Embed Size (px)
Citation preview

Plant Physiol. (1970) 46, 831-838
Synthesis and Transport of Hydroxyproline-rich Components inSuspension Cultures of Sycamore-Maple Cells1
Received for publication October 10, 1969
W. V. DASHEK2Michigan State University, Michigan State University-Atomic Energy Commission Plant Research Laboratory,East Lansing, Michigan 48823
ABSTRACT
Plant cell walls contain a glycoprotein rich in hydroxy-proline. To determine how Acer pseudoplatanus L. cellstransport this glycoprotein to the wall, the pulse-chasetechnique was used to follow changes in specific radio-activity of hydroxyproline and proline in isolated, mito-chondrial, Golgi, microsomal, soluble protein, and wallfractions. The turnover rates or changes in specific radio-activity of cytoplasmic hydroxyproline in these cell frac-tions indicated that the bulk of this hydroxyproline wastransferred not by the Golgi apparatus but by a smoothmembranous component.
Plant cell walls contain a glycoprotein rich in hydroxyproline(9, 10). Labeling experiments (9, 14) suggest that this glycopro-tein is synthesized in the cytoplasm and subsequently transferredto the wall. Dashek and Rosen (4) exposed pollen tubes to pulsesof 'H-proline and then employed electron microscope radioautog-raphy. Their results suggested that the transfer of the glycopro-tein might occur via the Golgi apparatus and its associated vesi-cles.The present paper reports the results of an attempt to determine
whether or not the Golgi apparatus does indeed transport a hy-droxyproline-rich glycoprotein. The pulse-chase technique wasused to follow changes in the specific radioactivity of hydroxypro-line and proline as a function of time of labeling or chase in or-ganelles isolated from cells exposed to '4C-proline.
MATERIALS AND METHODS
Growth Conditions. Sycamore-maple (Acer pseudoplatanus L.)cells were cultured as previously described (8).
Preparation of Cell Fractions. Ten-milliliter aliquots, with-drawn from 100-ml cultures (grown for 7 days), were suspendedin 4.5 ml of isolation medium which consisted of 0.1 M NaCl,0.001 M CaCl2, 1% dextran (molecular weight 86,000), 0.1% glu-taraldehyde, and 0.5 M sucrose (12, 13). The addition of glutar-aldehyde helps to prevent fragmentation of Golgi bodies during
1 The results reported herein were obtained while the author wasa National Institute of Health post-doctoral fellow (1-F2-GM22,422-01). The work was supported by the United States Atomic EnergyCommission.
2Present address: Department of Biology, Virginia CommonwealthUniversity, Richmond, Virginia 23220.
isolation (13), but to check the possibility that glutaraldehydemight cause adsorption of soluble proteins onto the isolated or-ganelles, the aldehyde was omitted from some experiments. Cellswere usually ruptured by a 90 sec sonication with a BronsonSonifier (setting No. 3, 40-60 milliamps). Figures 1 and 2 detailthe methods for isolation and purification of walls, organelles andsoluble proteins. The 7,000 and 10,000 rpm centrifugations werecarried out with a Sorvall preparative centrifuge and were equiva-lent to 6,040 and 12,100g respectively. Speeds of 20,000 (43,000g)and 37,000 (150,000g) rpm were reached with Spinco SW-39 andSW-50 swinging bucket rotors. Golgi bodies were prepared byboth differential and sucrose gradient centrifugation as describedby Morre and Mollenhauer (12) and Morre et al. (13).
Electron Microscopy. Intact cells were fixed for 30 min in 6.25%glutaraldehyde buffered with 0.1 M phosphate at pH 7.2 (18) andpostfixed for 1 hr in either phosphate-buffered 2% OS04 or 2%aqueous KMnO4. Some cells were fixed only in KMnO4. Or-ganelles isolated in sucrose-rich medium which either containedor lacked glutaraldehyde were suspended in OS04 or KMnO4 for1 hr. Following fixation, intact cells and isolated organelles weredehydrated through ethanol or acetone, embedded in Epon, andpolymerized in Beem capsules. Gold and silver sections weresometimes counterstained with lead citrate (16) before viewing ineither Phillips 100 or Siemens Elimskop 1A electron micro-scopes.
Labeling Procedures: To trace the cytoplasmic movement ofhydroxyproline-rich material from its site of synthesis to its siteof deposition, pulse-chase experiments were carried out and thechanges in specific radioactivity of hydroxyproline and prolinefollowed as a function of time of labeling or chase in isolated or-ganelles. For each experiment, a 100-ml culture was exposed to10 to 20 ,uc of "4C-proline (specific radioactivity 200 Ac/mole)and 10-mil aliquots were withdrawn at intervals. Following a 2-min centrifugation (500g) to remove growth medium, cells werefiltered, rapidly washed with ice-cold distilled water, and then re-suspended in ice-cold isolation medium for 15 min to allow "light",prefixation with glutaraldehyde. The cells were then rupturedand the walls and organelles were prepared as described above.Assays for Radioactivity and Amino Acids. Walls and isolated
organelles were hydrolyzed with 6 N HC1 for 36 hr at 105 C insealed tubes. The filtered hydrolyzates were evaporated to dry-ness, redissolved in distilled water, and then passed through anion exchange resin (AG 50W-X4, 200-400 mesh, H4 form). Afterelution of the amino acids with 2N NH40H, the eluates wereevaporated to dryness. Each dried sample was dissolved in 50/Aliter of water. Fifteen /Aliters were used for assaying radioactivityand 35 were used for colorimetric determinations of hydroxypro-line and proline. To determine the amount of radioactive hy-droxyproline and proline, 15 uliters were spotted on WhatmanNo. 4 paper, and the two amino acids were separated by electro-phoresis in a pH 1.9 buffer (acetic acid 8.7%, formic acid 2.5%
831 www.plantphysiol.orgon April 23, 2020 - Published by Downloaded from
Copyright © 1970 American Society of Plant Biologists. All rights reserved.
