Embed Size (px)
Citation preview

Supporting InformationReissner et al. 10.1073/pnas.0801639105SI Materials and MethodsMolecular Cloning. We generated a new control vector pCMV-GCfor producing secreted proteins with the human Fc region,consisting of signal peptide ASTLE-IgG1Fc with cloning sitesSalI-XbaI-BamHI after the signal peptide (1). Expression vectorpCMVIg�LNS6(-SS4) containing the Nrxn 1�LNS6 domain(boundaries: GTTYI … VGEVP) was generated by PCR frompCMVIg�-1 (1) and cloned with SalI and BamHI (Biolabs) intothe control vector, resulting in secreted fusion protein AST-(LNS6)-SSMTTIEGRE-IgG1Fc. The LNS6 domain of Nrxn 1�and the single LNS domain from Nrxn 1� were mutated withQuikChange (Stratagene) on templates pCMVIg�-1(-SS4) andpCMVIg�LNS6(-SS4). Mutations of cytosolic internal GFP-tagged full-length neurexin 1� (IENB; R.F. and M.M.) weregenerated by replacing PstI-SalI fragment in IENB with thosefrom mutated pCMVIg�1 L234S�I236S�N238D and pCM-VIg�-1 N238A, respectively. The expression vector of Nlgn 1without insert at splice site B, pCMVNL1-B, was obtained bydeleting the region coding for NRWSNSTKG in pCMVNL11(2). Site-directed mutations on pCMVNL1-B have been intro-duced by using QuikChange (Stratagene). The Nlgn 1 mutationH294V has been generated on pCMVNL11 containing spliceinsert B. All final constructs were verified by DNA sequencing.
IgG Control Vector. To avoid possible interference of a stretch ofhydrophobic residues present in an earlier version of the IgGcontrol vector (1), we generated a novel construct by deleting 16residues, using forward primer 5�-ATA AGC TTG ATA TCGAAT TCC GTG ATC TCT CTT GAG-3� (MM05–59) andreverse primer 5�-TGA AGC TTA GTA CTC ACC CTC GGGATC GGG ATC CTC TAG AGT CGA CGC CCC ACT GCCCAG CTC TGC CC-3� (MM05–90).
Expression Vector pCMVIg�LNS6(-SS4). The vector containing theNrxn 1�LNS6 domain (boundaries: GTTYI … VGEVP) wasgenerated by PCR from pCMVIg�-1 (1) using forward primer5�-GCG TCG ACT GGG ACG ACG TAT ATC TTT AGCAAA GGT GGT GG-3� (MM05–74) and reverse primer 5�-TCG GGA TCC TCT CTA CCT TCA ATA GTT GTC ATAGAG GAA GGC ACT TCA CCG ACC AGC-3� (MM05–75)and cloned with SalI and BamHI into the control vector,resulting in secreted protein AST-(LNS6)SSMTTIEGRE-IgG1Fc.
Site-Directed Mutagenesis Primers for NRXN Mutations. Residuestargeted and restriction enzyme site introduced for recognitionare in parentheses. 5�-CAG TTC CTC AGG CCT GGG TGCCTA CCT CGA GCT GCA CAT AC-3� (D137A, �XhoI,MM05–187), 5�-CAG TTC CTC AGG CCT GGG TCG GTACCT TGA GCT GCA CAT AC-3� (D137R, �KpnI, MM05–76),5�-GGA GTT AAA TTT AAT GTT GTG GCA GAT GATATC GCC ATT GAG GAG TC-3� (G155V, T156A, �EcoRV,MM05–78), 5�-GGG CGT CAG TCC ACA AGC TTC GATAGC CAA GCA ACC ATA ATA ATT G-3�(L234S,I236S,N238D, �HindIII, MM05–80), 5�-GGG CGTCAG CTC GCG ATC TTC AAT AGC CAA GCA ACC ATAATA ATT G-3� (T135A, �NruI, MM05–82), 5�-CAG GGCGTC AAC TCA CAA TCT TCG CTA GCC AAG CAA CCATAA TAA TTG-3� (N238A, �HindII, MM05–185), 5�-CAGGGC GTC AAC TCA CAA TCT TCC GTA GCC AAG CAACCA TAA TAA TTG-3� (N238R, �HindII, MM05–172), 5�-
CAG GGC GTC AAC TCA CAA TCT TCG GGA GCC AAGCAA CCA TAA TAA TTG-3� (N238G, �HindII, MM06–89).
Site-Directed Mutagenesis Primers for NLGN1 Mutations. Residuestargeted and restriction enzyme site introduced for recognitionare in parentheses. 5�-GGT TGA AGC TTG TGG AAA ACATAG TAG CTG TCT CTA ATT TTG TTG-3� (�EF1, �Hin-dIII, MM05–193), 5�-GTG ACC CAA ATC AAC CGG TACCTG AAG AAG TAG CAT GGA CCA G-3� (�L34, �KpnI,MM06–85), 5�-GTG ACC CAA ATC AAC CAC ATT CTCAAG AAG TAG CAT GGA CCA G-3� (�L34�HQS, �KpnI,MM06–85), 5�-GAA CAT CGA TTT CAG CCT CCT GAGTCA CCA TCT CCC TGG TCT GAT ATC-3� (P100S, �DdeI,MM07–50), 5�-GAT ATT CGC GAC AGT GGG GGT GCGAAA CCA GTG ATG GTG TAC ATC-3� (P192A, �NruI,MM07–86), 5�-GAT GGA CCA GCG AGA ACA TCG ATTTCT TTG GTG GTG ACC CCT TG-3� (G266D, �ClaI,MM07–52), 5�-CAT TGG GTT CTT TGG TGG TCG ACCCTT GCG AAT CAC CGT G-3� (D271R, �SalI, MM07–54),5�-GGT TCT TTG GTG GGG ATC CCG AGC GAA TCACCG TGT TTG GAT CAG G-3� (L273E, �BamHI, MM07–56),5�-CGA TTT CTT TGG TGG TGA CCC CGA TCG AAT CACCGT GTT TGG ATC-3� (G266D, L273D, � PvuI, MM07–82),5�-CTT TGG TGG TGA CCC CGG GCG AAT CAC CGTGTT TG-3� (L273G, � SmaI, MM07–84), 5�-GAT ACT GATGGA ACA AGG AGA ATT CTC CAA TTA TGA TAT CATGTT AGG-3� (L399S, �EcoRI, MM07–80), 5�-CCA AGAATG ATG TGA TGT TGA GCG ATC GAG TAA TGA CATACT GGA CG-3� (A557D, V558R, �PvuI, MM05–189).
The expression vector of Nlgn 1 without insert at splice site B,pCMVNL1-B, was obtained by deleting the region coding forNRWSNSTKG in pCMVNL11 (2), using QuikChange (Strat-agene) with primer 5�-GCT GAC TTT ATC CCA TTA TTCTGA AGG ACT TTT TCA ACG AGC AAT AGC-3� (-BstEII,MM05–191). The Nlgn 1 mutation H294V has been generatedon pCMVNL11 containing splice insert B by using the primer5�-GTC AAT CTG CTG ACT TTC TCA GTT TAT TCT GAAGGT AAC CGT TGG AGC-3� (H294V, � DdeI, MM07–58).
All final constructs were verified by DNA sequencing and areavailable from the authors upon request.
Protein Expression and Assays. pCMV vectors encoding solubleIgGFc fusion proteins of Nrxns and Nlgns were transfected intoCOS-7 cells using the DEAE method (1). For lysates, forebrainsof 6- to 12-week-old mice were homogenized with a Polytron inbuffer A (0.1 M NaCl/4 mM KCl/5 mM CaCl2/50 mM Tris�HCl,pH 7.5/0.1 �g/�l phenylmethylsulfonyl f luoride/1 �g/ml leupep-tin; Biomol). Triton X-100 (Fisher Scientific) was added to a finalconcentration of 1% (wt/vol), and lysis was performed for 1 h at4°C, followed by centrifugation at 120,000 � g. Supernatant wasadded to different IgG proteins in buffer A containing 0.1%Triton X-100, and binding was allowed to proceed for 1.5 h at4°C. After washing, bound proteins were solubilized in SDSsample buffer and analyzed by SDS/PAGE, Coomassie staining,and immunoblotting (Bio-Rad). Wild-type and mutated Nlgn 1expressed in COS-7 cells were analyzed accordingly by lysingcells in buffer B (50 mM Tris�HCl, pH 7.5/80 mM NaCl/2 mMCaCl2/1% Triton X-100). To assess Ca2� dependence, immobi-lized LNS-IgG fusion proteins were incubated with transfectedCOS-7 cell lysate in 50 mM Tris�HCl (pH 7.5), 80 mM NaCl, and1% Triton X-100 buffer. Different concentrations of free Ca2�
Reissner et al. www.pnas.org/cgi/content/short/0801639105 1 of 19

were calculated by MaxChelator (C. Patton, Stanford University,Stanford, CA) and adjusted by EDTA (Amersham)/CaCl2 (3).
Antibodies. Anti-Nlgn 4C12 antibody recognizes the extracellulardomain of Nlgn 1 (4), whereas antibodies 799 and 804 aremutually specific for the C terminus of Nlgn 2 and 3, respectively(kindly provided by Nils Brose, Max Planck Institute, Gottingen,Germany). Anti-neurexophilin (Nxph) 1 was raised in rabbitsusing peptide AQQTV IDAKD SKSC and affinity-purified(Eurogentec).
Calcium-45 Assay. Nrxn LNS-IgG fusion proteins were elutedfrom protein A beads (0.1 M glycine, pH 2.7/0.15 M NaCl).Supernatants were adjusted with 1 M Tris�HCl (pH 7.5) andblotted onto nitrocellulose paper using a Dot Blot 96 system(Whatman/Biometra). Each slot was washed with Ca2�-freewashing buffer (50 mM Tris, pH 7.5/0.15 M NaCl) and incubatedwith 30 �Ci calcium-45 chloride (PerkinElmer) for 30 min. Thenitrocellulose was washed with nonradioactive Ca2� buffer (5mM CaCl2/50 mM Tris, pH 7.5/0.15 M NaCl), and calcium-45binding was analyzed with a phospho-imager (Fuiji FLA-3000).
Cell Surface Labeling. Soluble Nrxn LNS-IgG fusion proteins wereincubated with Nlgn-GFP (Nlgn-A-GFP44)-expressing livetsA201 cells (ECACC) grown on coverslips in the presence of 2mM CaCl2. Cells were washed with 50 mM Tris�HCl (pH 7.5) and2 mM CaCl2, and fusion proteins were visualized with proteinA-conjugated Alexa Fluor 546 (Molecular Probes). Wild-typeand mutated proteins of Nlgn and EGFP-tagged Nrxn 1�(����) were also expressed in tsA201 for 48 h until their surfaceor whole cells were labeled with anti-Nlgn antibody. Results werevisualized using Axioskop 2 (Zeiss) epif luorescence and TCSSP2 (Leica) confocal microscopes.
