ1B in
vivo. (A) Immunofluorescent stainingof RUNX2 (Alexa Fluor 488, green)with RUVBL2, INTS3 or BAZ1B(Alexa Fluor 555, red) in Saos2 cellswas analyzed by confocalmicroscopy (z-sections were 0.2 mmthickness; images were acquiredusing a 63x objective with oilimmersion; 1.4 numerical aperture).Insets show differential interferencecontrast (DIC, lower right) and DAPIimages (upper right). (B) Thefunctional protein networks ofRUVBL2, INTS3 and BAZ1B wereanalyzed using STRING (version9.0). Boxes below the networkdiagrams show a simplified version ofthe model. (C) Reported biologicalfunctions of RUNX2, RUVBL2,INTS3 and BAZ1B, with references.
RESEARCH ARTICLE Journal of Cell Science (2015) 128, 728–740 doi:10.1242/jcs.160051
734
Jour
nal o
f Cel
l Sci
ence
2010). Human RUVBL2 is the apparent homolog of the bacterial
RUVB protein, which encodes a DNA helicase essential forhomologous recombination and DNA double-strand break repair.Recently, RUVBL2 has also been identified in chromatin
remodeling complexes including INO80, SRCAP, Uril andTip60 (Gorynia et al., 2011). The second protein, INTS3, wasinitially characterized as an RNA polymerase II C-terminaldomain binding factor involved in the 39 processing of small
nuclear RNAs (Inagaki et al., 2008). Recently, INTS3 has beenidentified as a key component of the DNA damage response(Huang et al., 2009; Skaar et al., 2009). BAZ1B, the third protein,
is a novel tyrosine protein kinase related to the bromodomainfamily. It contains a structural motif characteristic of proteins thatbind to acetylated lysines and are involved in the chromatin-
dependent regulation of transcription (Xiao et al., 2009). BAZ1Bphosphorylates H2AX at Tyr142, which plays a pivotal role inDNA repair and acts as a molecular marker that distinguishes
between apoptotic and repair responses to genotoxic stress. Thesimultaneous identification of these three proteins as prominentRUNX2-dependent nuclear-matrix proteins suggested that theymight share a RUNX2-related function.
RUVBL2, INTS3, BAZ1B and RUNX2 form a complex thatregulates DNA repair following UV-induced DNA damageIn these experiments, we found that the association ofendogenous RUNX2 with BAZ1B attenuated c-H2AX fociformation, and resulted in higher rates of cell death after UV
treatment than in RUNX2-depleted cells. The DNA-damage
marker c-H2AX relays signals from DNA lesions and recruits
DNA damage repair machinery as well as chromatin remodelers tomaintain genome stability. When DNA damage signal transductionis disrupted, however, cells fail to repair damaged DNA. We
propose a model in which RUNX2 binds to the c-H2AX complexwith BAZ1B in response to DNA damage and inhibits BAZ1B.Although cellular DNA damage responses mediated by BAZ1Bhave not been well characterized, the molecular function of
BAZ1B in response to DNA damage has been described (Xiaoet al., 2009). According to the authors, phosphorylation of H2AX atTyr142 by BAZ1B inhibits phosphorylation of H2AX at Ser139.
So, the balance between survival and apoptosis can be maintainedby two different but neighboring phosphorylation sites, Ser139 andTyr142, of H2AX. However, our data indicated that in spite of
increased phosphorylation at Tyr142, RUNX2-knockdown cellsdid not show higher levels of apoptosis as reported by others.Instead, these cells had higher survival rates after UV treatment
than cells transfected with nontargeting siRNA. We attribute thisresult to the fact that the Saos2 cells used in our study do notexpress p53. The tumor suppressor p53 plays a key role in the DNAdamage response, including arresting cells in G1 and G2 phases
(Decraene et al., 2001) and activating wild-type p53-inducedphosphatase 1 (WIP1, also known as protein phosphatase 1D), toreduce Ser139 phosphorylation of H2AX (Cha et al., 2010). For
future studies, it would be interesting to evaluate the effects oftreatments that induce double-strand DNA breaks, rather than therelatively benign single-strand breaks expected from the UV-
irradiation levels used in the experiments described here.
Fig. 6. UV effect on RUNX2 andinteracting proteins. Saos2 cells weretransfected with nontargeting (siNS) orRUNX2-targeting siRNA (siRUNX2), andwere UV irradiated (300 J/m2) or not treatedwith radiation (NT). (A,B) Confocalmicroscopy analysis of co-immunofluorescent staining. Non-irradiated(NT) cells and 30 or 60 min post-UV-irradiation cells were permeabilized, fixedand stained with RUNX2-specific antibodyand RUVBL2, INTS3, BAZ1B (A) or c-H2AX(B) antibodies. Blue arrows point toRUNX2-associated RUVBL2 or BAZ1Bfoci. The thickness of z-sections is 0.2 mm.Scale bar: 5 mm. (C) Western blot analysisof H2AX, c-H2AX, RUNX2, RUVBL2,INTS3, BAZ1B and GAPDH in whole-celllysates from non-irradiated and UV-irradiated Saos2 cells. UV-irradiated cellswere harvested at 5 or 30 min after UVirradiation (300 J/m2).
