Embed Size (px)
Citation preview

Substance P Innervation of the RatHippocampal Formation
ZSOLT BORHEGYI1 AND CSABA LERANTH1,2*1Department of Obstetrics and Gynecology, Yale University School of Medicine,
New Haven, Connecticut, 06520-80632Section of Neurobiology, Yale University School of Medicine,
New Haven, Connecticut 06520-8063
ABSTRACTLight and electron microscopic substance P (SP) immunostaining was performed on
hippocampal sections of colchicine-pretreated, control, untreated fimbria-fornix-transected (5days), as well as perforant path-stimulated Sprague-Dawley rats fixed in 5% acrolein.Numerous SP-immunoreactive neurons could be observed in the stratum oriens of theAmmon’s horn and subiculum, fewer were seen in the dentate hilar area and stratumradiatum of CA2 and CA3, and even fewer were seen at the border between the CA1 strataradiatum and the lacunosum moleculare of CA1 subfield. A higher dose of colchicine resultedin SP immunoreactivity in a large population of granule cells and mossy axon terminals. Theentire CA2 region, the stratum oriens of CA1, CA3, and the subiculum were denselyinnervated by SP-containing axon terminals. Ahomogeneous SP innervation was found in thestratum radiatum of CA1. Only a few SP fibers were seen adjacent to the granule cell layer.
Symmetric axosomatic contacts were seen between SP-containing boutons and somata inthe stratum oriens of theAmmon’s horn. However, throughout the hippocampal formation, themajority of SP-containing axons formed axodendritic symmetric synapses. Adense populationof SP-immunoreactive boutons that formed axodendritic asymmetric synapses was observedin the strata oriens and radiatum of the CA3a and CA2 regions, and a few were found inthe supragranular and subgranular layers of the dentate gyrus. Fimbria-fornix tran-section resulted in amarked loss of SP fibers in the strata oriens, pyramidale, and radiatum ofthe CA3a and CA2 subfields. In perforant pathway-stimulated animals, a population ofgranule cells and a large number of mossy axon terminals were immunoreactive for SP.These observations suggest two sources of SP innervation to the hippocampal formation: onearising from intrinsic sources (interneurons and granule cells) and one arising from ex-trinsic sources, most likely the supramammillary region. J. Comp. Neurol. 384:41–58, 1997.r 1997 Wiley-Liss, Inc.
Indexing terms: immunohistochemistry; dentate gyrus; Ammon’s horn; interneurons;
supramammillohippocampal tract
Numerous studies have demonstrated the importance ofsubstance P (SP) in the hippocampus. In the hippocampusof the human, SP cells were found to be susceptible toepileptic episodes, with the number of cells being greatlyreduced in the hippocampus of patients suffering fromtemporal lobe epilepsy (de Lanerolle et al., 1992). Kainicacid (KA) provides a well-known model for epilepsy (forreview, see Sperk, 1994). One of the effects of KA is onhippocampal SP synthesis. KA administration increasesthe preprotachykinin-A mRNA (SP precursor) level in therat hippocampus (Brene et al., 1992). SP is also involved inmnemonic functions. It has been shown that SP injected
peritoneally has memory-promoting and reinforcing ef-fects in rats (for review, see Huston et al., 1993). SP alsoinfluences the electrical activity of the hippocampus, be-cause, in slice preparations, 104 out of 115 nonpyramidal
Grant sponsor: NIH; Grant number: NS 26068.*Correspondence to: Csaba Leranth, M.D., Ph.D., Yale University, School
of Medicine, Department of Obstetrics and Gynecology, 339 FarnamMemorial Building, P.O. Box 208063, New Haven, CT 06520-8063.E-mail: [email protected] 12 July 1996; Revised 19 February 1997; Accepted 7 March
1997
THE JOURNAL OF COMPARATIVE NEUROLOGY 384:41–58 (1997)
r 1997 WILEY-LISS, INC.

cells responded with a reversible increase in firing (Drei-fuss and Raggenbass, 1986).Numerous SP-containing fibers and cell bodies have
been visualized by immunocytochemistry in the hippocam-pal formation of the guinea pig, domestic pig, cat, monkey,and even in that of human (Gall and Selawski, 1984;Bouras et al., 1986; Del Fiaccio et al., 1987; Sakamoto etal., 1987; Ino et al., 1988; Iritani et al., 1989; Gallagher etal., 1992; Holm et al., 1993; Seress and Leranth, 1996).Furthermore, ligand-binding studies have found a strongexpression of SP receptors in the guinea pig hippocampus(Dam and Quirion, 1986). This method as well as theapplication of in situ hybridization techniques (Shults etal., 1984; Mohini et al., 1985; Dam et al., 1990; Maeno etal., 1993) or immunocytochemical detection of the receptor(Nakaya et al., 1994) have provided the same results in therat hippocampus.In contrast to these physiological and pharmacological
data from rats, the immunocytochemical demonstration ofSP-containing elements in the hippocampal formation ofthe rat either failed to show (Cuello and Kanazawa, 1978;Shults et al., 1984) or showed only a low density of fibersand a few faintly stained, SP-containing cell bodies (Ljung-dhal et al., 1978; Vincent et al., 1981; Roberts et al., 1984;Davies and Kohler, 1985; Shults et al., 1987; Iritani et al.,1989). Former studies in this laboratory (Seress andLeranth, 1996) demonstrated that the use of acrolein inthe fixative as well as a longer incubation in the primaryantiserum (Davies and Kohler, 1985) could enhance the SPstaining in hippocampal sections.A recent study demonstrated the presence of g-aminobu-
tyric acid (GABA) in excitatory transmitter-containinggranule cells in the dentate gyrus of rats and monkeys,and, after perforant pathway stimulation in rats, GABAlevels were increased in these neurons (Sloviter et al.,1996). Furthermore, in a previous experiment, we wereable to show that intrinsic SP-containing neurons of thehippocampal formation of monkeys are GABAergic (Seressand Leranth, 1996). Taken together, we speculated thatgranule cells, in addition to expressing glutamate andGABA, are able to produce SP, and the level of the lattercan be increased by stimulation. Therefore, this studyemployed acrolein-fixed tissue incubated for a longer timein primary antiserum to describe the distribution andmorphology of the SP-immunoreactive cell bodies andfibers at both the light and electron microscopic levels incontrol, colchicine-treated, and fimbria-fornix-transectedand perforant path-stimulated rats.
MATERIALS AND METHODS
Animals and surgery
Adult female Sprague-Dawley rats (weighing 300–400 g)were used in these experiments. Animals were kept understandard laboratory conditions, with tap water and regu-lar rat chow ad libitum and on a 12-hour light/12-hourdark cycle. Surgical procedures were performed underdeep ketamine anesthesia (ketamine in a cocktail ofxylazine and acepromazine at a concentration of 25 mg/ml;injected i.m. at a dose of 3 ml/kg) through the use of a stereo-taxic apparatus (David-Kopf Instruments, Tujunga, CA).Fimbria-fornix transection was performed on four rats.
From a dorsal penetration, after aspiration of overlying
cortical areas and the corpus callosum, under visualcontrol, the right fimbria-fornix was completely transected(Jakab and Leranth, 1991). Animals were killed after a5-day survival time.Colchicine treatment was used on six rats to enhance
perikaryal labeling (Dahlstrom, 1971). Twenty-four hoursprior to euthanasia, a normal (125 µg/25 µl) or double (250µg/50 µl; three out of six rats) dose of colchicine was placedin the lateral ventricle by using a Hamilton syringe. Aftersurgery, the greatest risk was from dehydration. There-fore, preventive measures were taken by injecting 2-3 mli.p. of normal saline.To investigate the result of electrical stimulation of the
perforant pathway on SP-immunoreactivity, brains fromrats (generous gift from Dr. Sloviter) that had been perfo-rant path stimulated (for protocol, see Sloviter, 1983;Sloviter et al., 1996) and acrolein fixed according to ourprotocol, were sent to us for SP immunocytochemistry.
Tissue preparation
Fixation and tissue processing. Under deep etheranesthesia, animals were transcardially perfused with50–60 ml isotonic heparinized saline followed by 200–250ml fixative containing 5% acrolein in 0.1 M phosphatebuffer (PB), pH 7.4 (King et al., 1983). Following perfusion,the brains were removed, and the hippocampal complexwas dissected out. This tissue block was postfixed in thesame fixative for 30 minutes. Forty micrometer sectionswere cut from the septal, middle, and temporal parts of thehippocampal formation on a Vibratome (Lancer, St. Louis,MO) and rinsed in several changes of ice-cold PB. Sectionsfor electronmicroscopy were transferred into vials contain-ing 0.5 ml 10% sucrose (in PB) and rapidly frozen byimmersing the vial in liquid nitrogen. They were thenthawed to room temperature and repeatedly washed inPB. In order to eliminate the unbound aldehydes (Kosakaet al., 1986), sections for both light and electron micros-copy were incubated in 1% sodium borohydride for 30minutes followed by washing in PB.Light and electron microscopic single immunostain-
ing for SP. Light and electron microscopic single immu-nostaining for SP was performed on hippocampal Vibra-tome sections by using the avidin-biotin peroxidase (ABC;Vector Laboratories, Burlingame, CA; Hsu et al., 1981)technique. The sections were incubated first with 10%normal goat serum for 45 minutes, followed by 60 hours at4°C in a polyclonal rabbit anti-SP antibody [Leeman;1:5,000 dilution in PB containing 0.04% Triton X-100 (noTriton for electron microscopy) and 0.1% sodium azide].After this, the sections were incubated in the secondaryantibody [biotinylated goat anti-rabbit immunoglobulin(IgG), 1:250 in PB; Vector Laboratories] then in ABC Elite(1:250 in PB; Vector Laboratories), each for 2 hours at20°C. The tissue-bound peroxidase was visualized by usinga brown diaminobenzidine (DAB) reaction (15 mg DAB,165 µl 0.3% H2O2 in 30 ml PB) or by a nickel-intensifiedDAB (Ni-DAB) reaction (15 mg DAB, 12 mg NH4Cl, 0.12mg glucose oxidase, 600 µl 0.5 M Ni(NH4)2SO4, and 600 µl10% b-D-glucose in 40 ml PB) for 5–10 minutes. TheNi-DAB reaction (performed on light microscopic sections)resulted in a dark-blue to black reaction product. Betweeneach incubation step, sections were thoroughly rinsed inseveral changes of ice-cold PB. After the immunostaining
42 Z. BORHEGYI AND C. LERANTH

procedure, sections for light microscopic observation wereplaced on gelatin-coated slides, dehydrated, and mountedin Permount.Electron microscopy. For electron microscopy, sec-
tions were processed in the same way as for light micros-copy, except that no Triton X-100 was used in the primaryantisera. Following immunostaining, the sections wereosmicated (1% OsO4 in PB for 15 minutes), dehydratedthrough increasing ethanol concentrations (using 1% ura-nyl acetate in 70% ethanol for 30 minutes), flat embeddedinAraldite between liquid release-coated (ElectronMicros-copy Sciences, Fort Washington, PA) slides and coverslips,and placed in a 60°C oven for 48 hours. After this,flat-embedded sections were fixed with a drop of embed-ding medium on the top of cylindrical araldite blocks andcured again for 48 hours at 60°C. Sections were cut on aReichert-Jung Ultracut E ultramicrotome (Leica, Deer-field, IL), collected on Formvar-coated single-slot grids,and examined in a Philips CM-10 electron microscope.