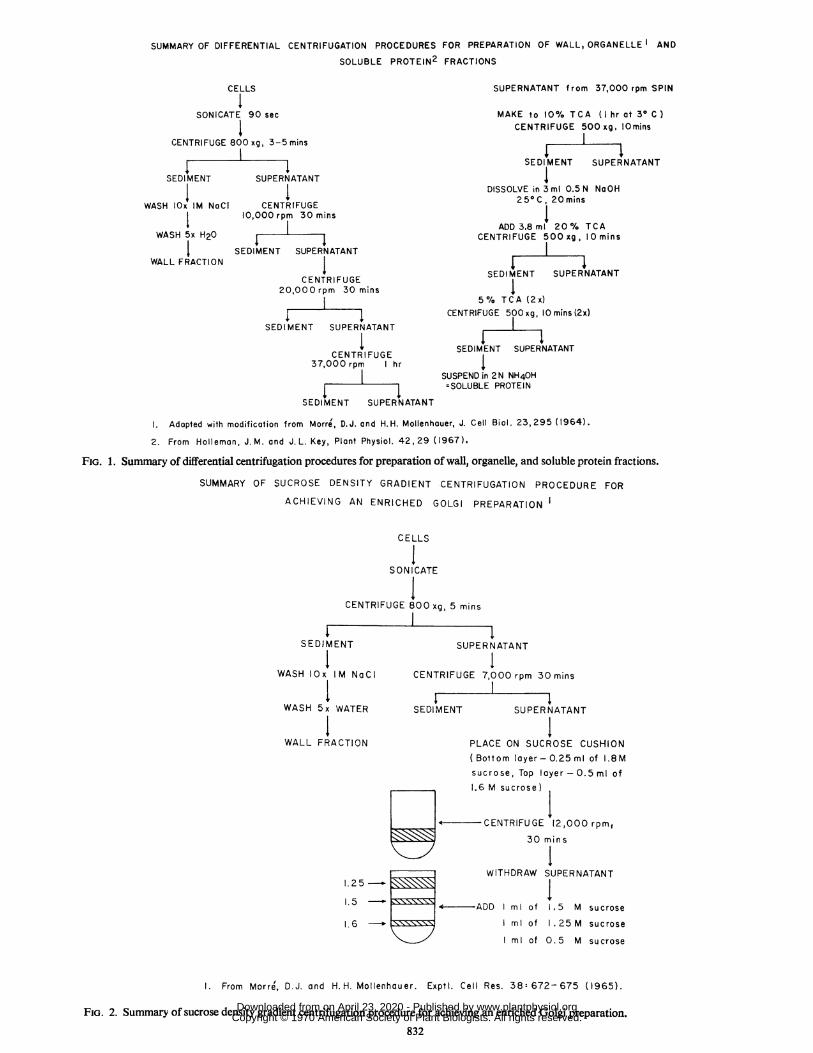
SUMMARY OF DIFFERENTIAL CENTRIFUGATION PROCEDURES FOR PREPARATION OF WALL, ORGANELLE AND
SOLUBLE PROTEIN2 FRACTIONS
CELLS
ISONICATE 90 sec
ICENTRIFUGE 800 xg, 3-5 mins
rI
SUPERNATANT from 37,000 rpm SPIN
MAKE to 10% TCA (I hr at 3° C )CENTRIFUGE 500 xg, IOmins
cF I I
SFr)I MFNT 91IIPFPNATANT
SEDI MENT SUPERNATANT 4
I I DISSOLVE in 3ml 0.5 N NaOHWASH lOx IM NaCI CENTRIFUGE 25°C, 20mins
10,000 rpm 30 mins I; ~~~~~~~~~~~~ADD3.8 ml 20% TCA
WASH 5x H20 CENTRIFUGE 500xg, lOminsSEDIMENT SUPERNATANT
WALL FRACTION ICENTRIFUGE SEDIMENT SUPERNATANT
20,000 rpm 30 mins I5% TCA (2x)
CENTRIFUGE 500xg, 10mins(2x)SEDIMENT SUPERNATANT I
I SEDIMENT SUPERNATANTCENTRI FUGE
37,000 rpm hrSUSPEND in 2 N NH40H=SOLUBLE PROTEIN
SEDIMENT SUPERNATANT
1. Adopted with modification from Morre', D.J. and H.H. Mollenhouer, J. Cell Biol. 23,295 (1964).2. From Holleman, J. M. and J. L. Key, Plant Physiol. 42,29 (1967).
