Embed Size (px)
Citation preview

Gen. Pharmac., 1976, Vol. 7. pp. 185 to 188. Pergamon Press. Printed in Great Britain
TACHYPHYLAXIS AND ANGIOTENSIN RECEPTORS*
T. B. PAIVA, J. ABOULAFIA, V. L. A. NOUAILHETAS and A. C. M. PAIVA
Department of Biophysics and Physiology, Escola Paulista de Medicina, 04023 S~to Paulo, S. P., Brazil
(Received 18 December 1975)
Abstract--1. Angiotensin tachyphylaxis was studied in the isolated guinea pig ileum under isometric conditions.
2. In tachyphylactic organs, the initial phasic response to angiotensin was abolished, while the tonic component of the response became faster in its onset and in its reversal (relaxation).
3. The relaxation of non-tachyphylactic organs after angiotensin washout was significantly slower than with bradykinin, des-amino-angiotensin and hexanoyl-angiotensin. This difference did not occur in tachyphylactic organs.
4. There was no correlation between production of tachyphylaxis or rate of relaxation after washout and the partition coefficient of the four peptides in an organic:aqueous solvent system,
5. It is suggested that tachyphylaxis depends on a slowly reversible alteration of a calcium transloca- tion step in the stimulus-response coupling.
SMOOTH muscle contracting substances are generally believed to form a reversible complex with a specific receptor located at the cell membrane, originating a stimulus that triggers a chain of reactions leading to an increase in the intracellular calcium concentration and consequent contraction of the myofibrils (Hur- witz & Suria, 1971). The linking of the agonist-recep- tor interaction with the final contractile response may be thought to involve components that are sequen- tially switched between resting (responsive) and acti- vated (unresponsive) states. Some of these com- ponents may be common to several agonists, but for each type of agonist one or more specific links in the stimulus-response chain may be present.
In an effort to better understand the stimulus-res- ponse coupling in the myotropic action of angiotensin II (angiotensin), we have tried to explore a specific property of this action: the acute desensitization (tachyphylaxis) produced by the repeated adminis- tration of the hormone (for a recent review see Stew- art, 1974). The analysis of this phenomenon has become particularly interesting after the finding that the N-terminal amino groups of angiotensin, in its protonated form, is essential for the manifestation of tachyphylaxis (Paiva, Juliano, Nouailhetas & Paiva, 1974; Paiva & Paiva, 1975).
In the present paper we discuss some observations regarding tachyphylaxis in the isolated guinea pig ileum under isometric conditions.
* Supported by grants from Financiadora Especial de Projetos (FINEP), Conselho Nacional de Pesquisas (CNPq) and Funda?ao de Amparo /t Pesquisa do Estado de Sag Paulo (FAPESP).
MATERIALS AND METHODS
[-5-isoleucine]Angiotensin II (angiotensin), r l-succinic acid, 5-isoleucine]angiotensin II (des-amino-angiotensin), hexanoyl-[5-isoleucine]angiotensin II 0aexanoyl-angioten- sin) and bradykinin were synthesized by the solid phase method (Merrifield, 1963; Stewart & Young, 1969) as de- scribed elsewhere (Paiva, Nouailhetas, Miyamoto, Mendes & Paiva, 1973 ; Paiva, Goissis, Juliano, Miyamoto & Paiva, 1974; Tominaga, Stewart, Paiva & Paiva, 1975).
The isolated guinea pig ileum was prepared by decapi- tating a starved (24 hr) animal of 150-200 g body wt and immediately removing a 15cm segment of its terminal ileum and washing it with Tyrode solution at room tem- perature. A 4 cm portion was then suspended in Tyrode solution in a chamber of 5.0 ml capacity maintained at 37°C, constantly bubbled with a stream of air. The compo- sition of the Tyrode solution was the following: NaC1, 0.14M; KCI, 2.7 mM; CaCI2, 1.36 mM; MgCI2, 0.53 mM; NaHCO3, 12 mM; NaHzPO4, 0.36 mM; glucose, 5.6 mM; atropine sulfate, 1 mg/l. The pH of this solution, when equilibrated with air, was 8.0. The isometric contractions were recorded with a Sanborn force transducer through a Hewlett-Packard model 8805 B amplifier and a Heath- Schlumberger model EU205-11 recorder.
