Embed Size (px)
Citation preview

REVIEWS
Taking dendritic cells into medicineRalph M. Steinman1 & Jacques Banchereau2
Dendritic cells (DCs) orchestrate a repertoire of immune responses that bring about resistance to infection and silencing ortolerance to self. In the settings of infection and cancer, microbes and tumours can exploit DCs to evade immunity, but DCsalso can generate resistance, a capacity that is readily enhanced with DC-targeted vaccines. During allergy, autoimmunityand transplant rejection, DCs instigate unwanted responses that cause disease, but, again, DCs can be harnessed to silencethese conditions with novel therapies. Here we present some medical implications of DC biology that account for illness andprovide opportunities for prevention and therapy.
Immunology is a major force in medicine. It is needed to under-stand how prevalent diseases (Fig. 1) come about and how todevelop preventions and treatments. This broad reach of theimmune system reflects its two functions: to recognize diverse
substances termed antigens, and to generate many qualitatively dis-tinct responses. Dendritic cells (DCs), named for their probing, tree-like or dendritic shapes (from the greek ‘dendron’, meaning tree)1
(Fig. 1), are pivotal for both recognition of a universe of antigens andcontrol of an array of responses.
Previously, we reviewed some biological features of DCs2. Here weillustrate medical implications of DCs, which control a spectrum ofinnate and adaptive responses. Innate immunity encompasses manyrapid reactions to infection and other challenges3. Adaptive immun-ity, in contrast, is learned or acquired more slowly, in days to weeks; ithas two hallmarks—exquisite specificity for antigens, and a durablememory to develop improved function on re-exposure to antigen.Adaptive responses are either immunogenic, providing resistance ininfection and cancer, or tolerogenic, leading to silencing as is desirablein transplantation, autoimmunity and allergy. To date, the successesof immunology in the clinic have largely been based on antibodiesmade by B cells, but T-cell-mediated immunity, which has enormous,yet untapped, therapeutic potential, will be stressed here.
DCs are specialized to capture and process antigens in vivo4,5,converting proteins to peptides that are presented on major histo-compatibility complex (MHC) molecules and recognized by T cells.DCs also migrate to T-cell areas of lymphoic organs, where the twocell types interact to bring about clonal selection6–8. The DC system is
thus designed to harness the recognition repertoire of T cells,consisting of billions of different lymphocytes, each with a distinctbut randomly arranged antigen receptor. This repertoire in turnrepresents a virtually infinite ‘drug library’ for specific therapies thatincrease or decrease T-cell function.
Following clonal selection, DCs control many T-cell responses.Antigen-selected T cells undergo extensive expansion, a thousand-fold or more, as a result of division at a rate as high as 2–3 cell cycles aday9. Clones of lymphocytes are also subject to silencing or toleranceby so-called ‘tolerogenic DCs’10, which either eliminate (delete)9,10 orblock (suppress) T cells11. If deletion is avoided, the clone undergoesdifferentiation to bring about an array of potential helper, killer andsuppressive activities. For example, under the control of DCs, helperT cells acquire the capacity to produce powerful cytokines likeinterferon-c to activate macrophages to resist infection by facultativeand obligate intracellular microbes (T helper (TH)1 cells)12–14; orinterleukin (IL)-4, -5 and -13 to mobilize white cells that resist hel-minths (TH2 cells)15; or IL-17 to mobilize phagocytes at body surfacesto resist extracellular bacilli (TH17 cells)16. Alternatively, DCs canguide T cells to become suppressive by making IL-10 (T regulatory(Treg)1 cells)17 or by differentiating into FOXP31 cells11. Finally, DCsinduce the T-cell clone to acquire memory, allowing it to persist forprolonged periods and to respond rapidly to a repeated exposure toantigen18,19.
There are four specialized features (Fig. 2) of DCs that contributeto their capacity to control T-cell recognition and responsivenessand, in turn, either prevent or generate disease. Recent advances inDC biology are outlined in Boxes 1–4. Briefly, (1) DCs are positionedto capture disease-causing antigens and to present these to lympho-cytes in lymphoid organs20–22, the sites for the generation of immun-ity and tolerance; (2) DCs have an endocytic system that is dedicated
1The Rockefeller University, New York 10065, USA. 2Baylor Institute for Immunology Research, Dallas, Texas 75204, USA.
• Infection
• Cancer
• Transplantation
• Autoimmunity and chronic inflammation
• Allergy
• Vaccines
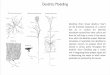
Figure 1 | One of the first views of DCs in mouse spleen1 (reproduced withpermission from The Journal of Experimental Medicine; copyright 1973Rockefeller University Press) and the conditions in which DCs and theimmune system are most studied.
Antigen uptake receptorsand processing pathwaysfor presentation of peptide–MHC complexes
Maturation or differentiationin response to microbial andother stimuli
Dendritic cells
T cellsand other lymphocytes
Subsets with distinctpattern recognitionreceptors and functions
Location at body surfaces and in the T-cell areas of lymphoid organs
Figure 2 | Innate features of DCs, each an intricate subject.
Vol 449j27 September 2007jdoi:10.1038/nature06175
419Nature ©2007 Publishing Group

to antigen capture and processing, creating ligands for different classesof lymphocytes4,5; (3) DCs differentiate or mature in response to aspectrum of stimuli23 (Table 1), allowing them to bring about innateand adaptive responses that are potent and qualitatively matched tothe disease-causing agent; and (4) DCs are comprised of subsets thatdiffer from one another in terms of location, antigen presentation, andmaturation24,25. Location (Box 1) and antigen presentation (Box 2)allow DCs to efficiently select specific clones from the diverse recog-nition repertoire. Maturation (Box 3) and subsets (Box 4) allow DCsto control the diverse response repertoire of T cells and other classes oflymphocytes, such as B cells and natural killer cells.
Dendritic cells in infectious diseases
DCs induce resistance to infection. When microbial antigens areinjected in association with DCs into mice, the animals acquireadaptive immunity to Borrelia burgdorferi, chlamydiae, Leishmaniamajor, fungi, Toxoplasma gondii, malaria and HIV. Conversely, DCdepletion reduces defences to viruses like CMV26, HSV-2 andLCMV27. In humans, a lack of circulating DCs during bacterial sepsisand dengue virus infection is associated with a poor prognosis.
A key concept is that DCs mature in distinct ways in responseto different microbial components, thereby launching alternativeversions of host immunity. The microbial ligands act on patternrecognition receptors3, including externally disposed Toll-likereceptors (TLRs) and lectins28, and the cytoplasmic NOD/NALP
family, RIG-I/DDX58 and MDA5/IFIH1 molecules. These pattern-recognition receptors can function synergistically14,29,30.
In contrast, several microbes have the capacity to actively block DCmaturation, for example, Coxiella burnettii, Salmonella typhi, anthraxlethal factor protein, Plasmodia, a Mycobacterium ulcerans mycolac-tone31 and viruses like vaccinia, herpes simplex, HIV, CMV, varicellazoster, HCV, Ebola/Marburg/Lassa fever and measles. An interestingexception is the effective attenuated yellow fever virus vaccine thatmay work by infecting and maturing DCs, allowing for antigen pre-sentation to T cells32.
Furthermore, pathogens can alter other levels of DC physiology toevade an immune response. For example, the agent of plague,Yersinia pestis, and of typhoid fever, Salmonella typhi, selectivelyinject toxins into phagocytes, including DCs, and destroy the cellsrequired for innate and adaptive protection. Influenza, measles andHSV-2 can induce apoptotic cell death in DCs33. With some viruses,cell death occurs through the intermediate formation of giant cells orsyncytia. CMV, herpes and Mycobacterium tuberculosis inhibit themigration step of DC function by blocking expression of CCR7(ref. 34), a chemokine receptor that guides DCs into lymphatic ves-sels and onwards to lymphoid tissues6,8 (Box 1).
