Embed Size (px)
Citation preview

SzénhidrátokSZERKEZET, REAKCIÓK, FUNKCIÓIK

Biomass
1. Carbohydrates
2. Lignans
3. Fats, Terpenoids, Proteins, Nucleic acids
A biomassza ökológiai (környezettudományi) fogalom, jelentése: biológiai úton létrejövő szervesanyagtömeg. A Földön évente újratermelődő biomassza mintegy 200 milliárd tonna/év tömegűnek becsülhető.
A biomassza összetétele

Szénhidrátok
A szénhidrátok nevüket onnan kapták, hogy régebben azt gondolták, ezek a vegyületek a szénnek vízzel alkotott vegyületei, azaz a szén „hidrátjai”.
A szénhidrátokat szerkezetük szerint két nagy csoportra oszthatjuk:
• monoszacharidok: E vegyületekre jellemző, hogy savas hidrolízissel már nem bonthatók kisebb molekulatömegű szénhidrátokra.
• összetett szénhidrátok: Jellemzőjük, hogy savas hidrolízissel egyszerű cukrokká bonthatók. Az összetett szénhidrátok egy részének fizikai és kémiai tulajdonságai még nagyon hasonlítanak a monoszacharidokéra. Ezeket oligoszacharidoknaknevezzük. Az összetett szénhidrátok másik csoportját, melyek tulajdonságai számottevően eltérnek a mono- és oligoszacharidokétól poliszacharidoknak hívjuk.

Szénhidrátok szerepe
•Energiahordozók•Növényekben: fotoszintézis•Vázanyagok: a szénhidrátokból keletkező polimerek (cellulóz, kitin)•Biológiai információk hordozói: A szénhidrátok és konjugátumaik (glikopeptidek, glikolipidek) az élő sejt felületén gondoskodnak a sejt adhézióról (tapadás), részt vesznek a sejt osztódás gátlásában, szerepük van a vírusok, baktériumok és hormonok valamint toxinok sejten való megkötődésében és irányítják az immunválaszt. Szerepük van az ivarsejtek egymásra találásában. Mint nukleozidok alkotóelemei közvetői a genetikai folyamatoknak (RNS, DNS, receptorok a sejt membrán felületén / glikolipidek, glikoproteidok)•Királis kiindulási anyagok
Szervezetben: cukorlebontás
(CO2)n + (H2O)n h
klorofil
(CH2O)n + (O2)n
iparban
keményítõEtOHüzemanyagpótló
glükóz H2O + CO2 675 kcal / mol

A monoszacharidok mint polihidroxi-oxovegyületek az oxocsoport jellege szerint aldózokra és ketózokra oszthatók fel,melyek külön-külön tovább csoportosíthatók szénatomszámuk szerint. Tekintettel arra, hogy ezek a vegyületek többkiralitás centrumot is tartalmaznak, így több sztereo izomerrel is számolnunk kell (a lehetséges sztereoizomerek száma 2n,ahol n az aszimmetrikus szénatomok számát jelöli.
Monoszacharidok
CHO
OH
HO
CHO
OH
HO
HO
CHO
OH
HO
HO
OH
CHO
OH
HO
HO
OH
OH
aldotrióz
aldotetróz
aldopentóz
aldohexóz
O
HO
O
HO
HO
O
HO
HO
OH
O
HO
HO
OH
OH
ketotrióz
ketotetróz
ketopentóz
ketohexóz
OH
OH
OH
OH
A MONOSZACHARIDOK CSOPORTOSÍTÁSA ÉS FIZIKAI TULAJDONSÁGAI
Az azonos összegképletű aldózok és ketózok egymás izomerjei. Általános összegképletük (CH2O)n alakban is felírható.
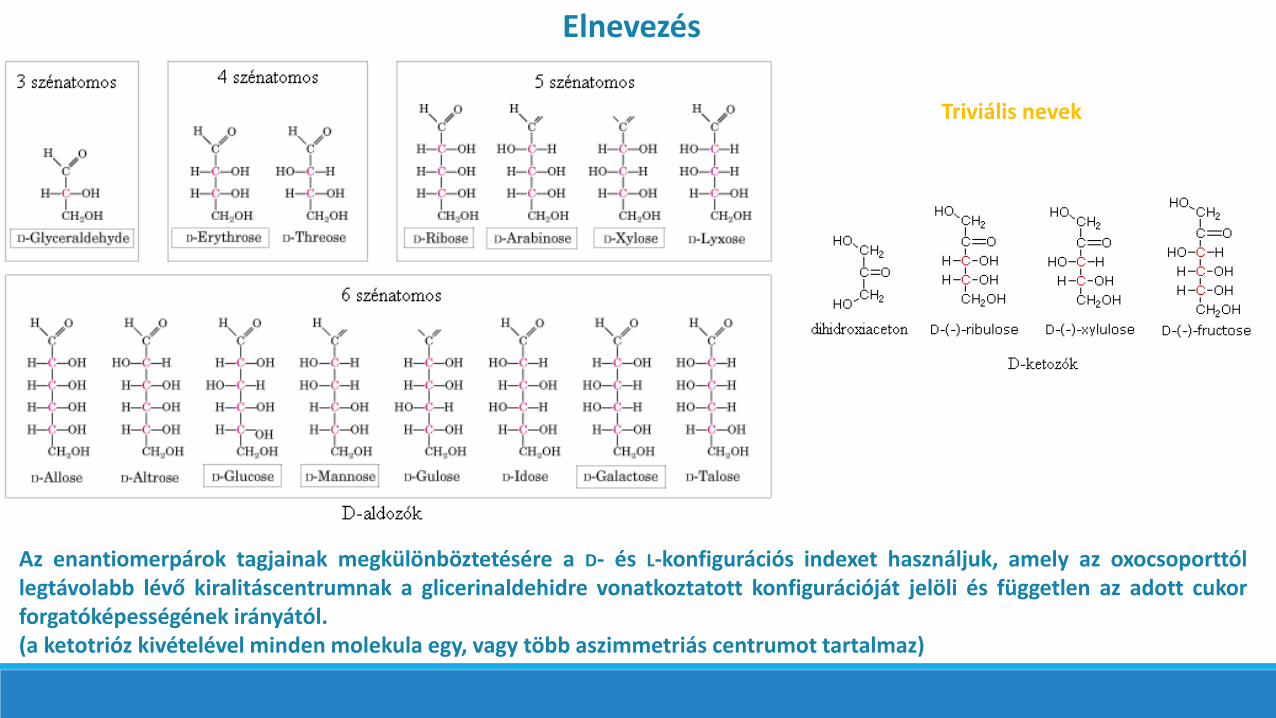
Az enantiomerpárok tagjainak megkülönböztetésére a D- és L-konfigurációs indexet használjuk, amely az oxocsoporttóllegtávolabb lévő kiralitáscentrumnak a glicerinaldehidre vonatkoztatott konfigurációját jelöli és független az adott cukorforgatóképességének irányától.(a ketotrióz kivételével minden molekula egy, vagy több aszimmetriás centrumot tartalmaz)
Triviális nevek
Elnevezés

A D-aldózok Fischer projekciójaEgyszerű cukrok (monoszacharidok)
Királis polihidroxialdehidek (aldózok)vagy ketonok (ketózok).
Híg savas hidrolízissel tovább nem bonthatók.
Glikóz
Trióz, tetróz, pentóz, hexóz, heptóz, stb.
A sorozatba
(D vagy L) tartozást
meghatározó
sztereogén centrum.

A D-ketózok Fischer projekciója
Egyszerű cukrok (monoszacharidok)
A sorozatba
(D vagy L) tartozást
meghatározó
sztereogén centrum.

A cukrok szerkezetének, konfigurációjának jelölésére a szakirodalom különböző írásmódokat használ. Az ismertebb és az általunk is használt jelölés szerint a királis szénatomok mindegyikét szubsztituenseikkel együtt feltüntetik (A). Szokás azonban az is, hogy a hidrogén szubsztituenst csupán egy vonallal jelölik és csak a hidroxilcsoportokat írják ki (B). Végül az a jelölésmód is elterjedt, hogy a hidrogén atomokat egyáltalán nem jelzik és a hidroxicsoportokat vonal szimbolizálja (C).Az utóbbi két esetben a szénlánc a Fischer-féle projekció alapján függőleges vonal, s a szénatomok metszéspontjában a szénatomok helyezkednek el.
C OHH
CHO
C
C
H HO
OHH
C
CH2OH
OHH
A
1
2
3
4
5
6
B
OH
CHO
HO
OH
CH2OH
OH
CHO
CH2OH
C
D-glükóz D-glükóz D-glükóz
A monoszacharidok szerkezete

10
Szénhidrátok térszerkezetének ábrázolása
C
ox
red
CHO
CH2OH
OHH
CHO
CH2OH
HO H
D L
C
CHO
CH2OH
H OH 1
2
3
4 C
H
CHO
CH2OHHO2 x
CH2OH
HO CHO
R
Fischer-féle vetítés: A Fischer-projekció a kémiában használt ábrázolásmód, melynek segítségével egyháromdimenziós molekulaszerkezet vetítéssel (projekcióval) két dimenzióban ábrázolható. A kötéseket függőleges ésvízszintes vonalak jelképezik oly módon, hogy a szénlánc mindig egy függőleges vonal, a szénatomok helyét pedig a vonalakmetszéspontja jelzi. Az egyes számú szénatomot rajzolják legfelülre. A módszert kidolgozójáról, Emil Fischerről nevezték el.

Fischer projection
A tetrahedral carbon atom is represented in a Fischer projection by two crossed lines. The horizontal lines representbonds coming out of the page, and the vertical lines represent bonds going into the page.

The chiral molecule can be drawn in many ways, it’s sometimes necessary to compare two projections to see if they representthe same or different enantiomers. To test for identity, Fischer projections can be moved around on the paper, but only twokinds of motions are allowed; moving a Fischer projection in any other way inverts its meaning.
A Fischer projection can be rotated on the page by 180°, but not by 90°or 270°. Only a 180°rotation maintains the Fischerconvention by keeping the same substituent groups going into and coming out of the plane. In the following Fischer projectionof (R)glyceraldehyde, for example, the -H and -OH groups come out of the plane both before and after a 180°rotation.
A 90°rotation breaks the Fischer convention by exchanging the groups that go into the plane and those that come out. In the following Fischer projections of (R)glyceraldehyde, the ] H and ] OH groups come out of the plane before rotation but go into the plane after a 90°rotation. As a result, the rotated projection represents (S)glyceraldehyde.
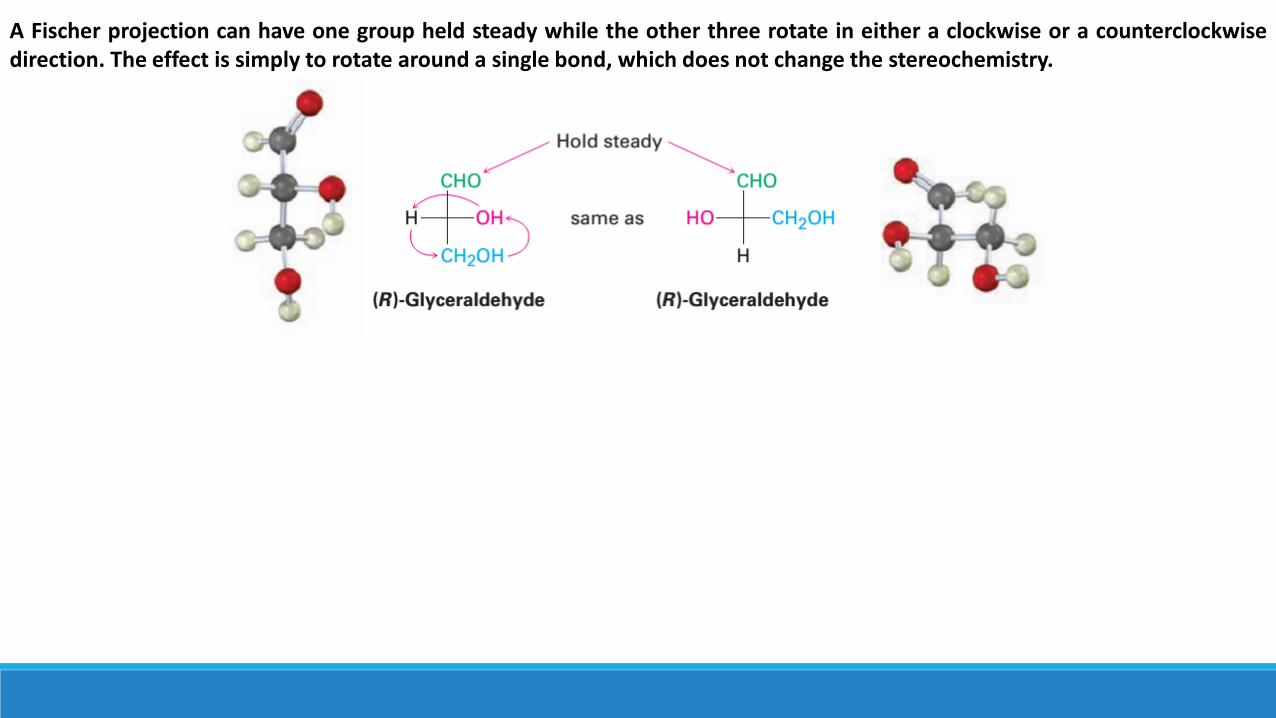
A Fischer projection can have one group held steady while the other three rotate in either a clockwise or a counterclockwisedirection. The effect is simply to rotate around a single bond, which does not change the stereochemistry.

