Embed Size (px)
Citation preview

DOI: 10.1161/CIRCGENETICS.115.001217
1
Synthesis of an Endogenous Steroidal Na Pump Inhibitor Marinobufagenin,
Implicated in Human Cardiovascular Diseases, Is Initiated by CYP27A1
via Bile Acid Pathway
Running title: Fedorova et al.; Mammalian marinobufagenin starts from bile acids
Olga V. Fedorova, PhD; Valentina I. Zernetkina, MD; Victoria Y. Shilova, PhD;
Yulia N. Grigorova, MD; Ondrej Juhasz, PhD; Wen Wei, PhD; Courtney A. Marshall, BS;
Edward G. Lakatta, MD; Alexei Y. Bagrov, MD, PhD
Laboratory of Cardiovascular Science, National Institute on Aging, NIH, Baltimore, MD
Correspondence:
Alexei Y. Bagrov, MD, PhD Olga Fedorova, PhD
Laboratory of Cardiovascular Science Laboratory of Cardiovascular Science
National Institute on Aging, NIH National Institute on Aging
Biomedical Research Center Biomedical Research Center
251 Bayview Blvd. 251 Bayview Blvd.
Baltimore, MD 21224-6825 Baltimore, MD 21224-6825
Tel: 410-558-8209 Tel: 410-558-8022
Fax: 410-558-8150 Fax: 410-558-8150
E-mail: [email protected] E-mail: [email protected]
Journal Subject Terms: Hypertension; Etiology
Yulia N. Grigorova, MD; Ondrej Juhasz, PhD; Wen Wei, PhD; Courtney A. Marararshshalalll, , , BSBS; ; ;
Edward G. Lakatta, MD; Alexei Y. Bagrov, MD, PhD
Laboratory oooff f Carrrdiioiovascscscuuular SSSccciennnccce, NaNaNationnnalll Insnssttitituuuttete on AgAgAging, NNNIHHH, BBBaltimimmoorore,, MMMD
Correspondence:
AlAlexexeiei YY BaBagrgrovov MDMD PhPhDD OlOlgaga FFededororovovaa PhPhDD
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
2
Abstract:
Background - The bioactive steroid, marinobufagenin (MBG), is an endogenous Na/K-ATPase
bufadienolide inhibitor that is synthetized by adrenocortical and placental cells. MBG binding to
Na/K-ATPase initiates pro-fibrotic cell signaling, and heightened MBG levels are implicated in
the pathogenesis of hypertension, preeclampsia and chronic kidney disease. Steroids are derived
from cholesterol through the “traditional” steroidogenesis pathway initiated by enzyme
CYP11A1, and via the “acidic” bile acid pathway, which is controlled by enzyme CYP27A1.
The mechanism of MBG biosynthesis in mammals however remains unknown.
Methods and Results - Here we show that post-transcriptional silencing of the CYP27A1 gene in
human trophoblast and rat adrenocortical cells reduced the expression of CYP27A1 mRNA by
70%, reduced total bile acids 2-fold, and MBG levels by 67%, compared to non-treated cells or
cells transfected with non-targeting siRNA. In contrast, silencing of the CYP11A1 gene did not
affect MBG production in either cell culture, but suppressed production of progesterone 2-fold in
human trophoblast cells, and of corticosterone by 90% in rat adrenocortical cells, compared to
cells transfected with non-targeting siRNA. In vivo, in a high salt administration experiment,
male and female Dahl-S rats became hypertensive after 4 weeks on a high NaCl diet, their
plasma MBG levels doubled, and adrenocortical CYP27A1 mRNA and protein increased 1.6-
fold and 2.0-fold.
Conclusions - Therefore, the endogenous steroidal Na/K-ATPase inhibitor, MBG, is synthesized
in mammalian placenta and adrenal cortex from cholesterol through the novel “acidic” bile acid
pathway. These findings will help to understand the role of MBG in highly prevalent human
cardiovascular diseases.
Key words: cholesterol; enzymes; gene; hormones; molecular biology; biosynthesis of steroids; natriuretic hormone; Na/K-ATPase inhibitors; CYP27A1; salt-sensitive hypertension
, , y , p
cells transfected with non-targeting siRNA. In contrast, silencing of the CYP11AA111 gggennne e e dididid d d nnon tt
affect MBG production in either cell culture, but suppressed production of progesterone 2-fold in
huuumamamannn trt oppphohohobllasasasttt cells, and of corticosterone by 909090% in rat adrennnocoo ortitiiccacal cells, compared to
ccecelllls transfecteeed dd wwwithhh nnnononon---tatatargrgrgetetetining g g sisis RNRNRNA.A IIInn vivooo, inn aa hhigggh h h saltltlt aaaddmdminininissstratattioioionn exexxpepepeririimemementntnt,
mmamalelele and femalee DDDahl---SSS rats bbbecece ameee hyyyppperteenensssive affterrr 444 weeeeeeks ooonn n a highghgh NNaCaCaCl diettt, tttheeeirr r
plasmamaa MMMBBBG levev llelss dodd bbubllel d, aandndnd adrenococcorortitiicac l CYCYCYPP2P27A7A7A111 mRmRRNANAA aa dndd pprorottein iiinncncrer assededd 111.666-
fold and 2.0-fold.
CCConclllu iisions - ThThTher ffefore, ttthhhe e ddndogenous ttster iioidddalll NNNa/K/K/K AA-ATPTPTPase iiinhihihibibibitttor, MMMBGBGBG, iiis synthththesiiiz dded
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
3
Introduction
The bioactive steroid, marinobufagenin (MBG) (Figure 1A), is an endogenous bufadienolide
Na/K-ATPase ligand and inhibitor.1-5 Recent studies demonstrate that increased levels of
endogenous Na/K-ATPase ligands including MBG are implicated in the pathogenesis of chronic
kidney disease, essential hypertension and preeclampsia.1-10 Thus, hypertensive Dahl salt-
sensitive rats (Dahl-S) have an increased plasma and adrenocortical levels of MBG, which was
accompanied by inhibition of Na/K-ATPase and blood pressure elevation.1,11 Angiotensin II
stimulates MBG production in adrenal cortex of Dahl-S rats in vitro and in vivo.12 Previously it
was demonstrated that adrenalectomy in rats resulted in the reduction of plasma digoxin-like
immunoreactivity13 and MBG levels,14 which indicated that adrenals are the major source of
MBG and other cardiotonic steroids in mammals.
Increasing evidence indicates that, in addition to inhibition of Na/K-ATPase, MBG is
capable to initiates potent cell signaling via binding to Na/K-ATPase.5,15 MBG via modulation of
Na/K-ATPase activity not only participates in blood pressure regulation, but also induces cardiac
and vascular fibrosis,10 a hallmark of cardiovascular aging, resistant hypertension, and chronic
kidney disease.16,17 Thus, administration of an anti-MBG monoclonal antibody in salt-sensitive
hypertension models lowers blood pressure,9 reverses preeclampsia-induced Na/K-ATPase
inhibition,9 and potently reverses cardiac fibrosis in uremic rats,2,6 states where the MBG levels
are increased.
Despite the fact that bufadienolide sodium pump ligands are emerging as potentially
important therapeutic targets, mechanisms of their biosynthesis are not understood, and the
paucity of knowledge regarding the biosynthesis of the steroidal Na/K-ATPase inhibitor, MBG,
has hampered the research on therapeutic targeting of MBG-driven Na/K-ATPase regulatory
mmunoreactivity13 and MBG levels,14 which indicated that adrenals are the majooor r soss urururcecece ooof f f
MBG anand d otheh r cacardiotonic steroids in mammals.
Increaaasiinnnggg evvidididenenencecece indddicicicatatatesese ttthahahat,t iiin n n adada ditiiooon tooo inininhihihibibb tiiononon ooof f f NaNaNa/K/K/K-ATATTPaPaPasesese, , , MBMBMBG G G isisis
cacaapapapablb e to initiatttesss potttennnt cellll sssignalililingngg vvvia bbbinnndinnng to NaNaNa K/K/K-AAATPPaPases .5,1555 MMMBGBGBG via mmmoooduuulaaationnn oof
Na/K-ATPase activiviitytyty nnot onlnlly y y papap rticippates in blooooood d pressusuurerere regegulattiooon,n,n, but also induces cardiac
anand d vavascsculularar ffibibrorosisis,s,,101010 aa hhalallmlmarark k ofof ccarardidiovovasascuculalar r agaggining,g,g, rresesisistatantnt hhypypyperertetensnsioion,n,, aandnd cchrhrononicic
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
4
signaling. Steroids are derived from cholesterol, and the “traditional” biosynthesis of biologically
active steroids begins via the side-chain cleavage of cholesterol by the cytochrome P450 enzyme
CYP11A1 and conversion into pregnenolone18,19 (Figure 2), although this pathway is not
involved in MBG biosynthesis.20
The biosynthesis pathways, other than “traditional” steroidogenesis, which also generate
physiologically active steroids, include “classical” and “acidic” bile acid pathways21-23 (Figure
2), in which cholesterol is cleaved, respectively, by CYP7A1, an enzyme, expressed only in the
liver,24 and by CYP27A1, an enzyme, expressed in the “extra-hepatic” tissues, including adrenal
glands.22,25-27 Because MBG is synthesized in non-hepatic tissues,1,4,12 and CYP7A1 is expressed
only in the liver,24 we hypothesized that mammalian MBG is a bioactive bile acid derivative
steroid, synthesized via the “acidic” extra-hepatic pathway that oxidizes cholesterol into bile
acids by CYP27A1 enzyme. To test this hypothesis, we studied (i) the levels of MBG, produced
by human choriocarcinoma JEG-3 cells; (ii) the role of post-transcriptional CYP27A1 gene
silencing on MBG production in human trophoblast and primary culture of rat adrenocortical
cells; (iii) the possible participation of adrenocortical CYP27A1 in MBG production in the
model of salt-sensitive hypertension in Dahl-S rats.
Materials and Methods
High sodium chloride diet in Dahl salt-sensitive model (animal study)
Dahl-S rats (5 weeks old, both genders; Charles River, Frederick, MD) were used for dietary
high NaCl administration. This experiment was approved by Institutional Animal Care and Use
Committee (IACUC). The rats were maintained in a 26°C environment with a 12:12 hour light-
dark cycle on a low salt diet (0.3% NaCl, Harlan Teklad, Madison, WI) and tap water ab libitum
for an adaptation for 1 week. Six animals of each gender were then placed on a high NaCl diet
only in the liver,24 we hypothesized that mammalian MBG is a bioactive bile aciddd dededeririivavavatititiveveve
teroid,d,, synyny thhese izzede via the “acidic” extra-hepatic papap thway that oxidizes chc olesterol into bile
acacciddds by CYPPP27277A1AA eenznznzymymyme.e.e TTooo teteteststst tthihihisss hyhyypopopothtt esiiis,, weee ssstututudididiede (((iii) tttheheh lllevevevelee s ofofof MMMBGBGBG, prprprodododucucucedede
bybyy hhhumu an chorioococaarcinononoma JJJEGEGEG-3 ceceellsss; (ii) thhhe roooleee of f f popopost--trrranssscrrriptionnnaaal CCCYYYP27A1A1A1 genenene
ilencing on MBG ppprororoduduction n n ininin human trophp oblalaastst and pppriririmamam ryy cultutuurerere oof rat adrenocortical
cecelllls;s;; (((iiiii)i)) tthehe ppposossisiblble e papap rtrticicipippatatioion n ofof aadrdrenenococorortiticacal l CYCYP2P27A7A11 inin MMBGBG ppproroduductctioion n inin tthehe
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
5
(8% NaCl, Harlan Teklad, Madison, WI). Control animals (males, n=6; and females, n=6) were
maintained on a low salt diet for 4 weeks. Blood pressure was measured in conscious animals by
tail-cuff plethysmography (IITC Life Science, Woodland Hills, CA) at baseline and after 4
weeks of a high NaCl diet. The adrenal glands were collected for the measurement of CYP11A1,
CYP27A1, CYP11B1 and CYP11B2 mRNA expression in adrenal cortex (real-time quantitative
PCR; below), for Western blotting and histochemistry analyses (below). Plasma was extracted
using Sep-Pak C-18 reverse-phase cartridges (Waters, Milford, MA), as reported previously,9 and
used for measurement of MBG (immunoassay; below).
