Embed Size (px)
Citation preview

A highly specialized mammalian skull from the Late Cretaceous of South America.
Guillermo W. Rougier 1, Sebastián Apesteguía2,3 & Leandro C. Gaetano 3,4
1 Department of Anatomical Sciences and Neurobiology, University of Louisville, Louisville, Kentucky, USA.
2 CEBBAD - Fundación de Historia Natural "Félix de Azara", Universidad Maimónides, Hidalgo 775, 7°p, Buenos Aires (1405), Argentina.
3 CONICET, Consejo Nacional de Investigaciones Científicas y Técnicas.
4
Departamento de Cs. Geológicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Pabellón II, Ciudad Universitaria, Buenos Aires, Argentina.
Supplementary information
Part 1: Materials, systematics, and phylogeny
Cronopio dentiacutus materials and expanded diagnosis.
Type: MPCA PV 454: A partial skull that is missing the skull roof, basicranium and squamosals. The
septomaxilla is not preserved, but its presence is deduced from a facet for this bone in the premaxilla.
Hypodigm: MPCA PV 450: partial left lower jaw with damaged teeth preserving posterior premolars
and damaged molars. MPCA PV 453: an incomplete skull with a relatively complete right lower jaw
missing all the dental positions mesial to and including p1. This specimen has suffered from bone
dissolution of the mesocranium. The petrosal remains in anatomical position but has no link with the
rostral part of the skull. The same phenomenon seems to account for the absence of the rear skull in
the type. Age and locality data: The fossils come from the Candeleros Formation, the basal unit of the
traditional Neuquén Group (i.e., see Ramos, 1988). For the Candeleros Formation there is no absolute
date available. Leanza et al. (2004) considered the Formation of a likely early Cenomanian age and
briefly summarized the known vertebrates. The same year Corbella et al. (2004) dated the conformably
SUPPLEMENTARY INFORMATIONdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 1

overlying Huincul Formation with a floor of approximately 92My (88 + 4); this date has been recently
challenged (Garrido, 2010) and it is now considered that the Candeleros Formation is wholly
Cenomanian, likely lower Cenomanian (Garrido 2010). Coordinates of the La Buitrera Locality are
available upon request.
Expanded diagnosis:
The upper incisors of Cronopio are simple and peg-like. Unlike many other dryolestoids, the first two
premolars of Cronopio lack sizable anterior accessory cusps but they are present posteriorly. In the
molars the parastylar hook is small and parastyle is absent, except on the M1. Both Cronopio and
Leonardus share the abrupt mesial narrowing of the rostrum and the posteriorly incurving of the
maxilla with the major axis of the molars directed mesiolingually; the larger Peligrotherium shares this
condition to some degree. We believe the single rooted molars of Cronopio derive from molars with
unequal roots similar to the condition seen among dryolestids and is therefore a derived condition. The
jaw of Cronopio is unusual among dryolestoids, with a small coronoid process, small angular process
and a very elevated condyle. The angular process is hook-like, short, but heavily inflected without
forming a medial shelf as seen in metatherians. The most distinctive feature is a large and prominent
masseteric process that extends laterally from the ventral edge of the jaw forming a solid lateral shelf.
A masseteric process is known among some “symmetrodonts” (Cifelli and Madsen, 1999) but it is much
smaller and likely not homologous.
Taxonomic definitions:
Meridiolestida: any dryolestoid more closely related to Peligrotherium than to Dryolestes or Paurodon.
The name is intended to reflect the Southern provenance of the members of this group that includes
several South American families: Mesungulatidae, Brandoniidae, Barbereniidae, Peligrotheridae,
Reigitheriidae, the formerly regarded triconodonts Austrotriconodontidae and possible Donodontidae
from the Cretaceous of Morocco. The validity of some of these families is problematic and depends on
the revision of the isolated teeth from Los Alamitos (Bonaparte and Soria, 1985; Bonaparte 1986; 1990,
1992, 1994, 2002), Allen Formations (Rougier et al., 2009a) and the remains from North Africa
(Sigogneau Russell, 1989; Sigogneau-Russell and Ensom, 1998).
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 2

Dryolestida: any dryolestoid more closely related to Dryolestes than to Peligrotherium.
Mesungulatoidea: the last common ancestor of Reigitherium, Mesungulatum and Peligrotherium plus
all its descendants.
Phylogenetic analysis and methods
The matrix below was compiled specifically for this project and it is an original data set. The matrix,
however, recovers and utilizes several previous matrices that were heavily modified for the goals of
our study here. Primary among those previous matrices is that presented by Rougier et al., 2007, which
in turn is a heavily modified version of Luo et al., (2002); substantial additions of characters and
character states used Prothero, (1981); Martin, (1999), and Wible et al. (2007; 2009) as sources.
Finally, original characters were added.
The authors have studied first hand most of the taxa included and when possible, scoring was based on
direct observations; otherwise the cast collection or the references were used for scoring. The data
was compiled in NDE (http://taxonomy.zoology.gla.ac.uk/rod/NDE/nde.html) for management,
commenting and editing.
The phylogenetic trees were obtained by using tree-search maximum parsimony ratchet analyses
(Nixon, 1999) using the program NONA and TNT (Goloboff, 1999; Goloboff et al., 2005), spawned
through the program Winclada (Asado, version 1.1 beta, by K. Nixon, Cornell University). Two runs
were performed: the results presented here (figure S1) included a set of additive characters (Farris
optimization), which included those used by Rougier et al., (2007) plus additional characters original to
this study (see below under assumptions). The second run considered all the characters as non-
additive (Fitch optimization). The results of this later exercise are similar to the one presented here,
however the dryolestoids are not a monophyletic group; the Meridiolestida and the Dryolestida are
successive sister groups of the stem therians.
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 3
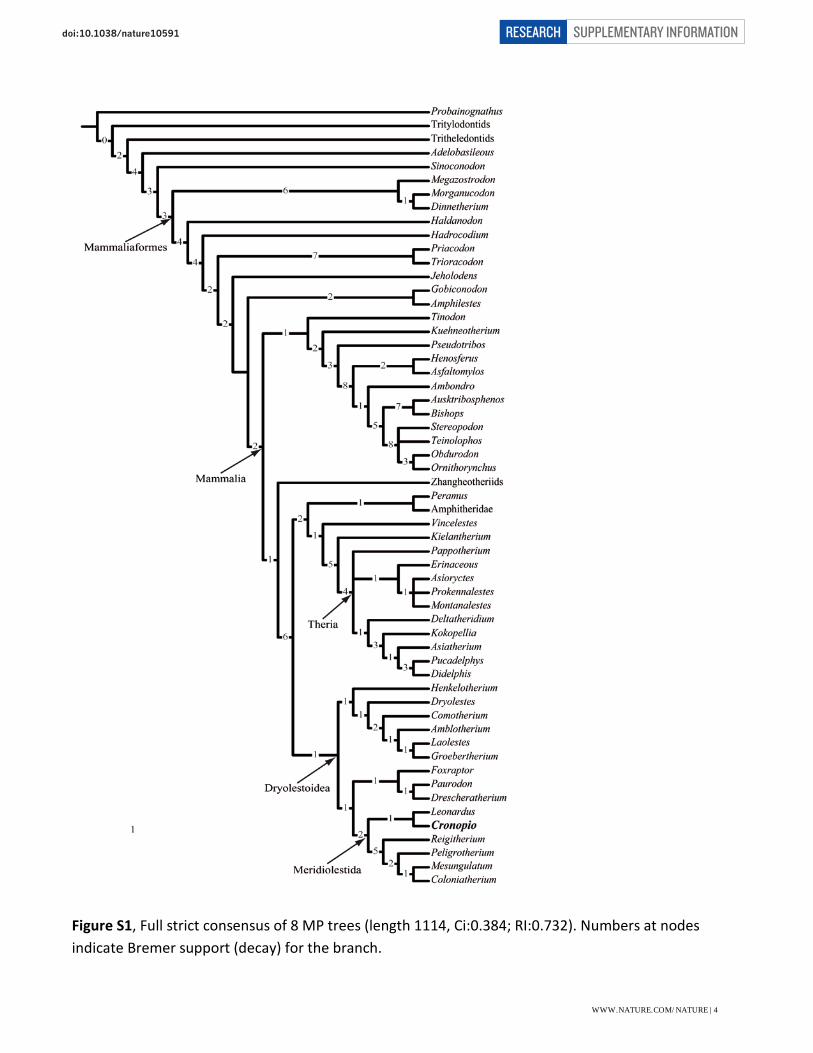
Figure S1, Full strict consensus of 8 MP trees (length 1114, Ci:0.384; RI:0.732). Numbers at nodes indicate Bremer support (decay) for the branch.
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 4

Figure S2, Detail of the dryolestoid portion of the consensus tree.
Sinapomorphy list of selected nodes of the strict consensus tree:
Dryolestoidea
Character 73: 1 --> 0 - Interlocking mechanism between two adjacent lower molars absent.
Character 93: 1 --> 0 - Labial cingulum of the upper molars absent or weak.
Character 116: 1 --> 2 - Metacristid (protocristid) orientation: transverse.
Character 119: 0 --> 1 - Distal metacristid absent.
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 5

Dryolestidae
Character 26: 2 --> 3 - Tilting of the coronoid process of the dentary: coronoid process is near vertical with an angle of 95-105 degrees.
Character 27: 1 --> 0 - Retromolar space absent.
Character 49: 0 --> 1 - Lower premolars lingual cingulid present.
Character 127: 2 --> 3 - Number of molars or molariforms postcanines: 6 or more.
Character 139: 0 --> 1 - Cingula pre and post cingula/cingulid height: close to crown base.
Node 1: unnamed node: Paurodontidae + Meridiolestoidea
Character 17: 2 --> 1 - Ventral border of the masseteric fossa present as a low and broad crest.
Character 44: 0 --> 1 - Last lower premolar- arrangement of principal cusp a, cusp b (if present) and cusp c: distinctive triangulation.
Character 121: 0 --> 1 - Position of stylocone in posterior molariforms: separated.
Character 122: 0 --> 1 - Stylocone relationship in triangular teeth: stylocone distal to labial ending of paracrista.
Meridiolestoidea
Character 5: 1 --> 2 - Degree of development of the Meckelian groove in adults: vestigial or absent.
Character 45: 1 --> 2 - Last lower premolar distinct distal cingulid cusp d: present as part of a continuous distal cingulid.
Character 123: 1 --> 2 - Stylocone size in triangular teeth: prominent cusp subequal or larger than paracone.
Character 126: 0 --> 1 – Metacone absent.
Character 127: 2 --> 1 - Number of molars or molariforms: 3 molars/molariforms.
Character 133: 0 --> 1 - Lower molar root cross section: anteroposteriorly compressed.
Character 210: 1 --> 0 - Sharp constriction of rostrum in front of molariform premolar present.
Character 277: 1 --> 0 - Maximum vertical depth of the zygomatic arch relative to the length of the skull between 10-20%.
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 6

