Embed Size (px)
Citation preview

TAURINE STIMULATION OF CALCIUM UPTAKE
IN THE RETINA: MECHANISM OF ACTION
by
JULIUS D. MILITANTS, B.S., M.S.
A DISSERTATION
IN
PHARMACOLOGY
Submitted to the Graduate Faculty of Texas Tech University Health Sciences Center
in Partial Fulfillment of the Requfrements for the Degree of
DOCTOR OF PHILOSOPHY
Advisory Committee
John B. Lombardini (Chairperson) Michael P. Blanton
Howard K. Strahlendorf Jean C. Strahlendorf Thomas E. Tenner
Accepted
Associate D § ^ of the Gradu§ft>echool of Biomedical Sciences Texas Tech University Health Sciences Center
May, 2003

ACKNOWLEDGEMENTS
I would like to thank God, our father in heaven, who has taken care of
me all these years and who has helped me be hopeful in spite of adversity
and happy in spite of difficulty. Through my long journey. He has always
truly made His loving presence known to me, and for this I feel so
privileged. I can only wish that everyone allows Him into their heart as He
has come into mine. All that we see will pass, and God's love will live
forever.
Next, I would like to thank Dr. John B. Lombardini, who gave me a
job when I had nothing else to do and nowhere else to go. Working for him
as his technician and his PhD student was reward enough in it by itself It is
only right that he be the one to mentor me to the most important
achievement I have in my life. I truly hope I have made him proud.
As with Dr. Lombardini, Dr. Tina K. Machu gave me a job when I
needed it, and yet again, my work had become replete with fulfillment and
satisfaction. It is akin to having lightning strike twice. I thank her for all
her help and friendship, and for the wonderful shine she has added to my
life and my future.
The members of my committee have been most encouraging,
especially in light of the unusual circumstances of my education. It was
very easy for me to give up on nodon that I should get the PhD, but my
committee made cle tr their support and urged me on. At rimes, I feel it is
more of their will that propelled me forward than it was mine. Without
them, I truly would have given up in despair and friastration. Thank you so
11

much, Dr. Michael P. Blanton, Dr. Howard K. Strahlendorf, Dr. Jean C.
Strahlendorf and Dr. Thomas E. Tenner. I pray that in the fiiture, I can live
up to the faith you had in me.
I must thank all my fi-iends in Lubbock, in the department, among the
students, in the whole school and outside. There are too many to mention.
Your love and fi"iendship give me life and fill my life. I cannot imagine not
having y'all there... imagining all of you there makes all hardship seem so
small. Just the thought of my friends truly makes me happy. Thank you so
much for being there.
Lastly, to my family, my mom, sister and brothers especially, your
love is unfailing, and I know it isn't because you are family. It is truly
wonderful to have you always with me, in thought and spirit. I hope my
work brings you joy and pride, for you deserve all the good that comes your
way. My future I offer to you and I hope that there will be greater things for
us all. To my departed father and brother, I offer this little achievement,
too, and hope you can be proud of me there in heaven, if it be allowed. I do
wish that you both were here.
Ill

TABLE OF CONTENTS
ACKNOWLEDGEMENTS ii
ABSTRACT vii
LIST OF TABLES x
LIST OF FIGURES xi
CHAPTER
L INTRODUCTION 1
Background 1
Physiologic significance of taurine 1
Taurine depletion experiments 3
Immunolocalization of taurine 6
Taurine uptake in retinal tissue 8
Taurine binding to retinal dssue 11
Taurine and calcium uptake in the retina 12
Calcium uptake and calcium binding
in the retina 14
Calcium flux and phototransduction 15
Taurine and protein phosphorylation 16 Hypotheses 17
iv

Main Interest 17
ATPase activity in the retina 18
Modulation of taurine uptake 19
Possible modulation of calcium channels 19
Calcium uptake and calcium binding 20
IL METHODS 22
Preparation of dssue samples 22
Protein assay 23
Calcium uptake assay 23
ATPase assay 24
Taurine uptake assay 25
Calcium binding assay 25
Stadstical analysis 26
m. RESULTS 29
Preliminary data 29
Hypothesis 1 37
Hypothesis 2 43
Hypothesis 3 59
Hypothesis 4 68