RESEARCH ARTICLE Journal of Cell Science (2015) 128, 728–740 doi:10.1242/jcs.160051
735
Jour
nal o
f Cel
l Sci
ence
Histone modification in the DNA damage response –histone acetylationEukaryotic cells protect their genome from UV-induced DNAdamage through a sequence of molecular processes that includedamage recognition, chromatin opening, DNA repair and chromatinsealing. Because chromatin is a multi-component complex of DNA
and proteins, and the arrangement of nucleosomes throughout thegenome is highly variable, this repair process has not beenexhaustively characterized. Recently, however, histone acetylation
was identified as a key player involved in opening chromatin toprovide access for the DNA repair machinery. Histone H3 and H4acetylation changes were observed after UV irradiation in yeast as
well as human cells (Yu et al., 2011). Cells can modulate histoneacetylation levels by recruiting either histone acetyltransferases(HATs) or histone deacetylases (HDACs), depending on thecellular context. H3K79 methylation is another important post-
translational modification involved in opening and sealing
chromatin following UV irradiation. DNA-damage-dependenthistone modification happens not only in the main nucleosome
components, but also in a variant histone H2AX withphosphorylation on S139 (c-H2AX).
In conclusion, we have defined RUNX2-dependent protein–protein interactions that are associated with the nuclear matrix
and discovered three proteins involved in the DNA damageresponse – BAZ1B, RUVBL2 and INTS3 – that require RUNX2for scaffolding into a multi-functional complex. This complex
supports histone displacement, DNA unwinding and stabilizationof single-stranded DNA to mount an integrated response to DNAdamage in breast, prostate and bone cancer cells.
MATERIALS AND METHODSCell culture and UV irradiationCells were grown at 37 C under 10% CO2. Saos2 and U2OS cells were
cultured with McCoy’s 5A medium with 15% FBS (Saos2) or 10% FBS
Fig. 7. RUNX2 association with c-H2AX byUV irradiation. Saos2 cells transfected withnontargeting siRNA (siNS) or RUNX2-targeting siRNA (siRUNX2) were not treatedwith radiation (NT) or were UV-irradiated(300 J/m2). (A) After UV irradiation, cells wereincubated for 30 or 60 min, thenpermeabilized, fixed and co-stained withRUNX2 (green) and c-H2AX (red) antibodiesand analyzed by confocal microscopy.(B) Cells showing c-H2AX foci in the nucleuswere counted. The graph shows the ratio ofthe number of cells with nuclear c-H2AX focito the total cells in the field; 3–14 cells werecounted per field, and at least 9 fields werecounted per treatment. (C) The number of c-H2AX foci per nucleus was counted andplotted; §36 cells were analyzed for eachdata point. The data show the mean6s.d.(D) Representative confocal images from30 min after UV irradiation are shown.(E) Saos2 cells were transfected with theFLAG–H2AX expression construct, UVirradiated or left untreated, then FLAG–H2AXprotein was immunoprecipitated (IP) andanalyzed by western blotting (WB). Becausethe intensity of ECL signal from Ser139phosphorylation on H2AX from input laneswas low, we included longer exposures of theblot for Ser139 phosphorylation (P-S139-H2AX) and total H2AX input lanes. The upperband from the P-S139-H2AX and H2AX blotscorresponds to overexpressed FLAG–H2AXand the lower band is from endogenousH2AX. *Long indicates data from a longerexposure of blots for ECL detection. Bandscorresponding to INTS3 and RUNX2 areindicated with red arrows.
RESEARCH ARTICLE Journal of Cell Science (2015) 128, 728–740 doi:10.1242/jcs.160051
736
Jour
nal o
f Cel
l Sci
ence
(U2OS). MDA-MB-231 cells were cultured with a-MEM with 10% FBS.
PC3 cells were cultured with T-medium with 10% FBS. L-glutamine and
penicillin-streptomycin mixture (100 units/ml penicillin plus 100 mg/ml
streptomycin) were added to the culture media for all cells (all cell
culture reagents were from Thermo Fisher, Waltham, MA). For UV
irradiation, cells were washed with PBS and irradiated in the UV-
crosslinker model XL-1000 (Spectronics, Westbury, NY) with 300 J/m2.
Culture media were added immediately after irradiation.
siRNAs, expression plasmids and transfection conditionsNontargeting control siRNA (siNS) (D-001210–01-20) and RUNX2-
targeting siRNA (siRUNX2) #4 were purchased from Thermo Fisher.
The target sequence for RUNX2 (#4) is 59-AAGGUUCAACGAUCU-
GAGAUUUU-39. siRNAs (20 nmol) were diluted in Opti-MEM (0.5 ml,
Thermo Fisher) and incubated with Oligofectamine (Thermo Fisher) for
30 min at room temperature. Cells were washed twice with PBS, and then
washed in Opti-MEM just prior to transfection. Cells were incubated for
4 h at 37 C with the siRNA-Oligofectamine transfection mixture. 36FBS-containing growth media were then added to cultures and siRNA-
transfected cells were harvested 48 h later.
For transient transfection, appropriate plasmids were transfected into
subconfluent Saos2 or U2OS cells using FuGENE6 or X-tremeGENE
transfection reagent (Roche, Basel Switzerland). The expression plasmid
for FLAG–H2AX was a gift from Dr Michael Rosenfeld (University of
California, San Diego, CA). The expression plasmid for FLAG–
RUVBL2 was purchased from Addgene. All transfections were
equalized for total DNA by adding empty plasmid.