Structural Analysis. SwissProt entries NRX1A�RAT (Q63372)and NRX1B�RAT (Q63373), containing all known alternativesplice inserts, were used to annotate LNS domains from Nrxn 1�and 1�, consistent with the annotation used for the crystalstructure of Nrxn 1� (5). NLGN1�RAT (Q62765) was used tomap and compare hot-spot residues on all three Nrxn/Nlgncrystal structures (PDB entries 2VH8, 3BIW, and 3B3Q). Neur-exin 1�LNS6 domain and its mutations were structurally alignedon coordinates from Nrxn 1�LNS (PDB entry 1C4R) andmodeled by using program SwissPDB Viewer (N. Guex, GlaxoSmithKline RandD, http://expasy.org/spdbv). Ca2� coordinationwas positionally refined using program X-plor (v3.1, A. Brunger,Howard Hughes Medical Institute, Department of MolecularBiophysics and Biochemistry, Yale University, New Haven, CT,http://xplor.csb.yale.edu/xplor/). The boundaries of the coredomain of LNS structures were derived by X-plor using struc-tural coordinates from laminin (1QU0, 1DYK, 1OKQ, and2JD4), agrin (1PZ7, 1PZ8, 1PZ9, and 1Q56), Gas6 (1H30), Nrxn(1C4R and 2H0B), SHBG (1D2S, 1KDK, 1KDM, 1LHN, 1LHU,and 1LHV), pentraxin/SAP (1SAC, 1QTJ, 1B09, 1GYK, 1LJ7,and 1GNH). Absolute values of each distance matrix werecolor-coded (RGB rainbow) for the range of 0 Å (blue) to 8 Å(red) using NIH ImageJ (http://rsb.info.nih.gov/ij). Electrostaticsurface maps for laminin 2�LNS4 (1DYK), laminin 2�LNS5(1QU0), agrin LNS3 (1PZ9), Gas6 LNS2 (1H30), and Nrxn1�LNS2 (2H0B) and 1�LNS (1C4R) were calculated by usingAPBS (N. Baker, Washington University, http://apbs.sourcefor-ge.net/). Free binding energies of wild-type and mutant Nrxn/Nlgn complexes have been calculated by using the FoldX pro-gram (http://foldx.crg.es/). Protein structures and surface mapswere visualized by using PyMol (DeLano Scientific, San Carlos,CA; www.pymol.org).
1. Missler M, Hammer RE, Sudhof TC (1998) Neurexophilin binding to alpha-neurexins. Asingle LNS domain functions as an independently folding ligand-binding unit. J BiolChem 273:34716–34723.
2. Nguyen T, Sudhof TC (1997) Binding properties of neuroligin 1 and neurexin 1betareveal function as heterophilic cell adhesion molecules. J Biol Chem 272:26032–26039.
3. Bers DMA (1982) A simple method for the accurate determination of free [Ca] inCa-EGTA solutions. Am J Physiol 242:C404–C408.
4. Song JY, Ichtchenko K, Sudhof TC, Brose N (1999) Neuroligin 1 is a postsynapticcell-adhesion molecule of excitatory synapses. Proc Natl Acad Sci USA 96:1100–1105.
5. Rudenko G, Nguyen T, Chelliah Y, Sudhof TC, Deisenhofer J (1999) The structure of theligand-binding domain of neurexin Ibeta: Regulation of LNS domain function byalternative splicing. Cell 99:93–101.
6. Sasaki T, et al. (2006) Structural basis for Gas6-Axl signalling. EMBO J 25:80–87.7. Grishkovskaya I, et al. (2000) Crystal structure of human sex hormone-binding globulin:
Steroid transport by a laminin G-like domain. EMBO J 19:504–512.8. Kang Y, Zhang X, Dobie F, Wu H, Craig AM (2008) Induction of GABAergic postsynaptic
differentiation by alpha-neurexins. J Biol Chem 283:2323–2334.9. Sheckler LR, Henry L, Sugita S, Sudhof TC, Rudenko G (2006) Crystal structure of the
second LNS/LG domain from neurexin 1alpha: Ca2� binding and the effects of alter-native splicing. J Biol Chem 281:22896–22905.
10. Shen KC, et al. (2008) Regulation of neurexin 1beta tertiary structure and ligandbinding through alternative splicing. Structure 16:422–431.
11. Chen X, Liu H, Shim AH, Focia PJ, He X (2008) Structural basis for synaptic adhesionmediated by neuroligin-neurexin interactions. Nat Struct Mol Biol 15:50–56.
12. Tsigelny I, Shindyalov IN, Bourne PE, Sudhof TC, Taylor P (2000) Common EF-handmotifs in cholinesterases and neuroligins suggest a role for Ca2� binding in cell surfaceassociations. Protein Sci 9:180–185.
13. Dean C, et al. (2003) Neurexin mediates the assembly of presynaptic terminals. NatNeurosci 6:708–716.
14. Comoletti D, et al. (2004) The Arg451Cys-neuroligin-3 mutation associated with autismreveals a defect in protein processing. J Neurosci 24:4889–4893.
15. Graf ER, Kang Y, Hauner AM, Craig AM (2006) Structure function and splice site analysisof the synaptogenic activity of the neurexin-1 beta LNS domain. J Neurosci 26:4256–4265.
16. Comoletti D, et al. (2003) Characterization of the interaction of a recombinant solubleneuroligin-1 with neurexin-1beta. J Biol Chem 278:50497–50505.
17. Chubykin AA, et al. (2005) Dissection of synapse induction by neuroligins: Effect of aneuroligin mutation associated with autism. J Biol Chem 280:22365–22374.
18. Comoletti D, et al. (2006) Gene selection, alternative splicing, and post-translationalprocessing regulate neuroligin selectivity for beta-neurexins. Biochemistry 45:12816–12827.
19. Boucard AA, Chubykin AA, Comoletti D, Taylor P, Sudhof TC (2005) A splice code fortrans-synaptic cell adhesion mediated by binding of neuroligin 1 to alpha- and beta-neurexins. Neuron 48:229–236.
20. Arac D, et al. (2007) Structures of neuroligin-1 and the neuroligin-1/neurexin-1betacomplex reveal specific protein-protein and protein-Ca(2�) interactions. Neuron56:992–1003.
21. Fabrichny IP, et al. (2007) Structural analysis of the synaptic protein neuroligin and itsbeta-neurexin complex: Determinants for folding and cell adhesion. Neuron 56:979–991.
Reissner et al. www.pnas.org/cgi/content/short/0801639105 2 of 19
β7
β4β3
β1
β14
β5-β10 β9
β12
100 290250200150
∆phi
∆psi
136156
251279
23399
134 157 251 283233100
β9 β8 β3
β4
β5β6β7 β11
β2
Ca2+
β10
BA
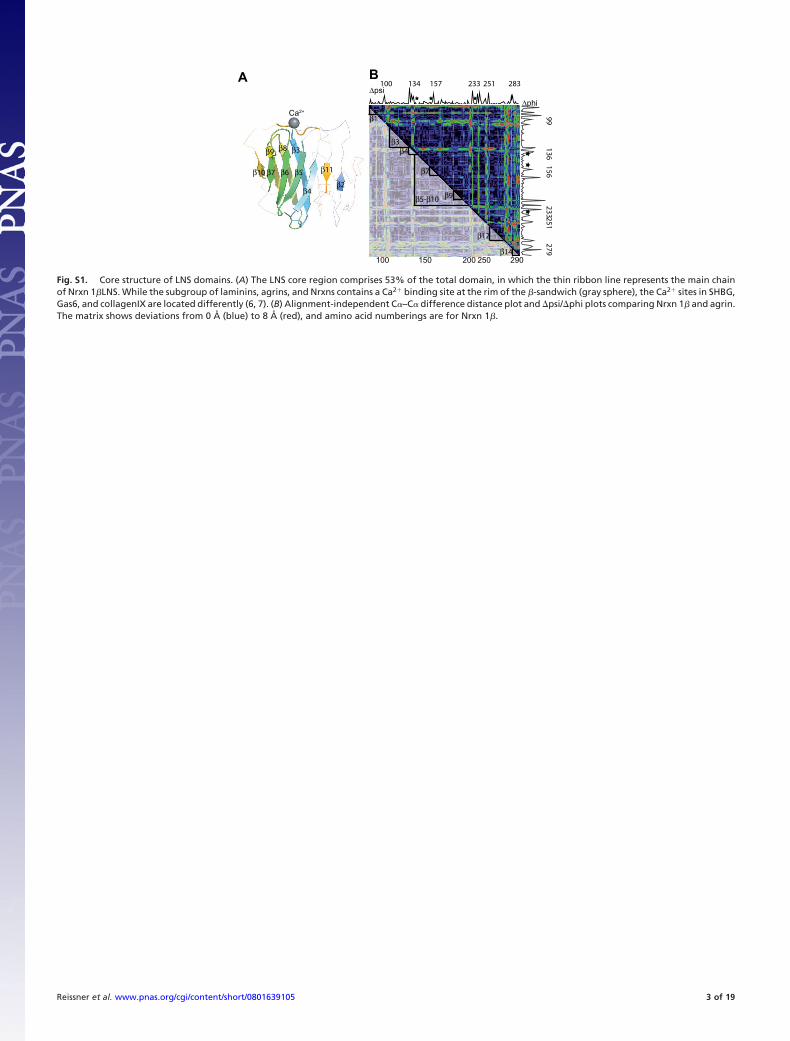
Fig. S1. Core structure of LNS domains. (A) The LNS core region comprises 53% of the total domain, in which the thin ribbon line represents the main chainof Nrxn 1�LNS. While the subgroup of laminins, agrins, and Nrxns contains a Ca2� binding site at the rim of the �-sandwich (gray sphere), the Ca2� sites in SHBG,Gas6, and collagenIX are located differently (6, 7). (B) Alignment-independent C�–C� difference distance plot and �psi/�phi plots comparing Nrxn 1� and agrin.The matrix shows deviations from 0 Å (blue) to 8 Å (red), and amino acid numberings are for Nrxn 1�.
Reissner et al. www.pnas.org/cgi/content/short/0801639105 3 of 19

ON
O
H
O
O
H
N
O
H
OO
OH
H H
Ca2+
L346
G416
L415
M414
D329
O
NO
OO
OO-OH
O
H
N
O
H
H
Ca2+
N2999
D3055
A3054
S3055
D2982
O
NO
OO
NHO-OH
O
H
N
O
H
H
Ca2+
V1200
N1284
F1283
I1282
D1183
Nrxn 1α LNS6 Laminin 2α LNS5 Nrxn 1α LNS2
C ED
Laminin 2α LNS5
B
Ca2+
I1282
N1284
D1183
G1201T1202
L1280
T1281
A
Fig. S2. Ca2� coordination is conserved in LNS domains. (A) Modeled Ca2� binding site of �LNS6 using coordinates identical to crystal structure of �-Nrxn (PDBentry 1C4R). Ca2� is coordinated by the two acidic side chains of �D1183 and �N1284. The corresponding residues in Nrxn 1�LNS are �D137 and �N238 (data notshown). (B) Electrostatic potentials mapped on the surface of laminin 2� LNS5 outlining the ‘‘standard’’ Ca2� binding surface of the laminin/neurexin/agrinsubtype from the LNS domain family. (C and D) Crystal structures of LNS from Nrxn (C), laminin 2� (D), and agrin (data not shown) reveal coordination of theCa2� ion by six oxygen atoms, forming an octahedral complex. Two main-chain carbonyl groups and the carboxylate group of an aspartate are placed at identicalpositions. (E) In Nrxn1� LNS2 the second aspartate in laminin (red in D) or asparagine in Nrxn1� LNS6 (red in C) is replaced by a water molecule (green) and Gly-416(red), but Ca2� binding is maintained (8).
