-
www.sciencemag.org/cgi/content/full/1120225/DC1
Supporting Online Material for
A Role for the Phagosome in Cytokine Secretion Rachael Z.
Murray, Jason G. Kay, Daniele G. Sangermani, Jennifer L. Stow*
*To whom correspondence should be addressed. E-mail:
[email protected]
Published 10 November 2005 on Science Express DOI:
10.1126/science.1120225
This PDF file includes:
Materials and Methods SOM Text Figs. S1 to S3 Table S1
References
Other Supporting Online Material for this manuscript includes
the following: (available at
www.sciencemag.org/cgi/content/full/1120225/DC1)
Movies S1 to S7
-
1
Supporting Online Material Materials and Methods Antibodies and
reagents. Rabbit polyclonal and rat monoclonal anti-mouse TNFα
antibodies were purchased from Calbiochem and Auspep respectively.
Mouse monoclonal PE-conjugated anti-TNFα antibody was purchased
from BD Biosciences. Rabbit polyclonal antibodies specific for
VAMP3 were purchased from AbCam, while rabbit polyclonal antibodies
specific for Rab11 and mouse monoclonal antibodies specific for
transferrin receptor were purchased from Zymed. Rat monoclonal
antibodies specific for LAMP1 were purchased from Southern Biotech,
while rabbit polyclonal antibodies specific for VAMP8 were
purchased from Synaptic Systems and rabbit polyclonal anti-TACE
antibodies were purchased from Chemicon. Mouse monoclonal
antibodies specific for syntaxin 6 and Vti1b were purchased from
Transduction Laboratories. Rabbit polyclonal antibodies specific
for syntaxin 4 were a kind gift from David James (Garvan Institute
of Medical Research, Sydney, Australia). Mouse monoclonal
antibodies specific for tubulin, Alexa-488 conjugated phalloidin
(used to label F-actin), Texas Red and Alexa 647-conjugated
transferrin (Tfn) and DilC18(5)-DS lipophilic dye were purchased
from Molecular Probes. HRP-conjugated Tfn was purchased from
Rockland. 3,3’-diaminobenzidine (DAB) was purchased from
Sigma-Aldrich. TACE inhibitor, TAPI-1, was purchased from
Calbiochem. LPS from Salmonella minosota Re595 was purchased from
Sigma. IFNγ was purchased from R&D Systems. GFP-tagged
constitutively active Rab11a (Rab11Q70L-GFP) and dominant
negative-Rab11a (Rab11S25N-GFP) were provided by Rob Parton
(University of Queensland, Australia) with permission from Marino
Zerial (Max Planck Institute for Molecular Cell Biology and
Genetics, Germany). GFP-tK was a kind gift from John Hancock
(University of Queensland, Australia) and ApoE-GFP was a kind gift
from Len Kritharides and Wendy Jessup (Centre for Vascular
Research, NSW, Australia). Candida albicans was kindly provided by
Robert Ashman (University of Queensland, Australia). Cell culture,
phagocytosis, molecular cloning and electroporation. RAW264.7 mouse
macrophages were cultured, electroporated and activated with LPS
(1-2 hours) as previously described (1). In some experiments
macrophages were primed for 18 hours in the presence of 500 pg/ml
IFNγ prior to their incubation with LPS (100 ng/ml) for 2 h or
incubated with live Candida albicans at a ratio of 10:1
(yeast:macrophage) for 15-40 min. Where stated 10 µM TACE inhibitor
was added to the cells at the same time as the LPS or Candida
albicans. VAMP3 (full-length GFP-VAMP3 and truncated GFP-VAMP3, aa
1-81) was subcloned from a NIA clone (NIA ID H3025H09) into the
pEGF-C2 vector (Clontech, BD Biosciences) to produce an N-terminal
GFP-tagged protein. TNFα was subcloned into pEGF-C2 vector
(Clontech, BD Biosciences) from an existing TNFα clone (2) to
produce an N-terminal GFP-tagged protein. Macrophages were
electroporated, using the Gene Pulser II (Bio-Rad), for transient
expression of cDNAs using 2.5 × 107 cells with 10 µg DNA, with a
high capacitance setting (280 mV and 950 µF). Cells were washed and
typically cultured for 4-24 hrs.