IV. DISCUSSION 72
Taurine effects of calcium uptake 72
ATPase activity in the retina 76
Taurine uptake versus taurine binding 79
The experimental use of chelerythrine (CHT) 83
Modulation of calcium channels 84
Calcium uptake versus calcium binding 90
Conclusions 93
REFERENCES 95
VI

ABSTRACT
Taurine is a fi-ee amino acid found in millimolar concentrations inside
most animal cells, and the retina appears to possess the greatest amount of
taurine compared to the other cell types. Taurine modulates calcium uptake
in retinal dssue, suggesting that the physiologic function of taurine may be
related to calcium. Taurine is known to stimulate calcium uptake in the
presence of low calcium levels (-10-500 |iM) in the presence of ATP, and
the mechanism behind this effect of taurine was studied. Much is unknown
about this specific effect of taurine. What is the site of acdon of taurine?
What is the nature of the calcium uptake? Most importantly, is this
pardcular type of calcium uptake, and in turn the effects of taurine,
physiologically relevant? Truly, the main interest of this thesis is the
possible physiologic relevance of the sdmulation produced by taurine. The
specific questions that were addressed relate to the nature of the ATP-
dependence of stimulation by taurine, the site of action of taurine, and the
nature of the calcium uptake that taurine increases.
Chelerythrine (CHT) is a potent protein kinase C (PKC) and ATPase
inhibitor that has been previously shown to inhibit taurine-related effects,
specifically in vitro CHT treatment produced an increased in the
phosphorylation of proteins that taurine specifically inhibited. The
discovery of the possible interaction between taurine and CHT suggested
the use of CHT as a possible pharmacological tool in the study of the effects
of taurine. Experiments using CHT were conducted that tested the
hypothesis that taurine modulates ATPase activity in the retina. CHT
VII

inhibited the stimulatory effects of taurine on retinal calcium uptake, and it
was used to help define the mechanism of acdon of taurine.
Among other effects, CHT inhibits ATPase activity in the retina, and
because the stimulatory effects of taurine are ATP-dependent, the data
suggested that ATPase activity may be involved in taurine stimulation of
calcium uptake. Thus, the role of ATPase activity in the mechanism of
action of taurine was studied. Taurine had no direct effect on ATPase
acdvity and so the involvement of ATPase activity was discounted.
CHT also inhibited taurine uptake, suggesting another mechanism by
which CHT may antagonize the stimulatory effects of taurine on calcium
uptake. Taurine uptake was studied relative to its stimulatory effects. The
kiendcs of taurine uptake were determined in both whole retinal
homogenate and in isolated rod outer segments (ROS). In the whole retina,
two uptake components were defined, one of low-affinity and the other of
high-affinity. In contrast, only one uptake system of high-affinity was
observed in ROS. Another series of experiments were conducted to address
the second hypothesis that the inhibidon of taurine uptake abolishes or
attenuates the stimulatory effects of taurine on calcium uptake. An
analogue of taurine, guanidinoethane sulfonic acid (GES) was found to
effecdvely inhibit taurine uptake. Inhibidon of taurine uptake with GES
surprisingly did not produce any effects, eliminating taurine uptake as a
necessary event behind taurine-dependent sdmulation of calcium uptake.
The data suggested that taurine binds to the membrane to produce its
effects.
Vll l

The nature of the calcium uptake was a logical succeeding question to
the stimulatory effects of taurine. Reference literature described the
modulatory effects of taurine on ion channels in the heart and in the skeletal
muscle, which suggested the possible involvement of calcium channel
activation in the mechanism of action of taurine. Experiments were
conducted to test the third hypothesis that specific inhibition of calcium
channels abolishes or attenuates the stimulatory effects of taurine. The
effects of taurine were antagonized by cation channel blockers, specifically
by pharmacologic blockers of cGMP-gated channels. The data strongly
suggested that taurine exerts a stimulatory effect on this channel to increase
calcium uptake. The mechanism behind this effect on ion channels is
unknown.
Lasdy, experiments were conducted to test the fourth hypothesis that
taurine does not affect calcium binding to rednal membranes. Taurine is
known to modulate calcium binding to sarcolemmal membranes and so the
stimulatory effects of taurine may include the stimuladon of calcium
binding. The involvement of calcium binding was ruled out with the use of
binding experiments and calcium ionophore treatments, suggesdng that the
increase in calcium uptake induced by taurine is solely due to increased flux
through calcium channels. The mechanism of taurine, thus, can be
summarized: Taurine binds to membranes to modulate the activadon of
calcium channels and increase calcium uptake, a process which does not
involve ATPase acdvity, taurine uptake or calcium binding.
IX