Tachyphylaxis was produced by repeated adminis- trations, at 5-min intervals, of an angiotensin concentration that produced maximum response of the organ (10 -~ M). The organ was washed after 90 sec of contact with the agonist, and left to rest for 3.5 rain before the next treat- ment. The same procedure was followed with the other agonists, also administered in concentrations that pro- duced maximum responses.
The partition coefficients were determined by shaking the peptide in the solvent system n-butanol-acetic acid- water (4:1 : 5) for 90 min, after which the phases were separ- ated by centrifugation for 90 min at 2500 rev/min and the peptide concentration in the aqueous phase was deter- mined spectrophotometrically at 275 nm.
185
186 T.B. PAIVA, J. ABOULAFIA, V. L. A. "NOUAILHETAS AND A. C. M. PAIVA
R E S U L T S
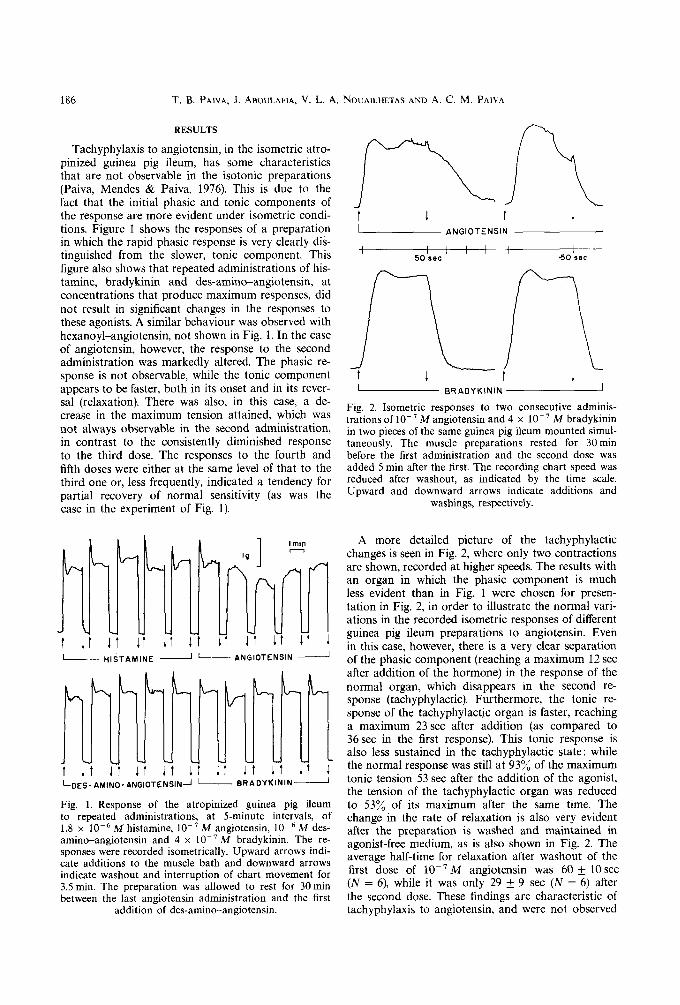
Tachyphylaxis to angiotensin, in the isometric atro- pinized guinea pig ileum, has some characteristics that are not observable in the isotonic preparations (Paiva, Mendes & Paiva, 1976). This is due to the fact that the initial phasic and tonic components of the response are more evident under isometric condi- tions. Figure 1 shows the responses of a preparation in which the rapid phasic response is very clearly dis- tinguished from the slower, tonic component. This figure also shows that repeated administrations of his- tamine, bradykinin and des-amino-angiotensin, at concentrations that produce maximum responses, did not result in significant changes in the responses to these agonists. A similar behaviour was observed with hexanoyl-angiotensin, not shown in Fig. 1. In the case of angiotensin, however, the response to the second administration was markedly altered. The phasic re- sponse is not observable, while the tonic component appears to be faster, both in its onset and in its rever- sal (relaxation). There was also, in this case, a de- crease in the maximum tension attained, which was not always observable in the second administration, in contrast to the consistently diminished response to the third dose. The responses to the fourth and fifth doses were either at the same level of that to the third one or, less frequently, indicated a tendency for partial recovery of normal sensitivity (as was the case in the experiment of Fig. 1).
