Embed Size (px)
Citation preview

FINE-STRUCTURE CHANGES IN ACHIALASIAOF THE ESOPHAGUS
I. VAGUS NERVES
ROBERT R. CASSELLA, M.D.*; F. HENRY ELLIS, JR., M.D.,AND ARNOLD L. BROWN, JR., M.D.
From the Section of Surgery and Section of Experimental andAnatomic Pathology, Mayo Clinic and Mayo Foundation, Rochester, Minn.
Achalasia of the esophagus is generally regarded as a disease of neuro-muscular origin although its precise cause is unknown. Some of thepathologic studies done in the early part of this century revealed thepresence of significant lesions in the extra-esophageal vagus nerves."qSubsequent studies failed to confirm these findings.4'5 Experimentallyhowever, it has been shown in the laboratory animal that certain proce-dures which interrupt the extra-esophageal vagus nerves produce a con-dition which simulates human achalasia, although incompletely.-8An electron microscopic study of the extra-esophageal vagus nerves
in achalasia was made in an attempt to demonstrate lesions of thesenerves in the region of the esophageal plexus in a group of patients withthis disease.
MATERIAL AND METHODSSmall vagus nerve branches to the lower esophageal plexus were biopsied at the
time of operation, and these provided the specimens for this study. Tissue wasobtained during modified Heller procedures 9 in 9 patients with esophageal achalasiaconfirmed by motility studies. Control specimens of vagus nerve were obtained froma comparable level of the esophagus from 8 patients during thoracotomy for thetreatment of carcinoma of the lung, carcinoma of the esophagus or duodenal ulcer(transthoracic vagotomy). All material was immediately fixed in Dalton's solution,10dehydrated in graded alcohol solutions, embedded in both butylmethyl methacrylate(4: i) and Epon 8I 2 11 and stained with uranyl acetate.12 After mounting on coppergrids, sections were examined with an RCA EMU-3E electron microscope.
RESULTS
Normal Esophageal Vagus Nerves. Normal vagus nerve at the levelof the thoracic esophagus was composed for the most part of non-
This investigation was supported in part by Research Grant AM-o5457 from theNational Institutes of Health, Public Health Service.
Abridgement of portion of thesis submitted by Dr. Cassella to the Faculty of theGraduate School of the University of Minnesota in partial fulfillment of the requirementsfor the degree of Doctor of Philosophy in Surgery.
Accepted for publication, August 24, I964.*Fellow in Surgery, Mayo Foundation, Rochester, Minn.
279

CASSELLA, ELLIS AND BROWN
myelinated axons (Fig. i a). Groups of axons were surrounded by long,slender processes of the perineural sheath cells. Within the perineurium,axons were orientated longitudinally, and individual axons were sur-rounded by a layer of cytoplasm originating from the endoneuralSchwann cells.18
Individual nonmyelinated axons ranged from 0.4 to 1.5 & in trans-verse diameter (Fig. i b). The centrally situated axoplasm in both non-myelinated and myelinated axons (Figs. i b and c) contained thin,branching, longitudinally coursing neurofilaments, small clear vesicles,mitochondria and small osmiophilic granules. The axolemma, or limitingaxoplasmic membrane, was usually indistinct. A double membrane, theaxon-Schwann membrane, separated the axoplasm from the surround-ing, darker Schwann cytoplasm. A similar morphologic structure, themesaxon, connected the axon to the surface of the Schwann cell.
Myelinated axons ranged from 1.5 to 6.6 ,u in transverse diameter(Figs. i b, c, and d). They featured the same morphologic componentsas nonmyelinated axons except for conspicuous thick, electron-densemyelin rings.Schwann cells (Fig. i d) were distinguished from fibrocytic and
endothelial cells by the presence of axons within their cytoplasm.13Their nuclei were round or ovoid without distinct nucleoli. Within thecytoplasm of Schwann cells were tiny, clumped, homogeneous granules,small, rounded, slightly osmiophilic vesicles and mitochondria. The over-all granularity of the Schwann cells in control material, however, wasonly slight or moderate. The interstitial space (Figs. i b and d) con-tained collagen, blood vessels and amorphous material.
Vagal Esophageal Nerves in Achalasia. In specimens from all 9 pa-tients with achalasia, some morphologic abnormality of both myelinatedand nonmyelinated axons and of Schwann cells was seen. Specimensfrom individual patients differed only in the frequency with which theseabnormalities were demonstrated.