Tissue sampling
For light microscopic examination, all of the sections cutfrom the septal, middle, and temporal parts of the hippo-campal formation from both hemispheres of each rat wereanalyzed. In the unilateral fimbria-fornix-transected ani-mals, the staining pattern of the ipsilateral hippocampuswas compared with that of the contralateral hippocampusas well as with hippocampal sections of control animals.For electron microscopy, four blocks were cut from eachregion (i.e., septal, middle, and temporal) of the hippocam-pal formation from rats (n 5 2) of each experimentalgroup. The analysis was performed on at least 20 ultrathinsections prepared from eachmajor hippocampal subfield ofevery tissue block. Furthermore, in order to gain a compre-hensive view of the distribution pattern and type (symmet-ric vs. asymmetric) of synaptic contacts, at least 100SP-immunoreactive boutons were analyzed from eachhippocampal subfield.
Primary antiserum and control experiments
According to the characterization by Dr. S. Leeman, theantiserum binds 40% of [125I]SP trace at a 1:250,000 finaldilution. Half-maximal displacement by SP is 30 pM. Nomeasurable displacement is observed at 100 pM withsubstance K, physalaemin, met-enkephalin, or eledoisin(Carraway and Leeman, 1979). This SP antiserum hasbeen used in other experiments visualizing immunoreactiv-ity for SP in the rat septum (Szeidemann et al., 1995) andmonkey hippocampus (Leranth and Nitsch, 1994; Nitschand Leranth, 1994). To further test the specificity of theantiserum on the rat hippocampus, two control experi-ments were performed: 1) Hippocampal sections wereincubated with an SP antiserum (diluted to 1:5,000)preadsorbed (24 hours at 4°C) with a 23.5 µM solution ofSP (S6883; Sigma Chemical Corp., St. Louis, MO), and 2)the SP antiserum was replaced with normal rabbit serum.In both cases, no immunoreaction could be observed. Theprotocols for the animal care and treatments used in thisstudy were approved by the Yale Animal Care and UseCommittee.
RESULTS
Distribution and morphologyof SP-immunoreactive cell bodies
Only a few faintly stained SP-containing neurons couldbe observed in untreated rats. Therefore, the analysis wasperformed on colchicine-treated animals. The majority ofSP-immunoreactive neuronsweremultipolar (Fig. 1). How-ever, in the subgranular layer of the dentate gyrus (Figs.2a, 3a,b) and at the border between the alveus and thestratum oriens (Fig. 7b), some horizontally oriented, fusi-form neurons were seen. Occasionally, bipolar neuronscould also be detected in the alveus. The SP-immunoreac-tive cells of the hilar region were smaller than those of theAmmon’s horn.Dentate gyrus and the hilar area. Approximately 14
SP-immunopositive neurons were seen in the hilar area oneach 40-µm-thick horizontal section of the midbody of thehippocampus. Fusiform neurons occupied areas subjacentto the stratum granulosum (Fig. 2a). The majority ofmultipolar cells were found in the deeper hilar area (Fig.2a). However, some could be found among the fusiformcells. Electronmicroscopic examination revealed that theseSP-labeled cells exhibited infolded nuclei (Fig. 3b). Thus,they appeared to be interneurons.In animals receiving a higher dose of colchicine (250 µg
in 50 µl), substantial numbers of small (6–12 µm indiameter) SP-immunoreactive neurons were seen in thegranule cell layer (Figs. 1, 2a). Upon electron microscopicobservation, these cells did not exhibit infolded nuclei (Fig.2b). Their dendrites entered into the molecular layer, andtheir axon initial segments (swollen in colchicine-treatedanimals) and axons were directed toward the hilus. SP-immunoreactive terminals with a morphology resemblingmossy fibers were also seen in the stratum lucidum. On thebasis of this evidence, these SP-immunopositive neuronswere identified as granule cells.After unilateral stimulation of the entorhinal cortex,
SP-immunoreactive neurons appeared in the granule celllayer of both sides of the dentate gyrus. However, the SPstaining was much weaker on the contralateral side (Fig.1b,d). Moreover, a dense SP-immunoreactive fiber networkwas seen in the hilus (Fig. 1b,d) and in the stratumlucidum of CA3 (Fig. 1b,c) with giantmossy-like terminals.According to these light microscopic observations, wedetermined that these were granule cells.Ammon’s horn and subiculum. In a single Vibratome
section, approximately 50–60 SP-immunoreactive neuronswere seen in the Ammon’s horn (Fig. 1). The vast majorityof these cells (45–50) were found in the strata oriens andpyramidale. The SP cell density in the CA2 region was abit higher than in other CA regions. Fewer SP cells (6–9)were seen in the strata lucidum and radiatum of CA3-CA2.Occasionally, SP-immunoreactive cell bodies were ob-served on the border between the strata radiatum and thelacunosum moleculare (Fig. 1). In the subicular area,approximately 10–20 SP-immunoreactive neurons werevisible in the strata oriens and pyramidale.Almost all SP-labeled cells were multipolar. Fusiform
cells were seen to be horizontally oriented close to thealveus (Figs. 1, 7b) and, occasionally, on the border of thestrata radiatum and lacunosum moleculare in the CA1region. Bipolar neurons rarely stained in the alveus.
SP INNERVATION OF THE RAT HIPPOCAMPUS 43
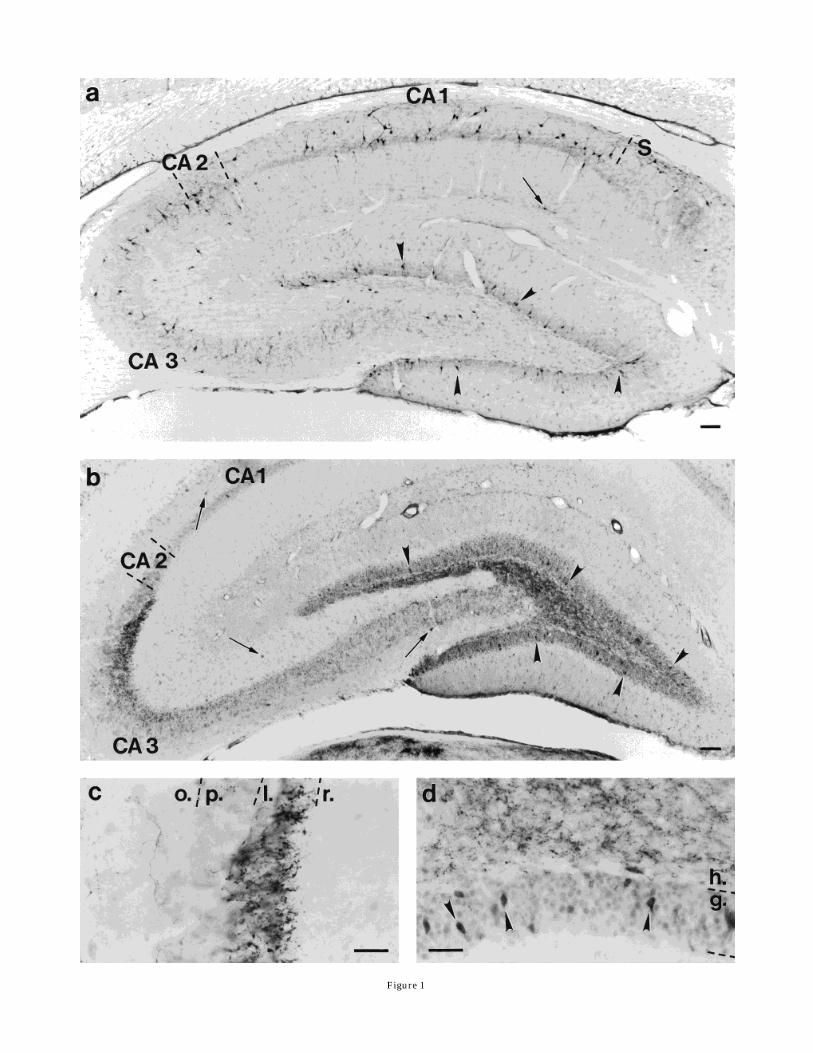
Figure 1

On the basis of their light microscopic morphology, theSP-immunopositive cells in the strata oriens and pyrami-dale can be divided into two principal groups (Figs. 1, 7b):first, fusiform cells, which were found close to the alveus.The dendrites of these neurons were oriented mostlyparallel to the alveus. The beginning of their axons ranhorizontally; they were then directed toward the pyrami-dal cell layer and sometimes passed through it. The othergroup, formed by multipolar neurons, was found in thepyramidal cell layer and the adjacent strata oriens andradiatum. Their dendrites were seen in stratum oriens aswell as in stratum radiatum. The axons of these cellsseemed to be oriented in different directions.In the subiculum, the pyramidal cell layer was much
wider, but the SP cells were distributed in the same way asin the Ammon’s horn described above. It must be notedthat, under the light microscope, in addition to the denselystained SP-immunoreactive neurons, several lightlystained glia cells could also occasionally be observed (Fig.1a,b). We interpreted this staining as a nonspecific label-ing, which is associated with the use of acrolein and,perhaps, with the longer Ni-intensified DAB reaction,because this labeling 1) was restricted only to the verysurface of the sections, and 2) it could not be seen on theelectron microscopic material, which was reacted in a DABsolution, or in paraformaldehyde-fixed (colchicine-pre-treated) tissue.