-
2
Small interfering RNA (siRNA) treatment. RAW264.7 cells, plated
on glass coverslips, were transfected with three different siRNA
constructs designed to silence mouse VAMP3, (Silencer Validated
siRNA ID#186988-90, Ambion) using Lipofectamine 2000 according to
the manufacturer’s instructions (Invitrogen) and cultured for 24
hours, retransfected under then same conditions then cultured for a
further 24 hours prior to LPS treatment in the presence of TACE
inhibitor. Microarray data analysis. RNA was harvested from RAW
264.7 macrophages, cultured in the presence of LPS (100 ng/ml) for
0, 2 or 12 hour. The RNA was probed using the Affymetrix 430A mouse
gene chip as previously described (1); results published at GEO,
accession number GSE1459,
http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE1459).
SDS-PAGE, immunoblotting, immunoprecipitation and
immunofluorescence staining. Raw264.7 cells were washed three times
with PBS, scraped and lysed in Buffer A (10 mM Tris, pH 7.4,
containing 1 mM EDTA, 0.5% Triton X-100, and Complete protease
inhibitors (Roche Applied Science)), by passage through a series of
successively smaller needles and centrifuged for 10 min at 17,000 X
g. The supernatant was assayed for protein content (BioRad protein
assay), subject to SDS-PAGE separation and analyzed by
immunoblotting (1). For immunoprecipitation RAW264.7 cell lysates
were incubated with antibody (5 µg) bound to protein A-Trisacryl
(Pierce) for 2 h at 4 C. Beads were then washed with excess Buffer
A containing 150 mM NaCl and bound proteins were solubilized in
SDS-PAGE sample buffer. Immunofluorescence staining was performed
as described previously (1). Assays for TNFα trafficking, surface
delivery and secretion. The trafficking of TNFα from the Golgi
complex to the cell surface was measured using an
immunofluorescence-based assay as previously described (3).
Briefly, macrophages were incubated in the presence of LPS (100
ng/ml) and TACE inhibitor (10 µM) for 2 hours, fixed with 4%
paraformaldehyde and immunostained to label surface TNFα, then
permeabilized with 0.1 % Trition X-100 and immunostained for
internal TNFα. A commercial enzyme-linked immunosorbant assay
(ELISA) kit (BD OptEIA, BD Biosciences) was used according to the
manufacturer’s instructions to determine levels of secreted TNFα.
HRP inactivation assay. An HRP inactivation assay was modified from
the protocol of Ang et al (4). RAW264.7 cells were incubated with
Tfn-HRP (10 µg/ml) and Tfn-Alexa-647 (10 µg/ml) in media for 1 hr
in the dark at 37°C in the presence of 100 ng/ml LPS. Cells were
washed twice in ice-cold PBS and surface-bound Tfn-HRP was removed
by two 5 min washes with 0.15 M NaCl and 20 mM citric acid, pH 5.
Cells were then washed twice with ice-cold PBS and incubated in the
dark for 1 hr with PBS containing 0.1 mg/ml DAB and 0.025 % H2O2 to
the inactivation sample (the control contained PBS alone). Cells
were washed twice in PBS containing 1 % BSA to stop the reaction
and then incubated in pre-warmed media containing LPS and TACE
inhibitor for 2 hrs at 37°C. Cells were washed, fixed and
immunostained for either surface or internal TNFα.