Human placental and rat adrenocortical cell cultures
The human choriocarcinoma cell line JEG-3 was obtained from ATCC (American Type Culture
Collection, Manassas, VA), and maintained in Eagle’s Minimum Essential Medium (EMEM,
ATCC) in the presence of 10% fetal bovine serum (FBS; Life Technologies/Invitrogen, Grand
Technologies/Invitrogen). Cells from passages 2-5 were used in the experiments.
A rat adrenocortical primary cell culture was prepared as described previously.12 Twelve
3-4 months old Dahl salt-sensitive rats (Charles River Laboratories International, Inc.,
Wilmington, MA) were euthanized by an overdose of sodium pentobarbital (100 mg/kg),
adrenals were removed, washed in 0.9% NaCl, the cortex was isolated, minced, and incubated
for 1 hour at 37°C placed in Dulbecco’s modified essential medium/F-12 medium (DMEM/F-12;
Life Technologies/GIBCO, Grand Island, NY) containing 0.5% FBS (Life
Technologies/Invitrogen), 1 mg/ml collagenase type IV (Worthington Biochemical Corp.,
-I from bovine pancreas, Sigma-Aldrich, St.
Louis, MO), and antibiotics penicillin and streptomycin (Life Technologies/Invitrogen). The
The human choriocarcinoma cell line JEG-3 was obtained from ATCC (Americaaan n TyTyTypepepe CCCululultutut re
Collectition,, MaM naasss as, VA), and maintained in Eagagglele’s Minimum Essentitiala Medium (EMEM,
AAATCCCC) in theee ppprereresess ncncceee ofofof 1110%0%% fffetetetalalal bbbovovovinii e ee seseserur mmm (FFFBSSS;;; LiLiLifefefe TTececechnhnhnololo ogogogieieies/Innnvivivitrtrtrogogogenenn, , , GrGrGranananddd
Technologies/Invitroroogegegen)). Ceellllllss s from ppassageg s 2---555 wew re uuusesesed dd in the eexpxpxperee iments.
A A rarat t adadrerenonococortrticicalal pppririmamaryryy ccelelll cucultlturure e wawass prprp epepparareded aass dedescscriribebed d prprp evevioioususlylyy..12 TwTwelelveve
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
6
Inc., Ranch Dominguez, CA), cells were collected, centrifuged at 200 g for 5 min, and cultured
in DMEM/F-12 media with 10% FBS. Media was replaced every 2-3 days. Cell from passages 2-
4 were used for the further analyses.
Concentrations of MBG, bile acids, progesterone or corticosterone produced by JEG-2
and adrenocortical cells were estimated by immunoassays of the extracted media. When the JEG-
3 cells reached the 80-90% confluence, the 10% FBS media was replaced by 2.5% FBS media.
JEG-3 cells were incubated for 0, 3 or 6 hours. The media was collected for MBG measurements
and for purification of MBG-immunoreactive material via high-performance liquid
buffer with protease/phosphatase inhibitors (radio-immuno-precipitation assay lysis buffer, Santa
Cruz Biotechnology, Inc., Santa Cruz, CA), and used for protein measurement and Western blot
analysis (below).
CYP27A1 and CYP11A1 genes silencing
Double stranded siRNAs were used to silence CYP27A1 and CYP11A1 genes. Oligonucleotides
for gene silencing in human cells were obtained from Qiagen, Valencia, CA (four siRNAs for
CYP27A1, catalog numbers SI00015533, SI00015540, SI00015547, SI00015554), and Thermo
Scientific / Dharmacon, (Pittsburg, PA) (siRNA for CYP11A1 ON-TARGETplus SMARTpool
L-008329-00-0005). Oligonucleotides for gene silencing in rat cells were obtained from
Dharmacon (CYP27A1rat ON-TARGETplus SMARTpool, and CYP11A1rat ON-TARGETplus
SMARTpool). ON-TARGETplus Non-targeting pool with no homology to mammalian genes
(Thermo Scientific / Dharmacon; catalog number D-001810-10-20) was used as a negative
bububuffffffer with pprororoteteteasasa e//phphphosososphphphatasasase e e inininhihih bibibitototorsss (((rararadio---immmmumuunonono-p-p-prer cicicippipitatatatitt ononon aaassayayy lllysysysisisis bbufufuffefefer,r,r, SSSanananta
CrCrCruzuzz Biotechnollogggy, IIInccc., Saaantntnta Cruzuzuz, CACACA), aaannnd usssed fofofor r r proototeein memem asurrremmmenennt tt and WeWeW sttterrrn blloot
analysis (below).
CYCYP2P27A7A11 anand d CYCYP1P11A1A1 1 gegeg neness sisilelencncining g g
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
7
control for both JEG-3 and adrenocortical cells.
Prior to silencing experiments JEG-3 and adrenocortical cells were cultured in 6-well
plates 24-48 hours at a density 2x10E5 cells/well in correspondent media with 10% FBS. When
cells reached the 60-65% confluence the transfection was performed with 100 nM siRNA in the
transfection reagent Gene Silencer (Genlantis, San Diego, CA) in 1
ml/well of culture media without FBS; 6 hours later an equal volume of correspondent media
with FBS was added to the final concentration 10% FBS.
Effects of gene silencing were assessed by real-time quantitative polymerase chain
reaction (qPCR) and western blot analyses. For qPCR, cells were cultured for 24 hours after
silencing, and for western blot cells were cultured for an additional 48 hours. Media for MBG
and other steroid measurements was collected 72 hours following silencing, during which the
10% FBS media was replaced by 2.5% FBS correspondent media, cells were incubated for 6
hours, culture media and cells were collected as above (Cell culture).
Real-time quantitative PCR
Real-time quantitative analysis of CYP11A1, CYP11B1, CYP11B2 and CYP27A1 mRNA levels
was performed by PCR amplification of the resulting cDNAs and normalized to expression of
housekeeping genes as the internal standards (rat and human 18S ribosomal mRNA for rat
adrenocortical cells and tissue and for human JEG-3 cells). In detail, total RNA was extracted
from JEG-3 cells and from adrenocortical cells or adrenocortical tissue, which were collected 24
hours after the transfection (Qiagen RNeasy mini-kit in the presence of DNase-I; Qiagen). Total
RNA samples were then reverse transcribed to cDNA using TaqMan Reverse transcription kit
(Life Technologies /Applied Biosystems, Grand Island, NY). Primer sets for real-time PCR for
rat CYP11A1, CYP11B1, CYP11B2, and both rat and human CYP27A1 genes, and housekeeping
ilencing, and for western blot cells were cultured for an additional 48 hours. Meeedididia aa fofofor r r MBMBMBGGG
and otheher r sterroid d mem asurements was collected 72 hohours following silencining, during which the
10100%%% FBS meeedididiaa a waww ss rerereplplplacacacede bbby y y 2.2.2.5%5%5% FFFBSSS cccoroo respspspondededentntnt mmmeddiaiaia, cececelllls s s weww re iiincncncubububatata eddd fffororor 666
hohooururrs, culture medededia aaandndd celllls wwwere cocc lllleeecteddd aaas abobobove (((CeCeC llll cccultuuureee).
Real-time quantitatatativivive e PCRRR
ReRealal-titimeme qqquauantntititatativive e ananalalysysy isis oof f CYCYP1P11A1A1,1,, CCYPYP1111B1B1,,, CYCYP1P11B1B22 aandnd CYCYP2P27A7A11 mRmRNANA llevevelelss
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
8
rat and human 18S ribosomal RNA genes were obtained from Qiagen (Table 1). Primers for
human CYP11A1 were designed using Primer-BLAST tool designed by NCBI
(http://www.ncbi.nlm.nih.gov/tools/primer-blast/primerinfo.html): Primer-BLAST is based on
Primer 3 software, and specificity of primers is confirmed using NCBI Blast database.
Quantitative real-time PCR was performed using QuantiFast SYBR Green PCR kit (Qiagen)
according to the manufacture’s protocol, and ABI 7300 Real Time PCR System (Life
Technologies/Applied Biosystems). For each sample, gene expression was analyzed using the
following protocol: activation at 95°C (8 min) followed by 40 cycles, consisting of a first phase
of denaturation at 95°C (10 sec), and a second phase of annealing/extending at 60°C (30 sec).
Each reaction was performed in triplicate with an inclusion of no-template controls in each
experiment. A dissociation curve analysis was performed in each experiment to eliminate non-
specific amplification, including primer dimers. The 18S Ct values were subtracted from the raw
sample Ct values to obtain the corrected Ct. Power conversion (power (2-(correctedCt)) was used to
convert corrected Ct to relative RNA quantity.
Immunoassays
72 hours following transfection with siRNA, both cell cultures were used to study the effect of
CYP27A1 and CYP11A1 silencing on production of MBG, total bile acids (TBA) and
progesterone (JEG-3 cells), and corticosterone (adrenocortical cells). Culture media samples
were collected as described above (“MBG production”), and extracted using Sep-Pak C-18
reverse-phase cartridges (Waters), as reported previously.9 MBG concentration was measured
using a DELFIA immunoassay kit based on 4G4 monoclonal anti-MBG antibody.9 Cross-
immunoreactivity of this antibody is (%): MBG – 100; marinobufotoxin – 43; cinobufotalin – 40;
telocinobufagin – 14; resibufagenin – 0.5; bufalin – 0.08; cinobufagin – 0.07; digoxin – 0.03;
Each reaction was performed in triplicate with an inclusion of no-template controololsss ininin eeeacacach h h
experimement. A A diissssociation curve analysis was perfrforo med in each experimem nt to eliminate non-
pppecccific amplililififificacacatitt ononn,, inininclclcluduu innng g g prprprimimimererer dddimmmererers.s Thehehe 188S S S CtCtCt vavv luuueees wwwerereee sususubtraraactctctededed fffrorom m m thththee e rararaw w
aaampmpmple Ct valuesss ttto obbbtaaain thhhe e coc rreccctetedd t Ct. PoPoPowerrr ccconvnvnveeersiionnn (poowwwer (222-((corrrrectcttedCt)) wawawas usssed tttoo
convert corrected CtCtt tototo relativvveee RNRNR A ququantityyy.
mmmumunonoasassasaysysy
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
9
ouabain – 0.005; ouabagenin – 0.001; digoxigenin – 0.004; proscillaridin A, digitoxin,
aldosterone, progesterone, prednisone, corticosterone, and thyroglobulin - <0.001.