LIST OF TABLES
1. Inhibitory potency (IC50) of CHT against calcium uptake and ATPase activity in rat retinal homogenate 33
2. Kinetic constants for taurine uptake calculated from data shown in Figures 12 and 14 46
3. Inhibitory potency (IC50) for CHT inhibition of high- and low-affinity uptake in whole retinal homogenate, and of high-affinity taurine uptake in isolated ROS 51
4. Half-maximal inhibition constant values (IC50) for GES inhibition of high- and low-affinity uptake in whole rednal homogenate, and of high-affinity taurine uptake in isolated ROS 55

LIST OF FIGURES
1. Calcium uptake in whole retinal homogenate in the presence of taurine alone 29
2. Calcium uptake in whole retinal homogenate in the presence of 1.2 mM ATP ± 32 mM 30
3. The inhibitory effects of CHT on calcium uptake in whole rednal homogenate in the absence and presence of 1.2 mM ATP ± 32 mM taurine.
(A) Raw uptake data (mean ± SEM, N = 5-7) 31
(B) Calcium uptake in the whole retina homogenate in the presence of 1.2 mM ATP presented in Figure 3(A) converted to % control, and best-fit non-linear regression curve (N = 6-7) 32
(C) Calcium uptake in the whole retina homogenate in the presence of 1.2 mM ATP and 32 mM taurine presented in Figure 3(A) converted to % control, and best-fit non-linear regression curve (N = 6-7) 32
4. Calcium uptake in whole retinal homogenate in the presence of increasing concentrations of ATP ± 100 |iM CHT 34
5. Calcium uptake in whole rednal homogenate in the presence of increasing concentradons oftaurineandl.2mMATP ilOOflMCHT 35
6. The effect of the PKC inhibitor K252b on calcium uptake in whole retinal homogenate in the presence of 1.2 mM ATP ± 32 mM taurine 36
XI

7. ATPase activity in whole retinal homogenate in the presence of increasing concentradons of ATP 37
8. ATPase activity in whole retinal homogenate in the presence of 1.2 mM ATP ± 32 mM taurine and of increasing concentradons of CHT.
(A) Raw ATPase data (mean ± SEM, N = 3) 39
(B) Data presented in Figure 8(A) expressed in terms of % control, and best-fit non-linear regression curve 39
9. ATPase acdvity in whole retinal homogenate in the presence of increasing concentradons of ouabain 41
10. ATPase acdvity in the presence of 100 |iM CHT, 1 mM ouabain (OUA), 1.2 mM ATP ± 32 mM taurine 41
11. ATPase activity in whole retinal homogenate in the presence of increasing concentradons of thapsigargin 42
12. Taurine uptake in whole retinal homogenate in the
presence of increasing concentradons of total taurine 43
13. Eadie Hofstee plot of data fi"om Figure 12 44
14. Taurine uptake in isolated ROS in the presence of increasing concentrations of total taurine 45
15. Eadie Hofstee plot ofdata from Figure 14 46
16. High-affinity taurine uptake in whole retinal homogenate in the presence of increasing concentradons of CHT.
(A) Raw uptake data (mean ± SEM, N = 3, *p < 0.05 vs. 0 liM CHT, Dunnett's posttest) 48
Xll

(B) Data presented in Figure 16(A) expressed in terms of % cond-ol, and best-fit non-linear regression curve (N = 3) 48
17. Low-affinity taurine uptake in whole retinal homogenate in the presence of increasing concentradons of CHT.
(A) Raw uptake data (mean ± SEM, N = 3, *p < 0.05 vs. 0 |iM CHT, Dunnett's posttest) 49
(B) Data presented in Figure 17(A) expressed in terms of % control, and best-fit non-linear regression curve (N = 3) 49
18. High-affinity taurine uptake in isolated ROS in the presence of increasing concentradons of CHT.
(A) Raw uptake data (mean ± SEM, N = 3, p < 0.05 vs. 0 |iM CHT, Dunnett's posttest) 50 *
(B) Data presented in Figure 18(A) expressed in terms of % control, and best-fit non-linear regression curve (N = 3) 50
19. Eadie-Hofstee transformation of taurine uptake data as measured in the presence of 25-750 |J,M total taurine ± 5 |iM CHT 52
20. High-affinity taurine uptake in whole retinal homogenate in the presence of phorbol myristate acetate (PMA) and staurosporine (STAU) 53
21. High-affinity taurine uptake in whole retinal homogenates inhibited by GES 54
22. Low-affinity taurine uptake in whole retinal homogenates inhibited by GES 55
23. High-affinity taurine uptake in isolated ROS inhibited by GES 56
Xll l