Reissner et al. www.pnas.org/cgi/content/short/0801639105 4 of 19
150
100
75
50
37
βD13
7A
βT23
5A
βN23
8AβL
234S
,
βI123
6S, β
N238D
βG15
5V, β
T156A
βLNS -
SS4
Contro
l
Brain
lysat
e
βLNS +
SS4
anti-Nlgn1
Coomassie
1 2 3 4 5 6 7 8 9
B
150
100
75
50
37
αT1281
A
αD1183
A
αN1284
AαL128
0S,
αI128
2S, α
N1284
D
αG1201
V, α
T1202
A
αLNS6
+ SS4
αLNS6
- SS4
Contro
l
Brain
lysat
e
anti-Nlgn1
Coomassie
1 2 3 4 5 6 7 8 9
A
Mr
Mr
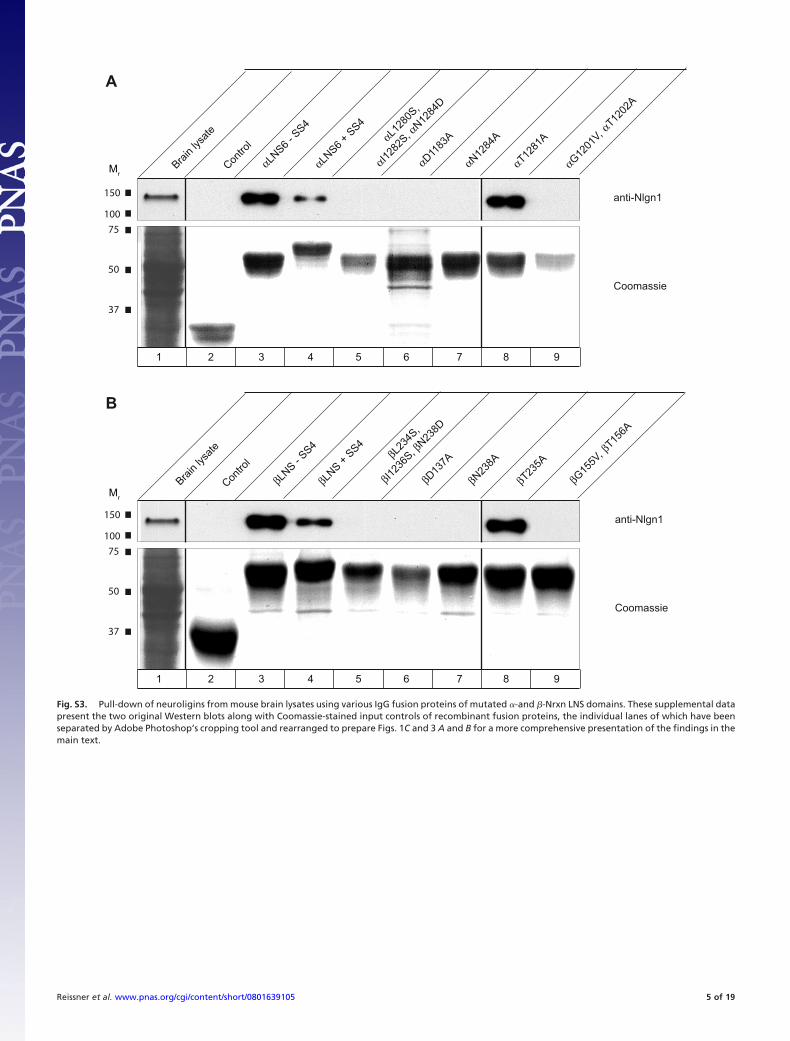
Fig. S3. Pull-down of neuroligins from mouse brain lysates using various IgG fusion proteins of mutated �-and �-Nrxn LNS domains. These supplemental datapresent the two original Western blots along with Coomassie-stained input controls of recombinant fusion proteins, the individual lanes of which have beenseparated by Adobe Photoshop’s cropping tool and rearranged to prepare Figs. 1C and 3 A and B for a more comprehensive presentation of the findings in themain text.
Reissner et al. www.pnas.org/cgi/content/short/0801639105 5 of 19
Ca2+ Ca2+
N
C
N
C
HH
H
H
H
H
H
αLNS6 βLNS
B C
A
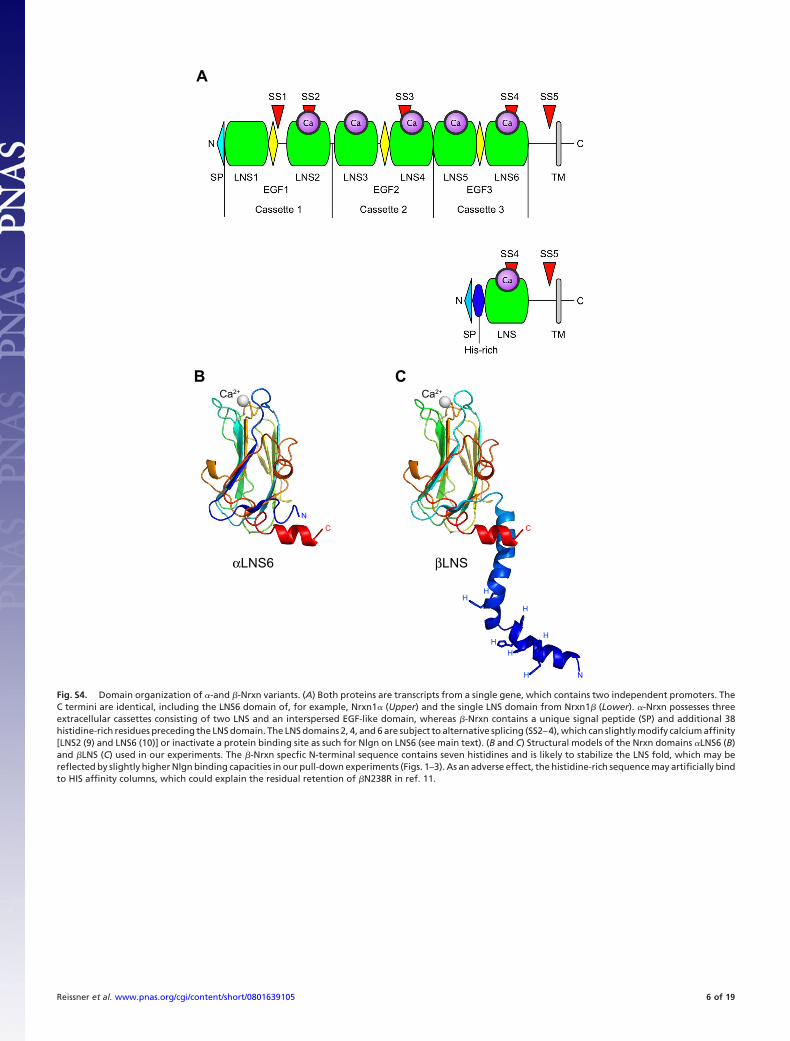
Fig. S4. Domain organization of �-and �-Nrxn variants. (A) Both proteins are transcripts from a single gene, which contains two independent promoters. TheC termini are identical, including the LNS6 domain of, for example, Nrxn1� (Upper) and the single LNS domain from Nrxn1� (Lower). �-Nrxn possesses threeextracellular cassettes consisting of two LNS and an interspersed EGF-like domain, whereas �-Nrxn contains a unique signal peptide (SP) and additional 38histidine-rich residues preceding the LNS domain. The LNS domains 2, 4, and 6 are subject to alternative splicing (SS2–4), which can slightly modify calcium affinity[LNS2 (9) and LNS6 (10)] or inactivate a protein binding site as such for Nlgn on LNS6 (see main text). (B and C) Structural models of the Nrxn domains �LNS6 (B)and �LNS (C) used in our experiments. The �-Nrxn specfic N-terminal sequence contains seven histidines and is likely to stabilize the LNS fold, which may bereflected by slightly higher Nlgn binding capacities in our pull-down experiments (Figs. 1–3). As an adverse effect, the histidine-rich sequence may artificially bindto HIS affinity columns, which could explain the residual retention of �N238R in ref. 11.
Reissner et al. www.pnas.org/cgi/content/short/0801639105 6 of 19
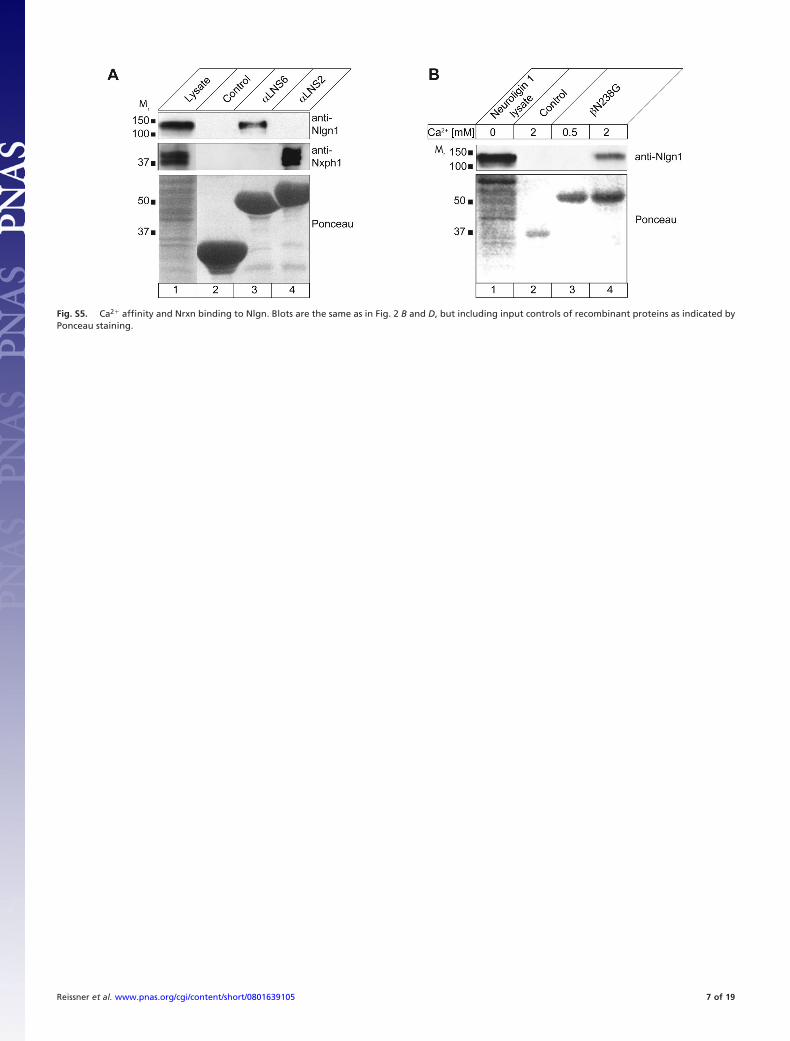
Fig. S5. Ca2� affinity and Nrxn binding to Nlgn. Blots are the same as in Fig. 2 B and D, but including input controls of recombinant proteins as indicated byPonceau staining.