Mesungulatoidea
Character 70: 0 --> 2 - Mesial transverse cingulid present, having occlusal contact with the upper molar.
Character 72: 0 --> 2 – Postcingulid present, horizontal above the gum level.
Character 103: 4 --> 5 - Outline of m1 rectangular or slightly rhomboidal.
Character 116: 2 --> 1 - Metacristid (protocristid) orientation: oblique.
Character 137: 0 --> 1 – Precingulum developed forming a lingual cusp.
Character 138: 0 --> 1 – Postcingulum developed forming a lingual cusp.
Character 139: 0 --> [12] - Cingula pre and post cingula/cingulid height: [1] close to crown base, or [2] elevated reaching occlusal surface.
Character 144: 0 --> 1 - Supernumerary roots on penultimate lower premolar present.
Character list, character states and matrix:
1. Symphysis height: 0. robust, verticalized 1. slender, oblique or approaching horizontal
2. Position of posterior-most mental foramen:
0. below the canine and anterior premolariform region 1. below the penultimate premolar 2. below the ultimate premolar 3. between the ultimate premolar and the first molar junction
3. Post-dentary trough: behind tooth row
0. present 1. absent
4. Mandibular alveolar border:
0. subequal in height to the lingual edge 1. labial border much lower than lingual
5. Degree of development of the Meckelian groove in adults:
0. well developed 1. weakly developed 2. vestigial or absent
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 7

6. Curvature of the Meckelian groove in adult: under the tooth row
0. parallel to the ventral border of mandible 1. convergent to the ventral border of mandible
7. Groove for the replacement dental lamina:
0. present 1. absent
8. Angular process presence:
0. absent 1. present
9. Angular process direction:
0. small knobby process, not projected 1. straight process posteriorly directed 2. transversely flaring 3. inflected
10. Antero-posterior position of angular process relative to the dentary condyle:
0. anterior position, the angular process is below the main body of the coronoid process 1. posterior position, the angular process is placed to the same level of the posterior end
of the coronoid process 11. Vertical position of angular process:
0. low, at or near the level of the ventral border of the mandibular horizontal ramus 1. high, at or near the level of the molar alveolar line
12. Coronoid, or its attachment scar, in adults:
0. present 1. absent
13. Medial fossa on the dentary angular process:
0. present 1. absent
14. Pterygoid fossa:
0. absent 1. present
15. Medial pterygoid ridge or shelf direction:
0. directed to angular process 1. reaching the dentary condyle via a low crest
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 8

16. Pterygoid shelf: 0. absent 1. present
17. Ventral border of the masseteric fossa:
0. absent 1. present as a low and broad crest 2. present as a well-defined and thin crest
18. Position of the mandibular foramen:
0. below or near to the base of the anterior border of the coronoid process 1. posterior to the anterior edge of the coronoid process
19. Masseteric foramen:
0. absent 1. present
20. Crest of the masseteric fossa along the anterior border of the coronoid process:
0. absent or weakly developed 1. present as a distinct anterior border
21. Mylohyoid process at the level of the anterior border of the coronoid process:
0. absent 1. present
22. Orientation of the dentary peduncle and condyle:
0. dentary peduncle is posteriorly directed, forms an angle of 35 degrees or less to the alveolar margin
1. vertically directed dentary peduncle, above 35 degrees 2. dentary condyle is continuous with the semicircular posterior margin of the dentary
23. Shape and relative size of the dentary articulation:
0. small and dorsoventrally compressed 1. condyle is massive and bulbous, transversely broad in its dorsal aspect 2. condyle mediolaterally narrow and vertically deep, forming a broad arc in lateral
outline, either ovoid or triangular in posterior view 24. Ventral border of the dentary peduncle:
0. posteriorly tapering without a condyle 1. columnar or ridge-like 2. ventrally flaring 3. robust and short
25. Position of the dentary condyle relative to the vertical level of the postcanine alveoli:
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 9

0. below or about the same level as the postcanine alveoli 1. above the level of the postcanine alveoli
26. Tilting of the coronoid process of the dentary: measured as the angle between the imaginary line of the anterior border of the coronoid
process and the horizontal alveolar line of all molars 0. coronoid process is strongly reclined forming an obtuse angle 1. coronoid process less reclined, 135-145 degrees 2. coronoid process less than vertical, 115-125 degrees 3. coronoid process is near vertical and the angle is small, 95-105 degrees
27. Retromolar space: at least half the length of the last molar
0. absent 1. present
28. Alignment of ultimate molar to the anterior margin of the dentary coronoid process:
0. ultimate functional molar is medial to the coronoid process 1. ultimate functional molar is in alignment with the anterior margin of coronoid process
29. Incisor replacement:
0. alternating and multiple replacement 1. diphyodont replacement or none
30. Number of lower incisors:
0. 3 or more 1. 2 or fewer
31. Maxillary incisor:
0. present 1. absent
32. Staggered incisor:
0. absent 1. present
33. Canine replacement:
0. multiple replacements 1. diphyodont
34. Canines:
0. present and large 1. present and small 2. absent
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 10

35. Upper canine height:
0. long, at least twice the height of tallest postcanine 1. short, less than twice the height of the tallest postcanine
36. Long upper canine height:
0. relatively short, less than 3 times the height of the tallest postcanine crown 1. tall, more than 3 times the height of tallest postcanine
37. Replacement of premolariform:
0. multiple replacement 1. one replacement or none
38. Total number of premolars: Lower premolar preferred, uppers used if no lowers are available
0. 2 or fewer 1. 3 premolars 2. 4 premolars 3. 5 or more
39. Diastema separating P1 from P2: distance equal or larger than half of the P1
0. absent 1. present
40. Penultimate lower premolar size:
0. small and subequal to other premolars 1. larger than any other premolar, longer and/or taller 2. hypertrophied, dominant tooth in the series
41. Penultimate lower premolar - paraconid (cusp b):
0. much smaller than metaconid (cusp c) of the same tooth, or absent 1. well developed as an important cusp of trigonid
42. Last lower premolar- symmetry of the main cusp a (protoconid):
0. asymmetrical, anterior edge of cusp a is more convex in outline than the posterior edge 1. symmetrical, anterior and posterior cutting edges are equal or subequal in length
43. Last lower premolar-anterior cusp b (paraconid):
0. present, at least subequal to cusp c, or posterior cingular cusp of the same tooth 1. small, much smaller than cusp c or posterior cingular cusp of the same tooth, or vestigial
to absent 44. Last lower premolar arrangement of principal cusp a, b (if present) and cusp c:
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 11

0. aligned straight or at a slight angle 1. distinctive triangulation
45. Last lower premolar distinct distal cingulid cusp d:
0. absent 1. posterior cingular cusp present 2. present as part of a continuous distal cingulid
46. Last lower premolar outline:
0. laterally compressed, crown outline longer than wide 1. transversely wide, crown outline subequal or wider than long
47. Last lower premolar size:
0. small and subequal to other premolars 1. large tooth, taller than first molar 2. hypertrophied, much larger than any molar
48. Labial cingulid of last lower premolar:
0. absent or vestigial 1. present along more than half of the crown length
49. Lower premolars lingual cingulid:
0. absent or vestigial 1. present
50. Relative height of the primary cusp a to c of the last lower premolar: measured as the height ratio of a and c from the bottom of the valley between the two adjacent
cusps 0. posterior cusp c is absent or very small 1. posterior cusp c is distinctive but less than 30% of primary cusp a 2. posterior cusp c and primary cusp a are equal or subequal in height (c is 40-100% of a)
51. Last upper premolar functional protocone:
0. absent 1. present
52. Last upper premolar parastylar hook:
0. present 1. absent or very small
53. Alignment of main cusps of the posterior lower molars: Reigitherium: lingual cusps are fused, m3 or more posterior if present
0. simple longitudinal row 1. obtuse angle (>90)
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 12

2. acute angle (<90) 54. Alignment of main cusps of the anterior lower molar (m1):
0. single longitudinal row 1. acute angle 2. obtuse angle
55. Development of postprotocrista on the upper molar for double rank postvallum shear
(applicable to molars with reversed triangulation of molar cusps): 0. postprotocrista is short and does not extend labially beyond metacone 1. postprotocrista is long and extends labially beyond metacone
56. Precise opposition of the upper and lower molars (either one-to-one, or occluding at the
opposing embrasure or talonid): 0. absence of precise opposition of upper and lower molars 1. present (either one-to-one, or occluding at the opposing embrasure or talonid)
57. Relationships between the cusps of the opposing upper and lower molars:
0. absent 1. present, lower primary cusp a occludes in the groove between upper cusp A and B 2. present, lower main cusp a occludes in front of upper cusp B and into the embrasure
between the opposing and preceding upper teeth 3. present, part of the talonid occluding with the lingual face (or any part) of the upper
molar 4. lower multicuspate rows alternately occlude between the upper multicuspate rows
58. Relative height of the primary cusp a (protoconid) to cusp c (metaconid) of the anterior lower
molars: measured as the height ratio of a and c from the bottom of the valley between the two adjacent
cusp, on m1 0. posterior cusp c is less than 40% of the primary cusp a (protoconid) 1. posterior cusp c is more than 40% of cusp a
59. Relative size/height of cusp b (paraconid) to cusp c (metaconid): based on m2 when possible
0. c taller than b 1. b and c subequal in height 2. b taller than c
60. Relative elevation of the bases of the paraconid (cusp b) and metaconid (cusp c):
0. almost or at the same level 1. base of the paraconid higher than base of the metaconid 2. base of metaconid higher than base of the paraconid
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 13