FIG. 1. Summary of differential centrifugation procedures for preparation of wall, organelle, and soluble protein fractions.
SUMMARY OF SUCROSE DENSITY GRADIENT CENTRIFUGATION PROCEDURE FOR
ACHIEVING AN ENRICHED GOLGI PREPARATION
CELLS
ISONICATE
CENTRIFUGE 800 xg, 5 mins
I1-
I
SEDIMENT
IWASH lOx IM NaCI
SUPERNATANT
ICENTRIFUGE 7,000 rpm 30 mins
WASH 5x Vv
WALL FRAC
(ATER SEDIMENT SUPERNATANT
TION PLACE ON SUCROSE CUSHION(Bottom layer-0.25ml of I.8Msucrose, Top layer-0.5ml of1.6 M sucrose)
CENTRIFUGE 12,000 rpm,30 mins
WITHDRAW SUPERNATANT1.25 I
1.51.5 4-Asxx+ ADD ml of 1.5 M sucrose
1.6 ml of .25 M sucrose
ml of 0.5 M sucrose
1. From Morre, D.J. and H. H. Mollenhauer. Exptl. Cell Res. 38: 672- 675 (1965).
FIG. 2. Summary of sucrose density gradient centrifugation procedure for achieving an enriched Golgi preparation.832
www.plantphysiol.orgon April 23, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.
HYDROXYPROLINE-RICH COMPONENTS
-t
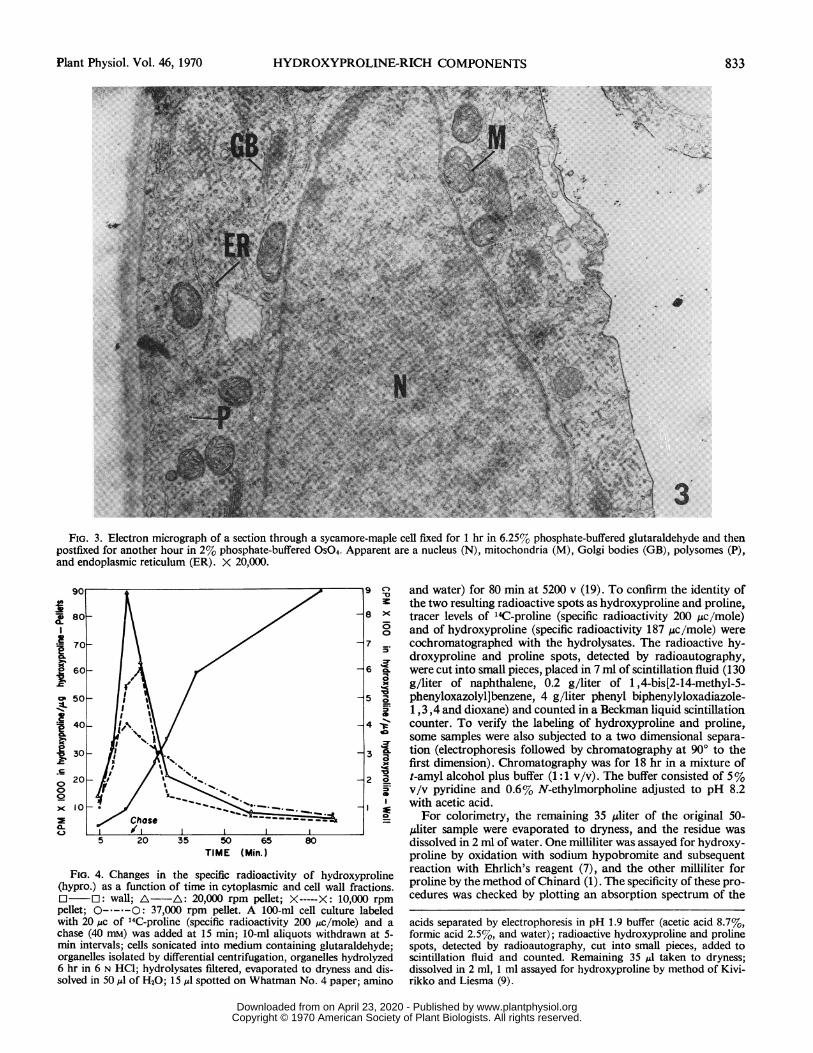
FIG. 3. Electron micrograph of a section through a sycamore-maple cell fixed for 1 hr in 6.25% phosphate-buffered glutaraldehyde and thenpostfixed for another hour in 2% phosphate-buffered OSO4. Apparent are a nucleus (N), mitochondria (M), Golgi bodies (GB), polysomes (P),and endoplasmic reticulum (ER). X 20,000.
I80~~~~~~~~~~~~~~~
70--7/.
060 6 IL
c0 50 8
FIG.4. Changes4inthespeificradioactivitofhydroxyprol
(hpr.30 3fucino iei yolsmcadcl alfatos
o20 wl:00 2 a
)c10 *
pele; -- Chase,00rp pelle.-A100-ml celclur aee
C-~ ~~hs5 20 35 50 65 80
TIME (Min.)
