Embed Size (px)
Citation preview

Mahrez et al. Supplemental material
1
Supplemental Material Supplemental Methods RNA isolation and RT-qPCR RNA extraction and reverse transcription were performed as described previously (Alexandre et al., 2009). For spruce RT-qPCR, total RNA was extracted using the RNeasy plant mini kit (Qiagen). RNA was treated DNase I twice on-column (Qiagen) and in solution (Fermentas) according to the manufacturer's instructions. DNA-free RNA was reverse-transcribed using a RevertAid First Strand cDNA Synthesis Kit (Fermentas, Helsingborg, Sweden) according to the manufacturer’s recommendations. Aliquots of the generated cDNA were used as template for PCR with gene-specific primers (Supplemental Tab. S3). qPCR was performed using SYBR green (Fermentas) on an IQ5 multicolor Real time PCR cycler (BIO-RAD, PA, USA). qPCR reactions were performed in triplicate, gene expression levels were normalized to a PP2A control gene, and results were analyzed as described (Simon 2003). Chromatin Immunopreciptation and ChIP-seq Cross-linked chromatin immunoprecipitation (X-ChIP) assays were performed as described (Exner et al., 2009). Approximately 1.5 g of 10-day-old cross-linked whole seedlings were used. For Norway spruce cells, X-ChIP was performed on chromatin isolated as described (Rehrauer et al., 2010). Isolated chromatin was incubated with antibodies at 4°C with rotation overnight. Protein A Dynabeads (Invitrogen) were added and incubated at 4°C with rotation for 2 hours. The beads were washed twice with high salt buffer (500 mM NaCl, 0.1% SDS, 1% Triton X-100, 2 mM EDTA, 20 mM Tris-HCl, pH 8.0), twice with low salt buffer (150 mM NaCl, 0.1% SDS, 1% Triton X-100, 2 mM EDTA, 20 mM Tris-HCl, pH 8.0) and once with TE. Precipitated chromatin was de-crosslinked in elution buffer (1% SDS, 0.1 M NaHCO3) at 65°C overnight. DNA was extracted with chloroform followed by ethanol precipitation. Antibodies used for ChIP were anti-H3 (Upstate/Millipore, cat. no. 07-690), anti-H3K36me3 (Diagenode, Belgium, cat. no. pAb-058-050), anti-H3K36ac (ActiveMotif, cat. no. 39379), anti-H3K36me3 (Diagenode, cat. no. pAb-058-050) and non-immune IgG (Sigma-Aldrich, cat. no. I5006). Quantitative real-time PCR (qPCR) using SYBR green (Fermentas, Helsingborg, Sweden) and gene-specific primers (Supplemental Tab. S3) was performed on an IQ5 multicolor Real time PCR cycler (BIO-RAD, PA, USA). Results were normalized to input and presented as the enrichment level of the given modification over the level of histone H3. ChIP was performed in two biological replicates.
Native Chromatin Immunoprecipitation (N-ChIP) was performed following a slightly modified procedure of (Bernatavichute et al., 2008). Isolated nuclei were digested with Micrococcal Nuclease (New England BioLabs, NEB, Massachusetts). The supernatant, which contained mono- and dinucleosomes, was precleared using non-immune rabbit IgG (Sigma-Aldrich, cat. no. I5006) and Dynabeads Protein A (Invitrogen, Stockholm, Sweden). One tenth of the precleared chromatin preparation was kept as input control, and one quarter was used for each immunoprecipitation with 2.5 μg antibody and collected with Dynabeads Protein A (Invitrogen). After washing, DNA was extracted by phenol-chloroform extraction and ethanol-salt precipitation. Antibodies for ChIP-seq were anti-histone (Upstate/Millipore, cat. no. MAB3422), anti-H3K36ac (ActiveMotif, cat. no. 39379) and non-immune IgG (Sigma-Aldrich, cat. no. I5006).
Biological duplicates were used for sequencing. ChIP-seq libraries were generated using the Microplex Library Preparation kit (Diagenode) following the manufacturer's protocol using 1 ng of starting material. Sequencing was performed on an Illumina HiSeq2000 in 50-bp single-end mode.

Mahrez et al. Supplemental material
2
Immunostaining Immunostaining was performed according to (Jasencakova et al., 2000) with some modifications. Paraformaldehyde prefixed Arabidopsis root tips were washed in MTSB buffer (50 mM PIPES, 5 mM MgSO4, and 5 mM EGTA, pH 7.9) and digested for 10 min at 37°C with a PCP enzyme mixture [2.5% pectinase, 2.5% cellulase Onozuka R-10, and 2.5% pectolyase Y-23 (w/v) dissolved in MTSB]. After washing with MTSB, root tips were squashed on a glass slide using a cover slip, and immediately frozen in liquid nitrogen. The cover slip was gently removed, and the nuclei were covered with blocking solution (MTSB containing 3% BSA) for 1 hour at 4°C. Next, the samples were incubated for 1 h at 4°C with anti-H3K36ac (39379 ActiveMotif) primary antibodies in MTSB (containing 1% BSA and 0.1% Tween 20) and kept in a humid chamber overnight. After 3 times washing with MTSB for 5 min each, the slides were covered for 2 h at 4°C with Rhodamine-conjugated anti-rabbit IgG antibody (Thermo Scientific) diluted in MTSB containing 1% BSA and 0.1% Tween 20. The slides were gently washed with MTSB, and the DNA was counterstained with 1 μg/ml of DAPI in mounting medium (Vectashield, Vector Labs). Fluorescence signal detection and documentation was performed with a Leica DMI 4000 microscope. Images were merged and processed using Photoshop and ImageJ. Sequence alignment of H3 amino-terminal tails Amino acid sequences of H3 from representative eukaryotes were selected using PSI-BLAST searches. Sequences were aligned using the ClustalW multiple sequence alignment program implemented in MEGA5 (Tamura et al., 2011). Visualization of the amino acid alignment was performed with GenDoc.