61. Cristid obliqua: 0. absent 1. present
62. Cristid obliqua orientation:
0. oriented to, or lingual to the metaconid-protoconid notch 1. hypertrophied, and directed to posterior part of the metaconid 2. short, and pointed anteriorly between the metaconid-protoconid notch and the
protoconid 63. Lower molar- pre-entocristid on the talonid heel:
0. talonid lacks medial and longitudinal crest 1. pre-entoconid cristid of talonid in alignment with the metaconid or with the
postmetacristid if the latter is present 2. pre-entocristid crest is offset from the metaconid and it is lingual to the base of the
metaconid 64. Labial curvature of the primary cusp a of lower molars (at the base level) relative to the
curvature of cusps b and c: 0. cusps a, b, and c have the same degree of bulging 1. cusp a is far more bulging than cusps b, c
65. Labial curvature of the main cusps A, B, C at the level of the cusp valley of the penultimate
and ultimate upper molars: 0. cusp A, B, and C have about the same degree of curvature 1. cusp A is slightly concave (or far less convex than either cusp B or C)
66. Labiolingual compression of the primary functional cusps of the lower molars (at the level of
the cusp base but above the cingulid): 0. absent 1. present
67. Posterior lingual cingulid of the lower molar:
0. absent or weak 1. distinctive 2. strongly developed, crenulated with distinctive cuspules
68. Cingular cuspule e on the lower molars:
0. present 1. absent
69. Cingular cuspule f:
0. absent 1. present
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 14

70. Mesial transverse cingulid:
0. absent 1. present as a continuous shelf bellow the trigonid without occlusal function 2. present, having occlusal contact with the upper molar
71. Cingulid shelf wrapping around the anterointernal corner of the molar to extend to the
lingual side of the trigonid below the paraconid: 0. absent 1. present, weakly developed restricted to the mesial aspect of the paraconid base 2. present, strongly developed, running along most of the lingual base of the paraconid
72. Postcingulid:
0. absent 1. present, oblique and connected to hypoconulid 2. present, horizontal above the gum level
73. Interlocking mechanism between two adjacent lower molars:
0. absent 1. present, posterior cingular cuspule d of the preceding molar fits in between cingular
cuspules e and f of the succeeding molar or flat surfaces of mesial cingula or cusp b 2. present, posterior cingular cuspule d fits between cingular cuspule e and cusp b of the
succeeding molar 3. present, posterior cingular cuspule d of the preceding molar fits into and embayment or
vertical groove of the anterior aspect of cusp b the succeeding molar 74. Size ratio of the posterior molars: Lower molar preferred when available
0. last three postcanines forming a series of posteriorly decreasing size 1. penultimate molar is the largest of molars 2. ultimate molar is the largest of molars
75. Orientation of the paracristid relative to the longitudinal axis molars:
0. longitudinal orientation 1. oblique 2. nearly transverse
76. Paraconid presence:
0. present 1. absent
77. Mesiolingual surface of paraconid on lower molars:
0. rounded 1. forming a keel
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 15

78. Procumbent paraconid:
0. absent 1. present, projected as a conical cusp beyond crown base
79. Proximity between paraconid and metaconid:
0. bases widely separated 1. bases approaching each other becoming confluent 2. single cusp (amphyconid)
80. Molar trigonid/talonid width ratio:
0. narrow (talonid <40% of trigonid) 1. wide (talonid is 40-70% of the trigonid) 2. talonid is equal or wider than trigonid (above 70% of the width of the trigonid)
81. Lower molar hypoflexid:
0. absent or shallow 1. deep (but less than 50% of the talonid width) 2. very deep (>60% of the talonid width)
82. Talonid basin:
0. absent 1. present
83. Morphology of rear portion of molariform:
0. single cusp (d) or cingulum 1. present as an incipient heel 2. present as a heel (with at least one functional cusp) 3. present as a transverse V-shaped basin two major cusps 4. rimmed with 3 major cusps
84. Hypoconulid (= cusp d):
0. hypoconulid at the cingulid level 1. hypoconulid elevated above the cingulid level
85. Hypoconid:
0. absent 1. present
86. Hypoconulid orientation:
0. cusp tip erect or procumbent 1. cusp tip recumbent (reclined posteriorly)
87. Entoconid:
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 16

0. absent 1. present but far from hypoconulid, at least equal to one cusp length 2. present and twinned with hypoconulid
88. Height of the entoconid as compared to other cusps of the talonid:
0. lower than the hypoconulid (or even vestigial) 1. subequal height as the hypoconulid
89. Alignment of the paraconid, metaconid and entoconid:
0. cusps not aligned 1. cusps aligned
90. Aspect ratio in occlusal view (length versus width) of the functional talonid basin at the
cingulid level: 0. longer than wide 1. length equals width 2. wider than long
91. Elevation of the talonid: 0. hypoconulid/protoconid height ratio less than 20% (hypoconulid or cusp d)
1. hypoconulid/protoconid height ratio between 25% and 35% 2. hypoconulid/protoconid height ratio between 40% and 50% 3. hypoconulid/protoconid height ratio: 50% or higher
92. Width of upper molar stylar shelf:
0. present and broad 1. present and narrow 2. absent
93. Labial cingulum of the upper molars:
0. absent or weak 1. distinctive cingulum, straight 2. distinctive cingulum with strong ectoflexus (but without hypertrophied stylar cusps) 3. wide cingulum with distinctive ectoflexus, plus individualized and hypertrophied stylar
cusps 4. cingulum with distinctive and even-sized multiple cuspules
94. Upper molars with a functional lingual protocone that grinds against a basin on the lowers:
0. absent 1. present
95. Transverse width of the protocone on upper molars: protocone must be present
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 17

0. narrow (distance from the protocone apex to paracone apex less than 0.60 of total tooth width)
1. strongly transverse (distance from the protocone apex greater than 0.60 of total width) 96. Anteroposterior development of the lingual region on the upper molars: Must have protocone
0. narrow (anteroposterior distance medial to the paracone and metacone less than 0.30 of total tooth length)
1. moderate development (distance between position of conules=0.31-0.50 of total tooth length)
2. long (distance between conules greater than 0.51 of total tooth length) 97. Conules on the upper molars:
0. absent 1. present but weak and without cristae 2. conule distinctive, with cristae
98. Relative height and size of the paracone (cusp B) and metacone of the upper molars:
0. paracone higher and larger than metacone 1. metacone is higher and larger than paracone
99. Centrocrista between the paracone and metacone of upper molars: Considered only for tribosphenic forms
0. straight 1. V-shaped, with labially directed postparacrista and premetacrista
100. Upper molars cuspule E:
0. present 1. absent
101. Upper molar interlock:
0. absent 1. tongue-in-groove interlock
102. Central crest (medianergrat) in triangular upper molariforms:
0. absent 1. present
103. Outline of m1:
0. oval-shaped 1. laterally compressed 2. oblong with slight labial bulge 3. oblong with strong labial bulge 4. triangular or tear-drop shaped
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 18

5. rectangular or slightly rhomboidal 104. Crown length width ratio among inferior molariforms:
0. crown longer than wide 1. crown length /width subequal 2. crown wider than long
105. Shape of lingual margin in lower molars:
0. notched 1. flat
106. Aspect ratio of M1:
0. laterally compressed 1. oval-shaped or spindle shaped 2. triangular outline 3. dumbbell shaped 4. rectangular or nearly so
107. Crown height difference between buccal and lingual sides in lower molariform:
0. buccal and lingual sides of similar height 1. buccal side much taller than lingual side
108. Functional development of occlusal facets on individual molar cusps:
0. absent for lifetime 1. absent at eruption but development later by extensive wearing of the crown 2. wear facet develops on a morphology approximately present upon eruption
109. Topographic relationships of wear facets to the main cusps:
0. lower cusps a, c support two different wear facets (1 and 4) that contact the upper main cusp A
1. lower cusps a, c support a single wear facet (4) that contacts the upper primary cusp B 110. Development and orientation of the prevallum/postvallid shearing:
0. absent 1. present and obtuse 2. present, hypertrophied and transverse
111. Upper molar- development of facet 1 and preprotocrista (or paracrista) on upper molar:
0. facet 1 (prevallum crest) is short, and does not extend to the stylocone area 1. wear facet 1 extends beyond into the hook-like area near the stylocone 2. long preprotocrista (below the paracone-stylocone crest) is added to the prevallum
shear and extends labially beyond paracone 112. Differentiation of wear facet 3 and facet 4:
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 19

0. absent 1. present 2. facet hypertrophied on the flanks of the strongly V-shaped talonid
113. Orientation of facet 4:
0. present and oblique to long axis of the tooth 1. present and forming a more transverse angle to long axis of the tooth
114. Wear pattern on talonid:
0. absent 1. present
115. Direction of jaw movement during occlusion:
0. dorsomedial movement 1. dorsomedial movement with a significant medial component 2. dorsoposterior movement
116. Metacristid (protocristid) orientation:
0. parallel to lower jaw axis 1. oblique 2. transverse
117. Bifid metaconid:
0. absent 1. present
118. Bifid metastyle:
0. absent 1. present
119. Distal metacristid:
0. present 1. absent
120. Superficial features on the occluding surface of facet 5 and 6 in the talonid for basined
talonid: 0. smooth surface on the talonid (or on cusp d) 1. multiple ridge within talonid basin
121. Position of stylocone in posterior molariforms:
0. along buccal edge 1. separated
122. Stylocone relationship in triangular teeth:
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 20

0. stylocone connected to paracrista or mesial to its end 1. stylocone distal to labial ending of paracrista 2. stylocone detached of preparacrista occupying central position on crown
123. Stylocone size in triangular teeth:
0. absent 1. small stylar cusp 2. prominent cusp subequal or larger than paracone
124. Parastylar hook in molars:
0. absent or poorly developed 1. present
125. Paracone orientation:
0. erect 1. recumbent 2. procumbent
126. Metacone:
0. present 1. absent
127. Number of molars or molariforms postcanines: Lowers preferred, upper used if no lowers known
0. 2 molars 1. 3 molars 2. 4 to 5 molars 3. 6 or more
128. Accessory cusps on buccal side:
0. absent 1. present
129. Deep ectoflexus:
0. present only on penultimate / ultimate molar 1. on penultimate and preceding molar 2. strongly reduced to absent
130. Lower molariform roots:
0. incipient or incomplete separation 1. root divided
131. Number of lower molariform roots:
0. one
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 21

1. two 2. three or more
132. Size of lower molar roots:
0. subequal 1. posterior molar root much smaller 2. single root (posterior root absent)
133. Lower molar root cross section, m2 when available:
0. circular/subcircular 1. anteroposteriorly compressed
134. Replacement of at least some functional molariforms:
0. present 1. absent
135. Procumbent and enlargement of the anteriormost lower incisor:
0. absent 1. present both procumbent and enlarged more than 50% the second
136. Trigon major axis orientation: as indicated by the distal wall of the trigonid
0. labially 1. mesially 2. sharply distal
137. Precingulum:
0. narrow and closely attached to crown, to absent 1. developed forming a lingual cusp
138. Postcingulum:
0. narrow and closely oppressed to crown, to absent 1. developed forming a lingual cusp
139. Cingula pre and post cingula/cingulid height:
0. absent or little differentiated 1. close to crown base 2. elevated reaching occlusal surface
140. Prehypoconulid crest: a crest connecting the metaconid with the hypoconulid along the lingual edge of the tooth
0. absent 1. present
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 22