Microbes also can alter the function of DCs so that they switchT-cell responses from protective TH1 to non-protective TH2, as ininfections with Aspergillus fumigatus, malaria and hepatitis C, or toIL-10 production in the case of Bordetella pertussis.
Box 1 j Location of DCs
DCs are a uniquely positioned, prime target for disease-relevantstimuli. DCs are abundant at body surfaces like the skin, pharynx,upper oesophagus, vagina, ectocervix and anus, and at so calledinternal or mucosal surfaces, such as the respiratory and gastro-intestinal systems21. DCs actually extend their processes through thetight junctions of epithelia, which probably involves DC expression ofthe tight junction proteins claudins and occludins, without alteringepithelial barrier function86. This increases DC capture of antigensfrom the environment even when there is no overt infection orinflammation, probably allowing for the silencing of the immunesystem to harmless environmental antigens.
DCs at body surfaces can function locally, for example, to convertvitamins A and D to active retinoic acid and 1,25-dihydroxy-vitamin D3.One consequence of the overlooked metabolic capacities of DCs is toincrease the homing of immune cells to that mucosal surface and, inthe case of retinoic acid, to help DCs differentiate suppressor T cells,which block autoimmune and inflammatory conditions87.
After leaving peripheral tissues, DCs migrate with environmental,self and microbial antigens to lymphoid organs—a process that isguided by chemokines6,8 and can be enhanced by vaccination.
DCs have now been studied in intact lymphoid tissues without theneed for cell isolation. These are the sites where immune resistanceand tolerance are initiated. The DCs create a labyrinthine systemwithin T-cell areas, while probing the environment through thecontinuous formation and retraction of processes22, and displayingantigens and other stimuli needed to initiate responses by appropriateclones of specific T cells88,89.
New research reveals interactions of DCs in lymphoid tissues withother major classes of lymphocytes, B cells and natural killer cells55.
Table 1 | Some stimuli for dendritic-cell maturation
Stimulus Details
Microbial products Via Toll-like, NOD, RIG-I, Mda5 receptorsLymphocytes (natural killer, NKT,cd T, ab T) and neutrophils
Via CD40, lymphotoxin ab receptors
Cytokines TNF, interferons, TSLP and IL-10
Endogenous ligands Uric acid, histamine, heat shock proteins, highmobility group box protein 1, defensins, ATP
Immune complexes Via activating and inhibitory FcR
These stimuli induce distinct pathways of DC differentiation, whereupon the conditioned DCsactivate distinct responses, for example, TH1, TH2, TH17, Treg cells. Two or more of these stimulican act synergistically14,29,30.
Box 2 jAntigen presentation by DCs
DCs contain a specialized endocytic system4,5, having many uptakereceptors that deliver antigen to processing compartments5,24. DCsthen present peptides from processed proteins to CD41 and CD81
T cells, self and microbial glycolipids to NKT cells, and native antigensto B cells.
Many uptake receptors on DCs are lectins with carbohydrate-recognition capacity28. Some, like LY75 (DEC-205/CD205) andmannose receptor MRC1/CD206, are type I transmembrane proteinswith multiple contiguous lectin domains. A majority to date, such asDC-SIGN/CD209, langerin/CD207, ASGPR, OLR1/LOX-1 (ref. 40)and CLEC4A/DCIR (ref. 24), are type II proteins with a single externallectin domain. Among these, CD209 has attracted wide interestbecause it binds a number of microbes including HIV35, dengue,cytomegalovirus, mycobacteria and candida, and because it can hinderDC maturation and contribute to immune evasion. Another lectin,langerin/CD207, is reported to degrade HIV in DCs, thereby reducingtransmission to T cells.
Early literature on DCs took antigen uptake for granted, withoutrealizing that ligation of uptake receptors increases the efficiency withwhich antigens are delivered to the immune system in vivo by ,100-fold9,19,38. Targeting vaccines to these DC receptors should significantlyimprove the efficacy of T-cell mobilizing vaccines.
Following uptake of antigen, DCs are able to ‘cross-present’ antigenson MHC I to elicit CD81 killer T cells33,90,91. During cross-presentation,non-replicating protein antigens are internalized and somehow gainaccess to the cytoplasm before being processed by the proteasome forpeptide presentation on MHC I. Critical steps seem to occur from less-acidic compartments. Cross-presentation allows DCs to induce CD81
T cell responses to immune complexes, non-replicating forms ofmicrobes and vaccines, and dying cells.
For dying cells, DC receptors for uptake remain to be defined in situ.Because cell death accompanies infection, cancer, transplantation, thenormal turnover of self tissues and some viral vaccines, the uptake ofdying cells is a starting point for DCs to capture antigens in manyclinical settings.
Fc receptors, which recognize antigen–antibody complexes,mediate antigen uptake and both activating and inhibitory signals inDCs47. For example, if inhibitory receptors are blocked, the binding oftumour cells coated with antibody leads to improved presentation oftumour antigens and production of the immune-stimulating cytokineIL-12 (ref. 47).
REVIEWS NATUREjVol 449j27 September 2007
420Nature ©2007 Publishing Group

At this time, there are no therapies that try to interrupt the micro-bial immune-evasion pathways that are summarized above.
Several microbes additionally can exploit DCs for purposes ofreplication and spread in the infected host. The lectin DC-SIGN/CD209 is used by dengue virus and Ebola virus to infect DCs. Inthe case of HIV-1, CMV and Ebola virus, the lectin additionallysequesters virus within DCs, which later transmit infectious virusto other targets, such as T cells35. DCs are also implicated in thespread of varicella zoster, measles virus, poliovirus, Aspergillus fumi-gatus, LCMV, Toxoplasma gondii, prions and Bacillus anthracisspores.
All these consequences of the microbial–DC interaction have beenanalysed on myeloid DCs, mainly in tissue culture but not in patients.We expect that microbes also evade and exploit plasmacytoid DCs,which also help to resist pathogens36 and are diminished in the bloodduring infections with HIV, HTLV-1 and RSV.
To counteract these mechanisms for pathogenesis of infectiousdisease, DCs are now being considered in the design of vaccines toprevent and treat infection by enhancing immunogenesis. A newconcept is to deliver vaccine antigens to specific receptors on DCs(Box 2), along with stimuli to control DC maturation. For example,microbial proteins are genetically engineered into anti-receptormonoclonal antibodies, which then quickly and selectively target to
large numbers of DCs within intact lymphoid tissues9,19,37. TheCD205 receptor, which is abundant on DCs in human lymphoidtissues, delivers antigen for processing onto both MHC class I andII (MHC I and II), increasing presentation efficiency .100-fold rela-tive to non-targeted antigen 19,38. TH1 responses, considered valuablefor protection against many intracellular pathogens and tumours,also are induced when antigens from HIV, malaria, Leishmania andtumours are targeted to maturing CD2051 DCs19,37,38. Significantly,these responses are broad; that is, they are capable of recognizingmany peptides from a given microbial protein and in several MHChaplotypes19,39, and the responses take place at mucosal surfaces—both key criteria in vaccine design. Other potential DC targets arebeing addressed, including LOX-1/OLR1 (ref. 40), MMR/CD206/MRC-1, DCIR/CLEC4A (ref. 24), DC-SIGN/CD209, toxin receptorsand CD40.
