Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, Nov. 1978, P. 582-5870021-9193/78/0136-0582$02.00/0Copyright © 1978 American Society for Microbiology
Vol. 136, No. 2
Printed in U.S.A.
Synthesis and Salvage of Purines During CellularMorphogenesis of Myxococcus xanthus
WEN-CHERNG TSAIt AND CARL A. WESTBY*Department of Microbiology, South Dakota State University, Brookings, South Dakota 57007
Received for publication 17 July 1978
Intact cells of Myxococcus xanthus were examined for de novo purine synthesisand salvage utilization. The cellular uptake rates of radioactive glycine (de novopurine precursor), adenine, and guanine were measured, and thin-layer chroma-tography and radioautography were used to examine cell extracts for de novo-synthesized purine nucleotides. Intact vegetative cells, glycerol-induced myxo-spores, and germinating cells of M. xanthus CW-1 were able to carry out de novopurine and salvage synthesis. Germinating cells and glycerol-induced myxosporeswere metabolically more active or as active as vegetative cells with respect topurine anabolism. We conclude that M. xanthus is capable of synthesizing purinenucleotides and salvaging purines throughout the glycerol version of its life cycle.
We previously reported on the discovery ofcertain de novo purine-synthesizing enzymes invegetative and myxospore extracts of Myxococ-cus xanthus (19). The scope of that report wastoo limited, however, to draw unequivocal con-clusions concerning purine synthesis in the in-tact myxospore. The possibility still existed thatthe enzymes, although present and measurablein myxospore extracts, did not actually functionin the whole cell because of stringent regulatorycontrols. Certain of our findings, when viewedagainst the more recent results of Hanson andDworkin (6), seem to rule out feedback inhibi-tion as a major deterring force of myxospore denovo purine synthesis. We found that AMP,GMP, ADP, GDP, and GTP were the mostpotent inhibitors of M. xanthus PP-ribose-Pamidotransferase but that the inhibition im-posed by AMP, GMP, and presumably othernucleotides dropped off sharply as the concen-tration was reduced from 6.25 mM to 1 to 2 mMand was very slight at the lower level. The lowerconcentration, 1 to 2 mM, is noteworthy becauseit would appear (6) from our calculations and byassuming a myxospore volume of 4 tLm3 to cor-respond very nearly to the additive intracellularconcentration in the myxospore of all of theabove important amidotransferase inhibitors.This means that major feedback inhibition of denovo purine synthesis is missing in the intactmyxospore unless there is a major synergisticeffect.To confirm the absence of large-scale feedback
inhibition of de novo purine synthesis in thet Present address: Department of Medical Microbiology,
National Yang-Ming Medical College, Shih-Pai, Taipei, Tai-wan, Republic of China.
myxospore and to rule out any dormancy-im-posed control due to lack of substrates or cellulardehydration, we have conducted intact-cell stud-ies on myxospores and vegetative and germinat-ing cells to measure de novo purine synthesis.Because purine salvage synthesis is interrelatedwith de novo synthesis, we also examined thisaspect of purine nucleotide anabolism duringour studies on intact cells.