Biochemical fractionationCells were washed twice with cold PBS, harvested by scraping, then
centrifuged for 10 min at 800 g, 4 C. After gentle aspiration of PBS from
the pellet, the cells were extracted in cytoskeletal buffer [10 mM PIPES,
pH 6.8, 100 mM NaCl, 300 mM sucrose, 3 mM MgCl2, 1 mM EGTA,
protease inhibitor cocktail (Roche), 1 mM AEBSF, 2 mM vanadyl
ribonucleoside complexes (VRC), 25 nM MG132] containing 0.1%
Triton X-100 for 5 min at 4 C to remove the soluble proteins. The
mixture was centrifuged for 5 min at 800 g, 4 C, and the supernatant was
collected as ‘cytoplasmic extract’. The pellet was resuspended in
cytoskeletal buffer with 10,000 U/ml RNase-free DNase I (Sigma-
Aldrich, St. Louis, MO) for 1 h at 30 C. Chromatin was then removed by
salt extraction with 2 M ammonium sulfate to a final concentration of
250 mM for 10 min at room temperature and centrifuged for 5 min at
800 g, 4 C. The supernatant was collected as ‘DNase I/salt extract’. The
remaining pellet was resuspended in protein sample buffer and collected
as the ‘nuclear matrix fraction’. For proteomic analysis, equal volume
fractions of each step of extraction and nuclear matrix fraction were
loaded on SDS-polyacrylamide gels.
In-gel trypsin digestionSDS-PAGE was performed with 4–15% linear-gradient polyacrylamide
gels containing Tris-HCl (Bio-Rad, Hercules, CA). After electrophoresis,
Coomassie-stained gel lanes were cut into 15 bands, each representing a
unique molecular mass region, and processed for in-gel digestion.
Briefly, control and siRUNX2 gel lanes were processed and transferred to
0.5-ml tubes. In some cases, a ‘short gel’ was run: nuclear matrix
fractions were electrophoresed ,2 cm by SDS-PAGE, Coomassie
stained and then the entire protein band was processed for in-gel
digestion. All gel pieces were de-stained twice with 200 ml of 25 mM
ammonium bicarbonate in 50% acetonitrile for 30 min at 37 C, and
reduced with 80 ml of 7.6 mg/ml dithiothreitol (DTT) for 10 min at 60 C.
Excess solvent was removed and gel pieces were alkylated with 80 ml of
18.5 mg/ml iodoacetamide for 1 h at room temperature. The excess
solvent was removed and gel pieces were washed twice with 200 ml of
25 mM ammonium bicarbonate in 50% acetonitrile for 15 min at 37 C
and shrunk with 50 ml of acetonitrile for 10 min at room temperature.
Excess acetonitrile was removed and the gel pieces were dried in a
speedvac. Finally, 10 ml of 10 ng/ml trypsin (Promega, Madison, WI) and
10 ml of 25 mM ammonium bicarbonate were added to each tube.
Fig. 8. RUNX2-dependent histonemodification in response to UV.(A) Saos2 cells transfected withnontargeting siRNA (siNS) or RUNX2-targeting siRNA (siRUNX2) were non-irradiated (cont) or were UV irradiated(300 J/m2). Then, cells were furtherincubated for 60 min, and K56 and K9acetylation of histone H3 and S139 andY142 phosphorylation of histone H2AXwere analyzed by western blotting.(B) The graph shows quantification ofthe western blot data shown in Aperformed using Image J software.Each band from acetylation orphosphorylation of H3 or H2AX wasmeasured and normalized to total H3 orH2AX levels. The image represents theanalysis of at least three independentwestern blots. Data show themean6s.d. (C) Saos2 cells transfectedwith siNS or siRUNX2were non-irradiated(Control) or UV irradiated (UV, 300 J/m2),and images of cells were analyzed byNikon phase-contrast microscopy (106).(D) Immunoprecipitation (IP) analysis ofUV irradiated (UV, 60 min) or non-irradiated (–) Saos2 cells co-transfectedwith FLAG–H2AX expression plasmidand the indicated siRNA using anti-FLAG antibody. Immunoprecipitationwith normal IgG was used as a control.
RESEARCH ARTICLE Journal of Cell Science (2015) 128, 728–740 doi:10.1242/jcs.160051
737

Jour
nal o
f Cel
l Sci
ence
Additional 25 mM ammonium bicarbonate was added until the gel pieces
were fully swollen (,10–50 ml). Samples were then incubated for 4 h at
37 C and peptides were recovered by collecting excess solvent. Gel
pieces were extracted twice more with 50 ml of 50% acetonitrile/5%
formic acid, vortexing, centrifuging and incubating for 15 min. Solvent
was removed and combined with the previously recovered solvent, for a
total of three extractions. Combined extracts were dried on a speedvac.
Nanoflow LC-MS/MS and database searchSamples were reconstituted in 20 ml of 2% acetonitrile, 0.1% (v/v) formic
acid, 0.01% (v/v) trifluoroacetic acid in water and analyzed using two
different instrument platforms. Peptide samples from one full set of SDS
gel pieces were injected (5 ml) using a NanoAcquity (Waters
Corporation, Milford, MA) UPLC and loaded onto a Waters Symmetry
C18 (180 mm i.d.62 cm) trapping column at a flow rate of 5 ml/min for
3 min. Peptides were then separated by in-line gradient elution using a
75 mm i.d.610 cm Waters BEH 130 (1.7 mm) analytical column, at a
flow rate of 300 nl/min using a linear gradient from 3 to 90% B over
95 min (mobile phase A – 2% acetonitrile, 0.1% (v/v) formic acid, 0.01%
(v/v) trifluoroacetic acid; mobile phase B – 98% acetonitrile, 0.1%
formic acid, 0.01% trifluoroacetic acid). Peptides were eluted into an
LTQ (Thermo Scientific) linear ion trap mass spectrometer operating in
positive-ion electrospray and data-dependent acquisition modes. One
mass spectrum was acquired over the m/z 400–2000 range, followed by
serial tandem mass spectrometry (i.e. MS/MS) on the seven most
abundant signals. Precursor ion isolation width was 2.0 Da, collision
energy was 35%, ion population targets were 10,000 for MS and 5000 for
MS/MS, and maximum ion fill times were 200 ms for both acquisition
types. Precursor ions analyzed were subjected to dynamic exclusion for
30 s using a window of 20.5 to +1.5 m/z. The repeat count was 1 with a
30 s delay; ions at m/z 371 and 445 were also excluded from MS/MS.