Nonmyelinated axons appeared distorted in several ways. Neurofila-ments were fragmented and dispersed in some axons (Fig. 2 a) whichappeared swollen. They were condensed in others in which the axoniclumen was contracted. Mitochondria were increased in number in someaxons and appeared swollen or darkened and had an indistinct innerstructure (Fig. 2 b). Axon-Schwann membranes and mesaxons wereoften discontinuous. The usually distinct axoplasm became indistinct,the picture suggesting a confluence of axoplasm and Schwann cell cyto-plasm. Uncommonly, nonmyelinated axons were seen without mesaxonsor with short mesaxons (Fig. 2 C).
Myelinated axons had similar axoplasmic changes (Fig. 3 a) and
280 Vol. 46, No. 2

ESOPHAGEAL ACHALASIA
distortion of myelin profiles. Myelin was seen folded into discrete masses(Fig. 3 b) associated with axoplasm outside the confines of the myelinlayer (Figs. 3 c and d). Nuclear material was found within the axoniclumen in one instance (Fig. 3 d); this probably resulted from break-down of the axolemma at the node of Ranvier and passage of the Schwanncell nucleus into this space. Axolemmal profiles were uncommonly seenin distorted myelinated axons (Fig. 3 c).Schwann cells appeared in increased numbers in nerve specimens from
2 patients. The cytoplasm of Schwann cells appeared either coarselygranular (Fig. 4 a) (because of the presence of increased numbers ofsmall osmiophilic granules, vesicles and mitochondria) or relativelyagranular (Fig. 4 b). On rare occasions, a single Schwann cell was parti-tioned into granular and agranular areas (Fig. 4 c).The interstitial space did not differ in achalasia patients compared
with similar areas in nerves obtained from controls.
COMMENT
The alterations seen in the esophageal vagal nerves in achalasia aredegenerative in nature. The lesions resemble most those changes seenin experimentally induced Wallerian degeneration. Fragmentation anddispersion of neurofilaments with loss of longitudinal orientation arefeatures of Wallerian degeneration in the albino rat.14 Contraction ofthe axonic lumen has been described by others.15 Mitochondrial swellingand degeneration occur.16 The collapse of myelin associated with de-struction of axis cylinders has been described,15 although the total dis-integration of myelin into globules as encountered in experimentalstudies was not encountered in the present study. The absence of mes-axons is reportedly a regenerative feature following Wallerian degenera-tion.17 Coarse granularity of Schwann cell cytoplasm is characteristic,17.18and the cytoplasm of some Schwann cells later in the course of Walleriandegeneration becomes almost clear.'8 Finally, cytoplasmic splitting isdescribed in Schwann cells containing degenerated fibers after nervetransection.19
It was not possible in the present study to determine whether thesemorphologic alterations were occurring in motor or sensory fibers orin the small numbers of sympathetic fibers which course along with thevagus nerve.20 The destruction of the dorsal motor nucleus of the vagusin achalasia reported by Kimura3 and recently substantiated 21 iS con-sidered to constitute suggestive evidence that some of the degenerativechanges occur in motor fibers of the extra-esophageal vagus nerves.The findings suggest that vagus nerve lesions outside the esophagus
are important in the motility disorder of achalasia. The relationship
28IFeb., I965

282 CASSELLA, ELLIS AND BROWN Vol. 46, No. 2
of the lesions to myenteric ganglion cell alterations usually described inesophageal achalasia is obscure.
SUMMARYThe fine structure of vagal branches to the esophageal plexus in 9
patients with achalasia revealed lesions resembling Wallerian degenera-tion in each case. Both myelinated and nonmyelinated axons were in-volved, but normal areas appeared in all specimens as well. Axoplasmexhibited either swelling with fragmentation and dispersion of neuro-filaments or contraction with indistinct neurofilaments. Discontinuitieswere found in axon-Schwann membranes and mesaxons. Myelin disrup-tion was a common feature. Schwann cells had either coarsely granularor relatively agranular cytoplasm.
It was not possible to determine whether these changes involved motoror sensory axons of the vagus, but previous reports describing the de-struction of the dorsal motor nucleus of the vagus in achalasia patientssuggested that some of the lesions involved motor axons. The relation-ship of the changes in fine structure to alterations of the esophagealmyenteric cells usually described in achalasia is unknown.
REFERENCESI. HIEYROVSKY, H. Casuistik und Therapie der idiopathischen Dilatation der
Speiserohre. Oesophagogastroanastomose. Arch. klin. Chir., I9I2-I3, I00,703-7 I5.
2. LOEPER, M., and FORESTIER, J. Les lesions nerveuses du pneumogastrique etle cardiospasme recurrent dans le cancer de l'estomac. Arch. mal. app. digest.,I92I, II, 307-3I2.