Supplemental References
Alexandre C, Möller-Steinbach Y, Schönrock N, Gruissem W, Hennig L (2009) Arabidopsis MSI1 is required for negative regulation of the response to drought stress. Mol Plant 2: 675-687
Bernatavichute YV, Zhang X, Cokus S, Pellegrini M, Jacobsen SE (2008) Genome-wide association of histone H3 lysine nine methylation with CHG DNA methylation in Arabidopsis thaliana. PLoS One 3: e3156
Exner V, Aichinger E, Shu H, Wildhaber T, Alfarano P, Caflisch A, Gruissem W, Köhler C, Hennig L (2009) The chromodomain of LIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development. PLoS ONE 4: e5335
Garcia BA, Hake SB, Diaz RL, Kauer M, Morris SA, Recht J, Shabanowitz J, Mishra N, Strahl BD, Allis CD, Hunt DF (2007) Organismal differences in post-translational modifications in histones H3 and H4. J Biol Chem 282: 7641-7655
Huang H, Sabari BR, Garcia BA, Allis CD, Zhao Y (2014) SnapShot: histone modifications. Cell 159: 458-458.e1
Jasencakova Z, Meister A, Walter J, Turner BM, Schubert I (2000) Histone H4 acetylation of euchromatin and heterochromatin is cell cycle dependent and correlated with replication rather than with transcription. Plant Cell 12: 2087-2100
Johnson L, Mollah S, Garcia BA, Muratore TL, Shabanowitz J, Hunt DF, Jacobsen SE (2004) Mass spectrometry analysis of Arabidopsis histone H3 reveals distinct combinations of post-translational modifications. Nucleic Acids Res 32: 6511-6518
Liu H, Galka M, Iberg A, Wang Z, Li L, Voss C, Jiang X, Lajoie G, Huang Z, Bedford MT, Li SS (2010) Systematic identification of methyllysine-driven interactions for histone and nonhistone targets. J Proteome Res 9: 5827-5836

Mahrez et al. Supplemental material
3
Morris SA, Rao B, Garcia BA, Hake SB, Diaz RL, Shabanowitz J, Hunt DF, Allis CD, Lieb JD, Strahl BD (2007) Identification of histone H3 lysine 36 acetylation as a highly conserved histone modification. J Biol Chem 282: 7632-7640
Oh S, Zhang H, Ludwig P, van Nocker S (2004) A mechanism related to the yeast transcriptional regulator Paf1c is required for expression of the Arabidopsis FLC/MAF MADS box gene family. Plant Cell 16: 2940-2953
Rehrauer H, Aquino C, Gruissem W, Henz SR, Hilson P, Laubinger S, Naouar N, Patrignani A, Rombauts S, Shu H, Van de PeY, Vuylsteke M, Weigel D, Zeller G, Hennig L (2010) AGRONOMICS1: A new resource for Arabidopsis transcriptome profiling. Plant Physiol 152: 487-499
Simon P (2003) Q-Gene: processing quantitative real-time RT-PCR data. Bioinformatics 19: 1439-1440
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28: 2731-2739
Tweedie-Cullen RY, Reck JM, Mansuy IM (2009) Comprehensive mapping of post-translational modifications on synaptic, nuclear, and histone proteins in the adult mouse brain. J Proteome Res 8: 4966-4982
Zhang K, Sridhar VV, Zhu J, Kapoor A, Zhu JK (2007) Distinctive core histone post-translational modification patterns in Arabidopsis thaliana. PLoS One 2
Zilberman D, Coleman-Derr D, Ballinger T, Henikoff S (2008) Histone H2A.Z and DNA methylation are mutually antagonistic chromatin marks. Nature 456: 125-129
Mahrez et al. Supplemental material
4
Supplemental Figures
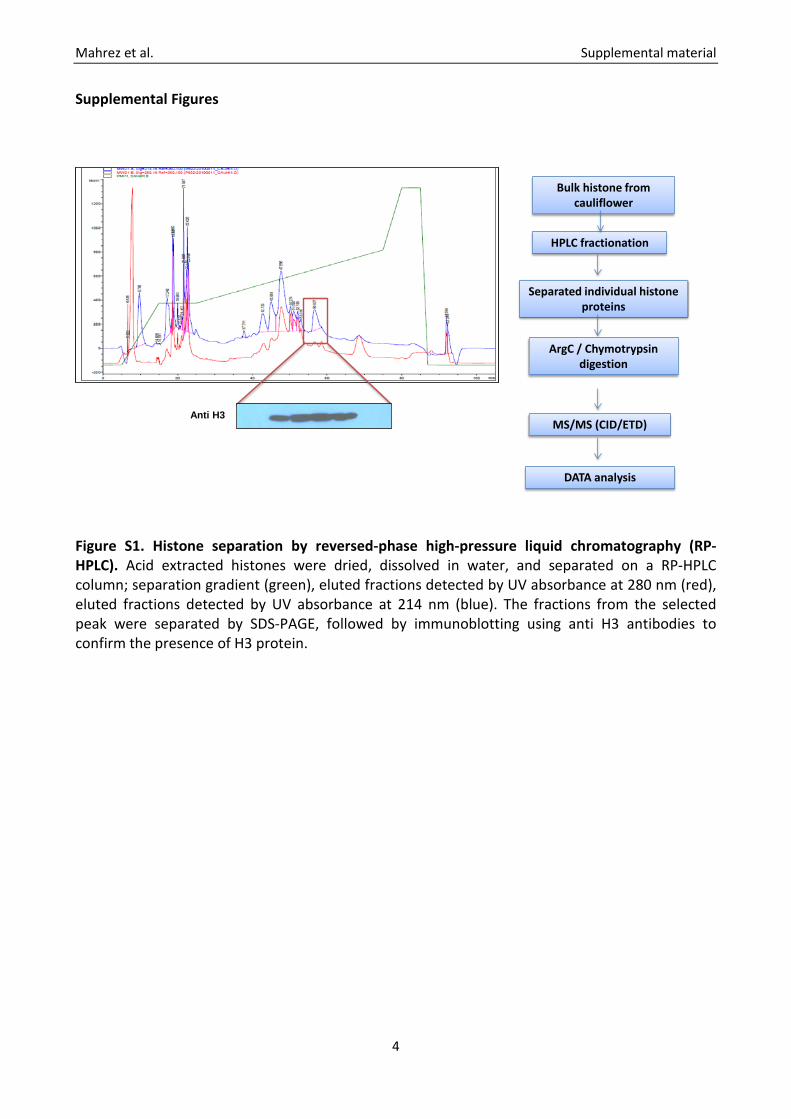
Figure S1. Histone separation by reversed-phase high-pressure liquid chromatography (RP-HPLC). Acid extracted histones were dried, dissolved in water, and separated on a RP-HPLC column; separation gradient (green), eluted fractions detected by UV absorbance at 280 nm (red), eluted fractions detected by UV absorbance at 214 nm (blue). The fractions from the selected peak were separated by SDS-PAGE, followed by immunoblotting using anti H3 antibodies to confirm the presence of H3 protein.