141. Number of upper molariform roots: 0. one 1. two 2. three 3. more than three
142. Position of lingual upper root:
0. under paracone 1. under protocone or trigon
143. Supernumerary roots on penultimate upper pm:
0. absent 1. present
144. Supernumerary roots on penultimate lower pm:
0. absent 1. present
145. Supernumerary roots on ultimate lower premolar:
0. absent 1. present
146. Supernumerary roots on ultimate upper premolar:
0. absent 1. present
147. Penultimate lower premolar distal root:
0. subequal to mesial root 1. large elongated root, more than 50% of crown length
148. Lower molar contact:
0. lower molars contact each other somewhere along the mesial and distal edges of the crown
1. lower molars do not contact each other, being separated by interdental spaces 149. Upper molar contact:
0. upper molars extensively contact each other 1. upper molars do not contact each other, or barely do so.
150. Fusion of atlas neural arch and intercentrum in adults:
0. unfused 1. fused
151. Atlas ribs in adults:
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 23

0. present 1. absent
152. Fusion of dens to axis:
0. unfused 1. fused
153. Rib of axis in adults:
0. free ribs present 1. ribs fuse to become transverse process
154. Postaxial cervical ribs in adult:
0. free ribs present 1. free ribs absent
155. Thoracic vertebrae:
0. 13 thoracic vertebrae 1. 15 or more vertebrae
156. Lumbar ribs:
0. unfused to vertebrae 1. synostosed to vertebrae to form transverse processes
157. Interclavicle in adults:
0. present 1. absent
158. Contact relationships in adults between the interclavicle and sternal manubrium:
0. posterior end of interclavicle abuts anterior border of manubrium 1. interclavicle broadly overlaps the ventral side of the manubrium 2. complete fusion of the embryonic membranous and endochondral elements
159. Cranial margin of the interclavicle:
0. anterior border is emarginated or flat 1. with a median process (assuming interclavicle is fused to the sternal manubrium in living
therians) 160. Claviculo-sternal apparatus joint:
0. inmobile 1. movile
161. Acromioclavicular joint:
0. extensive articulation 1. limited articulation
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 24

162. Curvature of the clavicule:
0. boomerang-shaped 1. slightly curved
163. Scapula- supraspinous fossa:
0. absent 1. weakly developed, present only along a part of the scapula 2. fully developed and present along the entire dorsal border of scapula
164. Scapula- acromion process:
0. short, even with or behind the glenoid 1. hook-like and extending below the glenoid
165. Scapula- fossa or process for the teres major muscle:
0. absent 1. present
166. Procoracoid:
0. present as a free element 1. fused to the sternal apparatus in adult
167. Procoracoid foramen:
0. present 1. absent
168. Coracoid:
0. large, with posterior process 1. small, without posterior process
169. Manubrium size relative to the succeeding sternebrae:
0. large 1. small
170. Orientation of the glenoid relative to the plane or axis of scapula:
0. nearly parallel to the long axis and facing posterolaterally 1. oblique and facing more posteriorly 2. perpendicular to the main plane of the scapular plate
171. Shape and curvature of the glenoid:
0. saddle-shaped, oval and elongated 1. uniformly concave and more rounded in outline
172. Medial surface of the scapula:
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 25

0. concave 1. flat
173. Humeral head:
0. subspherical, weakly inflected 1. spherical and strongly inflected
174. Intertubercular groove that separates deltopectoral crest from lesser tubercle:
0. shallow and broad 1. narrow and deep
175. Size of the lesser tubercle of the humerus:
0. wider than the greater tubercle 1. narrower than the greater tubercle
176. Torsion between the proximal and distal ends of the humerus:
0. strong (>30) 1. moderate (30-15) 2. weak (<15)
177. Ventral extension of the deltopectoral crest or position of the deltoid tuberosity:
0. not extending beyond the midpoint of the humeral shaft 1. extending ventrally (distally) past the midpoint of the shaft
178. Ulnar articulation on the distal humerus:
0. bulbous ulnar condyle 1. incomplete trochlea with vestigial ulnar condyle in anterior view 2. trochlea has extending to the anteroventral side
179. Radial articulation on the distal humerus:
0. distinct and rounded condyle separated from the ulnar articulation in the anteroventral view of the humerus
1. radial articulation forms a rounded condyle anteriorly but its posterior surface is nearly cylindrical
2. capitulum, radial articulating structure forms a continuous synovial surface with the ulnar trochlea
180. Entepicondyle and ectepicondyle of the humerus:
0. robust 1. weak
181. Rectangular shelf for the supinator ridge extended from the ectepicondyle:
0. absent 1. present
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 26

182. Styloid process of the radius:
0. weak 1. strong
183. Enlargement of the scaphoid with a distomedial projection:
0. absent 1. present
184. Size and shape of the hamate (unciform) in the wrist:
0. anteroposteriorly compressed (wider than longer in dorsal view) 1. mediolaterally compressed (longer than wide)
185. Acetabular dorsal emargination:
0. emarginated 1. with a complete rim
186. Sutures of ilium, ischium, and pubis within the acetabulum in adults:
0. unfused 1. fused
187. Ischiatic tuberosity:
0. dorsal margin with a small or absent ischiatic tuberosity 1. dorsal margin concave and ischiatic tubercle hypertrophied
188. Head of the femur is inflected and set off from the shaft by a neck:
0. neck absent, head oriented dorsally 1. neck present and head inflected medially
189. Fovea for the acetabular ligament on femoral head:
0. absent 1. present
190. Greater trochanter:
0. directed dorsolaterally 1. directed dorsally
191. Orientation of the lesser trochanter:
0. on the medial side of the shaft 1. on the ventromedial or ventral side of the shaft
192. Size of the lesser trochanter:
0. large 1. small
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 27

193. Patellar groove of the femur:
0. absent 1. shallow and weakly developed 2. well developed
194. Proximolateral tubercle or tuberosity of the tibia:
0. large and hook-like 1. indistinct
195. Distal tibial malleolus:
0. weak 1. distinct
196. Fibular contacting distal end of the femur:
0. present 1. absent
197. Distal fibular styloid process:
0. weak or absent 1. distinct
198. Fibula contacting the calcaneous:
0. extensive contact 1. reduced 2. mortise and tenon contact of fibula to the ankle
199. Superposition of the astragalus over the calcaneous:
0. little or absent 1. weakly developed 2. present
200. Orientation of the sustentacular facet of the calcaneous with regard to horizontal plane
astragalus: 0. nearly vertical 1. oblique (<70) to nearly horizontal
201. Astragalar neck:
0. absent 1. weakly developed 2. present
202. Astragalar trochlea:
0. absent
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 28

1. present 203. Calcaneal tubercle:
0. short, without terminal swelling 1. elongated with distal swelling
204. Peroneal process and groove of the calcaneous:
0. forming laterally directed shelf, and without a distinct process 1. weakly developed with shallow groove on the lateral side of process 2. with a distinct peroneal process
205. Contact of the cuboid on the calcaneous:
0. On the anterior end of the calcaneus, the cuboid is aligned with the long axis of the calcaneous
1. On the anteromedial aspect of the calcaneous, the cuboid is skewed to the medial side of the long axis of the calcaneous
206. Relationships of the proximal end of metatarsal V to the cuboid:
0. metatarsal V is offset from the cuboid 1. metatarsal V is far offset from the cuboid, so that it contacts the calcaneous 2. metatarsal V is aligned with the cuboid
207. Angle of metatarsal III to the calcaneous:
0. metatarsal III is aligned with (or parallel to) an imaginary line through the long axis of the calcaneous
1. metatarsal III is oriented oblique to an imaginary line through the long axis of the calcaneous
208. Sesamoid bones in flexor tendons:
0. absent 1. present and unpaired 2. present and paired
209. Tarsal spur:
0. absent 1. present
210. Sharp constriction of rostrum in front of molariform premolar:
0. present 1. absent
211. External size of the cranial moiety of squamosal:
0. narrow 1. broad
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 29

212. Participation of the cranial moiety of the squamosal to the braincase:
0. does not participate in the endocranial wall of the braincase 1. participates in the endocranial wall of the braincase
213. Neck between the glenoid and the cranial moiety of the squamosal:
0. absent 1. present
214. External auditory meatus:
0. present as the post-craniomandibular joint sulcus 1. absent 2. present as a groove on the squamosal, or a notch
215. Position of the craniomandibular joint:
0. posterior or lateral to the level to the fenestra vestibuli 1. anterior to the level of the fenestra vestibuli
216. Orientation of the glenoid:
0. on the inner side of the zygoma and facing ventromedially 1. on platform of the zygoma and facing ventrally
217. Postglenoid process:
0. absent 1. present as a distinctive process
218. Postglenoid foramen within the squamosal bone:
0. absent 1. present
219. Basisphenoid wing on the ventral aspect of the skull:
0. present, overlapping part of or the whole cochlear housing 1. absent
220. Relationship of the pars cochlearis to the lateral lappet of basioccipital:
0. pars cochlearis is entirely covered by basioccipital 1. pars cochlearis partially covered by basioccipital 2. pars cochlearis fully exposed as promontorium
221. Medial flat facet of the promontorium of the pars cochlearis:
0. present 1. it is inflated and convex
222. Ventral outline and morphology of the promontorium:
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 30

0. triangular, with steep and slightly concave lateral wall 1. elongated and cylindrical petrosal cochlear housing 2. bulbous and oval-shaped promontorium
223. Cochlea:
0. short and uncoiled 1. elongated and partly coiled 2. elongated and coiled to about 360 grade
224. Morphology of the internal acoustic meatus:
0. the floor is ossified and the meatus is a deep tube 1. present as a shallow depression 2. present and the floor is developed as the cribriform foramina for auditory nerve
225. Primary bony lamina within the cochlear canal:
0. absent 1. present
226. Secondary bony lamina for the basilar membrane within the cochlear canal:
0. absent 1. present
227. Crista interfenestralis:
0. horizontal and extending to base of paroccipital process 1. vertical, delimiting the back of the promontorium
228. Post-tympanic recess:
0. absent 1. present
229. Caudal tympanic process of the petrosal:
0. absent 1. present as a continuous crest 2. caudal tympanic process notched
230. Prootic canal presence:
0. present 1. absent
231. Prootic canal confluence:
0. prootic canal present, and its tympanic aperture is a distinct, separate foramen 1. prootic canal present, and its tympanic aperture is confluent with the
pterygoparoccipital foramen
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 31