3. KimuRA, K. The nature of idiopathic esophagus dilatation. Jap. J. Gastro-enterol., 1929, I, 199-207.
4. LENDRUM, F. C. Anatomic features of the cardiac orifice of the stomach: withspecial reference to cardiospasm. Arch. Int. Med., I937, 59, 474-5II.
5. ETZEL, E. Megaoesophagus and its neuropathology: a clinical and anatomo-pathological research. Guy's Hosp. Rep., I937, 87, I58-I74.
6. TAMiyA, C.; OKA, M., and KAWASHIMA, S. Experimentelle Studien ilber denOesophagus: Rontgenologische Untersuchungen der experimentellen Schluck-beschwerde des Oesophagus beim Hunde. (Einfluisse des Impulsausfailes deseinseitigen Vagus, des Sympathicus und des vagosympathischen Truncus).Jap. J. M. Sc. Tr., I927, VIII-I, IOI-I45.
7. MILHIET, H. Reflexions sur le mega-oesophage; a propos d'un cas traite parl'intervention chirurgicale et A propos des dilatations paralytiques experi-mentales de l'oesophage du chien. Presse mgd., I939, 2, I639-I641.
8. CARVETH, S. W.; SCHLEGEL, J. F.; CODE, C. F., and ELLIS, F. H., JR. Changesin the motor action of the esophagus and gastroesophageal sphincter aftervagotomy in dogs. Surg. Forum, I960, II, 3II-3I3.
9. ELLIS, F. H., JR.; OLSEN, A. M.; HOLMAN, C. B., and CODE, C. F. Surgicaltreatment of cardiospasm (achalasia of the esophagus); considerations ofaspects of esophagomyotomy. J.A.M.A., I958, I66, 29-36.

Feb., I965 ESOPHAGEAL ACHALASIA 283
IO. DALTON, A. J. A chrome-osimium fixative for electron microscopy. (Abstr.)Anat. Rec., 1955, 121, 28I.
xI. LUFT, J. H. Improvements in epoxy resin embedding methods. J. Biophys. &Biochem. Cytol., I96I, 9, 409-4I4.
I2. WATSON, M. L. Staining of tissue sections for electron microscopy with heavymetals. J. Biophys. & Biochem. Cytol., I958, 4, 475-478.
13. CAUSEY, G. The Cell of Schwann. E. & S. Livingstone, Ltd., Edinburgh andLondon, I960, I20 Pp.
14. ViAL, J. D. The early changes in the axoplasm during Wallerian degeneration.J. Biophys. & Biochem. Cytol., I958, 4, 55I-556.
I5. BARTON, A. A. An electron microscope study of degeneration and regenera-tion of nerve. Brain, I962, 85, 799-808.
I6. OHMI, S. Electron microscopic study on Wallerian degeneration of the periph-eral nerve. Z. Zellforsch., I96I, 54, 39-67.
I7. TERRY, R. D., and HARKIN, J. C. Regenerating peripheral nerve sheaths fol-lowing Wallerian degeneration. Exper. Cell Res., 1957, I13, I93-197.
M8. GRASSE, PmpER-P. etude au microscope electronique de la degenerescencewallerienne des fibres nerveuses amyeliniques. Compt. rend. Acad. sc., I959,248, 2796-2798.
z9. GLIMSTEDT, G., and WOHLFART, G. Electron microscopic studies on peripheralnerve regeneration. C. W. K. Gleerup, Lund, I960, 22 pp.
20. HOFFMAN, H. H., and SCHNITZLEIN, H. N. The numbers of nerve fibers in thevagus nerve of man. Anat. Rec., I96I, 139, 429-435.