Distribution and morphologyof the SP-immunoreactive fiber network
Sections of untreated rat hippocampi were analyzed.Specific axon patterns were seen in each subregion of thehippocampal formation. The majority of the SP-labeledterminals were found to form symmetric synapses, mostlyon dendrites. The type of synapses, symmetric vs. asymmet-ric, was determined by the criteria outlined by Peters et al.(1991).Dentate gyrus. Sparsely distributed, SP-immunoposi-
tive fibers and puncta were found in the supragranularand subgranular layers of the dentate gyrus and in the
hilar area (Fig. 3a). Thin SP-immunoreactive processesresembling axons passed through the granule cell layer(Fig. 3a). In all of the aforementioned areas, SP boutonswere found to form symmetric synapses, mostly on den-drites. However, contacts were found on cell bodies in thegranule cell layer. The ultrastructural features of theseneurons resembled granule cells. On rare occasions, SP-labeled terminals that formed asymmetric synapses wereseen, mostly on dendrites (Fig. 3c), in both the supragranu-lar layer and the subgranular layer.CA3c and CA3b. Amoderately dense SP-immunoreac-
tive axon network was found in the stratum oriens, close tothe pyramidal cell layer (Fig. 5a). Straight, varicose,winding axons and basket-like terminals were present inthis area. In order of greatest to least density, the stratumlucidum, alveus, and stratum pyramidale were also inner-vated by SP-immunoreactive fibers. A group of basket-likeSP axons was seen in the CA3c subfield next to the hilus inthe stratum radiatum (Fig. 4a). The other parts of thestratum radiatum and the stratum lacunosum molecularewere almost free of fibers. Upon electron microscopicexamination, SP-labeled terminals were found to formsymmetric synapses, mostly on proximal (stratum lu-cidum) but also with distal dendrites. Whether thesedendrites are processes of principal or nonprincipal neu-rons could not be determined. SP-labeled terminals mak-ing synaptic contact on cell bodies were uncommon. Thebasket-like SP terminals in the stratum radiatum of theCA3c subregion established symmetric synapses on cellbodies (Fig. 4b). Because of their location and morphologi-cal characteristics (neurons rich in endoplasmic organellesand containing infolded nuclei), these cells appeared torepresent nonprincipal neurons (Fig 4b).CA3a and CA2. SP fibers were found in all areas of
this region. In the CA3a subfield, a fiber pattern similar tothat in the adjacent CA3b area was seen, with two markeddifferences (Fig. 5b): Even more SP-labeled fibers andpuncta were observed in the stratum oriens near thestratum pyramidale, and winding fibers and boutons werefound in the stratum radiatum (Fig. 5c). This distributionof the SP fibers spread into the CA2 subregion, wheremorebouton-like puncta and thin axons were observed in thealveus, strata oriens, and radiatum. The pyramidal celllayer in the CA2 subfield was well innervated comparedwith other Ammon’s horn regions. Occasionally, SP-containing axons were observed in the stratum lacunosummoleculare. Electron microscopic examinations revealednumerous SP-labeled terminals that formed symmetricsynapses in the stratum oriens and the alveus on distaldendrites. Cell bodies of neurons that appeared to bepyramidal cells (Fig. 6b) and their proximal dendriteswere also postsynaptic targets of SP-immunoreactive bou-tons in the strata pyramidale and radiatum. Practically noSP synapses were found in the stratum lacunosummolecu-lare.Abundant SP-labeled terminals that formed asymmet-ric synapses were seen around the pyramidal cell layerand in the radiatum, mostly on proximal dendrites (Fig.6a) but also on distal dendrites.CA1 area. A homogeneous, dense, SP fiber network
innervated the strata oriens and radiatum and the alveusof this area (Fig. 7a). Fewer SP-containing axons werevisible in the pyramidal cell layer, whereas the stratumlacunosum moleculare was almost devoid of fibers. On the
Fig. 1. Distribution of substance P (SP)-immunoreactive cells inthe rat hippocampal formation after colchicine-treatment (a) or ento-rhinal stimulation (b–d). a: The majority of SP-immunoreactiveneurons are multipolar; however, some fusiform neurons are alsovisible in every hippocampal subfield. The strata oriens and pyrami-dale in all CA subfields contain the most dense population of labeledcells. Fewer SP neurons are seen in the strata lucidum and radiatumof the CA3 and CA2 areas and in the hilar region of the dentate gyrusas well as in the subiculum (S). Occasionally, there are immunostainedcell bodies (arrow) on the border between the CA1 strata radiatum andthe lacunosum moleculare. The stratum lacunosum moleculare of theentire Ammon’s horn and stratummoleculare of the dentate gyrus arefree of SP neurons. SP cells located in the granule cell layer (some areindicated by arrowheads) are visible only after an increased dose ofcolchicine.After stimulation of the entorhinal cortex, SP-immunoreac-tive neurons appear in the granule cell layer (some are indicated byarrowheads in b and d). In addition, SP-immunoreactive processesresembling mossy fiber axons and axon terminals can be seen in thehilus (a and d) and in the stratum lucidum of the CA3 region, which isshown in b and at higher magnification in c. Only a few cells areweakly stained without colchicine (some are indicated by arrows in b).g., Granule cell layer; h., hilus; l., stratum lucidum; o., stratum oriens;p., pyramidal cell layer; r., stratum radiatum. Scale bars 5 100 µm ina,b, 40 µm in c,d.
SP INNERVATION OF THE RAT HIPPOCAMPUS 45
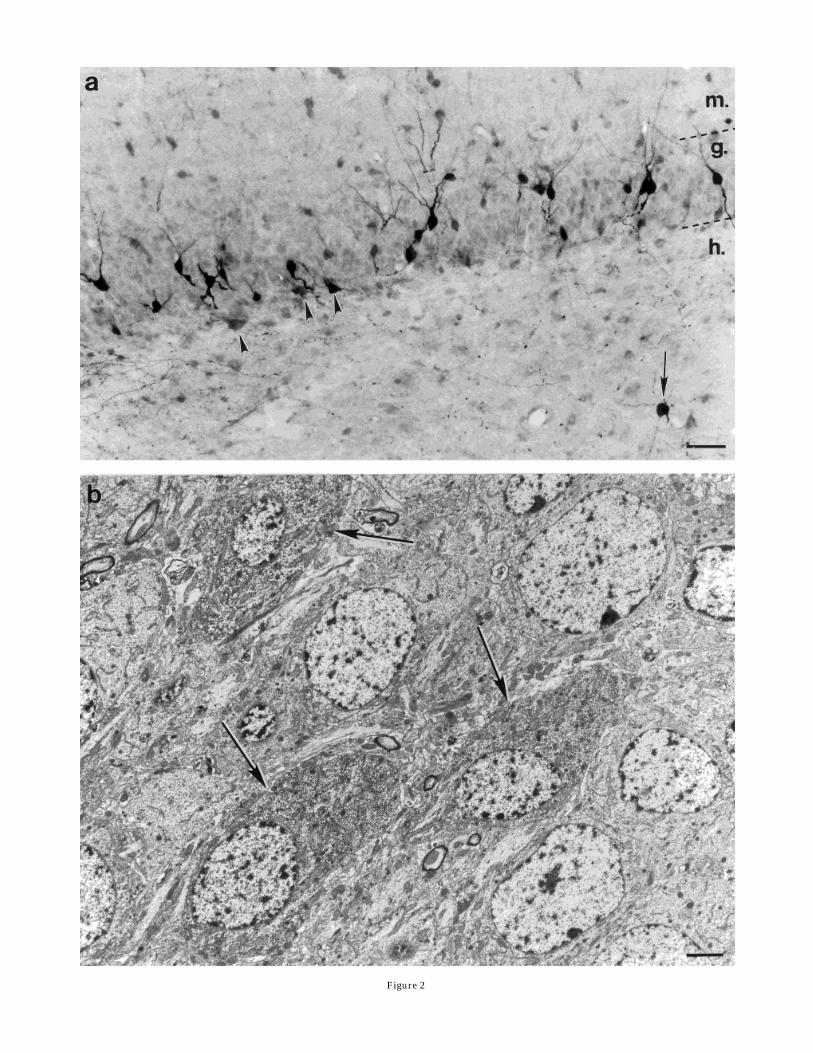
Figure 2

border of the strata radiatum and lacunosum moleculare,scattered fibers with larger boutons were visible (Fig. 7c).The electron microscopic observations showed SP-labeledterminals that formed only symmetric synapses in everyaforementioned area, predominantly on distal dendrites(Fig. 8d) but sometimes on proximal dendrites (Fig. 8a).The latter were designated as pyramidal cell dendrites.Occasionally, somata contacted by SP boutons were ob-served.
