Embed Size (px)
Citation preview

lable at ScienceDirect
Progress in Biophysics and Molecular Biology 107 (2011) 60e73
Contents lists avai
Progress in Biophysics and Molecular Biology
journal homepage: www.elsevier .com/locate/pbiomolbio
Original Research
Systematic characterization of the ionic basis of rabbit cellular electrophysiologyusing two ventricular models
Lucía Romero a, Beatriz Carbonell a, Beatriz Trenor a, Blanca Rodríguez b, Javier Saiz a, José M. Ferrero a,*
a Instituto de Investigación Interuniversitario en Bioingeniería y Tecnología Orientada al Ser Humano (I3BH), Universitat Politècnica de València (UPV), camino de Vera s/n,46022 Valencia, SpainbDepartment of Computer Science, University of Oxford, Wolfson Building, Parks Road, OX1 3QD, UK
a r t i c l e i n f o
Article history:Available online 3 July 2011
Keywords:Biological variabilityIonic currentModel validationArrhythmiaComputer simulation
* Corresponding author. Tel.: þ34 96 3877007x760E-mail addresses: [email protected] (L. Rome
(B. Carbonell), [email protected] (B. Trenor),(B. Rodríguez), [email protected] (J. Saiz), cferrero@g
0079-6107/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.pbiomolbio.2011.06.012
a b s t r a c t
Several mathematical models of rabbit ventricular action potential (AP) have been proposed to inves-tigate mechanisms of arrhythmias and excitation-contraction coupling. Our study aims at systematicallycharacterizing how ionic current properties modulate the main cellular biomarkers of arrhythmic riskusing two widely-used rabbit ventricular models, and comparing simulation results using the twomodels with experimental data available for rabbit. A sensitivity analysis of AP properties, Ca2þ and Naþ
dynamics, and their rate dependence to variations (�15% and �30%) in the main transmembrane currentconductances and kinetics was performed using the Shannon et al. (2004) and the Mahajan et al.(2008a,b) AP rabbit models. The effects of severe transmembrane current blocks (up to 100%) onsteady-state AP and calcium transients, and AP duration (APD) restitution curves were also simulatedusing both models. Our simulations show that, in both virtual rabbit cardiomyocytes, APD is significantlymodified by most repolarization currents, AP triangulation is regulated mostly by the inward rectifier Kþ
current (IK1) whereas APD rate adaptation as well as [Naþ]i rate dependence is influenced by the Naþ/Kþ
pump current (INaK). In addition, steady-state [Ca2þ]i levels, APD restitution properties and [Ca2þ]i ratedependence are strongly dependent on INaK, the L-Type Ca2þ current (ICaL) and the Naþ/Ca2þ exchangercurrent (INaCa), although the relative role of these currents is markedly model dependent. Furthermore,our results show that simulations using both models agree with many experimentally-reported elec-trophysiological characteristics. However, our study shows that the Shannon et al. model mimics rabbitelectrophysiology more accurately at normal pacing rates, whereas Mahajan et al. model behaves moreappropriately at faster rates. Our results reinforce the usefulness of sensitivity analysis for furtherunderstanding of cellular electrophysiology and validation of cardiac AP models.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
In the last years, some cellular electrophysiological propertiesrelated to action potential (AP) shape and duration and their ratedependence have been proposed in the scientific literature aspotential biomarkers of arrhythmic risk (Corrias et al., 2010;Lawrence et al., 2005). AP duration (APD) prolongation is the mainelectrophysiological property used for the preclinical assessment ofdrug cardiotoxicity and its predictive power is based on the existenceof clinical studies relating Torsade de Pointes episodes to theprolongationof theQT interval (YapandCamm,2003).More recently,
37; fax: þ34 96 3877093.ro), [email protected]@cs.ox.ac.ukbio.i3bh.es (J. M. Ferrero).
All rights reserved.
additional electrophysiological properties have been considered toovercome the limitations of this property to predict pro-arrhythmicrisk. Indeed, severalexperimentshave reported thathigher incidenceof early-afterdepolarizations is related to increased AP triangulation(Lawrence et al., 2005). Furthermore, experimental evidence showsthe important role played by intracellular sodium and calciumdynamics (Bers and Despa, 2006) in arrhythmia generation. Alter-ations in rate dependent properties have also been related to theoccurrence of alternans and the development of potentially lethalarrhythmias (Nolasco and Dahlen, 1968; Pastore et al., 2006; Pueyoet al., 2004, 2010; Watanabe et al., 2001). However, the preclinicaluseof thesebiomarkers topredict arrhythmic risk ishamperedby thebiological variability and complexity of the cellular electrophysio-logical mechanisms (Romero et al., 2009).
Mechanisms of ventricular electrophysiology, arrhythmias andexcitation-contraction coupling are often investigated using rabbits,due to its similarities with humans in terms of effective heart size

L. Romero et al. / Progress in Biophysics and Molecular Biology 107 (2011) 60e73 61
(Panfilov, 2006), repolarization reserve (Jost et al., 2007; Virag et al.,2001), calcium handling (Bers, 2002) and positive force frequencyrelationship (Maier et al., 2000). Thewealth of informationprovidedby those studies has been key in the development of mathematicalmodelsof the rabbit ventricularactionpotential (Aslanidi et al., 2010;Mahajan et al., 2008b; Puglisi and Bers, 2001; Shannon et al., 2004).In 2004, Shannon et al. (2004) proposed a rabbit ventricular APmodel with a novel mathematical formulation for calcium homeo-stasis. Most of the data used to build the calcium dynamics modelwere obtained from rabbit cardiomyocytes. However, the formula-tions of most sodium and potassium transmembrane currents werebased on the Luo and Rudy (1994) guinea pig AP model, with theexception of Ito based on rabbit experimental data by Bassani et al.(2004). Some of these currents were also scaled to reproduce theeffectsof IKr and IKs blockersonAPDasmeasured in rabbit ventricularcardiomyocytes (Shannon et al., 2004). More recently, Mahajan et al.(2008b) published a new rabbit model based on the Shannon et al.model but with updated formulations for L-type Ca2þ current (ICaL)and calcium cycling. Themain aimwas to correctly reproduce rabbitventricular APD restitution and calcium transient alternans at rapidheart rates. More recently, Aslanidi et al. (2010) have published fourmodified versions of the Shannon et al. model corresponding torabbit ventricular myocytes from different locations in the ventric-ular wall and to the rabbit Purkinje fibers in order to investigate APDheterogeneity at the Purkinje-ventricular junction.
The different rabbit AP models produced to date have beenformulated building on the increasing knowledge on rabbit ventric-ularelectrophysiologygenerated in recentyears. Theyhavebeenusedin the investigation of different and specific scientific questions,including but not limited to the mechanisms of the cardiac biologicalpacemaker generation in IK1-downregulated cardiac myocytes(Kurata et al., 2008), the stochastic and deterministic aspects ofintracellularcalciumdynamicsduringalternans (RestrepoandKarma,2009), the arrhythmogenic role of abnormal calcium handling inmany pathological settings (Restrepo et al., 2008; Xie et al., 2008), theorigin of cardiac alternans induced by fibroblast-myocyte coupling(Xie et al., 2009), the anti-fibrillatory effects of altering ICaL kinetics(Mahajan et al., 2008a), the pharmacological restoration of inactiva-tion of ICaL in the Timothy syndromemutation (Yarotskyyet al., 2009),the synchronization of chaotic early-afterdepolarizations in thegenesis of cardiac arrhythmias (Sato et al., 2009), the antiarrhythmicbenefits of targeting theNaþ/Ca2þ exchanger (Anumonwo, 2008) andthe electrical heterogeneity between purkinje fibers and ventricularmyocytes (Aslanidi et al., 2010).
The proliferation of cardiac AP models during the past decadehas led to a number of publications focussed on AP modelscomparison and analysis (Bueno-Orovio et al., 2008; Cherry et al.,2008; Cherry and Evans, 2008; Cherry and Fenton, 2004, 2007;Clayton et al., 2011; Fink et al., 2011; Nygren et al., 2001; TenTusscher et al., 2006; Wilders, 2007). Interestingly, it has beenshown that different ventricular APmodels of the same species mayexhibit different electrophysiological characteristics (Bueno-Orovioet al., 2008; Cherry et al., 2008; Cherry and Evans, 2008; Cherryand Fenton, 2004, 2007; Clayton et al., 2011; Fink et al., 2011; TenTusscher et al., 2006; Wilders, 2007). In the present study, weperform a systematic analysis of the sensitivity of the main cellularbiomarkers of arrhythmic risk in two rabbit models to variations of�15e30% in ionic current properties, similarly to our previous workin a human model (Romero et al., 2009). We specifically focus ontwo widely-used models, namely Shannon et al. (2004) and theMahajan et al. (2008b) models. Simulation results using bothmodels are compared between them and also to experimental dataobtained in rabbit. As drugs are usually applied at varying doses inexperimental studies, we also investigated the effects of severedegrees of current blocks up to 100% on electrophysiological
properties such as steady-state and restitution properties of AP, andcalcium transients. The systematic and comprehensive analysisperformed in this study provides new insights into the ionic basis ofthe main preclinical biomarkers of arrhythmic risk in rabbits, an in-depth characterization of the Shannon et al. and Mahajan et al.rabbit models and a comparison between both models and exper-imental findings. In addition, the analysis identifies knowledge gapsand controversies on the mechanismsmodulating rabbit ventricularelectrophysiology, which could suggest new experiments to furtherour understanding of rabbit ventricular electrophysiology.
2. Methods
2.1. Action potential models
The electrophysiological activity of an isolated rabbit ventricularmyocyte was simulated using the Shannon et al. (2004) and theMahajan et al. (2008b) ventricular AP models. The main differencesbetween these two models lie on the calcium cycling and the ICaLformulations, including the Naþ/Ca2þ exchanger (NCX) calciumdependence. The remaining transmembrane currents formulationsof the Mahajan et al. (2008b) model are based on the Shannon et al.model, except for the fact that ionic channels are no longerassumed to be distributed homogeneously throughout the cellmembrane and several current parameters were modified to fitexperimentally observed single cell dynamics.
2.2. Stimulation protocols
As in Romero et al. (2009), five protocols were applied in thesimulations to characterize the main cellular arrhythmic riskbiomarkers using the two rabbit models (Fig. 1).
2.2.1. Steady-state AP and [Ca2þ]i propertiesAPD, AP triangulation and systolic [Ca2þ]i were calculated from
the simulations following application of a train of 1000 squaretransmembrane current pulses of 2 ms duration and twice diastolicthreshold at a basic cycle length (BCL) of 400 ms, which is thenormal sinus pacing rate in rabbits. This protocol allowed modelvariables to reach steady-state, except for the cases of 50% INaKblock and 70% INaCa block, where 2000 pulses were required.Steady-state APD at 90% and 60% repolarization (APD90 and APD60)and AP triangulation (defined as the difference between APD60 andAPD90) were calculated (Fig. 1A, top panel).