Anti H3
Bulk histone from cauliflower
HPLC fractionation
Separated individual histone proteins
ArgC / Chymotrypsin digestion
MS/MS (CID/ETD)
DATA analysis
Mahrez et al. Supplemental material
5
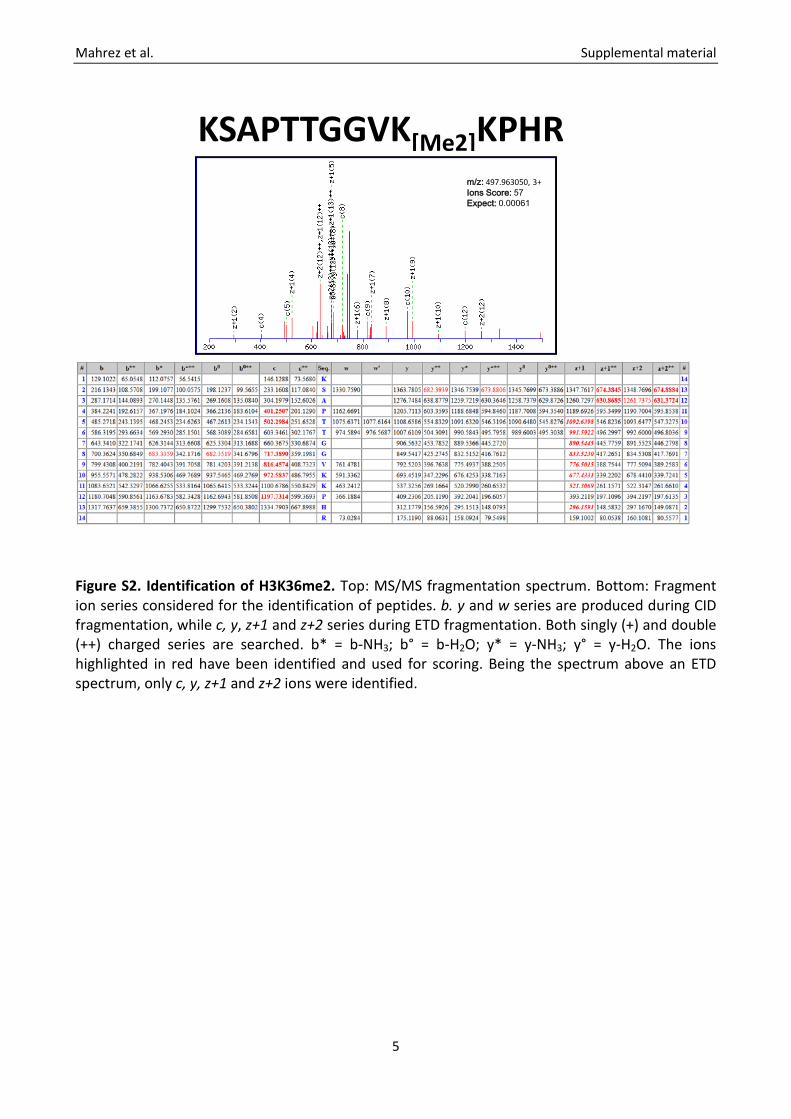
Figure S2. Identification of H3K36me2. Top: MS/MS fragmentation spectrum. Bottom: Fragment ion series considered for the identification of peptides. b. y and w series are produced during CID fragmentation, while c, y, z+1 and z+2 series during ETD fragmentation. Both singly (+) and double (++) charged series are searched. b* = b-NH3; b° = b-H2O; y* = y-NH3; y° = y-H2O. The ions highlighted in red have been identified and used for scoring. Being the spectrum above an ETD spectrum, only c, y, z+1 and z+2 ions were identified.
KSAPTTGGVK[Me2]KPHRm/z: 497.963050, 3+Ions Score: 57 Expect: 0.00061

Mahrez et al. Supplemental material
6
Figure S3. Identification of H3K36me3. Top: MS/MS fragmentation spectrum. Bottom: Fragment ion series considered for the identification of peptides. b. y and w series are produced during CID fragmentation, while c, y, z+1 and z+2 series during ETD fragmentation. Both singly (+) and double (++) charged series are searched. b* = b-NH3; b° = b-H2O; y* = y-NH3; y° = y-H2O. The ions highlighted in red have been identified and used for scoring. Being the spectrum above an ETD spectrum, only c, y, z+1 and z+2 ions were identified.
SAPTTGGVK[Me3]KPHRm/z: 345.204450 ,4+Ions Score: 54 Expect: 0.0011
Mahrez et al. Supplemental material
7
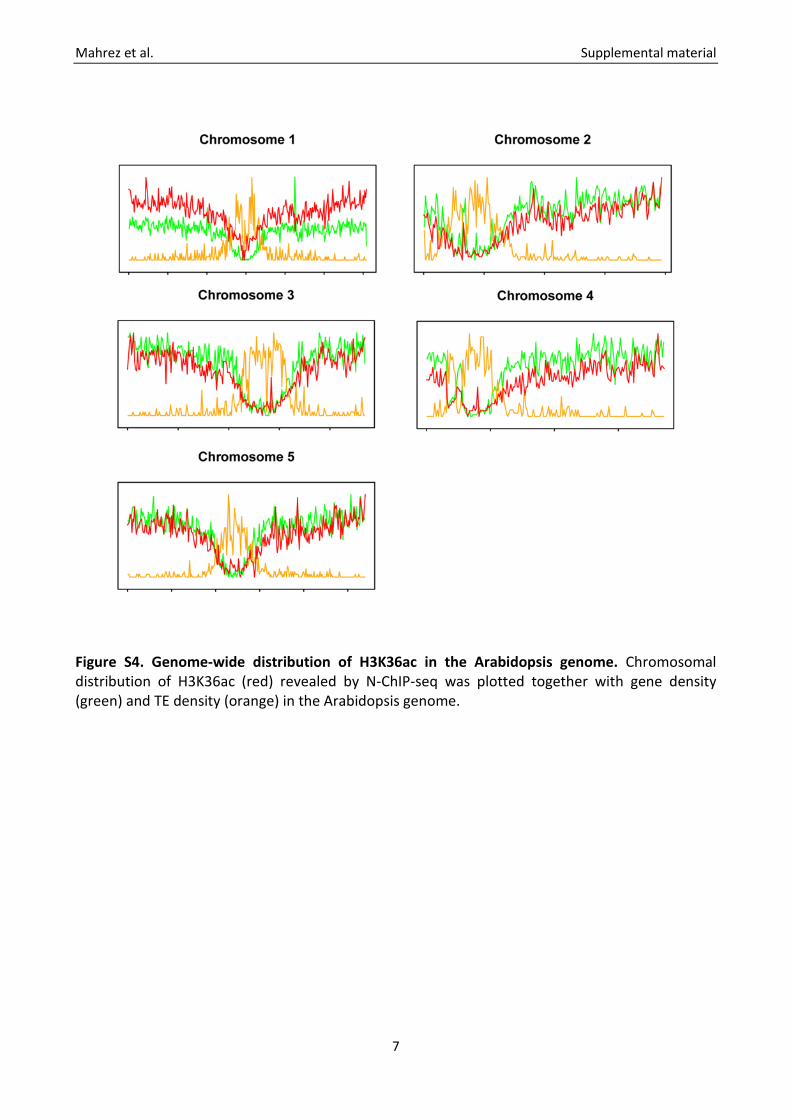
Figure S4. Genome-wide distribution of H3K36ac in the Arabidopsis genome. Chromosomal distribution of H3K36ac (red) revealed by N-ChIP-seq was plotted together with gene density (green) and TE density (orange) in the Arabidopsis genome.