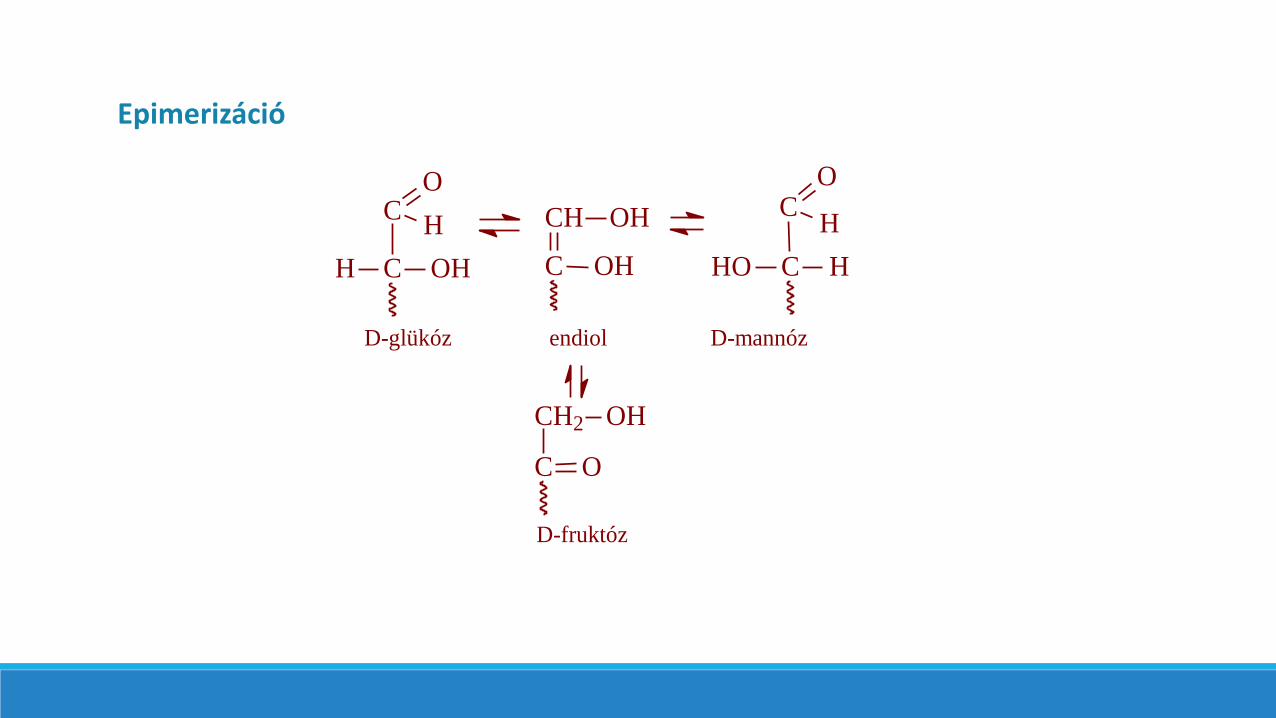
Az epimer cukrok egy szénatom konfigurációjában különböznek egymástól. Így például a D-glükóz C-2 epimerje a D-mannózés a C-4 epimerje pedig a D-galaktóz.
1
2
CHO
CH2OH
D-mannóz D-galaktóz
CHO
CH2OH
4
3
2
1
CHO
H OH
HO H
H OH
CH2OH
H OH
CHO
CH2OH
CHO
CH2OH
CHO
CH2OH
D-glükóz D-mannóz D-galaktóz
epimer cukrok
epimer cukrok
nem epimerek
24=168 db D
8 db L
Epimerek
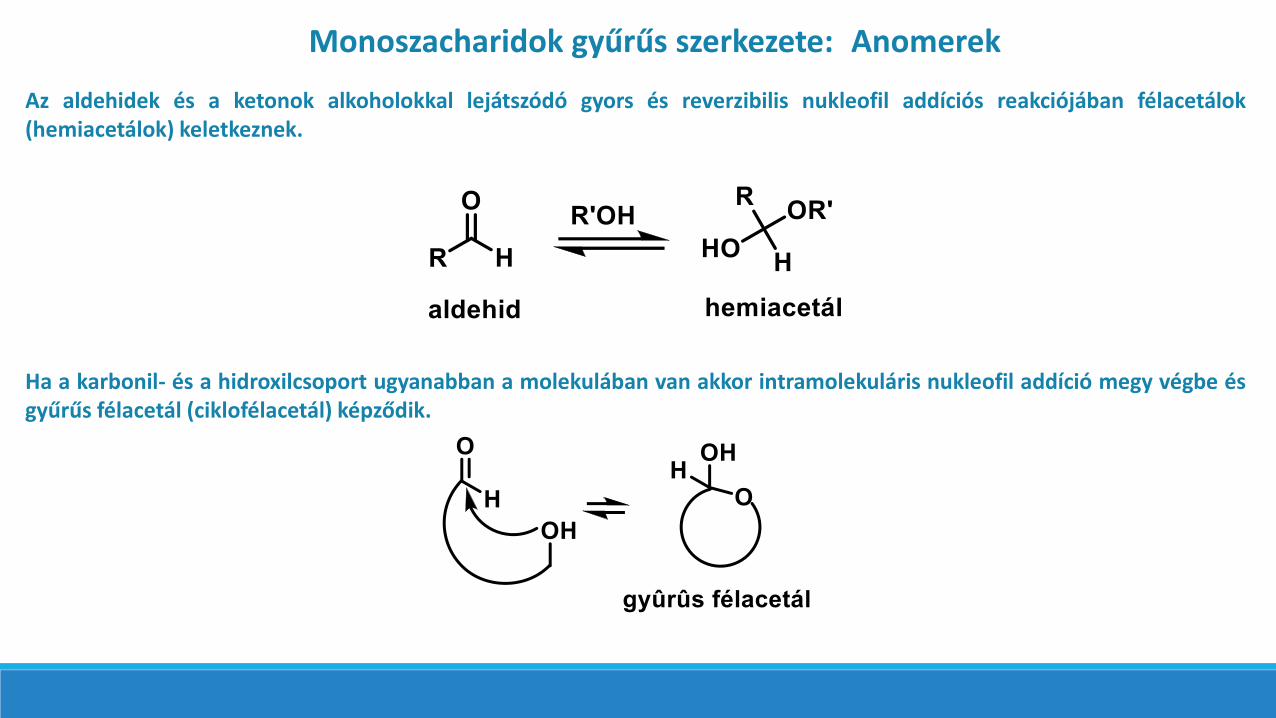
Monoszacharidok gyűrűs szerkezete: Anomerek
Az aldehidek és a ketonok alkoholokkal lejátszódó gyors és reverzibilis nukleofil addíciós reakciójában félacetálok(hemiacetálok) keletkeznek.
Ha a karbonil- és a hidroxilcsoport ugyanabban a molekulában van akkor intramolekuláris nukleofil addíció megy végbe ésgyűrűs félacetál (ciklofélacetál) képződik.

A szénhidrátok egyensúlyi elegyében anyílt láncú forma és a ciklofélacetálszerkezet (laktolgyűrű) öt- vagyhattagú gyűrűs állapotban található →Haworth-Böeseken-képlet (furanóz,illetve piranóz, a megfelelőheterociklusok analógiájára): IRspektroszkópiával ugyanis nemmutatható ki a karbonilcsoport, tehátspontán, reverzibilis intramolekulárisnukleofil addícióval ciklo-félacetálokként vannak jelen.A laktolgyűrű 5-tagú (furanóz) vagyhattagú (piranóz).A laktolgyűrű kialakulásakor a C-1(ketózoknál C-2) szénatom iskiralitáscentrummá válik és az ígyfellépő két epimert a cukrok körébenanomereknek nevezzük, és -, illetve-jelzéssel különböztetjük megegymástól.