2.2.2. APD restitution curvesBoth S1S2 and dynamic APD90 restitution curves were obtained
using the two models. The S1S2 restitution protocol was appliedafter the virtual cell reached steady-state and consisted of a train of10 square current pulses (S1) at a BCL of 400 ms, followed by anextra-stimulus (S2) applied at coupling intervals (CIs) ranging from5000 ms to 50 ms with a CI step of 1 ms. The S1S2 restitution curvewas generated by plotting the APD90 elicited by S2 versus thediastolic interval (DI) for each S2 (Fig. 1B, top panel). The dynamicrestitution curve, which is used to characterize drug reverse ratedependence (Guo et al., 2008), describes the dependence of thesteady-state APD90 on the DI for BCLs ranging from 1000 ms to50 ms (Fig. 1B, bottom panel). DIs were calculated as the differencebetween BCL and APD90 for every simulated BCL. The maximalslopes of both restitution curves (slopemax,S1S2 and slopemax,DYN)were calculated as in Romero et al. (2009).
2.2.3. APD rate adaptation to abrupt changes in BCLIn order to analyze the APD90 rate adaptation, the virtual rabbit
cardiomyocyte was paced at a BCL of 1000 ms for 8 min, then at

Fig. 1. Simulated electrophysiological activity of a rabbit ventricular cardiomyocyte using the Shannon et al. model (thick lines) and the Mahajan et al. model (thin lines). A: Timecourse of the steady-state action potential (top) and [Ca2þ]i transient (bottom) at 2.5 Hz. APD60 and APD90 refer to APD at 60% and 90% repolarization, respectively. Triangulation wasdefined as the interval between APD60 and APD90. B: S1S2 (top) and dynamic (bottom) APD restitution curves. C: APD heart rate adaptation after sudden changes in cycle length (CL).D: Normalized systolic [Ca2þ]i (top) and [Naþ]i (bottom) relative to value at 0.25 Hz as a function of the pacing frequency.
L. Romero et al. / Progress in Biophysics and Molecular Biology 107 (2011) 60e7362
a BCL of 600 ms for 8 min and finally the BCL was set back to1000 ms for 8 min. The slow time constant (sslow) of the temporalAPD90 adaptation to the accelerating rate was computed (Pueyoet al., 2010; Romero et al., 2009) (Fig. 1C).
2.2.4. Rate dependence of [Naþ]i and [Ca2þ]iVentricular myocytes were paced at increasingly fast frequen-
cies (0.25, 0.5, 1, 1.5, 2, 2.5 and 3 Hz) for 10 min and systolic [Ca2þ]iand [Naþ]i levels were monitored for each frequency and normal-ized to the respective values at 0.25 Hz (Fig. 1D).
2.3. Sensitivity analysis
Investigation into the impact of alterations in ionic currentproperties in the modulation of preclinical biomarkers ofarrhythmic risk was carried out by performing a sensitivity study,
as in our previous study (Romero et al., 2009). The ionic currentproperties considered in this workwere the maximal conductancesand time constants of the main transmembrane ionic currents,namely, maximal conductance (GNa) of the fast sodium current(INa), activation gate time constant (sm) and inactivation gate timeconstants (sh and sj); maximal conductance (GCaL) of the L-typeCa2þ current (ICaL), activation gate time constants (sd in the Shan-non et al. model) and transition rates involved in channel opening(r1 and r2 and a and b in the Mahajan et al. model), as well as itsvoltage-dependent inactivation time constant (sf) in the Shannonet al. model and transition rates involved in inactivation (s10 and s20,k30 and k40, k50 and k60 together with k5 and k6) in the Mahajan et al.model; maximal conductance (GKr), activation (sXr) and inactiva-tion gate time constant (sRr in Shannon) of the rapid component ofthe delayed rectifier current (IKr); maximal current conductance(GKs) and activation time constants (two sXs in the Shannon et al.

L. Romero et al. / Progress in Biophysics and Molecular Biology 107 (2011) 60e73 63
model and sXs1 and sXs2 in the Mahajan et al. model) of the slowcomponent of the delayed rectifier current (IKs); maximal conduc-tance (Gto,f) and activation and inactivation gate time constant(sXto,f and sYto,f, respectively) of the fast component of the transientoutward potassium current (Ito,f); maximal conductance (Gto,s) andactivation and inactivation gate time constants (sXto,s, sYto,s andsRto,s respectively) of the slow component of the transient outwardpotassium current (Ito,s); maximal conductance of the inwardrectifier Kþ current (IK1), maximal activity of Naþ/Kþ pump current(INaK) and Naþ/Ca2þ exchanger (NCX) current (INaCa), namely GK1,GNaK and GNaCa respectively.
The sensitivity analysis was performed by varying one param-eter at a time, by �30%, �15%, þ15% and þ30%, respectively, as inour previous study (Romero et al., 2009), being the magnitude ofthese modifications in line with natural variability reportedexperimentally in rabbit ventricular tissues (Sims et al., 2008).Simulations using the five stimulation protocols described in theprevious section were run for each parameter data set and resultswere compared with experimental studies performed in rabbitventricular myocytes available in the literature.
In addition, severe channel blocks up to 100% were also simu-lated using both models. To determine the maximum degree ofblock, we looked for the maximum percentages of current blockreported in experimental works. 100% block of IKr, IKs and Ito can beforced in rabbit myocytes by adding 100 nM E-4031, 100 nM L-735,821 and 3 mM 4-aminopyridine (Lengyel et al., 2001; Szigligetiet al., 1998), respectively. To the best of our knowledge, themaximum INaCa block studied in rabbit is 70% approximately, whichis provoked by 1.0 mM SEA0400 (Farkas et al., 2008). In addition,10�5 M BaCl2 can be added in rabbit myocytes to selectively blockIK1, a concentration that does not block IKr, but blocks IK1 currentsover 70% (Lu et al., 2008). Unfortunately, maximum percentages ofICaL and INaK block are not easily found in the literature. Indeed,most papers provide the drug concentration that is applied withoutmentioning the percentage of block that is achieved. Simulatedcurrent reductions corresponded to complete IKr and Ito block, 70%block for IK1 and INaCa, 60% block for ICaL and 50% block for INaK.Finally, IKs was completely blocked in the Shannon et al. model,while only 80% block was possible in the Mahajan et al. model, asfurther IKs reductions resulted in repolarization failure in the latter
Fig. 2. Comparison of the relative sensitivities of the electrophysiological characteristics (firs(right triangles) and the Mahajan et al. (left triangles) models depicted in a gray color scalecharacteristic among all model parameters, while black indicates that characteristic andmaximum absolute sensitivity of the characteristic correspondent to that row and negative
model. These block percentages are close to the maximum currentreductions that can be simulated without producing artifacts in thesimulation results and they are also in the range of severe blocksreported experimentally (Farkas et al., 2008; Lengyel et al., 2001;Ranu et al., 2002; Schillinger et al. 2003; Szigligeti et al., 1998;Tolkacheva et al. 2006). A total of 33,864 simulations was run.
2.4. Data analysis
For each electrophysiological characteristic ‘c’ and modelparameter ‘p’, the indexes percentage of change (Dc,p,a), sensitivity(Sc,p) and relative sensitivity (rc,p) were calculated as
Dc;p;a ¼�cp;a � ccontrol
�
ccontrol$100 (1)
Sc;p ¼ DDc;p;a
Da¼ Dc;p;þ30% � Dc;p;�30%
0:6(2)
rc;p ¼ Sc;pjSc;pjmax;c
(3)
with cp,a and ccontrol being the magnitude of the electrophysiologicalcharacteristic ‘c’when the parameter ‘p’ is increased by ‘a’ and undercontrol conditions, respectively and jSc;pjmax;c being the maximumabsolute value among all the sensitivities obtained for the charac-teristic ‘c’. When a characteristic showed a biphasic percentage ofchange with a certain model parameter, the Sc,p was calculated as
Sc;p ¼ DDc;p;a
Da¼ �Dc;p;�30%
0:3(4)
3. Results
Fig. 2 shows the sensitivity of electrophysiological characteristics(biomarkers) (first column) to changes in parameter values (toprow) for each of the twomodels. Left and right triangles of each boxshow results for the Mahajan et al. and the Shannon et al. models,respectively. Results corresponding toparameters suchasGNa, sm, sh,sj, r1 and r2, a and b, s10 and s20, k30 and k40, sXr, sRr, sXto,f, Gto,s, sXro,s,sYto,s and sRto,s exert a negligible effect on repolarization-related
t column) to variations in ionic current properties (first row) between the Shannon et al.. White indicates the maximum relative sensitivity of a particular electrophysiologicalparameter are completely independent. Percents in each white triangle indicate thesign indicates that characteristic and parameter vary inversely.

L. Romero et al. / Progress in Biophysics and Molecular Biology 107 (2011) 60e7364
electrophysiological characteristics and thus are not shown. A greyscale is used in Fig. 2: the strongest relative sensitivities are depictedin white, whereas the lack of relationship between parameter andcharacteristic is shown in black. Negative sensitivities are markedwith a negative sign. For each electrophysiological property, themaximum absolute sensitivity among all parameters is also indi-cated as a percent value. In addition, Fig. 3 illustrates the time courseof the steady-state AP for control and for 30% reduction of indicatedparameter value for Shannon et al. (top panels) and Mahajan et al.(bottom panels) models. Only AP waveforms significantly differentfrom control AP waveforms were depicted.
Furthermore, Figs. 4e6 show, for both models, the effect of �15%and �30% changes in parameter values on steady-state APD, APtriangulation and systolic [Ca2þ]i (Fig. 4), maximum slopes of APDrestitutioncurves and slow timeconstantofAPDadaptation to abruptchanges inpacing rate (Fig. 5) and [Ca2þ]i and [Naþ]i rate dependence(Fig. 6). Results for the Shannon et al. and the Mahajan et al. modelsare shown in the left and right panels of eachfigure, respectively. Theelectrophysiologically-reported range of each property is delimitedby dotted horizontal lines in the corresponding panels, when dataunder normal conditions are available in the literature.
Finally, Figs. 7 and 8 depict the time course of steady-state APsand calcium transients, as well as APD restitution curves for controland under severe current blocks using both models, respectively.Only the cases for which calcium transients were significantlydifferent from control were depicted.
In the following sections, our results are further described anddiscussed.