232. Lateral trough floor anterior to the tympanic aperture of the prootic canal and/or the primary facial foramen: 0. open lateral trough but no bony floor 1. present as a bony shelf 2. lateral trough absent
233. Enclosure of the geniculate ganglion by bony floor of the petrosal:
0. absent 1. present
234. The anteroventral opening of the cavum epiptericum:
0. fully open ventrally 1. partially enclosed by petrosal or lateral flange 2. enclosed by both the alisphenoid and the petrosal
235. The anterior lamina of the petrosal and ascending process of the alisphenoid and their
relationships to the exit of the mandibular brach (V3) of the trigeminal nerve: 0. V3 foramen placed at the suture of the alisphenoid ascending process and the anterior
lamina of petrosal 1. V3 placed within the enlarged anterior lamina of the petrosal 2. double trigeminal foramina within the anterior lamina in addition to the trigeminal
foramen at the anterior lamina border with alisphenoid 3. V3 within the ascending process of the alisphenoid
236. Quadrate ramus of the alisphenoid:
0. forming a rod overlapping with the anterior part of the lateral flange 1. present, mostly laminar process in the vicinity of the oval foramen 2. absent
237. Orientation of the anterior part of the lateral flange:
0. horizontal shelf 1. ventrally directed 2. vestigial or absent
238. Vascular foramen in the posterior part of the lateral flange anterior to the pterygoparoccipital
foramen: 0. present 1. absent
239. Relationship of the petrosal lateral flange to the crista parotica:
0. widely separated 1. narrow separated 2. continuous bone formed by petrosal
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 32

240. Morphology of the pterygoparoccipital foramen: i.e. ramus superior foramen
0. laterally open notch 1. foramen enclosed by the petrosal or squamosal or both
241. Position of the pterygoparoccipital foramen relative to the fenestra vestibuli:
0. foramen posterior or lateral to the level of the fenestra vestibuli 1. foramen anterior to the level of the fenestra vestibuli
242. Bifurcation of paroccipital process:
0. absent 1. present
243. Posterior paroccipital process of the petrosal:
0. no ventral projection below the level of its surrounding structures 1. projecting below the surrounding structures
244. Morphological differentiation of the anterior paroccipital region:
0. anterior paroccipital region is indistinct from surrounding structures 1. anterior paroccipital region is bulbous and distinctive from the surrounding structures 2. anterior paroccipital region has a distinct crista parotica
245. Epitympanic recess lateral to the crista parotica:
0. absent 1. present
246. Relationship of the squamosal on the paraoccipital process:
0. squamosal covers the entire paroccipital region 1. no squamosal cover on anterior paroccipital region 2. squamosal covering a part of the paroccipital region, but not on the crista parotica (the
squamosal wall and the crista parotica are separated by the epitympanic recess) 247. Medial process of the squamosal reaching toward the foramen ovale:
0. absent 1. present
248. Stapedial artery sulcus on the petrosal:
0. absent 1. present
249. Transpromontorial sulcus for the internal carotid artery on the pars cochlearis:
0. absent 1. present
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 33

250. Bullar process of the alisphenoid: 0. absent 1. present
251. Hypotympanic recess in the junction of the alisphenoid, squamosal and petrosal:
0. absent 1. present
252. Separation of the fenestra cochleae from the jugular foramen:
0. fenestra cochleae and jugular foramen within the same depression 1. separate
253. Channel of the perilymphatic duct:
0. open channel and sulcus 1. channel partially or fully enclosed
254. Tensor tympani fossa:
0. indistinct or very shallow 1. deep recess on lateral trough anterior to hiatus Fallopii
255. Stapedial muscle fossa:
0. absent 1. present and in alignment with the crista interfenestralis 2. present and lateral to the crista interfenestralis
256. Hypoglossal foramen:
0. indistinct, either confluent with the jugular foramen or sharing a depression with the jugular foramen
1. separated from the jugular foramen 257. Shape of the incudo-mallear contact:
0. trochlear surface on the incus 1. trough or saddle-shaped contact on the incus 2. flat surface
258. Quadrate-incus neck: separation of the dorsal plate and the trochlea; this represents the differentiation between body
and crus brevis of the incus 0. absent 1. present
259. Stapedial process of the incus-quadrate:
0. absent 1. present
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 34

260. Dorsal plate (=crus brevis) of the quadrate-incus:
0. broad plate 1. pointed triangle 2. reduced
261. Incus, angle of the crus brevis to crus longus:
0. alignment or obtuse angle between stapedial process (crus longum) and the dorsal plate (crus brevis)
1. perpendicular 2. acute angle of the crus brevis and crus longus
262. Primary suspension of the incus-quadrate on the basicranium:
0. by squamosal and quadratojugal 1. by squamosal only 2. by petrosal (either by the preserved direct contact of incus, or by the inference from the
presence of a well-defined crista parotica 263. Quadratojugal notch in the squamosal:
0. present as an independent element in adult 1. absent
264. Morphology of the stapes:
0. columniform-macroperforate 1. columelliform-imperforate (or microperforate) 2. bicrurate-perforate
265. Bony secondary palate:
0. ending anterior to the posterior end of the tooth row 1. level with the posterior end of the tooth row 2. extending posterior to the tooth row
266. Relationship of the maxilla to the subtemporal margin of the orbit:
0. participating in the rounded subtemporal margin of the orbit 1. forming a well defined edge along the subtemporal margin
267. Pterygopalatine ridge:
0. present 1. absent
268. Transverse process of the pterygoid:
0. present and massive 1. present as the hamulus 2. greatly reduced or absent
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 35

269. Palatal width anterior to the basisphenoid:
0. very narrow anterior to the basisphenoid 1. intermediate width anterior to the basisphenoid 2. palatal width is as broad at the basisphenoid as the internal choanae
270. Vault of the naso-oral pharyngeal passage near the pterygoid-basisphenoid junction:
0. roof of the pharynx is V-shaped in transverse section, narrowing toward the basisphenoid
1. roof of the pharynx is U-shaped in transverse section 271. Complete ossification of the orbital floor:
0. absent 1. present
272. Pattern of orbital mosaic as exposed externally:
0. alisphenoid contacts the frontal and parietal 1. the petrosal anterior lamina contacts the orbitosphenoid, thereby separating the
alisphenoid from the front and the parietal 273. Outline of the facial part of the lacrimal:
0. large, triangular and pointed anteriorly 1. small and rectangular or crescentic 2. excluded from the facial (and pre-orbital) part of the skull
274. Pila antotica:
0. present 1. absent (in adult)
275. Fronto-parietal suture on the alisphenoid:
0. dorsal plate of alisphenoid contacting the frontal by the anterior corner 1. dorsal plate of alisphenoid has more extensive contact to the frontal (aprox. 50% of its
dorsal border) 276. Jugal on the zygoma:
0. anterior part of the jugal extends on the facial part of the maxillary and forming a part of the anterior orbit
1. anterior part of jugal does not reach the facial part of the maxillary and is excluded from the anterior part of the orbit
277. Maximum vertical depth of the zygomatic arch relative to the length of the skull:
0. between 10-20% 1. between 5-7%
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 36

278. Posterior opening of the post-temporal canal: 0. at the junction of the petrosal, squamosal and the tabular 1. between the petrosal and the squamosal
279. Anterior ascending vascular channel for the arteria diploetica magna in the temporal region:
0. open groove 1. partially enclosed in a canal 2. completely enclosed in a canal or endocranial
280. Lambdoidal crest:
0. crest overhanging the concave or straight supraocciput 1. weak crest with convex dorsal part of the occipital plate
281. Sagital crest:
0. prominently developed 1. weakly developed 2. absent
282. Tabular bone:
0. present 1. absent
283. Shape of the occipital condyle:
0. bulbous 1. ovoid 2. subcylindrical
284. Occiput slope:
0. occiput slopes posterodorsally, or vertically from the occipital condyles 1. occiput slopes anterodorsally from the occipital condyles
285. Foramina on the dorsal surface of the nasal:
0. absent 1. present
286. Septomaxilla:
0. present, with a ventromedial shelf 1. present, without the septomaxillary shelf 2. absent
287. Premaxillary internarial process:
0. present 1. absent
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 37

288. Facial part of the premaxillary borders on the nasal: 0. absent 1. present
289. Ossified ethmoidal cribriform plate of nasal cavity:
0. absent 1. present
290. Posterior excavation of the nasal cavity into bony sphenoid complex:
0. absent 1. present, confluent with the nasal cavity 2. present and partitioned from the nasal cavity
291. External bulging of braincase in parietal region:
0. absent 1. expanded, the parietal part of the cranial vault is wider than the frontal part, but
expansion does not extend to the lambdoidal region 2. greatly expanded, expansion of cranial vault extends to lambdoidal region
292. Interparietal:
0. present as a separate element in adult 1. absent
293. Bony tentorium septum:
0. present 1. absent
294. Overall size of the vermis:
0. small 1. enlarged
295. Lateral cerebellar hemisphere (excluding the paraflocculus):
0. absent 1. present
296. Lateral extension of the paraflocculus:
0. less than 30% of total cerebellar width 1. more than 30% of the cerebellar width
297. External division on the endocast between the olfactory lobe and the cerebral hemisphere
(circular sulcus): 0. absent 1. present
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 38

298. Anterior expansion of the cerebral hemisphere: 0. absent 1. developed
299. Expansion of the posterior cerebral hemisphere:
0. absent 1. present
300. Prism shape:
0. absent 1. arc 2. enclosed
301. Prism seams:
0. present 1. absent
302. Prism packing:
0. hexagonal 1. erratic 2. in rows
303. Interprismatic matrix:
0. on all sides, widely separated prisms 1. distinct inter-row sheets 2. prisms "shoulder to shoulder", little IPM
304. Outer aprismatic zone:
0. present 1. absent
Modified Nexus File
To facilitate the reading of the matrix the multistate characters have been replaced by letters as follows:
Multistate values: A:(01) B:(12) C:(123) D:(23) E:(012) F:(13) G:(02)
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 39