Beyond the value of antigen-targeting, there is a need to matureDCs during vaccination in a way that is appropriate to the pathogenat hand. Here, one needs to define correlates of immunogenicityin vivo; that is, the specific changes in DCs required to generateprotective responses from the response repertoire.
Dendritic cells in cancer
DCs are found in tumours in mice and patients. Yet tumours sup-press immunity, especially locally and by many pathways41. Tumoursexpress cytokines, like IL-6, vascular endothelial growth factor andIL-10, which suppress DCs through STAT3 signalling42. Tumoursmay condition local DCs to form suppressive T cells, such asFOXP31 (ref. 43) and IL-13-producing CD41 T cells (ref. 44) andnatural killer T cells (NKT cells). DCs even support the clonogenic
Box 3 jMaturation of DCs
DCs differentiate or mature in distinct ways in response to a spectrumof environmental and endogenous stimuli (Table 1). The maturationpathway then helps to designate which lymphocyte functions will beinduced, and which products will be made by both DCs andlymphocytes23,92.
In the steady state in the overt absence of maturation stimuli, DCscan induce tolerance when they capture self and environmentalantigens9. Maintenance of tolerance can require PD-L1/CD274 (ref.10) as well as FAS (ref. 93) on DCs. On infection or other causes ofmaturation, the DCs redeploy, but it is still not understood why DCdifferentiation is so rapid and extensive. One factor may be the highlevels of required NF-kB family proteins, with each family member(s)able to control different DC responses.
Maturing DCs can induce different types of CD41 T cells (see text),such as TH1, TH2 or TH17, to increase resistance. Other stimuli can yield‘tolerogenic’ DCs, which induce Tr1 and FOXP31 Treg cells11,17,75 tosilence immunity. Maturing DCs also express more IL-15 and activateinflammation and natural killer cells in vivo55,94
When DCs mature in response to microbial products, the expressionof hundreds of genes is altered95, leading to synthesis of cytokines, forexample, IL-12 and type I interferons, which enhance innate andadaptive resistance26,96. Although cytokines in turn can induce somecomponents of DC maturation, the DCs that directly interact withmicrobial ligands are the immunologically more-active ones97. Thetypes of cytokines are influenced by the DC subset and the mode of DCactivation. Several chemokines are also secreted in groups at a time98,attracting different cells in succession to the site of DC maturation:phagocytes, memory lymphocytes and naive T cells99. Numerousmechanisms also dampen the DC response to microbial products.
Maturation regulates antigen-processing by lowering the pH ofendocytic vacuoles, activating proteolysis, and transportingpeptide–MHC complexes to the cell surface5.
Importantly, maturing DCs remodel their surface, typicallyexpressing many membrane-associated co-stimulatory molecules65,97;these include members of the B7, TNF and Notch families.
A critical unknown is the definition of changes in DCs that link innateto adaptive immunity in vivo. The standard view that a combination ofMHC-peptide (‘signal one’) and high B7-2/CD86 is sufficient to driveT-cell immunity is oversimplified. To influence T-cell differentiation,DCs additionally need to produce cytokines like IL-12 and type Iinterferons, or membrane-associated TNF family receptors—likeCD40 (ref. 65) and lymphotoxin receptors—and TNF familymembers—like CD70 and OX40L to induce TH1 (ref. 38) and TH2 (ref.100) cells, respectively.
Box 4 jSubsets of DCs
There are several types of DCs, each with distinct markers andfunctions25. Plasmacytoid DCs, so named because of cytologicsimilarities to antibody-producing plasma cells, can be involved intolerance in their immature state. For maturation, these DCsselectively express activating FcR as well as TLR7 and TLR9. Whenimmune complexes containing DNA and RNA are bound and ingested,they signal potentially pathologic levels of type I interferons68,72
Other DCs, termed ‘myeloid’, also produce type I interferons.Myeloid DCs in different tissues can be further subdivided on the basisof expression of certain markers and functions. In the case of skin, DCsin the epidermis (Langerhans cells) express langerin/CD207 and DEC-205/CD205/LY75, and induce strong killer T-cell responses; someDCs in the dermis express DC-SIGN/CD209 and mannose receptor/CD206 and can activate antibody-forming B cells. Distinct skin DCpopulations also migrate to different areas of the draining lymphnodes101. Thus the outcome of skin vaccination may depend on thelectin and DC subset that pick up the vaccine.
In mouse spleen, a DC subset expresses DEC-205 and is particularlyefficient for the cross-presentation of antigens on MHC I 24, includingtumour cells, and also for the induction of IFN-c-producing, TH1 helperT cells12,13,38. A second subset expresses DCIR2/CLEC4A4 and otherlectins and is more efficient at processing antigens for presentation onMHC II24. In lymph nodes draining mucosal tissues, the presence of anintegrin, CD103/ITGAE, distinguishes a DC subset that cross-presentson MHC I102 but also synthesizes retinoic acid for the differentiation ofFOXP31 Treg cells87.
DC subsets communicate with each other, for example,plasmacytoid DCs produce interferons and membrane-bound co-stimulators that recruit other DCs to participate in immunity103.
The production of most DC subsets is controlled in the steady stateby the cytokine FLT3 ligand (FLT3LG)104,105, whereas duringinflammation and infection, another cytokine, GM-CSF, mobilizesincreased numbers of monocyte-derived DCs106. In the steady state,DCs in lymphoid tissues emanate from marrow progenitors in theblood107 but not monocytes25,108, although monocytes give rise to DCsin some non-lymphoid tissues in the steady state108 and in many sitesduring inflammation 25.
NATUREjVol 449j27 September 2007 REVIEWS
421Nature ©2007 Publishing Group

growth of tumours in multiple myeloma. Therefore, like someinfections, cancers have ways to evade and exploit DCs45.
Nevertheless, immunology is providing treatments for cancer,mainly in the form of monoclonal antibodies. Antibodies, by virtueof their antigen-binding variable Fab regions, can block critical func-tions on cancer cells; and by virtue of their constant or Fc regions,antibodies can mobilize an attack by other Fc receptor (FcR)-bearingcells such as innate phagocytes and natural killer cells. Antibodiesmediate the uptake and processing of tumour cells by DCs and alsocan trigger DC maturation (Table 1)46,47. This portends the design ofantibodies that harness select FcRs to induce better innate and adapt-ive anti-tumour immunity.
More emphasis is needed on cell-mediated immunity that also hasa clear capacity to resist cancer48. In phase I clinical research, adoptivetransfer of killer T cells leads to regressions of melanoma and othercancers. There is also a major survival benefit from allogeneic bonemarrow transplantation, in which lymphocytes from the marrowdonor resist leukaemia and other haematologic malignancies (the‘graft versus leukaemia’ reaction). In multiple myeloma, T cells thatrecognize glycolipids and peptides in the tumour are found in apremalignant stage of disease but are not found when the tumourgrows out of control. In paraneoplastic diseases, T cells respond toantigens shared by the tumour and the nervous system, and probablyresist the tumour but at the same time cause severe neurologic seque-lae. Examination of colorectal cancers indicates that the presence of aTH1-type immune response in the tumour correlates with a betterprognosis. On the other hand, immune suppression predisposes tohigher frequencies of several cancers. All of the above examples implya role for immune surveillance by T cells against human cancer, as isalso seen in mice49.
DCs can be marshalled for the prevention and treatment of cancerfor the following reasons: (1) tumours are replete with potentialantigens, and they can become immunogenic when presented byDCs50,51—this means that the immune attack on cancer can be broadenough to encompass multiple targets, including mutant proteinsexpressed by the cancer, and not just one target, where the latterfavours immune escape; (2) likewise, DCs can activate and expandthe different arms of cell-mediated resistance such as natural killer,NKT, cd T and ab T cells, each of which recognizes different altera-tions in cancer cells; and (3) DCs in systemic lymphoid organs andDCs generated ex vivo from progenitors in blood probably retaintheir immunizing capacities in cancer patients52,53. Therefore onecan test whether the specific features of DCs (Fig. 2) can be harnessedeither to generate therapeutic immunity or to prevent cancer duringpremalignant or minimal residual disease stages.