t 1 [ A N G I O T E N S l N
I I J I J - SO s e c
t 1 L B R A O Y K I N I N
t I
J I- • 5 0 sec
I
Fig. 2. Isometric responses to two consecutive adminis- trations of 10- 7 M angiotensin and 4 x 10- 7 M bradykinin in two pieces of the same guinea pig ileum mounted simul- taneously. The muscle preparations rested for 30min before the first administration and the second dose was added 5 min after the first. The recording chart speed was reduced after washout, as indicated by the time scale. Upward and downward arrows indicate additions and
washings, respectively.
t 1 t 1 - - H I S T A M I N E - -
L D E S - AMINO" ANGIOTENSIN - I
') I rain Ig J r ~
t ~t ~t IT ANGIOTENSIN
- - B R A D Y K I N I N - -
Fig. 1. Response of the atropinized guinea pig ileum to repeated administrations, at 5-minute intervals, of 1.8 × 10 -6 M histamine, 10 -7 M angiotensin, 10 - 6 M des- amino-angiotensin and 4 x 10-TM bradykinin. The re- sponses were recorded isometrically. Upward arrows indi- cate additions to the muscle bath and downward arrows indicate washout and interruption of chart movement for 3.5 min. The preparation was allowed to rest for 30 min between the last angiotensin administration and the first
addition of des-amino-angiotensin.
A more detailed picture of the tachyphylactic changes is seen in Fig. 2, where only two contractions are shown, recorded at higher speeds. The results with an organ in which the phasic component is much less evident than in Fig. 1 were chosen for presen- tation in Fig. 2, in order to illustrate the normal vari- ations in the recorded isometric responses of different guinea pig ileum preparations to angiotensin. Even in this case, however, there is a very clear separation
J of the phasic component (reaching a maximum 12 sec after addition of the hormone) in the response of the normal organ, which disappears in the second re- sponse (tachyphylactic). Furthermore, the tonic re- sponse of the tachyphylactjc organ is faster, reaching a maximum 23 sec after addition (as compared to 36 sec in the first response). This tonic response is also less sustained in the tachyphylactic state: while the normal response was still at 93~/o of the maximum tonic tension 53 sec after the addition of the agonist, the tension of the tachyphylactic organ was reduced to 53~o of its maximum after the same time. The change in the rate of relaxation is also very evident after the preparation is washed and maintained in agonist-free medium, as is also shown in Fig. 2. The average half-time for relaxation after washout of the first dose of 10 -TM angiotensin was 60- t -10sec (N = 6), while it was only 29 _+ 9 sec (N = 6) after the second dose. These findings are characteristic of tachyphylaxis to angiotensin, and were not observed

Angiotensin receptors
Table 1. Rate of relaxation of guinea pig ileum after washout of submaximal doses of angiotensin and two analogues
PEPTIDE BIOLOGICAL pARTITION DOSE RELAXATION
* HALF TIME ~ ACTIVITY COEFFICIENT ~ (x 108 M) -
(seconds)
187
Angiotensin I00 0.34 ± 0.06 5 35 ± 13 (15)
Des-amino-angiotensin 11 ± 2 1.19 ± 0.04 50 6 ± 1 (8)
Hexanoyl-angiotensln 12 ± 2 3.9 ± 0.6 50 9.5 ± 0.5 (8)
* Activity in the guinea pig ileum in percentage of the activity of angiotensin, com-
pared on a molar basis (z standard deviations).
t Solvent system: ~-butanol:acetic acid:water (4:1:5). Averages of 5 measurements ±
s~andard deviations.
- ± one standard deviation. Number of measurements of relaxation rates, made in dif-
ferent organs, shown in parentheses.
with Other agonists such as des-amino-antiotensin, hexanoyl-angiotensin, histamine and bradykinin.