The concentration of progesterone and TBA in the cell culture media was measured after
C18 extraction using a Progesterone DELFIA immunoassay kit (Perkin Elmer, Waltham, MA),
and TBA colorimetric enzymatic assay kit (Bio-Quant, Inc., San Diego, CA). Adrenocortical cell
corticosterone production was measured in the media by an enzyme-linked immunosorbent assay
(ELISA; Cayman Chemical Company, Ann Arbor, MI). The total amount of steroids per sample
was normalized to the total amount of cell protein per sample.
High Performance Liquid Chromatography (HPLC)
For the time course of MBG production, 6 ml of media conditioned by JEG-3 cells for 0
(baseline), 3 or 6 hours were extracted by 80% acetonitrile using Sep-Pak C-18 reverse-phase
cartridges (Waters),9 and the resultant extract was fractionated on an Agilent 1100 series HPLC
system using Agilent Zorbax Eclipse XDB-C18 (Agilent Technologies, Palo Alto, CA), 4.6 x
- 85.5 %) gradient of
acetonitrile against 0.1% trifluoroacetic acid (TFA) for 45 min.9 Thirty 1.5-min fractions were
collected and analyzed for MBG-immunoreactivity (MBG immunoassay, above).
Western blotting
Cell or adrenocortical tissue lysates in RIPA buffer (Santa Cruz Biotechnology) were pretreated
in sample buffer (Life Technologies / Invitrogen) for 5 min at 90°C, and 30 - 40 g protein per
lane was loaded on 10% SDS-PAGE gel (Life Technologies/Invitrogen). After electrophoresis,
proteins were transferred to nitrocellulose membranes (Life Technologies / Invitrogen).
Membranes were blocked for 1 hour with 5% nonfat dry milk in PBS (Phosphate-Buffered
Saline) with 0.1% Tween-20, and subsequently incubated with a 1:250 rabbit polyclonal
For the time course of MBG production, 6 ml of media conditioned by JEG-3 cellllls s fofof rrr 0 0 0
baselinine)e)),,, 3 oro 66 hhours were extracted by 80% aceetotonitrile using Sep-Paak k C-18 reverse-phase
cacaartttrrridges (WaWW tetetersrsrs),9 ananand d d thththee rereesususultltltanana t tt exexextracacactt t waww s frrractiiionononatatatedee ooonnn ananan AAgigigilelelent 1111010100 0 0 seseseriieseses HHHPLPLPLCCC
yyystttememe usingg Agggileent ZZZooorbaxxx EEcEclipse ee XDXDXDB-CCC118 (AgAgAgilenenent t t TeTeTechhhnollologig es, PaPaalooo AAAlto, CCCAAA), 4..6 xx
- 85.5 %) gradient of
acacetetononititririlele aagagag ininstst 00.1.1% % trtrififluluororoaoacecetitic c acacidid (((TFTFA)A)) fforor 4455 mimin.n.999 TThihirtrty y y 1.1.5-5-mimin n frfracactitionons s wewerere
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
10
CYP27A1 antibody (Abcam, Cambridge, MA) or with a 1:500 rabbit polyclonal CYP11A1
antibody (Abcam), following by the incubation with an anti-rabbit-HRP antibody (1:1000, Fisher
Scientific, Pittsburgh, PA) for JEG-3 cells. For rat adrenocortical cells and adrenocortical tissue,
proteins were detected with CYP11A1 and CYP27A1 goat polyclonal antibodies followed by the
secondary anti-goat-HRP antibodies (Santa-Cruz Biotechnology). The gel loading control was
performed by anti-GAPDH mouse antibody (Santa-Cruz Biotechnology). Protein bands were
visualized with Pierce SuperSignal West Pico Chemiluminescent Substrate or with ECL Western
Blotting System (Fisher Scientific), and protein amounts were measured by densitometric
analysis, using Kodak molecular imaging software, Version 5.0 (Molecular Imaging Systems,
Carestream Health, Inc., Rochester, NY).
Immunohistochemistry
Immunohistochemical staining was performed on formalin fixed, paraffin embedded 6-μm
adrenal tissue sections, which were mounted on Superfrost/plus slides (Fischer Scientific,
Pittsburgh, PA), de-paraffinized in xylene and rehydrated through a graded series of ethanol.
Sections were pre-heated to 90°C in antigen unmasking citrate buffer (Thermo Scietific,
Freemont, CA) and slowly cooled down to room temperature. Endogenous peroxidase was
quenched by incubation in 3% hydrogen peroxide in PBS (phosphate buffered saline; Fischer
Scientific) for 10 minutes. After 10 minutes of blocking (10% non-immune serum and 1% BSA
in PBS; 10 min), the slides were incubated overnight at 4°C in humidified chamber with rabbit
anti-CYP27A1 antibody (Abcam; 1:100 in 1% BSA in PBS). Then the slides were incubated
with biotinylated secondary antibody followed by horseradish peroxidase conjugated streptavidin
(LAB-SA detection system; Invitrogen, Camarillo, CA). The immunoreactivity was visualized
with 0.05% 3,3’-diaminobenzidine, followed by 5-min counterstaining with hematoxylin to
Carestream Health, Inc., Rochester, NY).
mmunonohihisttoco hehemim stry
mmmmmmunohistococochehehemimm cacaall l stststaiaiaininin nggg wawawasss pepeperfrfrforoo mememed dd on fooormamaalililin nn fififixex d,d,d, ppparararafafaffififinnn embebebeddddddededed 66--μmμmμm
adaddreeenann l tissue seccttionsss, wwwhichhh wwwere momom ununntedd d ooon Supupuperfrfrfrooost/pplpluuus sslidddes (FFFissschhhererr Sciennntititificc,
Pittsburgh, PA), de--papapararar ffinizzzededed in xyyleene and rehhhydydydrated thththrorr ugugh a grgrgradadadede series of ethanol.
SeSectctioionsns wwerere e prprp ee-hheaeateted d toto 990°0°C C inin aantntigiggenen uunmnmasaskikingngg ccititraratete bbufuffefer r (T(T( hehermrmo o ScScieietitifific,c,,
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
11
visualize nuclei (American Mastertech Scientific, Lodi, CA). The specificity of the
immunostaining was evaluated by omission of the primary antibody and processed as above. A
brown reaction product indicated localization of CYP27A1.
Color images were captured with the ZEISS Axioplan microscope (Thornwood, NY)
using a QCAM FAST 1394 digital camera (QImaging, Surrey, Canada). Quantitative analysis of
CYP27A1 content was performed with MetaMorph Image Analysis Software (Molecular
Devices Corporation, Sunnyvale, CA), and was calculated as the ratio of area stained with
CYP27A1 to total adrenal cortex area. Total 4-5 slides of adrenocortical tissue from each of 6
rats per group were analyzed.
Statistics
The results are presented as mean ± SEM (standard error of the means) or mean ± standard
deviation (SD), as specified in the table and figure legends. Shapiro-Wilks normality tests
(GraphPad Prism Software, San Diego, CA) were conducted for each sample and for each
variable. Practically all of the samples passed the normality test (a=0.05). Next, the Bartlett’s test
for equal variances detected that the variances did not differ significantly among the groups.
Because our data were consistent with normal distributions with constant variance, the
parametric ANOVA was applied for data analyses: one-way analysis of variance (ANOVA)
followed by Bonferroni or Newman-Keuls tests (intra-group analysis) or repeated measures
ANOVA followed by Newman-Keuls test (inter-group analysis) as specified in the table and
figure legends (GraphPad Prism Software). A two-sided P value of less than 0.05 was considered
to be statistically significant.
Results
Because substantial quantities of bufadienolides are synthesized in the placenta,4 we first studied
Statistics
The resusultl s arre prprp ese ented as mean ± SEM (standardrd error of the means) oor r mean ± standard
dededeviviviation (SD)D)D , , , asasas spepepecicicifififiededed in n thththeee tatatablblle e e anaa d dd fififigugg reee leeegenenndsdsds.. ShSS apppiiiro-o-o-WiWiWilklklksss noormrmrmalalalititity y y teeestststsss
GGGraaaphpp Pad Prismmm SSSofttwware, SaSaSan n Diegegego,,, CCCA) wwwere cooondududucctcteddd fffor eeeaacach sampmm leee aaandn fooror eeeaccchhh
variable. Practically y y alalalll ofo theee sssamama plp es ppassed the e nonon rmalittty y y tett sts ((a=0.0 050505).)) Next, the Bartlett’s test
fofor r eqeqquaual l vavaririanancecess dedetetectcteded tthahat t ththe e vavaririanancecess didid d nonot t didifffferer ssigiggninifificacantntlylyy aamomongngg tthehe gggroroupupps.s.
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
12
MBG production by human trophoblast JEG-3 cells. Figure 1B shows that the level of MBG-
immunoreactivity, detected by a 4G4 anti-MBG monoclonal antibody in the conditioned media
of JEG-3 cells, increased exponentially during 6 hours of observation. Figure 1C,D shows that
following HPLC-fractionation of C18-extracted media conditioned by JEG-3 cells, the maximum
of MBG-immunoreactivity co-eluted with MBG standard in a single peak at 24 minutes (fraction
16).
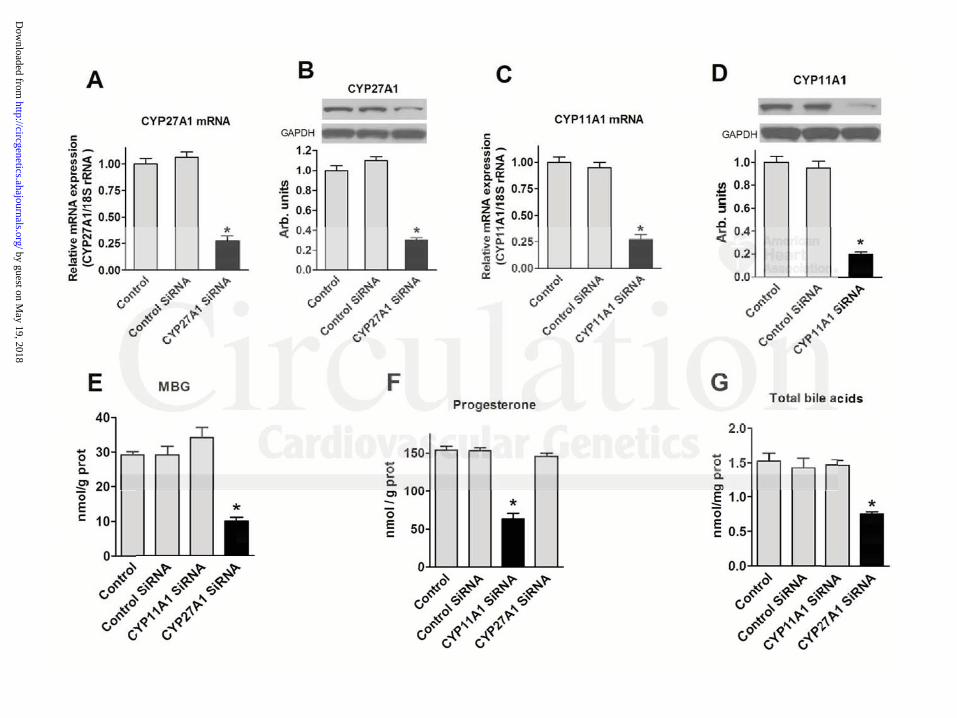
Figure 3 summarizes the results of experiments in which CYP27A1 and CYP11A1 genes
in JEG-3 cells were silenced. Following transfection of a CYP27A1-specific small interfering
RNA (RNAi), expression of CYP27A1 mRNA was reduced by 76% (Figure 3A). Silencing of
CYP27A1 gene reduced the level of CYP27A1 protein by 70% (Figure 3B), induced more than a
two-fold reduction in bile acid levels (Figure 3G), and led to a 67% reduction in the MBG level
in the conditioned medium (Figure 3E), but did not affect level of progesterone (Figure 3F). In
contrast, silencing of CYP11A1 gene (Figure 3C, D) reduced the expression of CYP11A1 mRNA
by 80%, and levels of CYP11A1 protein by 75%, did not affect the levels of bile acids or MBG
in the conditioned medium (Figure 3E, G), and markedly reduced the level of progesterone
(Figure 3F), a product of cholesterol side-chain-cleavage (Figure 2). Transfection with non-
targeting siRNA did not affect production of any steroids.