Two DC-based immune therapy approaches are currently avail-able. In one, DCs are generated ex vivo, loaded with tumour antigens,and re-injected to take advantage of the ability of DCs to migrate tothe T-cell areas of lymphoid organs to induce strong T-cell and,perhaps, natural killer immunity54,55 (Fig. 3). Less explored is theevidence that DCs themselves can acquire killer activity for humantumours and express granzyme and perforin killer molecules56. Inanother strategy, tumour cells or tumour antigens are targeteddirectly to DCs in the T-cell regions (Fig. 3), for example, withinmonoclonal anti-DC antibodies as discussed above.
Already a large number of phase I studies in humans have usedthe first approach, in which DCs are generated from precursorsex vivo. These early trials have only occasionally yielded significanttumour regressions57,58, and there are no studies yet showingimproved survival.
The current deficiencies in DC therapy need to be addressed inpatients to deal with significant scientific and other obstacles. First,DC vaccines are being tried only in late-stage cancer patients who areimmunosuppressed as a result of extensive radiation, chemotherapyand/or large tumour burdens. Why limit research to these patients,especially when DC therapy is nontoxic, including in the few patientswho have experienced tumour regressions57,58? Second, most injected
DCs remain at the injection site; only a few migrate to the draininglymphoid tissue, ,1% in mice and humans59. Research in patientswill be required to overcome this limitation. One example is to con-dition the injected site with cytokines like tumour necrosis factor(TNF), so that the injected DCs receive needed cues for migrationinto lymphatics. Third, more emphasis is needed on DC quality, bytesting DCs for their capacity to induce helper and killer T cells with ahigh avidity for tumour antigens but few Treg cells. Fourth, manyinitial vaccine studies used DCs charged with one or a few tumourantigens, whereas the potential of DCs rests with their still untappedcapacity to elicit a strong and broad immune attack to lessen thechances of tumour escape. Fifth, vaccine studies need to be accom-panied by in-depth immune monitoring to define assays for protec-tive lymphocytes, the presence of which correlates with tumourregression and/or improved survival. For example, in HIV vaccines,it is thought that protective T cells will produce several and higherlevels of cytokines like IFN-c, IL-2 and TNF-a; express low levels ofthe PD-1/CD279/PDCD1 regulatory protein; and have high func-tional avidity for antigen. Sixth, DC-based immune therapies requirea coordinated research effort to generate DCs for clinical use and tohelp investigators systematically evaluate the many variables pertin-ent to efficacy.
Even with these limitations, early trials with ex vivo DCs have, inpatients with advanced cancer, expanded T cells that recognize mul-tiple tumour antigens60 and that make the protective cytokine, IFN-c(ref. 52). These observations provide a starting point to use DCs in amore concerted way to study their immune capacities in cancerpatients, and to combine immunization with other therapeutic mod-alities. Chemotherapy61, radiotherapy and removal of tumour regu-latory mechanisms, for example, CTLA4, B7-H4/VTCN1, IL-10,TGF-b (ref. 43), IL-13 (ref. 44), could be more specific and less toxicif combined with DC-based therapies.
The ex vivo technique is being developed so that it is easier toperform, including in collaborative studies. This approach allowsthe basic features of DCs (Fig. 2) to be controlled, for example: toload them appropriately with multiple antigens, including tumourcells57,58, tumour RNA62, or, potentially, cancer stem cells; to selectfrom many options for DC maturation (Box 3); and to specify the DCsubset for clinical use60 (Box 4).
A second DC-based strategy in cancer therapy would be to mobi-lize DCs directly within the patient, either within the tumour orwithin lymphoid organs. In mice, irradiated tumour cells injectedintravenously are taken up by DCs. Alternatively, one can inject
Blood
Dendritic cells
Afferentlymphatics
Peripheral tissues
Blood
Bone marrow
Figure 3 | DC localization in T-cell areas of immune organs such as lymphnodes, as shown here. Lymphocytes, including T cells, move from the bloodinto the node through special vessels (yellow). T cells (blue) remain in T-cellareas (blue), whereas B cells (pink) move to the B-cell areas (pink). DCs(orange stellate profiles) are primarily in the T-cell areas, whereasmacrophages (green and orange profiles) are enriched in the periphery of thelymph node. To exploit this positioning in immunotherapy, DCs can begenerated ex vivo and injected, whereupon homing via lymphatics bringssmall numbers of DCs to the T-cell areas (top). Alternatively antigens can bedelivered directly to large numbers of DCs in the T-cell areas, for example, byincorporating the antigen within anti-DC monoclonal antibodies.
REVIEWS NATUREjVol 449j27 September 2007
422Nature ©2007 Publishing Group

irradiated tumour cells that are transduced to express GM-CSF,which recruits the patient’s DCs to capture and present tumourantigens63,64. A new approach to the maturation of antigen-capturingDCs in vivo uses innate NKT cells that recognize glycolipids65. Theactivated NKT cells mimic the effects of a combination of TLR andCD40 ligation on DCs and elicit long-lasting, protective CD41 andCD81 T-cell resistance. Another example is to selectively target can-cer antigens within monoclonal antibodies to receptors on DCs, thusensuring the delivery of antigen to large numbers of DCs in the T-cellareas of lymphoid organs, as opposed to the small numbers thatsuccessfully home from an injection site to these areas (Fig. 3).
Unfortunately, DC-mediated immunization against cancer is stillan underdeveloped field. Yet this approach can exploit the patient’sresponses to cancer to produce more specific, less toxic, broader andlonger-lasting therapies. There is an important unmet need toexpand and coordinate research on DC-based therapies against pro-liferating and initiating cancer cells.
Dendritic cells in autoimmunityInappropriate responses to self constituents, in select genetic back-grounds, can lead to chronic inflammatory conditions, termed auto-immune diseases. DCs bearing self antigens are able to induceautoreactive T cells in mouse models of multiple sclerosis, cardio-myopathy and systemic lupus erythematosus.
It is increasingly appreciated that a pivotal step leading to humanautoimmunity is an overproduction of a particular cytokine(s)66, andsubsets of DCs can be a major source. For example:
$ TNF-a is a key cytokine in rheumatoid arthritis and other dis-eases, like psoriasis, and TNF-a blockade is a powerful therapy formany patients. In psoriasis, a major source of TNF-a is a DC subsetinfiltrating the affected skin67.
$ Lupus erythematosus, a systemic disease in which antibodiesare formed against self constituents especially nucleoproteins, isaccompanied by what is termed an ‘interferon signature’ in whiteblood cells68. Type I interferon is made in large amounts by plasma-cytoid DCs69,70, which infiltrate the skin lesions of systemic lupuserythematosus. Viral nucleic acids, as well as self nucleoproteinsinternalized in the form of immune complexes, trigger TLR7 andTLR9, leading to type I interferon production. Moreover, theDNA-binding protein and cytokine, HMGB1, can deliver self DNAto TLR9 and ligate the RAGE/AGER receptor on DCs71. Interferon, inturn, drives the mobilization of activated granulocytes68 and differ-entiates monocytes into DCs, which may present dying cells in animmunogenic rather than tolerogenic manner72. Ligation of activ-ating FcR by the immune complexes that are formed during auto-immunity also drives DC maturation.