Mahrez et al. Supplemental material
8
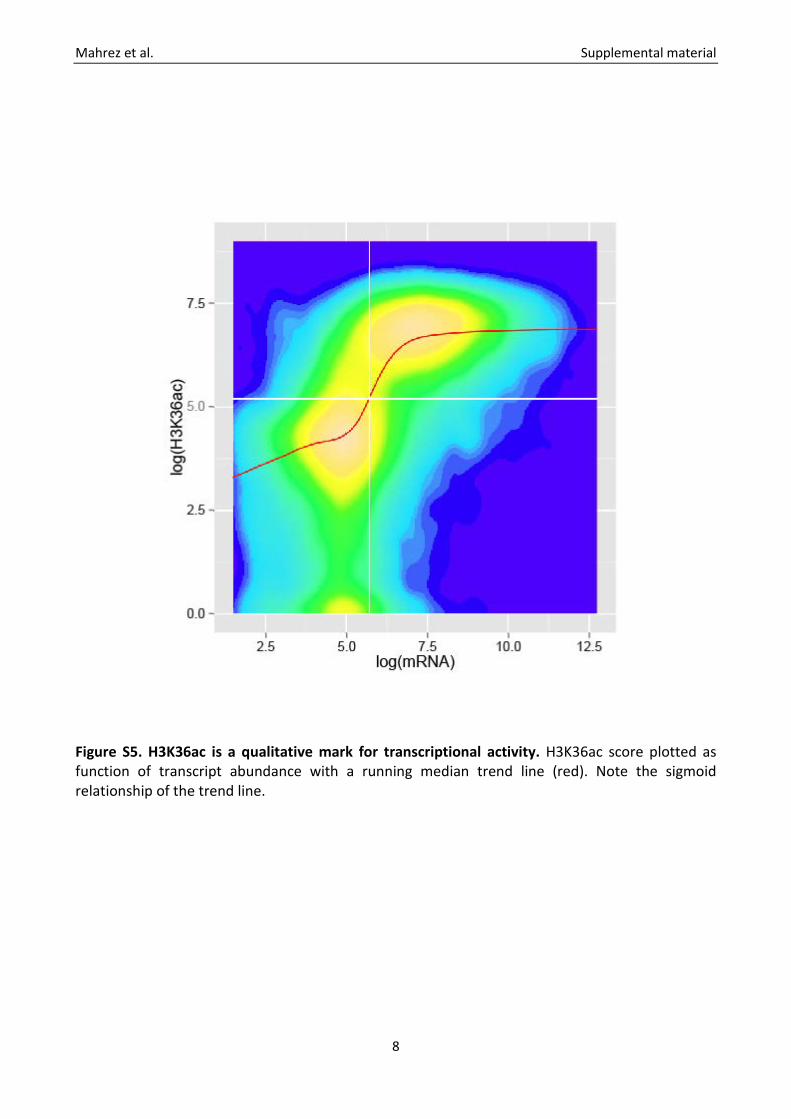
Figure S5. H3K36ac is a qualitative mark for transcriptional activity. H3K36ac score plotted as function of transcript abundance with a running median trend line (red). Note the sigmoid relationship of the trend line.
Mahrez et al. Supplemental material
9
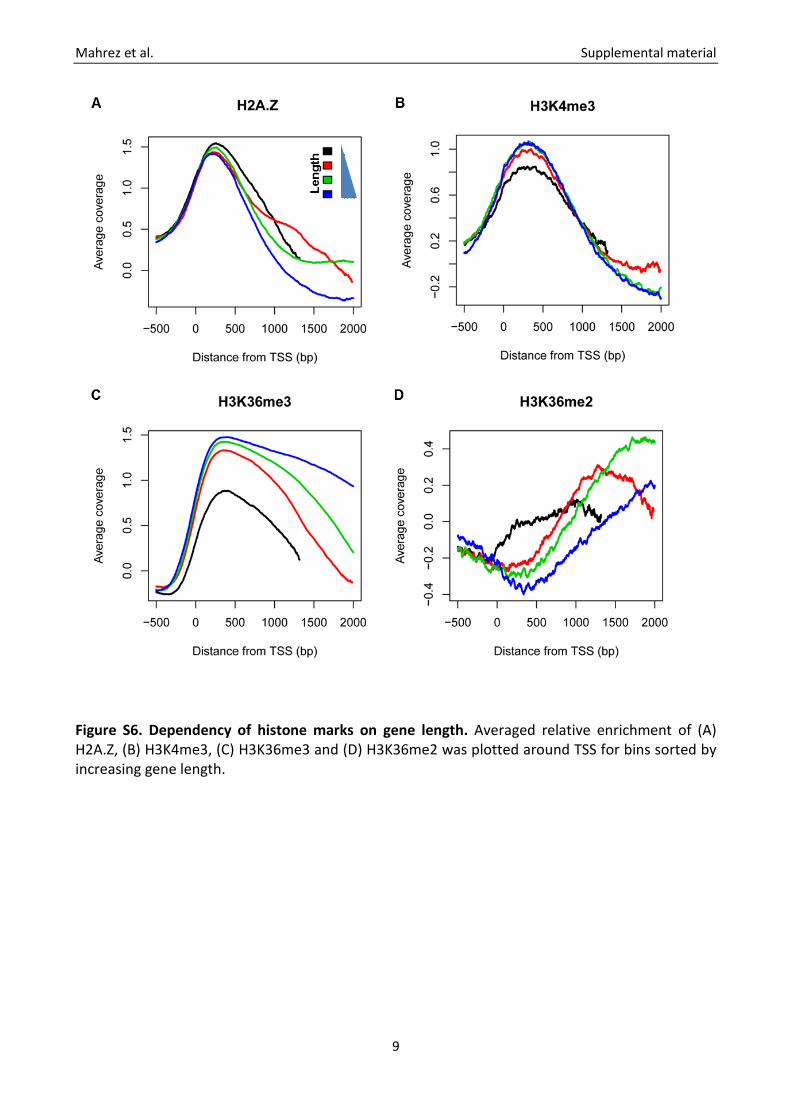
Figure S6. Dependency of histone marks on gene length. Averaged relative enrichment of (A) H2A.Z, (B) H3K4me3, (C) H3K36me3 and (D) H3K36me2 was plotted around TSS for bins sorted by increasing gene length.