Fig. 3. Light (a) and electron (b,c) micrographs of the SP-immunostained dentate gyrus of a noncolchicine-treated rat. a: SP-containing fibers and boutons in (arrowhead) and adjacent to (smallarrows) the granule cell layer (g.) and the dentate hilus (h.). Occasion-ally, in the hilar area and the subgranular layer, a few weaklyimmunostained SP-immunoreactive neurons are also seen (a, large
arrow). An electron micrograph of one of these cells is shown in b. Notethe deep nuclear infoldings of this neuron (arrowheads). A high-powerelectron micrograph (c) shows that, in the dentate hilus, an SP-immunoreactive bouton forms a symmetric synapse. m., molecularlayer. Scale bars 5 20 µm in a, 2 µm in b, 0.4 µm in c.
Fig. 2. Light (a) and electron (b) micrographs demonstrate SP-containing neurons in the dentate gyrus of a colchicine-treated rat.SP-immunoreactive multipolar neurons in the hilus (h. in a, arrow)and multipolar and fusiform cells in the subgranular layer (arrow-heads) can be visualized without colchicine treatment. However, SPcells in the granule cell layer (g.) are visible only after an intracerebro-ventricular colchicine injection. A group of these cells is shown on theelectron micrograph (b; arrows). The morphological features of theseimmunolabeled granule cells are identical to SP-immunonegativegranule cells. Scale bars 5 40 µm in a, 2 µm in b.
SP INNERVATION OF THE RAT HIPPOCAMPUS 47
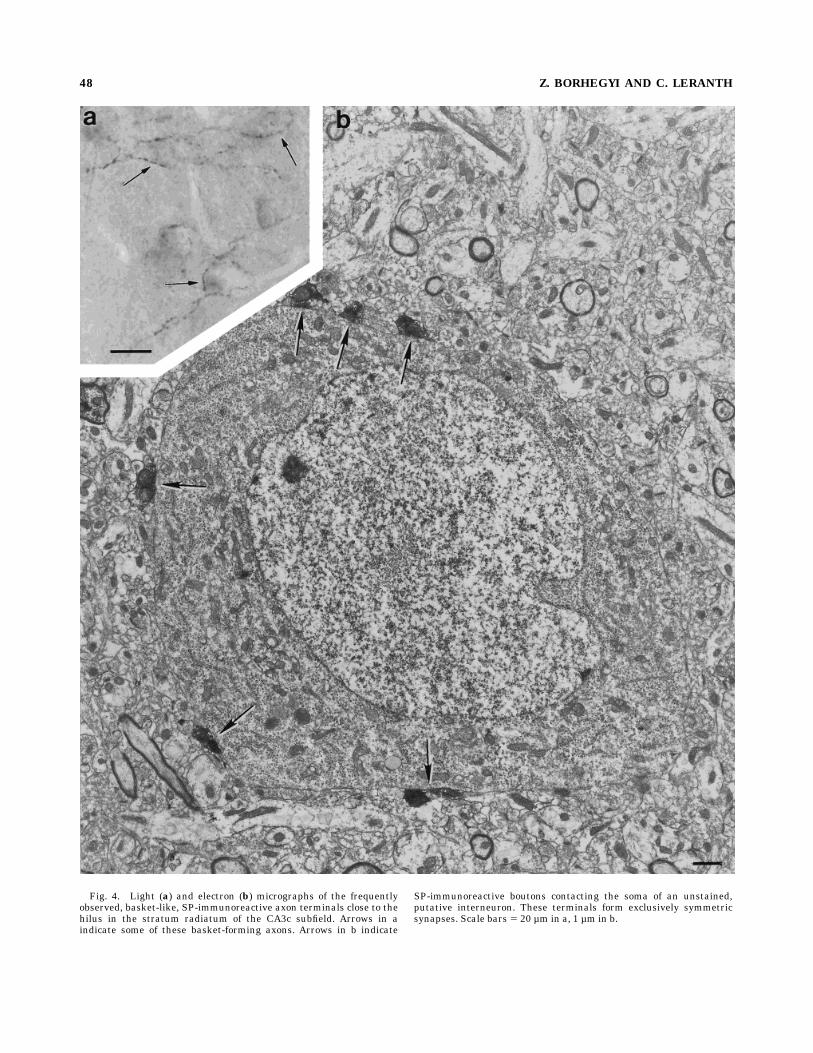
Fig. 4. Light (a) and electron (b) micrographs of the frequentlyobserved, basket-like, SP-immunoreactive axon terminals close to thehilus in the stratum radiatum of the CA3c subfield. Arrows in aindicate some of these basket-forming axons. Arrows in b indicate
SP-immunoreactive boutons contacting the soma of an unstained,putative interneuron. These terminals form exclusively symmetricsynapses. Scale bars 5 20 µm in a, 1 µm in b.
48 Z. BORHEGYI AND C. LERANTH

Subiculum. Many fibers were stained for SP in thisarea. The majority of the SP-immunoreactive axons andbouton-like puncta were found in the pyramidal cell layer(Fig. 9a). Winding fibers often formed basket-like termi-nals. Fewer labeled axons were seen in the strata oriensand radiatum, with hardly any in the alveus and stratumlacunosummoleculare. SP-immunopositive terminals thatformed symmetric synapses were found on proximal anddistal dendrites and on the cell bodies of pyramidal cellsand interneurons (Fig. 9b,c) throughout the area.
Fimbria-fornix transections
Because the transection of the fimbria-fornix was unilat-eral, we were able to use the contralateral hippocampus asa control. Under the light microscope, a marked decreasein the number of SP-containing fibers was detected aroundthe border between the CA3 and CA2 regions. Windingaxons (shown on Fig. 5c) disappeared from the radiatum,and only puncta-like SP staining remained in the strataoriens and pyramidale (Fig. 5d). No decrease in thenumber of SP fibers in the supragranular layer of thedentate gyrus was detected. Furthermore, no differenceswere observed in the contralateral (control) hippocampalformation of fimbria-fornix-transected rats compared withthat of untreated animals. In the electron microscope, onlysymmetric SP contacts could be detected in all hippocam-pal areas of the ipsilateral hippocampal formation.
Contacts between SP-immunoreactiveelements
Light microscopic examination of sections from normal,colchicine-treated animals occasionally showed SP-immu-nopositive, putative terminals on SP-stained proximaldendrites as well as on perikarya (Fig. 10). These homolo-gous contacts were found in the stratum oriens of everysubregion of the Ammon’s horn and subiculum. Some ofthese sections were embedded and processed for electronmicroscopy. The SP-immunoreactive fibers were found toform symmetric synaptic contacts on SP-labeled proximaldendrites and cell bodies (Fig. 10a,b).
DISCUSSION
By using acrolein fixation and an extended incubation inthe primary antiserum, this study showed an abundantnetwork of SP-containing axons and boutons as well as alarge population of SP-immunoreactive neurons locatedmostly in the stratum oriens of theAmmon’s horn. Further-more, whereas, in other studies, the use of colchicine wasalways indispensable in the visualization of SP-containingneurons (Cuello and Kanazawa, 1978; Ljungdhal et al.,1978; Vincent et al., 1981; Davies and Kohler, 1985; Shultset al., 1987), in this experiment, a population of neuronscould be visualized without colchicine treatment (Figs. 3a,7a, 9a).