$ Type I interferon and plasmacytoid DCs are also proposed tobe pathogenic in other diseases, such as psoriasis73, dermatomyositisand Sjogren’s syndrome.
$ IL-23 is a cytokine that drives disease, including psoriasis andinflammatory bowel disease74, and DCs are major IL-23 producers.
Despite their role in inducing autoimmunity, DCs are also relevantto the therapy of these diseases: (1) various treatments, especiallyglucocorticosteroids, can reduce DC numbers and functions; (2)because DCs seem to be a source of pathogenic cytokines, the stimulifor cytokine production need to be identified and obviated; and (3)one potential antigen-specific strategy relates to the capacity of DCsto expand and induce T cells that suppress immunity, usually termedTreg cells. Treg cells can suppress other DCs that present disease-producing antigens. In the case of a spontaneous model of autoim-munity—diabetes in NOD mice—Treg cells that recognize antigensin insulin-producing b cells can be generated by DCs and provide atherapeutic benefit even after the onset of disease11,75.
Dendritic cells in allergyDCs normally ensure the immune system is tolerant on exposure toharmless environmental antigens including the pollens, dust mites,
and foods that cause respiratory and intestinal allergy. The DCs atmucosal surfaces capture proteins and silence the correspondingT cells, possibly by expressing ICOSL, which in turn differentiatesantigen-specific, IL-10-producing Treg cells. In allergy, instead of thisimmune tolerance, CD41 helper T cells develop along the atopic TH2pathway and yield interleukins (IL-4, -5 and -13) responsible formany aspects of disease, such as the formation of IgE-type antibodies.DCs both initiate the formation of pro-allergic TH2 T cells and boostsymptom-producing responses in already allergic mice76.
How might the normal tolerizing roles of DCs be altered duringallergy? Cytokines may again be pivotal, for example, recent studiesreveal a role for thymic stromal lymphopoietin (TSLP) in atopicdermatitis77 and asthma. TSLP matures human DCs from blood77
and skin to elicit unusual TH2 cells that secrete not only IL-4, -5and 13 but also high levels of TNF. TSLP-treated mature DCs expressOX40L/CD252/TNFSF4, a TNF family member. OX40L can instructT cells to develop a TH2-type of memory along with the production ofallergic mediators like prostaglandin D2.
Several new directions for allergy research involve DCs: (1) DCmaturation by TSLP needs to be understood and blocked; likewise forTSLP production by epithelial cells, which is controlled by retinoidreceptors as well as IL-1 and other inflammatory cytokines; (2) thesubstances that cause allergy may modify DC function directly byblocking IL-12 production by DCs and in turn favouring formationof allergic TH2 cells78; (3) in humans, new synthetic oligonucleotidescan suppress the presentation of allergens by DCs to TH2 cells; thesecompounds have shown promise in treating ragweed-induced aller-gic rhinitis; (4) DCs might be targeted to allow for formation ofallergen-specific Treg cells, which can treat allergy as observed inmouse models; and (5) new drugs can interfere with the pro-allergicfunctions of DCs and treat experimental asthma. The sphingosine-1-phosphate receptor agonist FTY720 blocks the migration of DCsfrom lung to lymph nodes where the DCs could present antigensto disease-causing TH2 cells, whereas agonists for the D prostanoid1 receptor condition DCs to induce disease-reducing Treg cells79.
Dendritic cells in transplantation
DCs play a key role in the outcome of organ and haematopoietictransplantation. DCs in grafted organs mature and migrate into therecipient, where they stimulate alloreactive T cells that bring aboutgraft rejection. Recipient DCs also can capture portions of thegraft80,81 and elicit organ rejection. Interestingly, suppressive drugsin current clinical use may act on the rejection-inducing DCs as wellas the rejecting T cells82. In haematopoietic transplantation, recipientDCs are key initiators of T-cell-induced graft versus host reac-tions83,84. Mechanisms have yet to be pinpointed to explain the mat-uration and migration of DCs that accompany transplantation.
It seems reasonable to propose that strategies to block these DCfunctions during transplantation will promote acceptance. However,recent discoveries show that DCs in grafted tissues can regeneratelocally85, thus providing a long-term source of antigen to stimulaterejection84. In this light, alternative pathways to use DCs to inducetransplantation tolerance are being assessed. One involves the activa-tion of recipient natural killer cells, which reject donor DCs in tissueculture models and in vivo. Another pathway is to induce DCs tobecome tolerogenic, for example, to express the tolerogenic ILT3/LILRB4 molecule, or to induce graft-specific FOXP31 Treg cells tosuppress graft-rejecting T cells81. In haematopoietic cell transplanta-tion, strategies that lead to recipient DC depletion are currently beingtested in the clinic.
Extending DC biology into medicine
Immunology, including T-cell-mediated immunity, has a central rolein understanding how disease develops and in designing new treat-ments (Fig. 4). Here we suggest therapies aimed at the upstreamevents initiated by DCs. DCs can either intensify or subdue T-cell
NATUREjVol 449j27 September 2007 REVIEWS
423Nature ©2007 Publishing Group

responses, depending on whether resistance or tolerance needs to beincreased.
Considerable evidence from studies in mice, which predominatesin this review, shows how DCs act in a disease-specific manner.Increased emphasis on the antigens that elicit disease should providemedical approaches that are longer-lasting and less subject to sideeffects than current antigen non-specific ones. To repeat, DC-basedanalyses and therapies emphasize initial events in complex diseasecascades, but more patient-based research is needed to take DCs intomedicine.
The scientific rationale is that a patient’s response to diseaseinvolves not only antigens and lymphocytes but also DCs that controlantigen presentation and clonal selection (the immune recognitionrepertoire) as well as lymphocyte growth, differentiation and mem-ory (the immune response repertoire). Each feature of DCs in Fig. 2 isan intricate area for research, the findings of which are facilitating adeeper analysis of disease and how to interrupt its progress. However,major gaps exist to take this science into medicine.
In mice, the current research emphasis has been on model anti-gens, isolated DCs, and highly selected populations of T cells with asingle transgenic antigen receptor. Instead, research on DCs needs tobe directed to the control of immune responses (1) in situ in intactanimals, (2) within the natural immune repertoire, (3) to clinicallyrelevant antigens, and (4) with new immune enhancers or adjuvantsthat act in defined ways on DCs. Even so, the immune systems of miceand men differ in many aspects. A demanding subject, but one whichwill enhance disease research, is the development of mice withimmune systems derived from human sources.
More research needs to be done with patients. The patient sets thestandards for the quality of knowledge that is required to understandmany aspects of disease and its treatment. Often the pathogen(tumour, microbe, allergen, stimulus for autoimmune or autoin-flammatory disease) is not easily or completely modelled in mice.
Although scientists and the public both desire ‘translation’ frommice to humans, there is an underappreciated need to create asizeable limb of the scientific enterprise that will bring new methods,concepts and coordination to the basic study of disease with patients.This need is illustrated by the paucity of DC-based studies on theimmunotherapy of cancer. Basic research with patients differs sub-stantially from current outcome studies and drug-licensing trials,which test whether existing practices and concepts are clinicallyeffective. In contrast, disease research yields new ideas and therapies.The biology of DCs is ready to be extended to dissect disease path-ways and to direct its prevention and treatment. Despite theobstacles to research in patients, physicians and scientists have theknowledge and tools to think systemically about diseases, plan theirtherapies, and investigate how humans respond. DCs are an early
player in disease development and an unavoidable target in thedesign of treatments.
1. Steinman, R. M. & Cohn, Z. A. Identification of a novel cell type in peripherallymphoid organs of mice. I. Morphology, quantitation, tissue distribution. J. Exp.Med. 137, 1142–1162 (1973).