Mahrez et al. Supplemental material
10
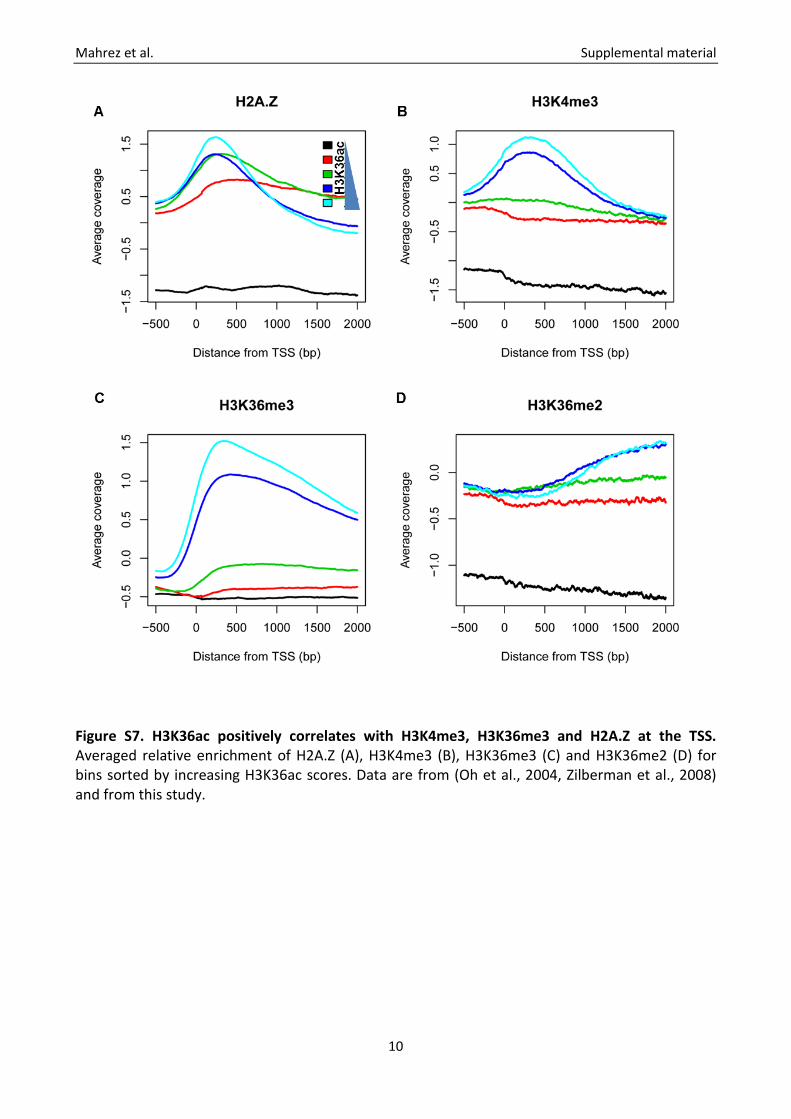
Figure S7. H3K36ac positively correlates with H3K4me3, H3K36me3 and H2A.Z at the TSS. Averaged relative enrichment of H2A.Z (A), H3K4me3 (B), H3K36me3 (C) and H3K36me2 (D) for bins sorted by increasing H3K36ac scores. Data are from (Oh et al., 2004, Zilberman et al., 2008) and from this study.
Mahrez et al. Supplemental material
11
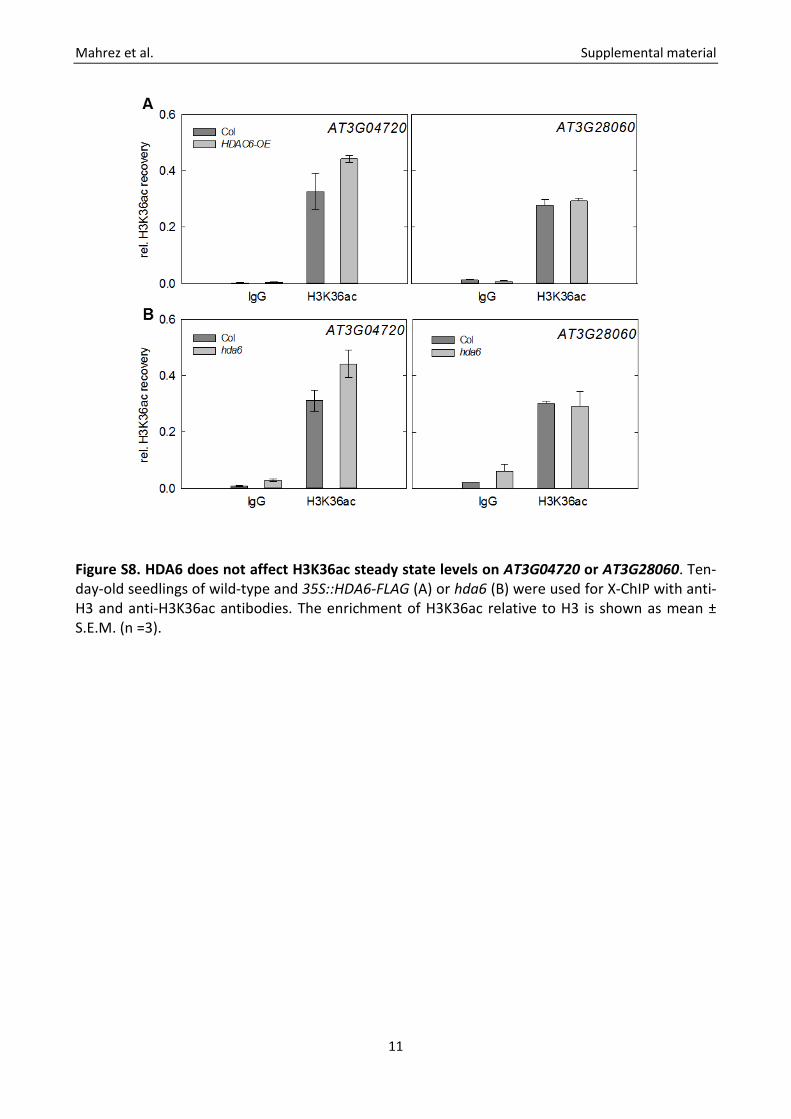
Figure S8. HDA6 does not affect H3K36ac steady state levels on AT3G04720 or AT3G28060. Ten-day-old seedlings of wild-type and 35S::HDA6-FLAG (A) or hda6 (B) were used for X-ChIP with anti-H3 and anti-H3K36ac antibodies. The enrichment of H3K36ac relative to H3 is shown as mean ± S.E.M. (n =3).
Mahrez et al. Supplemental material
12
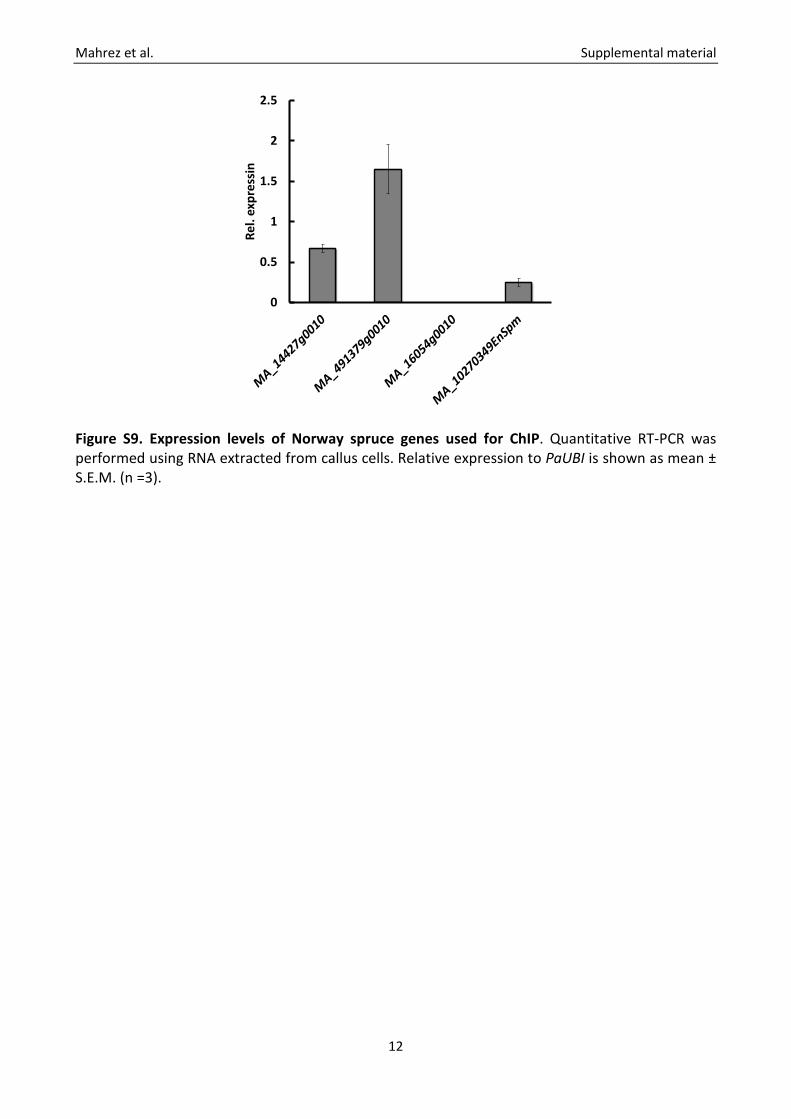
Figure S9. Expression levels of Norway spruce genes used for ChIP. Quantitative RT-PCR was performed using RNA extracted from callus cells. Relative expression to PaUBI is shown as mean ± S.E.M. (n =3).