#NEXUS
[ File saved by NDE version 0.5.0, Thu Jun 23 17:49:01 2011 ] BEGIN TAXA; DIMENSIONS NTAX=57; BEGIN CHARACTERS; DIMENSIONS NCHAR=304; FORMAT DATATYPE=STANDARD MISSING=? GAP=- SYMBOLS="012345"; MATRIX Probainognathus 0C000000?0 0000??0000 00?0?00000 0000000?0? ?????????? 0?00?00??? ?????00??0 ?00?0000?? ?00??????? ?200???0?0 00001100?? ????0000?? ??0010?0?0 0?000???0? 0??????000 0?00?0???? 0?000000?0 0000000000 000?0??000 000?00000? ??00???0?1 00000??000 ??00000000 0000001101 0000?000?0 0?00000000 0000000000 0000000000 0000000000 000?000000 0??0 Tritylodontids 0C000010?0 0000??0000 00?0?300?1 10?2??1??0 ??????0?0? ??????4??? ?????00??0 ?00??0???? ?????????? ??00?????1 00511402?? ????2????? ??????D0?1 20011???0? 3??????000 0000?00000 0000000000 000000A000 000?000000 0000000000 00000??001 00000??000 ??00000001 ?000100000 0101?100?0 0000100010 0000000A00 1000000000 0000000A01 010?000000 0??0 Tritheledontids 1?000000?0 00000?0000 0001?20000 00001?0?0? ????????0? 0?00?00??? ?????00??0 ?00?0?000? ?00??????? ?240???0?0 0?001100?? ????000??? ??0010?0?0 0?001???0? 0??????00? 0??0?00?11 01000000?0 0000000000 000?0??0?0 00???0???? ?????????1 000100?000 ??0?000001 ?000000100 0000?000?0 0000000100 ?110100000 0000000000 0010?001?0 01???????1 0001 Adelobasileous 1????????? ?????????? ?????????? ?????????? ?????????? 0??0?????? ???0?00??? ?????0???? ?????????? ???????0?? ??0??0???? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? 10??????01 ?0????0000 0100201011 0000?00000 000001???? 0?????0?11 10?00??110 1120?????? 01???????? ???? Sinoconodon 1?000000?0 0000000000 0011000000 00001?1010 0100000000 0100?00000 ???0010A00 000100000? ?000??0??? ?200???0?0 0?101000?0 ?0?0000??? ??00A020?A 10000???00 1?00000000 00?00?0000 0000000000 0000000000 000????000 000??0???? ?????????1 0001000011 0000000000 0111200000 0101?00000 0100111000 0110110110 1000001100 0000000001 0100000001 0100 Morganucodon 10000100?0 0000000000 00A1000010 00?01?1300 1000000001 0100?11000 ???0012001 202000000? ?000??0??? ?240???0?0 0?10100100 ?0?0000??? ??000020?1 10010???00 1?00000000 0000?10?1?
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 40

0010?000?0 0000000000 000?000010 0010?0?000 00000????1 00110AA012 1101000000 0111201010 0112010000 0101110110 ?210110110 1000001100 1110110001 0100000001 0100 Megazostrodon 10000100?0 0000000000 00020110?0 00?01??300 1010000001 0100?12000 ???0012000 102000000? ?000??0??? ?240???0?0 0?10100100 ?0?0000??? ??000020?1 10000???00 1?0000000? 0000?1???? 0?100000?0 0000000000 0?00000010 00?0000000 0000?00001 0?110??012 01010?0000 0111101?10 0112010000 01011101?0 ?21?11?11? 10??001?00 1?10?????? ?1???????1 0001 Dinnetherium 1000011100 0000000000 00220110?0 ?0?0???300 1000000001 0100?11020 ???0012011 201000000? ?000??0??? ?240???0?0 0?10100100 ?0?0000??? ??000020?1 10010???00 1?0000000? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ????????12 110???0000 01111?1?10 0112010000 0101110?10 ?21??1??1? ?????????? ?????????? ?1???????1 0100 Haldanodon 10000010?0 0000000000 0011011010 00001??100 1000000010 0100?11??0 ???0012002 100000000? ?000??0??? ?210???0?0 0150130100 ?0?00100?? ?0000020?1 100100001? 100000000? ?????????? ??100????0 0?00?01000 00?????0?0 00?0?????? ?????????1 0012011012 0001000000 0111001??? 0102010000 0110110??? ?2?0211221 1000000?10 1111110000 000???10?0 0??0 Hadrocodium 1?101???30 0?10000?00 000102???0 ?0?01??0?0 ?1100010?0 0100?12000 ???001?000 000000000? ?000??0??? ?200???0?0 0?10100100 ?0?0000??? ??000010?1 100?0???00 1?0000000? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????1 1002101012 12????0000 0111?11021 0002110000 010?11???? ?2??211221 1001001111 212101?00? 21??11111? ???? Kuehneotherium 10000110?0 0000000000 00?10010?0 ?01???13?0 ?010000001 0?12012020 0?01111011 2010100000 0000000??? 0110??00?0 0020010111 00?0010000 ?0100030?A 1001??0010 1??00?000? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????0 0??? Pseudotribos 1F00011100 00?00000?? 001?1001?0 ?0?11??310 0100100111 1021112010 0?01101012 2000100000 0011000??2 0020010000 0030120211 00?0110010 00100020A1 100?100010 110000000? ????000000 0000101000 0000001000 ????0?10?0 00???????? ?????????1 ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Ambondro ???0??1??? ?????????? ?????????? ?????????? ?0?0200011 ??21?131?1 1021?00011 200?100002 114110100B 1????????? ??301?0211 ?11?110?00 ???????0?1 100???0010 ???00??0?? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Ausktribosphenos 1010011121 A010001001 0??11201?? ???????3?0 1101210011 ??21?13102 1221?00001 2000200002 1141102112 3????????? ??501?0212 ???0110?11 ??????10?1 100?????00 ???00?00?? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ????
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 41

Henosferus 1000011121 A000002001 00011211?0 ?0?1???3?0 01001000?0 0?22?13121 1021?00110 0010100002 114110BA02 1????????? ?0301?0211 ?100110?00 ?01???10?1 100?00??00 ??000000?? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????0 1??1 Asfaltomylos 10000?1121 0??0002001 01A11211?? ?????????0 ???0100011 ??21?13??1 12?1?00110 0011100002 114110B002 1????????? ???01?0211 ??00110?10 ??1???10?1 100???0000 ???00000?? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Bishops 1010111121 1010001001 01111211?? ?0?????3?0 1101210011 ??21?13102 1221?00001 2001100002 1141102112 3????????? ??501?0212 ???0110?11 ??????10?1 100???0000 ???00?00?? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Stereopodon 1E?0101??? ?????????? ??????01?? ?????????? ??????0??? ??21?13112 11?1?00101 1200200012 2031100??2 3????????? ??501?0212 ?210020?00 ??????10?1 100???0011 ?0??0??0?? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Teinolophos ??10111121 0010002001 11111201?? ?????????? ?????????? ??21?13112 11?1?00001 12012000B2 2031100??2 3????????? ??501?0212 ?210020?00 ??????20?1 100??20011 ???????0?? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Obdurodon ??10201031 0011002011 10?11211?1 ???2???0?0 0110200110 0121?13112 11?1?00101 A200210?02 2031100??2 301??????? 0?50140212 ?210G20010 0?100010?1 200??20011 3?00000001 ?????????? ?????????? ?????????? ?????????? ?????????? ?????????0 0001110012 12????0000 1111111121 0012010000 000000???? ?2??211221 1121?11101 212111101? 211??????2 121? Ornithorynchus 10102?10?1 111100201? 11111?11?1 ???2???0?? ??????0??? 01???1???? ?????0???1 ?200210??2 20?1100??2 301??????? 0050040??? ????G2???? ??100010?1 2001?2??0? 30??00?11? 1101110000 0000101000 0000001000 000?111000 0020100000 0002111110 0001010012 1212000000 1111111121 0012010000 0000002012 ?211211221 1121?11101 2121111000 2111101110 1??? Gobiconodon 13101110?? ?0?1102011 02110100?1 10111?1200 01?0100001 0100?12010 ???0111010 001000?00? ?0000?0??? ?210?????0 0010100110 ?0?00000?? ???0202011 10001???00 1?0000000? ????00???? 1?21?1?1?1 1100001100 00??000000 00100100?? ????????11 0012011012 ?10?0?000? ?1?1?11??? ????0?0??0 0????????? ????21???? ??0??10120 01?0?1?0?? 01???????1 0100 Amphilestes 13101110?? ?1?110200? 02110210?0 ?0?0??1200 0110100001 0?00?12010 ??00?11010 001000?00? ?0000?0??? ?2???????? 0?10100110 ?0?0000??? ??????20?1 100?????00 1?000000?0 ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????1???? ?????????? ?????????? ????
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 42

Jeholodens 1?10B?10?? ???110?0?1 0221010??0 10?11??010 ?100000001 0100?12010 ???00110?0 001?00000? ?000000??? ?2A0?????0 0?10?00110 ?0?00000?? 00?0G02011 100?1???00 1?0000000? ?0??10???? 112111??01 110??01101 00000?0000 001000001? 000010100? 0012?110?? ?A?1?????? ?????????? ?????????? ????????12 2????1???? ??0??11??? 11??01?0?? 1????????? ???? Priacodon 11101010?? ?1?1102011 02210110?0 10?000?100 0010101002 0100?11110 ???0011100 003100?00? ?0000?0??? ?210?????1 1?10100100 ?0?00000?? 00?0002021 10010???00 1?0000000? ?????????? ?????????? ??????1101 00???????? ?????????? ?????????1 ?????????2 1101??0000 01111?1110 0??20??00? ?10121???? ?2??21???? ???????11? ??2??????? ?????????1 0100 Trioracodon 11101010?? ?1?1102011 022101001? ?010??1200 0010100002 0100?11110 ???0011A00 003100?00? ?0000?0??? ?210?????1 1?10100100 ?0?00000?? 00?0001021 10010???00 1?0000000? ?????????? ?????????? ?????????? ?????????? ?????????? ???0?????? ????????12 1101??0000 01111?1110 011201?00? ?1012????? ?2???1???? ???????11? ?????????? 1?1101110? ???? Tinodon 10101110?? ?0?11010?? 02111311?0 ?0?1???100 0100100001 0?12?12010 0?01101011 2010100000 0010000??0 0110??00?0 0020010111 ?0?001010? 0010002021 100??0??00 1??00?00?? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Peramus 1B10111111 0011002101 00111211?0 ?010???300 0011101001 0121013120 1001100010 001010A000 1021110??0 1010000000 0030110211 110011010? 0011A01011 100??0??00 100000000? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????1 ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Vincelestes 12102?1111 0011001101 01111211?1 101000?000 0010101011 0122013010 1001100100 0000100001 01210000?0 10110000?0 0030110211 1100110000 0010001021 100?10??00 2100000100 1111?11211 1121111101 1110100111 1000010111 0011101011 00121??0?1 1002111012 1210?11110 0111111021 1012120110 011021???? ?21?110?11 101000A120 01211110?? 11???????1 0000 Kielantherium 1B10101111 00?1001111 0???1311?? ???0???200 001000?000 0?21013020 1011100A10 0010100001 11411000?0 1001000000 0030110212 1101120100 00110020A1 100??0??00 21?00??0?? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Deltatheridium 13102?1131 0111011101 01131311?0 1110001100 0010101001 0121013120 1011100110 0011100001 1141111010 10210A1000 0030120212 2101120100 0011002011 100100??00 210000000? ?????????? ?????????? ?????????? ?????????? ???????121 1?12?????1 11?2111112 1222??1110 02123221?? ?00212?000 111021???? ?21?211110 101??0112? 1?200211?? 1????0???1 0000 Asiatherium 13101?1131 0111011101 011?1311?? 11??00?100 0010101001 0121113100 1211100110 0111101002 1141111111 3011022000 0030120212 2101120010 0011002001 100??0??00 210000000? ???101????
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 43