3.1. Steady-state AP properties
3.1.1. Action potential durationSimulated APD90 in control are 177 ms and 185 ms for the
Shannon et al. and the Mahajan et al. models, respectively. Asshown in Fig. 4, A and B, simulated APD90 are within the electro-physiological range observed in rabbit ventricular tissues (Biagettiand Quinteiro, 2006; Szigligeti et al., 1996; Yan et al., 2001),ranging from 123 ms to 190 ms. Such a wide electrophysiologicalrange raises from accounting for a wide variety of experimentalconditions that modify the APD, such as animal gender, age and
Fig. 3. Time course of the simulated rabbit action potentials (top panels and bottom panels,solid line) and for 30% reduction in parameters exerting a relevant effect on membrane potenlong dashed line, only Mahajan et al.); B and F: GKs (thin long dashed line, only Mahajan et aldotted line, only Mahajan et al.); C and G: Gto,s (thin long dashed line, only Shannon et al.), Gto,f
solid line) and GNaCa (thin long dashed line), as labeled next to each curve.
location of the myocyte (transmural and distance to the apex).Indeed, several experiments performed in rabbit tissue at 2.5 Hzhave shown APD differences up to 30 ms and 10 ms approximatelydepending on the transmural location (Biagetti and Quinteiro,2006; Yan et al., 2001) and on age (Idriss and Wolf, 2004), as wellas on sex (Biagetti and Quinteiro, 2006), respectively. As shown inFig. 2, 1st row, Fig. 3 and Fig. 4, A and B, APD90 is moderatelysensitive to changes in many repolarization current parameterswith maximum sensitivity of 45% obtained for sf in the Shannonet al. model and 33% for GNaK in the Mahajan et al. model. Signifi-cant differences between both models exist in the sensitivity ofAPD90 to changes in most parameters, as shown in Fig. 2, 1st row.
In both models, decrease in GNaK results in APD90 shortening(Fig. 2, 1st row, Fig. 4, A and B and Fig. 7, B and F), which is consistentwith experimental recordings in rabbit exposed to sodium potassiumpump inhibitors, such as ouabain (Kodama et al., 1981; Sanchez-Serrano et al., 1980) and acetylstrophanthidin (Bers et al., 1988). Ourresults also show that GCaL reduction also provokes APD90 shorteningwhen using the Shannon et al. model and when simulating severeblocks in the Mahajan et al. model (Fig. 2, 1st row, Fig. 4, A and B andFig. 7, A and E). Simulation results are also consistent with the pro-longed APDs reported experimentally in rabbit myocytes with highL-type calcium channel density (Sims et al., 2008). However, GCaL
variations up to �30% cause a slight prolongation of the APD90 whenusing the Mahajan et al. model. Experiments performed in rabbitventricles have traditionally reported APD shortening after theapplication of ICaL blockers, such as verapamil (Mahajan et al., 2008b;Wu et al., 2002), Bay-K 8644 (Lu et al., 2008), nisoldipine (Tolkachevaet al., 2006) and nifedipine (Bers et al., 1988). The prolongation in APDcaused by 15% and 30% ICaL blockwhen using theMahajan et al. modelis due to a significant reduction in INaK and IKs provoked by alterationsin intracellular concentrations, which counteract the effect of ICaLreduction in the APD90.
In both models, a decrease in INaCa results in APD90 shortening.This effect is however more prominent in the Shannon et al. modelthan in theMahajan et al.model (Fig. 2,1st row, and Fig. 3B and Fig. 7,B and F). To the best of our knowledge, no rabbit experimentssupport the APD90 prolongation observed in our simulations whenGNaCa is reduced. On the contrary, APD prolongation following INaCablock has been reported in some rabbit experiments (Ranu et al.,
Shannon et al. and Mahajan et al. models, respectively) under control conditions (thicktial. A and E: GCaL (thin solid line), sf (thin long dashed line, only Shannon et al.), sm (thin.), GKr (thin long dashed line, only Shannon et al.), GK1 (thin solid line), sXs1 (thin dashed(thin solid), and sYto,f (thin long dashed line, only Mahajan et al.); and D and H: GNaK (thin
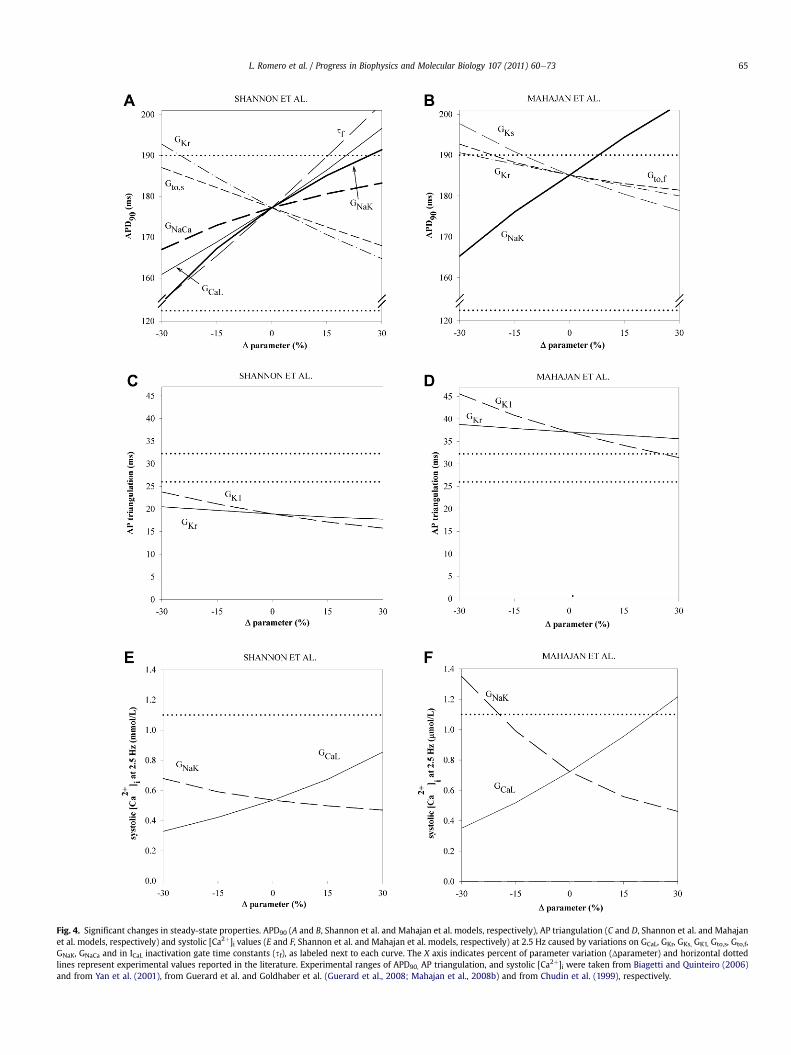
Fig. 4. Significant changes in steady-state properties. APD90 (A and B, Shannon et al. and Mahajan et al. models, respectively), AP triangulation (C and D, Shannon et al. and Mahajanet al. models, respectively) and systolic [Ca2þ]i values (E and F, Shannon et al. and Mahajan et al. models, respectively) at 2.5 Hz caused by variations on GCaL, GKr, GKs, GK1, Gto,s, Gto,f,GNaK, GNaCa and in ICaL inactivation gate time constants (sf), as labeled next to each curve. The X axis indicates percent of parameter variation (Δparameter) and horizontal dottedlines represent experimental values reported in the literature. Experimental ranges of APD90, AP triangulation, and systolic [Ca2þ]i were taken from Biagetti and Quinteiro (2006)and from Yan et al. (2001), from Guerard et al. and Goldhaber et al. (Guerard et al., 2008; Mahajan et al., 2008b) and from Chudin et al. (1999), respectively.
L. Romero et al. / Progress in Biophysics and Molecular Biology 107 (2011) 60e73 65

Fig. 5. Significant changes in restitution curves slopes and in the slow time constant of APD accommodation to abrupt changes on pacing rate. Slopemax,S1S2 (A and B, Shannon et al.and Mahajan et al. models, respectively), slopemax,DYN (C and D, Shannon et al. and Mahajan et al. models, respectively) and sslow (E and F, Shannon et al. and Mahajan et al. models,respectively) with changes in GCaL, sf, GKr, GK1, Gto,f, sYto,f, GNaK and GNaCa, as labeled next to each curve. The X axis indicates percent of parameter variation (Δparameter) andhorizontal dotted lines represent experimental values reported in the literature (Bell et al., 2007; Goldhaber et al., 2005; Mahajan et al., 2008b; Pitruzzello et al., 2007).
L. Romero et al. / Progress in Biophysics and Molecular Biology 107 (2011) 60e7366

Fig. 6. Significant changes in [Ca2þ]i and [Naþ]i rate dependence. Maximum systolic [Ca2þ]i (A and B, Shannon et al. and Mahajan et al. models, respectively) and maximum systolic[Naþ]i (C and D, Shannon et al. and Mahajan et al. models, respectively) with changes in GCaL, sf, GKs, sXs1, GNaK and GNaCa, as labeled next to each curve. The X axis indicates percent ofparameter variation (Δparameter) and horizontal dotted lines represent experimental values reported in the literature (Baartscheer et al., 2003;Maier et al., 2000)
Fig. 7. Simulated rabbit steady-state AP and steady-state calcium transients waveforms (top panels and bottom panels, Shannon et al. and Mahajan et al. models, respectively) undercontrol conditions (thick solid line) and for severe transmembrane current blocks as labeled next to each curve. A and E: action potentials for 100% block of IKr (0�GKr) and IKs(0�GKs) for the Mahajan et al. model and 80% block for the Shannon et al. model (0.2�GKs) and 60% block of ICaL (0.4�GCaL); B and F: action potentials for 100% block of Ito(0�Gto), 70% block of IK1 (0.3�GK1) and INaCa (0.3�GNaCa) and 50% block of GNaK (0.5�GNaK); C and G, and D and H calcium transients for the same current blocks as panels A and E,and B and F, respectively.