Fio;. 4. Changes in the specific radioactivity of hydroxyproline(hypro.) as a function of time in cytoplasmnic and cell wall fractions.j3 ~: wall; A A 20,000 rpm pellet; X----X: 10,000 rpmpellet; O-*---O: 37,000 rpm pellet. A 100-ml cell culture labeledwith 20,c of 14C-proline (specific radioactivity 200 ,uc/mole) and achase (40 mM) was added at 15 min; 10-ml aliquots withdrawn at 5-min intervals; cells sonicated into medium containing glutaraldehyde;organelles isolated by differential centrifugation, organelles hydrolyzed6 hr in 6 N HCI; hydrolysates filtered, evaporated to dryness and dis-solved in 50,l of H20; 15 ul spotted on Whatman No. 4 paper; amino
and water) for 80 min at 5200 v (19). To confirm the identity ofthe two resulting radioactive spots as hydroxyproline and proline,tracer levels of '4C-proline (specific radioactivity 200 Ac/mole)and of hydroxyproline (specific radioactivity 187 ,uc/mole) werecochromatographed with the hydrolysates. The radioactive hy-droxyproline and proline spots, detected by radioautography,were cut into small pieces, placed in 7 ml of scintillation fluid (130g/liter of naphthalene, 0.2 g/liter of 1 ,4-bis[2-14-methyl-5-phenyloxazolyl]benzene, 4 g/liter phenyl biphenylyloxadiazole-1,3,4 and dioxane) and counted in a Beckman liquid scintillationcounter. To verify the labeling of hydroxyproline and proline,some samples were also subjected to a two dimensional separa-tion (electrophoresis followed by chromatography at 90° to thefirst dimension). Chromatography was for 18 hr in a mixture oft-amyl alcohol plus buffer (1 :1 v/v). The buffer consisted of 5%v/v pyridine and 0.6% N-ethylmorpholine adjusted to pH 8.2with acetic acid.For colorimetry, the remaining 35 ,uliter of the original 50-
,4diter sample were evaporated to dryness, and the residue wasdissolved in 2 ml of water. One milliliter was assayed for hydroxy-proline by oxidation with sodium hypobromite and subsequentreaction with Ehrlich's reagent (7), and the other milliliter forproline by the method of Chinard (1). The specificity of these pro-cedures was checked by plotting an absorption spectrum of the
acids separated by electrophoresis in pH 1.9 buffer (acetic acid 8.7%,formic acid 2.5%, and water); radioactive hydroxyproline and prolinespots, detected by radioautography, cut into small pieces, added toscintillation fluid and counted. Remaining 35 Al taken to dryness;dissolved in 2 ml, 1 ml assayed for hydroxyproline by method of Kivi-rikko and Liesma (9).
Plant Physiol. Vol. 46, 1970 833
'a., . .,.:.I
www.plantphysiol.orgon April 23, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 46, 1970
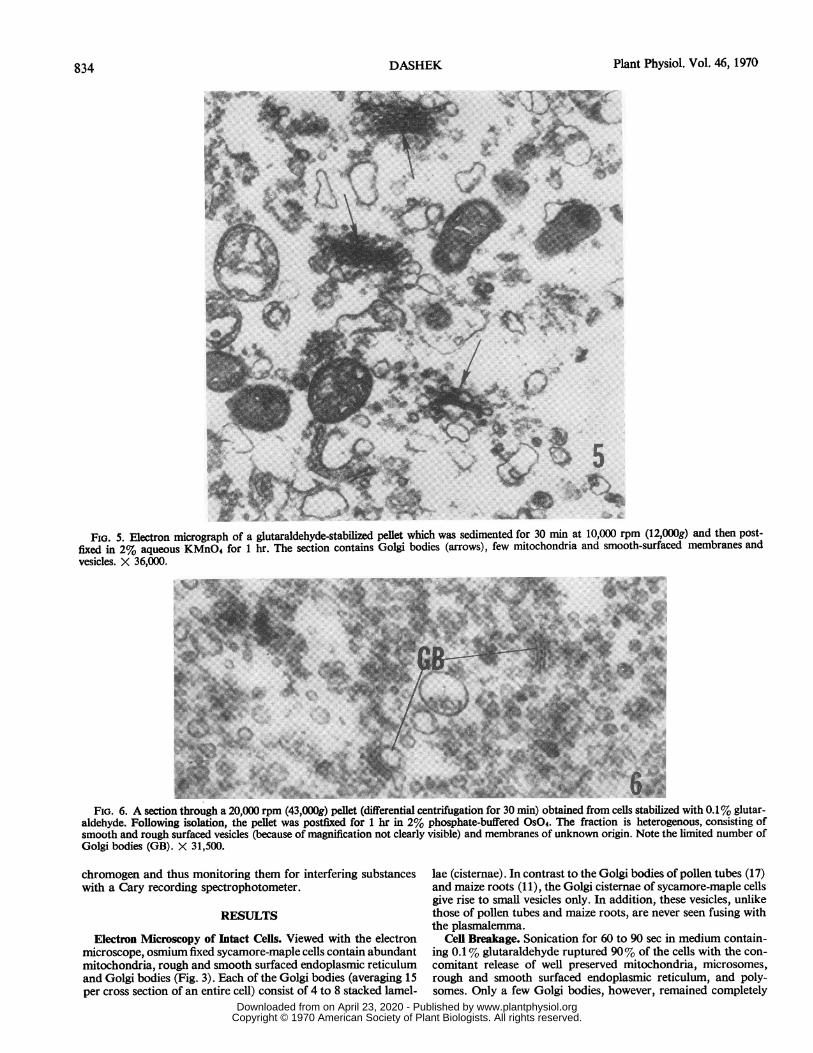
FIG. 5. Electron micrograph of a glutaraldehyde-stabilized pellet which was sedimented for 30 min at 10,000 rpm (12,000g) and then post-fixed in 2% aqueous KMnO4 for 1 hr. The section contains Golgi bodies (arrows), few mitochondria and smooth-surfaced membranes andvesicles. X 36,000.