In order to see whether there was a correlation between the rate of relaxation after washout and the solubility properties of the agonist, we have compared these properties in agiotensin, des-amino-angiotensin and hexanoyl-angiotensin. Table 1 shows that remo- val of the amino group (in des-amino--angiotensin) or its blocking by acylation with hexanoic acid (hex- anoyl-angiotensin) give rise to angiotensin analogues which are equipotent towards the guinea pig ileum, in spite of a marked difference in their relative solubi- lities in the organic and aqueous phases. It can also be seen in Table 1 that the free amino group is more important that the partition coefficient in determining the rate of the organ's relaxation after agonist remo- val. Indeed, while there is an approx 3-fold increase in the partition coefficient in going from angiotensin to its des-amino analog, and another 3-fold increase from the latter to the hexanoyl derivative, a large dif- ference in relaxation behaviour is observed between angiotensin and the other two peptides, while only a small difference is found between the two analogues.
DISCUSSION
The available evidence (Paiva et al., 1976) indicates that tachyphylaxis is caused by the sluggishness of a specific component of the angiotensin stimulus- effect chain in returning from an unresponsive to a responsive state, after a normal response. The evi- dence also points to this component being involved in calcium translocation. The fact that des-amino-- angiotensin, which acts on the same receptor as angiotensin (Paiva, Goissis, Juliano, Miyamoto & Paiva, 1974), does not induce tachyphylaxis, suggests that this phenomenon arises from a direct interaction of angiotensin (in particular its amino group) with the postulated calcium translocation component. This
interaction might be with the same receptor site in- volved in the peptide's agonistic action, as first pro- posed by Khairallah, Page, Bumpus & Tiirker (1966). In this case, tachyphylaxis would result from a slow dissociation of angiotensin from its normal receptor sites. This implies that the occupied receptors would be active and, consequently, the response would depend on the agonist-receptor reaction rate rather than on receptor occupation, as proposed by Paton (1961). If this were the case, one should expect that, in the tachyphylactic organ, the free receptors avail- able for binding to new angiotensin molecules would be smaller in number, but unchanged in their proper- ties. This should lead to a decrease in the response, but not in a qualitative change. However, the iso- metric recording of the guinea pig ileum's response to repeated angiotensin treatments showed that a change in the time course of the response is the most striking' sign of tachyphylaxis (Fig. 1). This indicates a qualitative alteration of the component of the sti- mulus-effect chain which is involved in tachyphylaxis.
Chang & Triggle (1973) have presented evidence that the initial phasic contraction of the guinea pig ileum is caused by intracellular release of membrane- bound Ca 2 +, while translocation of free extracellular Ca 2+ would be responsible for the tonic component. If this is the case, the tachyphylactic organ has lost its ability to release membrane-bound Ca 2 + into the cell, in response to angiotensin. This is a specific pro- perty of the angiotensin stimulus-effect chain, since the phasic response to other agonists is unaltered. The agonist-induced translocation of extracellular Ca 2+ into the cell is also affected in tachyphylaxis, as indi- cated by the faster onset and reversal of the tonic response (Fig. 2). This is supported by the enhance- ment of tachyphylaxis in low-calcium media (Stewart, 1974; Paiva et al., 1976).
Our results thus indicate that tachyphylaxis arises from the interaction of angiotensin with a component

188 T.B. PAIVA, J. ABOULAFIA, V. L. A. NOUAILHETAS AND A. C. M. PAIVA
of the stimulus-effect chain that is involved in calcium translocation. This component may or may not be the same one that contains the "receptor site" for angiotensin peptides, but the tachyphylactic interac- tion must involve a different site, since angiotensin analogues lacking a free amino group interact with the "receptor site" but do not induce tachyphylaxis.
An important characteristic of tachyphylaxis is the slowness of the recovery from the tachyphylactic state, which may be related to the slow relaxation ob- served after washout of the first dose of angiotensin as compared to other agonists (including desamino- angiotensin and hexanoyl-angiotensin). Kalsner & Nickerson (1968) have found that relaxation of rabbit aortic strips after washout is controlled by the rate of diffusion of agonist from the hydrophobic "bio- phase" into the aqueous external phase. This does not seem to occur in the case of angiotensin on the guinea pig ileum, since no correlation between parti- tion coefficient and relaxation rate was found (Table 1). Furthermore, the relaxation was faster in the case of analogues with higher solubility in organic (relative to aqueous) solvents, contrary to what should be expected if diffusion from the biophase were rate- limiting.