L. Romero et al. / Progress in Biophysics and Molecular Biology 107 (2011) 60e73 67
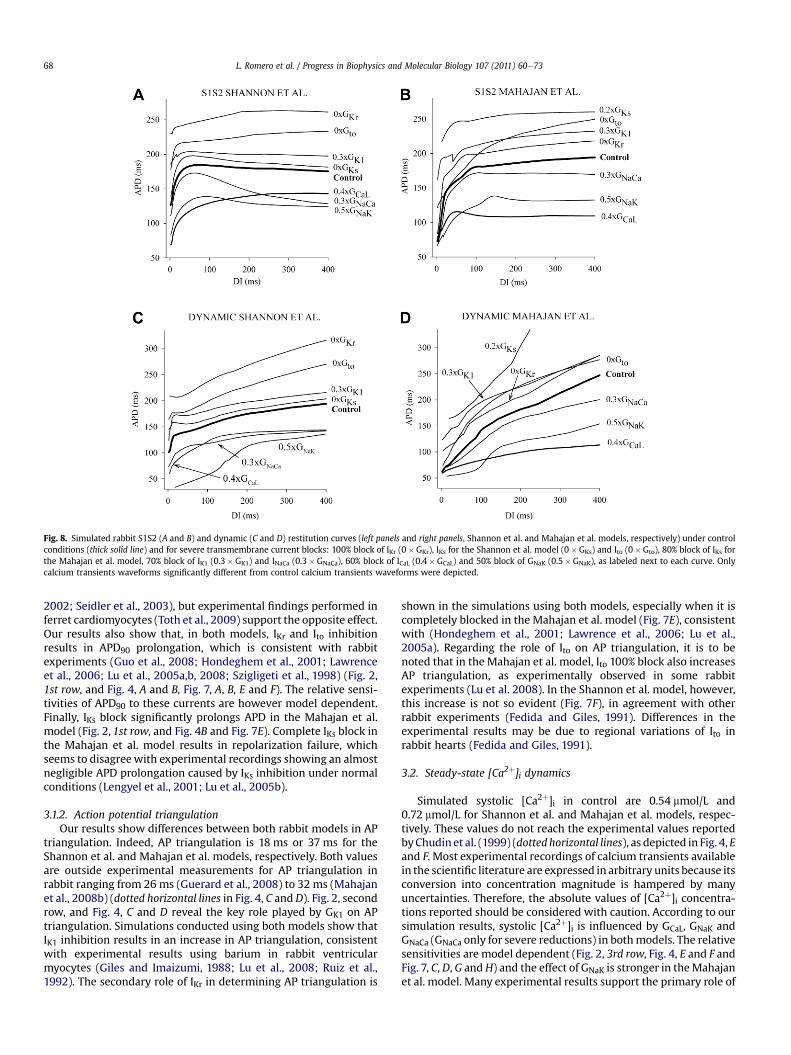
Fig. 8. Simulated rabbit S1S2 (A and B) and dynamic (C and D) restitution curves (left panels and right panels, Shannon et al. and Mahajan et al. models, respectively) under controlconditions (thick solid line) and for severe transmembrane current blocks: 100% block of IKr (0�GKr), IKs for the Shannon et al. model (0�GKs) and Ito (0�Gto), 80% block of IKs forthe Mahajan et al. model, 70% block of IK1 (0.3�GK1) and INaCa (0.3�GNaCa), 60% block of ICaL (0.4�GCaL) and 50% block of GNaK (0.5�GNaK), as labeled next to each curve. Onlycalcium transients waveforms significantly different from control calcium transients waveforms were depicted.
L. Romero et al. / Progress in Biophysics and Molecular Biology 107 (2011) 60e7368
2002; Seidler et al., 2003), but experimental findings performed inferret cardiomyocytes (Toth et al., 2009) support the opposite effect.Our results also show that, in both models, IKr and Ito inhibitionresults in APD90 prolongation, which is consistent with rabbitexperiments (Guo et al., 2008; Hondeghem et al., 2001; Lawrenceet al., 2006; Lu et al., 2005a,b, 2008; Szigligeti et al., 1998) (Fig. 2,1st row, and Fig. 4, A and B, Fig. 7, A, B, E and F). The relative sensi-tivities of APD90 to these currents are however model dependent.Finally, IKs block significantly prolongs APD in the Mahajan et al.model (Fig. 2, 1st row, and Fig. 4B and Fig. 7E). Complete IKs block inthe Mahajan et al. model results in repolarization failure, whichseems to disagreewith experimental recordings showing an almostnegligible APD prolongation caused by IKs inhibition under normalconditions (Lengyel et al., 2001; Lu et al., 2005b).
3.1.2. Action potential triangulationOur results show differences between both rabbit models in AP
triangulation. Indeed, AP triangulation is 18 ms or 37 ms for theShannon et al. and Mahajan et al. models, respectively. Both valuesare outside experimental measurements for AP triangulation inrabbit ranging from 26 ms (Guerard et al., 2008) to 32 ms (Mahajanet al., 2008b) (dotted horizontal lines in Fig. 4, C and D). Fig. 2, secondrow, and Fig. 4, C and D reveal the key role played by GK1 on APtriangulation. Simulations conducted using both models show thatIK1 inhibition results in an increase in AP triangulation, consistentwith experimental results using barium in rabbit ventricularmyocytes (Giles and Imaizumi, 1988; Lu et al., 2008; Ruiz et al.,1992). The secondary role of IKr in determining AP triangulation is
shown in the simulations using both models, especially when it iscompletely blocked in the Mahajan et al. model (Fig. 7E), consistentwith (Hondeghem et al., 2001; Lawrence et al., 2006; Lu et al.,2005a). Regarding the role of Ito on AP triangulation, it is to benoted that in the Mahajan et al. model, Ito 100% block also increasesAP triangulation, as experimentally observed in some rabbitexperiments (Lu et al. 2008). In the Shannon et al. model, however,this increase is not so evident (Fig. 7F), in agreement with otherrabbit experiments (Fedida and Giles, 1991). Differences in theexperimental results may be due to regional variations of Ito inrabbit hearts (Fedida and Giles, 1991).
3.2. Steady-state [Ca2þ]i dynamics
Simulated systolic [Ca2þ]i in control are 0.54 mmol/L and0.72 mmol/L for Shannon et al. and Mahajan et al. models, respec-tively. These values do not reach the experimental values reportedbyChudin et al. (1999) (dotted horizontal lines), as depicted in Fig. 4, Eand F. Most experimental recordings of calcium transients availablein the scientific literature are expressed in arbitrary units because itsconversion into concentration magnitude is hampered by manyuncertainties. Therefore, the absolute values of [Ca2þ]i concentra-tions reported should be considered with caution. According to oursimulation results, systolic [Ca2þ]i is influenced by GCaL, GNaK andGNaCa (GNaCa only for severe reductions) in bothmodels. The relativesensitivities are model dependent (Fig. 2, 3rd row, Fig. 4, E and F andFig. 7, C, D, G andH) and the effect of GNaK is stronger in theMahajanet al. model. Many experimental results support the primary role of

L. Romero et al. / Progress in Biophysics and Molecular Biology 107 (2011) 60e73 69
GCaL, GNaK and GNaCa on systolic [Ca2þ]i shown in our simulations.The important role played by GNaK in modulating systolic [Ca2þ]ilevels in both rabbit models is supported by the marked increase incell force of contraction and calcium levels caused by INaK inhibitors,such as ouabain (Giles and Shimoni, 1989; Kodama et al., 1981;McCans et al., 1974; Schillinger et al., 2003) and strophantidine(Loughrey et al., 2003; Ranu et al., 2002; Szigligeti et al., 1996).Furthermore, experimental results showing reduction of cellcontraction (related to systolic [Ca2þ]i) caused by the ICaL blockerverapamil (Mahajan et al., 2008a), are consistent with the impor-tance of GCaL in determining systolic [Ca2þ]i levels. Experimentalrecordings in rabbit also show that INaCa partial blockade tends toenhance the contractile force (related to the systolic [Ca2þ]i)although the impact of this modification is confusing. Indeed, somestudies report a strongeffect of INaCa inmodulating [Ca2þ]i transientsamplitude (Ranu et al., 2002; Schillinger et al., 2000; Toth et al.,2009) similarly to our simulation results for severe INaCa block(Fig. 7, D and H). However, others report only a moderate incrementof maximum systolic pressures when INaCa is severely inhibited(Farkas et al., 2008). Simulations with bothmodels showalterationsin steady-state calcium transients when Ito is completely blocked(Fig. 7, D and H), although they are markedly model dependent.Indeed, in the Mahajan et al. model the systolic [Ca2þ]i is drasticallyreduced, whereas in the Shannon et al. model this magnitude isslightly increased. Experimental evidence supports the importantrole played by Ito in modulating cardiac calcium transients throughits effects on ICaL and possibly INaCa (Sah et al., 2003). Indeed, ICaL andINaCa are influenced by the early repolarization period of the AP(phase 1), which is determined by Ito. This fact is in accordancewithrabbit experiments showing that APs with reduced overshootsdecrease ICaL amplitude (Linz and Meyer, 2000).
3.3. Rate adaptation
3.3.1. S1S2 restitution curveSimulated values of slopemax,S1S2 in control are 3.8 and 4.3 for
Shannon et al. and Mahajan et al. models, respectively. Bothcontrol slopemax,S1S2 are within the range reported experimen-tally (Goldhaber et al., 2005) (Fig. 5, A and B). However, the ionicmechanisms determining slopemax,S1S2 are different for each ofthe models: IK1, GNaK, sf, and GCaL, and GNaCa only for severereductions, are the key parameters in modulating slopemax,S1S2 inthe Shannon et al. model, whereas in the Mahajan et al. model,GNaK, GKs, sYto,f and Gto,f, together with GCaL (especially for severeblocks), all play a key role in slopemax,S1S2 (Fig. 2, 4th row, Fig. 5, Aand B and Fig. 8, A and B). Several experiments in rabbit prepa-rations support the strong influence exerted by GCaL (Tolkachevaet al., 2006;Wu et al., 2002), and Gto,f (Szigligeti et al., 1998) onslopemax,S1S2 found in the Mahajan et al. model, although theinfluence of GKs seems to be too large in this model (Bell et al.,2007). To the best of our knowledge, the effect of GK1 on S1S2restitution curves has not been studied in rabbit myocytes butthe influence of this parameter in the Shannon et al. model isconsistent with experimental studies in pigs reporting a flat-tening of the S1S2 restitution curve by bretilium (IK1 blocker)administration (Garfinkel et al., 2000). Moreover, the negligiblerole of GKr on the slopemax,S1S2 observed in both models is inaccordance with rabbit experiments (Bell et al., 2007), except forthe reduction of the slopemax,S1S2 produced by 100% IKr block inthe Shannon et al. model (Fig. 8A).