Reissner et al. www.pnas.org/cgi/content/short/0801639105 7 of 19
50
37
150
100
1 2 3 4 5
Ca2+ [mM] 0 2 0.01 0.1 1
Neuro
ligin
1
lysat
e
Contro
l
βLNS +
SS4
anti-Nlgn1
Ponceau
B
Mr
1 2 3 4 5 6 7
Neuro
ligin
1
lysat
e
Contro
l
βLNS -
SS4
anti-Nlgn1
Ca2+ [mM] 20 0.005 0.01 0.1 0.5 2
Ponceau
150
100
50
37
A
Mr
150
100
50
37
1 2 3 4 5 6 7 8
Lysa
te
Contro
l
αLNS6 - SS4
αLNS6 + SS4
Lysa
te
Contro
l
αLNS6 - SS4
αLNS6 + SS4
D
anti-Nlgn1
Coomassie
Nlgn1 - B Nlgn1 + B
Mr
150
100
50
37
1 2 3 4 5 6
Brain
lysat
e
Contro
l
αLNS6
- SS4
βLNS -
SS4
βLNS +
SS4
αLNS +
SS4
C
Mr
anti-Nlgn1
Coomassie
150
10075
50
37
250150
1 2 3 4
E
Neure
xin 1
α
lysat
e
Contro
l
Nlgn 1 +
B
Nlgn 1 − B
Ponceau
anti-Nrxn
Mr
10075
50
37
150
100
Coomassie
Neuro
ligin
1
lysat
e
Contro
l
Contro
l
αLNS6
αLNS5
_E_ L
NS6
αLNS5
_E_ L
NS6
αLNS6
- DTT + DTT
anti-Nlgn1
1 2 3 4 5 6 7
Mr
F
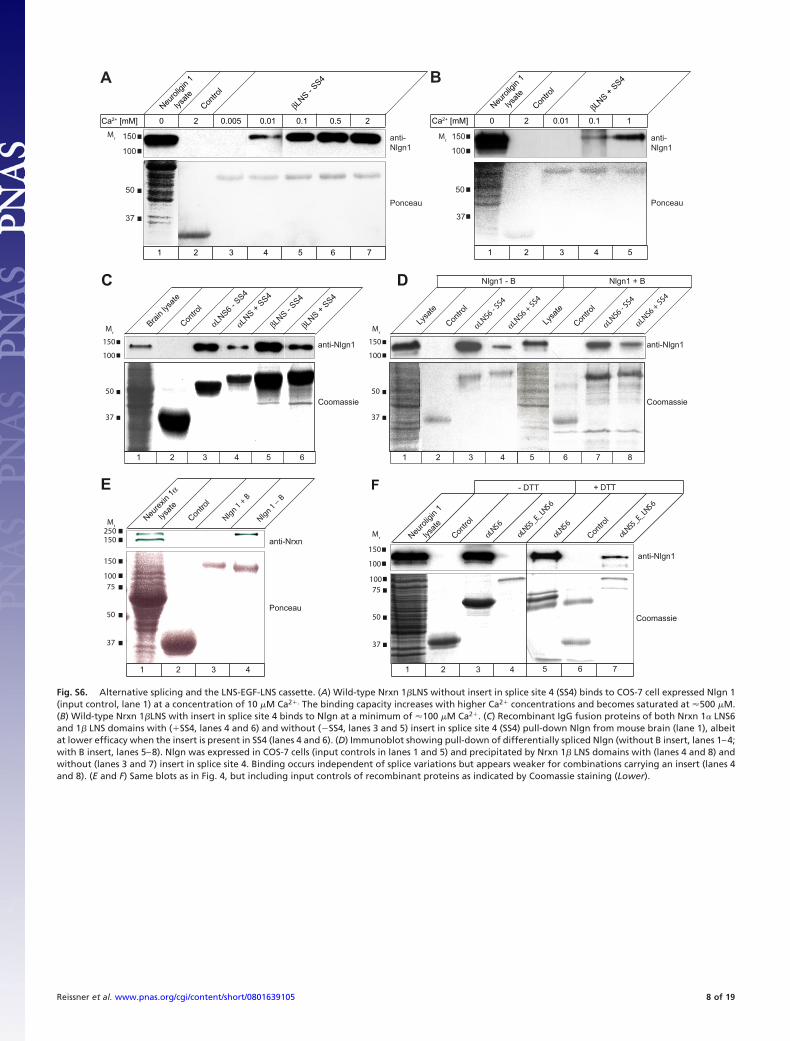
Fig. S6. Alternative splicing and the LNS-EGF-LNS cassette. (A) Wild-type Nrxn 1�LNS without insert in splice site 4 (SS4) binds to COS-7 cell expressed Nlgn 1(input control, lane 1) at a concentration of 10 �M Ca2�. The binding capacity increases with higher Ca2� concentrations and becomes saturated at 500 �M.(B) Wild-type Nrxn 1�LNS with insert in splice site 4 binds to Nlgn at a minimum of 100 �M Ca2�. (C) Recombinant IgG fusion proteins of both Nrxn 1� LNS6and 1� LNS domains with (�SS4, lanes 4 and 6) and without (SS4, lanes 3 and 5) insert in splice site 4 (SS4) pull-down Nlgn from mouse brain (lane 1), albeitat lower efficacy when the insert is present in SS4 (lanes 4 and 6). (D) Immunoblot showing pull-down of differentially spliced Nlgn (without B insert, lanes 1–4;with B insert, lanes 5–8). Nlgn was expressed in COS-7 cells (input controls in lanes 1 and 5) and precipitated by Nrxn 1� LNS domains with (lanes 4 and 8) andwithout (lanes 3 and 7) insert in splice site 4. Binding occurs independent of splice variations but appears weaker for combinations carrying an insert (lanes 4and 8). (E and F) Same blots as in Fig. 4, but including input controls of recombinant proteins as indicated by Coomassie staining (Lower).
Reissner et al. www.pnas.org/cgi/content/short/0801639105 8 of 19
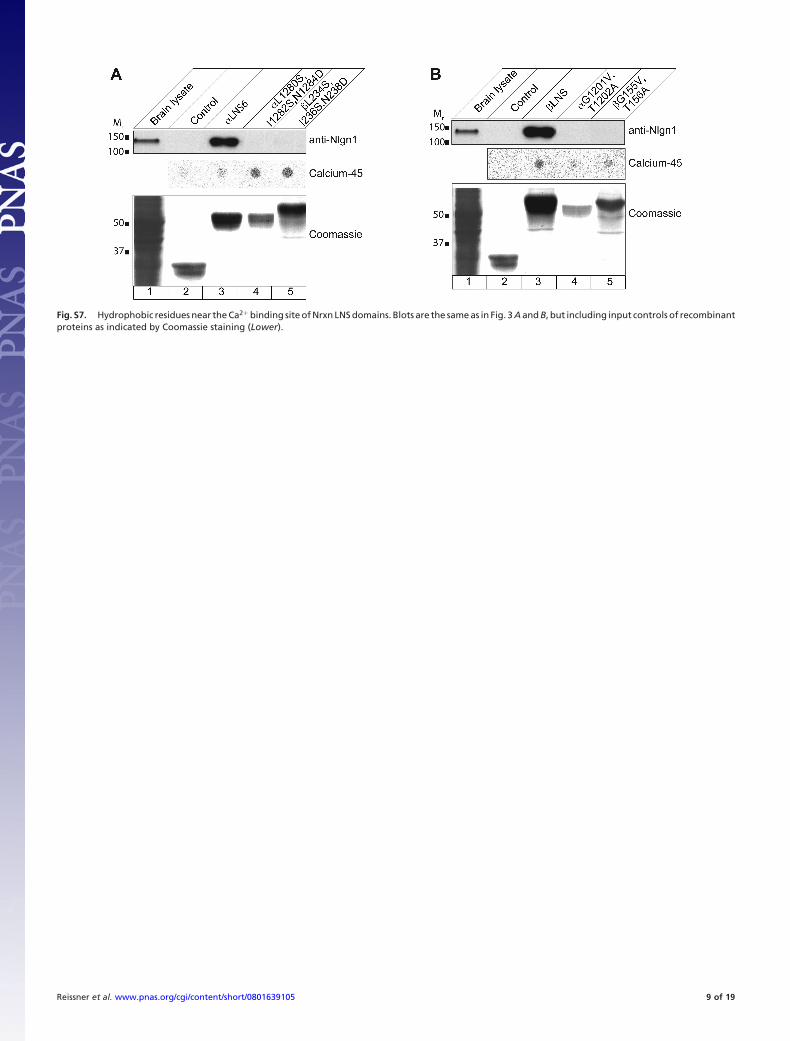
Fig. S7. Hydrophobic residues near the Ca2� binding site of Nrxn LNS domains. Blots are the same as in Fig. 3 A and B, but including input controls of recombinantproteins as indicated by Coomassie staining (Lower).