SP-containing axons
Similar to the monkey hippocampal formation (Seressand Leranth, 1996), two types of SP-immunoreactive axonterminals could be observed in the rat: those that formsymmetric or asymmetric synaptic membrane specializa-tions. SP-containing axon terminals that form symmetricsynapses appear to be intrinsic, whereas SP boutons that
establish asymmetric synaptic contacts seem to be extrin-sic and may originate in the supramammillary area. Thisview is supported by the following observations: 1) Similarto other species (Gall and Selawski, 1984; Nitsch andLeranth, 1994), the density of SP axons in the rat CA2-CA3a areas (that contained the majority of asymmetric SPcontacts) was greatly reduced after fimbria-fornix transec-tion; 2) following hippocampal injection of a retrogradetracer in rats (Borhegyi and Leranth, unpublished observa-tion) as well as in other species, including cat (Ino et al.,1988), guinea pig (Gall and Selawski, 1984), and monkey(Nitsch and Leranth, 1994), retrogradely labeled SP-immunoreactive neurons were observed in the supramam-millary area only; 3) in the monkey supramammillaryarea, SP neurons cocontain calretinin (Nitsch and Leranth,1993), and, both in monkeys (Nitsch and Leranth, 1994) andin rats (Magloczky et al., 1994), supramammillohippocam-pal-projecting CR neurons form exclusively asymmetricsynaptic contacts in the hippocampus; 4) anterogradelylabeled axon terminals that originate in the supramammil-lary area form exclusively asymmetric synapses (Magloc-zky et al., 1994); and 5) after a fimbria-fornix transection,electron microscopic analysis revealed no asymmetric SPsynapses in the ipsilateral hippocampal formation (pres-ent study).The aforementioned studies also indicate that the par-
ent neurons of the extrinsic SP fibers are located in thesupramammillary area. In contrast, there are biochemicaland morphological data indicating their septal location.Septal lesion significantly decreased the SP content of thehippocampus (Vincent and McGeer, 1981), and SP-immunoreactive neurons located in the border betweenthe lateral and medial septal areas were shown to beretrogradely labeled following injection of wheat germagglutinin-conjugated horseradish peroxidase into the hip-pocampal formation (Peterson and Shurlow, 1992). How-ever, septal lesion can destroy the supramammillohippo-campal fiber tract; Lamour et al. (1989) could not findretrogradely labeled SP cells in the septum followinghippocampal injection of a retrograde tracer; and, accord-ing to Szeidemann et al. (1995), although the lateralseptum, particularly the dorsolateral septal area, containsSP neurons, all of these cells are intrinsic, GABAergicneurons that form exclusively symmetric synapses in thelateral septum.In addition to similarities in the SP innervation of the
hippocampal formation of different species, there aremajor differences. Whereas, in the dentate gyrus of guineapig (Gall and Selawski, 1984), cat (Ino et al., 1988),monkey (Iritani et al., 1989; Nitsch and Leranth, 1994;Seress and Leranth, 1996), and human (Del Fiaccio et al.,1987), the supragranular layer is densely innervated bySP axons (forming asymmetric contacts in the monkey;Nitsch and Leranth, 1994; Seress and Leranth, 1996), onlya few SP fibers could be found in the rat dentate gyrus(Davies and Kohler, 1985; present study). Another differ-ence between monkeys and rats is in the SP innervation ofthe dentate distal molecular layer and stratum lacunosummoleculare of the Ammon’s horn. In monkeys, these areascontain an abundant network of intrinsic SP-containingaxons that form symmetric synaptic contacts (Iritani et al.,1989; Seress and Leranth, 1996), whereas, in rats, this
SP INNERVATION OF THE RAT HIPPOCAMPUS 49
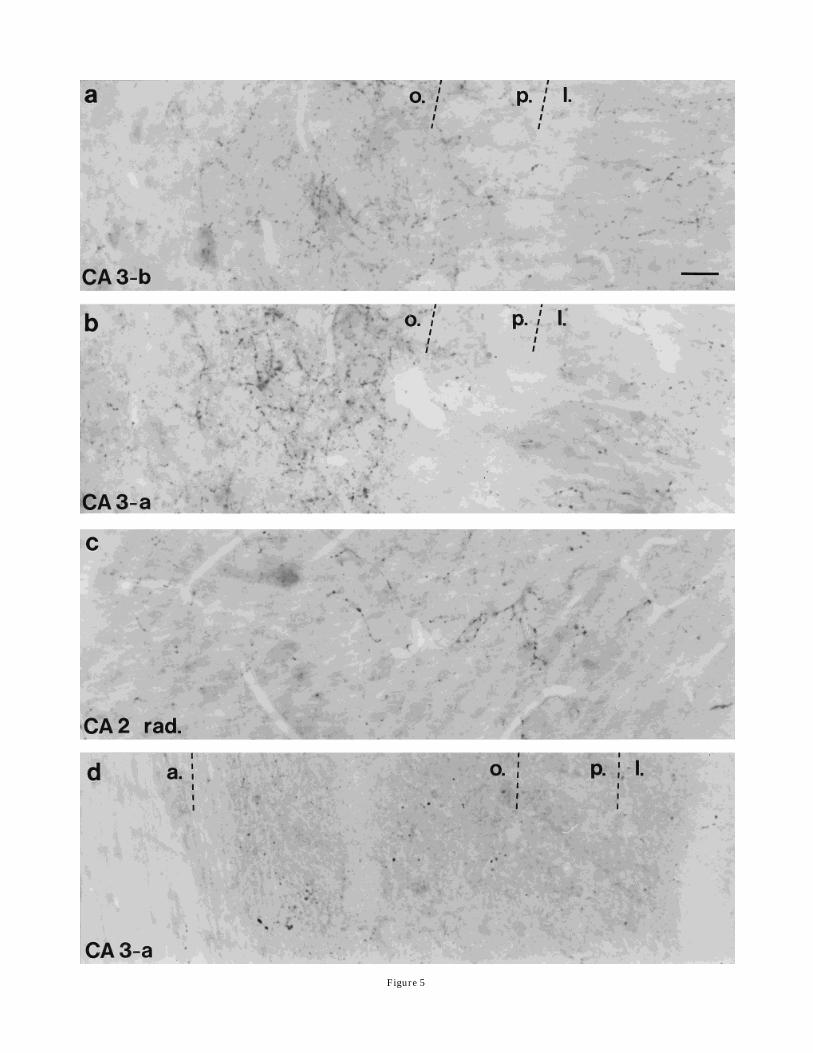
Figure 5
plexus cannot be observed (Davies and Kohler, 1985;present study).
Postsynaptic targets of SP-immunoreactiveboutons
Compared with the SP innervation of theAmmon’s horn,only a moderate number of SP axons could be observed inthe dentate gyrus. The very low density of SP-immunoposi-tive axons in the dentate gyrus and the very high densityof SP receptors in this area (Liu et al., 1994; Nakaya et al.,1994; Acsady et al., 1997) is another example of thewell-known mismatch in the location of neuroactive sub-stances and their receptors (Nakaya et al., 1994; Jakabet al., 1996). It has been suggested that SP and otherpeptides can reach their receptors via a nonsynapticdiffusion (Herkenham, 1987; Liu et al., 1994). This view issupported by the observation that SP receptors are notonly associated with postsynaptic structures but are homo-
geneously distributed on entire perikaryal and dendriticmembranes (Liu et al., 1994;Acsady et al., 1997). Thus, thepossibility exists that areas with high densities of SPreceptors are those in which the role of SP is mostimportant and may reflect a system primed for maximalresponse to any SP that is released.Another significant disparity is evident between SP
receptor-containing and SP-innervated neurons. Accord-ing to light and electron microscopic SP receptor studies,only the nonprincipal neurons of the rat hippocampalformation contain SP receptors (Acsady et al., 1997).However, in the present study, SP-containing boutonswere found to contact both principal cells (Fig. 8) andnonprincipal cells (Figs. 4, 9, 10). Furthermore, SP has adramatic effect on dispersed granule cells (I. Mody, per-sonal communication).
SP-containing neurons
Despite the use of colchicine, only moderate numbers ofSP-immunoreactive neurons have been reported in thealdehyde-fixed rat hippocampal formation (Ljungdhal etal., 1978; Vincent et al., 1981; Davies and Kohler, 1985;Shults et al., 1987). The use of an acrolein fixative greatlyincreased the numbers of detectable SP cells in all hippo-campal areas. The majority of SP neurons were seen in theAmmon’s horn. One population of these cells was foundadjacent to the pyramidal cells, and their dendrites ex-tended throughout the strata radiatum, pyramidale, andoriens. The location and dendritic arbor of these neurons(Fig. 7b) resembled that of the bistratified neurons (Sik et
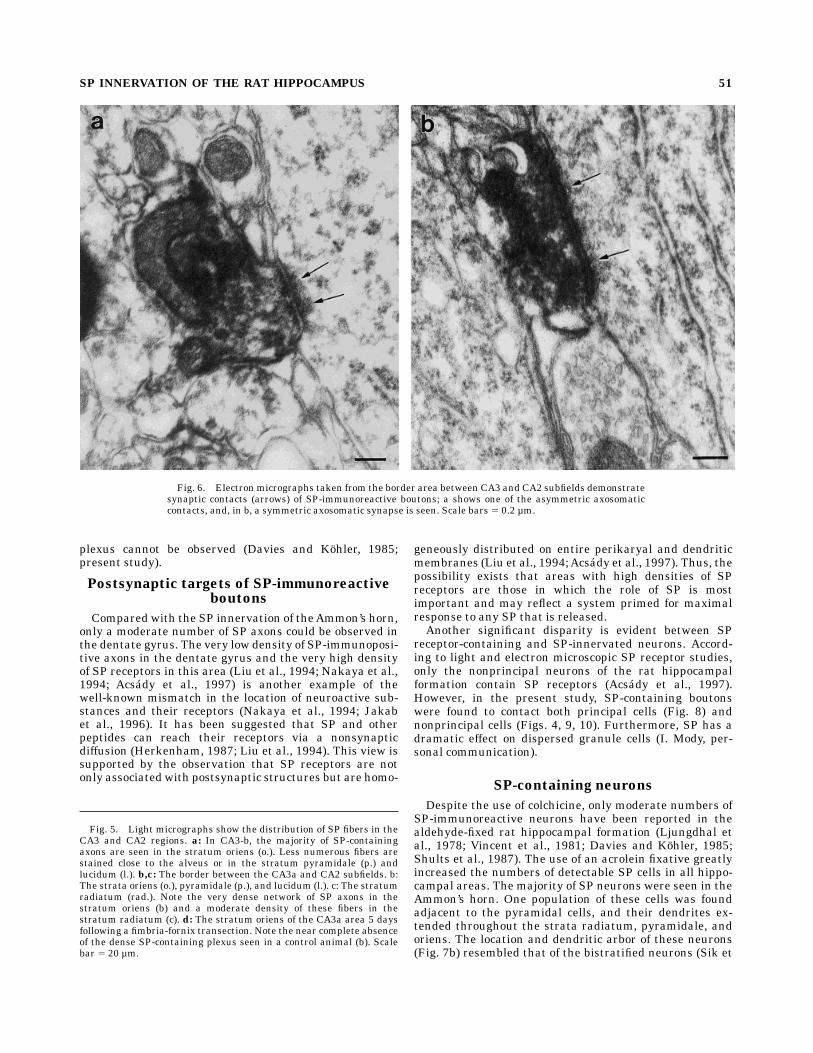
Fig. 6. Electron micrographs taken from the border area between CA3 and CA2 subfields demonstratesynaptic contacts (arrows) of SP-immunoreactive boutons; a shows one of the asymmetric axosomaticcontacts, and, in b, a symmetric axosomatic synapse is seen. Scale bars 5 0.2 µm.
Fig. 5. Light micrographs show the distribution of SP fibers in theCA3 and CA2 regions. a: In CA3-b, the majority of SP-containingaxons are seen in the stratum oriens (o.). Less numerous fibers arestained close to the alveus or in the stratum pyramidale (p.) andlucidum (l.). b,c: The border between the CA3a and CA2 subfields. b:The strata oriens (o.), pyramidale (p.), and lucidum (l.). c: The stratumradiatum (rad.). Note the very dense network of SP axons in thestratum oriens (b) and a moderate density of these fibers in thestratum radiatum (c). d: The stratum oriens of the CA3a area 5 daysfollowing a fimbria-fornix transection. Note the near complete absenceof the dense SP-containing plexus seen in a control animal (b). Scalebar 5 20 µm.