3.3.2. Dynamic restitution curveMarked differences between both rabbit models in simulated
control slopemax,DYN were found. Indeed, simulated control valuesof slopemax,DYN are 4.1 and 0.96 for Shannon and Mahajan et al.
models, respectively. As shown in Fig. 5, C and D, slopemax,DYN forcontrol is outside the experimentally-reported range for bothmodels (Mahajan et al., 2008b). However, it must be noted thatquantitative comparison of maximal restitution slopes is prob-lematic due to the strong dependency of slopemax,DYN on CI step,minimum CI and the fitting of the curves. Our results show thatslopemax,DYN strongly depends on INaK for both models and also onother parameters but in a model-dependent manner (Fig. 2, 5throw, and Fig. 5, C and D). Indeed, GKr, GNaK, sf, GKs, GNaCa, Gto,s andGCaL (GCaL only for severe blocks) also determine slopemax,DYN in theShannon et al. model whereas in the Mahajan et al. model, it isGNaK, GCaL (especially for severe blocks), Gto,f, sYto,f, GKs and GNaCa(GNaCa only for severe blocks). There is experimental evidence onthe reduction of slopemax,DYN in rabbit hearts by ICaL blockers, suchas nisoldipine (Tolkacheva et al., 2006) and verapamil (Lawrenceet al., 2006), although this reduction depends on drug concentra-tion (Lawrence et al., 2006). To the best of our knowledge, no rabbitexperiments have been conducted to study the role of INaCa ondynamic restitution curves. However, flattening of the dynamicrestitution curve by INaCa inhibition has also been experimentallyreported in ferret ventricular myocytes (Janvier and Boyett, 1996),which is consistent with our results using both models. Theincrease in slopemax,DYN caused by GKs inhibition is also supportedby experiments performed on rabbit tissues perfused withchromanol-293b (Bell et al., 2007). However, other experimentsreport little effect of IKs block on APD at a wide range of stimulationfrequencies (Lengyel et al., 2001; Lu et al., 2005b). Finally, theincrement of the slopemax,DYN caused by GKr inhibition with theShannon et al. model is in accordance with the reverse frequencydependence exerted by IKr blockers in experiments (Bell et al.,2007; Eckardt et al., 2002; Lawrence et al., 2006; Lengyel et al.,2001). However, Lu et al. (2005a) observed that, in rabbit papil-lary muscles, dofetilide slightly prolonged APD90 at normal heartrates whereas in a condition of tachycardia it prolonged APD90,which is in contrast of most experimental results.
3.3.3. APD adaptation to sudden sustained changesin stimulation rate
Control values of sslow are 52 s and 84 s for Shannon et al. andMahajan et al. models, respectively. sslow values are moderately (forShannon et al. model) or significantly (for Mahajan et al. model)longer than the ones observed experimentally (Bell et al., 2007;Pitruzzello et al., 2007) (Fig. 5, E and F). Fig. 2, 6th row, and in Fig. 5,E and F reveal the critical influence of GNaK on sslow. To the best of ourknowledge, the effect of GNaK on sslow has not been experimentallystudied in rabbit cardiomyocytes, but it has been reported for humanand canine ventricular cells (Boyett and Fedida, 1984; Pueyo et al.,2010). In the Shannon et al. model, sslow is also affected by changesin GCaL, consistent with rabbit experiments (Tolkacheva et al., 2006).
3.3.4. Rate dependence of ionic concentrationsMaximum systolic [Ca2þ]i normalized to the value at 0.25 Hz
was 240% and 527% for Shannon et al. and Mahajan et al. models,respectively, being both values in line with experiments performedin rabbit myocytes (Maier et al., 2000) (Fig. 6, A and B, dottedhorizontal lines). However, the dependency of systolic [Ca2þ]i withfrequency is stronger in theMahajan et al. than in the Shannon et al.model. Fig. 2, 7th row shows that frequency dependence of systolic[Ca2þ]i is highly sensitive to INaK, and ICaL in bothmodels and INaCa inthe Shannon et al. model. Unfortunately, conflicting experimentalresults about the effect of the INaK blocker ouabain on rabbitcontraction force at different frequencies have been obtained,especially at high drug concentrations (Koch-Weser and Blinks,1963; Komai et al., 1996). However, higher force increments areusually reported at lower frequencies (Koch-Weser and Blinks,

L. Romero et al. / Progress in Biophysics and Molecular Biology 107 (2011) 60e7370
1963; Komai et al., 1996), which is in contrast with the resultsobtained with both models. The influence of ICaL on the frequencydependence of systolic [Ca2þ]i is also consistent with experimentsperformed in rabbit ventricular preparations (McCans et al., 1974).The increase in the frequency dependence of systolic [Ca2þ]i withINaCa overexpression is in accordance with some experimentsreported in adult rabbit myocytes (Ranu et al., 2002). Othershowever report the opposite effect (Schillinger et al., 2000).
Themaximumvalue of [Naþ]i normalized to the value at 0.25 Hzin control is 164% and 881% for the Shannon et al. andMahajan et al.models, respectively. The value obtained in the Shannon model isvery close to the experimental modulation observed by Baartscheeret al. (2003) but in theMahajan et al. model it is too large. Fig. 2, lastrow, and Fig. 6, C and F reveal the importance of INaK in modulatingthe frequency dependence of [Naþ]i in both models, followed by IKs(GKs and sXs1) only in the Mahajan et al. model.
4. Discussion
4.1. Major findings
In the present study, a systematic analysis was performed tocharacterize the role of ionic current properties in modulatingcellular electrophysiological biomarkers in two widely-used rabbitAPmodels. Simulation results were compared to experimental dataobtained in rabbit for normal conditions and following pharma-cological or protein over-expression interventions. Our simulationsshow that both models correctly reproduce many electrophysio-logical characteristics, but some of them are out of experimentalrange. In addition, several differences in the output of the twomodels in terms of the mechanisms regulating each of thebiomarkers were identified. These discrepancies could be due tothe high variability in the experimental recordings used to build themodels possibly caused by sex, age, and location differences inrabbit electrophysiology or variations in the experimental condi-tions used in each laboratory. They could however also be due toa necessarily incomplete knowledge of rabbit electrophysiologyand the need for better model building techniques and procedures.
Comparison of our simulation results for the two rabbit modelswith our previous findings using a similar analysis on a humanmodel (Romero et al., 2009) provides novel insights into the inter-species differences that should be considered when extrapolatingrabbit investigations to humans. Our study also supports the needto perform sensitivity analysis on models of ventricular electro-physiology and to compare to experimental data for in-depthquantitative characterization and validation of the models, ashighlighted in previous studies (Nygren et al., 1998; Rodriguezet al., 2010; Romero et al., 2009; Sobie, 2009). Furthermore, thissensitivity analysis could be used for the fine-tuning andimprovement of the models. Finally, our analysis provides impor-tant insights on the electrophysiological characteristics each modelis able to reproduce and is key for a better understanding of whateach model is useful for.
4.2. Sensitivity of electrophysiological biomarkersof arrhythmic risk
Our rabbit simulations show that APD, the main preclinicalbiomarker used for drug cardiotoxicity evaluation, is moderatelysensitive to changes in many repolarization current parameters inboth AP rabbit ventricular models. Therefore, APD prolongationmay not be useful for arrhythmic risk stratification because thesame APD can be obtained for different ionic current alterations, asshown in experimental studies (Hondeghem et al., 2001; Lawrenceet al., 2006) and in our previous theoretical work in humans
(Romero et al., 2009). Our study reaffirms the need of using addi-tional electrophysiological characteristics to assess pro-arrhythmicrisk preclinically, as previously suggested (Corrias et al., 2010;Hondeghem et al., 2001; Lawrence et al., 2005; Romero et al.,2009). The electrophysiological characteristics studied in thiswork are usually considered in preclinical and basic scienceinvestigations (Hondeghem et al., 2001; Lawrence et al., 2005;Lengyel et al., 2001; McCans et al., 1974; Szigligeti et al., 1996,1998; Tolkacheva et al., 2006). Clinical evaluation of cardiacarrhythmicity requires the use of ECG-based biomarkers, such asthe QT interval (Corrias et al., 2010).
Our results show that GK1 reduction results in AP triangulationprolongation in rabbit, as experimentally reported (Giles andImaizumi, 1988; Lu et al., 2008; Ruiz et al., 1992), which is relatedto an increase in the risk of Torsades de Pointes (TdP) occurrence,a polymorphic ventricular arrhythmia. In addition, steady-state[Ca2þ]i levels, APD rate adaptation, restitution properties and[Ca2þ]i and [Naþ]i frequency dependence are strongly dependenton INaK, INaCa and ICaL properties. Our results support the relevantrole of these currents in cell homeostasis and in arrhythmiasrelated to alternans and afterdepolarizations. The influence of thefast sodium current on the properties studied in this paper wasnegligible (results not shown), consistent with a previous study(Boyett and Jewell, 1980). However, it is well known that its effectson AP depolarization, conduction velocity or safety factor of prop-agation (Romero et al., 2009; Shaw and Rudy,1997) could be criticalfor arrhythmia generation.
As previously stated, the sensitivity analysis performed herecould be used to identify avenues for model improvement, con-cerning for example the determination of model parameters indi-rectly determined in experiments, such as those characterizingNaþ/Kþ pump and NCX activity. For example, as sslow is basicallydetermined by GNaK (see Fig. 2), the effect of this model parameteron sslow could be considered in the characterization of GNaK.
4.3. Differences in biomarkers using the rabbit ventricularAP models
In both models, steady-state APD, calcium rate dependence aswell as their modulation bymodification of ionic current propertiesare overall in good agreement with experimental recordings.Exceptions are the exaggerated role of IKs on APD and the effects ofsmall variations of ICaL on APD in the Mahajan et al. model. Ourresults also show that AP triangulation, systolic [Ca2þ]i, slope-max,DYN and sslow in bothmodels andmaximum systolic [Naþ]i in theMahajan et al. model are outside experimentally-reported ranges.
Regarding APD modulation, both models are markedly sensitiveto INaK alterations and severe blocks of ICaL and Ito. However, someunderlying mechanisms differ between the two models: sf, GKr andGto,s, and GNaCa (only for severe blocks) play a relevant role in APDmodulation in the Shannon et al. model, whereas, alterations in GKsand Ito,f modify the APD in the Mahajan et al. model. ICaL and INaK, aswell as INaCa when it is severely reduced, are determinant forsteady-state systolic [Ca2þ]i. However, the effects of Ito on thisproperty are model dependent.
Furthermore, control values and ionic mechanisms modulatingsslow, sodium rate dependence in control and ionic mechanisms ofAP triangulation reproduce experimental data more faithfully inthe Shannon et al. model than in the Mahajan et al. model.Conversely, control values of AP triangulation and APD restitutioncurves, as well as the ionic mechanisms modulating restitutionproperties are reasonably well reproduced by the Mahajan et al.model, except for the fact that IKs impact in electrophysiologicalproperties seems to be too large. In addition, comparison of therelative sensitivities between bothmodels reveals some differences

L. Romero et al. / Progress in Biophysics and Molecular Biology 107 (2011) 60e73 71
on the impact of the main transmembrane currents on thebiomarkers of pro-arrhythmic risk. Indeed, alterations of IKr andINaCa more effectively modify the electrophysiological properties inthe Shannon et al. model. Conversely, the influence of IKs, Ito,f andINaK is more significant in the Mahajan et al. model. The differentimpact of the ionic currents on the electrophysiological propertiesbetween both models is related to the modifications that Mahajanet al. included in the formulation of the membrane currents (GKrwas divided by 2.4 and Gks and Gtof were two- and six-foldincreased, respectively) and the differences in calcium dynamicsthat affect the activity of the NCX and the Naþ/Kþ pump.