2I. CASSELLA, R. R.; BROWN, A. L.; SAYRE, G. P., and ELLIS, F. H., JR. Achalasiaof the esophagus; pathologic and etiologic considerations. Ann. Surg., I964,i60, 474-487-
[Illustrations follow]

CASSELLA, ELLIS AND BROWN
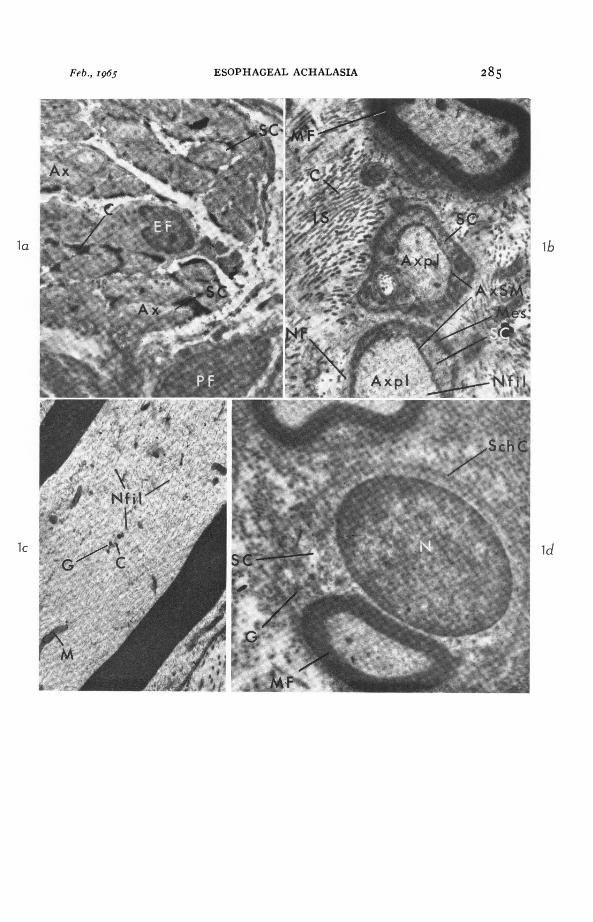
LEGENDS FOR FIGURESFIG. i. Normal vagal esophageal nerve.
(a) Entirely nonmyelinated nerve. Group of axons (Ax) are surrounded byslender processes of perineural fibrocytes (PF). Central area contains an endo-neurial fibrocyte (EF) and a fibrocytic process. Individual axons are uniformlysurrounded by Schwann cytoplasm (SC). Collagen (C). X 4,200.(b) Transverse section through a myelinated (MF) and two nonmyelinatedfibers (NF). Axoplasm (Axpl) is bright compared with surrounding Schwanncell cytoplasm (SC). Axon-Schwann membrane (AxSM). Neurofilaments (Nfil).Mesaxons (Mes) and formed elements of axoplasm are indistinct. Interstitialspace (IS) contains collagen (C) and amorphous material. X I5,6oo.(c) Longitudinal section through myelinated fiber. Axoplasm contains thinbranching neurofilaments (Nfil), mitochondria (M), clear vesicles (C) andosmiophilic granules (G). X 28,200.(d) Normal Schwann cell (Sch C) containing myelinated axon (MF) in itscytoplasm (SC). Ovoid nucleus (N). Schwann cell cytoplasm contains smallgranules (G) in aggregates. Mitochondria and vesicles not seen or indistinct.X I 5,840.
284 Vol. 46, No. 2
ESOPHAGEAL ACHALASIA 285
lb
id
Feb., I965
. .~~~A

CASSELLA, ELLIS AND BROWN
2a
1'IG. 2. Vagal esophageal nerves in achalasia.(a) Transverse section through a nonmyelinated axon (NF) showing fragmenta-tion and dispersion of neurofilaments (Nfil). Mitochondria (M). Axon-Schwannmembrane (AxSM). Schwann cytoplasm (SC). Collagen (C). X 37.670.(b) Longitudinal sections through two nonmyelinated axons (NF). Mitochondria(M) are increased in number and have lost their characteristic inner structure.Schwann cytoplasm (SC). Axon-Schwann membrane (AxSM) is fragmented.X 227,400.(c) Transverse sections through several nonmyelinated axons (NF) showing"hugging" of axons to surface membrane (SM ) of Schwanin cell so that mesaxons(Mes) are indistinct. X I5,700.