11210111?2 1111120221 111?0101?1 11111101?? ???????201 1102111112 122???111? ?2?2322??? ?00?121??1 111??1???? ?21?211110 10???01??? 11200????? 21???????? ???? Kokopellia 13?01?113? ?1?1??1111 0????301?0 ?0?0???1?0 0010101001 ??21113110 1211100110 0111101002 1141111110 3021012000 0030120212 2101120010 00210020A1 100100??00 21?00?000? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Pucadelphys 13101?1131 0111011101 02131311?0 1110001100 0010101001 0121113100 1211100110 0111101002 1141112111 3031022110 0030120212 2101120010 0021002001 100100??00 2100000001 1111011211 1121011112 1111120221 111?110111 1111110121 1112020?01 1102111112 1222??1110 02123221?? ?002121000 111021???? ?21?211110 1011101120 11200211?2 10???????? ???? Didelphis 12102?1131 0111011101 0213130110 1110001111 0010101001 0121113100 1211100110 0111101002 1141112111 3031020110 0030120212 2101120010 0020002001 1001000?00 2100000001 1111011211 1121011112 1111120221 1111110111 1111110121 1112020201 1102111112 1222111110 02123221?? ?002120001 1110211111 1212211110 1011101120 0120021112 1001101111 1221 Pappotherium ???0??1??? ?????????? ?????????? ?????????0 ?010001000 ??21013100 1011100110 001?100001 1141111100 2031001000 0030120212 2101110110 001100?001 100??0??00 21??00000? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Erinaceous 11102?1111 0111001101 0113131111 10111?1000 0001100000 1121113000 1211100100 0010100002 1141101111 3011122000 0050120212 2111120010 0010001021 1001101110 2100000001 1111011211 1121011112 1111120221 0101110111 1121101221 2111020201 1102111112 1222111121 ?2123221?1 1002120110 0110211111 1212211110 1011101120 1121021112 2101101111 1221 Asioryctes 1B102?1131 0011001101 01131311?0 0010001200 0110100000 1021113000 1011100110 0012100001 1141111100 3021112000 0030120212 2101120110 0011001001 100100??00 2100000000 1111?????? ?????????? ?????????? 0101?????? ????10122? 2111020?01 1102111112 122???1121 ?2123221?1 1002120100 011021???? ?21?211110 1011101120 11210211?? 110110111? ???? Prokennalestes 1B10101111 0011001111 00131211?0 ?0?01??300 0100100000 1021113000 1011100110 0012100001 1141111100 3031111000 0?30120212 2101110110 0011001001 1001?00?00 210000000? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????1 ????????12 1222111120 1212??2121 10021??11? ?11021???? ?2???????? ???1?????? ?????????? ?????????1 0000 Montanalestes 13102?1131 0011001101 01131311?? ?0?????300 0110100010 ??21?13010 1011100111 0012100001 1141111100 3????????? ??301?0212 ?101120?10 ??1???10?1 100?????00 2??00?00?? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ????
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 44

Zhangheotheriids 12101010?? ?0?1102001 02111011?0 10111?1110 1100100101 0122012010 0?01101001 2010100000 0000000??0 0020??00?0 0042120111 00?011000? 0011103011 1011110010 100000000? 1100010101 1121111101 11101A1111 0000000111 1011000011 001211?211 0012111012 11????1110 01?11?1121 0012120000 01?121???? ?21??1???? 1?0??01??? 1???0111?? ?????????? ???? Amphiteridae 1011111111 0011002001 01111211?0 ?0?0???3?0 0010100010 ??21?13100 1001100110 0010100100 1021110??0 1110??00?0 0040110211 110011010? 00110030?1 100??0??00 200000000? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Henkelotherium 1011101111 0011002001 0111130110 ?010??1100 0000101011 0121012100 0?01100100 0000100100 0000000??0 0000??0000 0040121211 10?012011? 0010103021 1001000010 2000000000 ?????1???? 112111?1?1 1110?00111 000??00111 10111010?? ??1?????10 1?12?11012 121111?110 A11A???121 ?002??011? ?110B????? ????B1???? ??11?01??? ?????????? ?????????1 000? Dryolestes 1011111111 0011002001 0111130110 00101?1200 0000101011 0121012100 0?01100100 0000100100 0000000??0 0000??0000 0042121211 10?012011? 0120103021 1111000010 200000000? ?????????? ?????????? ??????1111 00???????? ?????????? ?????????1 ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????1 0000 Laolestes 1011111111 0011002001 01111301?0 ?010???2?0 0000101011 0121012100 0?01100100 0000100100 0000000??0 0000??0000 0142121211 10?012111? 1121103021 1111110010 200000000? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????1 ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????1 0000 Amblotherium 1011111111 0011002001 01111301?0 ?010???200 0000101011 0?21012100 0?01100100 0000100100 0000000??0 0000??0000 0142121211 10?012011? 0011?03011 1111110010 200000000? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Comotherium ???1?????? ????????0? ?????????? ?????????? ?????????? ????012??? ????10???? ?????0???? ?????????? ?000??0000 004??21211 10??1??1?? 0021003011 11?1?1???? 200??0??0? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Mesungulatum ??11??1??? ?????0???? 0????????? ???0?????2 ?1012?2011 0121012100 0?01100002 0200100010 0000000??0 00A0??00?0 ?151121211 10?0?1111? 1220111021 101??01110 20???1100? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Coloniatherium 11112?1111 01?1001101 0????310?? ??1000?102 1101212001 0121012100 0?01100002 0200100010 0000000-?0 0000??00?0 0151121211 10?011011? 1220111021 1011?01110 201111100? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????2 1220??11B0 0111??1111 010212?11? 01112????? ?2???????? ?????????? ?????????? ?????????1 0000
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 45

Reigitherium 11112?1??? ?1???01??1 0????211?? ???A???B?1 0111211000 01210121?0 0?01000102 0200100020 0000000??0 0040??00?0 0051121210 10?0110?1? 122001B121 1011??1120 20?11??00? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????2 12201111B0 0111?????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Peligrotherium 10112?1111 0111001101 01?F1310?? 1?1000?102 1111212102 0121012100 0?01100002 0200100010 0000000??0 0040??00?0 0151141211 10?01B011? 1220111121 1011001120 201111000? ?????????? ?????????? ?????????? ????10???? ?????0???? ?????????0 ??0?0???12 1220??1100 111BB21101 011B?2?11? 01112????? ?2??21?2?? 1?0??00120 01?10A11?B 11??0011?? ???? Groebertherium ???1?????? ?????????? ?????????? ?????????? ?????????? 0?21012100 0?01100101 010?100100 0000010??0 0000??00?0 0A42121211 10?0?2101? 022111?0A1 111???0110 20????0??? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????0???? ???? Leonardus ???12?1??? ?????????? ?????????? ?????????0 ?111011002 0121012100 0??1100100 0000200000 00000?0??? ?000??00?0 0142121211 10?0?2001? 122011B021 111??1000? 100?10?11? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????0 ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Foxraptor 0??01011?? ?1???01??1 0?????10?0 ?0?0???100 0111101001 ??21013100 0?01?00100 000B100100 0020000??2 0????????? ??500?0211 ?0?0?20?1? ???0??20?1 100?0???00 ??000?00?? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Paurodon 0?101111?? ?0?1?01??1 0????D11?0 ??10???010 0011101011 0?21013000 0?01100100 0000100100 0020010--2 0??0-????? ??40120211 10?0?20?1? ??????20?1 100?????00 ??000?01?? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ???? Cronopio 10112?1131 0111001001 02111110?? 1?1001?111 0011201002 0021012100 0?01100100 0000200010 0000000--0 0000--00?0 0042021211 10?012011? 0020111020 0211?10?00 000000111? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????0 0011011112 12B0??1110 A11???1011 010212?11? 01112????? ?21G0???0? 100?100??? ??1?0A10?? 2????01?1? ???? Drescheratherium ?????????? ?????????? ?????????? 0??101?11? ?????????? 00??01???? ????10???? ???010???? ?????????? ?000??00?0 01???2?211 1??????1?? 1110002001 1????10?0? 200??0??0? ?????????? ?????????? ?????????? ?????????? ?????????? ?????????1 ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? ?????01??? ?????????? ?????????? ???? ; ENDBLOCK; BEGIN ASSUMPTIONS; OPTIONS DEFTYPE=UNORD POLYTCOUNT=MINSTEPS;
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 46

TYPESET * default = ORD: 2 5 26 38 40 47 53 - 54 62 75 79 - 80 90 96 - 97 110 - 111 116 122 127 130 - 132 139 141 163 170 176 178 - 179 193 199 201 220 222 - 223 229 232 234 236 265 268 - 269 273 279 281 283 286 291 300 303; ENDBLOCK; BEGIN NOTES; [Taxon comments] TEXT TAXON=45 TEXT='Melanodon is a junior synonym'; TEXT TAXON=46 TEXT='Kurtodon is a junior synonym'; TEXT TAXON=53 TEXT='MACN RN 1097(Chornogubsky, 2011) is referred as lowers of Leonardus';
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 47