24, Calcium uptake in whole retinal homogenate in the presence of GES, a taurine uptake inhibitor 57
25, Calcium uptake in isolated ROS in the presence of GES, a taurine uptake inhibitor 58
26, Calcium uptake in whole retinal homogenate in the presence of 1.2 mM ATP ±32 mM taurine, and cadmium, a non-specific cation channel inhibitor 59
27, Calcium uptake in whole retinal homogenate in the presence of 1,2 mM ATP ± 32 mM taurine, and of increasing concentrations of nifedipine, a blocker of voltage-gated L-type calcium channels 60
28, Calcium uptake in whole retinal homogenate in the presence of 1.2 mM ATP ± 32 mM taurine, and of LY83583, a blocker of cGMP-gated channels 61
29, Calcium uptake in isolated ROS in the presence of 1.2 mM ATP ± 32 mM taurine, and of LY83586, a blocker of cGMP-gated channels 62
30, Calcium uptake in isolated ROS in the presence of 1,2 mM ATP ± 32 mM taurine, and Rp-8-Br-PET-cGMPS, a specific blocker of cGMP-gated channels 63
31, Calcium uptake in isolated ROS in the presence of 1,2 mM ATP ±32 mM taurine, and of increasing concentrations of dibutyryl cGMP, a membrane-permeant analogue of cGMP 65
32, Calcium uptake in the isolated ROS in the presence of 1,2 mM ATP and increasing concend-adons of Rp-cGMP, an analogue of cGMP 65
XIV

33. Calcium uptake in the isolated ROS in the presence of 1,2 mM ATP ± 32 mM taurine, and of increasing concentrations of zaprinast, an inhbitor of cGMP-specific phosphodiesterase 67
34, Calcium binding in whole retinal homogenate in the presence of 1,2 mM ATP and/or 32 mM 69
35, Calcium binding in whole retinal homogenate in the presence of increasing concentrations of ATP 69
36, Calcium binding in whole retinal lysate in the presence of ATP and increasing concentrations of taurine 70
37, Calcium uptake in whole retinal lysate in the presence of A23187, a calcium ionophore 71
38, Calcium uptake in lysed ROS in the presenceofA23187, a calcium ionophore 71
XV

CHAPTER I
INTRODUCTION
Background
Physiologic significance of taurine
The bulk of the first chapter was adapted fi-om a review published in
Nutritional Neuroscience (Militante and Lombardini, 2002). Taurine (2-
aminoethanesulfonic acid) is a fi-ee amino acid found in high millimolar
concentradons in animal dssue to which an ever growing number of cellular
and physiologic functions have been atd-ibuted (Wright et al., 1986;
Huxtable, 1992; Sturman, 1993). Taurine has been sUidied as a
neurodansmitter, as an osmoregulator and as an antioxidant, among others.
In addition, the effects of taurine to protect against hepatotoxicity and to
modulate ion channels have been reviewed (Timbrell et al,, 1995; Satoh and
Sperelakis, 1998). However, reviews on the significance of taurine seemed
to have decreased considerably among peer-reviewed journals this past
decade.
Special attendon has been paid to the mechanism of action of taurine
in the retina, especially in the decades of the 70s and 80s, mainly because of
experiments which suggest that taurine is most abundant in the retina
compared to other dssue types and because of studies which linked taurine
deficiency with visual dysflincdon (reviewed in Lombardini, 1991), More
importandy, the simple supplementation of the diet with taurine proved to
be sufficient in alleviating the vision problem, theoretically through the
replenishment of the depleted taurine levels. Taurine is a naUiral product