0
0.5
1
1.5
2
2.5
Rel.
expr
essi
n
Mahrez et al. Supplemental material
13
Supplemental Tables
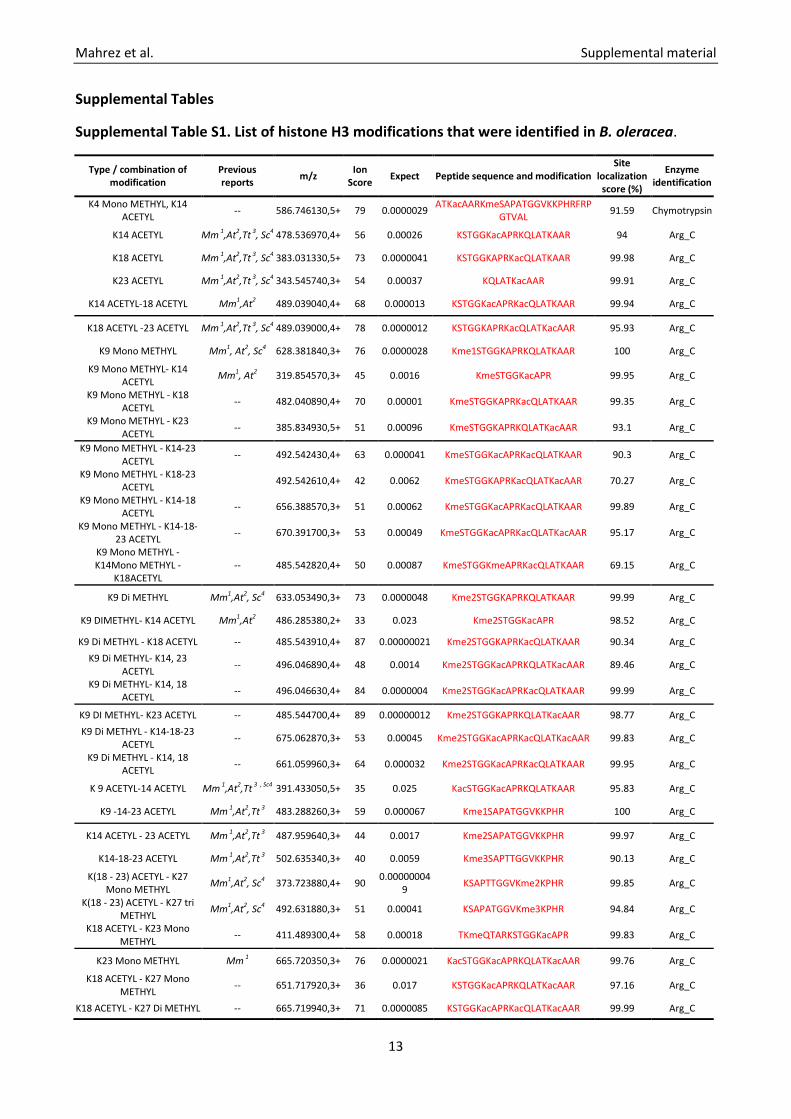
Supplemental Table S1. List of histone H3 modifications that were identified in B. oleracea.
Type / combination of modification
Previous reports m/z Ion
Score Expect Peptide sequence and modification Site
localization score (%)
Enzyme identification
K4 Mono METHYL, K14 ACETYL -- 586.746130,5+ 79 0.0000029 ATKacAARKmeSAPATGGVKKPHRFRP
GTVAL 91.59 Chymotrypsin
K14 ACETYL Mm 1,At2,Tt 3, Sc4 478.536970,4+ 56 0.00026 KSTGGKacAPRKQLATKAAR 94 Arg_C
K18 ACETYL Mm 1,At2,Tt 3, Sc4 383.031330,5+ 73 0.0000041 KSTGGKAPRKacQLATKAAR 99.98 Arg_C
K23 ACETYL Mm 1,At2,Tt 3, Sc4 343.545740,3+ 54 0.00037 KQLATKacAAR 99.91 Arg_C
K14 ACETYL-18 ACETYL Mm1,At2 489.039040,4+ 68 0.000013 KSTGGKacAPRKacQLATKAAR 99.94 Arg_C
K18 ACETYL -23 ACETYL Mm 1,At2,Tt 3, Sc4 489.039000,4+ 78 0.0000012 KSTGGKAPRKacQLATKacAAR 95.93 Arg_C
K9 Mono METHYL Mm1, At2, Sc4 628.381840,3+ 76 0.0000028 Kme1STGGKAPRKQLATKAAR 100 Arg_C
K9 Mono METHYL- K14 ACETYL Mm1, At2 319.854570,3+ 45 0.0016 KmeSTGGKacAPR 99.95 Arg_C
K9 Mono METHYL - K18 ACETYL -- 482.040890,4+ 70 0.00001 KmeSTGGKAPRKacQLATKAAR 99.35 Arg_C
K9 Mono METHYL - K23 ACETYL -- 385.834930,5+ 51 0.00096 KmeSTGGKAPRKQLATKacAAR 93.1 Arg_C
K9 Mono METHYL - K14-23 ACETYL -- 492.542430,4+ 63 0.000041 KmeSTGGKacAPRKacQLATKAAR 90.3 Arg_C
K9 Mono METHYL - K18-23 ACETYL 492.542610,4+ 42 0.0062 KmeSTGGKAPRKacQLATKacAAR 70.27 Arg_C
K9 Mono METHYL - K14-18 ACETYL -- 656.388570,3+ 51 0.00062 KmeSTGGKacAPRKacQLATKAAR 99.89 Arg_C
K9 Mono METHYL - K14-18-23 ACETYL -- 670.391700,3+ 53 0.00049 KmeSTGGKacAPRKacQLATKacAAR 95.17 Arg_C
K9 Mono METHYL - K14Mono METHYL -
K18ACETYL -- 485.542820,4+ 50 0.00087 KmeSTGGKmeAPRKacQLATKAAR 69.15 Arg_C
K9 Di METHYL Mm1,At2, Sc4 633.053490,3+ 73 0.0000048 Kme2STGGKAPRKQLATKAAR 99.99 Arg_C
K9 DIMETHYL- K14 ACETYL Mm1,At2 486.285380,2+ 33 0.023 Kme2STGGKacAPR 98.52 Arg_C
K9 Di METHYL - K18 ACETYL -- 485.543910,4+ 87 0.00000021 Kme2STGGKAPRKacQLATKAAR 90.34 Arg_C K9 Di METHYL- K14, 23
ACETYL -- 496.046890,4+ 48 0.0014 Kme2STGGKacAPRKQLATKacAAR 89.46 Arg_C
K9 Di METHYL- K14, 18 ACETYL -- 496.046630,4+ 84 0.0000004 Kme2STGGKacAPRKacQLATKAAR 99.99 Arg_C
K9 DI METHYL- K23 ACETYL -- 485.544700,4+ 89 0.00000012 Kme2STGGKAPRKQLATKacAAR 98.77 Arg_C K9 Di METHYL - K14-18-23
ACETYL -- 675.062870,3+ 53 0.00045 Kme2STGGKacAPRKacQLATKacAAR 99.83 Arg_C
K9 Di METHYL - K14, 18 ACETYL -- 661.059960,3+ 64 0.000032 Kme2STGGKacAPRKacQLATKAAR 99.95 Arg_C