Reissner et al. www.pnas.org/cgi/content/short/0801639105 9 of 19
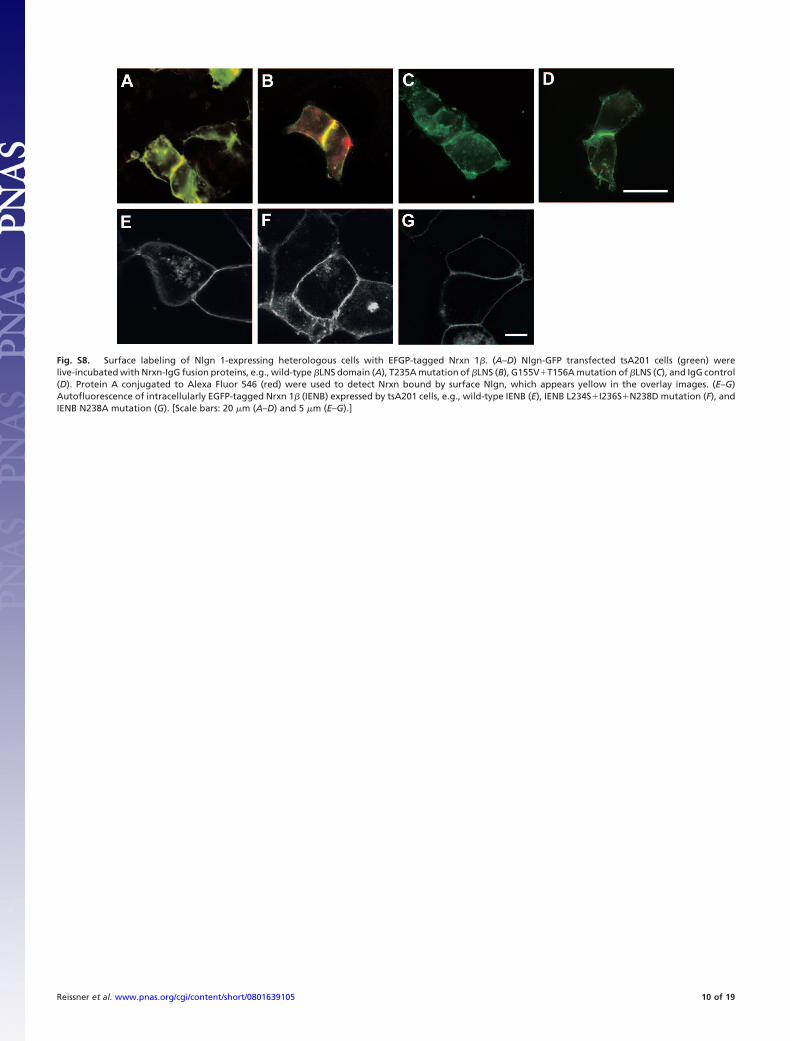
Fig. S8. Surface labeling of Nlgn 1-expressing heterologous cells with EFGP-tagged Nrxn 1�. (A–D) Nlgn-GFP transfected tsA201 cells (green) werelive-incubated with Nrxn-IgG fusion proteins, e.g., wild-type �LNS domain (A), T235A mutation of �LNS (B), G155V�T156A mutation of �LNS (C), and IgG control(D). Protein A conjugated to Alexa Fluor 546 (red) were used to detect Nrxn bound by surface Nlgn, which appears yellow in the overlay images. (E–G)Autofluorescence of intracellularly EGFP-tagged Nrxn 1� (IENB) expressed by tsA201 cells, e.g., wild-type IENB (E), IENB L234S�I236S�N238D mutation (F), andIENB N238A mutation (G). [Scale bars: 20 �m (A–D) and 5 �m (E–G).]
Reissner et al. www.pnas.org/cgi/content/short/0801639105 10 of 19

+SS4
αEGF3
αLNS6 αLNS5
+B
Nlgn
α−Nrxn
Fig. S9. The �EGF3 domain is a negative factor for �-Nrxn binding to Nlgn. Proposed structure of �-Nrxn LNS5 (green), EGF3 (yellow), and LNS6 (cyan) boundto Nlgn (pink) based on crystal data from �-Nrxn LNS domain (PDB entry 1C4R), EGF domain (1EGF), and �-Nrxn/Nlgn complex (3B3Q), respectively. While Nlgnsplice insert B (red) does not interfere with binding to �-Nrxn, the presence of Nrxn splice insert SS4 blocks complex formation. Reducing cysteine bridges withinthe �EGF3 restored Nlgn binding preferences of the �LNS6�E�LNS5 cassette similar to that of the isolated �LNS6 domain (Fig. 4). This could be explained by themodel in which �EGF3 is responsible for the orientation of �LNS5, which reduces the freedom for SS4 within the complex. Binding of Nlgn is not completelyblocked by this domain arrangement but sterically hindered, resulting in only a few possible positions for SS4.
Reissner et al. www.pnas.org/cgi/content/short/0801639105 11 of 19
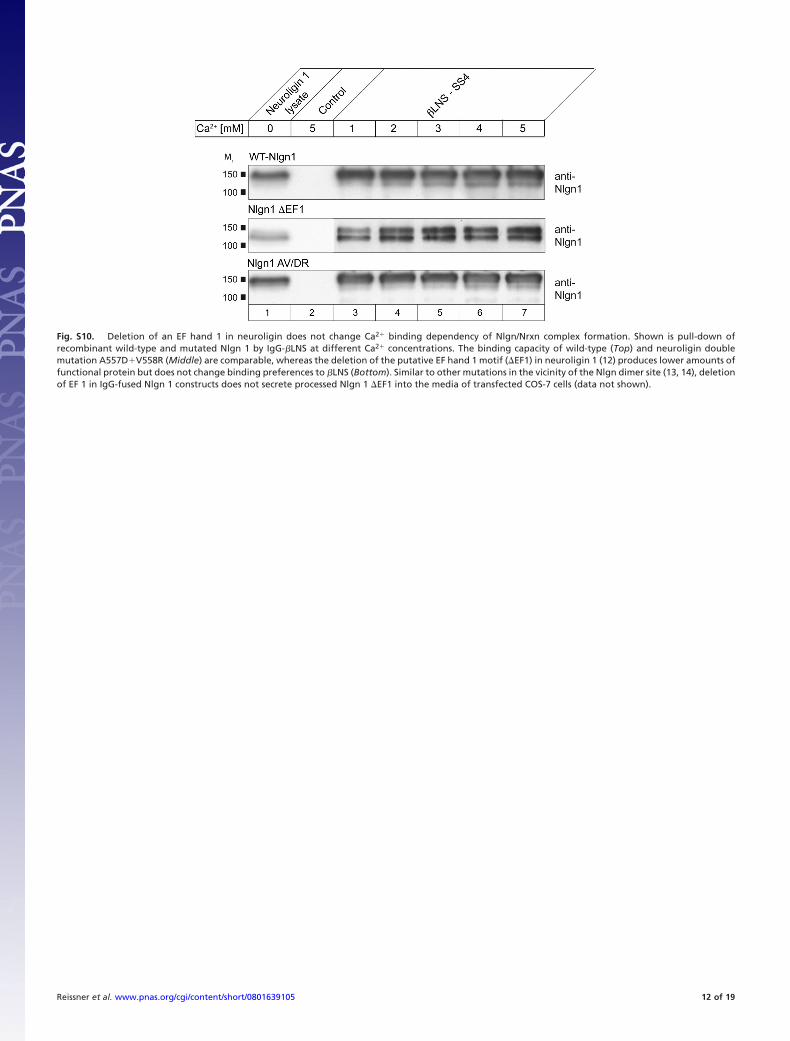
Fig. S10. Deletion of an EF hand 1 in neuroligin does not change Ca2� binding dependency of Nlgn/Nrxn complex formation. Shown is pull-down ofrecombinant wild-type and mutated Nlgn 1 by IgG-�LNS at different Ca2� concentrations. The binding capacity of wild-type (Top) and neuroligin doublemutation A557D�V558R (Middle) are comparable, whereas the deletion of the putative EF hand 1 motif (�EF1) in neuroligin 1 (12) produces lower amounts offunctional protein but does not change binding preferences to �LNS (Bottom). Similar to other mutations in the vicinity of the Nlgn dimer site (13, 14), deletionof EF 1 in IgG-fused Nlgn 1 constructs does not secrete processed Nlgn 1 �EF1 into the media of transfected COS-7 cells (data not shown).
Reissner et al. www.pnas.org/cgi/content/short/0801639105 12 of 19
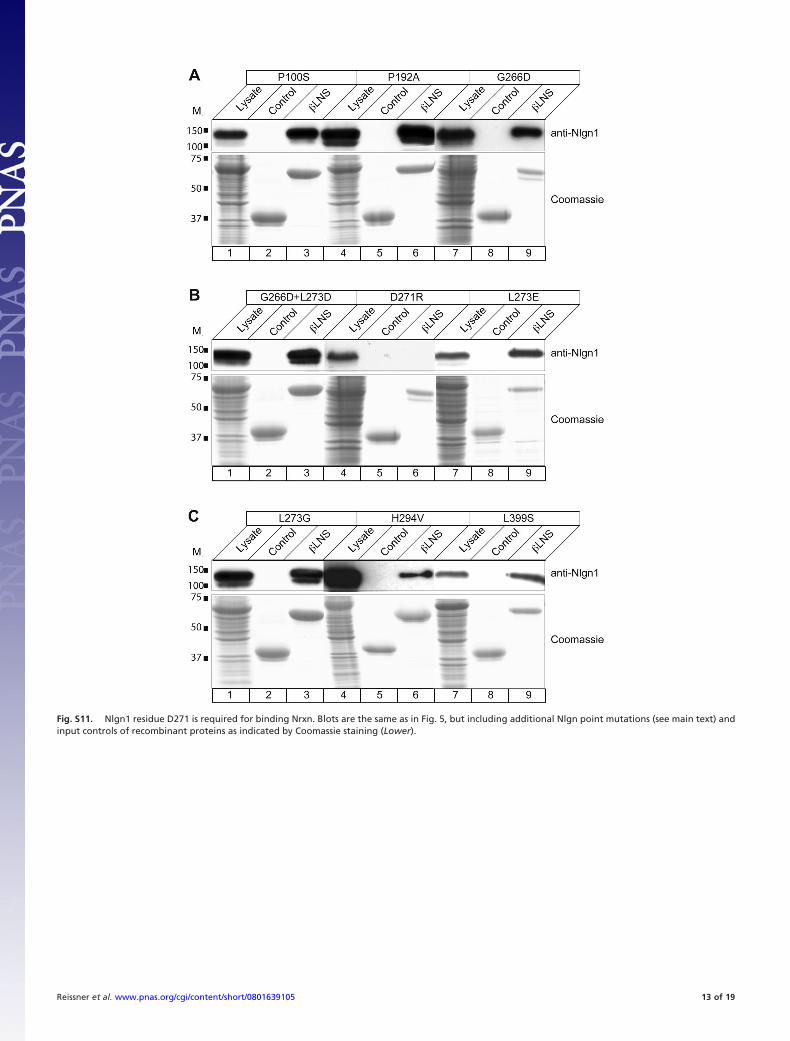
Fig. S11. Nlgn1 residue D271 is required for binding Nrxn. Blots are the same as in Fig. 5, but including additional Nlgn point mutations (see main text) andinput controls of recombinant proteins as indicated by Coomassie staining (Lower).