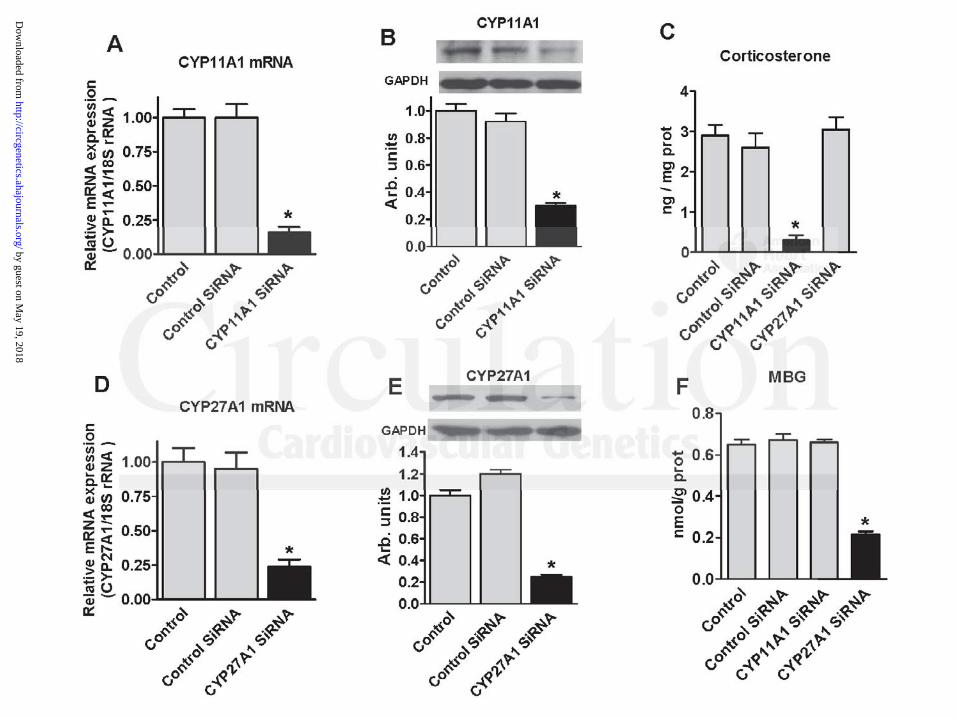
Because the adrenal cortex is a major site for steroidogenesis in mammals,12 it was
important to establish whether CYP27A1 is also implicated in biosynthesis of MBG in
adrenocortical cells. CYP11A1 and CYP27A1 genes were silenced in a primary culture of
adrenocortical cells from Dahl salt-sensitive rats, i.e., cells known to produce substantial
amounts of MBG.12 Figure 4 shows that transfection of adrenocortical cells with a CYP27A1-
specific small interfering RNA reduced expression of CYP27A1 mRNA by 76%, reduced the
CYP27A1 gene reduced the level of CYP27A1 protein by 70% (Figure 3B), induuuceced dd momomorerere ttthahahan a
wo-folold d reduuctioon n in bile acid levels (Figure 3G),), aandn led to a 67% reduuctc ion in the MBG level
nnn thhhe conditiiionononededed mmededediuiuiummm (F(Figigigururureee 3E3E3E),),), bbbututt dddididid nooot aaffececect t t lelelevevv l ofofof ppprororogegegestststeree onneee (F(F(Figigiguru e e e 3F3F3F).).). IIInn n
cocoonntrtrtrast, silencingngng of CYCYCYP11AAA111 geneee (FFFigggureee 333C, D)D)D) redededuuuceddd ttthe eeexxxpressssiooon ofofof CYPP11111AAA1 mmRNRNRNA
by 80%, and levels ooof f f CYCC P11A1A1A111 prp otein byy 75%,,, didid d not afafaffefef ctc the levevvelelels ss of bile acids or MBG
nn tthehe cconondidititiononeded mmedediuium m (F(F( igiggururee 3E3E,,, G)G)),,, anand d mamarkrkededlylyy rrededucuceded tthehe llevevelel oof f prprp ogoggesesteteroronene
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
13
level of CYP27A1 protein by 73%, and also reduced MBG production by 70%, but not that of
corticosterone (Figure 4C-F), a product of cholesterol side-chain cleavage. Conversely, silencing
of CYP11A1 gene in adrenocortical cells reduced the expression of CYP11A1 mRNA by 84%,
level of CYP11A1 protein by 78%, and resulted in 10-fold reduction in the level of
corticosterone (Figure 4A-C), but did not affect the levels of MBG in the conditioned medium
(Figure 4F).
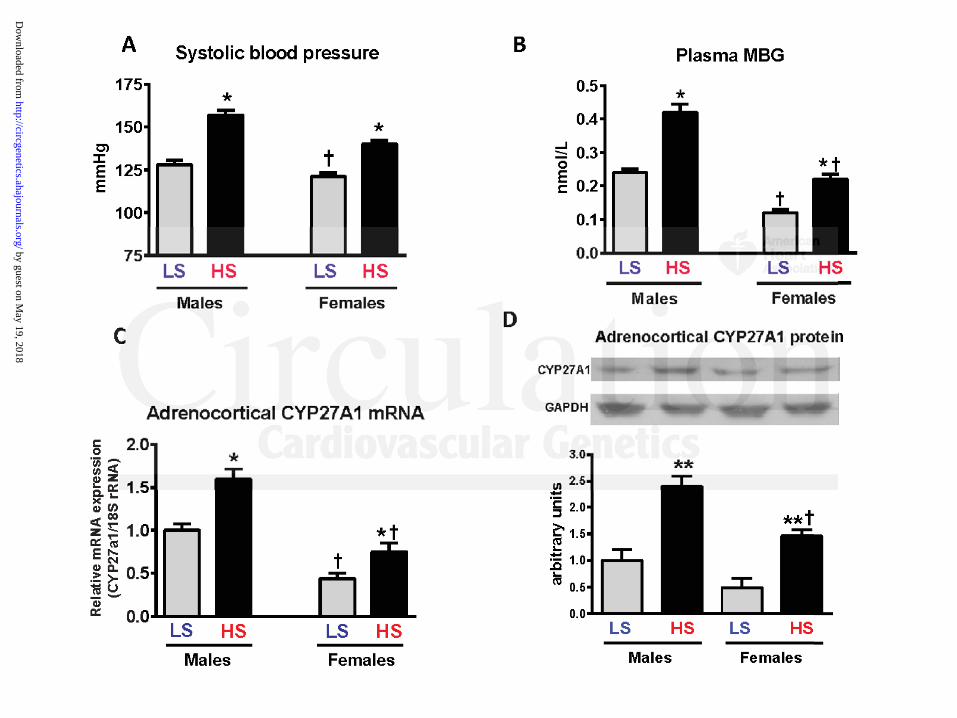
The effect of a high salt diet in Dahl-S rats on blood pressure, plasma MBG levels, and
adrenocortical CYP27A1 and CYP11A1 protein and mRNA is presented in Figures 5-7. After 4
weeks of a high salt diet, male Dahl-S rats demonstrated elevated systolic blood pressure (SBP)
(Figure 5A), increased plasma MBG levels (1.75-fold; Figure 5B), increased adrenocortical
CYP27A1 mRNA and CYP27A1 protein, estimated by Western blotting (Figure 5C,D), and by
immunohistochemistry (Figure 6), but adrenocortical CYP11A1 protein and mRNA levels did
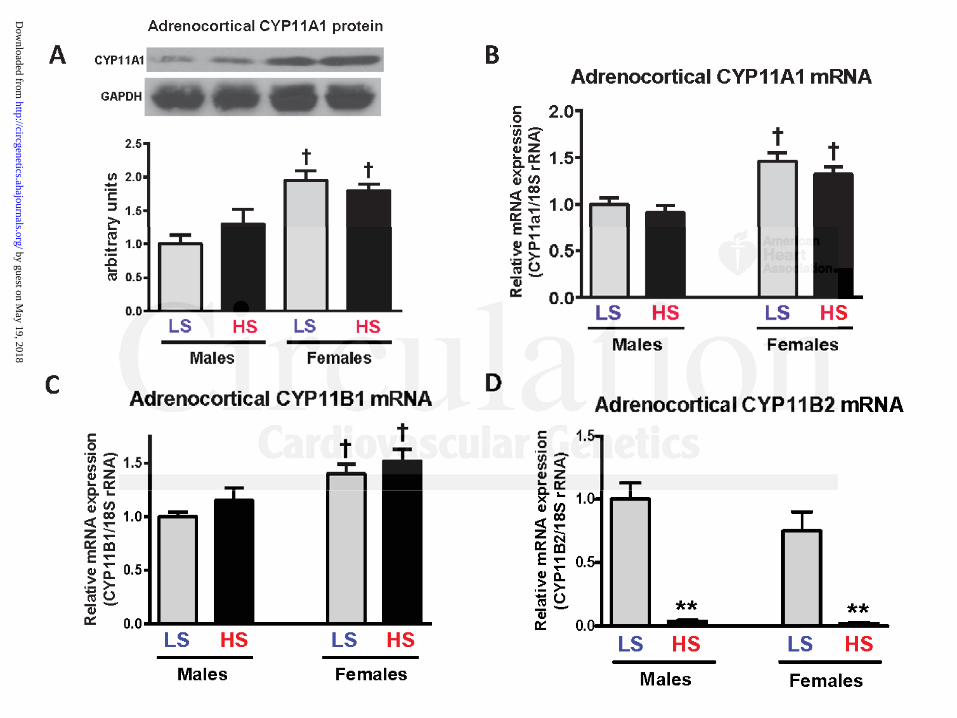
not change (Figure 7A and 7B) vs. the low-salt-fed group. Female Dahl-S rats on a high salt diet
also demonstrated elevated SBP (Figure 5A), increased plasma MBG levels (1.83-fold, Figure
5B), increased adrenocortical CYP27A1 protein level (Figure 5C, Western blotting; Figure 6,
immunohistochemistry), and CYP27A1 mRNA (Figure 5D), but adrenocortical CYP11A1
protein and mRNA levels did not change (Figures 7A and 7B). Notably, that SBP, plasma MBG
levels, and adrenocortical CYP27A1 protein abundance and CYP27A1 mRNA expression were
significantly lower in females than in males on both low and high salt diets (Figure 5). In
contrast, adrenocortical CYP11A1 mRNA expression and CYP11A1 protein levels were higher
in Dahl-S females compared to these parameters in males on both diets (Figure 7A and 7B).