...B.._.IN~*
'4~~~~~~~~~~~~~~~~~~4-
FIG. 6. A section through a 20,000 rpm (43,000g) pellet (differential centrifugation for 30 min) obtained from cells stabilized with 0.1% glutar-aldehyde. Following isolation, the pellet was postfixed for 1 hr in 2% phosphate-buffered OsO4. The fraction is heterogenous, consisting ofsmooth and rough surfaced vesicles (because of magnification not clearly visible) and membranes of unknown origin. Note the limited number ofGolgi bodies (GB). X 31,500.
chromogen and thus monitoring them for interfering substanceswith a Cary recording spectrophotometer.
RESULTS
Electron Microscopy of Intact Cells. Viewed with the electronmicroscope, osmium fixed sycamore-maple cells contain abundantmitochondria, rough and smooth surfaced endoplasmic reticulumand Golgi bodies (Fig. 3). Each of the Golgi bodies (averaging 15per cross section of an entire cell) consist of 4 to 8 stacked lamel-
lae (cisternae). In contrast to the Golgi bodies of pollen tubes (17)and maize roots (11), the Golgi cisternae of sycamore-maple cellsgive rise to small vesicles only. In addition, these vesicles, unlikethose of pollen tubes and maize roots, are never seen fusing withthe plasmalemma.
Cell Breakage. Sonication for 60 to 90 sec in medium contain-ing 0.1% glutaraldehyde ruptured 90% of the cells with the con-comitant release of well preserved mitochondria, microsomes,rough and smooth surfaced endoplasmic reticulum, and poly-somes. Only a few Golgi bodies, however, remained completely
DASHEK834
www.plantphysiol.orgon April 23, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.

HYDROXYPROLINE-RICH COMPONENTS
FIG. 7. Electron micrograph of a section through a glutaraldehyde-stabilized pellet sedimented at 37,000 rpm (150,000g) for 1 hr. The pelletis homogenous, lacking membranous contamination. X 20,100.
Table 1. Summary of Components of Glutaraldehyde-stabilizedFractions Isolated from Sycamore-Maple Cells
Fraction Components
rpm
10,000 12, lOOg Golgi bodiesMitochondriaSmooth surfaced membranes
20,000 43,000g Few Golgi bodiesSmooth surfaced microsomesRough surfaced microsomes
37,000 150,000g Free and membrane-bound ribosomesSmooth surfaced microsomes
Table II. Summary of Turnover and Accumulation Rates ofHydroxyproline in Wall and Organelle Fractions
Fraction Designation Transport Accumu-lation
rpm pg hydroxyprolinel
10,000 12, 100g Crude Golgi-smooth 0.12membrane
20,000 43,000g Microsomal 0.0337,000 150,OOOg Ribosomal 0.04
800g Wall 0.1
intact. Sonication for 15 to 30 sec ruptured only 25% of the cellsbut the number of intact Golgi bodies was more than doubled.It should be noted however that the proline pulse-chase kineticswere not dependent on optimal preservation of the Golgi bodies.The use of glutaraldehyde allowed positive identification of theGolgi fractions. Except where mentioned, cells were sonicatedfor 90 sec to ensure maximal cell breakage and optimal yield ofmaterial.
Pulse-chase Experiment 1, Changes in Specific Radioactivity ofHydroxyproline as a Function of Time in Cell Fractions. To tracethe movement of hydroxyproline-rich material from its place offormation to its site of deposition, we followed the time-depend-
ent changes in specific radioactivity of hydroxyproline in subcellu-lar fractions. Cells were pulsed with labeled proline and thenchased with unlabeled proline at 15 min. Samples were withdrawnand cell fractions were obtained as in "Materials and Methods."Figure 4 summarizes the changes in specific radioactivity of hy-droxyproline in the glutaraldehyde-stabilized cell fractions. Thespecific radioactivities of hydroxyproline in the 20,000 (micro-some) and 37,000 (ribosome) rpm pellets decrease on addition ofunlabeled proline. In contrast, the specific radioactivity of hy-droxyproline in the 10,000 (Golgi smooth membrane) rpm pelletcontinues to rise for 5 min after the addition of 40 mm prolineand then falls. The hydroxyproline turnover (expressed as ,ghypro per hr per mg dry wt cells) in each of these fractions wascalculated from the half-life of labeled hydroxyproline (derivedfrom the changes in specific radioactivity and the total amountof hydroxyproline in each fraction). The 37,000 rpm super-natant contained insufficient trichloroacetic acid-precipitableradioactive hydroxyproline for analysis.