Overall, our results suggest that the Shannon et al. model ismore appropriate to reproduce rabbit electrophysiology at sinuspacing rate, while the Mahajan et al. model mimics rabbit elec-trophysiology at faster rates. However, this conclusion should beconsidered with caution due to the high dispersion of rabbitexperimental data (Lu et al., 2008).
4.4. Experiments for further understanding of rabbit cellularelectrophysiology
Our study shows how computational physiology can be used forfurther understanding of cardiac electrophysiology and the mech-anisms of arrhythmias. Indeed, our simulation study aims at com-plementing experimental studies, helping to raise new questions,and suggests new experiments to fill specific knowledge gapsrelated to rabbit ventricular electrophysiology.
In this work, we have altered maximum conductances andkinetics of the main transmembrane currents and we have inves-tigated the effect of the alterations on the main cellular electro-physiological properties at different levels, membrane potential,concentrations and their rate dependence. Our simulations revealthat, according to the models, the Naþ/Kþ pump is the mostinfluential current in most simulated cellular biomarkers. To thebest of our knowledge, no experimental results have studied theeffect of this current on the dynamic or standard AP restitutioncurves, on the accommodation of the rabbit APD to sudden changesin frequency pacing, neither on the [Naþ]i rate dependence.Moreover, there are some uncertainties regarding the role of theNaþ/Kþ pump on calcium rate dependence. In addition, our simu-lations show that NCX is also relevant in many electrophysiologicalproperties. Unfortunately, controversial experimental results aboutthe impact of NCX on APD and calcium levels have been found andno experiments have been undertaken to dilucidate its role onrabbit APD restitution curves to our knowledge. In addition, there isalso some uncertainty regarding the impact of IKs and IKr alterationon rabbit electrophysiology under normal conditions. Finally, theheterogeneous experimental conditions and the natural variabilitydifficult the quantitative comparison between simulation resultsand experiments of the effects of the main transmembranecurrents on cardiac electrophysiology.
We believe that experiments performed in rabbit ventriculartissues analyzing the roles of the Naþ/Kþ pump using blockers(such as strophantidine and ouabain), and the NCX in over-expressing NCX proteins on the biomarkers selected for this studywould be valuable for further understanding of rabbit cellularelectrophysiology and AP model improvement and validation. Inaddition, experiments evaluating the impact of the natural vari-ability of the main transmembrane currents on the biomarkers orarrhythmic risk would also be very relevant. For this purpose,tissues from different regions of the heart (apex and base, left andright ventricle), transmural positions (epi, endo and mid-myocardium), sex and age should be studied under similar exper-imental conditions (Lu et al., 2005b; Sims et al., 2008).
4.5. Human and rabbit inter-species comparison
In our previous study (Romero et al., 2009), we performeda similar sensitivityanalysis on theTenTusscherandPanfilovhumanventricular AP model (TP06) to the one performed in the currentstudy using two rabbit models. Comparison of the relative sensi-tivities of the cellular electrophysiological properties related toarrhythmic risk to changes in ionic current properties betweenrabbit and humans reveals interesting insights that could beconsidered when extrapolating results from rabbits to humans. Wefound that similar mechanismsmodulate sslow and AP triangulationin both species. The ionic mechanisms of systolic [Ca2þ]i levels atelectrophysiological normal rates and calcium rate dependence arevery similar in the TP06 model and the Shannon et al. model.However, modulation of both APD restitution curves in the TP06model resembles the results observedwhenusing theMahajan et al.model (Romero et al., 2009). The main differences between rabbitand human are observed in the ionic mechanisms of APD andsodium rate dependence. Indeed, our previous results showa stronginfluence of small variations of IKs on APD in the TP06 model that isnot observed in rabbit models. Conversely, the pivotal role of INaK onAPD in both rabbit ventricular models is not shownwhen using theTP06 model. In addition, sodium rate dependence is modulated bymany transmembrane currents in the TP06 model, namely, ICaL,which is themost influential current, but also IKs, INaK and IKr. Sodiumrate dependence is however significantly modified by GNaK whenusing rabbit models. These discrepancies can arise from true speciesdependent mechanisms or from model deficiencies. The stronginfluence of IKs on APDobserved in the TP06model could be amodeldeficiency as human experiments show a small influence of thiscurrent on APD under normal conditions (Jost et al., 2005).Regarding the roleof INaKonAPD, it is consistentwithexperiments indifferent animal species (Bers et al., 1988; Kodama et al., 1981;Sanchez-Serrano et al., 1980; Pueyo et al., 2010). The INaK formula-tion of the TP06 model is not fitted to human data but it is possiblethat the role of INaK onAPD could be different in these two species. Inthe TP06model, ICaL, the current exerting the strongest influence onsodium rate dependence, and IKr, weremostly fitted to human data.However, calciumdynamicswerebasedon theShannonet al.model,and INaK was adopted from the LuoeRudy model. Therefore, thedifferential role of ICaL, IKr and possibly INaK, in sodium rate adapta-tion observed in our simulations when using rabbit and humanmodels could be a true inter-species difference. Unfortunately,experiments reporting the changes in intracellular sodiumconcentration with varying pacing rate are scarce and no experi-mental data of these two species on the mechanisms modulatingsodium rate dependence have been found in the literature.
The evidence of different APD ionic mechanisms in rabbit andhuman myocytes reinforce the need of biomarkers exertinga similar behavior in both species to assess pro-arrhythmic riskpreclinically due to the fact that experimental studies of pro-arrhythmic mechanisms are scarcely performed in humans.
5. Conclusions
In this study, a systematic analysis of the sensitivity of the maincellular biomarkers of arrhythmic risk in rabbits to variations in ioniccurrent properties using the Shannon et al. (2004) and the Mahajanet al. (2008b) models has been performed. In addition, modeloutputs have been compared to experimental findings available in theliterature. Our results show that both models reproduce manyelectrophysiological characteristics but, as expected due to gaps ofknowledge in ventricular electrophysiology, differences betweensimulations and experiments are also found. In addition, this analysisprovides new insights into the ionic basis of rabbit cellular

L. Romero et al. / Progress in Biophysics and Molecular Biology 107 (2011) 60e7372
electrophysiology, identifying controversies that could suggest theneed of further experiments. Finally, our results reinforce theusefulness of this sensitivity analysis for further understanding ofcellular electrophysiology and validation of cardiac AP models.
6. Limitations of the study
We have performed a systematic analysis on the role of ioniccurrent properties in modulating preclinical arrhythmic riskbiomarkers in rabbit myocytes using two widely-used rabbitventricularmodels, the Shannonet al. and theMahajanet al.models.For this purpose, we have studied the influence of the maximumconductance andkinetics of themain transmembrane ionic currentsin the main cellular electrophysiological characteristics related toarrhythmic risk in both ventricular rabbit models. Other modelparameters, like steady-state values of the gates, may also affectthese properties and have not been included in this study.
In addition, it is well known that electrotonic currents can affectcellular electrophysiological properties. Some simulation studieshave shown differences in AP amplitude and shape and in therestitution curves, alternans and memory between isolated cellsand coupled cells (Bueno-Orovio et al., 2008; Cherry et al., 2008;Cherry and Evans, 2008; Cherry and Fenton, 2007; Nygren et al.,1998; Ten Tusscher et al., 2006). Although arrhythmic episodesare generated at the tissue level, our simulations were performed inan isolated virtual cell for computational purposes. Therefore, theionic basis of the cellular biomarkers of pro-arrhythmic riskdescribed in this paper may be modified when coupling the virtualcells into a tissue.
Editors’ note
Please see also related communications in this issue by Cooperet al. (2011) and Winslow and Greenstein (2011).
Acknowledgments
This work was partially supported by the European CommissionpreDiCT grant (DG INFSO-224381), Plan Nacional de InvestigaciónCientífica, Desarrollo e Innovación Tecnológica del Ministerio deCiencia e Innovación of Spain (TEC2008-02090, TIN2004-03602),Programa de Apoyo a la Investigación y Desarrollo (PAID-06-09-2843)de la Universidad Politécnica de Valencia, Dirección Generalde Política Científica de la Generalitat Valenciana (GV/2010/078),a Consellería de Educación de la Generalitat Valenciana grant(BEST/2010/102) (to J.M.F), a Universidad Politécnica de Valenciagrant (PAID-00-10) (to L.R.) and an UK Medical Research CouncilCareer Development Award (to B.R.).
References
Anumonwo, J., 2008. Antiarrhythmic benefits of targeting the Naþ/Ca2þ exchanger.Heart Rhythm 5, 1453e1454.
Aslanidi, O.V., Sleiman, R.N., Boyett, M.R., Hancox, J.C., Zhang, H., 2010. Ionicmechanisms for electrical heterogeneity between rabbit Purkinje fiber andventricular cells. Biophys. J. 98, 2420e2431.
Baartscheer, A., Schumacher, C.A., van Borren, M.M., Belterman, C.N., Coronel, R.,Fiolet, J.W., 2003. Increased Naþ/Hþ-exchange activity is the cause of increased[Naþ]i and underlies disturbed calcium handling in the rabbit pressure andvolume overload heart failure model. Cardiovasc. Res. 57, 1015e1024.
Bassani, R.A., Altamirano, J., Puglisi, J.L., Bers, D.M., 2004. Action potential durationdetermines sarcoplasmic reticulum Ca2þ reloading in mammalian ventricularmyocytes. J. Physiol. 559, 593e609.
Bell, J., Rouze, N., Krassowska, W., Idriss, S., 2007. The electrocardiogram restitutionportrait quantifying dynamical electrical instability in young myocardium.Comput. Cardiol. 34, 789e792.
Bers, D.M., 2002. Cardiac excitation-contraction coupling. Nature 415, 198e205.
Bers, D.M., Christensen, D.M., Nguyen, T.X., 1988. Can Ca entry via Na-Caexchange directly activate cardiac muscle contraction? J. Mol. Cell Cardiol.20, 405e414.
Bers, D.M., Despa, S., 2006. Cardiac myocytes Ca2þ and Naþ regulation in normaland failing hearts. J. Pharmacol. Sci. 100, 315e322.
Biagetti, M.O., Quinteiro, R.A., 2006. Gender differences in electrical remodeling andsusceptibility to ventricular arrhythmias in rabbits with left ventricularhypertrophy. Heart Rhythm 3, 832e839.
Boyett, M.R., Fedida, D., 1984. Changes in the electrical activity of dog cardiacPurkinje fibres at high heart rates. J. Physiol. 350, 361e391.
Boyett, M.R., Jewell, B.R., 1980. Analysis of the effects of changes in rate and rhythmupon electrical activity in the heart. Prog. Biophys. Mol. Biol. 36, 1e52.