MATERIALS AND METHODS
Chemicals. ATP (disodium), AMP (type V), GTP(trisodium, type I), GMP (sodium), CMP (disodium),UTP (sodium, type III), cocarboxylase (aneurine py-rophosphate: thiamine pyrophosphate chloride) andS-adenosyl-L-methionine iodine were obtained fromSigma Chemical Co. Guanine-hydrochloride and thia-mine-hydrochloride were purchased from NutritionalBiochemicals Co. Diethylamine was from EastmanOrganic Chemical Co., and [2-14C]glycine (5.97mCi/mmol), [8-'4C]adenine (53.5 mCi/mmol), and [8-"C]guanine (52.01 mCi/mmol) were obtained fromNew England Nuclear Corp.Organism and cultivation conditions. M. xan-
thus strain CW-1 was obtained from M. Dworkin,University of Minnesota, as strain FB.CW-1 cells were grown in low levels of free purine
and were assumed to be partially repressed and onlypartly feedback inhibited for de novo purine synthesis.The cells were obtained by using CT medium (3)containing pretested Casitone (2% solution that repro-ducibly supported a 4.5-h generation time) which hadbeen previously dialyzed to remove free purines. Themedium was prepared by dialyzing 4% Casitone for 22h against running tap water and for 2 h against runningdeionized water, diluting the residue to 2% dissolvedsolids (wt/vol) with deionized water, and finally addingthe other CT components. This 2/ dialyzed CT me-dium contains (unpublished data) a very low level of
582
on Decem
ber 4, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

M. XANTHUS CELL CHANGE AND PURINE ANABOLISM 583
free purines, 1.0 ig of adenine equivalents per ml. Itwas sterilized by filtration (Nalge 0.45-am disposablefilter unit) rather than by autoclaving to prevent theformation of a precipitate that was found to resultupon steam sterilization of dialyzed Casitone. Thedialyzed CT medium was prepared in 100-ml quanti-ties in 300-ml screw-capped side-arm (14-mm diame-ter) flasks, and at the beginning of an experiment theflasks were inoculated 1:14 (vol/vol) from CT culturesincubated for 20 to 24 h in roller tubes. The inoculumwas obtained from deep-freezer stocks of vegetativecells or myxospores. The cultures were incubated at30°C in a gyratory-shaking water bath (Metabolytewater bath shaker, New Brunswick Scientific Co.,speed setting of 175 rpm) until the turbidity of theflask contents had reached an absorbancy of 0.4(Bausch & Lomb, Spectronic 20) at 520 nm. Thiscorresponds to 1.1 x 108 log-phase cells per ml growingat a generation time of 4.5 to 5.5 h.
Harvesting and preparation of vegetativecells. CW-1 vegetative cells were harvested by cen-trifugation (5,900 x g for 15 min at 4°C), washed oncewith 160 ml of chilled potassium phosphate buffer(0.01 M, pH 7.2) containing 0.008 M MgSO4, and thenresuspended in 10 ml of the same phosphate-MgSO4buffer.Myxospore production. Vegetative cells were
washed once with 100 ml of dialyzed and chilled 1%Casitone containing 0.008 M MgSO4. The cells werethen suspended for 1 to 8 h in 200 ml of myxosporeinduction medium with the same ingredients as thewash medium plus 0.5 M glycerol. To ensure adequateaeration for induction (4), the suspended cells werevigorously shaken (as above) in a vessel with a largesurface-to-volume ratio (1,200-ml Fleaker flask) andbubbled with air (MicroFerm Laboratory fermentorair supply system, New Brunswick Scientific Co., airpressure set at 4.4 instrument units). The various lotsof myxospores were harvested by centrifugation asabove.Those batches not to be germinated were washed
in 160 ml of chilled potassium phosphate buffer (asabove) containing 0.5 M glycerol and were suspendedin 10 ml of the same solution.Myxospore germination. Eight-hour myxospores
were germinated by washing and suspending (10 ml)in chilled buffer (as above) lacking glycerol, followedby incubation at 30°C for 35 to 50 min (12) and dilution(see below).
All the batches of myxospores except 1-h myxo-spores were sonically disrupted once for 20 s afterwashing and suspension (Branson Ultrasonifier, model9110, with a small probe with 3-mm-diameter tip,power setting of 3) to break up sticky clumps of sporesand produce a uniform suspension. Individual myxo-spores are resistant to such treatment and do notrupture (15). Each lot of myxospores was checked byphase microscopy for cellular uniformity before beingused in glycine, adenine, or guanine uptake studies.Measurement of glycine, adenine, and guanine
uptake. A slight modification of the method of White(personal communication) and Ramsey and Dworkin(12) was employed to measure separately the rates ofuptake of glycine, adenine, and guanine by vegetativecells, myxospores, and germinating cells of CW-1.