K 9 ACETYL-14 ACETYL Mm 1,At2,Tt 3 , Sc4 391.433050,5+ 35 0.025 KacSTGGKacAPRKQLATKAAR 95.83 Arg_C
K9 -14-23 ACETYL Mm 1,At2,Tt 3 483.288260,3+ 59 0.000067 Kme1SAPATGGVKKPHR 100 Arg_C
K14 ACETYL - 23 ACETYL Mm 1,At2,Tt 3 487.959640,3+ 44 0.0017 Kme2SAPATGGVKKPHR 99.97 Arg_C
K14-18-23 ACETYL Mm 1,At2,Tt 3 502.635340,3+ 40 0.0059 Kme3SAPTTGGVKKPHR 90.13 Arg_C
K(18 - 23) ACETYL - K27 Mono METHYL Mm1,At2, Sc4 373.723880,4+ 90 0.00000004
9 KSAPTTGGVKme2KPHR 99.85 Arg_C
K(18 - 23) ACETYL - K27 tri METHYL Mm1,At2, Sc4 492.631880,3+ 51 0.00041 KSAPATGGVKme3KPHR 94.84 Arg_C
K18 ACETYL - K23 Mono METHYL -- 411.489300,4+ 58 0.00018 TKmeQTARKSTGGKacAPR 99.83 Arg_C
K23 Mono METHYL Mm 1 665.720350,3+ 76 0.0000021 KacSTGGKacAPRKQLATKacAAR 99.76 Arg_C
K18 ACETYL - K27 Mono METHYL -- 651.717920,3+ 36 0.017 KSTGGKacAPRKQLATKacAAR 97.16 Arg_C
K18 ACETYL - K27 Di METHYL -- 665.719940,3+ 71 0.0000085 KSTGGKacAPRKacQLATKacAAR 99.99 Arg_C
Mahrez et al. Supplemental material
14
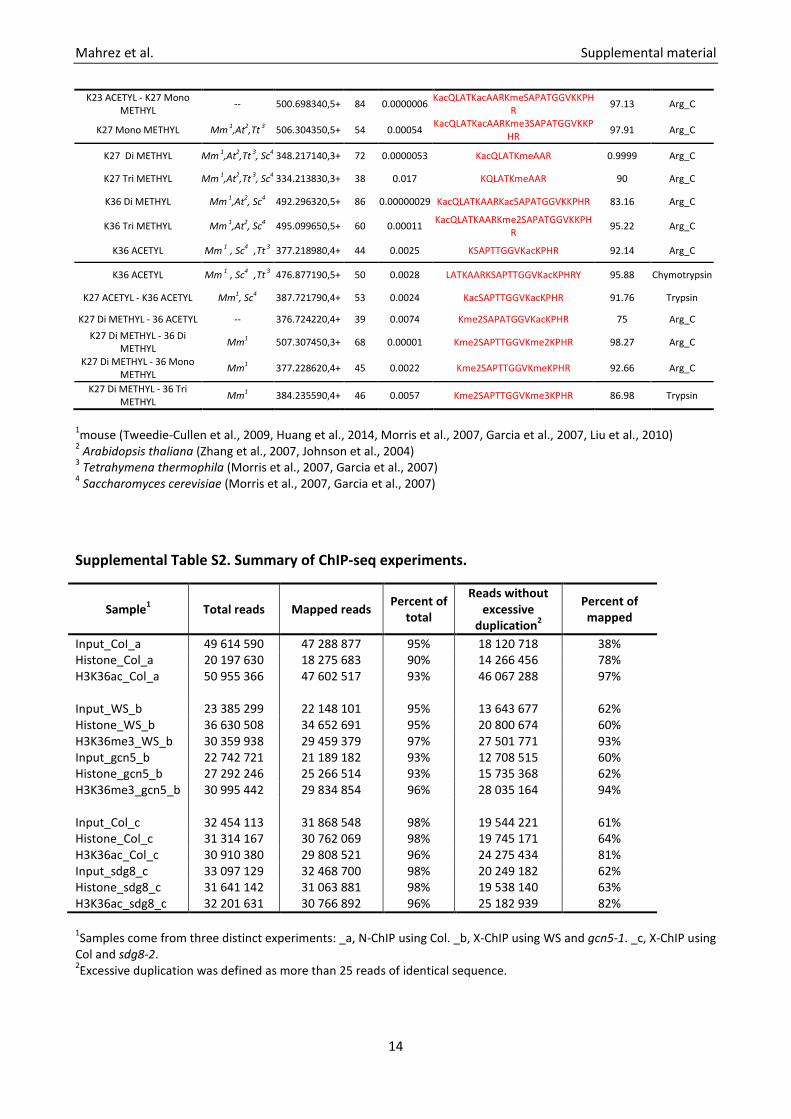
K23 ACETYL - K27 Mono METHYL -- 500.698340,5+ 84 0.0000006 KacQLATKacAARKmeSAPATGGVKKPH
R 97.13 Arg_C
K27 Mono METHYL Mm 1,At2,Tt 3 506.304350,5+ 54 0.00054 KacQLATKacAARKme3SAPATGGVKKPHR 97.91 Arg_C
K27 Di METHYL Mm 1,At2,Tt 3, Sc4 348.217140,3+ 72 0.0000053 KacQLATKmeAAR 0.9999 Arg_C
K27 Tri METHYL Mm 1,At2,Tt 3, Sc4 334.213830,3+ 38 0.017 KQLATKmeAAR 90 Arg_C
K36 Di METHYL Mm 1,At2, Sc4 492.296320,5+ 86 0.00000029 KacQLATKAARKacSAPATGGVKKPHR 83.16 Arg_C
K36 Tri METHYL Mm 1,At2, Sc4 495.099650,5+ 60 0.00011 KacQLATKAARKme2SAPATGGVKKPHR 95.22 Arg_C
K36 ACETYL Mm 1 , Sc4 ,Tt 3 377.218980,4+ 44 0.0025 KSAPTTGGVKacKPHR 92.14 Arg_C
K36 ACETYL Mm 1 , Sc4 ,Tt 3 476.877190,5+ 50 0.0028 LATKAARKSAPTTGGVKacKPHRY 95.88 Chymotrypsin
K27 ACETYL - K36 ACETYL Mm1, Sc4 387.721790,4+ 53 0.0024 KacSAPTTGGVKacKPHR 91.76 Trypsin
K27 Di METHYL - 36 ACETYL -- 376.724220,4+ 39 0.0074 Kme2SAPATGGVKacKPHR 75 Arg_C K27 Di METHYL - 36 Di
METHYL Mm1 507.307450,3+ 68 0.00001 Kme2SAPTTGGVKme2KPHR 98.27 Arg_C
K27 Di METHYL - 36 Mono METHYL Mm1 377.228620,4+ 45 0.0022 Kme2SAPTTGGVKmeKPHR 92.66 Arg_C
K27 Di METHYL - 36 Tri METHYL Mm1 384.235590,4+ 46 0.0057 Kme2SAPTTGGVKme3KPHR 86.98 Trypsin
1mouse (Tweedie-Cullen et al., 2009, Huang et al., 2014, Morris et al., 2007, Garcia et al., 2007, Liu et al., 2010) 2 Arabidopsis thaliana (Zhang et al., 2007, Johnson et al., 2004) 3 Tetrahymena thermophila (Morris et al., 2007, Garcia et al., 2007) 4 Saccharomyces cerevisiae (Morris et al., 2007, Garcia et al., 2007) Supplemental Table S2. Summary of ChIP-seq experiments.