Reissner et al. www.pnas.org/cgi/content/short/0801639105 13 of 19
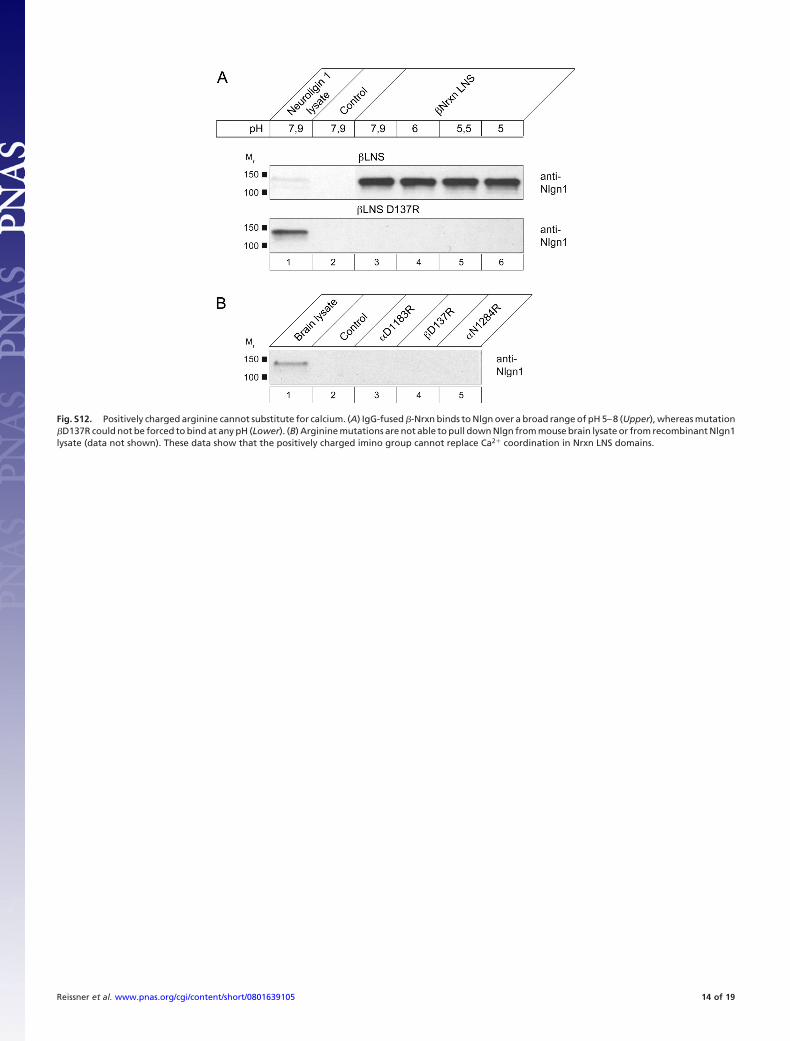
Fig. S12. Positively charged arginine cannot substitute for calcium. (A) IgG-fused �-Nrxn binds to Nlgn over a broad range of pH 5–8 (Upper), whereas mutation�D137R could not be forced to bind at any pH (Lower). (B) Arginine mutations are not able to pull down Nlgn from mouse brain lysate or from recombinant Nlgn1lysate (data not shown). These data show that the positively charged imino group cannot replace Ca2� coordination in Nrxn LNS domains.
Reissner et al. www.pnas.org/cgi/content/short/0801639105 14 of 19
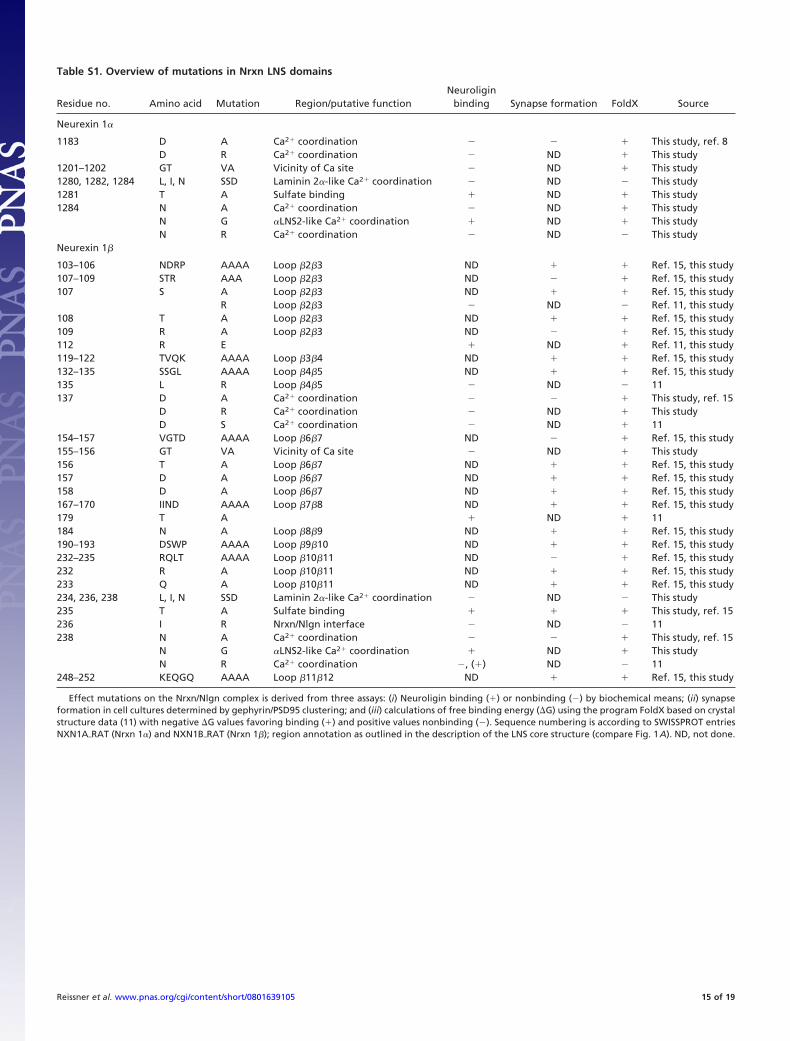
Table S1. Overview of mutations in Nrxn LNS domains
Residue no. Amino acid Mutation Region/putative functionNeuroligin
binding Synapse formation FoldX Source
Neurexin 1�
1183 D A Ca2� coordination � This study, ref. 8D R Ca2� coordination ND � This study
1201–1202 GT VA Vicinity of Ca site ND � This study1280, 1282, 1284 L, I, N SSD Laminin 2�-like Ca2� coordination ND This study1281 T A Sulfate binding � ND � This study1284 N A Ca2� coordination ND � This study
N G �LNS2-like Ca2� coordination � ND � This studyN R Ca2� coordination ND This study
Neurexin 1�
103–106 NDRP AAAA Loop �2�3 ND � � Ref. 15, this study107–109 STR AAA Loop �2�3 ND � Ref. 15, this study107 S A Loop �2�3 ND � � Ref. 15, this study
R Loop �2�3 ND Ref. 11, this study108 T A Loop �2�3 ND � � Ref. 15, this study109 R A Loop �2�3 ND � Ref. 15, this study112 R E � ND � Ref. 11, this study119–122 TVQK AAAA Loop �3�4 ND � � Ref. 15, this study132–135 SSGL AAAA Loop �4�5 ND � � Ref. 15, this study135 L R Loop �4�5 ND 11137 D A Ca2� coordination � This study, ref. 15
D R Ca2� coordination ND � This studyD S Ca2� coordination ND � 11
154–157 VGTD AAAA Loop �6�7 ND � Ref. 15, this study155–156 GT VA Vicinity of Ca site ND � This study156 T A Loop �6�7 ND � � Ref. 15, this study157 D A Loop �6�7 ND � � Ref. 15, this study158 D A Loop �6�7 ND � � Ref. 15, this study167–170 IIND AAAA Loop �7�8 ND � � Ref. 15, this study179 T A � ND � 11184 N A Loop �8�9 ND � � Ref. 15, this study190–193 DSWP AAAA Loop �9�10 ND � � Ref. 15, this study232–235 RQLT AAAA Loop �10�11 ND � Ref. 15, this study232 R A Loop �10�11 ND � � Ref. 15, this study233 Q A Loop �10�11 ND � � Ref. 15, this study234, 236, 238 L, I, N SSD Laminin 2�-like Ca2� coordination ND This study235 T A Sulfate binding � � � This study, ref. 15236 I R Nrxn/Nlgn interface ND 11238 N A Ca2� coordination � This study, ref. 15
N G �LNS2-like Ca2� coordination � ND � This studyN R Ca2� coordination , (�) ND 11
248–252 KEQGQ AAAA Loop �11�12 ND � � Ref. 15, this study
Effect mutations on the Nrxn/Nlgn complex is derived from three assays: (i) Neuroligin binding (�) or nonbinding () by biochemical means; (ii) synapseformation in cell cultures determined by gephyrin/PSD95 clustering; and (iii) calculations of free binding energy (�G) using the program FoldX based on crystalstructure data (11) with negative �G values favoring binding (�) and positive values nonbinding (). Sequence numbering is according to SWISSPROT entriesNXN1A�RAT (Nrxn 1�) and NXN1B�RAT (Nrxn 1�); region annotation as outlined in the description of the LNS core structure (compare Fig. 1A). ND, not done.
Reissner et al. www.pnas.org/cgi/content/short/0801639105 15 of 19
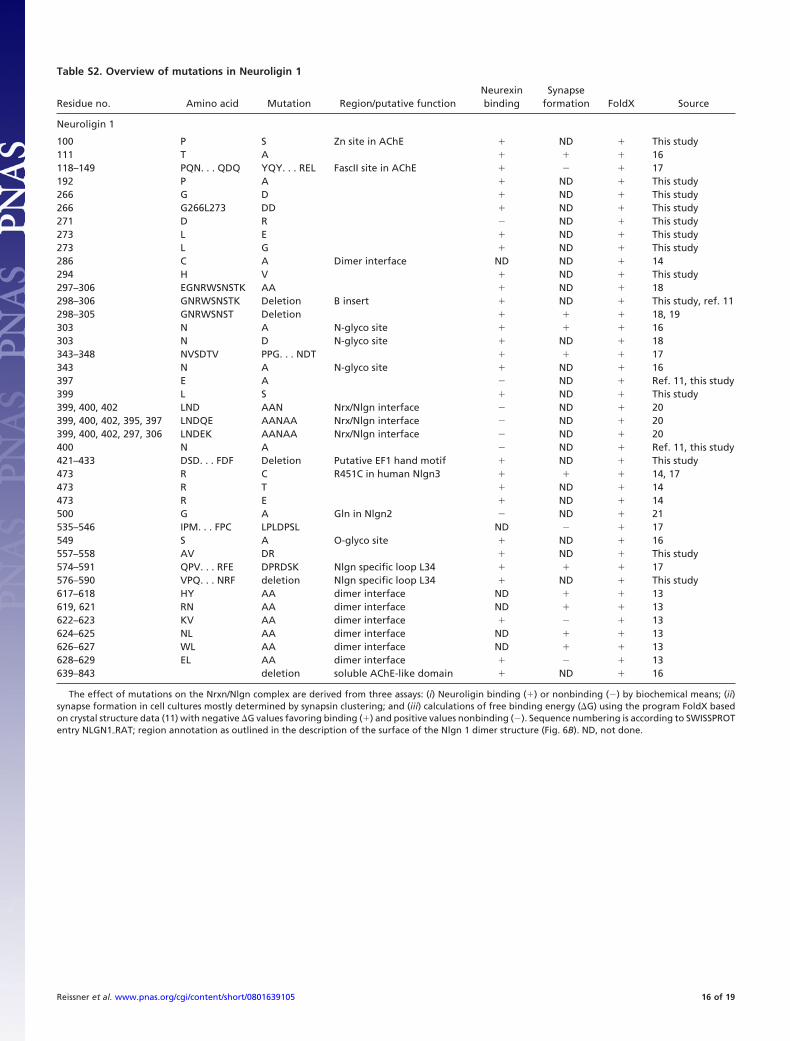
Table S2. Overview of mutations in Neuroligin 1
Residue no. Amino acid Mutation Region/putative functionNeurexinbinding
Synapseformation FoldX Source
Neuroligin 1
100 P S Zn site in AChE � ND � This study111 T A � � � 16118–149 PQN. . . QDQ YQY. . . REL FascII site in AChE � � 17192 P A � ND � This study266 G D � ND � This study266 G266L273 DD � ND � This study271 D R ND � This study273 L E � ND � This study273 L G � ND � This study286 C A Dimer interface ND ND � 14294 H V � ND � This study297–306 EGNRWSNSTK AA � ND � 18298–306 GNRWSNSTK Deletion B insert � ND � This study, ref. 11298–305 GNRWSNST Deletion � � � 18, 19303 N A N-glyco site � � � 16303 N D N-glyco site � ND � 18343–348 NVSDTV PPG. . . NDT � � � 17343 N A N-glyco site � ND � 16397 E A ND � Ref. 11, this study399 L S � ND � This study399, 400, 402 LND AAN Nrx/Nlgn interface ND � 20399, 400, 402, 395, 397 LNDQE AANAA Nrx/Nlgn interface ND � 20399, 400, 402, 297, 306 LNDEK AANAA Nrx/Nlgn interface ND � 20400 N A ND � Ref. 11, this study421–433 DSD. . . FDF Deletion Putative EF1 hand motif � ND � This study473 R C R451C in human Nlgn3 � � � 14, 17473 R T � ND � 14473 R E � ND � 14500 G A Gln in Nlgn2 ND � 21535–546 IPM. . . FPC LPLDPSL ND � 17549 S A O-glyco site � ND � 16557–558 AV DR � ND � This study574–591 QPV. . . RFE DPRDSK Nlgn specific loop L34 � � � 17576–590 VPQ. . . NRF deletion Nlgn specific loop L34 � ND � This study617–618 HY AA dimer interface ND � � 13619, 621 RN AA dimer interface ND � � 13622–623 KV AA dimer interface � � 13624–625 NL AA dimer interface ND � � 13626–627 WL AA dimer interface ND � � 13628–629 EL AA dimer interface � � 13639–843 deletion soluble AChE-like domain � ND � 16
The effect of mutations on the Nrxn/Nlgn complex are derived from three assays: (i) Neuroligin binding (�) or nonbinding () by biochemical means; (ii)synapse formation in cell cultures mostly determined by synapsin clustering; and (iii) calculations of free binding energy (�G) using the program FoldX basedon crystal structure data (11) with negative �G values favoring binding (�) and positive values nonbinding (). Sequence numbering is according to SWISSPROTentry NLGN1�RAT; region annotation as outlined in the description of the surface of the Nlgn 1 dimer structure (Fig. 6B). ND, not done.