In the modified version, the same solution used forthe final suspension was employed to dilute (1:10) twoto four replicate samples of the different 10-ml cellsuspensions. The uptake substances as "C-isotopeswere present at a final concentration of 1 ,iCi/ml, andafter their addition to cells, 1-mi samples were mea-sured for uptake at 0, 5, 10, 15, and 30 min.The zero-time background for each cell type was
always rather high, about 600 to 10,000 pmol/109washed cells contained on a filter pad rinsed with 3 mlof suspension buffer. This may reflect the technicaldifficulty in our system of obtaining exact zero-timesamples and the immediate high rate of isotope uptakeby rather large populations of the different cell types.Data processing of scintillation and cell count
data. A computer program (in BASIC) was devised toconvert counts per minute of [2-'4C]glycine, [8-'4C]-adenine, or [8-'4C]guanine taken up per number ofcells per time of incubation to picomoles of substancetaken up per 109 cells per min. A cell count value of109 was chosen because the results of a recent relatedstudy (6) are reported in these terms.
Labeling of vegetative cell and myxospore pu-rines with [2-'4C]glycine. Certain batches of vege-tative cells, 1- to 8-h myxospores and germinating cellswere exposed to [2-'4C]glycine to allow for uptake andincorporation of this purine nucleotide precursor intotrichloroacetic acid-soluble purine nucleotides andthiamine. To each 10 ml of cell suspension of 1.1 x 109cells per ml was added 0.045 mCi of [2-'4C]glycine andthe 14C-treated cells were incubated at 30°C in ashaking water bath (as above) for 90 min to allow forglycine uptake and utilization in de novo purine syn-thesis. Germinating myxospores were incubated foronly 50 min. After the incubation, each cell suspensionwas harvested by centrifugation (as above) andwashed once with 160 ml of the same buffer used forfinal suspension.
Extraction of labeled purines. After washing,cells that had been exposed to [2-'4C]glycine weretreated with 3 ml of cold 10% trichloroacetic acid in30% methanol in an ice bath with occasional mixing toliberate trichloroacetic acid-soluble components fromthe cells. After 50 min of such treatment, a precipitatethat formed was removed by centrifugation (27,000 xg for 30 min at 0°C) and was discarded. The superna-tant fluid was extracted five times with 9 ml of cold,water-saturated diethyl ether to remove the trichlo-roacetic acid (6). Various samples so obtained werefrozen overnight at -70°C and lyophilized on the nextday. The lyophilized residues were diluted with 0.15to 0.2 ml of distilled water and were either useddirectly for thin-layer chromatography or frozen(-70°C) and used later.
Partial purification of cellular ATP. Portions(10 to 30 pl) of the lyophilized, trichloroacetic acid-soluble extracts of vegetative cells, myxospores, andgerminating cells prepared as described above werespotted on untreated thin-layer chromatoplates (20 by20 cm, plastic backed, Mackerey-Nagel Polygram Cel300). Control chromatoplates were spotted with mix-tures containing 10 ,l of 5 mM AMP, ATP, GMP,GTP, CMP, UTP, thiamine-hydrochloride, thiaminepyrophosphate, or S-adenosyl-methionine and 10Ml ofthe extract.
VOL. 136, 1978
on Decem
ber 4, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

584 TSAI AND WEST'BY
The dried plates were chromatographed (ascend-ing) in the first dimension with n-butanol:ace-tone:diethylamine:water (10: 10:2:5) and then, after thetop part of each chromatogram had been cut off justbelow the pH front, in the second dimension with 1.5M potassium phosphate, pH 3.4. Purines were partiallyseparated from one another by this procedure.
Hydrolysis of [14C]ATP and separation ofproducts. ['4C]ATP spots wherever identifiable byUV absorbance and radioautography were cut fromthe two-dimensional chromatograms. These spotswere not always distinct, and the ATP mav have beencontaminated with UTP, thiamine pyrophosphate,and other purines. Enough chromatograms were avail-able so that four ATP spots from four separate chro-matograms were obtained for each cell type. Thechromatogram spots from like cell types were com-
bined and extracted with 2.5 ml of deionized water.The extracted ['4C]ATP was dephosphoribosylatedwith perchloric acid (10) to free adenine from theribose of ATP. '4C in the ribose of ATP would notnecessarily indicate de novo synthesis but could beexplained by salvage synthesis. The glycine carboncould have been deployed into ribose by the glyoxalatecycle, gluconeogenesis, and transketolase reactionsand then into purine nucleotides by salvage enzymes
(18). '4C in the adenine could only be explained by denovo synthesis. After the acidified product containingfree adenine had been neutralized with dilute NaOH,20 to 30 pl of the resultant solution was co-chromato-graphed with 5 mM adenine in one dimension againstdeionized water (ascending) on a thin-layer plate(Polygram Cel 300). After drying, the chromatogramwas examined for ['4C]adenine spots by checking forUV spots corresponding to known adenine and byradioautography.