Egyszerű cukrok
(monoszacharidok)
Gyűrűs félacetál
szerkezet
A: Új aszimmetria centrum:
anomer szénatom
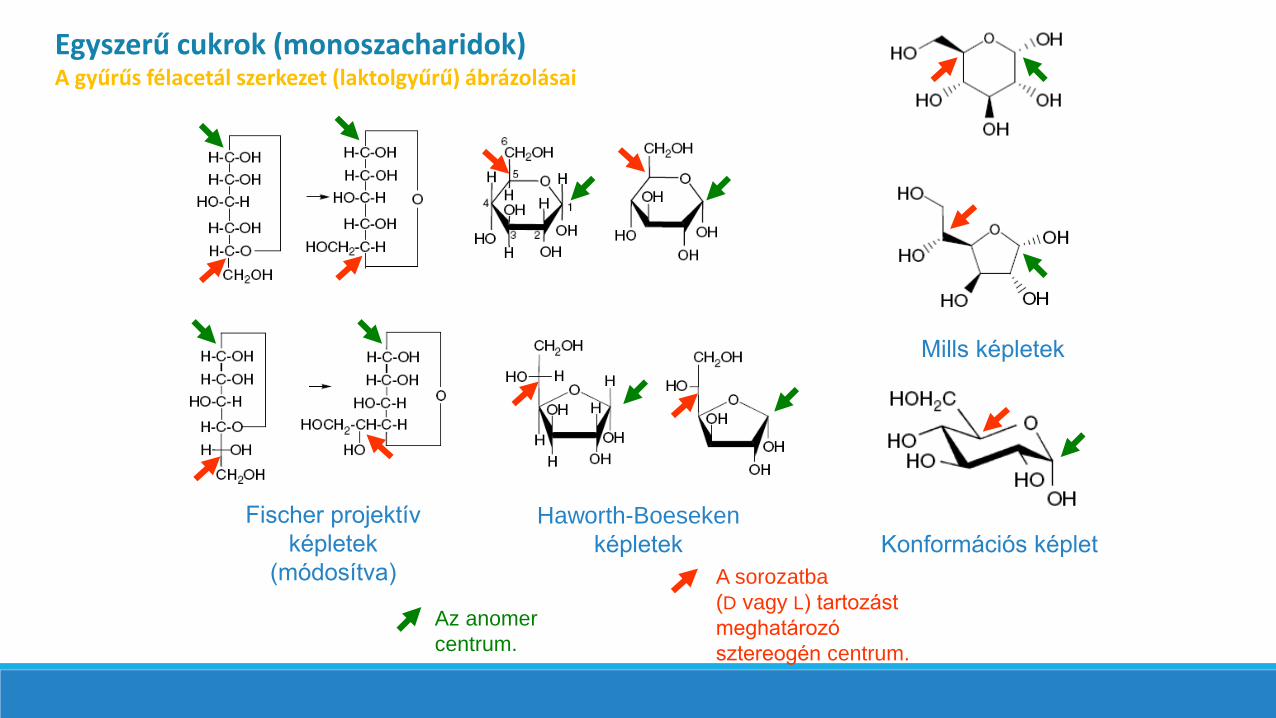
Egyszerű cukrok (monoszacharidok)A gyűrűs félacetál szerkezet (laktolgyűrű) ábrázolásai
Fischer projektív
képletek
(módosítva)
Haworth-Boeseken
képletek
Mills képletek
Konformációs képlet
A sorozatba
(D vagy L) tartozást
meghatározó
sztereogén centrum.
Az anomer
centrum.
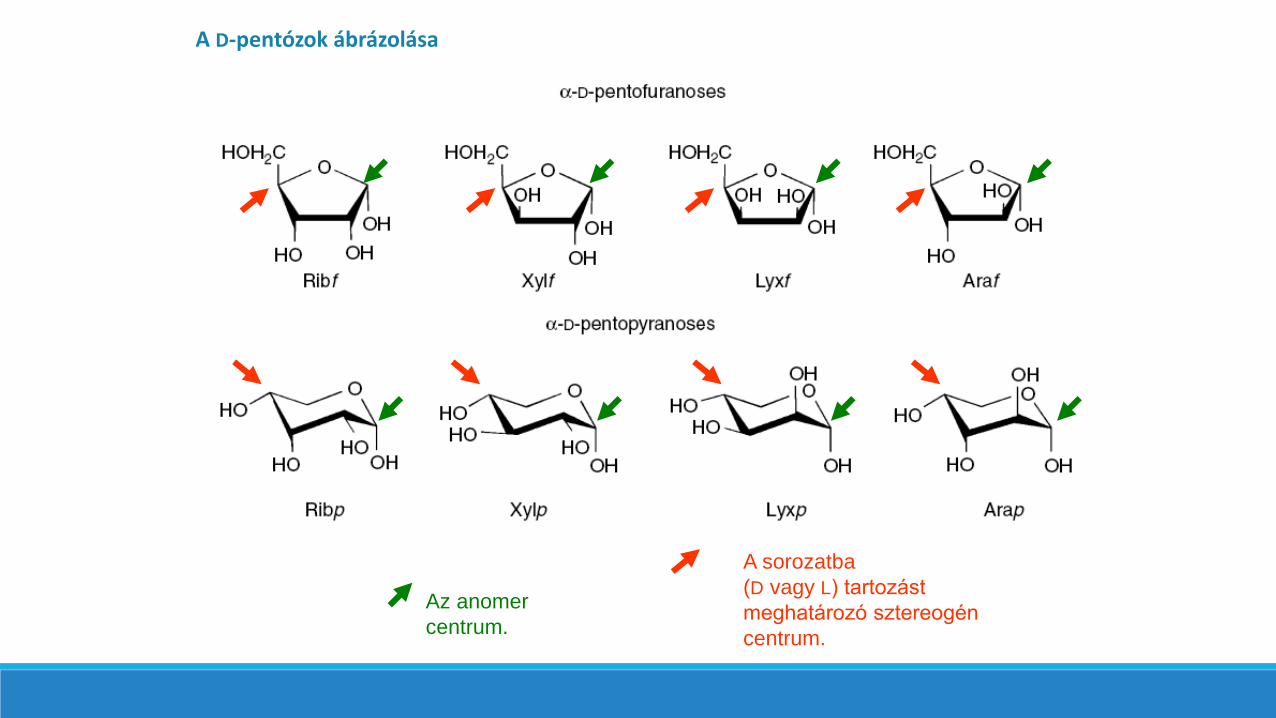
A D-pentózok ábrázolása
Az anomer
centrum.
A sorozatba
(D vagy L) tartozást
meghatározó sztereogén
centrum.

A D-hexózok ábrázolása
Az anomer
centrum.
A sorozatba
(D vagy L)
tartozást meghatározó
sztereogén centrum.

Fischer projekció, projektív képlet: Molekulák térszerkezetének síkbeli ábrázolása, melynek előállítása során szigorúan meghatározott vetítési szabályok alkalmazásával biztosítjuk az egy-egy értelmű megfelelést.
Haworth-Boeseken képlet: A szénhidrátok gyűrűs formáinak ábrázolására alkalmazott perspektivikus képlet, ahol a gyűrű atomjait egy síkban levőnek tételezzük fel, amely az ábrázolás síkjára merőlegesen helyezkedik el. A szubsztituensek agyűrű síkja alatt és fölött találhatók.
Mills képlet: A szénhidrátok gyűrűs formáinak ábrázolása oly módon, hogy a gyűrű a papír síkjában van, és az e sík elé, illetve mögé mutató kötésirányokat vastagított, illetve szaggatott vonallal jelöljük. Előnyös pl. több gyűrűt tartalmazó származékok esetén.
Konformációs képlet: A szénhidrátok tényleges térszerkezetét a legjobban visszaadó perspektivikus ábrázolás.
Anomer szénatom: Egyszerű cukrok laktolgyűrűs formájának képződésekor keletkező új aszimmetria centrum.
Anomer: Az anomer szénatom konfigurációjában különböző epimer monoszacharid származékok.
Fogalmak

Mutarotáció
Mutarotáció: (Szó szerint: az optikai forgatóképesség változása) Szacharidok esetén az anomerek egymásba alakulását (egészen pontosan a mutarotációs egyensúlyi elegy kialakulását) kísérő forgatóképesség változás.
22
Az anomerek keletkezésére (és tulajdonképpen a gyűrűs szerkezet kialakulására) a mutarotáció jelensége hívta fel afigyelmet. A kristályosítás módjától függően kétféle forgatóképességű D-glükóz (vagy egyéb cukor) ismeretes.
jégecet oldat alkohol
[] = + 112,2° [] = +52,7o [] = + 18,7o
-D-glükóz -D-glükóz
A mutarotáció jelensége úgy értelmezhető,hogy akármelyik anomerből is indulunk ki,oldatban a ciklofélacetál-gyűrű felnyílik és anyíltláncú alakon keresztül a piranóz ésfuranóz enantiomerek egymásba átalakulnakadott egyensúlyi állapot eléréséig. Amutarotáció jelensége nemcsak a D-glükózoldatára jellemző, hanem minden másegyszerű szénhidrátéra is.
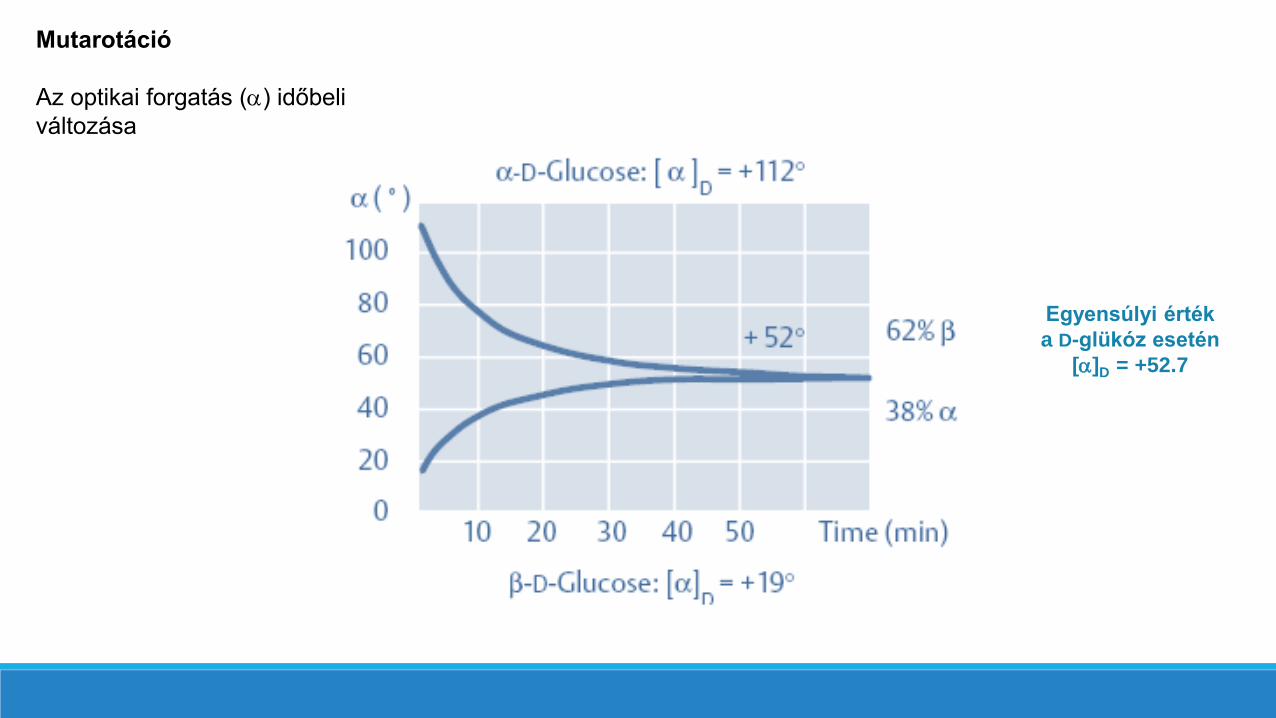
Mutarotáció
Az optikai forgatás () időbeli
változása
Egyensúlyi érték
a D-glükóz esetén
[]D = +52.7

Mutarotációs egyensúlyi elegyek összetétele

A piranóz gyűrű konformációi
A szacharidokban C–O–C kötésszög és kötéstávolság csak kissé tér el a C–C–C fragmensétől, ezért a konformációs
sajátságaik hasonlóak a ciklohexánéhoz, illetve a ciklopentánéhoz.
A C, H, S és B konformációk esetén négy atom, az E esetén pedig öt atom egy síkban van.

Boríték
(Envelope)
Csavart forma
(Twist)
A furanóz gyűrű konformációi

A piranózok téralkata székalkatú, a két lehetséges szék konformer energiatartalma azonban nem azonos. Így például aD-glükopiranóz -anomerje kizárólag a 4C1 szimbólummal (a felső és alsó index a C-1 és C-4 szénatomok relatív helyzetéreutal) jelzett szék konformációban fordul elő, mivel ebben a nagy térkitöltésű hidroximetil- és valamennyi hidroxilcsoport azenergetikailag kedvező ekvatoriális helyzetben van. A másik szék konformáció (4C
1) nem létképes, mivel ez esetben a
nevezett csoportok mindegyike axiális helyzetbe kerülne, és az így fellépő 1,3-diaxiális kölcsönhatások () a molekula
számottevő energianövekedését eredményeznék.
O
OH
O
OH
OH
OH
H1
23
-D-glükopiranóz
O
OH
OH
OH
CH2OH
O
HH
4
H
4C1 4C1
O
OH HO
HO
OH
HOH
4C1 konformer 4C1 konformer
(stabilabb)
ae OOO

Anomer effektus
Az -anomer esetében is a 4C1-konformáció a kedvezményezett, jóllehet ilyenkor a glikozidos hidroxilcsoport már nemekvatoriális, hanem axiális állású. A 4C1-konformációban a glikozidos hidroxilcsoport és a C-3, valamint a C-5 szénatomokhozkapcsolódó hidrogénatomok közelsége (vagyis az 1,3-diaxiális kölcsönhatás) a molekula energiatartamát növeli. Azenergiatartalom növekedését az ún. anomer effektus, mint energianyereség viszont ellensúlyozza.Az anomer effektus ugyanis azt jelenti, hogy a gyűrű oxigénatomjának axiális térhelyzetű nemkötő elektronpárja (mint HOMO-pálya), kölcsönhatásba lép a vele azonos síkban levő glikozidos OH-csoport szén-oxigén kötésének lazító pályájával (mintLUMO-pálya). A megfelelő előjelű pályarészek átlapolása jelentős energia nyereséggel jár (delokalizációs modell).
-D-glükopiranóz
O
OH
HO HO
OH
OH
H HH O
C
O
n
*
E
n
*
C5 O C1
HOMO
LUMO
C1 OH