Another set of similarly prepared samples were analyzed using a
Proxeon Easy nanoLC (Thermo Scientific) system directly configured to
an LTQ-Orbitrap Velos (Thermo Scientific) hybrid mass spectrometer.
Peptide samples (2 ml) were loaded at 4 ml/min for 7 min onto a custom-
made trap column (100 mm i.d. fused silica with Kasil frit) containing
2 cm of 200-A, 5-mm Magic C18AQ particles (Michrom Bioresources,
Auburn, CA). Peptides were then eluted using a custom-made analytical
column (75 mm i.d. fused silica) with a gravity-pulled tip and packed
with 25 cm of 100-A, 5-mm Magic C18AQ particles (Michrom). Peptides
were eluted with a linear gradient as described above. Mass spectrometry
data were acquired using a data-dependent acquisition routine of
acquiring one mass spectrum from m/z 350–2000 in the Orbitrap
(resolution, 60,000; ion population, 1.06106; maximum ion injection
time, 500 ms) followed by tandem mass spectrometry in the linear ion
trap (LTQ) of the ten most abundant precursor ions observed in the mass
spectrum. MS/MS data were acquired using a precursor isolation width of
2.0 Da, a collision energy of 35%, an ion population of 5000 and a
maximum ion fill time of 50 ms. Charge-state rejection of singly charged
ions and dynamic exclusion was utilized [20.1 to +1.1 Da window,
repeat count 1 (30-s delay)] to minimize data redundancy and maximize
peptide identification.
The raw data files were processed using Extract MSN software
(Thermo Scientific) and searched against the human index of the
SwissProt database (version 09/24/11) with Mascot (version 2.3.02;
Matrix Science, London, UK) and X! Tandem [The GPM (www.thegpm.
org); version Cyclone (2010.12.01.1)] software packages. LTQ Orbitrap
Velos data were searched using a parent mass tolerance of 15 ppm and a
fragment mass tolerance of 0.5 Da. LTQ data utilized a parent tolerance
of 1.2 Da and a fragment tolerance of 1.0 Da. Full tryptic specificity
with two missed cleavages was considered, and variable modifications
of acetylation (protein N-term), cyclization of N-terminal S-
carbamoylmethylcysteine (peptide N-term) and oxidation (methionine)
and fixed modification of carbamidomethylation (cysteine) were
considered. All search results were loaded into Scaffold software
(Version 3.3.1; Proteome Software, Portland, OR) for comparative
analyses using spectral counting of tandem mass spectra and full
annotation of the data (Searle, 2010). Peptide identifications were
accepted if they could be established at .95% probability by the Peptide
Prophet algorithm (Keller et al., 2002) following Scaffold delta-mass
correction. Protein identifications were accepted if they could be
established at .99% probability and contained at least two identified
peptides; protein probabilities were assigned by the Protein Prophet
algorithm (Nesvizhskii et al., 2003). Normalized spectral counts were
calculated by dividing the spectral counts for an identified protein by the
sum of the spectral counts per sample.
Quantitative gene expression analysisRNA was isolated using Trizol reagent (Thermo Fisher Scientific) and
trace DNA was removed using the DNA-free RNA kit (Zymo Research,
Irvine, CA). cDNA was synthesized using Superscript III (Thermo Fisher
Scientific) and amplified using gene-specific primers (supplementary
material Table S2) and iTAQ SYBR green supermix (Bio-Rad
Laboratories). Reactions were run and data collected on an ABI
PRISM 700 system (Thermo Fisher Scientific). Primers for PCR are
displayed in supplementary material Table S1.
ImmunoprecipitationConfluent cells on 100-mm plates were harvested and solubilized in lysis
buffer (10 mM Tris-HCl pH 8.0, 100 mM NaCl, 1 mM EDTA, 0.1%
Nonidet P40, 1 mM DTT and 1 mM PMSF). Insoluble material was
removed by centrifugation. The supernatants were incubated for 12 h at
4 C with 2 mg of anti-RUNX2 (M70 or S19; Santa Cruz Biotechnology,
Santa Cruz, CA), anti-FLAG (M2; Sigma-Aldrich) and a mixture of anti-
rabbit- and anti-goat-IgG (Santa Cruz Biotechnology). Following the
addition of 30 ml of Protein-A/G–agarose beads (Santa Cruz
Biotechnology), mixtures were incubated for 2 h at 4 C with rotation.
Immune complexes were washed three times with lysis buffer; the
agarose beads were then boiled for 10 min in sample buffer.
Immunoprecipitates were run on 8% acrylamide SDS-PAGE or 4–20%
gradient precast gels (Bio-Rad), followed by Western blot analysis with
antibodies against RUVBL2, INTS3, BAZ1B, H2AX or phosphorylated
Ser139-H2AX.
GST pulldown assaysThe proteins containing N-terminal glutathione S-transferase (GST)
fused in-frame to the RUNX homologs were obtained by expression in
Escherichia coli BL21 strain as reported previously (Pande et al., 2013).