Data on the effect of a high NaCl diet for 4 weeks vs. low salt diet on CYP27A1
abundance, detected by immunostaining in adrenocortical tissue from Dahl-S male and female
Figure 5A), increased plasma MBG levels (1.75-fold; Figure 5B), increased adrreenenocoo ororortititicacacal l l
CYP27A7A1 mRmRNAA and CYP27A1 protein, estimateed d by Western blotting g g (F(( igure 5C,D), and by
mmmmmmunohistococochehehemimm sttryryry (((FiFiFigugug reee 666),),), bbbututt aaadrdd enenenocococortiicaaal CYCYYP1P1P11A1A1 11 prprprotototeieie n n n ananand mRmRmRNANANA levevvelelelsss didididdd
nonoot chchc ange (Figuuureee 7AAA aaand 77B7B) ) ) vs. thhhe loloow-ssalltt-feddd gggrouououppp. FFFemmmaleee DDDahl---S raattsts oon a hhihighghgh saaalt dddieeet
also demonstrated eelelelevavav ted SBSBBPPP (F( iggure e 5A),), increrereasa ed plalaasmsmsma a MBG G lelelevevv ls ((1.83-fold, Figure
5B5B),),), iincncrereasaseded aadrdrenenococorortiticacal l CYCYP2P27A7A11 prprp ototeiein n lelevevell (F(F( igiggurure e 5C5C,,, WeWeststerern n blblotottitingngg;;; FiFigugug rere 66,,,
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
14
rats is presented in Figure 6. The representative immunohistochemical images of adrenocortical
tissue from each Dahl-S experimental groups are shown in Figure 6A-D. CYP27A1 is stained
brown by anti-CYP27A1 antibody. The intensity of CYP27A1 staining was more pronounced in
adrenocortical zona fasciculata compared to other adrenocortical zones, and it was greater in
both males and females fed a high salt diet for 4 weeks compared to the correspondent low-salt-
fed control groups (Figure 6E). No significant gender difference in CYP27A1 abundance was
detected by immunohistochemistry technique (Figure 6).
Previously it was demonstrated that CYP11B1 is predominantly expressed in zona
fasciculata of adrenal cortex, and CYP11B2 in zona glomeruloza,28 therefore, the ratio of
adrenocortical CYP11B1 and CYP11B2 mRNA expression may be used as a marker of a relative
contribution of tissue material from these adrenocortical zones to the production of the steroid of
interest. Expression of adrenocortical CYP11B1 mRNA was significantly higher than CYP11B2
mRNA in both sexes on both diets (Figure 7C,D). The expression ratio of CYP11B1 mRNA to
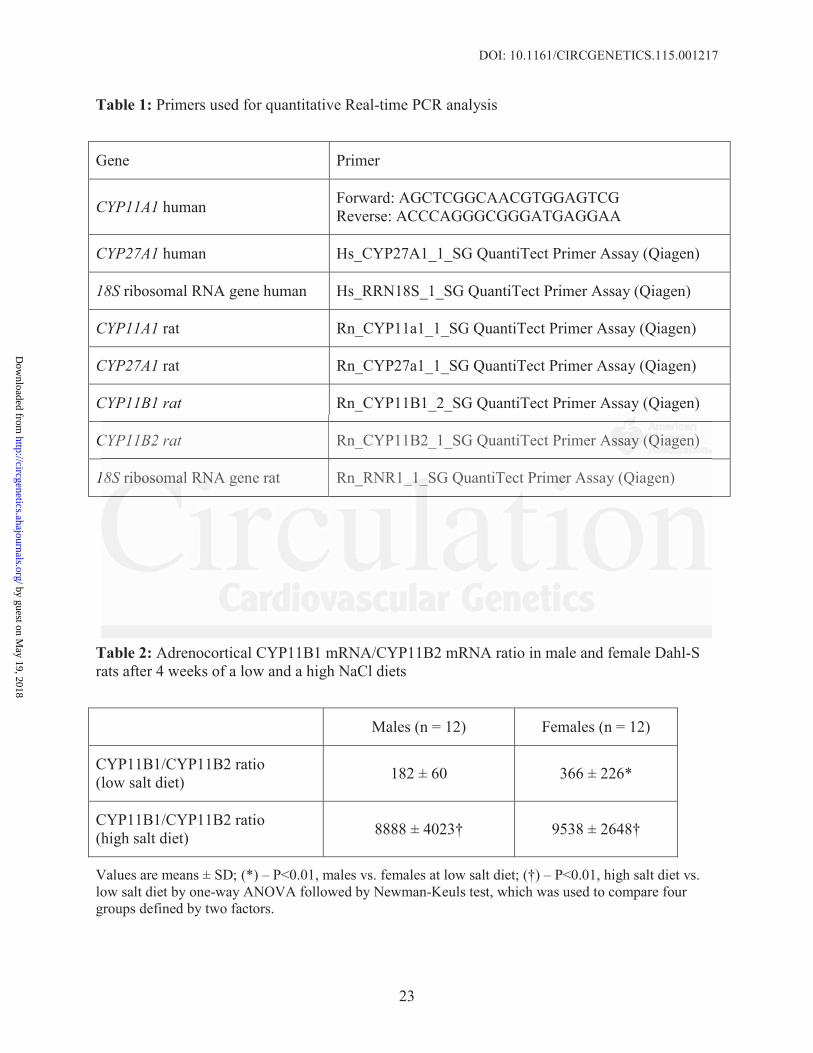
CYP11B2 mRNA in males and females on a low and high NaCl diets are given in Table 2.
Adrenocortical CYP11B1 mRNA did not change after 4 weeks of a high salt diet in both males
and females (Figure 7C), and CYP11B2 mRNA dramatically decreased in both males and
females on a high salt diet vs. low salt intake (Figure 7D). Because the ratio of
CYP11B1/CYP11B2 mRNA expression in adrenocortical samples is >150 (Table 2), and
because CYP11B1 is predominantly expressed in zona fasciculata of adrenal cortex,28 we
conclude that the adrenocortical material in our experiment, used for CYP27A1 mRNA and
protein measurements, was represented by the zona fasciculata.
Discussion
Our results demonstrate, for the first time that the mammalian steroid MBG, an endogenous
adrenocortical CYP11B1 and CYP11B2 mRNA expression may be used as a maaarkrkrkeree ooof f f a a a rererelalalatit ve
contribubutiion oof tiissssue material from these adrenocoortrtical zones to the prododuction of the steroid of
nnnteeerrest. Exprrresesssisisionoo oooff f adadadrererenonn cococortrtrticicicalala CCCYPYPY 111111B1BB mmmRRRNA A A wawawasss signgngnififficicicananantltltly y y highghhererer ttthahahan n CYCYCYP1P1P11B1B1B2
mmRmRNNNA in both ssexxxes ooonnn bothhh dddiets (F(FFigggurrre 7CCC,,,D). TTThe exexexpreesessionnn rrrata io ooof f CYYYPPP11B111 mRmRmRNNNA ttto
CYP11B2 mRNA in n n mamam les annnd d d females s on a loww aaand high h h NaNN ClC dietss aaarerere ggiven in Table 2.
AdAdrerenonococortrticicalal CCYPYP1111B1B1 mmRNRNA A didid d nonot t chchanangegeg aaftfterer 44 wweeeeksks oof f a a hihighghg ssalalt t didietet iin n bobothth mmalaleses
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
15
sodium pump ligand of bufadienolide nature, is derived from bile acids, and its biosynthesis is
initiated by CYP27A1 enzyme. This observation was made in rat adrenocortical and human
placental cells, which were chosen as models for bile acid pathway studies, because these tissues
are known to produce MBG. Thus we cannot exclude that MBG can be produced in other
mammalian tissues and organs. Bile acid synthesis originally was thought to be limited to the
liver.24 Later, the extra-hepatic synthesis of bile acids via an “acidic” pathway was described,
and bile acids were identified not merely as a product of cholesterol elimination, but also as
regulatory signaling molecules implicated in genesis of several diseases including diabetes and
cancer.21,22,29 Growing evidence indicates the physiological importance of regulatory enzyme
CYP27A1 for “acidic” bile acid pathway, which participates in oxysterol formation, cholesterol
transport and homeostasis, and activates metabolic signaling pathways.21,22,30,31 Activation of
CYP27A1 is an adaptive mechanism for cholesterol utilization in human adipocytes following by
the de novo biosynthesis of steroids.25 CYP27A1 also increased the expression of markers of
EMT and fibrosis in mouse breast cancer cells.26 The results of the present study indicated that
the production of MBG, which participates in initiation of the pro-fibrotic pathways in different
tissues in clinical settings and experimental models,2,3,10 is also controlled by CYP27A1 in both
rat and human tissues.