SP INNERVATION OF THE RAT HIPPOCAMPUS 51
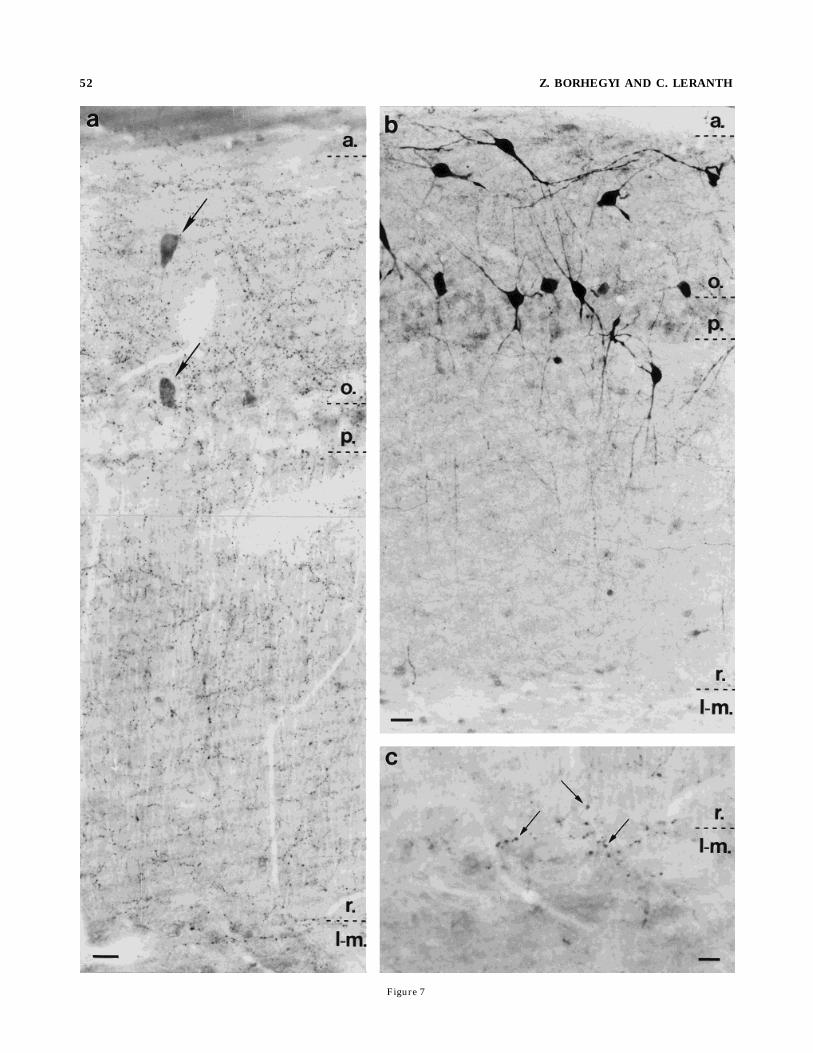
Figure 7
52 Z. BORHEGYI AND C. LERANTH

al., 1995). The homogeneous SP innervation of the strataoriens and radiatum of the CA1 region and the symmetricsynapses that were found in this area would support thishypothesis.Another important observation of this study is the
presence of immunoreactivity for SP in a population ofgranule cells and mossy axon terminals in animals treatedwith high doses of colchicine. The possibility that this is anonspecific staining of these profiles can be excluded,because, on the same ultrathin sections, several otherunstained granule cells and mossy terminals could be seen(Fig. 2b). It is widely accepted that colchicine blocks axonaltransport, resulting in the accumulation of substancesproduced in the soma that enables their immunodetection.However, one can speculate on another action of colchicine.Earlier studies showed that colchicine administration canalter themRNAlevel of cholecystokinin, somatostatin, andneuropeptide Y in the hippocampus (Cortes et al., 1990)and SP mRNA levels in the hypothalamus (Boyer et al.,1993). This view is also supported by the observation thatchemical excitation of the granule cells by KA can increasethe SP mRNA level 20-fold over the detection level (Breneat al., 1992). Moreover, systemic KA application can in-crease the mRNA level of neuropeptide Y (Sperk et al.,1992) or glutamic acid decarboxylase (Schwarzer andSperk, 1995; Sloviter et al., 1996) in granule cells. On theother hand, it is also possible that, in nonphysiologicalconditions, hippocampal granule cells are capable of produc-ing different peptides. Although SP-immunoreactive,mossy-like axon terminals in the CA3 region of the un-treated domestic pig have been reported (Holm et al.,1993), it is still uncertain whether this ‘‘SP production’’ inrats is a de novo synthesis in special circumstances or onlya problem of the sensitivity of our techniques.
Functional considerations
Very little is known about the physiological action of SPon hippocampal neurons. Dodd and Kelly (1981) concludedthat SP has no action on hippocampal pyramidal cells.However, an excitatory action by SP has been shown inother projecting systems (for review, see Nicoll et al.,1980). Furthermore, a more recent study demonstratedthat, out of 115 nonpyramidal neurons, 104 responded toSP administration with a reversible, concentration-dependent, increased firing (Dreifuss and Raggenbass,1986). This stimulatory action can be realized throughdifferent second-messenger systems, including a phospha-tidylinositol hydrolysis and cyclic AMP cascade (Nakajimaet al., 1992), which results in calcium release from intracel-lular stores (Womack et al., 1988).
Despite the above considerations, our findings suggestthat the intrinsic SP system is inhibitory, whereas theextrinsic SP fibers may elicit stimulatory effects, because1) intrinsic SP fibers form symmetric and extrinsic axonterminals establish asymmetric membrane specializa-tions, which are believed to represent inhibitory andexcitatory connections, respectively (Eccles, 1964); 2) ithas been demonstrated that hippocampal SP neurons, atleast in the monkey, cocontain GABA (Seress and Leranth,1996), whereas supramammillary SP cells projecting tothe monkey hippocampus are not GABAergic neurons(Nitsch and Leranth, 1994); and 3) Mizumori et al. (1989)provided strong evidence of a direct excitatory projectionfrom the supramammillary area to the hippocampal forma-tion that mediates enhancement of perforant path-elicitedpopulation spikes.The distribution pattern of symmetric SP synapses in
the hippocampal formation indicates that the intrinsic SPneurons exert most of their action on the Ammon’s horn.Because the great majority of these SP terminals contactdendrites (Figs. 6b, 8a,b), they may be involved in den-dritic inhibitory processes. In addition to the inhibitoryrole that intrinsic SP neurons may play, they could beinvolved in disinhibitory functions by innervating otherinterneurons. One such example of a supposed disinhibi-tory connection appears to affect the electric activity of theentire Ammon’s horn, because the SP interneurons arehomogeneously distributed in all CA areas and are con-tacted by putatively inhibitory, intrinsic SP fibers (Fig. 10).On the other hand, in a very distinct area, the stratumradiatum of the CA3c subfield, intrinsic SP fibers formvery dense pericellular baskets on a large population ofimmunonegative interneurons (Fig. 4).Extrinsic SP axons that form asymmetric contacts seem
to have particular influence on the electric activity of theAmmon’s horn rather than on that of the dentate gyrus.This putative excitatory effect, in contrast to the moregeneral inhibitory SP action theorized above, seems to beassociated exclusively with the CA2–CA3a area, because,in other CA subfields, asymmetric SP synapses could notbe detected. The nearly complete lack of extrinsic SP fibersin the rat dentate gyrus is in sharp contrast to thesituation in other species, in which the bulk of extrinsic SPneurons can stimulate the granule cells (Gall and Selaw-ski, 1984; Ino et al., 1988; Nitsch and Leranth, 1994). Thisalso indicates that, in contrast to nonhuman primates inwhich the supramammillohippocampal-projecting SP neu-rons cocontain calretinin (Nitsch and Leranth, 1993), thesupramammillohippocampal calretinin-containing (Ma-gloczky et al., 1994) and SP-containing neurons in the ratmay represent two different populations of projecting cells,and only a small population of these neurons, whichproject to the CA2 area, contain both substances. This ideais substantiated by the facts that: 1) in the rat hippocam-pus, similar to the monkey (Nitsch and Leranth, 1993,1994), the extrinsic calretinin fibers target both the supra-granular layer and the CA2 subfield (Magloczky et al.,1994); and 2) in the supramammillary area, not all of theneurons that were retrogradely labeled from the hippocam-pus were immunoreactive for calretinin (Magloczky et al.,1994).
Fig. 7. a–c: Light micrographs of the CA1 region of the Ammon’shorn of a colchicine-treated (b) and a control rat (a,c). In a, a dense,homogeneous network of SP-containing axons is visible in the strataoriens (o.) and radiatum (r.). The SP innervation of the stratapyramidale (p.) and lacunosum moleculare (l-m.) is less extensive. c:Characteristic large axons at the border between the strata radiatum(r.) and lacunosum moleculare (l-m.). Despite the lack of colchicinetreatment, SP-immunoreactive cells are also visible (a, arrows). b:Heavily stained SP neurons in the CA1 region of a colchicine-treatedrat. Horizontally oriented cells are at the border between the alveus(a.) and the stratum oriens (o.), and multipolar neurons are visible inthe stratum oriens (o.) and adjacent to the pyramidal cell layer (p.).Scale bars 5 20 µm in a,b, 10 µm in c.