A total of 1 mg of GST alone, GST-fused N-terminal RHD domain or C-
terminal RUNX2 proteins was incubated with Glutathione Sepharose 4B
(GE Healthcare) beads in 500 ml of binding buffer (20 mM Tris-HCl
pH 8.0, 100 mM KCl, 0.5% NP-40, 10 mM EDTA, 0.05 mM PMSF,
1 mM DTT) for 30 min at 4 C with rotation. The beads were washed four
times with 500 ml of binding buffer for 5 min at 4 C and incubated with
1 mg of whole-cell lysates from confluent Saos2 cells for 12 h at 4 C
with rotation. After four washes with binding buffer, each for 5 min at
4 C, the beads were resuspended in 50 ml of sample buffer and boiled for
10 min at 95 C. The proteins that were retained with the beads were
separated by 8% acrylamide SDS-PAGE and analyzed by western
blotting with specific antibodies against RUVBL2 (Santa Cruz
Biotechnology), INTS3 (Abcam, Cambridge, UK) and BAZ1B (Abcam).
Confocal microscopyTo prepare cells for immunofluorescence confocal microscopy analysis,
Saos2 cells were washed in Hanks balanced salt solution (Thermo Fisher
Scientific), and were incubated in 0.5% Triton X-100 in cytoskeletal
buffer for 10 min. This step removes soluble proteins from both the
cytoplasm and nucleus. Cells were later fixed in 4% formaldehyde in
cytoskeletal buffer for 50 min on ice. For antibody staining, cells were
stained with antibodies as described previously (Wagner et al., 2003).
The primary antibodies were as follows: rabbit polyclonal antibody
against RUNX2 (M70; 1:100; Santa Cruz Biotechnology); mouse
monoclonal antibody against RUVBL2 (B9; 1:40; Santa Cruz
Biotechnology); mouse monoclonal antibody against BAZ1B (1:40;
Santa Cruz Biotechnology); goat polyclonal antibody against INTS3
(1:500; Abcam); mouse monoclonal antibody against phosphorylated
RESEARCH ARTICLE Journal of Cell Science (2015) 128, 728–740 doi:10.1242/jcs.160051
738

Jour
nal o
f Cel
l Sci
ence
S139 of H2AX (1:100; Millipore). All antibodies were incubated
overnight at 4 C. The following secondary antibodies (Thermo Fisher
Scientific) were used: Alexa-Fluor-488-conjugated goat anti-rabbit-IgG
(1:2000); Alexa-Fluor-568-conjugated goat anti-mouse-IgG (1:2000);
Alexa-Fluor-594-conjugated donkey anti-goat-IgG (1:2000); Alexa-
Fluor-488-conjugated donkey anti-rabbit-IgG (1:2000). Coverslips were
mounted with ProlongGold (Thermo Fisher Scientific). Images were
collected using a Leica TCS SP5 confocal microscope using a 663 oil
lens (numerical aperture 1.4) at optimum zoom, fixed pinhole size
(100 mm) and optimum Z-plane interval (from 0.2–0.3-mm Z-stack),
depending on the sample. The 405-nm, 488-nm and 561-nm excitation
lasers were used to excite DAPI, Alexa Fluor 488, Alexa Fluor 568 and
Alexa Fluor 594, respectively, and were activated sequentially during
image collection. Images were analyzed using ImageJ and Photoshop 5.0.
AcknowledgementsWe thank the members of our laboratory, especially Jason Dobson and ShirwinPockwinse, for stimulating discussions and/or general support.
Competing interestsThe authors declare no competing or financial interests.
Author contributionsS.Y., J.A.N., A.N.I., J.B.L., J.L.S., A.J.v.W. and G.S.S. planned and designed thestudy. S.Y. performed the experiments, with help from A.J.C.Q. and J.A.N. forconfocal microscopy, and K.M.G. and S.A.S. for mass spectrometry. Data wereanalyzed and interpreted by S.Y., K.M.G., S.A.S., A.J.C.Q., J.A.N., A.N.I., J.B.L.,J.L.S., A.J.v.W. and G.S.S. S.Y. drafted the manuscript, with help from J.A.N.,A.N.I., J.B.L., J.L.S., A.J.v.W., L.A.M.-B., S.A.S. and G.S.S. All authors read andapproved the final manuscript.
FundingThis work was supported, in whole or in part, by the National Institutes of Health[grant numbers P01 CA082834, P01 AR048818, R01 AR049069 and R01AR039588]. The contents of this manuscript are solely the responsibility of theauthors and do not necessarily represent the official views of the National Centerfor Research Resources or the National Institutes of Health. Deposited in PMC forrelease after 12 months.
Supplementary materialSupplementary material available online athttp://jcs.biologists.org/lookup/suppl/doi:10.1242/jcs.160051/-/DC1
ReferencesAlbrethsen, J., Knol, J. C., Piersma, S. R., Pham, T. V., de Wit, M., Mongera, S.,Carvalho, B., Verheul, H. M. W., Fijneman, R. J. A., Meijer, G. A. et al. (2010).Subnuclear proteomics in colorectal cancer: identification of proteins enriched inthe nuclear matrix fraction and regulation in adenoma to carcinoma progression.Mol. Cell Proteomics 9, 988-1005.
Atanassov, B., Gospodinov, A., Stoimenov, I., Mladenov, E., Russev, G.,Tsaneva, I. and Anachkova, B. (2005). Repair of DNA interstrand crosslinksmay take place at the nuclear matrix. J. Cell. Biochem. 96, 126-136.