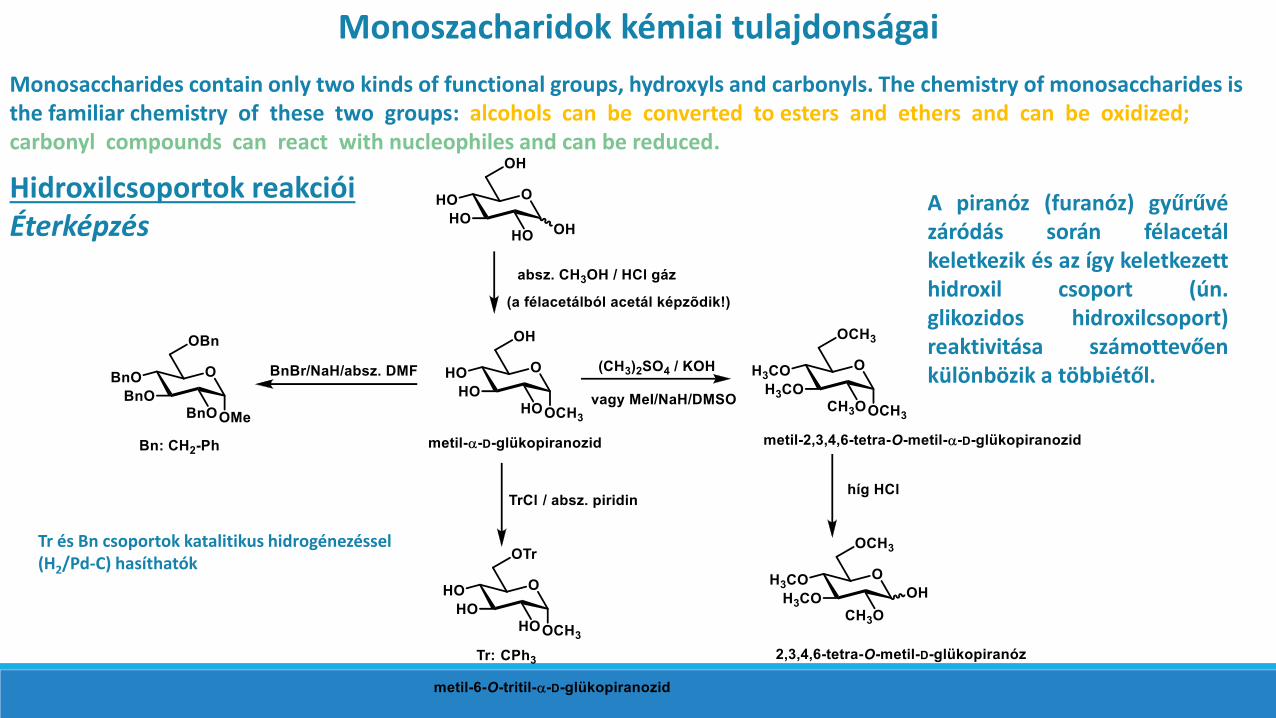
Monoszacharidok kémiai tulajdonságai
Monosaccharides contain only two kinds of functional groups, hydroxyls and carbonyls. The chemistry of monosaccharides is the familiar chemistry of these two groups: alcohols can be converted to esters and ethers and can be oxidized; carbonyl compounds can react with nucleophiles and can be reduced.
Hidroxilcsoportok reakcióiÉterképzés
A piranóz (furanóz) gyűrűvézáródás során félacetálkeletkezik és az így keletkezetthidroxil csoport (ún.glikozidos hidroxilcsoport)reaktivitása számottevőenkülönbözik a többiétől.
Tr és Bn csoportok katalitikus hidrogénezéssel(H2/Pd-C) hasíthatók

H O CH2 PhBr2 / h
H O CH Ph
Br
OHH O CH Ph
OH
H / H2O
H OH PhCHO
A benzilcsoport eltávolítás történhet a következőképpen is:

Észterképzés

Zemplén-féle dezacetilezés és mechanizmusa

Szulfonátok előállítása és átalakítási lehetőségeik

Acetál védőcsoportok
Vicinális cisz-helyzetű hidroxilcsoportot tartalmazó cukrokból acetonnal savkatalizált reakcióban gyűrűs izopropilidénszármazékok, ún. acetonidok állíthatók elő.
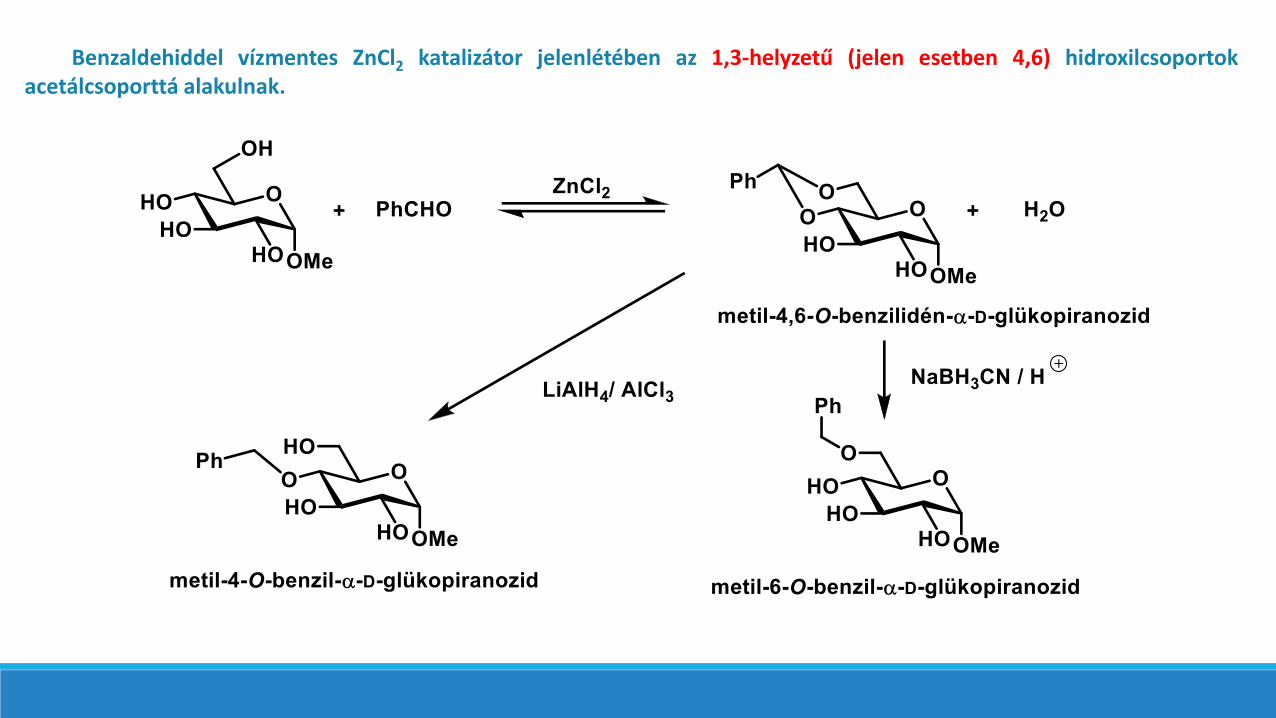
Benzaldehiddel vízmentes ZnCl2 katalizátor jelenlétében az 1,3-helyzetű (jelen esetben 4,6) hidroxilcsoportokacetálcsoporttá alakulnak.
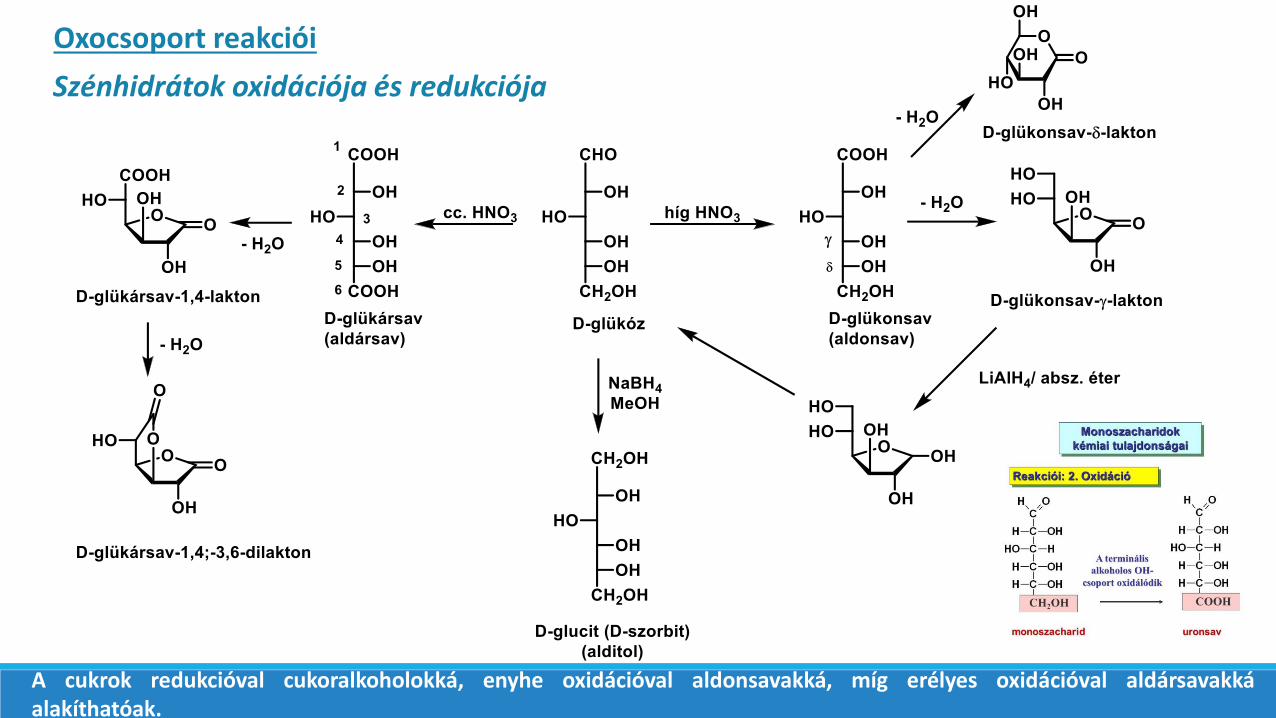
Szénhidrátok oxidációja és redukciója
Oxocsoport reakciói
A cukrok redukcióval cukoralkoholokká, enyhe oxidációval aldonsavakká, míg erélyes oxidációval aldársavakkáalakíthatóak.