SP INNERVATION OF THE RAT HIPPOCAMPUS 53
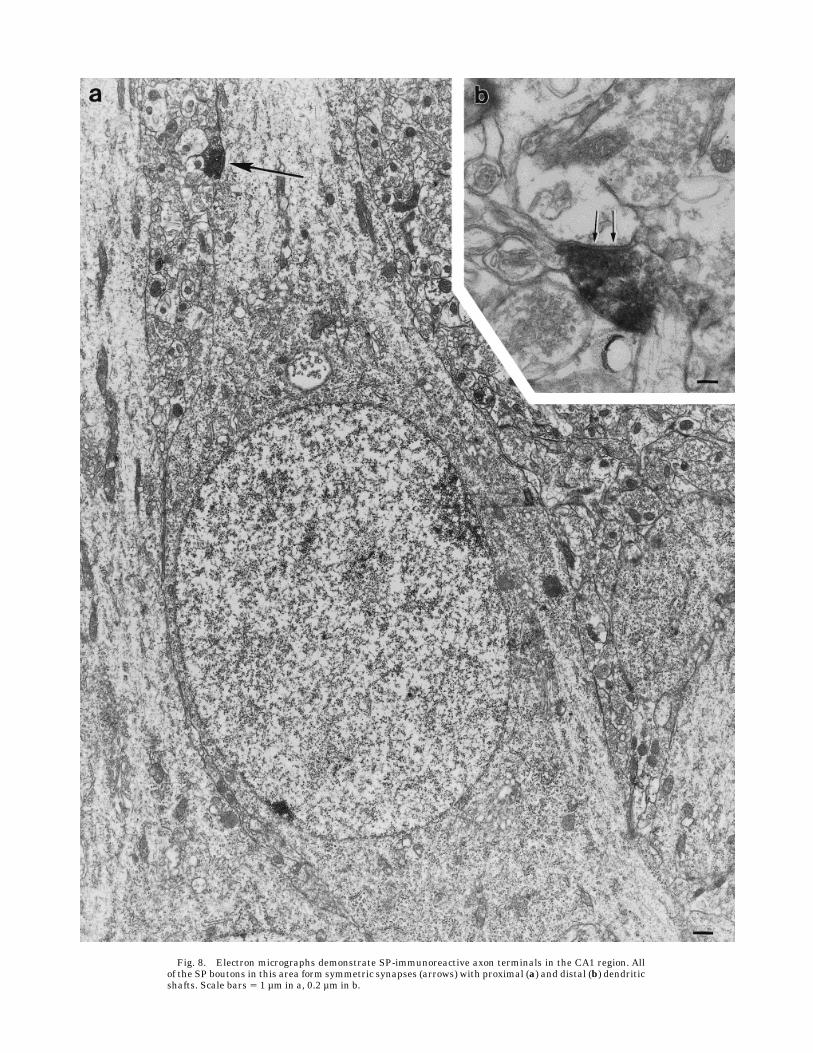
Fig. 8. Electron micrographs demonstrate SP-immunoreactive axon terminals in the CA1 region. Allof the SP boutons in this area form symmetric synapses (arrows) with proximal (a) and distal (b) dendriticshafts. Scale bars 5 1 µm in a, 0.2 µm in b.
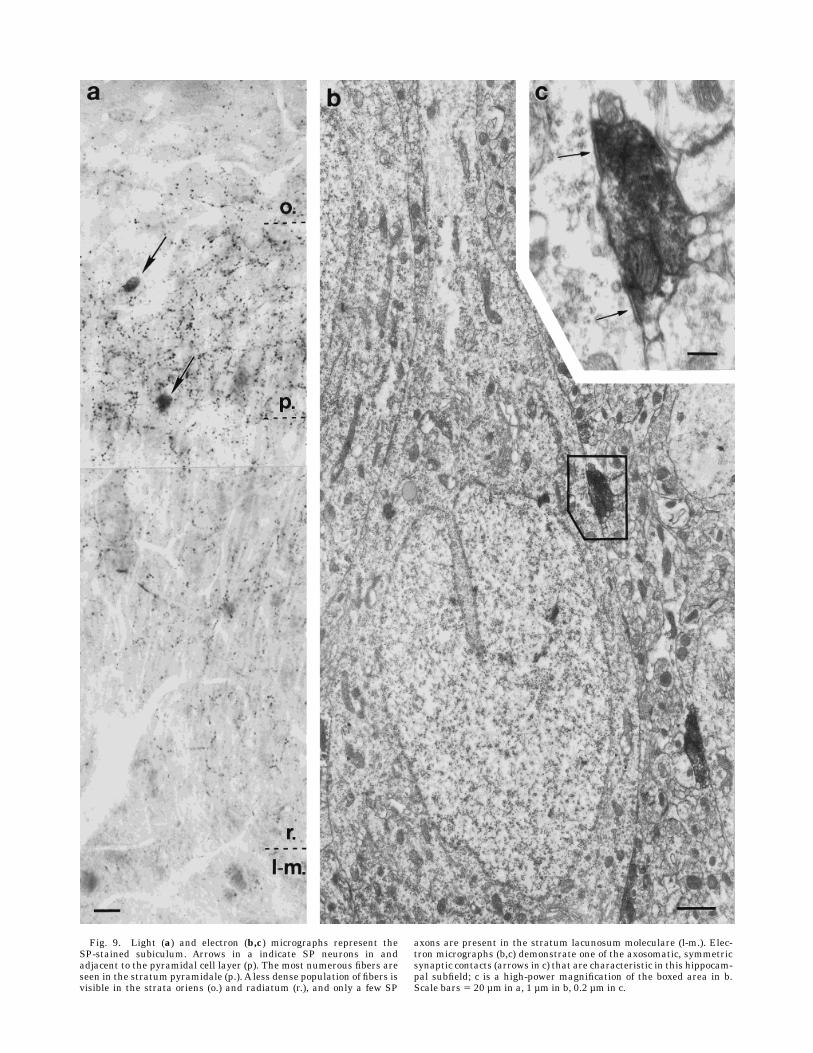
Fig. 9. Light (a) and electron (b,c) micrographs represent theSP-stained subiculum. Arrows in a indicate SP neurons in andadjacent to the pyramidal cell layer (p). The most numerous fibers areseen in the stratum pyramidale (p.). A less dense population of fibers isvisible in the strata oriens (o.) and radiatum (r.), and only a few SP
axons are present in the stratum lacunosum moleculare (l-m.). Elec-tron micrographs (b,c) demonstrate one of the axosomatic, symmetricsynaptic contacts (arrows in c) that are characteristic in this hippocam-pal subfield; c is a high-power magnification of the boxed area in b.Scale bars 5 20 µm in a, 1 µm in b, 0.2 µm in c.

Fig. 10. a,b: Electron micrographs demonstrate an axosomatic,symmetric, synaptic contact between an SP-immunoreactive neuronand an SP-containing bouton in the stratum oriens of the CA1 area.This type of homologous contact is characteristic in the stratum oriens
of all subfields of Ammon’s horn. b: High-power magnification of thesynaptic contact (arrows) seen in the boxed area in a. Scale bars 5 0.4µm in a, 0.2 µm in b.

ACKNOWLEDGMENTS
The authors thank Dr. S. Leeman for kindly providingthe SP antiserum and Dr. Sloviter for his contribution ofperforant path-stimulated rat brains. We are grateful toMarya Shanabrough for critically reading the paper.
LITERATURE CITED
Acsady, L., I. Katona, A.I. Gulyas, R.Y. Shigemoto, and T.F. Freund (1997)Immunostaining for substance P receptor labels GABAergic cells withdistinct termination patterns in the hippocampus. J. Comp. Neurol.378:320–336.
Bouras, C., P.G. Vallet, H. Dobrinov, S. de St.-Hilaire, and J. Constantinidis(1986) Substance P neuronal cell bodies in the human brain: Completemapping by immunohistofluorescence. Neurosci. Lett. 69:31–36.
Boyer, P.A., A. Trembleau, V. Leviel, and M. Arluision (1993) Effects ofintranigral injections of colchicine on the expression of some neuropep-tides in the rat forebrain: An immunohistochemical and in situ hybrid-ization study. Brain Res. Bull. 33:541–560.
Brene, S., N. Lindefors, M. Ballarin, and H. Person (1992) Kainic acid-mediated increase of preprotachykinin-Amessenger RNAexpression inthe rat hippocampus and a region-selective attenuation by dexametha-sone. Neuroscience 50:611–618.
Carraway, R., and S.E. Leeman (1979) The amino acid sequence of bovinehypothalamic substance P. Identity to substance P from colliculi andsmall intestine. J. Biol. Chem. 254:2944–2945.
Cortes, R., S. Ceccatelli, M. Schalling, and T. Hokfelt (1990) Differentialeffects of intracerebroventricular colchicine administration on theexpression of mRNAs for neuropeptides and neurotransmitter en-zymes, with special emphasis on galanin: An in situ hybridizationstudy. Synapse 6:369–391.
Cuello, A.C., and I. Kanazawa (1978) The distribution of substance Pimmunoreactive fibers in the rat central nervous system. J. Comp.Neurol. 178:129–156.
Dahlstrom, A. (1971) Effects of vinblastine and colchicine on monoaminecontaining neurons of the rat with special regard to the axoplasmictransport of amine granules. Acta Neuropathol. (Berlin) 5:226–237.
Dam, T.V., and R. Quirion (1986) Pharmacological characterization andautoradiographic localization of substance P receptors in guinea pigbrain. Peptides 7:855–864.
Dam, T.V., B. Martinelli, and R. Quirion (1990) Autoradiographic distribu-tion of brain neurokinin-1/substance P receptors using highly selectiveligand [3H]-[Sar9,Met(O2)11]-substance P. Brain Res. 531:333–337.
Davies, S., and C. Kohler (1985) The substance P innervation of the rathippocampal region. Anat. Embryol. 173:45–52.
de Lanerolle, C.N., M. Brines, A. Williamson, and H.J. Kim (1992)Neurotransmitters and their receptors in human temporal lobe epi-lepsy. In C.E. Ribac, C.M. Gall, and I. Mody (eds): The Dentate Gyrusand Its Role in Seizures. Amsterdam: Elsevier, pp. 235–250.
Del Fiacco, M., M.C. Levanti, M.L. Dessi, and G. Zucca (1987) The humanhippocampal formation and parahippocampal gyrus: Localization ofsubstance P-like immunoreactivity in newborn and adult postmortemtissue. Neuroscience 21:141–150.
Dodd, J., and J.S. Kelly (1981) The actions of cholecystokinin and relatedpeptides on pyramidal neurons of the mammalian hippocampus. BrainRes. 338:97–113.