Excessive production of MBG causes inhibition of vascular Na/K-ATPase and
accompanies elevated blood pressure in essential hypertension,1 chronic kidney disease,2,6 and
preeclampsia,3 a major cause of maternal and fetal morbidity worldwide.8 An increasing body of
evidence indicates, that in addition to its traditional transport function, Na/K-ATPase is a potent
generator of cell signaling.5,15 In addition to increasing vascular tone, MBG induces cardiac and
vascular fibrosis,10 which is a hallmark of cardiovascular aging, resistant hypertension, chronic
CYP27A1 for “acidic” bile acid pathway, which participates in oxysterol formatioioion,n,n cchohoholelelestststeere ol
ransppporort and d homemeostasis, and activates metabolicc ssignaling pathways.21,22,30,31 Activation of
CYCYCYPPP27A1 is aaan n n adadadappptititiveveve mmmecee hahahanininismsmsm fffororor chohoholelelesterololol utiilililizazazatititionoo iiinnn huhuhumamamann n adaa ippocococytytyteseses fololollololowiwiwingngng bby
hhhe e dedd novo biosyyynnnthesssisss of sssteteeroror ids.25 CCCYYYP2227AAA1 aaalssso ininincccreaaaseeed thhheee exprrressssiiononon of mmamarrkrkeeersss of
EMT and fibrosis in n n momom use brbrreaeaeast cancec r cells.26 TTThe resululltststs of the prp esesesenenent study indicated that
hhe e prprp ododucuctitionon oof f MBMBG,G,, wwhihichch ppparartiticicipapap tetes s inin iininititiatatioion n ofof tthehe ppproro-fifibrbrototicic pppatathwhwayayys s inin ddififfefererentnt
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
16
kidney disease, and preeclampsia.16,17,32 In preeclampsia, heightened MBG levels contributed to
blood pressure increase3,9,32 and development of vascular remodeling.3 Notably, that infusion of
MBG to rats produced effects similar to cardiovascular diseases in animal models of salt-
sensitive hypertension and chronic renal failure.6,10,14 In our previous study the infused MBG
increased blood pressure, changed ventricular hemodynamics, and increased cardiac and aortic
deposition of collagen-1 in parallel with cardiovascular remodeling.14 Physiologically relevant
concentrations of MBG stimulate synthesis of collagen-1 in vitro and induce cardiac fibrosis in
rats with renal failure,10 and an anti-MBG monoclonal antibody potently reverses cardiac
fibrosis.2,6
In the present study, we have shown that mammalian MBG production is controlled by
CYP27A1 enzyme, but not by CYP11A1. Silencing of CYP27A1 mRNA in adrenocortical cell
cultures reduced total bile acids and MBG levels, indicating the participation of the “acidic” bile
acid pathway in MBG production. In contrast, silencing of CYP11A1 mRNA did not affect
MBG, but caused a decrease in progesterone (JEG-3 cells) and in corticosterone (rat
adrenocortical cells), which require side-chain cleavage of cholesterol. This is in agreement with
the previous observation that the “traditional” steroidogenesis via the side-chain cleavage of
cholesterol is not involved in the biosynthesis of MBG.20 In this study, the de novo biosynthesis
of MBG in murine Y1 adrenocortical cells required cholesterol as a precursor, but did not
involve the conversion of cholesterol to pregnenolone via side chain cleavage by CYP11A1.20
A discovery made more than 40 years age in Bufo marinus toads,33,34 which produce
large quantities of bufadienolides, including MBG, for protection against predators, indicated
that amphibian bufadienolides are synthesized from bile acids.33,34 Those authors demonstrated
that in toads, the in vivo administration of radioactive label from cholesterol-14 -
In the present study, we have shown that mammalian MBG production iss cocoontntntrororollllllededed bbby
CYP27A7A1 ennzyzymeme, but not by CYP11A1. Silencinng g g of CYP27A1 mRNA A in adrenocortical cell
cucuultttuures reducuccededd tttotoo alall bbbililileee acaca iddds s s ananand d d MBMBMBG GG lelelevevevels, iiinddicaaatititingngng ttthe ppparrrtititicicic papapatitition ooof f f thththe e e “acicicidididic”c”c bbbililile
acaccididd ppathwayyy in MMMBGG ppproduucuctitition. Innn cononontrassst, silenncccingngng oooff CYCYCYP11AAA1 mRRRNANANA dddid nootot aaffffeccct
MBG, but caused a dededecrcrc ease iiin n n prprp ogogesteterone ((JEGGG-3 3 cells)s)s) aaandndn in cortrticicicosoo terone (rat
adadrerenonococortrticicalal ccelellsls),),), wwhihichch rreqeqquiuirere ssidide-e chchaiain n clcleaeavavagegeg oof f chchololesesteterorol.l. TThihiss isis iin n agaggrereememenent t wiwithth
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
17
hydroxy- -cholanate-24-14C, but not from radiolabeled pregnenolone, was incorporated into
bufadienolide, including MBG.31 Notably, a product of acidic pathway of bile acid biosynthesis
in h -hydroxy-5-cholestenoic acid.35 Interestingly, some researches
-hydroxy- -cholanate in the “acidic” bile acid pathway.25,30,36,37 It was previously
demonstrated that a major biological function of CYP27A1 in human tissues is conversion of
cholesterol into 27- -hydroxy-5-cholestenoic acid,38 which regulate the
expression of nuclear receptors and modulate cholesterol metabolism.35 -
hydroxy- -cholanate was found in amniotic fluid of human fetuses, human newborn urine,39,40
and in plasma of pregnant women,41 which is consistent with our observation of elevated plasma
MBG levels in pregnancy.3,9,32 The present findings, that human choriocarcinoma cells produced
MBG (Figure 1), and that MBG production is affected by silencing of CYP27A1 (Figure
3A,B,E), are in agreement with the previous data.
27-hydroxycholesterol (Figure 2) is an endogenous selective estrogen receptor modulator
and adversely affects estrogen-related cardiovascular protection.42 The upregulation of
CYP27A1, which is important for the production of 27-hydroxycholesterol, may have a dramatic
impact upon the cardiovascular system, bone biology and cancer.42,43 Circulating levels of 27-
hydroxycholesterol increase with age, and are lower in females than in males,42 indicating that
CYP27A1 activity and CYP27A1 gene expression may be lower in females,44 which is consistent
with our present observation, that mRNA CYP27A1 level in adrenocortical tissue from female
rats is 2.5-fold lower than in males. Notably, in the present study CYP11A1 protein abundance,
CYP11A1 mRNA and CYP11B1 mRNA are higher in females than in males Dahl-S rats, which
is also in agreement with the previous finding.44 This indicates that in females, higher expression
of genes involved in sex steroid and mineralocorticoid hormone synthesis is accompanies by
MBG levels in pregnancy.3,9,32 The present findings, that human choriocarcinommaaa cececelllllss prprprodododucuu ed
MBG (F(F( igiggure e 1))),,, ana d that MBG production is affecectet d by silencing of CYCYP27A1 (Figure
3A3AA,B,B,B,E), are iiin n agagagrer emememenenenttt wiww ththh ttthehehe ppprerereviviviouuusss dadadata.
27-hyyydroooxyyychoooleeesteroolol (((Figuurerer 222)) is aaan endddogggenononouuus sssellectiiivveve estrrrogggennn rrereceptttororr mmmooddulaaatooor
and adversely affectttsss esese trt ogggennn---rererelaated cac rdiovascculuu ara pproteteectctctioion.42 Thehee uuuprprp eggulation of
CYCYP2P27A7A1,1,, wwhihichch iiss imimpopop rtrtanant t fofor r ththe e prprp ododucuctitionon oof f 2727-hyhyydrdroxoxycycy hoholeleststererolol,,, mamay y y hahaveve aa ddraramamatiticc
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
18
lower expression of genes regulating basal cell function, which may include production of
natriuretic steroid MBG. Interestingly, circulating levels of MBG in female rats were also 2-fold
lower than in males, and were associated with lower systolic blood pressure in females at
baseline than in males. It is tempting to speculate that a lower circulating MBG level in females
is an adaptive mechanism, because, during pregnancy, additional MBG will be synthetized in
placenta, as demonstrated in our previous4 and in the present studies.
Expression of CYP27A1 mRNA in adrenocortical tissue in both males and females
increased 1.6-fold after 4 weeks on a high NaCl diet, and was associated with increase in plasma
MBG and elevation of blood pressure in both sexes. The observation, that a high salt diet-
induced upregulation of adrenocortical CYP27A1 mRNA and increase in CYP27A1 protein
abundance was accompanied by an increase in circulating MBG, as well as the finding that in
vitro silencing of CYP27A1 mRNA in rat adrenocortical cell culture was accompanied by a
decrease in produced MBG levels, provides a strong evidence for a relationship between
CYP27A1 activation and production of the steroid MBG. Previous observations that bilateral
adrenalectomy resulted in partial reduction of plasma MBG levels,14 indicated that adrenocortical
tissue is the predominant, but not the sole, source of MBG in mammals. In the present and our
previous studies4 we demonstrated that MBG is also produced by placental cells.
Thus, the present results demonstrate that mammalian bufadienolide steroid MBG, an
endogenous ligand of the Na/K-ATPase and a pro-hypertensive factor, is very likely to be
synthesized via an extra-hepatic “acidic” bile acid pathway similar to amphibian MBG.34
Specific chemical reactions in the transformation of bile acids into bufadienolide molecules have
been described in amphibians35; however, these reactions remain unknown in mammals and
merit further investigation. Our present finding, that biosynthesis of pro-hypertensive and pro-
nduced upregulation of adrenocortical CYP27A1 mRNA and increase in CYP2777A1A1A1 ppprororoteteteininin
abundaancnce wawas acaccoc mpanied by an increase in circculu ating MBG, as welll aas the finding that in
vivivitrrooo silencinnnggg ofofof CCCYPYPYP272727A1A1A1 mmmRNRNRNA A A ininn rrrataa adadadrererenocccorrrticalalal cccelelell l l cuultltltururreee wawawasss acaa cooompmpmpanananieiei d d d bybyby aaa
dedeecrrreae se in prp odducucuced MMMBG lllevevvels, ppprorr vvvidddes aaa ssstrongngng evivividededencceee forrr aa a relatttiooonshshhipipp betweweweenenn
CYP27A1 activationonn aaandn pprooduduductcc ion of the steroididd MBG. PrPrPreveve ious obsbssererervav tions that bilateral
adadrerenanalelectctomomy y y reresusultlteded iin n papap rtrtiaial l rereduductctioion n ofof ppplalasmsma a MBMBG G lelevevelsls,,,14144 inindidicacateted d ththatat aadrdrenenococorortiticaca
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
19
fibrotic steroid MBG is controlled by CYP27A1, gives a new direction for future studies that will
likely enable the emergence of novel therapeutic strategies to block MBG production in order to
reduce its impact on highly prevalent human diseases involving heightened levels of MBG.
Acknowledgments: The authors are grateful to Dr. Christopher H. Morrell for statistical
analyses and Ruth Sadler for editorial assistance.
Funding Sources: Supported by Intramural Research Program, National Institute on Aging, NIH
Conflict of Interest Disclosures: None.
References:
1. Fedorova OV, Talan MI, Agalakova NI, Lakatta EG, Bagrov AY. An endogenous ligand of alpha-1 sodium pump, marinobufagenin, is a novel mediator of sodium chloride dependent hypertension. Circulation. 2002;105:1122-1127.
2. Kolmakova EV, Haller ST, Kennedy DJ, Isachkina AN, Budny GV, Frolova EV, et al. Endogenous cardiotonic steroids in chronic renal failure. Nephrol Dial Transplant.2011;26:2912-2919.
3. Nikitina ER, Mikhailov AV, Nikandrova ES, Frolova EV, Fadeev AV, Shman VV, et al. In preeclampsia endogenous cardiotonic steroids induce vascular fibrosis and impair relaxation of umbilical arteries. J Hypertens. 2011;29:769-776.
4. Fedorova OV, Tapilskaya NI, Bzhelyansky AM, Frolova EV, Nikitina ER, Reznik VA, et al. Interaction of Digibind with endogenous cardiotonic steroids from preeclamptic placentae. JHypertens. 2010;28;361-366.
5. Bagrov AY, Shapiro JI, Fedorova OV. Endogenous cardiotonic steroids: physiology, pharmacology and novel therapeutic targets. Pharmacol Rev. 2009;61:9-38.
6. Haller ST, Kennedy DJ, Shidyak A, Budny GV, Malhotra D, Fedorova OV, et al. Monoclonal antibody against marinobufagenin reverses cardiac fibrosis in rats with chronic renal failure. Am J Hypertens. 2012;25:690-696.
7. Bagrov AY, Fedorova OV, Dmitrieva RI, Howald WN, Hunter AP, Kuznetsova EA, et al. Characterization of a urinary bufodienolide Na+,K+-ATPase inhibitor in patients after acute myocardial infarction. Hypertension. 1998;31:1097-1103.
References:
1. Fedororovo a OVO ,,, TaT lan MI, Agalakova NI, Lakatta a EGE , Bagrov AY. An n ene dogenous ligand of alphphpha-a-a-111 sodididiuumum pppuuump, marinobufagenin, is a novelele mmmediator of sodddium chchchlol ride dependent hyhyhyppepertensionn. CiCiCircrcrcullatatatiiiononon. 2022 020202;1;1;1050505:1111212122--1111112722 .