Tollens és Fehling próbaEzüst tükör póba: az aldózok az Ag(NH3)2
+ oldatából fém ezüstöt választanak ki miközben aldonsavakká oxidálódnak.
Fehling póba: az aldózok Cu(II)SO4-nak K-Na-tartaráttal képzett komplexéból vörös színű Cu2O-t választanak le miközbenaldonsavakká oxidálódnak.
K-Na-tartarát
D-glükóz endiol D-mannóz
D-fruktóz
CO
H
CH OH
CO
H
CHO H
CH2
C O
OH
C OH
CH OH
Epimerizáció

Nukleofilekkel lejátszódó reakciókOszazon képződés
Wohl – Zemplén lebontás

Kiliani- Fischer féle lánchosszabbítás
OCH2HO
OH
OH
2-dezoxi-D-ribóz
L-ramnóz6-dezoxi-L-mannóz
O CH3
HOOH
OH
OH
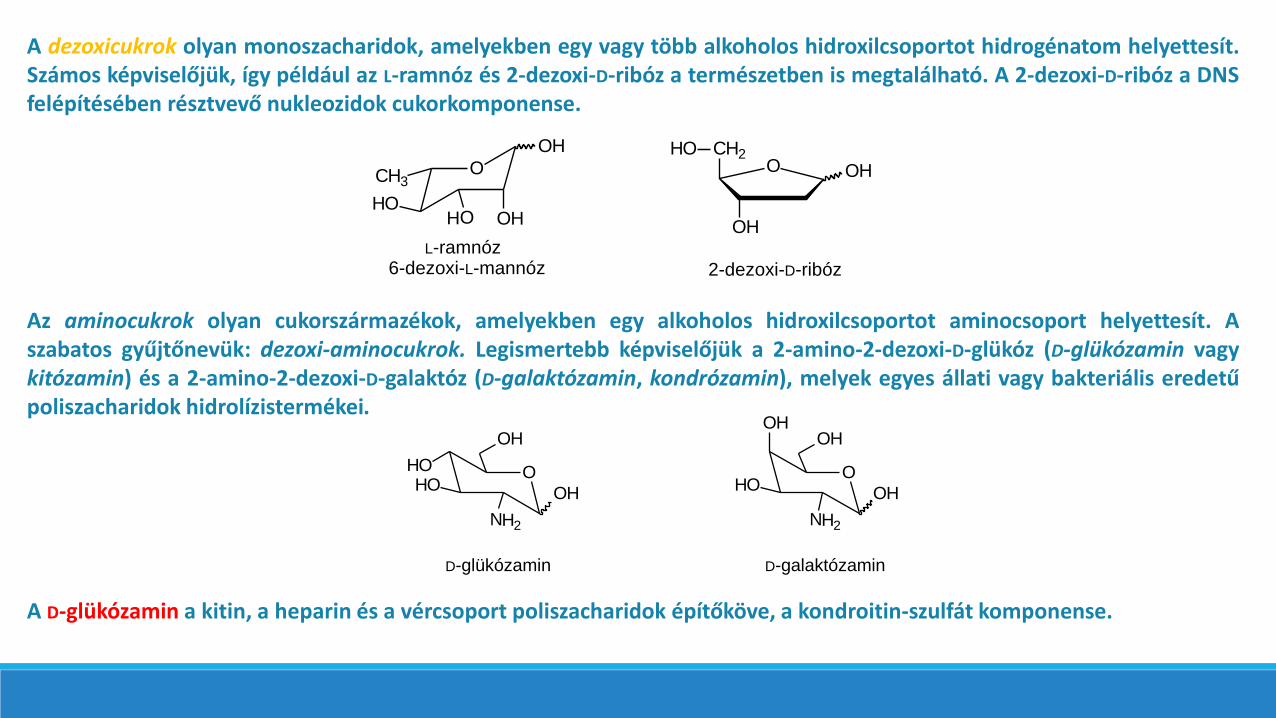
A dezoxicukrok olyan monoszacharidok, amelyekben egy vagy több alkoholos hidroxilcsoportot hidrogénatom helyettesít.Számos képviselőjük, így például az L-ramnóz és 2-dezoxi-D-ribóz a természetben is megtalálható. A 2-dezoxi-D-ribóz a DNSfelépítésében résztvevő nukleozidok cukorkomponense.
Az aminocukrok olyan cukorszármazékok, amelyekben egy alkoholos hidroxilcsoportot aminocsoport helyettesít. Aszabatos gyűjtőnevük: dezoxi-aminocukrok. Legismertebb képviselőjük a 2-amino-2-dezoxi-D-glükóz (D-glükózamin vagykitózamin) és a 2-amino-2-dezoxi-D-galaktóz (D-galaktózamin, kondrózamin), melyek egyes állati vagy bakteriális eredetűpoliszacharidok hidrolízistermékei.
D-glükózamin
O
OHHO
HO
OH
N
O
OH HO
OHOH
N
D-galaktózamin
H2H2
A D-glükózamin a kitin, a heparin és a vércsoport poliszacharidok építőköve, a kondroitin-szulfát komponense.

Fontos monoszacharidokHumans need to obtain eight monosaccharides for proper functioning. Although it can be biosynthesized from simplerprecursors if necessary, it’s more energetically efficient to obtain them from the diet.The eight are: L-fucose (6-deoxylgalactose), D-galactose, D-glucose, D-mannose, N-acetyl-D-glucosamine, N-acetyl-D-galactosamine, D-xylose, and N-acetyl-D-neuraminic acid
All are used for the synthesis of the glycoconjugate components of cell walls, and glucose is also the body’s primary source ofenergy.

Diszacharidok
Redukáló és nem redukáló diszacharidok
A redukáló diszacharidok adják az ezüst tükör és a Fehling próbát.

Nem redukáló diszacharid: Nem található bennük szabad glikozidos hidroxilcsoport.
Redukáló diszacharid: Szabad glikozidos hidroxilcsoportot tartalmaznak.
Nem redukáló vég: Oligo/poliszacharid láncok végén található, szabad glikozidos hidroxilcsoportot nem tartalmazó monoszacharid egység.
Redukáló vég: Oligo/poliszacharid láncok végén található, szabad glikozidos hidroxilcsoportot tartalmazó monoszacharidegység.
Oligoszacharid: Glikozidos kötésekkel összekapcsolódó, 2-20 azonos vagy eltérő monoszacharidból álló vegyületek gyűjtő-neve. Lehetnek egyenes vagy elágazó láncúak.
Diszacharid: Glikozidos kötéssel összekapcsolódó, 2 monoszacharidból álló vegyületek gyűjtőneve. Lehet redukáló és nem redukáló.
Fogalmak

A legjelentősebb nem redukáló diszacharid a szacharóz (répa- vagy nádcukor).
A szacharóz híg ásványi savakkal vagy enzimatikus úton D-glükózra és D-fruktózra hidrolizál. A hidrolízis során acukoroldat forgatóképessége folyamatosan csökken, eléri a nulla értéket, majd balra forgatóvá válik, mivel a fruktózerősebben forgat balra, mint a glükóz jobbra. A forgatóképesség előjelének megváltozása miatt a szacharóz hidrolízisétinvertálásnak, és az így nyert egyszerű cukor keveréket invertcukornak („műméz”) nevezik.A mézhamisítás leggyakoribb módja a virágmézként árult műméz. A műméz invertcukorból aromás anyagok (néhazamatos méz) hozzákeverésével és festékanyagok színezésével készül. A hamisítást könnyen felismerhetjük, ha egykávéskanálnyi mézet tiszta szeszben feloldunk. Amennyiben teljesen oldódik, úgy a méz tiszta. Ellenkező esetbenhomályos lesz és rövid idő múlva a belekevert anyag leülepedik.
Szacharóz, invertcukor, műméz

Maltóz, cellobióz és laktózA redukáló diszacharidok közül a maltóznak (malátacukor), a cellobióznak és laktóznak (tejcukor) van gyakorlati
jelentősége.
O
OH HO
OH
HO
O
HHO
HO
OH
HO
O
/-maltóz
A cellobióz [4-O-(-D-glükopiranozil)-D-glükóz] a természetben legelterjedtebbszénvegyületnek, a cellulóznak azépítőköve; innen származik a neve is.Savas hidrolízisekor hasonlóan amaltózhoz, kizárólag D-glükóz képződik. Ekét vegyület szerkezete a glikozidkötés -konfigurációját kivéve azonos.
A maltóz [4-O-(-D-glükopiranozil)-D-glükóz]a keményítő és a glikogén alkotórésze. Atermészetben szabad állapotban mindigolyan növényi részekben jelenik meg,amelyekben a keményítő enzimeshidrolízise játszódik le (pl.burgonyacsírában, zöld levelekben stb.).Előállítása is a keményítő enzimeshidrolízisével történik. A maltóz kétglükózegységet tartalmaz, mégpedig piranózformában, és a glikozil komponens -glikozidos kötéssel kapcsolódik a másik D-glükóz (aglikon) C-4-es oxigénatomjához.
O
OH HO
OHOHO
HO
OH
O
HO
HOH
-cellobióz
A laktóz [4-O-(-D-galaktopiranozil)-D-glükóz] az emlősök tejénekcukorkomponense; az anyatej 5-8%laktózt (tejcukrot) tartalmaz.Szerkezetileg a cellobiózzal rokon: aD-glükóz C-4 hidroxilcsoportjához ezesetben -D-galaktozilcsoportkapcsolódik.
O
OH HO
OHOHO
H
OH
O
OH
HO
HO
H
,-laktóz

Diszacharidok szintézise Koenigs-Knorr módszerrel
Gentiobiose is a disaccharide composed of two units of D-glucose joined with a β(1->6) linkage. It is a white crystallinesolid that is soluble in water or hot methanol. Gentiobiose isincorporated into the chemical structure of crocin, thechemical compound that gives saffron (sáfrány) its color. It is aproduct of the caramelization of glucose.
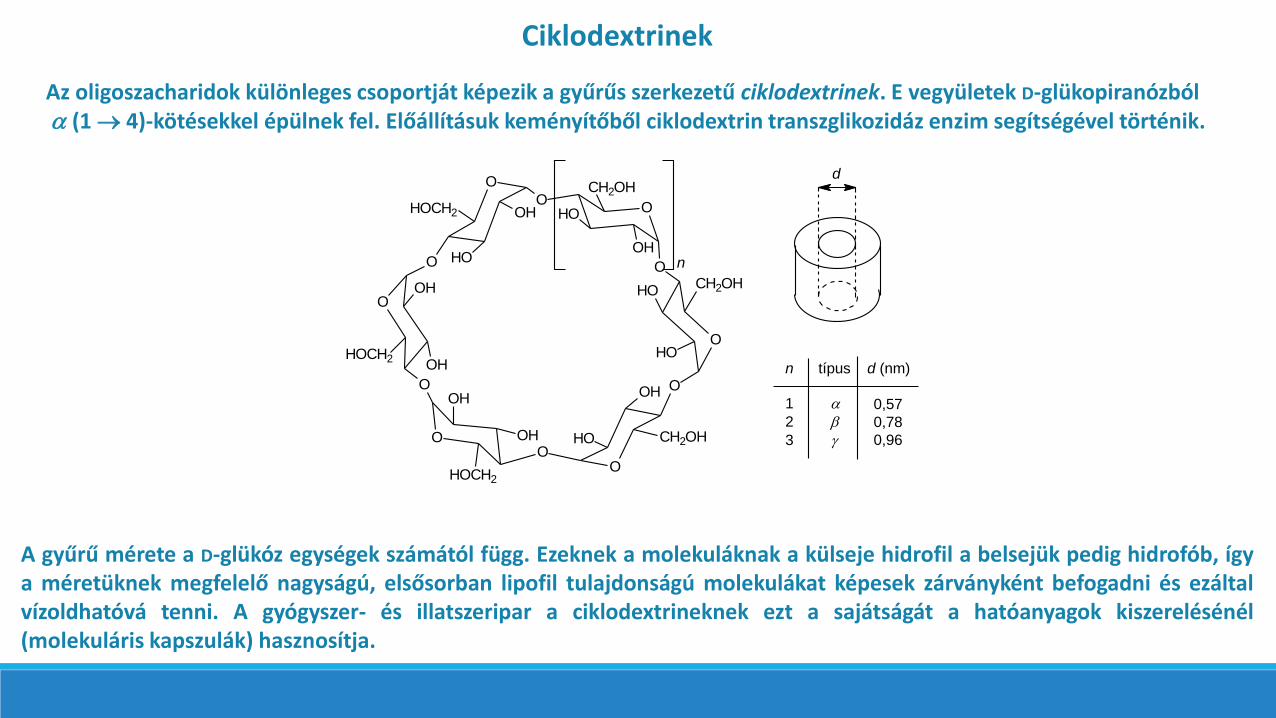
Az oligoszacharidok különleges csoportját képezik a gyűrűs szerkezetű ciklodextrinek. E vegyületek D-glükopiranózból (1 4)-kötésekkel épülnek fel. Előállításuk keményítőből ciklodextrin transzglikozidáz enzim segítségével történik.
Ciklodextrinek
CH2OH
O
CH2OH
O
O
HO
HO
O
OH
OH
O
HOCH2
O
OHO
OH
HOCH2
O
HO
OHO
HOCH2
O OH
OH
OCH2OH
O
OH
HO
n
d
n típus d (nm)
1
2
3
0,57
0,78
0,96
A gyűrű mérete a D-glükóz egységek számától függ. Ezeknek a molekuláknak a külseje hidrofil a belsejük pedig hidrofób, ígya méretüknek megfelelő nagyságú, elsősorban lipofil tulajdonságú molekulákat képesek zárványként befogadni és ezáltalvízoldhatóvá tenni. A gyógyszer- és illatszeripar a ciklodextrineknek ezt a sajátságát a hatóanyagok kiszerelésénél(molekuláris kapszulák) hasznosítja.