Dreifuss, J.J., and M. Raggenbass (1986) Tachykinins and bombesin excitenonpyramidal neurons in rat hippocampus. J. Physiol. 379:417–428.
Eccles, J.C. (1964) The Physiology of Synapses. Heidelberg: Springer.Gall, C., and L. Selawski (1984) Supramammillary afferents to guinea pig
hippocampus contain substance P-like immunoreactivity. Neurosci.Lett. 51:171–176.
Gallagher, A.W., L.A. Chahl, and M. Lynch (1992) Distribution of substanceP-like immunoreactivity in guinea pig central nervous system. BrainRes. Bull. 29:199–207.
Herkenham, M. (1987) Mismatches between neurotransmitter and recep-tor localizations in brain: Observations and implications. Neuroscience23:1–38.
Holm, I.E., F.A. Geneser, and J. Zimmer (1993) Cholecystokinin-, enkepha-lin-, and substance P-like immunoreactivity in the dentate area,hippocampus and subiculum of the domestic pig. J. Comp. Neurol.331:310–325.
Hsu,M., L. Raine, andH. Fanger (1981) The use of avidin-biotin-peroxidasecomplex (ABC) in immunoperoxidase techniques: A comparison be-tween ABC and unlabeled antibody (peroxidase) procedures. J. Histo-chem. Cytochem. 29:577–590.
Huston, J.P., R.U. Hasenohrl, F. Boix, P. Gerhardt, and R.K.W. Schwarting(1993) Sequence-specific effects of neurokinin substance P on memory,reinforcement, and brain dopamine activity. Psychopharmacology 112:147–162.
Ino, T., K. Itoh, T. Sugimoto, T. Kaneko, H. Kamiya, and N. Mizuno (1988)The supramammillary region of the cat sends substance P-like immuno-reactive axons to the hippocampal formation and the entorhinal cortex.Neurosci. Lett. 90:259–264.
Iritani, S., M. Fujii, and K. Satoh (1989) The distribution of substance P inthe cerebral cortex and hippocampal formation: An immunohistochemi-cal study in the monkey and rat. Brain Res. Bull. 22:295–303.
Jakab, R.L., and C. Leranth (1991) Synaptology and origin of somatostatinfibers in the rat lateral septal area; convergent somatostatinergic andhippocampal inputs of somatospiny neurons. Brain Res. 565:123–134.
Jakab, R.L., L.-N. Nazrati, and P. Goldman-Rakic (1996) Distribution andneurochemical character of substance P receptor (SPR)-immunoreac-tive striatal neurons of the macaque monkey: Accumulation of SP fibersand SPR neurons and dendrites in ‘‘striatocapsules’’ encircling strio-somes. J. Comp. Neurol. 369:137–149.
King, J.C,. R.M. Lechan, G. Kugel, and E.L.P. Anthony (1983) Acrolein: Afixative for immunocytochemical localization of peptides in the centralnervous system. J. Histochem. Cytochem. 31:62–68.
Kosaka, T., H. Nagatsu, J.Y. Wu, and K. Hama (1986) Use of highconcentrations of glutaraldehyde for immunocytochemistry of transmit-ter-synthesizing enzymes in the central nervous system. Neuroscience18:975–990.
Lamour, Y., M.C. Senut, P. Dutar, and M.H. Bassant (1989) Neuropeptidesand septo-hippocampal neurons: Electrophysiological effects and distri-butions of immunoreactivity. Peptides 9:1351–1359.
Leranth, C., and R. Nitsch (1994) Hypothalamic substance P-containingafferents can filter the signal flow in the monkey hippocampal forma-tion: Morphological evidence for stimulation and feed-forward inhibi-tion of different populations of granule cells and indirect inhibition ofCA3 pyramidal neurons. J. Neurosci. 14:4079–4094.
Liu, H., J.L. Brown, L. Jasmin, J.E. Maggio, S.R. Vigna, P.W. Mantyh, andA.I. Basbaum (1994) Synaptic relationship between substance P andthe substance P receptor: Light and electron microscopic characteriza-tion of mismatch between neuropeptides and their receptors. Proc.Natl. Acad. Sci. USA 91:1009–1013.
Ljungdhal, Å., T. Hokfelt, and G. Nilson (1978) Distribution of substanceP-like immunoreactivity in the central nervous system of the rat. I. Cellbodies and nerve terminals. Neuroscience 3:861–943.
Maeno, H., H. Kiyama, and M. Tohyama (1993) Distribution of thesubstance P receptor in the central nervous system. Mol. Brain. Res.18:43–58.
Magloczky, Z., L. Acsady, and T.F. Freund (1994) Principal cells are thepostsynaptic targets of supramammillary afferents in the hippocampusof the rat. Hippocampus 4:322–334.
Mizumori, S.J.Y., B.L. McNaughton, and C.A. Barnes (1989) A comparisonof supramammillary and medial septal influences on hippocampal fieldpotentials and single-unit activity. J. Neurophysiol. 61:15–31.
Mohini, P., S.W. Bahouth, D.E. Brundish, and J.M. Musacchio (1985)Specific labeling of rat brain substance P receptor with [3H]physalae-min. J. Neurosci. 5:2078–2085.
Nakajima, Y., K. Tsuchida, M. Negishi, S. Ito, and S. Nakanishi (1992)Direct linkage of three tachykinin receptors to stimulation of bothphosphatidylinositol hydrolysis and cyclic AMP cascade in transfectedChinese hamster ovary cells. J. Biol. Chem. 267:2437–2442.
Nakaya, Y., T. Kaneko, R. Shigemoto, S. Nakanishi, and N. Mizuno (1994)Immunohistochemical localization of substance P receptor in the cen-tral nervous system of the adult rat. J. Comp. Neurol. 347:249–274.
Nicoll, R.A., C. Schenker, and S.E. Leeman (1980) Substance P as aneurotransmitter candidate. Annu. Rev. Neurosci. 3:227–268.
Nitsch, R., and C. Leranth (1993) Calretinin immunoreactivity in themonkey hippocampal formation. II. Intrinsic GABAergic and hypotha-lamic non-GABAergic systems: An experimental tracing and coexist-ence study. Neuroscience 55:797–812.
Nitsch, R., and C. Leranth (1994) Substance P-containing hypothalamicafferents to the monkey hippocampus:An immunocytochemical, tracingand coexistence study. Exp. Brain Res. 101:231–240.
SP INNERVATION OF THE RAT HIPPOCAMPUS 57

Peters, A., S.L. Palay, and H.F. Webster (1991) The Fine Structure of theNervous System: Neurons and Their Supporting Cells. Oxford: OxfordUniversity Press.
Peterson, P.M., and C.L. Shurlow (1992) Morphological evidence for asubstance P projection from medial septum to hippocampus. Peptides13:509–517.
Roberts, G.W., P.L. Woodhams, J.M. Polak, and T.J. Crow (1984) Distribu-tion of neuropeptides in the limbic system of the rat: The hippocampus.Neuroscience 11:35–77.
Sakamoto, N., J.-P. Michel, N. Kopp, M. Tohyama, and J. Pearson (1987)Substance P- and enkephalin-immunoreactive neurons in the hippocam-pus and related areas of the human infant brain. Neuroscience 22:801–811.
Schwarzer, C., and G. Sperk (1995) Hippocampal granule cells expressglutamic acid decarboxylase-67 after limbic seizures in the rat. Neuro-science 69:705–709.
Seress, L., and C. Leranth (1996) Distribution of substance P-immunoreac-tive neurons and fibers in themonkey hippocampal formation. Neurosci-ence 71:633–650.
Shults, C.W., R. Quirion, B. Chronwall, T.N. Chase, and L. O’Donohue(1984) A comparison of the anatomical distribution of substance P andsubstance P receptors in the rat nervous system. Peptides 5:1097–1128.
Shults, C.W., P. Johnston, and F.H. Gage (1987) Comparison of substanceK-like fibers and cells in the rat hippocampus. Brain Res. 426:290–296.
Sik, A., M. Penttonen, A. Ylinen, and G. Buzsaki (1995) Hippocampal CA1interneurons: An in vivo intracellular labeling study. J. Neurosci.15:6651–6665.
Sloviter, R.S. (1983) ‘‘Epileptic’’ brain damage in rats induced by sustainedelectrical stimulation of the perforant path. I. Acute electrophysiologi-cal and light microscopic studies. Brain Res. Bull. 10:675–697.
Sloviter, R.S., M.A. Dichter, T.L. Rachinsky, E. Dean, J.H. Goodman, A.L.Dollas, and D.L. Martin (1996) Basal expression and induction ofglutamate decarboxylase and GABA in excitatory granule cells of therat and monkey hippocampal dentate gyrus. J. Comp. Neurol. 373:593–618.
Sperk, G. (1994) Kainic acid seizures in the rat. Progr. Neurobiol. 42:1–32.
Sperk, G., J. Marksteiner, B. Gruber, R. Bellmann, M. Mahata, and M.Ortler (1992) Functional changes in neuropeptide Y- and somatostatin-containing neurons induced by limbic seizures in the rat. Neuroscience50:831–846.
Szeidemann, Z., R.L. Jakab, M. Shanabrough, and C. Leranth (1995)Extrinsic and intrinsic substance P innervation of the rat lateral septalarea calbindin cells. Neuroscience 69:1205–1221.
Vincent, S.R., and E.G. McGeer (1981) A substance P projection to thehippocampus. Brain Res. 215:349–351.
Vincent, S.R., H. Kimura, and E.G. McGeer (1981) Organization of sub-stance P fibers within the hippocampal formation demonstrated with abiotin-avidin immunoperoxidase technique. J. Comp. Neurol. 199:113–123.
Womack,M.D.,A.B.MacDermott, and T.M. Jessell (1988) Sensory transmit-ters regulate intracellular calcium in dorsal horn neurons. Nature334:351–353.
58 Z. BORHEGYI AND C. LERANTH