Electron Microscopy of Cell Fractions Resulting from Differen-
Table III. Distribution of Radioactive Hydroxyproline in ThreeLayers Resulting from Discontinuous Sucrose Gradient
Purification of Golgi BodiesA 100-ml cell subculture was labeled with 20,c of 14C-proline;
cells were ruptured by sonication; walls were separated fromcytoplasm by centrifugation (2 min at 500g); cytoplasm wascentrifuged at 7,000 rpm (6,040g) for 30 min and the supernatantwas processed as in Figure 2 to yield three bands corresponding to1.25, 1.5, and 1.6 to 1.8 M sucrose; organelles of three bands werehydrolyzed with HCl; radioactive hydroxyproline and prolinewas determined as in Figure 4.
Sucrose ConcnTime
Top Middle Bottom
min 1.25 M 1.5 H 1.6-1.8 M
10 10,0601 356 18315 18,565 723 60320 22,370 776 613
l cpm in hydroxyproline.
Plant Physiol. Vol. 46, 1970 835
www.plantphysiol.orgon April 23, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 46, 1970DASHEK
FIG. 8. Electron micrograph of a section through top band of organelles (1.25 M sucrose) obtained by discontinuous sucrose gradient cen-trifugation methods. The fraction is enriched with respect to Golgi bodies, many of which consist of stacked lamellae with associated vesicles.X 35,000.
Table IV. Comparison of Specific Radioactivity of Hydroxyprolinefor 7,000 rpm and Enriched Golgi Fractions
Specific Radioactivity of HydroxyprolineTime
7,000 rpm Pellet Enriched Golgi
min cpm in hydroxyproline/Ag of hydroxyproline
5 6,600 7,24310 9,500 5,70015 16,30020 17,900 10,90225 15,951 4,69430 4,25340 9,287 3,70060 6,819 2,170
tial Centrifugation. The major constituents of the 10,000, 20,000,and 37,000 rpm fractions are shown in Figure 5, 6, and 7, andsummarized in Table I.
Pulse-chase Experiment 2, Comparison of Changes in Turnoverof Hydroxyproline as a Function of Time in Crude and EnrichedGolgi Fractions. The turnover data obtained from pulse-chaseexperiment 1 (Table II) showed the Golgi smooth membranefraction as the chief vehicle of hydroxyproline transport (enoughhydroxyproline moves through this fraction to account for thataccumulating in the wall over the same period of time). Furtherpurification of a Golgi smooth membrane fraction was obtainedby taking a 7,000 rpm supernatant (rather than the 10,000 rpmpellet in pulse-chase experiment 1) and subjecting it to a discon-tinuous sucrose gradient centrifugation (Fig. 2 [13]). This gave
three discrete layers (top, middle, and bottom) with greater than95%0 of the hydroxyproline in the top layer (Table III). Electronmicroscopy (Fig. 8) showed that the top layer was enriched withGolgi bodies often structurally intact but still apparently contam-inated by smooth membranous material. (The organelle contentof the middle and bottom layers was not examined because theycontained very little hydroxyproline). However, the hydroxypro-line turnover rate in this purified fraction was much lower (0.04)than in the crude Golgi (0.09) fraction (10,000 rpm pellet, experi-ment 1), indicating that purification had removed a hydroxypro-line component from the crude Golgi fraction.
Pulse-chase Experiment 3, Comparison of Changes in SpecificRadioactivity of Hydroxyproline in an Enriched Golgi Fractionand a Smooth Membrane Fraction (7,000 rpm pellet). The specificradioactivity of hydroxyproline in the 7,000 rpm pellet removedby differential centrifugation from those sonicates which weresubjected to sucrose gradient centrifugation was much higherthan that of the enriched Golgi fraction (Table IV). Electronmicroscopic examination of the 7,000 rpm pellet showed that itconsisted mainly of smooth membranes together with mito-chondria but was essentially devoid of Golgi bodies (Fig. 9).
Pulse-chase Experiment 4, Changes in Specific Radioactivity ofHydroxyproline as a Function of Time in Cell Fractions Isolatedin the Absence of Glutaraldehyde. To rule out the possibility thatglutaraldehyde might cause adsorption of soluble proteins ontoisolated organelles, a single pulse-chase experiment was carriedout in which the aldehyde was deleted from the isolation medium.Table V shows that in the absence of glutaraldehyde (a) the bulkof cytoplasmic l4C-hydroxyproline is organelle-bound, and (b)the specific radioactivities of hydroxyproline in the 10,000 and20,000 rpm pellets (smooth membranes mixtures of disrupted
836
www.plantphysiol.orgon April 23, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.

HYDROXYPROLINE-RICH COMPONENTS
FIG. 9. Electron micrograph of a section through an aldehyde-stabilized 7,000 rpm pellet. Apparent are some mitochondria and an abundantbackground of smooth surfaced membranes. Upon careful examination some of these membranes appear to be vesiculate. X 27,500.
Table V. Changes in Specific Radioactivity of Hydroxyproline as aFunction of Time in Cytoplasmic and Wall Fractions
Isolated without Glutaraldehyde Stabilization
Experimental details as in Figure 4 except that a 100-ml cellculture was exposed to 10 ,c of 14C-proline, and organelles weresonicated into homogenization medium lacking glutaraldehyde.