Ciklooligoszacharidok
Ciklodextrinek

Ciklodextrinek


Az anyatej oligoszacharidjai (human milk oligosaccharides, HMO)Prebiotikus potenciál– Nem szívódik fel és ellenáll a gyomor és bélrendszerben az enzimatikusbontásnak– A bél mikroflóra fermentálni tudja– Szelektíven stimulálja a jótékony hatású bélbaktériumok növekedését.
Az anyatej oligoszacharidjai (HMO)
Ezáltal • gátolja a kórokozók megtelepedését (adhéziós és nutritívkompetíció, baktericidek termelése) • erősíti a bél védelmi funkcióját – SCFA termelés (tápanyag az epithelsejtjeinek) jobb epithel funkciók • támogatja a bélhez kapcsolódó immunrendszert
Kb. 130 féle oligoszacharid struktúrát tudtak kimutatni Bode L. Nutrition Reviews 2009, 67(Suppl. 2):S183–S191



A poliszacharidok monoszacharid-részekből felépített nagy molekulatömegű vegyületek. Általában 80-10000monomer egységből állnak. Fizikai tulajdonságaikban jelentősen eltérnek a mono- és oligoszacharidoktól.Nem édes ízűek, vízben nem vagy csak kolloidálisan oldódnak. A poliszacharidok szerkezeti elve ugyanaz, mintaz oligoszacharidoké, ezért érthető, hogy savas hidrolízisük során monoszacharidokra esnek szét. Sokpoliszacharid hidrolízisével csak egyetlen monoszacharid-féleség keletkezik, de ismerünk olyanpoliszacharidokat is, amelyeknek a hidrolitikus lebontása kétféle monoszacharidhoz vagy monoszacharidszármazékhoz (pl. uronsavhoz) vezet.
Poliszacharidok
Kizárólag D-glükóz egységekből épül fel három nagyon fontos és elterjedt poliszacharid: a cellulóz, a keményítőés a glikogén.

A cellulóz a Földön a legnagyobb mennyiségben előforduló szénvegyület, a magasabb rendű növények sejtfalának főalkotórésze. A levelek szárazanyagának 10-20%-át, a fás növényi részek 50%-át alkotja. A cellulóz átlagos polimerizációsfoka az eredettől függően 3000 és 8000 között van. Savas hidrolízise (tömény sósav hidegen) csak D-glükózhoz vezet. Azegyik glükóz egység -glikozidos kötéssel kapcsolódik a másik egység C-4 helyzetű hidroxilcsoportjához, és így lineárisszerkezetű fonalstruktúra alakul ki.
n
n = 3000-8000cellulóz
HOCH2 O
OH
H
O
OH HOCH2 O
OOH
O
HOCH2 O
OH
H
O
OH HOCH2 O
OOH
O
A fonalstruktúrájú cellulózmolekulákat az intramolekuláris hidrogénkötések merevítik, ugyanakkor az intermolekulárishidrogénhidak pedig a hosszú fonalak szoros egymáshoz illeszkedését biztosítják. Ez rendkívüli stabilitást, rugalmasságot,valamint oldhatatlanságot eredményez és ezáltal a cellulóz ideális alapanyaga a növények sejtfalának.
Szerkezeti (váz) poliszacharidok – cellulóz (növények)
O
AcNHHO
HO
OH
O
AcNHHOO
OH
OH
O
OH
HOAcNH
OHO
AcNHHO
HO
OH
O1
4
4
1
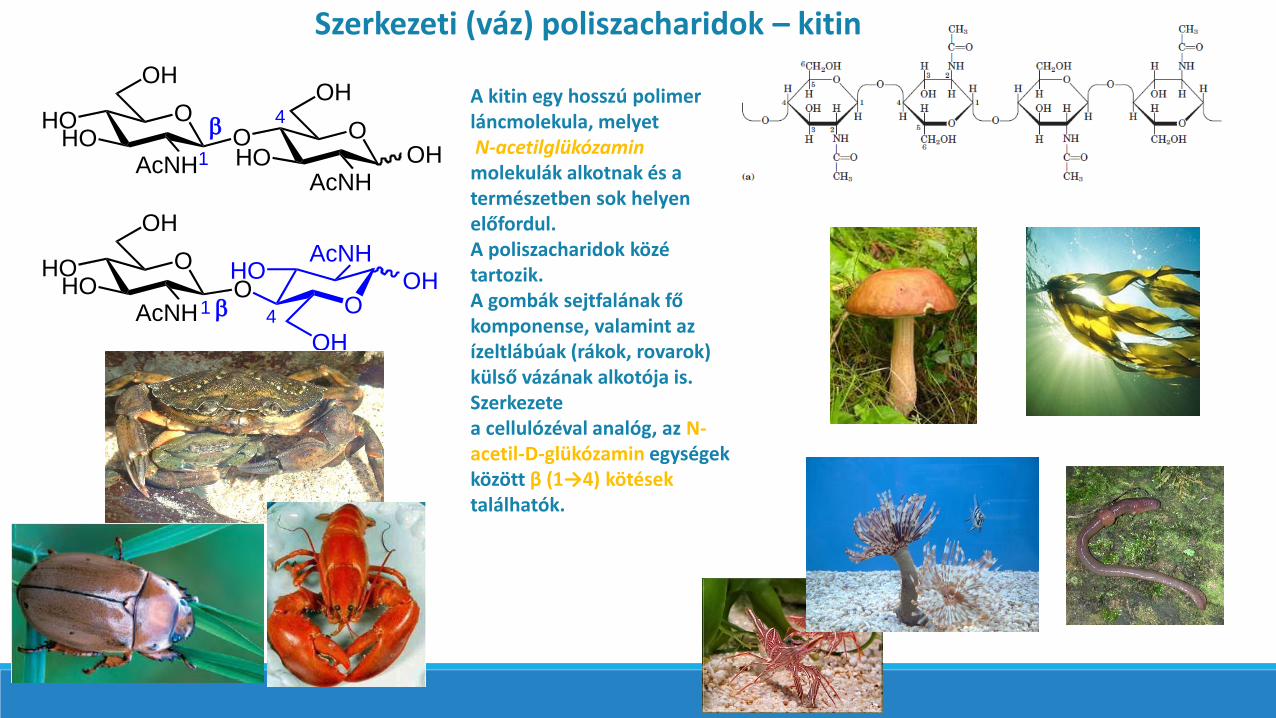
Szerkezeti (váz) poliszacharidok – kitin
A kitin egy hosszú polimer láncmolekula, melyetN-acetilglükózaminmolekulák alkotnak és a természetben sok helyen előfordul. A poliszacharidok közé tartozik. A gombák sejtfalának fő komponense, valamint az ízeltlábúak (rákok, rovarok)külső vázának alkotója is. Szerkezete a cellulózéval analóg, az N-acetil-D-glükózamin egységekközött β (1→4) kötések találhatók.

Typical levels of pectin in plants are (fresh weight):
apples 1–1.5%, apricot 1% , cherries 0.4%, oranges 0.5–3.5%,
carrots approx. 1.4%, citrus peels, 30%.
One of the
most promi-
nent charac-
teristics of
pectins is their
ability to form
gels at
concentrations
as low as 0.3–
0.7% (w/v).
Szerkezeti (váz) poliszacharidok – pektin (gyümölcsök)
Pectins are polysaccharides particularly prevalent in fruits such as apple pulp (pép) (10–15%) and orange and lemon rinds(héj) (20–30%). Pectins are composed of D-galactopyranuronic acid units linked α-1→4.
O
HOHO
OCOOH
O
HOHO
OCOOH
O
HOHO
OCOOH
O
Az élelmiszeriparban elsősorban gyümölcszselékben és lekvárokban, E440 néven széles körben alkalmazzák. Előfordulhat továbbá gyümölcsitalokban, tejtartalmú élelmiszerekben, zselés cukorkákban, valamint egyes diétás ételekben.
A pektint az élelmiszeriparban zselésítőanyagként, térfogatnövelőként,valamint stabilizálószerként alkalmazzák. A legtöbb estben lekvárok,dzsemek zselésítésére használják, így javítva annak minőségét. Házibefőzéshez cukorral keverve kapható (egy kevés citromsav hozzáadásávala pH-egyensúly megtartása miatt). Egyes országokban a befőzéshez a pektinttiszta, porított állapotban lehet beszerezni. A hagyományos jellegű lekvár(60%-nál nagyobb cukor- és gyümölcstartalom) előállításához magasésztertartalmú pektint használnak. Alacsony észter-tartalmú pektin alacsonycukortartalmú (diétás) lekvárok készítéséhez ajánlott. Az élelmiszerekben apektint általában 0,5-1,0% koncentrációban alkalmazzák (a frissgyümölcsökben is ilyen arányban található meg).Pékáruk esetén zsírok helyettesítésére használják.Kozmetikumok elsősorban stabilizáló hatása miatt tartalmazzák.
(uronsav!)

A keményítő a cellulóz mellett a másik legelterjedtebb poliszacharid a növényvilágban. A keményítő a széndioxid-asszimiláció terméke, amely keletkezésének helyén a növényi nedvekben feloldódik és más növényi részekbe (pl. gumóba,gyökérbe) vándorol, és ott mint tartaléktápanyag szemcsés formában raktározódik. A növény csökkent asszimilációjaidején a keményítőt tápanyagként hasznosítja. A keményítő polimer szerkezetét tekintve két anyag az amilóz és azamilopektin keveréke. A legtöbb keményítő kb. 10-20% amilózt és 80-90% amilopektint tartalmaz.
CH2OHO
OO
HOCH2
O
O
O
HOCH2
O
O
O
H
O
O
H
O
HOCH2
OO
O
H
O
H HH
CH2OH
O
O
O
O
H
H O
HOCH2 O
OO
HO
H
CH2OH
O
OH
O
H
O
Az amilopektin a keményítő másik komponense, ezernél több D-glükózegységből épül fel. Szerkezete az amilózéhoz nagyon hasonló. Egyetlen, denagyon lényeges különbség, hogy az (1 4) glikozidos kötéssel a D-glükózbólfelépülő főlánchoz minden 20-25 glükóz egységnél (1 6) glikozidos kötésselújabb „amilózszerű” oldallánc kapcsolódik.
Az amilóz mintegy ezer D-glükopiranózegységből (1 4) glikozidos kötéssel épül fel ésezáltal a hosszú lánc spirális alakú hélixet képez,melyet intramolekuláris hidrogénkötések tartanakössze.
A keményítő Tartaléktápanyag poliszacharidok – keményítő és glikogén
Asszimiláció (anabolizmus) az a folyamat, amelynek során az élő szervezetek a környezetükből felvett anyagokból saját testük anyagait felépítik.