Part 2: Dental position and taxonomic diversity of other Late Cretaceous South
American dryolestoids.
Because of its completeness Cronopio serves as a model in reinterpreting some of the dental
homologies and redundancies among the Late Cretaceous taxa that have been erected based mostly
on isolated teeth (Bonaparte and Soria, 1985; Bonaparte, 1986; 1990, 1992, 1994, 2002; Rougier et al.,
2009a, b).
We argue here that the taxa from the Late Cretaceous Los Alamitos Formation described by Bonaparte
as Quirogatherium and Barberenia (figue 2 SI, C, E) are in fact superior premolars of other taxa which
were described based on molars. The morphology of these premolars is characterized by a mesial lobe
with a low cusp and a small sub-circular root. The distal portion of the tooth is dominated by two cusps
that determine a narrow and oblique basin. In the more derived, herbivorous, mesungulatid
dryolestoids a distal cingulum is present. The overall morphology is similar to the deciduous premolars
of Late Jurassic dryolestoids from Europe and North America (Martin, 1999), a point noted by Martin
(1999) and supported by Kielan-Jaworowska et al. (2004) and tentatively by Bonaparte and Migale
(2010). This is a reasonable interpretation of the morphology of Quirogatherium and relatives;
however, the frequency of these teeth in the collection from Los Alamitos Fm. is too high to regard
them as shed deciduous teeth. Cronopio and the Paleocene dryolestoid Peligrotherium show that teeth
with a morphology reminiscent of that present in the deciduous premolars of Jurassic dryolestoids
were retained into adulthood. The Late Cretaceous dryolestoid Coloniatherium (Rougier et al., 2009b)
also presents a similar morphology (figure S3, D). The presently available material of either Cronopio or
Peligrotherium does not allow us to determine if the tooth in question is a retained deciduous or a
second generation tooth. The position of the tooth in question (P3 in Cronopio, P2 in Coloniatherium
and Peligrotherium) coincides with a strong narrowing of the rostrum. It is likely that this highly derived
morphology is related to a proportionally slender snout, a feature proportionately retained in the
otherwise massive Peligrotherium.
Given that most taxa from Los Alamitos Fm. are based on isolated teeth, it is hard to determine exactly
what premolars correspond to a given molar morphology. It has been suggested that Quirogatherium
is probably a junior synonym of Mesungulatum (Martin, 1999; Kielan-Jaworowska et al., 2004); when
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 48

the teeth assigned to Quirogatherium are compared with the molars of Mesungulatum (using either
Cronopio or Peligrotherium as reference) it appears that Quirogatherium is too small. Mesungulatum
and allies have low bunodont cusps covered by thick enamel; a tooth regarded as undetermined by
Bonaparte and Migale (2010, page 336) fits the morphology and size expected for Mesungulatum. It is
therefore likely that both Quirogatherium and Barberenia are upper premolars of smaller dryolestoids
from Los Alamitos Fm.
Figure S3. Deciduous upper premolar of a Late Jurassic dryolestid (A) and a variety of penultimate
upper premolars in SA dryolestoids (B-F). A, Dryolestes dP3; B, photo and line drawing of the P3 of
Cronopio; C, Barberenia; D, Coloniatherium; E, Quirogatherium; F, Peligrotherium. A, modified from
Martin, 1999, C and E modified from Bonaparte (1990).
Similarly, Bonaparte (1986, 1992) described a handful of isolated teeth from Los Alamitos as
triconodonts and erected for them the family Austrotriconodontidae. The main features differentiating
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 49

them is the presence of a hypertrophied central cusp a and very reduced posterior cusps. This
morphology is also present among upper (figure S43) and lower premolars of some of the dryolestoids
from the Cretaceous of Argentina. An upper molar purportedly showing a groove on the mesial root
and a palisade arrangement of minor cusps is in our opinion damaged and the morphology as
presented artifactual. It is likely that most, or all, of the teeth described as triconodonts from the Late
Cretaceous Los Alamitos Fm. are in fact dryolestoid specimens and the only bona fide triconodonts
from SA would be those from the Jurassic of Patagonia (Rougier et al., 2007; Gaetano and Rougier,
2011).
Figure S4. A, Austrotriconodon, A1 Austrotriconodon sepulvedai; A2 Austrotriconodon mckennai, both
described as a lower molar; B, Coloniatherium silinskii, premolar; C, Cronopio dentiacutus, C1 upper
premolar, C2 lower premolar (A1 modified from Bonaparte,1992; A2 modified from Bonaparte 1986)
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 50

Bibliography
Bonaparte, J. F. Sobre Mesungulatum houssayi y nuevos mamíferos cretácicos de Patagonia. Actas IV
Congreso Argentino de Paleontología y Bioestratigrafía 2, 48–61 (1986).
Bonaparte, J. F. New Late Cretaceous mammals from the Los Alamitos Formation, northern Patagonia.
National Geographic Research 6, 63–93 (1990).
Bonaparte, J. F. Una nueva especie de Triconodonta (Mammalia) de la Formación Los Alamitos,
Provincia de Río Negro y comentarios sobre su fauna de mamíferos. Ameghiniana 29, 99–110
(1992).
Bonaparte, J. F. Approach to the significance of the Late Cretaceous mammals of South America.
Berliner geowissenschafliche Abhandlungen E 13, 31–44 (1994).
Bonaparte, J. F. New dryolestidae (Theria) from the Late Cretaceous of Los Alamitos, Argentina and
paleogeographical comments. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen,
224, 229–271 (2002).
Bonaparte, J. F. & Migale. Protomamíferos y mamíferos Mesozoicos de América del Sur. Museo de
Ciencias Naturales Carlos Ameghino (2010).
Bonaparte, J. F. & Soria, M. F. Nota sobre el primer mamífero del Cretácico Argentino, Campaniano–
Maastrichtiano (Condylarthra). Ameghiniana 21, 178–183 (1985).
Chornogubsky, L. New remains of the dryolestoid mammal Leonardus cuspidatus from the Los Alamitos
Formation (Late Cretaceous, Argentina). Paläontologische Zeitschrift 85, 343-350 (2011).
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 51

Cifelli, R. L. & Madsen, S. K. Spalacotheriid symmetrodonts (Mammalia) from the medial Cretaceous
(Upper Albian or lower Cenomanian) Mussentuchit local fauna, Cedar Mountain Formation,
Utah, USA. Geodiversitas 21, 167–214 (1999).
Corbella, H., Novas, F. E., Apesteguía, S. & Leanza, H. A. First fission track-age for the dinosaur-bearing
Neuquén Group (Upper Cretaceous) Neuquén Basin, Argentina. Revista Museo Argentino de
Ciencias Naturales (N.S.) 6, 1–6 (2004).
Gaetano, L. C. & Rougier, G. W. New materials of Argentoconodon fariasorum (Mammaliaformes,
Triconodontidae) from the Jurassic of Argentina and its bearing on triconodont phylogeny.
Journal of Vertebrate Paleontology 31, 829-843 (2011).
Garrido, A. C. 2010. Estratigrafía del Grupo Neuquén, Cretácico Superior de la Cuenca Neuquina
(República Argentina): Nueva propuesta de ordenamiento litoestratigráfico. Revista del Museo
Argentino de Ciencias Naturales, Nueva Serie, 12 (2).
Goloboff, P. A. Analyzing large data sets in reasonable times: Solutions for composite optima. Cladistics
15, 415–428 (1999).
Goloboff, P., S. Farris, and K. Nixon. 2005. TNT (Tree analysis using New Technology) (BETA) ver. 0.2
Published by the authors, Tucumán, Argentina.
Kielan-Jaworowska, Z., Cifelli, R. L. & Luo, Z.-X. Mammals from the Age of Dinosaurs—Origins,
Evolution, and Structure (Columbia Univ. Press, New York, 2004).
Leanza, H. A., Apesteguía, S., Novas, F. E. & de la Fuente, M. S. Cretaceous terrestrial beds from the
Neuquén Basin (Argentina) and their tetrapod assemblages. Cretaceous Research 25, 61–87
(2004).
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 52

Luo, Z. X., Kielan-Jaworowska, Z. & Cifelli, R. L. In quest for a phylogeny of Mesozoic mammals. Acta
Paleontologica Polonica 47, 1–78 (2002).
Martin, T. Dryolestidae (Dryolestoidea, Mammalia) aus dem Oberen Jura von Portugal. Abhandlungen
Senckenbergische Naturforschende Gesellschaft 550, 1–119 (1999).
Nixon, K. C. The Parsimony Ratchet, a new method for rapid parsimony analysis. Cladistics 15, 407–414
(1999).
Ramos, V. Late Proterozoic-Early Paleozoic of South America. A collisional history. Episodes 11, 168–
175 (1988).
Rougier, G.W., Chornogubsky, L., Casadio, S. Paéz Arango, N. & Giallombardo, A. Mammals from the
Allen Formation, Late Cretaceous, Argentina. Cretaceous Research. 30:223-238 (2009).
Rougier, G. W., Martinelli, A. G., Forasiepi, A. M. & Novacek, M. J. New Jurassic mammals from
Patagonia, Argentina: a reappraisal of australosphenidan morphology and interrelationships.
American Museum Novitates 3566,1–54 (2007).
Rougier, G. W., Forasiepi, A. M., Hill, R. V. & Novacek, M. J. New Mammalian Remains from the Late
Cretaceous La Colonia Formation, Patagonia, Argentina. Acta Paleontologica Polonica 54, 195–
212 (2009).
Sigogneau-Russell, D. Découverte du premier Symmétrodonte (Mammalia) du continent africain.
Comptes Rendus de l'Académie des Sciences, Paris 309:921-926 (1989).
Sigogneau-Russell, D & Ensom.P.C., Thereuodon (Theria, Symmetrodonta) from the Lower Cretaceous of North Africa and Europe, and a brief review of symmetrodonts Cretaceous Research 19:445-470 (1998).
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 53

Wible, J. R., Rougier, G. W., Novacek, M. J. & Asher, R. J. Cretaceous eutherians and Laurasian origin for
placental mammals near the K-T boundary. Nature, 447, 1003–1006 (2007).
Wible, J. R., Rougier, G. W., Novacek, M. J. & Asher, R. J.. The eutherian mammal Maelestes gobiensis
from the Late Cretaceous of Mongolia and the phylogeny of Cretaceous eutherians. Bulletin of
the American Museum of Natural History 327, 1–123 (2009).
SUPPLEMENTARY INFORMATIONRESEARCHdoi:10.1038/nature10591
WWW.NATURE.COM/NATURE | 54





