Fraction (rpm X 1000)Time Wall
10 20 37 37
min 12,000g 43,000g 1SO,OOOg supernatant5 23541 1961 8666 233 16
15 10580 11160 23450 591 4825 13267 14328 11783 654 9240 8702 9236 8416 630 33960 4010 2196 3380 631 404
The cpm in hydroxyproline/,ug hydroxyproline.
organelles) decrease later than that of the 37,000 rpm (ribosomal)fraction.
DISCUSSION
Limitations of Techniques Employed. Glutaraldehyde, a com-monly used fixative in electron microscopy, was incorporated intothe homogenization medium because it stabilizes Golgi bodies(13) during isolation so that they could be easily identified. How-ever, glutaraldehyde can cause adsorption of soluble proteins ontoorganelles. In addition, the kinetics reported herein could be arti-facts reflecting differential rates for glutaraldehyde-induced inac-
tivation of the metabolism of a hydroxyproline-containingcomponent(s) in each cell fraction. To test this possibility, a 14Cjproline pulse-chase experiment was carried out in which organ-elles were isolated without the aid of aldehyde stabilization. Thefindings (see "Results," pulse-chase experiment 4) revealed littlesoluble hydroxyproline (the bulk of cytoplasmic hydroxyprolinebeing organelle bound), indicating that glutaraldehyde does notlead to the soluble hydroxyproline being bound or trapped by theorganelles. Also, the observation that smooth membrane fractions(10,000 and 20,000 rpm) chase later than a ribosomal fraction inthe absence of aldehyde stabilization argues against the view thatthe kinetics for hydroxyproline-containing component(s) in alde-hyde-stabilized fractions are glutaraldehyde-induced artifacts.
Table VI. Organelle Content of Three Fractions Implicated inTransport of a Wall Destined Hydroxyproline-containing
Component
Fraction (rpm or position Preparative Procedure Organelles Present
7,000 rpm (6,040g) Differential centri- Smooth surfacedfugation membranes,
mitochondria10,000 rpm (12,000g) Differential centri- Smooth surfaced
fugation membranes, fewGolgi bodies,mitochondria
1.25 M Sucrose Sucrose gradient Smooth surfacedcentrifugation membranes,
many Golgibodies
Plant Physiol. Vol. 46, 1970 837
www.plantphysiol.orgon April 23, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.

8Plant Physiol. Vol. 46, 1970
Unfortunately, by electron microscopy, recognizable Golgi bodieswere not obtained when glutaraldehyde was deleted from thehomogenization medium. Recently, Ray and his coworkers (15)reported isolation of Golgi bodies from mung bean without priorglutaraldehyde stabilization. These Golgi bodies possessedf,-glucan synthetase activity and were isolated by coupled velocity/isopycnic gradient centrifugations following homogenization pro-cedures involving minimal stress. Thus, to provide additionalproof that the findings reported herein are not aldehyde-producedartifacts, pulse-chase experiments utilizing the improved methodsof Ray et al. (15) for Golgi body isolation will be carried out.Although our present techniques do not rule out the possibility
of several hydroxyproline-rich components, the data presentedhere do show that the bulk of the hydroxyproline is on its way tothe cell wall. Evidence that organellar-bound hydroxyproline isin a protein can be summarized as follows: (a) the rapid appear-ance of high specific radioactivity hydroxyproline in ribosomalpreparations during pulse-chase experiments (e.g. experiment 1),(b) cycloheximide, a potent inhibitor of protein synthesis at theribosomal level (20) inhibits hydroxyproline formation in syca-more-maple cells (5), (c) proteases release both high and lowmolecular weight hydroxyproline-containing glycopeptides fromcell walls (10), (d) examination of the amino acid composition ofpurified cell walls shows a wide range of amino acids (9), and (e)the hydroxyproline-containing component(s) of cytoplasmic or-ganelles can be solubilized as high molecular weight material.
Interpretation of Results. The results presented show that threefractions are involved in transport, i.e., they possess a hydroxy-proline-containing component(s) whose specific radioactivity de-clines later than that of the ribosomal fraction. Table VI lists theorganelles which characterize these fractions and demonstratesthat the only component common to each is a smooth membra-nous component. These results therefore point to the involvementof a smooth membranous component rather than the Golgi bodiesin the transport of hydroxyproline-containing components. Thisis consistent with the recent report (2) that wall bound hydroxy-proline of phloem parenchyma cells derived from carrot disks alsoappears to be secreted by membraneous organelles.The identity of the smooth surfaced element(s) remains an ex-
citing but unanswered question, although it may simply be smoothendoplasmic reticulum.
Villemez and coworkers (22) recently isolated a particulatefraction from Phaseolus aureus shoots at velocities comparable tothose reported here and found that these particles could incorpo-rate nucleotide sugars. The particles of Villemez et al. (22) andthe smooth membranous component reported here have similarsedimentation rates and could be identical. Thus, the nucleotide-mediated sugar incorporation observed by Villemez et al. mightat least in part be the glycosylation of a hydroxyproline-contain-ing component(s). In this connection, it is of interest that glyco-sylationof variousanimal proteins is mediatednot by Golgi bodiesbut by other smooth surfaced membranes (3, 21).