that is readily available in food and nutrition stores, and is relatively
inexpensive. Thus, the understanding of its actions may provide an
inexpensive health option for many people.
There are other arguments for the physiologic importance of taurine
in the retina. Numerous reports validate the effect of taurine to modulate
cellular processes in vitro. The synthesis, uptake and release of taurine are
tightly regulated, suggesting that the maintenance of taurine levels is crucial
in the normal funcdon of the redna (reviewed in Lombardini, 1991; in
Militante and Lombardini, 2002). Also, taurine exhibits d-ophic effects in
animal models of rednal regeneration (reviewed in Lima, 1999). Moreover,
the decrease in retinal taurine levels appears to be one of the earliest lesions
observed in the degeneration of the retina in rat models of rednitis
pigmentosa (Okada et al,, 2000), However, scientific interest and research
on the physiologic function of taurine have been, for all intents and
purposes, insufficient to the extent that there is still more speculation than
certainty as to the exact mechanism of action of taurine, in the redna or in
other tissues. More studies have to be done, especially in light of the
potential taurine has to affect cell function.
Calcium ions provide a crucial signal in the photodansducdon
process in the rod outer segments (ROS) of the retinal photoreceptor cells
(Baylor, 1996), The acdvadon and deacdvadon of the cGMP-gated cation
channels found in the plasma membrane of the ROS are involved in this
calcium flux. Taurine concend-adons in retinal tissues have been estimated
to be as high as 79 mM (Voaden et al,, 1977), and so it is of great interest
that taurine is known to exert some condol over calcium flux in the redna

(reviewed in Lombardini, 1991). This may be an important physiologic
funcdon of taurine. The research presented in this thesis concerns the
mechanism of acdon behind the specific sdmulatory effect of taurine under
condidons of low calcium concentradon.
Taurine depletion experiments
An important model through which the native function of taurine in
the retina may be established is through the use of animals which have been
nutritionally deprived or pharmacologically depleted of taurine. In fact, this
is the most common methodology used to study the importance of taurine in
the retina. One of the earliest and more well-known models is that of the cat
which is fed a taurine-free casein diet (Schmidt et al., 1976). In this model,
the animal exhibited taurine levels of only 6% and 60% of normal in the
plasma and in the retina, respectively. A similar nutritional protocol has
been used in Rhesus monkeys which utilized taurine-fi-ee human infant
formula (Imaki et al,, 1987; Neuringer and Sturman, 1987), In human
padents, taurine deficiencies were found to occur with total parenteral
nudition that did not include taurine in the food formula (Geggel et al,
1985), and this particular model has also been used to sdidy the physiologic
function of taurine in the retina.
The rat model presents a different metabolic type as the use of a
taurine-fi-ee diet is not effecdve in decreasing the level of taurine in the
retina (Lake, 1982), Antagonists of taurine transport have been used in the
rat to deplete taurine in the redna, specifically, guanidinoethanesulfonic
acid (GES) and p-alanine (Lake, 1981, 1982; Lake and Cocker, 1983;

Quesada et al,, 1984; Lake and de Marte, 1988). The inclusion of GES in
the drinking water (1%) of rats causes a significant decrease in the amino
acid level (>50%) and in the uptake of radiolabeled taurine in the retina
(-60%). It is interesting to note that the retina appears to be particulariy
sensitive to taurine depletion as compared to other neural-type tissues as
GES d-eatment had no effect on the taurine levels in the cerebellum and in
the cerebral cortex (Quesada et al, 1984). Also, other amino acids (GABA,
glycine, glutamate, aspartate, glutamine) do not appear to be affected by
GES d-eatment (Lake, 1981, 1982). Taurine uptake in the retina is also
inhibited by P-alanine but tissue taurine levels are only slighdy decreased
(Lake and de Marte, 1988).
The depledon of taurine has been invariably linked to retinal cell
damage and elecfroretinogram (ERG) abnormalities. Depletion of taurine
results in cellular degeneration and death in the rat (Pasantes-Morales et al,,
1983; Lake, 1986; Lake and Malik, 1987; Cocker and Lake, 1987; Quesada
et al,, 1988; Shimada et al,, 1992; Lombardini et al,, 1996), the cat (Berson
et al,, 1976; Jacobson et al,, 1987; Leon et al, 1995) and in the Rhesus
monkey (Neuringer and Sturman, 1987; Imaki et al , 1993), ERG patterns
were studied in the rat and in the cat, and expectedly, abnormalities were
observed. Behavioral studies revealed deficits in visual acuity in rhesus
monkeys nudidonally depleted of taurine (Neuringer and Sturman, 1987),
In all cases, the reind-oduction of taurine or the withdrawal of the taurine
uptake antagonist reverses or prevents the negadve effects of taurine
depletion. It is in these experiments that the nud-idonal value of taurine has
been most cleariy defined, as the use of taurine supplementadon to reverse