286 Vol. 46, No. 2
2b
ESOPHAGEAL ACHALASIA
3c
1AR;S,:,10 W
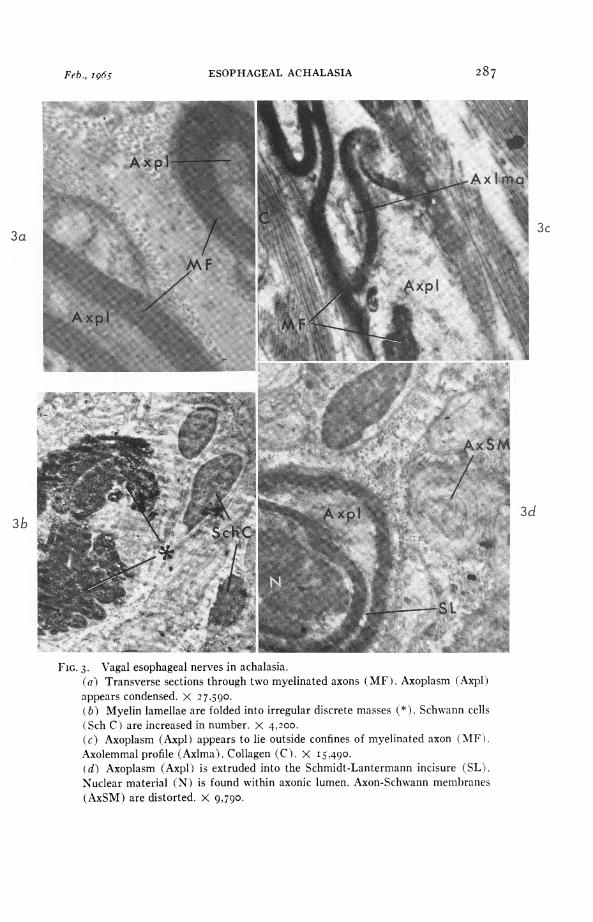
FIG. 3. Vagal esophageal nerves in achalasia.(a) Transverse sections through two myelinated axons (MF). Axoplasm (Axpl)appears condensed. X 27,590.(b) Myelin lamellae are folded into irregular discrete masses (*). Schwann cells(Sch C ) are increased in number. X 4,200.(c) Axoplasm (Axpl) appears to lie outside confines of myelinated axon (MF).Axolemmal profile (Axlma). Collagen (C). X I 5.490.(d) Axoplasm (Axpl) is extruded into the Schmidt-Lantermann incisure (SL).Nuclear material (N) is found within axonic lumen. Axon-Schwann membranes(AxSM) are distorted. X 9,790.
287Feb., I96-5
CASSELLA, ELLIS AND BROWN
4a
4b
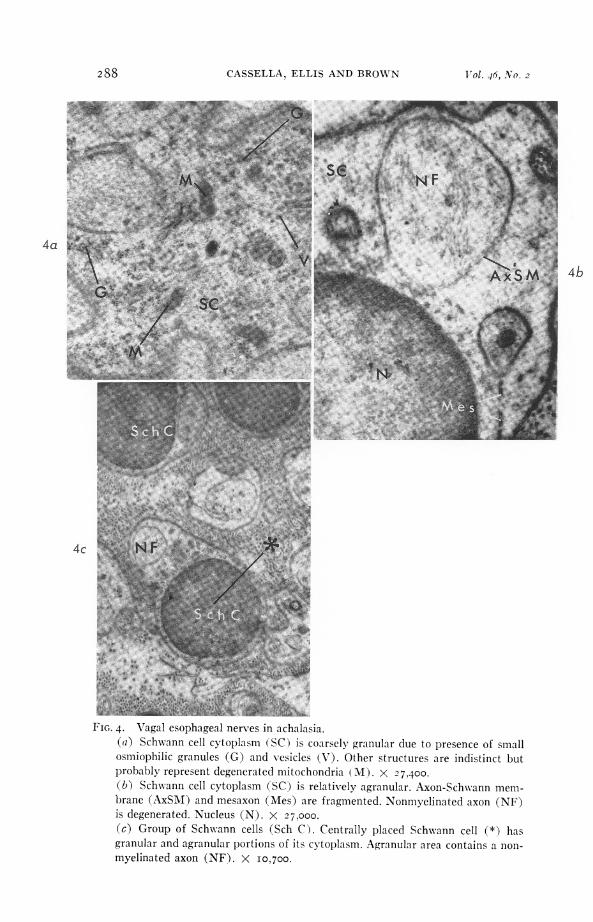
FIG. 4. Vagal esophageal nerves in achalasia.(a) Schwann cell cytoplasm (SC) is coarsely granular due to presence of smallosmiophilic granules (G) and vesicles (V). Other structures are indistinct butprobably represent degenerated mitochondria (M). X 27,400.(b) Schwann cell cytoplasm (SC) is relatively agranular. Axon-Schwann mem-brane (AxSM) and mesaxon (Mes) are fragmented. Nonmyelinated axon (NF)is degenerated. Nucleus (N). X 27.000.(c) Group of Schwann cells (Sch C). Centrally placed Schwann cell (*) hasgranular and agranular portions of its cytoplasm. Agranular area contains a non-myelinated axon (NF). X i0o700.
288 X ol. 46, No. 2










![*º&k ¼96 ]%4'5 m4(4E m ± g' >Ì Â £#è Êk-k-f.sakura.ne.jp/newsystem/wp-content/uploads/2019/04/1eb52e2d… · '¨>Þ>ä G D \ *º&k ¼96 ]%4'5 m4(4E m ± g' >Ì ` Û>à º#è](https://img.pdfslide.net/doc/110x75/6004e9f1463a96627107fc2b/k-96-45-m44e-m-g-oe-k-k-f-g-d.jpg)


