Radioautography. Radioautography was accom-
plished by exposing the chromatogram to X-ray film,(Kodak single-coated X-ray film, blue-sensitive codeSB54) for 7 to 30 days.
RESULTS
Uptake of purine de novo precursor gly-cine by M. xanthus. The results of a [2-14C]glycine uptake study employing vegetativecells and myxospores (germinating and nonger-minating) obtained from dialyzed CT growthand induction media are indicated in Table 1.The results show that glycerol myxospores atvarious stages of maturation, as well as vegeta-tive cells and 8-h germinating myxospores, takeup measurable amounts of glycine when extra-cellular free purine and glycine levels are low.We have observed in other uptake tests that ifthe cells, of whatever type, are grown or main-tained in CT medium supplemented with 100ytg of adenine or guanine-hydrochloride per ml,they take up glycine at a lower rate.Adenine and guanine uptake by vegeta-
tive cells, myxospores, and germinatingcells. Besides being able to take up glycine,purine-limited vegetative cells, myxospores, and
TABLE 1. Initial rates of glycine, adenine, andguanine uptake by M. xanthus CW-1 cellsmaintained at low external purine levels
Initial rate of compound uptake'
Cell type (pmol/min per 109 cells)Glycine Adenine Guanine
Vegetative 1,138 105 31-h Myxospore 552 113 202.5-h Myxospore 911 169 138-h Myxospore 1,114 93 35Germinating 8-h 1,695 188 214myxosporea Rate for initial 30-min exposure was determined
on minicomputer from polynomial regression analysisof slope of picomoles of uptake versus 0- to 30-mintime plot.
germinating cells can also take up adenine andguanine, as shown in Table 1. We found little orno delay in the start of uptake of the purines bythe different cell types.
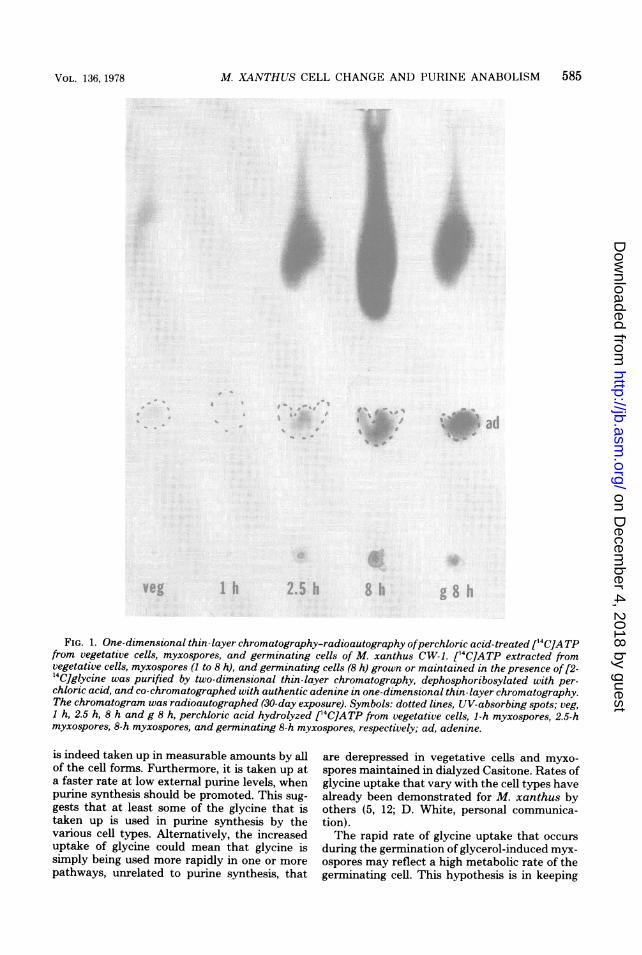
[2-1'C]Glycine incorporation into adeninering ofATP in intact cells. Figure 1 shows theresults of a one-dimensional thin-layer chroma-tography-radioautography of the perchlorate-hydrolyzed ATP pools of vegetative cells, 1- to8-h myxospores and 8-h germinating myxosporespreviously given [2-14C]glycine (perchloratefrees adenine from ATP). The extracted cellswere grown or maintained in solutions contain-ing low external levels of purines, such as di-alyzed CT medium, to promote purine de novosynthesis. The radioactive and UV-absorbingspots on the lower part of the radioautogram(Rf, 0.31) of Fig. 1 correspond to adenine. Theexistence of radioactivity in all of the adeninespots except that of 1-h myxospores indicatesthat myxospores of M. xanthus as well as vege-tative cells can deploy carbon-2 of glycine intothe adenine part of ATP. This means that M.xanthus can carry out purine de novo synthesisthroughout the glycerol version of its life cycle,except perhaps during the initial stages of myx-ospore formation.