2.2.. KKKolo makova EV,VV Haaalleeer STTT, KeKK nnedede yyy DDDJ, IIIsaaachkkkinnna ANANAN, BuBuBudnyyy GGGV, FFFroooloovovaaa EV, etett alll.Endododogegegenononoususus ccararardddiotototonononicicc sssteroooididids s ininin cchrronononicicic rrrenenenalalal fffaiiilululurerere. NeNeNephphphrorr l ll DiDiDialalal TTTraaansnsn plplplananantt.t.2011;26:2912-2919.9..
3.3. NNikikititinina a ERER, , , MiMikhkhaiailolov v AVAV, , , NiNikakandndrorovava EES,S,, FFrorololovava EEV,V,, FFadadeeeev v AVAV, , , ShShmaman n VVVV,,, etet aal.l. IIn n prpreeeeclclamampspsiaia eendndogogenenououss cacardrdioiototoninicc ststereroioidsds iindnducucee vavascsculularar ffibibrorosisiss anandd imimpapairir rrelelaxaxatatioionn ofof
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
20
8. Sibai B, Dekker G, Kupferminc M. Pre-eclampsia. Lancet. 2005;365:785-799.
9. Fedorova OV, Simbirtsev AS, Kolodkin NI, Kotov AY, Agalakova NI, Kashkin VA, et al. Monoclonal antibody to an endogenous bufadienolide, marinobufagenin, reverses preeclampsia-induced Na/K-ATPase inhibition and lowers blood pressure in NaCl-sensitive hypertension. JHypertens. 2008;26:2414-2425.
10. Kennedy DJ, Vetteth S, Periyasamy SM, Kanj M, Fedorova L, Khouri S, et al. Central role for the cardiotonic steroid, marinobufagenin, in the pathogenesis of experimental uremic cardiomyopathy. Hypertension. 2006;47:488-495.
11. Fedorova OV, Kolodkin NI, Agalakova NI, Lakatta EG, Bagrov AY. Marinobufagenin, an endogenous alpha-1 sodium pump ligand, in hypertensive Dahl salt-sensitive rats. Hypertension. 2001;37:462-466.
12. Fedorova OV, Agalakova NI, Talan MI, Lakatta EG, Bagrov AY. Brain ouabain stimulates peripheral marinobufagenin via angiotensin II signaling in NaCl loaded Dahl-S rats. J Hypertens.2005;23:1515-1523.
13. Doris PA. Immunological evidence that the adrenal gland is a source of an endogenous digitalis-like factor. Endocrinology. 1988;123:2440-2444.
14. Elkareh J, Kennedy DJ, Yashaswi B, Vetteth S, Shidyak A, Kim EG, et al. Marinobufagenin stimulates fibroblast collagen production and causes fibrosis in experimental uremic cardiomyopathy. Hypertension. 2007;49:215-224.
15. Cereijido M, Contreras RG, Shoshani L, Larre I. The Na+K+ATPase as self-adhesion molecule and hormone receptor. Am J Physiol Cell Physiol. 2012;302:C473-C481.
16. Briet M, Schiffrin EL. Aldosterone: effects on the kidney and cardiovascular system. Nat Rev Nephrol. 2010;6:261-273.
17. Laurent S, Alivon M, Beaussier H, Boutouyrie P. Aortic stiffness as a tissue biomarker for predicting future cardiovascular events in asymptomatic hypertensive subjects. Ann Med. 2012;44:S93-S97.
18. Payne AH, Hales DB. Overview of steroidogenic enzymes in the pathway from cholesterol to active steroid hormones. Endocr Rev. 2004;25:947-970.
19. Murrell JR, Randall JD, Rosoff J, Zhao JL, Jensen RV, Gullans SR, et al. Endogenous ouabain: upregulation of steroidogenic genes in hypertensive hypothalamus but not adrenal. Circulation. 2005;112:1301-1318.
20. Dmitrieva RI, Bagrov AY, Lalli E, Sassone-Corsi P, Stocco DM, Doris PA. Mammalian bufodienolide is synthesized from cholesterol in the adrenal cortex by a pathway which is independent of cholesterol side-chain cleavage. Hypertension. 2000;36;442-448.
2005;23:1515-1523.
13. Doris PA. Immunological evidence that the adrenal gland is a source of an endododogggenononoususus digitaliiss-llike faf cttoror. Endocrinology. 1988;123:244040-2444.
14144. EEElkareh J,JJ KKKenenenneedydydy DDDJJ,J, Yasasashahahaswswswi ii B,B,B, VVVetetetteteteth SSS, Shididdyayayak k k A,AA KKKimmm EEEG,G,G, eeet all. . . MaMaMariririnonn bububufafafagegegenininin nttimmmulates fibrobbblaaast ccoollllllagennn ppprodududuccctiooonnn anddd cccausesss fibrbrbrososisisis iiin exxxppperimeeennntalll uuuremiiccc
cacaardrddioi myopathyhy.. HHHypeeerttensiononon. 200777;4449::21555-22224.
15. Cereijido M, Cononontrtrtrere as RRG,G,G, Shoshana i L, Larreree I. The NaNaNa+K+ATPaPasesese aaas self-adhesion ffmolecule and hormooonenene rrrecececepepeptototor.r. AmAmAm JJJ PPPhyhyhysisisiololol CCCelelell l l PhPhPhyyysioioolll.. 202020121212;3;33020202:C:C:C4747473-3-3-CCC481.
1616 BrBrieiett MM SSchchififfrfrinin EELL AAldldososteteroronene:: efeffefectctss onon tthehe kkididneneyy anandd cacardrdioiovavascsculularar ssysystetemm NaNatt ReRevv
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
21
21. Javitt NB. Oxysteroids: a new class of steroids with autocrine and paracrine functions. Trends Endocrinol Metab. 2004;15:393-397.
22. Pikuleva IA. Cytochrome P450s and cholesterol homeostasis. Pharmacol Ther.2006;112:761-773.
23. Thomas C, Pellicciari R, Pruzanski M, Auwerx J, Schoonjans K. Targeting bile-acid signalling for metabolic diseases. Nat Rev Drug Discov. 2008;7:678-693.
24. Javitt NB. Bile acid synthesis from cholesterol: regulatory and auxiliary pathways. FASEB J.1994;8:1308-1311.
25. Li J, Daly E, Campioli E, Wabitsch M, Papadopoulos V. De novo synthesis of steroids and oxysterols in adipocytes. J Biol Chem. 2014;289:747-764.
26. Nelson ER, Wardell SE, Jasper JS, Park S, Suchindran S, Howe MK, et al. 27-Hydroxycholesterol links hypercholesterolemia and breast cancer pathophysiology. Science.2013;342:1094-1098.
27. Uno Y, Hosaka S, Yamazaki H. Identification and analysis of CYP7A1, CYP17A1, CYP20A1 and CYP51A1 in Cynomolgus Macaques. J Vet Med Sci. 2014;76:1647-1650.
28. Dringenberg T, Schwitalla M, Haase M, Scherbaum WA, Willenberg HS. Control of CYP11B2/CYP11B1 expression ratio and consequences for the zonation of the adrenal cortex. Horm Metab Res. 2013;45:81-85.
29. Hylemon PB, Zhou H, Pandak WM, Ren S, Gil G, Dent P. Bile acids as regulatory molecules. J Lipid Res. 2009;50:1509-1520.
30. Norlin M, Wikvall K. Enzymes in the conversion of cholesterol into bile acids. Curr Mol Med. 2007;7:199-218.
31. Nebert DW, Wikvall K, Miller W. Human cytochromes P450 in health and disease. Philos Trans R Soc Lond B Biol Sci. 2014;368:1-21.
32. Lopatin DA, Ailamazian EK, Dmitrieva RI, Shpen VM, Fedorova OV, Doris PA, Bagrov AY. Circulating bufodienolide and cardenolide sodium pump inhibitors in preeclampsia. JHypertens. 1999;17:1179-1187.
33. Chen C, Osuch MV. Biosynthesis of bufadienolides--3-beta-hydroxycholanates as precursors in Bufo marinus bufadienolides synthesis. Biochem Pharmacol. 1969;18:1797-1802.
34. Porto AM, Baralle FE, Gros EG. Biosynthesis of bufadienolides in toads. J Steroid Biochem.1972;3:11-17.
2013;342:1094-1098.
27. Uno Y, Hosaka S, Yamazaki H. Identification and analysis of CYP7A1, CYP1P117A7A7A1,11 CYP20A0A1 annd d CYCYP51A1 in Cynomolgus Macaququeses. J Vet Med Sci. 201414;76:1647-1650.
28288. DDDringenbbberererg g g T,TT SSchchchwiwiwitatatalll a a M,M,M, HHHaaaaasesese MMM,, ScScScherrrbaaaum m m WAWAWA,,, WiWiWillllenenenbebebergrgrg HHS.. CCCononontrtrtrolo ooofff CYCYCYP11B2/CYP1111B1 exexexprp essisisiooon ratatatiioio aaannnd coononsequuueenceeesss fofoorr r ththhe zzzonnnationnn ooof thththeee adrereenananal cocoortexxx. HHoHormrmrm Metab Resss. 2013;3;45:81-1--858 .
29. Hylemon PB, Zhhhououou H, Paaandndndakak WM, Ren S, GiGiGil G, Denennt t t P.P. Bile acaccidididsss as regulatory molecules. J Lipid ReReResss.. 202020090909;5;5;50:0:0:151515090909-1-1-15252520.0.0
3030 NoNorlrlinin MM WiWikvkvalalll KK EEnznzymymeses iinn ththee coconvnverersisionon ooff chchololesesteteroroll inintoto bbililee acacididss CuCurrrr MMolol
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
22
35. Pikuleva I, Javitt MB. Novel sterols synthesized via the CYP27A1 metabolic pathway. Arch Biochem Biophys. 2003;420:35-39.
36. Pikuleva IA, Babiker A, Waterman MR, Björkhem I. Activities of recombinant human cytochrome P450c27 (CYP27) which produce intermediates of alternative bile acid biosynthetic pathways. J Biol Chem. 1998;273:18153-18160.
37. Chiang JYL. Regulation of bile acid synthesis. Front Biosci. 1998;3:176-193.
38. Meaney S, Babiker A, Lütjohann D, Diczfalusy U, Axelson M, Björkhem I. On the origin of the cholestenoic acids in human circulation. Steroids. 2003;68:595-601.
39. Délèze G, Paumgartner G, Karlaganis G, Giger W, Reinhard M, Sidiropoulos D. Bile acid pattern in human amniotic fluid. Eur J Clin Invest. 1978;8:41-55.
40. Makino I, Sjövall J, Norman A, Strandvik B. Excretion of 3beta-hydroxy-5-cholenoic and3a-hydroxy-5a-cholanoic acids in urine of infants with biliary atresia. FEBS Lett. 1971;15:161-164.
41. Fulton IC, Douglas JG, Hutchon DJR, Beckeett GL. Is normal pregnancy cholestatic? Clin Chim Acta. 1983;130:171-176.
42. Umetani M, Shaul PW. 27-hydroxycholesterol: the first identified endogenous SERM. Trend Endocrinol Metab. 2011;22:130-135.
43. Lee WR, Ishikawa T, Umetani M. The interaction between metabolism, cancer and cardiovascular disease, connected by 27-hydroxycholesterol. Clin Lipidol. 2014;9:617-624.