Reissner et al. www.pnas.org/cgi/content/short/0801639105 16 of 19
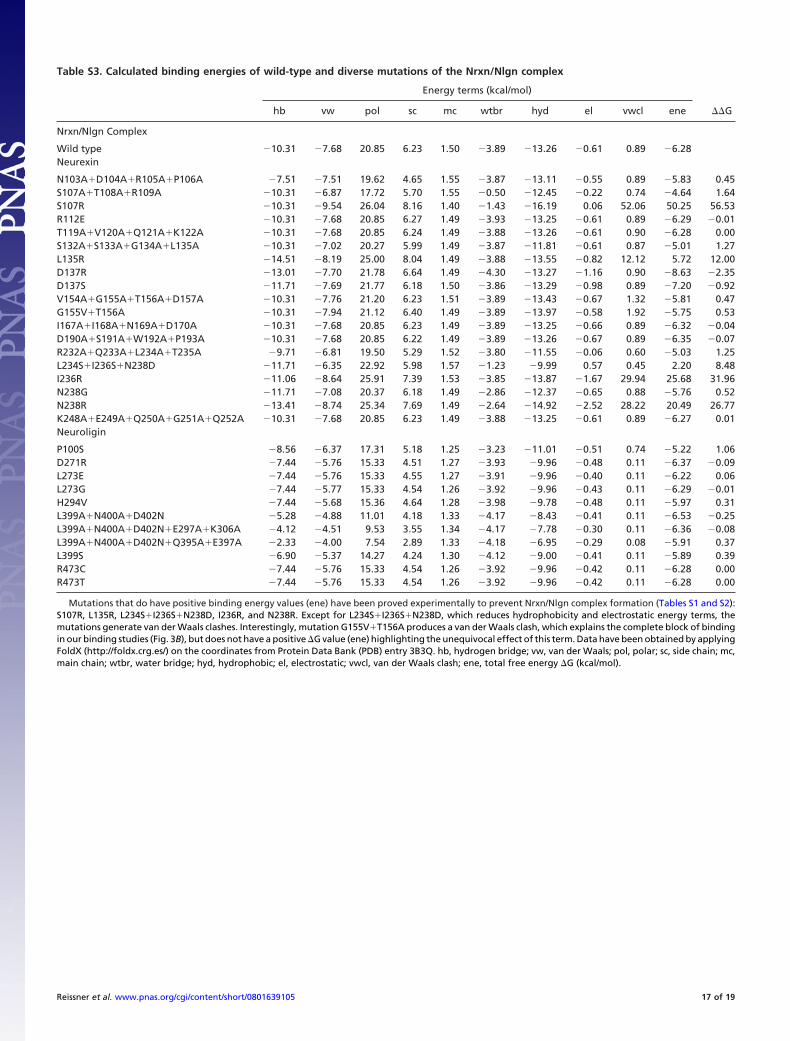
Table S3. Calculated binding energies of wild-type and diverse mutations of the Nrxn/Nlgn complex
Energy terms (kcal/mol)
hb vw pol sc mc wtbr hyd el vwcl ene ��G
Nrxn/Nlgn Complex
Wild type 10.31 7.68 20.85 6.23 1.50 3.89 13.26 0.61 0.89 6.28Neurexin
N103A�D104A�R105A�P106A 7.51 7.51 19.62 4.65 1.55 3.87 13.11 0.55 0.89 5.83 0.45S107A�T108A�R109A 10.31 6.87 17.72 5.70 1.55 0.50 12.45 0.22 0.74 4.64 1.64S107R 10.31 9.54 26.04 8.16 1.40 1.43 16.19 0.06 52.06 50.25 56.53R112E 10.31 7.68 20.85 6.27 1.49 3.93 13.25 0.61 0.89 6.29 0.01T119A�V120A�Q121A�K122A 10.31 7.68 20.85 6.24 1.49 3.88 13.26 0.61 0.90 6.28 0.00S132A�S133A�G134A�L135A 10.31 7.02 20.27 5.99 1.49 3.87 11.81 0.61 0.87 5.01 1.27L135R 14.51 8.19 25.00 8.04 1.49 3.88 13.55 0.82 12.12 5.72 12.00D137R 13.01 7.70 21.78 6.64 1.49 4.30 13.27 1.16 0.90 8.63 2.35D137S 11.71 7.69 21.77 6.18 1.50 3.86 13.29 0.98 0.89 7.20 0.92V154A�G155A�T156A�D157A 10.31 7.76 21.20 6.23 1.51 3.89 13.43 0.67 1.32 5.81 0.47G155V�T156A 10.31 7.94 21.12 6.40 1.49 3.89 13.97 0.58 1.92 5.75 0.53I167A�I168A�N169A�D170A 10.31 7.68 20.85 6.23 1.49 3.89 13.25 0.66 0.89 6.32 0.04D190A�S191A�W192A�P193A 10.31 7.68 20.85 6.22 1.49 3.89 13.26 0.67 0.89 6.35 0.07R232A�Q233A�L234A�T235A 9.71 6.81 19.50 5.29 1.52 3.80 11.55 0.06 0.60 5.03 1.25L234S�I236S�N238D 11.71 6.35 22.92 5.98 1.57 1.23 9.99 0.57 0.45 2.20 8.48I236R 11.06 8.64 25.91 7.39 1.53 3.85 13.87 1.67 29.94 25.68 31.96N238G 11.71 7.08 20.37 6.18 1.49 2.86 12.37 0.65 0.88 5.76 0.52N238R 13.41 8.74 25.34 7.69 1.49 2.64 14.92 2.52 28.22 20.49 26.77K248A�E249A�Q250A�G251A�Q252A 10.31 7.68 20.85 6.23 1.49 3.88 13.25 0.61 0.89 6.27 0.01Neuroligin
P100S 8.56 6.37 17.31 5.18 1.25 3.23 11.01 0.51 0.74 5.22 1.06D271R 7.44 5.76 15.33 4.51 1.27 3.93 9.96 0.48 0.11 6.37 0.09L273E 7.44 5.76 15.33 4.55 1.27 3.91 9.96 0.40 0.11 6.22 0.06L273G 7.44 5.77 15.33 4.54 1.26 3.92 9.96 0.43 0.11 6.29 0.01H294V 7.44 5.68 15.36 4.64 1.28 3.98 9.78 0.48 0.11 5.97 0.31L399A�N400A�D402N 5.28 4.88 11.01 4.18 1.33 4.17 8.43 0.41 0.11 6.53 0.25L399A�N400A�D402N�E297A�K306A 4.12 4.51 9.53 3.55 1.34 4.17 7.78 0.30 0.11 6.36 0.08L399A�N400A�D402N�Q395A�E397A 2.33 4.00 7.54 2.89 1.33 4.18 6.95 0.29 0.08 5.91 0.37L399S 6.90 5.37 14.27 4.24 1.30 4.12 9.00 0.41 0.11 5.89 0.39R473C 7.44 5.76 15.33 4.54 1.26 3.92 9.96 0.42 0.11 6.28 0.00R473T 7.44 5.76 15.33 4.54 1.26 3.92 9.96 0.42 0.11 6.28 0.00
Mutations that do have positive binding energy values (ene) have been proved experimentally to prevent Nrxn/Nlgn complex formation (Tables S1 and S2):S107R, L135R, L234S�I236S�N238D, I236R, and N238R. Except for L234S�I236S�N238D, which reduces hydrophobicity and electrostatic energy terms, themutations generate van der Waals clashes. Interestingly, mutation G155V�T156A produces a van der Waals clash, which explains the complete block of bindingin our binding studies (Fig. 3B), but does not have a positive �G value (ene) highlighting the unequivocal effect of this term. Data have been obtained by applyingFoldX (http://foldx.crg.es/) on the coordinates from Protein Data Bank (PDB) entry 3B3Q. hb, hydrogen bridge; vw, van der Waals; pol, polar; sc, side chain; mc,main chain; wtbr, water bridge; hyd, hydrophobic; el, electrostatic; vwcl, van der Waals clash; ene, total free energy �G (kcal/mol).