Substances such as ribose, uracil, certain nu-cleosides, and nucleotides that may have beenpresent in the perchlorate-hydrolyzed cell ex-tracts along with adenine, all chromatographwithin the region of the fast-moving radioactivespots observed in Fig. 1 (Rf, 0.6 to 0.99) muchabove adenine.
DISCUSSIONIf purine de novo synthesis were to occur to
any extent in the various cellular stages of M.xanthus, uptake of external glycine would pro-vide a convenient source of this necessary purineprecursor. Our results demonstrate that glycine
J. BACTERIOL.
on Decem
ber 4, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
M. XANTHUS CELL CHANGE AND PURINE ANABOLISM 585
FIG. 1. One-dimensional thin-layer chromatography-radioautography ofperchloric acid-treated [14C]ATPfrom vegetative cells, myxospores, and germinating cells of M. xanthus CW-1. [14C]ATP extracted fromvegetative cells, myxospores (I to 8 h), and germinating cells (8 h) grown or maintained in the presence of [2-'4C]glycine was purified by two-dimensional thin-layer chromatography, dephosphoribosylated with per-chloric acid, and co-chromatographed with authentic adenine in one-dimensional thin-layer chromatography.The chromatogram was radioautographed (30-day exposure). Symbols: dotted lines, UV-absorbing spots; veg,1 h, 2.5 h, 8 h and g 8 h, perchloric acid hydrolyzed ['4C]ATP from vegetative cells, 1-h myxospores, 2.5-hmyxospores, 8-h myxospores, and germinating 8-h myxospores, respectively; ad, adenine.
is indeed taken up in measurable amounts by allof the cell forms. Furthermore, it is taken up ata faster rate at low external purine levels, whenpurine synthesis should be promoted. This sug-gests that at least some of the glycine that istaken up is used in purine synthesis by thevarious cell types. Alternatively, the increaseduptake of glycine could mean that glycine issimply being used more rapidly in one or morepathways, unrelated to purine synthesis, that
are derepressed in vegetative cells and myxo-spores maintained in dialyzed Casitone. Rates ofglycine uptake that vary with the cell types havealready been demonstrated for M. xanthus byothers (5, 12; D. White, personal communica-tion).The rapid rate of glycine uptake that occurs
during the germination of glycerol-induced myx-ospores may reflect a high metabolic rate of thegerminating cell. This hypothesis is in keeping
VOL. 136, 1978
on Decem
ber 4, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

586 TSAI AND WESTBY
with the findings of Dworkin (personal commu-nication) that during their initial round of celldivision, germinated myxospores have a fastergrowth rate than vegetative cells. Another ex-planation for the increased glycine uptake is thatthe extra glycine is used to synthesize freshsupplies of purine nucleotides to replace thoselost from the cell during germination by leakageor purposeful excretion. The reported extracel-lular accumulation of AMP and GMP that ac-companies germination (6) is consistent withthis explanation.Although the glycine uptake results do not
conclusively prove that glycerol-induced myxo-spores conduct purine de novo synthesis, thethin-layer chromatography-radioautography re-sults do unequivocally demonstrate this formyxospores maintained in low purine concen-trations. The thin-layer chromatography-radioautographic results also indicate that veg-etative cells, myxospores of various ages, andgerminating cells also carry out de novo purinesynthesis.