44. Trejter M, Hochol A, Tyczewska M, Ziolkowska A, Jopek K, Szyszka M, et al. Sex-related gene expression profiles in the adrenal cortex in the mature rat: Microarray analysis with emphasis on genes involved in steroidogenesis. Int J Mol Med. 2015;35:702-714.
164.
41. Fulton IC, Douglas JG, Hutchon DJR, Beckeett GL. Is normal pregnancy cholelelestststataticicic??? ClClClininin Chim AActcta. 1919833;1;1; 30:171-176.
424242. UUUmetani M,M,M, SSShahh ululul PPPW.W.W. 227-7-hyhyhydrdrdroooxyxyxychchcholollesesestett rolll: ttthe fffiririrststst iiided ntntntiififieieied dd enenendodd geenononoususus SSSERERRM.M.M TrTrTrenenendEEnE dddocrinol Metababab. 2001111;1;2222:1113330-1353535.
433. LeLeLeee e WRWRWR, IsIsshhihikakakawawawa TTT, , , Ummetetetananiii M.MM TTThehehe iiintntnterereracacactttiononon bbbetetetweweweenenn mmetete abababolololiisism,m,m, ccananancececer r r aanand d d cardiovascular diseaaasesese,, coc nnecececteteted byy 27-7 hyydroxycycychoholesterrrololol... ClC in Lipippidididoloo . 2014;9:617-624.
4444. . TrTrejejjteter r M,M,, HHocochohol l A,A,, TTycycy zezewswskaka MM, , , ZiZiololkokowswskaka AA,,, JoJopepep k k K,K,, SSzyzyyszszkaka MM, , , etet aal.l. SSexex-rerelalateted dgegenene eexpxpreressssioionn prprofofilileses iinn ththee adadrerenanall cocortrtexex iinn ththee mamatuturere rratat:: MiMicrcroaoarrrrayay aananalylysisiss wiwithth
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from
DOI: 10.1161/CIRCGENETICS.115.001217
23
Table 1: Primers used for quantitative Real-time PCR analysis
Gene Primer
CYP11A1 human Forward: AGCTCGGCAACGTGGAGTCGReverse: ACCCAGGGCGGGATGAGGAA
CYP27A1 human Hs_CYP27A1_1_SG QuantiTect Primer Assay (Qiagen)
18S ribosomal RNA gene human Hs_RRN18S_1_SG QuantiTect Primer Assay (Qiagen)
CYP11A1 rat Rn_CYP11a1_1_SG QuantiTect Primer Assay (Qiagen)
CYP27A1 rat Rn_CYP27a1_1_SG QuantiTect Primer Assay (Qiagen)
CYP11B1 rat Rn_CYP11B1_2_SG QuantiTect Primer Assay (Qiagen)
CYP11B2 rat Rn_CYP11B2_1_SG QuantiTect Primer Assay (Qiagen)
18S ribosomal RNA gene rat Rn_RNR1_1_SG QuantiTect Primer Assay (Qiagen)
Table 2: Adrenocortical CYP11B1 mRNA/CYP11B2 mRNA ratio in male and female Dahl-Srats after 4 weeks of a low and a high NaCl diets
Males (n = 12) Females (n = 12)
CYP11B1/CYP11B2 ratio(low salt diet) 182 ± 60 366 ± 226*
CYP11B1/CYP11B2 ratio(high salt diet) 8888 ± 4023† 9538 ± 2648†
Values are means ± SD; (*) – P<0.01, males vs. females at low salt diet; (†) – P<0.01, high salt diet vs. low salt diet by one-way ANOVA followed by Newman-Keuls test, which was used to compare four groups defined by two factors.
CYP11B2 rat Rn_CYP11B2_1_SG QuantiTect Primer AsAsAssaaay y y (Q(Q(Qiaiaiagegegen)n)n)
18S ririribobobosososomamamal RNRNRNA gene rat Rn_RNR1__1_1_1_SG QuantiTect Primmmererer Assay (Qiagen)
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
24
Figure Legends:
Figure 1: Structure of marinobufagenin (MBG), and MBG production by JEG-3 cells. A,
Structure of marinobufagenin. B, Time course of release of MBG by JEG-3 cells in the medium.
Each bar represents means ± SEM from 18-24 observations. Statistical analysis: repeated-
measures ANOVA followed by Bonferroni test: (*) – P<0.01 vs. 0 hour, (†) – P<0.01 vs. 3
hours. C, Elution of MBG-immunoreactive material produced by JEG-3 cells following
fractionation of chloroform-extracted condition medium on reverse-phase HPLC column. D,
Elution of MBG standard purified from the Bufo marinus toad parotid gland secretion7 on
reverse-phase HPLC column. Both MBG-immunoreactive material and MBG standard eluted in
a single peak at 24 minutes (fraction 16).
Figure 2: Scheme of biosynthesis of bile acids and of traditional steroidogenesis. Enzymes
targeted by SiRNA method are marked.
Figure 3: Effect of silencing of CYP11A1 and CYP27A1 genes in JEG-3 cells on CYP27A1
mRNA (A), CYP27A1 protein (B), CYP11A1 mRNA (C) and CYP11A1 protein (D),
concentrations of marinobufagenin (MBG) (E), progesterone (F), and total bile acids (G) in the
conditioned medium. Each bar represents means ± SEM from 15-20 observations. Statistical
analysis: one-way ANOVA followed by Bonferroni test: (*) – P<0.01 vs. Control and Control
SiRNA.
Figure 4: Effect of silencing of CYP11A1 and CYP27A1 genes in primary culture of rat
adrenocortical cells on CYP11A1 mRNA (A), CYP11A1 protein (B), CYP27A1 mRNA (D),
CYP27A1 protein (E), corticosterone (C), and marinobufagenin (MBG) (F) in conditioned
everse-phase HPLC column. Both MBG-immunoreactive material and MBG standadadardd eelulutet d d ini
a single peak at 24 minutes (fraction 16).
FFFigguure 2: Scheeemememe of ff bbibiosososynynynththhesesesiisis oof ff bibibileee acicicidsdss anddd ooof trtrtradadaditititioioionanal ll stttereroioioidododogggeneneesisisis.s EEEnznznzymymymeseses
aaargrgrgetete ed by SiSiSiRNRNNAAA meeethhhod aaareree markekek ddd.
Figug re 3: Effect of sisiilllencinini g g offof CCCYPYPYP11111A1A1A1 a ddnd CCCYPYPYP272727A1A ggenes iiin JEJEJEG-GG 333 cecelllllls on CYP27A1
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

DOI: 10.1161/CIRCGENETICS.115.001217
25
media. Each bar represents means ± SEM from 15-20 observations. Statistical analysis: one-way
ANOVA followed by Bonferroni test: (*) – P<0.01 vs. Control and Control SiRNA.
Figure 5: Effect of a 4 week high (8%) NaCl diet on blood pressure (A), circulating MBG (B),
adrenocortical CYP27A1 mRNA (C), and CYP27A1 protein abundance (D) (Western blotting)
in male and female Dahl-S rat on a low and a high NaCl diets. Each bar represents means ±
SEM. Statistical analysis: one-way ANOVA followed by Bonferroni test: (*) – P<0.01, high salt
(HS) vs. low salt (LS) diet; (†) – P<0.01, females vs. males.
Figure 6: Immunohistochemical detection of CYP27A1 protein abundance in adrenocortical
tissue of male (A,B) and female DS (C,D) on a low salt (LS) and high salt (HS) diets. CYP27A1
is stained brown. E, Statistical analysis of individual images of adrenocortical CYP27A1
abundance. Grey bars represent CYP27A1 abundance at a low salt intake in males and females,
black bars represent CYP27A1 abundance at a high salt intake in both males and females. Each
bar represents means ± SEM of the percentage of brown stain of adrenal cortex for CYP27A1
from the total adrenocortical area; 4-5 slides of adrenocortical tissue from each of 6 rats per
group were analyzed. By one-way ANOVA followed by Bonferroni test: (*) – P<0.05, high salt
(HS) vs. low salt (LS) diet ZG – zona glomerulosa; ZF – zona fasciculata.
Figure 7: Adrenocortical expression of CYP11A1 protein (A), CYP11A1 mRNA (B), CYP11B1
mRNA (C), and CYP11B2 mRNA (D) in male and female Dahl-S rats. Each bar represents
means ± SEM. Statistical analysis: one-way ANOVA followed by Bonferroni test: (*) – P<0.01,
high salt (HS) vs. low salt (LS) diet; (†) – P<0.01, females vs. males.
Figure 6: Immunohistochemical detection of CYP27A1 protein abundance in adreeenononocoortrticicalal
issue of male (A,B) and female DS (C,D) on a low salt (LS) and high salt (HS) dididietetetss.s. CCCYPYPYP272727A1A1A1
s sstatatainininededed bbbrororownwnn.. EE,E Statistical analysis of individdduuual ll images of adrenocooortrtrtici al CYP27A1
abababuunundance. Greeey yy bbbarsrsrs repepeprerereseseentntnt CCCYPYPYP272727AAA1 aabububundaaanccce aaattt a a lololowww saaltl iiintntakakake e in mmmalalaless aaandndnd fffememmalalalesese ,,,
blbllacacack kk bars repepeprer ssesennnt CCCYYYP27A1A1A1 abuundndndanannccce attt aaa higghgh salallttt iinintaaakee in bbooth mmmallless aannnd femmmaaaleees. Eachchch
bar represents meansnsns ±±± SEM ooof f f tht e pepp rccentagegg of f f brbb own ststtaiaiain nn ofo adrenennalalal cortex for CYP27A1
from the total adrenooco trtiicic lall arerea;a; 4-555 slililidddes ffof aadrdrd enococortitiicalll titissuee fffrorom m eaea hch of 6 rats pper
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from

Wen Wei, Courtney A. Marshall, Edward G. Lakatta and Alexei Y. BagrovOlga V. Fedorova, Valentina I. Zernetkina, Victoria Y. Shilova, Yulia N. Grigorova, Ondrej Juhasz,
Human Cardiovascular Diseases, Is Initiated by CYP27A1 via Bile Acid PathwaySynthesis of an Endogenous Steroidal Na Pump Inhibitor Marinobufagenin, Implicated in
Print ISSN: 1942-325X. Online ISSN: 1942-3268 Copyright © 2015 American Heart Association, Inc. All rights reserved.
TX 75231is published by the American Heart Association, 7272 Greenville Avenue, Dallas,Circulation: Cardiovascular Genetics
published online September 15, 2015;Circ Cardiovasc Genet.
http://circgenetics.ahajournals.org/content/early/2015/09/15/CIRCGENETICS.115.001217World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circgenetics.ahajournals.org//subscriptions/
is online at: Circulation: Cardiovascular Genetics Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further information aboutnot the Editorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center,Circulation: Cardiovascular Genetics Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on May 19, 2018
http://circgenetics.ahajournals.org/D
ownloaded from








![A novel endogenous inhibitor of the secreted streptococcal ... · itis, impetigo) to life threatening (toxic shock syndrome, necrotizing fasciitis) [4]. The contribution that any](https://img.pdfslide.net/doc/110x75/5c9bc45d09d3f206138bc209/a-novel-endogenous-inhibitor-of-the-secreted-streptococcal-itis-impetigo.jpg)




