2. Banchereau, J. & Steinman, R. M. Dendritic cells and the control of immunity.Nature 392, 245–252 (1998).
3. Janeway, C. A. Jr & Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol.20, 197–216 (2002).
4. Guermonprez, P., Valladeau, J., Zitvogel, L., Thery, C. & Amigorena, S. Antigenpresentation and T cell stimulation by dendritic cells. Annu. Rev. Immunol. 20,621–667 (2002).
5. Trombetta, E. S. & Mellman, I. Cell biology of antigen processing in vitro and in vivo.Annu. Rev. Immunol. 23, 975–1028 (2005).
6. Cyster, J. G. Chemokines and the homing of dendritic cells to the T cell areas oflymphoid organs. J. Exp. Med. 189, 447–450 (1999).
7. Itano, A. A. & Jenkins, M. K. Antigen presentation to naive CD4 T cells in thelymph node. Nature Immunol. 4, 733–739 (2003).
8. Randolph, G. J., Angeli, V. & Swartz, M. A. Dendritic-cell trafficking to lymphnodes through lymphatic vessels. Nature Rev. Immunol. 5, 617–628 (2005).
9. Hawiger, D. et al. Dendritic cells induce peripheral T cell unresponsiveness understeady state conditions in vivo. J. Exp. Med. 194, 769–780 (2001).
10. Probst, H. C., McCoy, K., Okazaki, T., Honjo, T. & van den Broek, M. Restingdendritic cells induce peripheral CD81 T cell tolerance through PD-1 and CTLA-4.Nature Immunol. 6, 280–286 (2005).
11. Luo, X. et al. Dendritic cells with TGF-b1 differentiate naive CD41CD252 T cellsinto islet-protective Foxp31 regulatory T cells. Proc. Natl Acad. Sci. USA 104,2821–2826 (2007).
12. Pulendran, B. et al. Distinct dendritic cell subsets differentially regulate the class ofimmune responses in vivo. Proc. Natl Acad. Sci. USA 96, 1036–1041 (1999).
13. Maldonado-Lopez, R. et al. CD8a1 and CD8a2 subclasses of dendritic cells directthe development of distinct T helper cells in vivo. J. Exp. Med. 189, 587–592(1999).
14. Napolitani, G., Rinaldi, A., Bertoni, F., Sallusto, F. & Lanzavecchia, A. Selected Toll-like receptor agonist combinations synergistically trigger a T helper type1-polarizing program in dendritic cells. Nature Immunol. 6, 769–776 (2005).
15. Seder, R. A., Paul, W. E., Davis, M. M., Fazekas de St. & Groth, B. The presence ofinterleukin 4 during in vitro priming determines the lymphokine-producingpotential of CD41 T cells from T cell receptor transgenic mice. J. Exp. Med. 176,1091–1098 (1992).
16. Leibundgut-Landmann, S. et al. Syk- and CARD9-dependent coupling of innateimmunity to the induction of T helper cells that produce interleukin 17. NatureImmunol. 8, 630–638 (2007).
17. Jonuleit, H., Schmitt, E., Schuler, G., Knop, J. & Enk, A. H. Induction of interleukin10-producing, nonproliferating CD41 T cells with regulatory properties byrepetitive stimulation with allogeneic immature human dendritic cells. J. Exp. Med.192, 1213–1222 (2000).
18. Badovinac, V. P., Messingham, K. A., Jabbari, A., Haring, J. S. & Harty, J. T.Accelerated CD81 T-cell memory and prime-boost response after dendritic-cellvaccination. Nature Med. 11, 748–756 (2005).
19. Trumpfheller, C. et al. Intensified and protective CD41 T cell immunity at amucosal surface after a single dose of anti-dendritic cell HIV gag fusion antibodyvaccine. J. Exp. Med. 203, 607–617 (2006).
20. Huang, F.-P. et al. A discrete subpopulation of dendritic cells transports apoptoticintestinal epithelial cells to T cell areas of mesenteric lymph nodes. J. Exp. Med.191, 435–442 (2000).
21. Niess, J. H. et al. CX3CR1-mediated dendritic cell access to the intestinal lumenand bacterial clearance. Science 307, 254–258 (2005).
22. Lindquist, R. L. et al. Visualizing dendritic cell networks in vivo. Nature Immunol. 5,1243–1250 (2004).
23. Reis e Sousa, C. Dendritic cells in a mature age. Nature Rev. Immunol. 6, 476–483(2006).
24. Dudziak, D. et al. Differential antigen processing by dendritic cell subsets in vivo.Science 315, 107–111 (2007).
25. Shortman, K. & Naik, S. H. Steady-state and inflammatory dendritic-celldevelopment. Nature Rev. Immunol. 7, 19–30 (2007).
26. Dalod, M. et al. Interferon a/b and interleukin 12 responses to viral infections:pathways regulating dendritic cell cytokine expression in vivo. J. Exp. Med. 195,517–528 (2002).
27. Probst, H. C. & van den Broek, M. Priming of CTLs by lymphocytic choriomeningitisvirus depends on dendritic cells. J. Immunol. 174, 3920–3924 (2005).
28. Figdor, C. G., van Kooyk, Y. & Adema, G. J. C-type lectin receptors on dendriticcells and Langerhans cells. Nature Rev. Immunol. 2, 77–84 (2002).
29. Gautier, G. et al. A type I interferon autocrine-paracrine loop is involved in Toll-likereceptor-induced interleukin-12p70 secretion by dendritic cells. J. Exp. Med. 201,1435–1446 (2005).
30. Fritz, J. H. et al. Nod1-mediated innate immune recognition of peptidoglycancontributes to the onset of adaptive immunity. Immunity 26, 445–459 (2007).
31. Coutanceau, E. et al. Selective suppression of dendritic cell functions byMycobacterium ulcerans toxin mycolactone. J. Exp. Med. 204, 1395–1403 (2007).
Disease development
Immune evasionMicrobes and tumours can block DC function, for example, maturation, migration, or they can exploit DCs to replicate
Immune pathogenesisDCs instigate innate and adaptive responses leading to allergy,autoimmunity and graft rejection,for example, TH2 polarization of T cells, the overproduction of cytokines and cross-presentation of grafted tissues
Disease treatment
Immune resistanceMaturing DCs loaded ex vivowith tumour or microbialantigens, or by antigen targeting in vivo, induceimmunity
Immune silencingDCs loaded with antigens arerendered tolerogenic, forexample, with TGF-β,retinoic acid, rapamycin orsteroids
Figure 4 | Some potential roles for DCs in medicine.
REVIEWS NATUREjVol 449j27 September 2007
424Nature ©2007 Publishing Group

32. Querec, T. et al. Yellow fever vaccine YF-17D activates multiple dendritic cellsubsets via TLR2, 7, 8, and 9 to stimulate polyvalent immunity. J. Exp. Med. 203,413–424 (2006).
33. Albert, M. L., Sauter, B. & Bhardwaj, N. Dendritic cells acquire antigen fromapoptotic cells and induce class I-restricted CTLs. Nature 392, 86–89 (1998).
34. Khader, S. A. et al. Interleukin 12p40 is required for dendritic cell migration and Tcell priming after Mycobacterium tuberculosis infection. J. Exp. Med. 203,1805–1815 (2006).
35. Geijtenbeek, T. B. H. et al. DC-SIGN, a dendritic cell specific HIV-1 binding proteinthat enhances trans-infection of T cells. Cell 100, 587–597 (2000).
36. Colonna, M., Krug, A. & Cella, M. Interferon-producing cells: on the front line inimmune responses against pathogens. Curr. Opin. Immunol. 14, 373–379 (2002).