A glikogén az emberi és állati szervezetben ugyanazt a szerepet tölti be, mint a keményítő a növényi szervezetben.Tartalékszénhidrát szerepe miatt állati keményítőnek is nevezik. A szervezetben a glikogén az izmokban és főleg a májbanhalmozódik fel és az ún. glikolízis folyamán D-glükozil-foszfáttá való lebontást követően aerob (O2 jelenlétében) körülményekközött szén-dioxiddá és vízzé ég el, illetve anaerob módon L-(+)-tejsavvá alakul át.
OHO
HO
OH
HOOPO3H2
D-glükóz-1-foszfát
CO2 + H2O
glikogén
COOH
HO H
CH3
A szerkezete az amilopektinéhez hasonló, azonban a glikogénben – lévén hogy minden 10-12 glükózegységre esik egy-egy láncelágazás – a láncok sokkal elágazóbbak.
A glikogén
tejsav

Monoszacharidok bioszintézisének útvonalai

Monoszacharidok bioszintézisének kulcsreakciói
etil acetoacetát (ha 2 etil-acetát molekula reagál egymással)
(pl. ha 2 aldehid reagál egymással)

The light-independent reactions of photosynthesisare chemical reactions that convert carbondioxide and other compounds into glucose.Despite its name, this process occurs only whenlight is available. Plants do not carry out the Calvincycle by night.
Monoszacharidok bioszintézise –a Calvin-ciklus

H2C
C
HC
HC
H2C
O
O
OH
OH
O
P
P
H2C
C
C
HC
H2C
OH
O
OH
OH
O
P
P
P = P(=O)OH2
O
C
O
+
H2C
C
C
HC
H2C
OH
O
O
OH
O
P
P
OOC
H2C
C
C
HC
H2C
OH
O
O
OH
O
P
P
HOOC
H HHO
H2C
C OH
O P
HOOC
C
HC
H2C
O
OH
O P
H
HO+
COOH
HC
H2C
OH
O PCHO
HC
H2C
OH
O P
H2C
C
H2C
O
O P
OH
Fruktóz-1,6-biszfoszfát
CH
C
H2C
O
O P
HC
HC
H2C
OH
O P
OH
HO
Ribulóz-1,5-biszfoszfát
i: dihidroxiaceton-foszfát ii: glicerinsav-3-foszfátiii: glicerinaldehid-3-foszfát
i
ii
iii
Monoszacharidok bioszintézise – a CO2 fixálásának első lépései

A fotoszintézis fényreakciói során keletkező ATP és NADPH aszéndioxid fixálás folyamatában hasznosul. Mivel ezen folyamatoknem függenek közvetlenül a fény jelenlététől, a fotoszintézissötétreakcióinak is nevezik őket. A sötétreakciók a sztrómábanjátszódnak. le. A CO2 fixálás alapfolyamata az ún. Calvin ciklus, ami anövények jelentős részében a CO2 megkötés Melvin Calvin és mtsai.által feltárt kizárólagos mechanizmusa. A Calvin ciklus folyamánkeletkező elsődleges termékek 3 szénatomot tartalmaznak, ezért aszéndioxidot ezen mechanizmus alapján kötő növényeket C3-asnövényeknek nevezzük. A Calvin ciklus kulcsenzime a Rubisco(ribulóz-1,5-biszfoszfát-karboxiláz-oxigenáz), ami a tilakoid membránsztróma felöli oldalához tapad (azaz nincs a membránba ágyazódva).A ciklus első lépésében a ribulóz 1,5-biszfoszfát (RuBP) CO2-t köt meg,majd egy 6 szénatomos átmeneti termék képződése után 2 molekulaglicerinsav-3-foszfátra (3PG) esik szét. Ezek egy-egy ATPfelhasználásával glicerinsav-1,3-biszfoszfáttá alakulnak. Ezt afolyamatot a 3-foszfoglicerát kináz enzim katalizálja. A folyamatkövetkező lépésében glicerinaldehid-3-foszfát (G3P) keletkezik,NADPH felhasználásával. Így a megkötött széndioxid szénhidrát szintigredukálódott. A folyamat során 6 CO2 molekula megkötésénekeredményeként 12 molekula glicerinaldehid-3-foszfát keletkezik.Közülük kettő továbbalakul glukózzá, aminek átmeneti terméke afruktóz-1,6-biszfoszfát. A többi, ATP felhasználásával a Rubiscoregenerálására fordítódik.

Oligoszacharidok bioszintézise
Szacharóz Keményítő: A glükóz anomer centrumának aktiválása ADP-veltörténik. Az újabb monoszacharid egységek a lánc redukáló(!) végére épülnek be.
Glikogén: A glükóz anomer centrumának aktiválása UDP-vel történik. Az újabb monoszacharid egységek a lánc nem redukáló(!) végére épülnek be.
Szacharóz képződése UDP-vel aktivált D-glükóz anomercentrumán lejátszódó nukleofil szubsztitúcióval.
Kérdés: Miért nem változik a konfiguráció?

A glikogén bioszintézise
A glükóz anomer centrumának aktiválása UDP-vel történik. Az újabb monoszacharid egységek a lánc nem redukáló(!) végére épülnek be.

Oligo- és poliszacharidok (ipari) felhasználása – válogatás
Ciklodextrinekanyagok védelme oxidáció és UV-degradáció ellen feldolgozás vagy tárolás során; illat/aromaanyagok, fűszerekstabilizálása; élelmiszer/gyógyszer keserűségének vagy kellemetlen illatának elfedése; folyadékok száraz formába alakítása;vízoldhatóság növelése; szénhidrogének, szteroidok, zsírok, zsírsavak emulzifikálása; gyógyszerek, illat/aromaanyagokkontrollált kibocsátása; kémiai reakciók katalízise; elválasztástechnika (kromatográfia, kapilláris elektroforézis).
Cellulózpapíripar, textilipar (pamut, len, stb), „nitrocellulóz” (cellulóz-nitrát; lőgyapot, celluloid); cellulóz-acetát (szigetelő filmek,lakkok, ragasztók, szálak); viszkóz (műselyem, cellofán).
Keményítőélelmiszeripar (gélesítés, ételszerkezet módosítása, sörfőzés, stb); textilipar (szálak írezése: felületük lesimítása);gyógyszeripar (hordozó- és kötőanyag); bioetanol gyártás.
Kitingyógyszeripar (hordozó- és kötőanyag, antibakteriális sebkötöző anyagok); biodegradábilis csomagoló anyagok.

Glikokonjugátumok
A cukor és egyéb biomolekulák között kovalens kötéssel létrejövő összetett polimereket nevezzük glikokonjugátumoknak.
Az extracelluláris mátrix egyik főkomponense, a proteoglikánok esetében egyközponti fehérjéhez (core protein)kapcsolódnak GAG (glükózamin-glikán)láncok. A proteoglikánok jelentőségétmutatja, hogy emlősökben legalább 40-félefordul elő, melyek a központifehérjeegységben és a hozzá kapcsolódó GAGláncokban is különböznek. A GAG láncokáltalában tetraszacharid linkeren keresztülkapcsolódnaka polipeptidlánc egyik Ser oldalláncához (O-glikozidos kötéssel).
A baktériumok sejtfalát felépítő egyetlen„óriásmolekulában”, a peptidoglikánokban (régebbineve murein) poliszacharid láncokat kötnek összerövid peptidszakaszok.
A szindekán és glipikán proteoglikánok. Az ábra felső részén a kondroitin-szulfátés a linker oligoszacharid szerkezete látható.
Egy peptidoglikán vázlatos szerkezete
szerin

A glikoproteineknél a polipeptidlánchoz változatos felépítésűoligoszacharidok kapcsolódnak. A sejtmembrán külső oldalántalálhatók (integráns membránfehérjékben csak azextracelluláris oldalon lévő polipeptidlánc részhezkapcsolódnak!), az organellumoknál viszont asejtkompartmentum belseje felé néznek. A cukorrészekváltozatossága teszi lehetővé, hogy mind a proteoglikánok,mind a glikoproteinek információhordozó makromolekuláklehessenek.A glikoproteinekben egy vagy több oligoszacharid lánc (azanomer C-n keresztül) kovalensen kötődik apolipeptidlánchoz. A cukorrész a fehérje tömegének 1-70%-alehet. Az oligoszacharidok kétféle módon kapcsolódhatnak:N-glikozidos kötéssel Asn oldallánchoz (N-kapcsolt), O-glikozidos kötéssel pedig Ser vagy Thr hidroxilcsoporthoz (O-kapcsolt).
Glikoproteinek cukorrészének két kapcsolódási módja
elte.prompt.hu/sites/default/files/tananyagok/.../ch10s04.html
Aszparagin (Asp) Treonin (Thr) Szerin (Ser)

A peptidoglikán (vagy más néven murein), a valódi baktériumok (Eubacteria)sejtfalának egyik összetevőjét alkotó makromolekula; az eukariótákban nemtalálható meg. Benne aminocukrok (N-acetil-glükózamin és N-acetil-muraminsavβ1→4 glikozidos kötéssel összekapcsolt) láncai kapcsolódnak egy alaninból,glutaminból és lizinből vagy diamino-pimelinsavból álló tripeptidhez. Apeptidoglikánok erőt kölcsönöznek és alakot adnak a sejtfalnak. Azősbaktériumok (Archaea) egy hasonló poliszachariddal rendelkeznek, apszeudopeptidoglikánnal, amiben N-acetil-talozaminuronsav található az N-acetil-muraminsav helyett.
GlcNAc Mur2Ac
Szerkezeti (váz) poliszacharidok – peptidoglikán, murein(baktériumok)
Az ábrán a Staphylococcus aureus baktérium sejtfalának szerkezetét mutatja (ez egy gram-pozitív baktérium). A peptidláncok (színes körök) kovalensen kapcsolódnak az N-acetilmuraminsav egységhez a szomszédos poliszacharid láncban.
Az eukarióták (Eukaryota) olyan élőlények, amelyek valódi sejtmaggal rendelkező sejtekből állnak.A prokarióták, más néven elősejtmagosok vagy sejtmag nélküliegysejtűek (Prokaryota vagy Monera) egysejtű – ritka esetekben többsejtű –,körülhatárolt sejtmag nélküli élőlények.

A kötőszövetek egyfajta hálózatos szerkezetből és sejt közötti állományból állnak. A legfőbb alkotórészei a kollagének,melyek kollagén rostokat hoznak létre, az elasztin, mely rugalmas rostokat hoz létre és a glükozamin-glikánok (GAG) melyeka mátrix alapját alkotják.A rugalmas rostok szerteágazó struktúrát alkotnak a köztes tereket pedig proteoglikánok töltik ki. A kötőszövet jellegzetesszerkezeti egységét tehát a proteoglikánok jelentik, melyek fehérjéből és cukrokból épülnek fel és nagy vízmegkötőképességgel rendelkeznek.
Proteoglikánok: glikóz-aminoglikánok + extracelluláris proteinek
Glikánoknak nevezzük az egyféle cukor alkotta láncokat, homopolimereket (melyek glikozidos kötést tartalmaznak).

Proteoglycan aggregate of the extracellular
matrix
One very long molecule of hyaluronate is
associated noncovalently with about 100 molecules
of the core protein aggrecan. Each aggrecan
molecule contains many covalently bound
chondroitin sulfate and keratan sulfate chains. Link
proteins situated at the junction between each core
protein and the hyaluronate backbone mediate the
core protein–hyaluronate interaction.