lesions associated with taurine deficiency was proven to be easy, safe and
effecdve.
Of pardcular note is the work by Geggel et al. (1985) which described
ERG abnormalities in human patients suffering taurine deficiency. Patients
on total parenteral nutrition which did not include taurine were found to
have decreased levels of taurine in the blood. Moreover, ERG deficits were
observed in some of the subjects. The authors found that only young
children and not adult patients were affected. While the reason behind this
difference is unclear, it can be surmised that taurine metabolism changes
with age, Indoduction of taurine to the intravenous solutions proved
effective in normalizing ERG activity in three out of four children tested.
In various rat studies, it has been demonsdated that the ERG deficits
occur before retinal cell loss. Also, the deleterious effects of taurine
deficiency on cell survival and ERG acdvity have been demonsdated to be
either dependent on or synergistic with light exposure. Rapp et al. (1988)
described the interaction between light exposure and GES treatment in rats
exposed to either low- or high-intensity cyclic lighting for 10 weeks. No
outer nuclear layer (ONL) abnormalities were observed with GES treatment
and simultaneous low-intensity light exposure. However, ERG
abnormalities were observed, indicating the precedence of ERG dysfuncdon
over frank cell loss. Significant cell loss in the ONL of the retina and
greater reducdon in the a- and b-wave were observed with GES freatment
and simultaneous exposure to high-intensity light. Measurements of
taurine content demonstrated lower taurine levels with exposure to high-

intensity light, with and without GES treatment, suggesting light-induced
loss of taurine.
Other studies which used a light-cycle protocol against a dark-
maintenance protocol support the putative interaction between light
exposure and GES treatment in the deterioration of ERG activity (Quesada
et al , 1988; Cocker and Lake, 1989). GES-related ERG deficits were
significantly less in dark-maintained rats than in rats exposed to light.
Taurine levels were also higher with dark maintenance. Similar to the low-
and high-intensity light experiments previously discussed (Rapp et al,
1988), DNA measurements indicate no cell loss with GES deatment with
both light-cycle and dark maintenance protocols. In general, ERG deficits
preceded cell loss, were associated with decreased taurine levels and with
exposure to light.
Immunolocalization of taurine
The localizadon of a substance to specific cell types in histological
preparations is key to idendfying its physiological significance. Usually,
distribution to specific cell-types suggests a requirement for the substance in
the unique fijnction or funcdons of that cell type. The scientific proof of the
abundance of taurine in the redna nadirally led to great interest as to the
idendty of the specific rednal cells which contained this amino acid. Early
studies made use of radiolabeled taurine and autoradiography of retinal
dssue secdons. Through this method, the high-affinity uptake of taurine in
photoreceptor cells and pigment epithelium was demonsfrated in the rat,
mouse, guinea pig, baboon, pigeon, cat and frog (Lake et al , 1978), To a

lesser degree, taurine was also taken up in bipolar and amacrine neurons,
and in glial cells.
Taurine uptake studies readily identify cells that actively transport
taurine, but cells may slowly fransport taurine yet retain high amounts
infracellulariy. The labeling of taurine in fixed dssue through
immunochemical procedures provides a more accurate visualization of
steady-state levels of taurine. The use of andsera in specifically detecting
taurine was first indoduced with brain secdons in 1985 (Madsen et al,
1985; Ottersen et al , 1985), Taurine-like immunoreacdvity (TLI) was first
demonsfrated in insect photoreceptors in 1988 (Schafer et al, 1988) and in
the mammalian retina in 1989 (Lake and Verdone-Smith, 1989) with the cat,
rat and guinea pig. The procedure has since been used as the method of
choice in studying taurine distribution in the retina.
Various animal species have been studied, e.g., honeybee
(Apis)(SchsifeT et al , 1988), marine snail (Bulla gouldiana) (Michel et al,
2000), Australian lungfish (Neoceratodus forsteri){?ow, 1994), goldfish
(Marc, et al, 1995; Omura and Inagaki, 2000), eel (Anguilla
japonica){Omum, Inagaki, 2000), rat (Fletcher and Kalloniatis, 1996), cat
(Marc et al , 1998), cynomologous monkey (Kalloniads et al, 1996), and
human (Nag et al , 1998), Photoreceptor cells or analogous cell types
consistendy exhibit TLI, but other cell types like Muller glial cells also
exhibited reacdvity. Photoreceptor cells, specifically the rod outer segments
(ROS) of the rod cell, are responsible for visual sensation, fransforming
light signals into neuronal signals. It is particulariy noteworthy that very
few studies with human retina have been performed, and that the available