37. Boscardin, S. B. et al. Antigen targeting to dendritic cells elicits long-lived T cellhelp for antibody responses. J. Exp. Med. 203, 599–606 (2006).
38. Soares, H. et al. A subset of dendritic cells induces CD41 T cells to produce IFN-cby an IL-12-independent but CD70-dependent mechanism in vivo. J. Exp. Med.204, 1095–1106 (2007).
39. Bozzacco, L. et al. DEC-205 receptor on dendritic cells mediates presentation ofHIV gag protein to CD81 T cells in a spectrum of human MHC I haplotypes. Proc.Natl Acad. Sci. USA 104, 1289–1294 (2007).
40. Delneste, Y. et al. Involvement of LOX-1 in dendritic cell-mediated antigen cross-presentation. Immunity 17, 353–362 (2002).
41. Gabrilovich, D. Mechanisms and functional significance of tumour-induceddendritic-cell defects. Nature Rev. Immunol. 4, 941–952 (2004).
42. Kortylewski, M. et al. Inhibiting Stat3 signaling in the hematopoietic system elicitsmulticomponent antitumor immunity. Nature Med. 11, 1314–1321 (2005).
43. Ghiringhelli, F. et al. Tumor cells convert immature myeloid dendritic cells intoTGF-b-secreting cells inducing CD41CD251 regulatory T cell proliferation. J. Exp.Med. 202, 919–929 (2005).
44. Aspord, C. et al. Breast cancer instructs dendritic cells to prime interleukin 13-secreting CD41 T cells that facilitate tumor development. J. Exp. Med. 204,1037–1047 (2007).
45. Vicari, A. P., Caux, C. & Trinchieri, G. Tumour escape from immune surveillancethrough dendritic cell inactivation. Semin. Cancer Biol. 12, 33–42 (2002).
46. Dhodapkar, K. M., Krasovsky, J., Williamson, B. & Dhodapkar, M. V. Anti-tumormonoclonal antibodies enhance cross-presentation of cellular antigens and thegeneration of myeloma-specific killer T cells by dendritic cells. J. Exp. Med. 195,125–133 (2002).
47. Kalergis, A. M. & Ravetch, J. V. Inducing tumor immunity through the selectiveengagement of activating Fcc receptors on dendritic cells. J. Exp. Med. 195,1653–1659 (2002).
48. Blattman, J. N. & Greenberg, P. D. Cancer immunotherapy: a treatment for themasses. Science 305, 200–205 (2004).
49. Shankaran, V. et al. IFNc and lymphocytes prevent primary tumour developmentand shape tumour immunogenicity. Nature 410, 1107–1111 (2001).
50. Figdor, C. G., De Vries, I. J., Lesterhuis, W. J. & Melief, C. J. Dendritic cellimmunotherapy: mapping the way. Nature Med. 10, 475–480 (2004).
51. Banchereau, J. & Palucka, A. K. Dendritic cells as therapeutic vaccines againstcancer. Nature Rev. Immunol. 5, 296–306 (2005).
52. Schuler-Thurner, B. et al. Rapid induction of tumor-specific type 1 T helper cells inmetastatic melanoma patients by vaccination with mature, cryopreserved,peptide-loaded monocyte-derived dendritic cells. J. Exp. Med. 195, 1279–1288(2002).
53. Paczesny, S. et al. Expansion of melanoma-specific cytolytic CD81 T cellprecursors in patients with metastatic melanoma vaccinated with CD341
progenitor-derived dendritic cells. J. Exp. Med. 199, 1503–1511 (2004).54. Fernandez, N. C. et al. Dendritic cells directly trigger NK cell functions: cross-talk
relevant in innate anti-tumor immune responses in vivo. Nature Med. 5, 405–411(1999).
55. Lucas, M., Schachterle, W., Oberle, K., Aichele, P. & Diefenbach, A. Dendritic cellsprime natural killer cells by trans-presenting interleukin 15. Immunity 26, 503–517(2007).
56. Stary, G. et al. Tumoricidal activity of TLR7/8-activated inflammatory dendriticcells. J. Exp. Med. 204, 1441–1451 (2007).
57. Palucka, A. K. et al. Dendritic cells loaded with killed allogeneic melanoma cellscan induce objective clinical responses and MART-1 specific CD81 T-cellimmunity. J. Immunother. 29, 545–557 (2006).
58. O’Rourke, M. G. et al. Durable complete clinical responses in a phase I/II trialusing an autologous melanoma cell/dendritic cell vaccine. Cancer Immunol.Immunother. 52, 387–395 (2003).
59. De Vries, I. J. et al. Effective migration of antigen-pulsed dendritic cells to lymphnodes in melanoma patients is determined by their maturation state. Cancer Res.63, 12–17 (2003).
60. Banchereau, J. et al. Immune and clinical responses in patients with metastaticmelanoma to CD341 progenitor-derived dendritic cell vaccine. Cancer Res. 61,6451–6458 (2001).
61. Obeid, M. et al. Calreticulin exposure dictates the immunogenicity of cancer celldeath. Nature Med. 13, 54–61 (2007).
62. Gilboa, E. & Vieweg, J. Cancer immunotherapy with mRNA-transfected dendriticcells. Immunol. Rev. 199, 251–263 (2004).
63. Soiffer, R. et al. Vaccination with irradiated autologous melanoma cellsengineered to secrete human granulocyte-macrophage colony-stimulating factor
generates potent antitumor immunity in patients with metastatic melanoma.Proc. Natl Acad. Sci. USA 95, 13141–13146 (1998).
64. Laheru, D. & Jaffee, E. M. Immunotherapy for pancreatic cancer—science drivingclinical progress. Nature Rev. Cancer 5, 459–467 (2005).
65. Fujii, S., Liu, K., Smith, C., Bonito, A. J. & Steinman, R. M. The linkage of innate toadaptive immunity via maturing dendritic cells in vivo requires CD40 ligation inaddition to antigen presentation and CD80/86 costimulation. J. Exp. Med. 199,1607–1618 (2004).
66. Banchereau, J. & Pascual, V. Type I interferon in systemic lupus erythematosusand other autoimmune diseases. Immunity 25, 383–392 (2006).
67. Lowes, M. A. et al. Increase in TNFa and inducible nitric oxide synthase-expressing dendritic cells in psoriasis and reduction with efalizumab (anti-CD11a). Proc. Natl Acad. Sci. USA 102, 19057–19062 (2005).
68. Bennett, L. et al. Interferon and granulopoiesis signatures in systemic lupuserythematosus blood. J. Exp. Med. 197, 711–723 (2003).
69. Siegal, F. P. et al. The nature of the principal type 1 interferon-producing cells inhuman blood. Science 284, 1835–1837 (1999).
70. Asselin-Paturel, C. & Trinchieri, G. Production of type I interferons: plasmacytoiddendritic cells and beyond. J. Exp. Med. 202, 461–465 (2005).
71. Tian, J. et al. Toll-like receptor 9-dependent activation by DNA-containingimmune complexes is mediated by HMGB1 and RAGE. Nature Immunol. 8,487–496 (2007).
72. Blanco, P., Palucka, A. K., Gill, M., Pascual, V. & Banchereau, J. Induction ofdendritic cell differentiation by IFN-a in systemic lupus erythematosus. Science294, 1540–1543 (2001).
73. Nestle, F. O. et al. Plasmacytoid predendritic cells initiate psoriasis throughinterferon-a production. J. Exp. Med. 202, 135–143 (2005).