Bueno-Orovio, A., Cherry, E.M., Fenton, F.H., 2008. Minimal model for humanventricular action potentials in tissue. J. Theor. Biol. 253, 544e560.
Cherry, E.M., Evans, S.J., 2008. Properties of two human atrial cell models in tissue:restitution, memory, propagation, and reentry. J. Theor. Biol. 254, 674e690.
Cherry, E.M., Fenton, F.H., 2004. Suppression of alternans and conduction blocksdespite steep APD restitution: electrotonic, memory, and conduction velocityrestitution effects. Am. J. Physiol. Heart Circ. Physiol. 286, H2332eH2341.
Cherry, E.M., Fenton, F.H., 2007. A tale of two dogs: analyzing two models of canineventricular electrophysiology. Am. J. Physiol. Heart Circ. Physiol. 292, H43eH55.
Cherry, E.M., Hastings, H.M., Evans, S.J., 2008. Dynamics of human atrial cellmodels: restitution, memory, and intracellular calcium dynamics in single cells.Prog. Biophys. Mol. Biol. 98, 24e37.
Chudin, E., Goldhaber, J., Garfinkel, A., Weiss, J., Kogan, B., 1999. Intracellular Ca2þ
dynamics and the stability of ventricular tachycardia. Biophys. J. 77, 2930e2941.Clayton, R.H., Bernus, O., Cherry, E.M., Dierckx, H., Fenton, F.H., Mirabella, L.,
Panfilov, A.V., Sachse, F.B., Seemann, G., Zhang, H., 2011. Models of cardiac tissueelectrophysiology: progress, challenges and open questions. Prog. Biophys. Mol.Biol. 104, 22e48.
Cooper, J., Corrias, A., Gavaghan, D., Noble, D., 2011. Considerations for the use ofcellular electrophysiology models within cardiac tissue simulations. Prog. Bio-phys. Mol. Biol. 107, 74e80.
Corrias, A., Jie, X., Romero, L., Bishop, M.J., Bernabeu, M., Pueyo, E., Rodriguez, B.,2010. Arrhythmic risk biomarkers for the assessment of drug cardiotoxicity:from experiments to computer simulations. Philos. Transact. A Math. Phys. Eng.Sci. 368, 3001e3025.
Eckardt, L., Breithardt, G., Haverkamp, W., 2002. Electrophysiologic characterizationof the antipsychotic drug sertindole in a rabbit heart model of Torsade depointes: low torsadogenic potential despite QT prolongation. J. Pharmacol. Exp.Ther. 300, 64e71.
Farkas, A.S., Acsai, K., Nagy, N., Toth, A., Fulop, F., Seprenyi, G., Birinyi, P., Nanasi, P.P.,Forster, T., Csanady, M., Papp, J.G., Varro, A., Farkas, A., 2008. Na(þ)/Ca(2þ)exchanger inhibition exerts a positive inotropic effect in the rat heart, but failsto influence the contractility of the rabbit heart. Br. J. Pharmacol. 154, 93e104.
Fedida, D., Giles, W.R., 1991. Regional variations in action potentials and transientoutward current in myocytes isolated from rabbit left ventricle. J. Physiol. 442,191e209.
Fink, M., Niederer, S.A., Cherry, E.M., Fenton, F.H., Koivumaki, J.T., Seemann, G.,Thul, R., Zhang, H., Sachse, F.B., Beard, D., Crampin, E.J., Smith, N.P., 2011. Cardiaccell modelling: observations from the heart of the cardiac physiome project.Prog. Biophys. Mol. Biol. 104, 2e21.
Garfinkel, A., Kim, Y.H., Voroshilovsky, O., Qu, Z., Kil, J.R., Lee, M.H., Karagueuzian, H.S.,Weiss, J.N., Chen, P.S., 2000. Preventing ventricular fibrillation by flatteningcardiac restitution. Proc. Natl. Acad. Sci. U.S.A. 97, 6061e6066.
Giles, W., Shimoni, Y., 1989. Slow inward tail currents in rabbit cardiac cells.J. Physiol. 417, 447e463.
Giles, W.R., Imaizumi, Y., 1988. Comparison of potassium currents in rabbit atrialand ventricular cells. J. Physiol. 405, 123e145.
Goldhaber, J.I., Xie, L.H., Duong, T., Motter, C., Khuu, K., Weiss, J.N., 2005. Actionpotential duration restitution and alternans in rabbit ventricular myocytes: thekey role of intracellular calcium cycling. Circ. Res. 96, 459e466.
Guerard, N.C., Traebert, M., Suter, W., Dumotier, B.M., 2008. Selective block of IKsplays a significant role in MAP triangulation induced by IKr block in isolatedrabbit heart. J. Pharmacol. Toxicol. Methods 58, 32e40.
Guo, D., Zhou, J., Zhao, X., Gupta, P., Kowey, P.R., Martin, J., Wu, Y., Liu, T., Yan, G.X.,2008. L-type calcium current recovery versus ventricular repolarization:preserved membrane-stabilizing mechanism for different QT intervals acrossspecies. Heart Rhythm 5, 271e279.
Hondeghem, L.M., Carlsson, L., Duker, G., 2001. Instability and triangulation of theaction potential predict serious proarrhythmia, but action potential durationprolongation is antiarrhythmic. Circulation 103, 2004e2013.
Idriss, S.F., Wolf, P.D., 2004. Transmural action potential repolarization heteroge-neity develops postnatally in the rabbit. J. Cardiovasc. Electrophysiol. 15,795e801.
Janvier, N.C., Boyett, M.R., 1996. The role of Na-Ca exchange current in the cardiacaction potential. Cardiovasc. Res. 32, 69e84.
Jost, N., Papp, J.G., Varro, A., 2007. Slow delayed rectifier potassium current (IKs) andthe repolarization reserve. Ann. Noninvasive Electrocardiol. 12, 64e78.
Jost, N., Virag, L., Bitay, M., Takacs, J., Lengyel, C., Biliczki, P., Nagy, Z., Bogats, G.,Lathrop, D.A., Papp, J.G., Varro, A., 2005. Restricting excessive cardiac actionpotential and QT prolongation: a vital role for IKs in human ventricular muscle.Circulation 112, 1392e1399.
Koch-Weser, J., Blinks, J.R., 1963. The influence of the interval between beats onmyocardial contractility. Pharmacol. Rev. 15, 601e652.

L. Romero et al. / Progress in Biophysics and Molecular Biology 107 (2011) 60e73 73
Kodama, I., Toyama, J., Shibata, S., Norton, T.R., 1981. Electrical and mechanicaleffects of anthopleurin-A, a polypeptide from a sea anemone, on isolated rabbitventricular muscle under conditions of hypoxia and glucose-free medium.J. Cardiovasc. Pharmacol. 3, 75e86.
Komai, H., Chiou, K.Y., Rusy, B.F., 1996. Lack of inhibition by inhalational anestheticsof myocardial contraction dependent on intracellular sodium activity. Anes-thesiology 85, 1139e1146.
Kurata, Y., Matsuda, H., Hisatome, I., Shibamoto, T., 2008. Regional difference indynamical property of sinoatrial node pacemaking: role of Naþ channel current.Biophys. J. 95, 951e977.
Lawrence, C.L., Bridgland-Taylor, M.H., Pollard, C.E., Hammond, T.G., Valentin, J.P.,2006. A rabbit Langendorff heart proarrhythmia model: predictive value forclinical identification of Torsades de Pointes. Br. J. Pharmacol. 149, 845e860.
Lawrence, C.L., Pollard, C.E., Hammond, T.G., Valentin, J.P., 2005. Nonclinicalproarrhythmia models: predicting Torsades de Pointes. J. Pharmacol. Toxicol.Methods 52, 46e59.
Lengyel, C., Iost, N., Virag, L., Varro, A., Lathrop, D.A., Papp, J.G., 2001. Pharmaco-logical block of the slow component of the outward delayed rectifier current(IKs) fails to lengthen rabbit ventricular muscle QTc and action potential dura-tion. Br. J. Pharmacol. 132, 101e110.
Linz, K.W., Meyer, R., 2000. Profile and kinetics of L-type calcium current during thecardiac ventricular action potential compared in guinea-pigs, rats and rabbits.Pflugers Arch. 439, 588e599.
Loughrey, C.M., MacEachern, K.E., Cooper, J., Smith, G.L., 2003. Measurement of thedissociation constant of Fluo-3 for Ca2þ in isolated rabbit cardiomyocytes usingCa2þ wave characteristics. Cell Calcium 34, 1e9.
Lu, H.R., Vlaminckx, E., Gallacher, D.J., 2008. Choice of cardiac tissue in vitro plays animportant role in assessing the risk of drug-induced cardiac arrhythmias inhuman: beyond QT prolongation. J. Pharmacol. Toxicol. Methods 57, 1e8.
Lu, H.R., Vlaminckx, E., Teisman, A., Gallacher, D.J., 2005a. Choice of cardiac tissueplays an important role in the evaluation of drug-induced prolongation of theQT interval in vitro in rabbit. J. Pharmacol. Toxicol. Methods 52, 90e105.
Lu, H.R., Vlaminckx, E., Van de, W.A., Gallacher, D.J., 2005b. Both beta-adrenergicreceptor stimulation and cardiac tissue type have important roles in eluci-dating the functional effects of IKs channel blockers in vitro. J. Pharmacol.Toxicol. Methods 51, 81e90.
Luo, C.H., Rudy, Y., 1994. A dynamic model of the cardiac ventricular actionpotential. I. Simulations of ionic currents and concentration changes. Circ. Res.74, 1071e1096.
Mahajan, A., Sato, D., Shiferaw, Y., Baher, A., Xie, L.H., Peralta, R., Olcese, R., Garfinkel, A.,Qu, Z.,Weiss, J.N., 2008a.Modifying L-type calciumcurrent kinetics: consequencesfor cardiac excitation and arrhythmia dynamics. Biophys. J. 94, 411e423.
Mahajan, A., Shiferaw, Y., Sato, D., Baher, A., Olcese, R., Xie, L.H., Yang, M.J., Chen, P.S.,Restrepo, J.G., Karma, A., Garfinkel, A., Qu, Z., Weiss, J.N., 2008b. A rabbitventricular action potential model replicating cardiac dynamics at rapid heartrates. Biophys. J. 94, 392e410.
Maier, L.S., Bers, D.M., Pieske, B., 2000. Differences in Ca2þ-handling and sarco-plasmic reticulum Ca2þ-content in isolated rat and rabbit myocardium. J. Mol.Cell Cardiol. 32, 2249e2258.