Sample1 Total reads Mapped reads Percent of total
Reads without excessive
duplication2
Percent of mapped
Input_CoI_a 49 614 590 47 288 877 95% 18 120 718 38% Histone_Col_a 20 197 630 18 275 683 90% 14 266 456 78% H3K36ac_Col_a 50 955 366 47 602 517 93% 46 067 288 97% Input_WS_b 23 385 299 22 148 101 95% 13 643 677 62% Histone_WS_b 36 630 508 34 652 691 95% 20 800 674 60% H3K36me3_WS_b 30 359 938 29 459 379 97% 27 501 771 93% Input_gcn5_b 22 742 721 21 189 182 93% 12 708 515 60% Histone_gcn5_b 27 292 246 25 266 514 93% 15 735 368 62% H3K36me3_gcn5_b 30 995 442 29 834 854 96% 28 035 164 94% Input_Col_c 32 454 113 31 868 548 98% 19 544 221 61% Histone_Col_c 31 314 167 30 762 069 98% 19 745 171 64% H3K36ac_Col_c 30 910 380 29 808 521 96% 24 275 434 81% Input_sdg8_c 33 097 129 32 468 700 98% 20 249 182 62% Histone_sdg8_c 31 641 142 31 063 881 98% 19 538 140 63% H3K36ac_sdg8_c 32 201 631 30 766 892 96% 25 182 939 82% 1Samples come from three distinct experiments: _a, N-ChIP using Col. _b, X-ChIP using WS and gcn5-1. _c, X-ChIP using Col and sdg8-2.
2Excessive duplication was defined as more than 25 reads of identical sequence.

Mahrez et al. Supplemental material
15
Supplemental Table S3. Lists of primers used.
Gene-specific primers used for RT-qPCR
Gene /Alias Primer ID/ Primer sequence
AT2G10930, PP2A DK053: TCTTAATTTGAATCGGTTACTAGG DK054: CATACATAACAATTTCACAAGCA
AT5G09810, Actin7 LH1718: GGAACTGGAATGGTGAAGGCTG LH1719: CGATTGGATACTTCAGAGTGAGGA
AT4G03740, Ta2 LH1252: ATACTCTCCACTTCCCGTTTTTCTTTTTA LH1253: AAACGATGCGTTGGGATAGGTC
PaUBI LH3602: TTCAGAGAGCTTTTCGTTCAAAG LH3603: GTGCAGGGTAGATTCCTTTTGTA
MA_14427g0010 LH3604: CCAAGATCCAGGACAAGGAA LH3605: TTCCGGTAAGGGTTTTGACA
MA_491379g0010 LH3608: TGCCGATTCCTTCTATCGTT LH3612: TTGAGAGGTTCGAGGACGAG
MA_16054g0010 LH3617: CCCTCCATTCCTCAATTCAA LH3616: GTTGGAGAGCCGATGAAATC
MA_10270349EnSpm LH3622: ATGTGAAAGGGGCCAACAT LH3623: CCAGAAACCCTCCCATATCA
List of gene-specific primers used for ChIP-qPCR assays.
AT3G04720_1st region LH1960: AGCAATAAGCACTCACGGCTCTCA LH1961: TCCGACCAACAACTGTCAGAGCAA
AT3G04720_2nd region LH2004: TGGTCCACTATTCTCACAGTTAC LH2005: TGTTACCATTTACCAACGTCAAC
AT3G04720_3rd region LH2006: TACGAAACATATATCCCTCGACAATAC LH2007: GCCATTGATCGCGTTTGATATT
AT3G28060_1st region LH1666: ATTCTGTCAAAGAGACGGCGGGAT LH1667: ATAGAGTTGCCACAAGTGCACCCA
AT3G28060_2nd region LH2008: GTCTGCGTTTCAATCTCTTGTG LH2009: GCGACGATGCATATCAACATAAT
AT3G28060_3rd region LH2010: TAGCGCTTTAGCCAAATCCA LH2011: AAGGTCGTTGCCTCCATTAC
MA_14427g0010, upstream_TSS LH3594: TTCTGTGCATATTCCGTCGT LH3595: AAGCCAACGAACAAGACCTG
MA_14427g0010, downstream_TSS LH3596: CGGTAAGCAGCTCGAGGAT LH3597: GTCTTTCCGGTCAGGGTTTT
MA_491379g0010, upstream_TSS LH3610: ATTCCTAGCCCCAACTCAGG LH3611: AGTTGCTTGGGAAGGACGTT
MA_491379g0010, downstream_TSS LH3608: TGCCGATTCCTTCTATCGTT LH3609: TTCTTCTTTGGCTTGGGTTG
MA_16054g0010, upstream_TSS LH3613: GGTTCTATTTGGTCAAGTCCCTTA LH3614: GGGGTTTCTCCCTCTTCTCT
MA_16054g0010, downstream_TSS LH3615: GCCCACAGAGCTCGGTAATA LH3616: GTTGGAGAGCCGATGAAATC
MA_10270349EnSpm, upstream LH3618: CCCACTTGTTTCCATCATCA LH3619: TTGAAATTGAAATTTGATGCTTT
MA_10270349EnSpm, inside LH3620: CCGAACGGATTTTAAAAGGAA LH3621: ATTTCAATGTTGGCCCCTTT





