Barboro, P., D’Arrigo, C., Repaci, E., Bagnasco, L., Orecchia, P., Carnemolla,B., Patrone, E. and Balbi, C. (2009). Proteomic analysis of the nuclear matrix inthe early stages of rat liver carcinogenesis: identification of differentiallyexpressed and MAR-binding proteins. Exp. Cell Res. 315, 226-239.
Barnes, L., Dumas, M., Juan, M., Noblesse, E., Tesniere, A., Schnebert, S.,Guillot, B. and Moles, J. P. (2010). GammaH2AX, an accurate marker thatanalyzes UV genotoxic effects on human keratinocytes and on human skin.Photochem. Photobiol. 86, 933-941.
Barnett, C. and Krebs, J. E. (2011). WSTF does it all: a multifunctional protein intranscription, repair, and replication. Biochem. Cell Biol. 89, 12-23.
Berezney, R., Mortillaro, M. J., Ma, H., Wei, X. and Samarabandu, J. (1995).The nuclear matrix: a structural milieu for genomic function. Int. Rev. Cytol.162A, 1-65.
Bidwell, J. P., Torrungruang, K., Alvarez, M., Rhodes, S. J., Shah, R., Jones,D. R., Charoonpatrapong, K., Hock, J. M. and Watt, A. J. (2011). Involvementof the nuclear matrix in the control of skeletal genes: the NMP1 (YY1), NMP2(Cbfa1), and NMP4 (Nmp4/CIZ) transcription factors. Crit. Rev. Eukaryot. GeneExpr. 11, 279-297.
Bidwell, J. P., Van Wijnen, A. J., Fey, E. G., Dworetzky, S., Penman, S., Stein,J. L., Lian, J. B. and Stein, G. S. (1993). Osteocalcin gene promoter-bindingfactors are tissue-specific nuclear matrix components. Proc. Natl. Acad. Sci.USA 90, 3162-3166.
Calikowski, T. T., Meulia, T. and Meier, I. (2003). A proteomic study of thearabidopsis nuclear matrix. J. Cell. Biochem. 90, 361-378.
Cha, H., Lowe, J. M., Li, H., Lee, J. S., Belova, G. I., Bulavin, D. V. and Fornace,A. J., Jr (2010). Wip1 directly dephosphorylates gamma-H2AX and attenuatesthe DNA damage response. Cancer Res. 70, 4112-4122.
Decraene, D., Agostinis, P., Pupe, A., de Haes, P. and Garmyn, M. (2001).Acute response of human skin to solar radiation: regulation and function of thep53 protein. J. Photochem. Photobiol. B 63, 78-83.
Delcuve, G. P., Rastegar, M. and Davie, J. R. (2009). Epigenetic control. J. Cell.Physiol. 219, 243-250.
Dworetzky, S. I., Fey, E. G., Penman, S., Lian, J. B., Stein, J. L. and Stein, G. S.(1990). Progressive changes in the protein composition of the nuclear matrixduring rat osteoblast differentiation. Proc. Natl. Acad. Sci. USA 87, 4605-4609.
Dzieciatkowska, M., Hill, R. and Hansen, K. C. (2014). GeLC-MS/MS analysis ofcomplex protein mixtures. Methods Mol. Biol. 1156, 53-66.
Fey, E. G. and Penman, S. (1988). Nuclear matrix proteins reflect cell type oforigin in cultured human cells. Proc. Natl. Acad. Sci. USA 85, 121-125.
Getzenberg, R. H., Pienta, K. J., Huang, E. Y. and Coffey, D. S. (1991).Identification of nuclear matrix proteins in the cancer and normal rat prostate.Cancer Res. 51, 6514-6520.
Gorynia, S., Bandeiras, T. M., Pinho, F. G., McVey, C. E., Vonrhein, C., Round,A., Svergun, D. I., Donner, P., Matias, P. M. and Carrondo, M. A. (2011).Structural and functional insights into a dodecameric molecular machine – theRuvBL1/RuvBL2 complex. J. Struct. Biol. 176, 279-291.
Huang, J., Gong, Z., Ghosal, G. and Chen, J. (2009). SOSS complexesparticipate in the maintenance of genomic stability. Mol. Cell 35, 384-393.
Inagaki, Y., Yasui, K., Endo, M., Nakajima, T., Zen, K., Tsuji, K., Minami, M.,Tanaka, S., Taniwaki, M., Itoh, Y. et al. (2008). CREB3L4, INTS3, and SNAPAPare targets for the 1q21 amplicon frequently detected in hepatocellularcarcinoma. Cancer Genet. Cytogenet. 180, 30-36.
Izumi, N., Yamashita, A., Iwamatsu, A., Kurata, R., Nakamura, H., Saari, B.,Hirano, H., Anderson, P. and Ohno, S. (2010). AAA+ proteins RUVBL1 andRUVBL2 coordinate PIKK activity and function in nonsense-mediated mRNAdecay. Sci. Signal. 3, ra27.
Javed, A., Barnes, G. L., Pratap, J., Antkowiak, T., Gerstenfeld, L. C., vanWijnen, A. J., Stein, J. L., Lian, J. B. and Stein, G. S. (2005). Impairedintranuclear trafficking of Runx2 (AML3/CBFA1) transcription factors in breastcancer cells inhibits osteolysis in vivo. Proc. Natl. Acad. Sci. USA 102, 1454-1459.
Jha, S., Dutta, A. (2009). RVB1/RVB2: running rings around molecular biology.Mol. Cell 34, 521-33.