CONCLUSIONS
The results presented in this paper, together with previous findings, lead us to the following conclu- sions:
1. Tachyphylaxis to angiotensin in the isolated guinea pig ileum results from the slow return of a component of the stimulus-effect chain from an un- responsive to a responsive state, after the agonist is washed out.
2. The diffusion of angiotensin from the hydro- phobic biophase into the aqueous external phase is not rate-limiting for the relaxation after washout.
3. To exert its tachyphylactic effect angiotensin in- teracts with a site that is different from the agonistic receptor site, and the peptide's amino group is essen- tial for this interaction.
4. In the tachyphylactic state the angiotensin-pro- duced transient release of membrane-bound Ca 2÷ into the cell is probably depressed, while the translo-
cation of calcium from the extra- to the intracellular space occurs earlier than in the normal state.
Acknowledgemen~This work was done with the techni- cal assistance of Nelson A. Mora.
REFERENCES
CHANG K. J. & TRIGGLE D. J. (1973) Quantitative aspects of drug-receptor interaction~I. Ca 2+ and cholinergic receptor activation in smooth muscle: a basic model for drug-receptor interactions. J. theor. Biol. 40. 125 154.
HURWITZ L. & SURIA A. (1971) The link between agonist action and response in smooth muscle. Ann. Rev. Phar- mac. l l , 303-326.
KALSNER S. & NICKERSON M. (1968) A method for the study of mechanisms of drug disposition in smooth muscle. Can. J. Physiol. Pharmaeol. 46, 719-730.
KHA1RALLAH P. A., PAGE I. H., BUMPUS F. M. & Tt)RKER R. K. (1966) Angiotensin tachyphylaxis and its reversal. Circul. Res. 19, 247-254.
MERRIFIELD R. B. (1963) Solid phase synthesis I. The syn- thesis of a tetrapeptide. J. Am. Chem. Soc. 85, 2149-2154.
PAIVA A. C. M., NOUAILHETAS V. L. A., MIYAMOTO M. E., MENDES G. B. & PA1VA T. B. (1973) New specific angiotensin inhibitors: [ValS]-[IleS] - and Chlorambucil- [des-Asp I, ValS]angiotensins I. J. reed. Chem. 16, 6-9.
PAIVA A. C. M. & PAIVA T. B. (1975) A model for angioten- sin receptors in smooth muscle cells. In Concepts of Membranes in Regulation and Excitation (Edited by SILVA M. & StJAREZ-KURZ G.), pp. 145 154. Raven Press, New York.
PAIVA T. B., GOlSSIS G., JULIANO L., MIYAMOTO M. E. & PAIVA A. C. M. (1974) Angiotensin-like and antagonistic activities of N-terminal modified I-Leu8]-angiotensin II peptides. J. reed. Chem. 17, 238-241.
PAIVA T. B., JULIANO L., NOUAILHETAS V. L. A. & PAIVA A. C. M. (1974) The effect of pH on tachyphylaxis to angiotensin peptides in the isolated guinea pig ileum and rat uterus. Eur. J. Pharmac. 25, 191-196.
PAIVA T. B., MENDES G. B. & PAIVA A. C. M. (1976) The specific desensitization (tachyphylaxis) of the guinea pig ileum to angiotensin II. Am. J. Physiol. (In press).
PATO• W. D. (1961) A theory of drug action based on the rate of drug-receptor combination. Proc. R. Soc. Lond. B. 154, 21-69.
STEWART J. M. (1974) Tachyphylaxis to angiotensin. In Angiotensin (Edited by PAGE I. H. & BUMPUS F. M.). pp. 170-184. Springer-Verlag, Berlin.
STEWART J. M. & YOUNG J. (1969) Solid Phase Peptide Synthesis. Freeman, San Francisco.
TOMINAGA M., STEWART J. M., PAIVA T. B. & PMVA A. C. M. (1975) Synthesis and properties of new bradykinin potentiating peptides. J. reed. Chem. 18, 130-133.





