Although purine de novo synthesis seems tooccur consistently throughout most of the glyc-erol life cycle of M. xanthus, it apparently doesstop or is severely reduced at the first hour ofmyxospore induction (Fig. 1). This is of interestbecause it occurs at about the same time as a50% decline in glycine uptake (Table 1), a haltin intracellular purine pool buildup (6), and adecrease in the rate of net nucleic acid synthesis(1, 8, 13). These events suggest that little or noglycine is committed to making purine nucleo-tides and nucleic acids during the first hour ofglycerol induction. The supply of internal gly-cine may be diminished by reduced uptake andsynthesis (2) or by increased demands on theavailable glycine, and this may control purinesynthesis.An alternative explanation (D. White, per-
sonal communication) for these events is thatthe glyoxalate cycle, which is only fully operativein the 1- to 3-h myxospore (9, 11), generatesenough intracellular glycine to saturate the cel-lular pool of this amino acid. This would reducethe specific activity of any [2-14C]glycine takenup by the myxospore and lower the specificactivity of purine nucleotides formed from theglycine of this pool. The apparent decline inpurine synthesis at 1 h seen in Fig. 1 thereforemay not be a real decline or it may not be asexaggerated as would be indicated by Fig. 1.This alternative hypothesis implies that themyxospore, like the vegetative cell (2), is proto-trophic for glycine.
Regardless of which explanation of the eventsis accepted, feedback inhibition of the purine de
novo pathway certainly does not appear to beimportant in regulation.The first indication that purine salvage is pres-
ent in M. xanthus came from the finding thatpurine bases, nucleosides, or nucleotides cansupport the growth of purine-requiring mutants(7). Our finding that both adenine and guaninecan be taken up by vegetative cells, myxospores,and germinating cells presents evidence that allcell forms of M. xanthus are capable of purinesalvage. Adenine is taken up at a more rapidrate than guanine by all cell types, and bothpurines are taken up at a markedly lower ratethan glycine. It would appear from this thatboth exogenous adenine and guanine play a rolein the metabolism of M. xanthus but that quan-titatively adenine is more important than gua-nine. Presumably the purines taken up are con-verted to functional and interchangeable nucleo-tides. Because there occurs no lag period foreither adenine or guanine cellular incorporation,it seems likely that the enzymes responsible fortheir transport into cells are constitutive ratherthan inducible.Our results showing active glycine and purine
uptake by the myxospore as well as substantivepurine synthesis, salvage, and interconversionseem somewhat inconsistent with the supposeddormancy of this resting cell. The so-called rest-ing state of the myxospore from our vantagepoint seems to be typified by a reordering of themetabolism rather than an abrupt halt or indeedeven a significant slowdown. Active purine anab-olism appears to be as important to the maturingmyxospore as it undoubtedly is to the vegetativeand germinating cell. The emphasis and direc-tion of purine anabolism may indeed changeduring and after myxospore formation, but itsoverall importance seems to be undiminishedthroughout glycerol-induced morphogenesis inM. xanthus.The presence of active purine synthesis in the
"dormant" glycerol myxospore means that suf-ficient purine precursors are available intracel-lularly and the myxospore is hydrated enoughto allow this anabolic process to proceed. Incontrast, another dormant, dehydrated, procar-yotic cellular body, the Bacillus endospore, doesnot conduct purine synthesis under any circum-stances (14, 16, 17). Whether de novo purinesynthesis occurs during amino acid or polya-mine-induced myxospore formation (20) or dur-ing fruiting body formation is still unknown.
LITERATURE CITED
1. Bacon, K., and E. Rosenberg. 1967. Ribonucleic acidsynthesis during morphogenesis in Myxococcus xan-
thus. J. Bacteriol. 94:1883-1889.
J. BACTERIOL.
on Decem
ber 4, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

M. XANTHUS CELL CHANGE AND PURINE ANABOLISM 587
2. Bretscher, A. P., and D. Kaiser. 1978. Nutrition ofMyxococcus xanthus, a fruiting myxobacterium. J. Bac-teriol. 133:763-768.