-
3
Cell surface labelling and FACS analysis. Raw264.7 macrophages
were electroporated with either GFP-alone or GFP-VAMP3, cultured
for 24 hours, stimulated with LPS for 2 hours and the cell surface
was labeled, according to the manufacturer’s instructions, with 2
µM DilC18(5)-DS fluorescently labeled lipophilic dye (Molecular
Probes) for 2 min followed by a 15 min incubation on ice. Cells
were washed three times with ice-cold PBS and fixed using 4%
paraformaldehyde. Confocal microscopy confirmed that the lipophilic
dye labeling was restricted to the cell surface. Immunofluorescence
analysis was performed using the FACSVantage SE DiVa (Becton
Dickinson) and the subsequent data was analyzed using WinMDI
software (Joseph Trotter, The Scripps Research Institute, La Jolla,
CA). The relative amount of DilC18(5)-DS lipophilic dye on the
plasma membrane was used as an indicator of cell surface area and
was assessed on the basis of mean fluorescence intensity (MFI), in
arbitrary units. Transfection itself did not alter cell size. Live
cell imaging. RAW264.7 cells cultured on glass bottom microwell
dishes (MatTech, USA) were imaged in CO-2 independent media (Gibco,
Invitrogen) at 37°C using a heated stage on a Zeiss LSM510 META
confocal microscope (Carl Zeiss Microscope Systems, Germany)
equipped with a 60 or 100 X oil objective. Movie frame capture
rates were between 1.5 and 12 seconds, with total capture periods
ranging from 5 to 40 minutes. Movies were cropped, constructed and
analyzed using Zeiss LSM software version 3.2, and exported as
Quicktime movies with playback speeds of 8-10 frames per second.
Scanning Electron Microscopy. RAW264.7 cells cultured on 12mm glass
coverslips were fixed in 2% glutaraldehyde in cacodylate buffer,
post-fixed in 1% osmium tetroxide (ProSciTech, Queensland,
Australia) and dehydrated through a graded series of ethanols (5).
The coverslips were washed twice in hexamethyldisilazane (HMDS,
ProSciTech), air dried overnight and then platinum coated using a
Baltec Med020 coater (Bal-Tec, Liechtenstein) before viewing on a
JEOL 6300 Field Emission Scanning Electron Microscope (Jeol
Australasia, Brookswater, NSW, Australia) at 7kV. Images were
captured using ImageSlave software (ImageSlave, Sydney,
Australia).
-
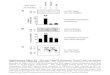
GFP-VAMP3 Rab11
Figure S1. VAMP3 localizes to the RE. Macrophages expressing
GFP-VAMP3 werestimulated with LPS for 2 hours, fixed, permeablized
and immunostained for the REprotein Rab11.
-
20
10
30No LPSPlus LPS
GFP alone GFP-VAMP3
Cel
l sur
face
fluo
resc
ence
DilC18(5)-DS
A B
C
Con
trol
LPS
Figure S2. Increased cell surface in activated macrophages. (A)
LPS-treated anduntreated macrophages expressing either GFP or
GFP-VAMP3 were briefly incubatedwith the lipophilic dye DilC to
label the cell surface. Cells were then fixed and analysedby FACS.
Increasing the level of VAMP3 increased cell surface area. (B)
LPS-activatedand control macrophages were fixed and analysed by
scanning electron microscopy.Control cells have sparse surface
features while activated cells have abundant surfaceruffles which
increase the surface area. (C) GFP-VAMP3 staining on the cell
surfaceis seen concentrated on ruffles. Bars = 5 μM.
GFP-VAMP3
5 μm
5 μm
-
Figure S3. Model showing delivery of TNFα from the RE to the
phagocytic cup and itssubsequent release from the cell surface. The
newly-synthesized, transmembrane precursor ofTNFα is delivered to
the RE which contains the R-SNARE VAMP3. The RE membrane fuses
withthe plasma membrane at the site of a nascent phagocytic cup
where the cognate Q-SNARE complex(syntaxin 4-SNAP23) for VAMP3 is
concentrated. This expands the membrane to accommodatephagocytosis
while simultaneously delivering transmembrane TNFα to the cell
surface. TACE, theenzyme that cleaves TNFα, is strategically
positioned at the cup to ensure rapid cleavage and releaseof
soluble TNFα. Soluble TNFα is released from the membrane before
closure of the cup and ittherefore does not appear with the
ingested microbe in the mature phagosome. This trafficking
stepensures delivery of both membrane and cytokine to the surface
for early inflammatory responses.