Reissner et al. www.pnas.org/cgi/content/short/0801639105 17 of 19
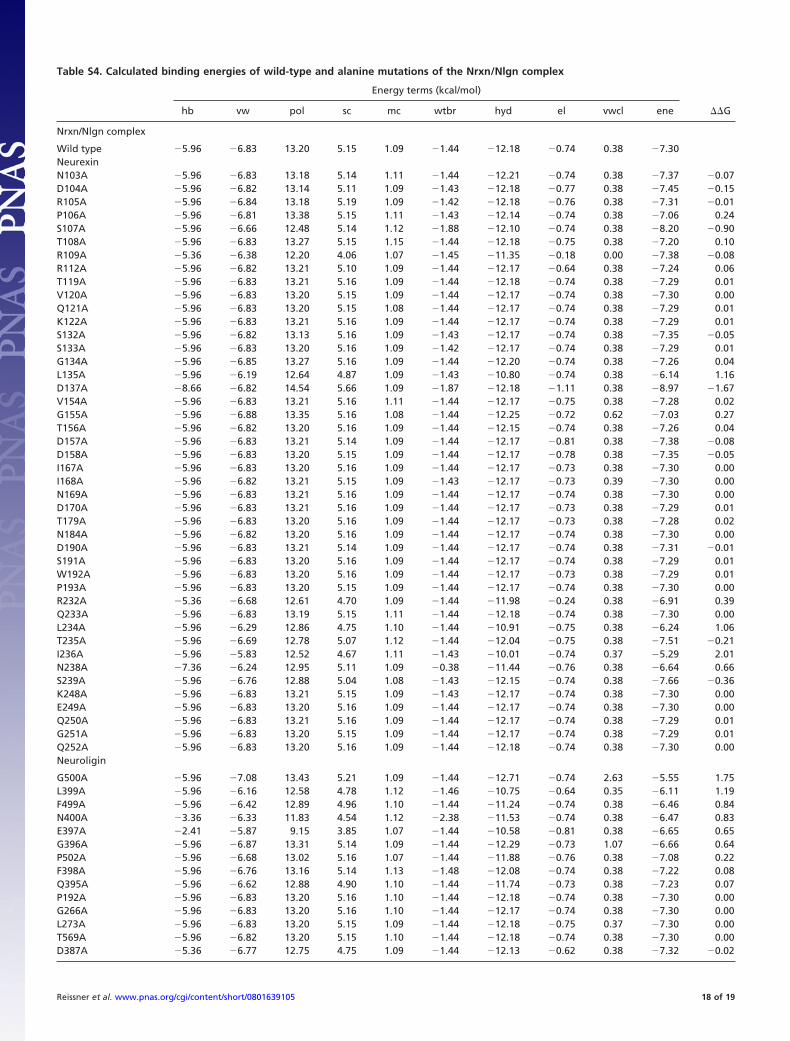
Table S4. Calculated binding energies of wild-type and alanine mutations of the Nrxn/Nlgn complex
Energy terms (kcal/mol)
hb vw pol sc mc wtbr hyd el vwcl ene ��G
Nrxn/Nlgn complex
Wild type 5.96 6.83 13.20 5.15 1.09 1.44 12.18 0.74 0.38 7.30NeurexinN103A 5.96 6.83 13.18 5.14 1.11 1.44 12.21 0.74 0.38 7.37 0.07D104A 5.96 6.82 13.14 5.11 1.09 1.43 12.18 0.77 0.38 7.45 0.15R105A 5.96 6.84 13.18 5.19 1.09 1.42 12.18 0.76 0.38 7.31 0.01P106A 5.96 6.81 13.38 5.15 1.11 1.43 12.14 0.74 0.38 7.06 0.24S107A 5.96 6.66 12.48 5.14 1.12 1.88 12.10 0.74 0.38 8.20 0.90T108A 5.96 6.83 13.27 5.15 1.15 1.44 12.18 0.75 0.38 7.20 0.10R109A 5.36 6.38 12.20 4.06 1.07 1.45 11.35 0.18 0.00 7.38 0.08R112A 5.96 6.82 13.21 5.10 1.09 1.44 12.17 0.64 0.38 7.24 0.06T119A 5.96 6.83 13.21 5.16 1.09 1.44 12.18 0.74 0.38 7.29 0.01V120A 5.96 6.83 13.20 5.15 1.09 1.44 12.17 0.74 0.38 7.30 0.00Q121A 5.96 6.83 13.20 5.15 1.08 1.44 12.17 0.74 0.38 7.29 0.01K122A 5.96 6.83 13.21 5.16 1.09 1.44 12.17 0.74 0.38 7.29 0.01S132A 5.96 6.82 13.13 5.16 1.09 1.43 12.17 0.74 0.38 7.35 0.05S133A 5.96 6.83 13.20 5.16 1.09 1.42 12.17 0.74 0.38 7.29 0.01G134A 5.96 6.85 13.27 5.16 1.09 1.44 12.20 0.74 0.38 7.26 0.04L135A 5.96 6.19 12.64 4.87 1.09 1.43 10.80 0.74 0.38 6.14 1.16D137A 8.66 6.82 14.54 5.66 1.09 1.87 12.18 1.11 0.38 8.97 1.67V154A 5.96 6.83 13.21 5.16 1.11 1.44 12.17 0.75 0.38 7.28 0.02G155A 5.96 6.88 13.35 5.16 1.08 1.44 12.25 0.72 0.62 7.03 0.27T156A 5.96 6.82 13.20 5.16 1.09 1.44 12.15 0.74 0.38 7.26 0.04D157A 5.96 6.83 13.21 5.14 1.09 1.44 12.17 0.81 0.38 7.38 0.08D158A 5.96 6.83 13.20 5.15 1.09 1.44 12.17 0.78 0.38 7.35 0.05I167A 5.96 6.83 13.20 5.16 1.09 1.44 12.17 0.73 0.38 7.30 0.00I168A 5.96 6.82 13.21 5.15 1.09 1.43 12.17 0.73 0.39 7.30 0.00N169A 5.96 6.83 13.21 5.16 1.09 1.44 12.17 0.74 0.38 7.30 0.00D170A 5.96 6.83 13.21 5.16 1.09 1.44 12.17 0.73 0.38 7.29 0.01T179A 5.96 6.83 13.20 5.16 1.09 1.44 12.17 0.73 0.38 7.28 0.02N184A 5.96 6.82 13.20 5.16 1.09 1.44 12.17 0.74 0.38 7.30 0.00D190A 5.96 6.83 13.21 5.14 1.09 1.44 12.17 0.74 0.38 7.31 0.01S191A 5.96 6.83 13.20 5.16 1.09 1.44 12.17 0.74 0.38 7.29 0.01W192A 5.96 6.83 13.20 5.16 1.09 1.44 12.17 0.73 0.38 7.29 0.01P193A 5.96 6.83 13.20 5.15 1.09 1.44 12.17 0.74 0.38 7.30 0.00R232A 5.36 6.68 12.61 4.70 1.09 1.44 11.98 0.24 0.38 6.91 0.39Q233A 5.96 6.83 13.19 5.15 1.11 1.44 12.18 0.74 0.38 7.30 0.00L234A 5.96 6.29 12.86 4.75 1.10 1.44 10.91 0.75 0.38 6.24 1.06T235A 5.96 6.69 12.78 5.07 1.12 1.44 12.04 0.75 0.38 7.51 0.21I236A 5.96 5.83 12.52 4.67 1.11 1.43 10.01 0.74 0.37 5.29 2.01N238A 7.36 6.24 12.95 5.11 1.09 0.38 11.44 0.76 0.38 6.64 0.66S239A 5.96 6.76 12.88 5.04 1.08 1.43 12.15 0.74 0.38 7.66 0.36K248A 5.96 6.83 13.21 5.15 1.09 1.43 12.17 0.74 0.38 7.30 0.00E249A 5.96 6.83 13.20 5.16 1.09 1.44 12.17 0.74 0.38 7.30 0.00Q250A 5.96 6.83 13.21 5.16 1.09 1.44 12.17 0.74 0.38 7.29 0.01G251A 5.96 6.83 13.20 5.15 1.09 1.44 12.17 0.74 0.38 7.29 0.01Q252A 5.96 6.83 13.20 5.16 1.09 1.44 12.18 0.74 0.38 7.30 0.00Neuroligin
G500A 5.96 7.08 13.43 5.21 1.09 1.44 12.71 0.74 2.63 5.55 1.75L399A 5.96 6.16 12.58 4.78 1.12 1.46 10.75 0.64 0.35 6.11 1.19F499A 5.96 6.42 12.89 4.96 1.10 1.44 11.24 0.74 0.38 6.46 0.84N400A 3.36 6.33 11.83 4.54 1.12 2.38 11.53 0.74 0.38 6.47 0.83E397A 2.41 5.87 9.15 3.85 1.07 1.44 10.58 0.81 0.38 6.65 0.65G396A 5.96 6.87 13.31 5.14 1.09 1.44 12.29 0.73 1.07 6.66 0.64P502A 5.96 6.68 13.02 5.16 1.07 1.44 11.88 0.76 0.38 7.08 0.22F398A 5.96 6.76 13.16 5.14 1.13 1.48 12.08 0.74 0.38 7.22 0.08Q395A 5.96 6.62 12.88 4.90 1.10 1.44 11.74 0.73 0.38 7.23 0.07P192A 5.96 6.83 13.20 5.16 1.10 1.44 12.18 0.74 0.38 7.30 0.00G266A 5.96 6.83 13.20 5.16 1.10 1.44 12.17 0.74 0.38 7.30 0.00L273A 5.96 6.83 13.20 5.15 1.09 1.44 12.18 0.75 0.37 7.30 0.00T569A 5.96 6.82 13.20 5.15 1.10 1.44 12.18 0.74 0.38 7.30 0.00D387A 5.36 6.77 12.75 4.75 1.09 1.44 12.13 0.62 0.38 7.32 0.02
Reissner et al. www.pnas.org/cgi/content/short/0801639105 18 of 19

Energy terms (kcal/mol)
hb vw pol sc mc wtbr hyd el vwcl ene ��G
D271A 5.96 6.83 13.21 5.16 1.10 1.44 12.18 0.82 0.38 7.37 0.07N498A 5.96 6.76 13.01 5.10 1.08 1.44 12.07 0.74 0.38 7.38 0.08E297A 5.36 6.58 12.29 4.18 1.10 1.44 11.75 0.27 0.34 7.46 0.16H294A 5.96 6.31 12.15 4.81 1.10 1.44 11.28 0.57 0.00 7.48 0.18D402A 5.96 6.78 12.85 5.14 1.09 1.44 12.14 0.79 0.38 7.65 0.35
These calculations have been done to estimate the energy contribution of each side chain at the binding interface without generating van der Waals clashes(see Table S3). Residues reducing ��G by �1 kcal/mol were classified as hot-spot residues of the Nrxn/Nlgn complex (Fig. 6B). Beside G500, which is stringentrequired to prevent a steric clash, all other hot-spot residues are hydrophobic. Data have been obtained by applying FoldX (http://foldx.crg.es/) on the coordinatesfrom PDB entry 3B3Q. hb, hydrogen bridge; vw, van der Waals; pol, polar; sc, side chain; mc, main chain; wtbr, water bridge; hyd, hydrophobic; el, electrostatic;vwcl, van der Waals clash; ene, total free energy �G (kcal/mol).
Reissner et al. www.pnas.org/cgi/content/short/0801639105 19 of 19





