A proteoglikánok poliszaharidláncait a glükózamin és ezek N-acetil és szulfát származékai alkotják. A legfontosabb glükózamin-glikánok a heparin, kondroitin-szulfát, keratán-szulfát, dermatán-szulfát és a hilauronsav. Izületeink szempontjából akondroitin-szulfát és a keratán-szulfát ami figyelmet érdemel. E szulfát tartalmú porc-proteoglikánok (kondroitin-szulfát)aggregációra képesek, vízkötésük 50-szerese a szárazanyag tartalmuknak. Ám nem egyforma mértékben. Az öregedés és azizületek túlterhelése során e makromolekulák szerkezete megváltozik. A keratán-szulfát tartalom nő és egyre csökken akondroitinlánc hossza. Ezek a változások a vízkötés csökkenésére, enzimatikus degradációra és vele degenerációra, arthrosisrahajlamosítnak.
Heparin

76
A porcszövetek degeneratív folyamataiban, tehát aporc kondroitin-szulfát tartalma jelentősen csökken,ennek következtében a vízmegkötő képessége iskisebb lesz, így a csökkent víztartalmú szövetekrugalmassága is csökken. Ez a folyamat azutánterhelés hatására mikro-sérülésekhez vezet, amitovábbi degradációt okoz.
kondroitin-szulfát
Természetes kopolimer, amelynek fő építőelemeit két diszacharidszármazék – a [4)-(β-D-glükopiranoziluronsav)-(1→3)-[2-(acetilamino)-2-dezoxi-β-Dgalaktopiranozil-4-[hidrogén-szulfát)]-(1→] és a [4)-(β-D-glükopiranoziluronsav)- (1→3)-[2-(acetilamino)-2-dezoxi-β-D-galaktopiranozil-6-[hidrogén-szulfát)]-(1→] – nátriumsói alkotják. Az anyag teljes hidrolízise során D-galaktózamin, D-glükuronsav, ecetsav és kénsav szabadul fel. Az anyag kinyerésére szárazföldi és tengeri állatok porcszöveteit egyaránt felhasználják. A 4-szulfát- és a 6-szulfátcsoportok aránya a kinyerésre felhasznált állatfajtól függően eltérő. Tartalom: 95–105% (szárított anyagra). SAJÁTSÁGOK Küllem: fehér vagy csaknem fehér, nedvszívó por. Oldékonyság: vízben bőségesen oldódik; acetonban és etanolban (96%) gyakorlatilag nem oldódik

Glikoproteinek – oligoszacharid-fehérje konjugátumok
A glikoproteinek olyan fehérjék, melyekhez kovalens kötéssel szénhid-rátok kapcsolódnak. Prokarióták, eukarióták (valódi sejtmaggal rendelkeznek), sőt vírusok isrendelkeznek rájuk jellemző glikoproteinekkel. Eukarióta sejtekben a glikolizáció (vagyis aszénhidrátok hozzákapcsolódása a fehérjéhez) a Golgi-készülék nevű sejt szervecskébentörténik meg. Szénhidrátok kötődhetnek szerin, treonin, hidroxi-lizin és hidroxi-prolin aminosavakhoz O-glikozidos kötéssel, valamint aszparaginhoz N-glikozidos kötéssel. Aglikolizációt glikozil-transzferáz enzimek végzik.

- megtermékenyülés- immunvédelem- vírusos, bakteriális fertőzések- sejtnövekedés- sejt-sejt adhézió- vérrögök feloldódása- gyulladásos folyamatok- tumor antigének
Glikoproteinek szerepe
plazmamembrán


Galaktolipidekben egy vagy két galaktóz egység kapcsolódik glikozidos kötéssel az 1,2-diacilglicerin C3 hidroxilcsoportjához. Anövényi sejtmembránok szulfolipideket is tartalmaznak, melyekben A C6 helyzetben szulfonált glükóz egységek találhatóak.Ezek a molekulák is amfipatikusak a foszfolipidekhez hasonlóan, és a molekula hidrofil része negatív töltésű.
Glikoszfingolipidek a plazmamembrán külső oldalán találhatóak, és a ceramid egység C1 hidroxil csoportja van glikozilezve.Szerepük a felismerésben (vírusok, baktériumok) és a sejtek közötti kommunikációban van.

Glikoszfingolipidek határozzák meg a vércsoportot. Az emberi vércsoportokat (O, A, B) meghatározó a glikoszinolipidekbentaláható eltérő oligoszacharid egységekben különböznek egymástól (glu: glükóz; Gal: galaktóz; GalNAc: N-acetil-galaktózamin;Fuc: fukóz)
Szfingozin: egy 20 C-atomos aminoalkohol
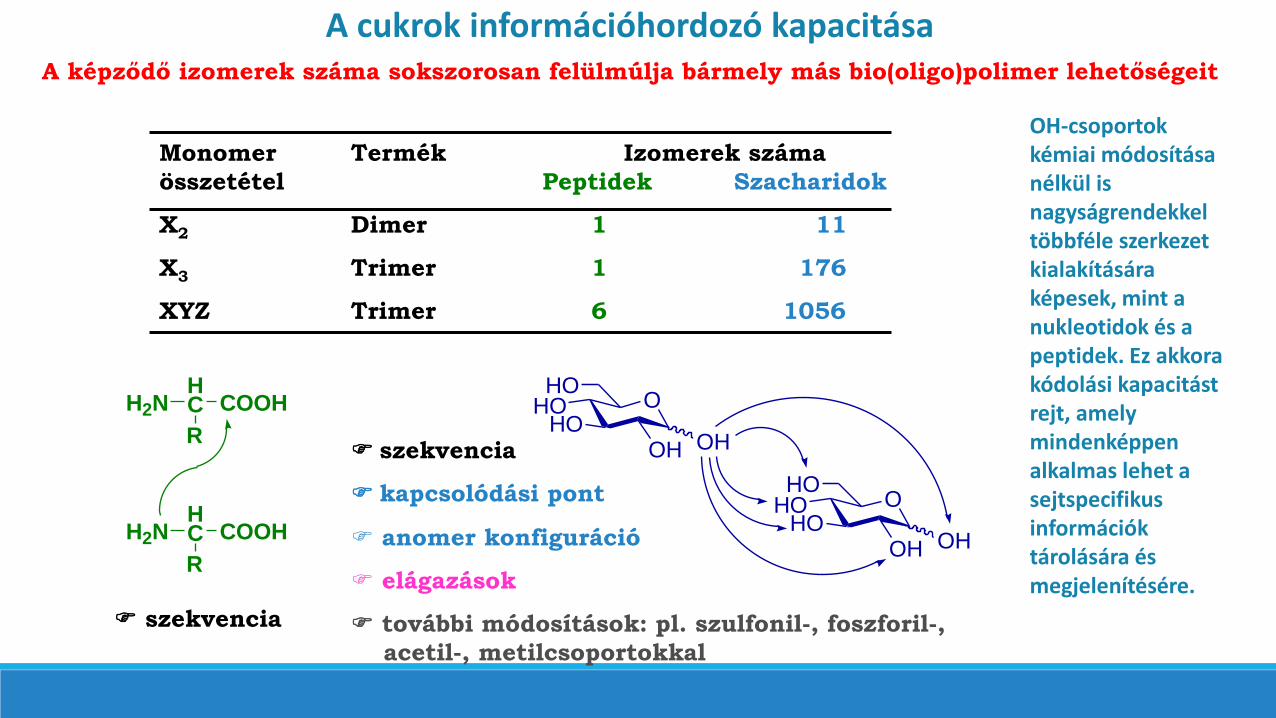
A cukrok információhordozó kapacitásaA képződő izomerek száma sokszorosan felülmúlja bármely más bio(oligo)polimer lehetőségeit
Monomer Termék Izomerek száma
összetétel Peptidek Szacharidok
X2 Dimer 1 11
X3 Trimer 1 176
XYZ Trimer 6 1056
O
OHOHHO
HOHO
O
OHOHHO
HOHO
H2NHC
R
COOH
H2NHC
R
COOH
szekvencia
szekvencia
kapcsolódási pont
anomer konfiguráció
elágazások
további módosítások: pl. szulfonil-, foszforil-,
acetil-, metilcsoportokkal
OH-csoportokkémiai módosítása nélkül is nagyságrendekkel többféle szerkezet kialakítására képesek, mint a nukleotidok és a peptidek. Ez akkora kódolási kapacitást rejt, amely mindenképpen alkalmas lehet a sejtspecifikusinformációk tárolására és megjelenítésére.

Oligonukleotidok, oligopeptidek és oligoszacharidok sokféleségeA szénhidrátkód
Forrás: P. H. Seeberger et al., ACS Chem. Biol., 2007, 2, 685.
O
OHOHHO
HOHO
O
OHOHHO
HOHO
O
OH
OH
HOHOHO
O
OHAcNHHO
HOHO
O
OHAcNHHO
HOHO
O
OHOHHO
HO
O
OHOHHO
HOHOOC
O
OHOHHO
HO
HOOC
OH3C
HOOH
OH
OH
OAcNH
OH
OH
COOHHO
HO
OH
D-GlcNAc
(31.8 %; - 8 %)
D-Gal
(24.8 %; - 23 %)
D-Man
(18.9 %; - 8.2 %)
D-Sia (8.3 %; - 26.1 %) L-Fuc (7.2 %; - 23.8 %)
D-GalNAc
(4.8 %; - 2.3; - 2.2 %)
D-Glc
(2.5 %; - 0.8 %)
D-GlcA (0.3 %)
D-Xyl (0.1 %)
L-IdoA (0.1 %)
Monoszacharid
Előfordulás (%)
Végcsoportként (%)
3299 emlős
oligoszacharid
statisztikai
elemzése alapján
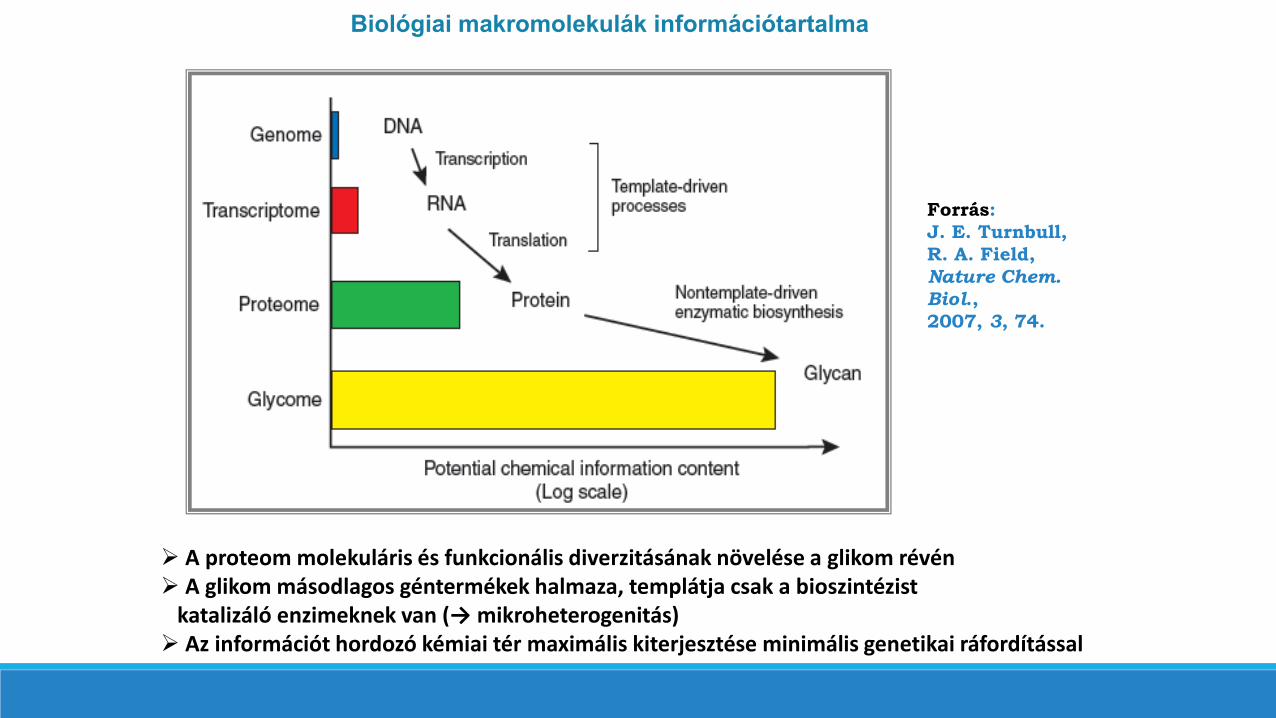
A proteom molekuláris és funkcionális diverzitásának növelése a glikom révén A glikom másodlagos géntermékek halmaza, templátja csak a bioszintézist
katalizáló enzimeknek van (→ mikroheterogenitás) Az információt hordozó kémiai tér maximális kiterjesztése minimális genetikai ráfordítással
Forrás:
J. E. Turnbull,
R. A. Field,
Nature Chem.
Biol.,
2007, 3, 74.
Biológiai makromolekulák információtartalma





![Magyarok a Lengyel Légiókban 1914–1918epa.oszk.hu/02900/02924/00036/pdf/EPA02924_valosag_2015_12_035-060.pdfközülük kik kapták azt meg. Nevezettek egy részét korábban [is]felterjesztettük,](https://img.pdfslide.net/doc/110x75/5e53fac323993e046079376a/magyarok-a-lengyel-lgikban-1914a-kzlk-kik-kaptk-azt-meg-nevezettek.jpg)