Kashiwaba, S., Kitahashi, K., Watanabe, T., Onoda, F., Ohtsu, M., Murakami,Y. (2010). The mammalian INO80 complex is recruited to DNA damage sites inan ARP8 dependent manner. Biochem. Biophys. Res. Commun. 402, 619-25.
Keller, A., Nesvizhskii, A. I., Kolker, E. and Aebersold, R. (2002). Empiricalstatistical model to estimate the accuracy of peptide identifications made by MS/MS and database search. Anal. Chem. 74, 5383-5392.
Kubben, N., Voncken, J. W. and Misteli, T. (2010). Mapping of protein- andchromatin-interactions at the nuclear lamina. Nucleus 1, 460-471.
Kuo, Y. H., Zaidi, S. K., Gornostaeva, S., Komori, T., Stein, G. S. and Castilla,L. H. (2009). Runx2 induces acute myeloid leukemia in cooperation withCbfbeta-SMMHC in mice. Blood 113, 3323-3332.
Lian, J. B., Javed, A., Zaidi, S. K., Lengner, C., Montecino, M., van Wijnen,A. J., Stein, J. L. and Stein, G. S. (2004). Regulatory controls for osteoblastgrowth and differentiation: role of Runx/Cbfa/AML factors. Crit. Rev. Eukaryot.Gene Expr. 14, 1-42.
Lian, J. B., Stein, G. S., Javed, A., van Wijnen, A. J., Stein, J. L., Montecino,M., Hassan, M. Q., Gaur, T., Lengner, C. J. and Young, D. W. (2006). Networksand hubs for the transcriptional control of osteoblastogenesis. Rev. Endocr.Metab. Disord. 7, 1-16.
Long, F. (2012). Building strong bones: molecular regulation of the osteoblastlineage. Nat. Rev. Mol. Cell Biol. 13, 27-38.
Lundgren, D. H., Hwang, S. I., Wu, L. and Han, D. K. (2010). Role of spectralcounting in quantitative proteomics. Expert Rev. Proteomics 7, 39-53.
Markaki, Y., Gunkel, M., Schermelleh, L., Beichmanis, S., Neumann, J.,Heidemann, M., Leonhardt, H., Eick, D., Cremer, C. and Cremer, T. (2010).Functional nuclear organization of transcription and DNA replication: atopographical marriage between chromatin domains and the interchromatincompartment. Cold Spring Harb. Symp. Quant. Biol. 75, 475-492.
Meldi, L. and Brickner, J. H. (2011). Compartmentalization of the nucleus. TrendsCell Biol. 21, 701-708.
Merriman, H. L., van Wijnen, A. J., Hiebert, S., Bidwell, J. P., Fey, E., Lian, J.,Stein, J. and Stein, G. S. (1995). The tissue-specific nuclear matrix protein,NMP-2, is a member of the AML/CBF/PEBP2/runt domain transcription factorfamily: interactions with the osteocalcin gene promoter. Biochemistry 34, 13125-13132.
Mullenders, L. H., Venema, J., van Hoffen, A., Mayne, L. V., Natarajan, A. T.and van Zeeland, A. A. (1990). The role of the nuclear matrix in DNA repair.Prog. Clin. Biol. Res. 340A, 223-232.
Nasrabadi, D., Larijani, M. R., Fathi, A., Gourabi, H., Dizaj, A. V., Baharvand, H.and Salekdeh, G. H. (2010). Nuclear proteome analysis of monkey embryonicstem cells during differentiation. Stem Cell Rev. 6, 50-61.
Nesvizhskii, A. I., Keller, A., Kolker, E. and Aebersold, R. (2003). A statisticalmodel for identifying proteins by tandem mass spectrometry. Anal. Chem. 75,4646-4658.
RESEARCH ARTICLE Journal of Cell Science (2015) 128, 728–740 doi:10.1242/jcs.160051
739

Jour
nal o
f Cel
l Sci
ence
Nickerson, J. A., Krockmalnic, G., Wan, K. M. and Penman, S. (1997). Thenuclear matrix revealed by eluting chromatin from a cross-linked nucleus. Proc.Natl. Acad. Sci. USA 94, 4446-4450.
Oehr, P. (2003). Proteomics as a tool for detection of nuclear matrix proteins andnew biomarkers for screening of early tumors stage. Anticancer Res. 23, 805-812.
Pande, S., Browne, G., Padmanabhan, S., Zaidi, S. K., Lian, J. B., van Wijnen,A. J., Stein, J. L. and Stein, G. S. (2013). Oncogenic cooperation betweenPI3K/Akt signaling and transcription factor Runx2 promotes the invasiveproperties of metastatic breast cancer cells. J. Cell. Physiol. 228, 1784-1792.
Pratap, J., Lian, J. B. and Stein, G. S. (2011). Metastatic bone disease: role oftranscription factors and future targets. Bone 48, 30-36.
Ruthenburg, A. J., Graybosch, D. M., Huetsch, J. C., Verdine, G. L. (2005). Asuperhelical spiral in the Escherichia coli DNA gyrase A C-terminal domainimparts unidirectional supercoiling bias. J. Biol. Chem. 280, 26177-26184.
Searle, B. C. (2010). Scaffold: a bioinformatic tool for validating MS/MS-basedproteomic studies. Proteomics 10, 1265-1269.
Simon, D. N. and Wilson, K. L. (2011). The nucleoskeleton as a genome-associated dynamic ‘network of networks’. Nat. Rev. Mol. Cell Biol. 12, 695-708.