McCans, J.L., Lindenmayer, G.E., Munson, R.G., Evans, R.W., Schwartz, A., 1974.A dissociation of positive staircase (Bowditch) from ouabain-induced positiveinotropism. Circ. Res. 35, 439e447.
Nolasco, J.B., Dahlen, R.W., 1968. A graphic method for the study of alternation incardiac action potentials. J. Appl. Physiol. 25, 191e196.
Nygren, A., Fiset, C., Firek, L., Clark, J.W., Lindblad, D.S., Clark, R.B., Giles, W.R., 1998.Mathematical model of an adult human atrial cell: the role of Kþ currents inrepolarization. Circ. Res. 82, 63e81.
Nygren, A., Leon, L.J., Giles, W.R., 2001. Simulations of the human atrial actionpotential. Phil. Trans. R. Soc. Lond. A 359, 1111e1125.
Panfilov, A.V., 2006. Is heart size a factor in ventricular fibrillation? Or how close arerabbit and human hearts? Heart Rhythm 3, 862e864.
Pastore, J.M., Laurita, K.R., Rosenbaum, D.S., 2006. Importance of spatiotemporalheterogeneity of cellular restitution in mechanism of arrhythmogenic discor-dant alternans. Heart Rhythm 3, 711e719.
Pitruzzello, A.M., Krassowska, W., Idriss, S.F., 2007. Spatial heterogeneity of therestitution portrait in rabbit epicardium. Am. J. Physiol. Heart Circ. Physiol. 292,H1568eH1578.
Pueyo, E., Husti, Z., Hornyik, T., Baczko, I., Laguna, P., Varro, A., Rodriguez, B., 2010.Mechanisms of ventricular rate adaptation as a predictor of arrhythmic risk.Am. J. Physiol. Heart Circ. Physiol. 298, H1577eH1587.
Pueyo, E., Smetana, P., Caminal, P., de Luna, A.B., Malik, M., Laguna, P., 2004. Char-acterization of QT interval adaptation to RR interval changes and its use asa risk-stratifier of arrhythmic mortality in amiodarone-treated survivors ofacute myocardial infarction. IEEE Trans. Biomed. Eng. 51, 1511e1520.
Puglisi, J.L., Bers, D.M., 2001. LabHEART: an interactive computer model of rabbitventricular myocyte ion channels and Ca transport. Am. J. Physiol. Cell Physiol.281, C2049eC2060.
Ranu, H.K., Terracciano, C.M., Davia, K., Bernobich, E., Chaudhri, B., Robinson, S.E.,Bin, K.Z., Hajjar, R.J., MacLeod, K.T., Harding, S.E., 2002. Effects of Naþ/Ca2þ-exchanger overexpression on excitation-contraction coupling in adult rabbitventricular myocytes. J. Mol. Cell Cardiol. 34, 389e400.
Restrepo, J.G., Karma, A., 2009. Spatiotemporal intracellular calcium dynamicsduring cardiac alternans. Chaos 19, 037115.
Restrepo, J.G., Weiss, J.N., Karma, A., 2008. Calsequestrin-mediated mechanism forcellular calcium transient alternans. Biophys. J. 95, 3767e3789.
Rodriguez, B., Burrage, K., Gavaghan, D., Grau, V., Kohl, P., Noble, D., 2010. Thesystems biology approach to drug development: application to toxicityassessment of cardiac drugs. Clin. Pharmacol. Ther. 88, 130e134.
Romero, L., Pueyo, E., Fink, M., Rodriguez, B., 2009. Impact of ionic current vari-ability on human ventricular cellular electrophysiology. Am. J. Physiol. HeartCirc. Physiol. 297, H1436eH1445.
Ruiz, P.E., Leblanc, N., deLorenzi, F., Allard, Y., Schanne, O.F., 1992. Effects of Kþ
channel blockers on the action potential of hypoxic rabbit myocardium. Br. J.Pharmacol. 106, 924e930.
Sanchez-Serrano, D., Alvarez, J.L., Garcia-Barreto, D., 1980. Enhancement of ouabainarrhythmias by fatty acids. J. Cardiovasc. Pharmacol. 2, 331e335.
Sah, R., Ramirez, R.J., Oudit, G.Y., Gidrewicz, D., Trivieri, M.G., Zobel, C., Backx, P.H.,2003. Regulation of cardiac excitationecontraction coupling by action potentialrepolarization: role of the transient outward potassium current (Ito). J. Physiol.546, 5e18.
Sato, D., Xie, L.H., Sovari, A.A., Tran, D.X., Morita, N., Xie, F., Karagueuzian, H.,Garfinkel, A., Weiss, J.N., Qu, Z., 2009. Synchronization of chaotic early after-depolarizations in the genesis of cardiac arrhythmias. Proc. Natl. Acad. Sci. U.S.A.106, 2983e2988.
Schillinger, W., Janssen, P.M., Emami, S., Henderson, S.A., Ross, R.S., Teucher, N.,Zeitz, O., Philipson, K.D., Prestle, J., Hasenfuss, G., 2000. Impaired contractileperformance of cultured rabbit ventricular myocytes after adenoviral genetransfer of Naþ-Ca2þ exchanger. Circ. Res. 87, 581e587.
Schillinger, W., Ohler, A., Emami, S., Muller, F., Christians, C., Janssen, P.M., Kogler, H.,Teucher, N., Pieske, B., Seidler, T., Hasenfuss, G., 2003. The functional effect ofadenoviral Naþ/Ca2þ exchanger overexpression in rabbit myocytes depends onthe activity of the Naþ/Kþ-ATPase. Cardiovasc. Res. 57, 996e1003.
Seidler, T., Miller, S.L., Loughrey, C.M., Kania, A., Burow, A., Kettlewell, S., Teucher, N.,Wagner, S., Kogler, H., Meyers, M.B., Hasenfuss, G., Smith, G.L., 2003. Effects ofadenovirus-mediated sorcin overexpression on excitation-contraction couplingin isolated rabbit cardiomyocytes. Circ. Res. 93, 132e139.
Shannon, T.R., Wang, F., Puglisi, J., Weber, C., Bers, D.M., 2004. A mathematicaltreatment of integrated Ca dynamics within the ventricular myocyte. Biophys. J.87, 3351e3371.
Shaw, R.M., Rudy, Y., 1997. Ionic mechanisms of propagation in cardiac tissue. Rolesof the sodium and L-type calcium currents during reduced excitability anddecreased gap junction coupling. Circ. Res. 81, 727e741.
Sims, C., Reisenweber, S., Viswanathan, P.C., Choi, B.R., Walker, W.H., Salama, G.,2008. Sex, age, and regional differences in L-type calcium current are importantdeterminants of arrhythmia phenotype in rabbit hearts with drug-induced longQT type 2. Circ. Res. 102, e86e100.
Sobie, E.A., 2009. Parameter sensitivity analysis in electrophysiological modelsusing multivariable regression. Biophys. J. 96, 1264e1274.
Szigligeti, P., Banyasz, T., Magyar, J., Szigeti, G., Papp, Z., Varro, A., Nanasi, P.P., 1998.Intracellular calcium and electrical restitution in mammalian cardiac cells. ActaPhysiol. Scand. 163, 139e147.
Szigligeti, P., Pankucsi, C., Banyasz, T., Varro, A., Nanasi, P.P., 1996. Action potentialduration and force-frequency relationship in isolated rabbit, guinea pig and ratcardiac muscle. J. Comp. Physiol. [B] 166, 150e155.
Ten Tusscher, K.H., Bernus, O., Hren, R., Panfilov, A.V., 2006. Comparison of elec-trophysiological models for human ventricular cells and tissues. Prog. Biophys.Mol. Biol. 90, 326e345.
Tolkacheva, E.G., Anumonwo, J.M., Jalife, J., 2006. Action potential duration resti-tution portraits of mammalian ventricular myocytes: role of calcium current.Biophys. J. 91, 2735e2745.
Toth, A., Kiss, L., Varro, A., Nanasi, P.P., 2009. Potential therapeutic effects of Naþ/Ca2þ exchanger inhibition in cardiac diseases. Curr. Med. Chem. 16, 3294e3321.
Virag, L., Iost, N., Opincariu, M., Szolnoky, J., Szecsi, J., Bogats, G., Szenohradszky, P.,Varro, A., Papp, J.G., 2001. The slow component of the delayed rectifier potas-sium current in undiseased human ventricular myocytes. Cardiovasc. Res. 49,790e797.
Watanabe, M.A., Fenton, F.H., Evans, S.J., Hastings, H.M., Karma, A., 2001. Mecha-nisms for discordant alternans. J. Cardiovasc. Electrophysiol. 12, 196e206.
Wilders, R., 2007. Computer modelling of the sinoatrial node. Med. Biol. Eng.Comput. 45, 189e207.
Winslow, R.L., Greenstein, J.L., 2011. Cardiac myocytes and local signaling in nano-domains. Prog. Biophys. Mol. Biol. 107, 48e59.
Wu, T.J., Lin, S.F., Weiss, J.N., Ting, C.T., Chen, P.S., 2002. Two types of ventricularfibrillation in isolated rabbit hearts: importance of excitability and actionpotential duration restitution. Circulation 106, 1859e1866.
Xie, L.H., Sato, D., Garfinkel, A., Qu, Z., Weiss, J.N., 2008. Intracellular Ca alternans:coordinated regulation by sarcoplasmic reticulum release, uptake, and leak.Biophys. J. 95, 3100e3110.
Xie, Y., Garfinkel, A., Camelliti, P., Kohl, P., Weiss, J.N., Qu, Z., 2009. Effects offibroblast-myocyte coupling on cardiac conduction and vulnerability to reentry:a computational study. Heart Rhythm 6, 1641e1649.
Yan,G.X., Rials, S.J.,Wu,Y., Liu, T., Xu, X.,Marinchak, R.A., Kowey, P.R., 2001.Ventricularhypertrophy amplifies transmural repolarization dispersion and induces earlyafterdepolarization. Am. J. Physiol. Heart Circ. Physiol. 281, H1968eH1975.
Yap, Y.G., Camm, A.J., 2003. Drug induced QT prolongation and Torsades de pointes.Heart 89, 1363e1372.
Yarotskyy, V., Gao, G., Peterson, B.Z., Elmslie, K.S., 2009. The Timothy syndromemutation of cardiac CaV1.2 (L-type) channels: multiple altered gating mech-anisms and pharmacological restoration of inactivation. J. Physiol. 587,551e565.







![arXiv:1406.1701v1 [cs.CE] 6 Jun 2014 · A computational study of the effects of remodelled electrophysiology and mechanics on initiation of ventricular fibrillation in human heart](https://img.pdfslide.net/doc/110x75/5f7617c066919e74003ab5de/arxiv14061701v1-csce-6-jun-2014-a-computational-study-of-the-eiects-of-remodelled.jpg)