3. Dworkin, M., and S. M. Gibson. 1964. A system forstudying microbial morphogenesis: rapid formation ofmicrocysts in Myxococcus xanthus. Science 146:243-244.
4. Dworkin, M., and W. Sadler. 1966. Induction of cellularmorphogenesis in Myxococcus xanthus. I. General de-scription. J. Bacteriol. 91:1516-1519.
5. Filer, D., D. White, S. H. Kindler, and E. Rosenberg.1977. Myxospore coat synthesis in Myxococcus xanthus:in vivo incorporation of acetate and glycine. J. Bacteriol.131:751-758.
6. Hanson, C. W., and M. Dworkin. 1974. Intracellularand extracellular nucleotides and related compoundsduring the development of Myxococcus xanthus. J.Bacteriol. 118:486-496.
7. Hemphill, H. E., and S. A. Zahler. 1968. Nutrition ofMyxococcus xanthus FBa and some of its auxotrophicmutants. J. Bacteriol. 95:1011-1017.
8. Kimchi, A., and E. Rosenberg. 1976. Linkages betweendeoxyribonucleic acid synthesis and cell division inMyxococcus xanthus. J. Bacteriol. 128:69-79.
9. Kottel, R. H., M. Orlowski, D. White, and J. Grutsch.1974. Presence of amino acid dehydrogenases and trans-aminases in Myxococcus xanthus during vegetativegrowth and myxospore formation. J. Bacteriol. 119:650-651.
10. Marshak, A., and H. J. Vogel. 1951. Microdetermina-tion of purines and pyrimidines in biological materials.J. Biol. Chem. 189:597-605.
11. Orlowski, M., P. Martin, D. White, and M. C.-W.Wong. 1972. Changes in activity of glyoxylate cycleenzymes during myxospore development in Myxococcus
xanthus. J. Bacteriol. 111:784-790.12. Ramsey, W. S., and M. Dworkin. 1968. Microcyst ger-
mination in Myxococcus xanthus. J. Bacteriol. 95:2249-2257.
13. Rosenberg, E., M. Katarski, and P. Gottlieb. 1967.Deoxyribonucleic acid synthesis during exponentialgrowth and microcyst formation in Myxococcus xan-thus. J. Bacteriol. 93:1402-1408.
14. Setlow, P., and A. Kornberg. 1970. Biochemical studiesof bacterial sporulation and germination. XXIII. Nu-cleotide metabolism during spore germination. J. Biol.Chem. 245:3645-3652.
15. Sudo, S. Z., and M. Dworkin. 1969. Resistance of vege-tative cells and microcysts of Myxococcus xanthus. J.Bacteriol. 98:883-887.
16. Switzer, R. L., C. L. Turnbough, Jr., J. S. Brabson,and L. M. Waindle. 1975. Enzyme inactivation of denovo nucleotide biosynthesis in sporulating Bacillussubtilis cells, p. 327-334. In P. Gerhardt, R. N. Costilow,and H. L. Sadoff (ed.), Spores VI. American Society forMicrobiology, Washington, D.C.
17. Turnbough, C. L., Jr., and R. L. Switzer. 1975. Oxygen-dependent inactivation of glutamine phosphoribosyl-pyrophosphate amidotransferase in stationary-phasecultures of Bacillus subtilis. J. Bacteriol. 121:108-114.
18. Watson, B. F., and M. Dworkin. 1968. Comparativeintermediary metabolism of vegetative cells and micro-cysts of Myxococcus xanthus. J. Bacteriol. 96:1465-1473.
19. Westby, C. A., and W.-C. Tsai. 1974. De novo purinesynthesis in vegetative cells and myxospores of Myxo-coccus xanthus. J. Bacteriol. 117:1099-1107.
20. Witkin, S. S., and E. Rosenberg. 1970. Induction ofmorphogenesis by methionine starvation ofMyxococcusxanthus: polyamine control. J. Bacteriol. 103:641-649.
VOL. 136, 1978
on Decem
ber 4, 2018 by guesthttp://jb.asm
.org/D
ownloaded from





