Skaar, J. R., Richard, D. J., Saraf, A., Toschi, A., Bolderson, E., Florens, L.,Washburn, M. P., Khanna, K. K. and Pagano, M. (2009). INTS3 controls thehSSB1-mediated DNA damage response. J. Cell Biol. 187, 25-32.
Stenoien, D., Sharp, Z. D., Smith, C. L. and Mancini, M. A. (1998). Functionalsubnuclear partitioning of transcription factors. J. Cell. Biochem. 70, 213-221.
van Driel, R., Wansink, D. G., van Steensel, B., Grande, M. A., Schul, W. andde Jong, L. (1995). Nuclear domains and the nuclear matrix. Int. Rev. Cytol.162A, 151-189.
Wagner, S., Chiosea, S. and Nickerson, J. A. (2003). The spatial targeting andnuclear matrix binding domains of SRm160. Proc. Natl. Acad. Sci. USA 100,3269-3274.
Warters, R. L., Cassidy, P. B., Sunseri, J. A., Parsawar, K., Zhuplatov, S. B.,Kramer, G. F. and Leachman, S. A. (2010). The nuclear matrix shell proteomeof human epidermis. J. Dermatol. Sci. 58, 113-122.
Xiao, A., Li, H., Shechter, D., Ahn, S. H., Fabrizio, L. A., Erdjument-Bromage,H., Ishibe-Murakami, S., Wang, B., Tempst, P., Hofmann, K. et al. (2009).WSTF regulates the H2A.X DNA damage response via a novel tyrosine kinaseactivity. Nature 457, 57-62.
Yu, S., Teng, Y., Waters, R. and Reed, S. H. (2011). How chromatin is remodelledduring DNA repair of UV-induced DNA damage in Saccharomyces cerevisiae.PLoS Genet. 7, e1002124.
Zaalishvili, T. M., Gabriadze, I. Y., Margiani, D. O., Philauri, V. R. andSurguladze, N. M. (2000). Participation of Poly(ADP-ribose)-polymerase ofnuclear matrix in DNA repair. Biokhimiia 65, 659-661.
Zaidi, S. K., Javed, A., Choi, J. Y., van Wijnen, A. J., Stein, J. L., Lian, J. B. andStein, G. S. (2001). A specific targeting signal directs Runx2/Cbfa1 tosubnuclear domains and contributes to transactivation of the osteocalcingene. J. Cell Sci. 114, 3093-3102.
Zaidi, S. K., Sullivan, A. J., van Wijnen, A. J., Stein, J. L., Stein, G. S. and Lian,J. B. (2002). Integration of Runx and Smad regulatory signals at transcriptionallyactive subnuclear sites. Proc. Natl. Acad. Sci. USA 99, 8048-8053.
Zaidi, S. K., Sullivan, A. J., Medina, R., Ito, Y., van Wijnen, A. J., Stein, J. L.,Lian, J. B. and Stein, G. S. (2004). Tyrosine phosphorylation controlsRunx2-mediated subnuclear targeting of YAP to repress transcription. EMBOJ. 23, 790-799.
Zaidi, S. K., Javed, A., Pratap, J., Schroeder, T. M., J Westendorf, J., Lian, J. B.,vanWijnen, A. J., Stein, G. S. and Stein, J. L. (2006). Alterations in intranuclearlocalization of Runx2 affect biological activity. J. Cell. Physiol. 209, 935-942.
Zaidi, S. K., Young, D. W., Javed, A., Pratap, J., Montecino, M., van Wijnen, A.,Lian, J. B., Stein, J. L. and Stein, G. S. (2007a). Nuclear microenvironments inbiological control and cancer. Nat. Rev. Cancer 7, 454-463.
Zaidi, S. K., Pande, S., Pratap, J., Gaur, T., Grigoriu, S., Ali, S. A., Stein, J. L.,Lian, J. B., van Wijnen, A. J. and Stein, G. S. (2007b). Runx2 deficiency anddefective subnuclear targeting bypass senescence to promote immortalizationand tumorigenic potential. Proc. Natl. Acad. Sci. USA 104, 19861-19866.
Zeng, C., van Wijnen, A. J., Stein, J. L., Meyers, S., Sun, W., Shopland, L.,Lawrence, J. B., Penman, S., Lian, J. B., Stein, G. S. et al. (1997).Identification of a nuclear matrix targeting signal in the leukemia and bone-related AML/CBF-alpha transcription factors. Proc. Natl. Acad. Sci. USA 94,6746-6751.
Zeng, C., McNeil, S., Pockwinse, S., Nickerson, J., Shopland, L., Lawrence,J. B., Penman, S., Hiebert, S., Lian, J. B., van Wijnen, A. J. et al. (1998).Intranuclear targeting of AML/CBFalpha regulatory factors to nuclear matrix-associated transcriptional domains. Proc. Natl. Acad. Sci. USA 95, 1585-1589.
Zink, D., Fischer, A. H. and Nickerson, J. A. (2004). Nuclear structure in cancercells. Nat. Rev. Cancer 4, 677-687.
Zybailov, B., Coleman, M. K., Florens, L. and Washburn, M. P. (2005).Correlation of relative abundance ratios derived from peptide ionchromatograms and spectrum counting for quantitative proteomic analysisusing stable isotope labeling. Anal. Chem. 77, 6218-6224.
RESEARCH ARTICLE Journal of Cell Science (2015) 128, 728–740 doi:10.1242/jcs.160051
740





