VAMP3TNFα
Syntaxin 4TACE
RE
TransmembraneTNFα is deliveredto the phagocytic
cup via the RE
Soluble TNFα isreleased from the cell
surface and isexcluded from thecup before closure
TNFα delivery to and release from the phagocytic cup
VAMP3
-
4
Movie S1. Newly synthesized TNFα is trafficked to the RE.
GFP-TNFα was expressed and imaged in live LPS-stimulated
macrophages at a focal plane through the Golgi complex on a Zeiss
LSM510 META confocal microscope. Newly synthesized GFP-TNFα
concentrates in the Golgi region and GFP-TNFα-containing carriers
move centripetally towards and fuse with a larger compartment (1-2
µm diam) that resembles in size and shape the VAMP3-positive REs
labelled elsewhere in this study (boxed area) (captured at 1.52
second intervals over 34 seconds). Movies S2 and S3. TNFα is
trafficked through a transferrin-positive RE. GFP- TNFα (green) was
imaged in live LPS-stimulated macrophages after uptake of
Alexa-647-conjugated Tfn (red) as a marker of the RE. GFP-TNFα
enters a Tfn-positive RE (boxed area), remains there transiently
and then another GFP-TNFα carrier exits the RE and moves along a
trajectory towards the plasma membrane (captured at 2.2 second
intervals over 30 seconds). Movie S4 and S5. Newly synthesized TNFα
is delivered to the phagocytic cup. GFP- TNFα was imaged live in
Candida albicans-phagocytosing, activated macrophages, movie frames
captured at 8.5 second intervals over 300 seconds (movie 4) and
1700 seconds (movie 5). Movie 4 shows a sequence with GFP-TNFα in
intracellular compartments and carriers and concentrated in the
phagocytic cup as it forms around a yeast particle. GFP-TNFα on the
membrane of the phagocytic cup surrounds the yeast and is then left
concentrated at an outer point as the yeast is ingested. Movie 5
shows GFP-TNFα moving towards and being delivered to a nascent
phagocytic cup. Movie S6. Newly synthesized ApoE is not delivered
to the phagocytic cup. GFP-ApoE was imaged in Candida
albicans-phagocytosing macrophages (frames captured at 1.4 second
intervals for 140 seconds), showing that GFP-ApoE leaving the Golgi
does not move towards the phagocytic cup during phagocytosis. Two
instances can be seen of carriers containing GFP-ApoE moving
towards an area of the plasma membrane away from the phagocytic
cup. Movie S7. GFP-VAMP3 membranes associate with the phagocytic
cup. GFP-VAMP3 was imaged in Candida alibicans-phagocytosing,
stimulated macrophages (frames captured at 7 second intervals for
1225 seconds). GFP-VAMP3 labeled membranes move towards and are
delivered to the phagocytic cup.
-
5
Table S1. R-SNARE gene regulation in macrophages. Changes in
gene expression after 2 and 12 hours stimulation of RAW264.7 cells
with LPS are shown for R-SNARE genes present on the array.
Gene Accession number 2 hours LPS 12 hours LPS VAMP1 NM_009496
ND ND VAMP2 NM_009497 NC NC VAMP3 NM_009498 I (+1.2) I (+2.8) VAMP4
NM_016796 NC D (-1.4) VAMP5 AK009266 ND ND VAMP7 BC003764 NC NC
VAMP8 NM_016794 NC NC
Key: NC is no change, ND is not detected, I is increased and D
is decreased, with fold increases indicated.
-
6
References and Notes
1. R. Z. Murray, F. G. Wylie, T. Khromykh, D. A. Hume, J. L.
Stow, J. Biol. Chem. 280, 10478 (2005).
2. W. Shurety, A. Merino-Trigo, D. Brown, D. A. Hume, J. L.
Stow, J. Interferon Cytokine Res. 20, 427 (2000).
3. J. K. Pagan et al., Curr. Biol. 13, 156 (2003). 4. A. L. Ang
et al., J. Cell Biol. 167, 531 (2004). 5. S. Seveau, H. Bierne, S.
Giroux, M. C. Prevost, P. Cossart, J. Cell Biol. 166, 743
(2004).