74. Hue, S. et al. Interleukin-23 drives innate and T cell-mediated intestinalinflammation. J. Exp. Med. 203, 2473–2483 (2006).
75. Tarbell, K. V. et al. Dendritic cell-expanded, islet-specific, CD41 CD251 CD62L1
regulatory T cells restore normoglycemia in diabetic NOD mice. J. Exp. Med. 204,191–201 (2007).
76. Lambrecht, B. N. & Hammad, H. Taking our breath away: dendritic cells in thepathogenesis of asthma. Nature Rev. Immunol. 3, 994–1003 (2003).
77. Soumelis, V. et al. Human epithelial cells trigger dendritic cell-mediated allergicinflammation by producing TSLP. Nature Immunol. 3, 673–680 (2002).
78. Traidl-Hoffmann, C. et al. Pollen-associated phytoprostanes inhibit dendritic cellinterleukin-12 production and augment T helper type 2 cell polarization. J. Exp.Med. 201, 627–636 (2005).
79. Hammad, H. et al. Activation of the D prostanoid 1 receptor suppresses asthma bymodulation of lung dendritic cell function and induction of regulatory T cells. J.Exp. Med. 204, 357–367 (2007).
80. Inaba, K. et al. Efficient presentation of phagocytosed cellular fragments on theMHC class II products of dendritic cells. J. Exp. Med. 188, 2163–2173 (1998).
81. Ochando, J. C. et al. Alloantigen-presenting plasmacytoid dendritic cells mediatetolerance to vascularized grafts. Nature Immunol. 7, 652–662 (2006).
82. Hackstein, H. & Thomson, A. W. Dendritic cells: emerging pharmacologicaltargets of immunosuppressive drugs. Nature Rev. Immunol. 4, 24–34 (2004).
83. Shlomchik, W. D. et al. Prevention of graft versus host disease by inactivation ofhost antigen-presenting cells. Science 285, 412–415 (1999).
84. Merad, M. et al. Depletion of host Langerhans cells before transplantation ofdonor alloreactive T cells prevents skin graft-versus-host disease. Nature Med. 10,510–517 (2004).
85. Merad, M. et al. Langerhans cells renew in the skin throughout life under steady-state conditions. Nature Immunol. 3, 1135–1141 (2002).
86. Rescigno, M. et al. Dendritic cells express tight junction proteins and penetrate gutepithelial monolayers to sample bacteria. Nature Immunol. 2, 361–367 (2001).
87. Coombes, J. L. et al. A functionally specialized population of mucosal CD1031 DCsinduces Foxp31 regulatory T cells via a TGF-b- and retinoic acid-dependentmechanism. J. Exp. Med. 204, 1757–1764 (2007).
88. Bousso, P. & Robey, E. Dynamics of CD81 T cell priming by dendritic cells in intactlymph nodes. Nature Immunol. 4, 579–585 (2003).
89. Shakhar, G. et al. Stable T cell–dendritic cell interactions precede thedevelopment of both tolerance and immunity in vivo. Nature Immunol. 6, 707–714(2005).
90. Jung, S. et al. In vivo depletion of CD11c1 dendritic cells abrogation priming ofCD81 T cells by exogenous cell-associated antigens. Immunity 17, 211–220(2002).
91. Heath, W. R. et al. Cross-presentation, dendritic cell subsets, and the generationof immunity to cellular antigens. Immunol. Rev. 199, 9–26 (2004).
92. Pulendran, B. Variegation of the immune response with dendritic cells andpathogen recognition receptors. J. Immunol. 174, 2457–2465 (2005).
93. Stranges, P. B. et al. Elimination of antigen-presenting cells and autoreactive Tcells by Fas contributes to prevention of autoimmunity. Immunity 26, 629–641(2007).
94. Ohteki, T. et al. Essential roles of DC-derived IL-15 as a mediator of inflammatoryresponses in vivo. J. Exp. Med. 203, 2329–2338 (2006).
95. Granucci, F. et al. Inducible IL-2 production by dendritic cells revealed by globalgene expression analysis. Nature Immunol. 2, 882–888 (2001).
96. Reis e Sousa, C. et al. In vivo microbial stimulation induces rapid CD40L-independent production of IL-12 by dendritic cells and their re-distribution to Tcell areas. J. Exp. Med. 186, 1819–1829 (1997).
NATUREjVol 449j27 September 2007 REVIEWS
425Nature ©2007 Publishing Group

97. Sporri, R. & Reis e Sousa, C. Inflammatory mediators are insufficient for fulldendritic cell activation and promote expansion of CD41 T cell populationslacking helper function. Nature Immunol. 6, 163–170 (2005).
98. Sallusto, F. et al. Rapid and coordinated switch in chemokine receptorexpression during dendritic cell maturation. Eur. J. Immunol. 28, 2760–2769(1998).
99. Piqueras, B., Connolly, J., Freitas, H., Palucka, A. K. & Banchereau, J. Upon viralexposure, myeloid and plasmacytoid dendritic cells produce 3 waves of distinctchemokines to recruit immune effectors. Blood 107, 2613–2618 (2006).
100. Ito, T. et al. TSLP-activated dendritic cells induce an inflammatory T helper type 2cell response through OX40 ligand. J. Exp. Med. 202, 1213–1223 (2005).
101. Kissenpfennig, A. et al. Dynamics and function of Langerhans cells in vivo: dermaldendritic cells colonize lymph node areas distinct from slower migratingLangerhans cells. Immunity 22, 643–654 (2005).
102. Del Rio, M. L., Rodriguez-Barbosa, J. I., Kremmer, E. & Forster, R. CD1032 andCD1031 bronchial lymph node dendritic cells are specialized in presenting andcross-presenting innocuous antigen to CD41 and CD81 T Cells. J. Immunol. 178,6861–6866 (2007).
103. Yoneyama, H. et al. Plasmacytoid DCs help lymph node DCs to induce anti-HSVCTLs. J. Exp. Med. 202, 425–435 (2005).
104. D’Amico, A. & Wu, L. The early progenitors of mouse dendritic cells andplasmacytoid predendritic cells are within the bone marrow hemopoieticprecursors expressing Flt3. J. Exp. Med. 198, 293–303 (2003).
105. Karsunky, H., Merad, M., Cozzio, A., Weissman, I. L. & Manz, M. G. Flt3 ligandregulates dendritic cell development from Flt31 lymphoid and myeloid-committed progenitors to Flt31 dendritic cells in vivo. J. Exp. Med. 198, 305–313(2003).
106. Leon, B., Lopez-Bravo, M. & Ardavin, C. Monocyte-derived dendritic cells formedat the infection site control the induction of protective T helper 1 responsesagainst Leishmania. Immunity 26, 519–531 (2007).
107. Liu, K. et al. Origin of dendritic cells in peripheral lymphoid organs of mice. NatureImmunol. 8, 578–583 (2007).
108. Varol, C. et al. Monocytes give rise to mucosal, but not splenic, conventionaldendritic cells. J. Exp. Med. 204, 171–180 (2007).
Acknowledgements The authors thank C. Moberg and M. Nussenzweig forextensive comments and J. Adams for assistance with the manuscript. We aregrateful to our patients and colleagues, for their many contributions, and to the NIHand several foundations for support. A version of the review with a more completebibliography is published on the authors’ websites: www.biir.org andwww.rockefeller.edu/labheads/steinman/generalReviews.php.
Author Information Reprints and permissions information is available atwww.nature.com/reprints. The authors declare competing financial interests:details accompany the full-text HTML version of the paper at www.nature.com/nature. Correspondence should be addressed to R.M.S.([email protected]).
REVIEWS NATUREjVol 449j27 September 2007
426Nature ©2007 Publishing Group