Acknowledgements-I am indebted to Drs. Derek T. A. Lamport and Anton Langfor reading the manuscript and to Dr. Lamport also for advice and encouragementduring a post-doctoral stay in his laboratory. Grateful appreciation is extended toDr. J. M. Holleman for supplying cell cultures, and Dr. G. Spink for the use of aPhillips 100 electron microscope. I am alsograteful to Miss Ruth Schmnitter who didthe major portion of the electron microscopy reported here. It is a pleasure to acknowl-edge the skilled assistance of Mrs. Linda Franzen.
REFERENCES CITED
1.CHINARD, F. P. 1952. Photometric estimation of proline and ornithine. J. Biol.Chem. 199: 91-95.
2. CHRISPEELS, M. J. 1969. Synthesis and secretion of hydroxyproline containingmacromolecules in carrots.I. Kinetic analysis. Plant Physiol. 44: 1187-1193.
3. COOK, G. M.W., M. T. LAIco, AND E. H.EYLAR. 1965. Biosynthesis of glyco-proteins of the Ehrlich ascites carcinona cell membranes. Proc. Nat. Acad.Sci. U.S.A. 54: 247-252.
4. DASHEK,W. V. ANDW. G. ROSEN. 1966. Electron microscopical localization ofchemical components in the growth zone of lily pollen tubes. Protoplasma. 61:192-204.
5. HOLLEMAN, J. M. 1967. Incorporation of hydroxyproline into protein of sycamorecells incubated at growth-inhibitory levels of hydroxyproline. Proc. Nat.Acad. Sci. U.S.A. 57: 50-54.
6. HOLLEMAN,J. M. AND J. L. Key. 1967. Inactive and protein precursor pools ofamino acids in the soybean hypocotyl. Plant Physiol. 42: 29-36.
7.KsviRiKKo, K.I. AND M. LIESMA. 1959. A colorimetric method for determinationof hydroxyproline in tissue hydrolysates. Scand. J. Clin. Lab. Invest. 11: 128-133.
8. LAMPORT. D. T. A. 1964. Cell suspension cultures of higher plants: isolation andgrowth energetics. Exp. Cell Res. 33: 195-206.
9. LAMPORT, D. T. A. 1965. The protein component of primary cell walls: In: R. D.Preston ed., Advances in Botanical Research 2:151-218.
10. LAMPORT, D. T. A. 1969. The isolation and partial characterization of hydroxy-proline-rich glycopeptides obtained by enzymic degradation of primary cellwalls. Biochemistry 8: 1155-1169.
11. MOLLENHAUER, H. H.,W. G.WHALEY, AND J. H. LEECH. 1961. A function ofthe Golgi apparatus in outer root cap cells.J. Ultrastruct. Res. 5:193-200.
12. MORRE, D.J. AND H. H. MOLLENHAUER. 1964. Isolation of the Golgi apparatusfrom plant cells.J. Cell. Biol. 23: 295-305.
13. MORRE, D. J., H. H. MOLLENHAUER, ANDJ. E. CHAMBERS. 1965. Glutaraldehydestabilization as an aid to Golgi apparatus isolation. Exp. Cell. Res. 38: 672-675.
14. OLSON, A. C. 1964. Proteins and plant cell walls. Proline to hydroxyproline intobacco suspension cultures. Plant Physiol. 39: 543-554.
15. RAY, P. M., T. L. SHININGER, AND M. M. RAY. 1969. Isolation of,-glucan synthe-tase particles from plant cells and identification with Golgi membranes. Proc.Nat. Acad. Sci. U.S.A. 64: 605-612.
16. REYNOLDS, E. S. 1963. The use of lead citrate at high pH as an electron opaquestain in electron microscopy. J. Cell Biol. 17: 208-212.
17. ROSEN,W. G. AND S. R. GAWLIK. 1966. Fine structure of lily pollen tubes followingvarious fixation and staining procedures. Protoplasma 61:181-191.
18. SABATINI, D., K. BENSCH, AND R. J. BARNETr. 1963. Cytochemistry and electronmicroscopy. J. Cell Biol. 17: 19-58.
19. SHEEHAN, J. C. AND J. G. WHITNEY. 1963. The synthesis of cis- and trans-3-hydroxyl-L-proline. Two new amino acids from the antibiotic telomycin. J.Amer. Chem. Soc. 85: 3863-3865.
20. SIEGEL, M. AND H. SISLER. 1965. Site of action of cycloheximide in cells of Saccharo-myces pastorianus. M. Further studies on the mechanism of resistance and themechanism of action of Saccharomyces species. Biochim. Biophys. Acta 103:558-567.
21. SIMKIN, J. L. AND J. C. JAMIESON. 1967. Studies on the nature of microsome-boundsubstances involved in the biosynthesis of acidic glycoproteins of guinea-pigserum. Biochem. J. 103: 38P-39P.
22. ViLLEMEZ, C. L., J. M. MCNAB, AND P. ALBERSHEIM. 1968. Formation of plantcell wall polysaccharides. Nature 218: 878-880.
838 DASHEK
www.plantphysiol.orgon April 23, 2020 - Published by Downloaded from Copyright © 1970 American Society of Plant Biologists. All rights reserved.





