data indicate that photoreceptor ROS do not contain significant levels of
taurine (Nag et al , 1998), in condast to data from lower animal forms like
the monkey (Kalloniatis et al , 1996), Data from the eel, however,
demonstrated that ROS were not immunostained in light-adapted
conditions, but were intensely stained in dark-adapted conditions, implying
that taurine flux in the ROS is dynamic in nadire. Cleariy, the experimental
conditions under which tissue sampling is done would influence the results
of taurine analysis, at least in the ROS.
Taurine uptake in retinal tissue
In the rat and cat, the uptake of taurine has been associated with
Muller, photoreceptor and amacrine cells (reviewed in Huxtable, 1989;
Lombardini, 1991), The affinity of the uptake system for taurine is
measured by the concenfration of taurine subsdate (Km) needed to produce
half-maximal activity. The kinedcs of taurine uptake in the retina have been
studied to a fairly significant degree. It is fairly well-accepted that there are
at least two sites for taurine uptake characterized by two discernible uptake
components with different affinities for taurine, a system that seems to also
exist in brain slices and synaptosomal preparations (reviewed in Huxtable,
1989; Lombardini, 1991). Usually, there is at least a 10-fold difference in
the affinity and the maximum velocity (Vmax) exhibited by the two uptake
systems. There is evidence that high-affinity taurine uptake is specifically
associated with photoreceptor cells (Schmidt and Berson, 1978), High-
affinity uptake was observed to disappear with the loss of photoreceptor
cells in rats with hereditary age-related rednal degeneradon called RCS

(Royal College of Surgeons) rats. However, the exact nadire of uptake in
the ROS vis-a-vis the whole retina was unknown.
Salceda and colleagues have done considerable work with taurine
uptake in the retinal pigment epithelium (RPE) cells (Salceda and Saldana,
1993). Experiments with RPE cells isolated from rat eyes revealed uptake
systems functioning with Km values of less than 100 |iM, Later sdidies
revealed the existence of another uptake system with lower affinity (Km =
-400 |iM) (Salceda, 1999). Thus, it appears that, in addition to the
photoreceptor cells, RPE cells may also account for high-affinity uptake in
the retina. RPE cells are intimately associated with the ROS and form an
important component of the rednal-blood barrier which separates the blood
from the outer retinal layer. It is possible that, in the RCS rats, RPE cells
degenerate along with photoreceptor cells as they age, hence, the
disappearance of high-affinity taurine uptake with age (Schmidt and Berson,
1978). Regardless, the data suggest that taurine uptake is tighdy regulated
as it is fransported from the intravascular space to the photoreceptor cells.
The taurine transporter has been cloned using DNA from the Mabin-
Darby canine kidney (MDCK) cell line (Uchida et al , 1992, 1993), mouse
brain (Liu et al , 1992), rat brain (Smith et al , 1992), human thyroid (Jhiang
et al , 1993), human placenta (Ramamoorthy et al , 1994), and bovine
endothelial cells (Qian et al , 2000). Using the DNA of the taurine
fransporter from the human thyroid, the taurine fransporter in human RPE
cell line was also cloned through reverse franscripdon-polymerase chain
reacdon (RT-PCR) (Miyamoto et al , 1996). The DNA contained 1863 base
pairs, predicdng a protein 620 amino acids in length. The DNA was also

found to be almost identical in sequence to the taurine transporter cloned in
human thyroid and placenta. The DNA sequence has been deposited as
human RPE taurine fransporter in GenBank under accession No, U09220.
The fransporter was expressed in frog oocytes, and the fiincdon of this
fransporter was found to be sodium- and chloride-dependent, and
susceptible to inhibition by P-alanine and GABA. In similar fashion, the
taurine fransporter in mouse retina (mTAUT) was cloned (accession No.
AF020194) using the DNA of the MDCK taurine fransporter (Vinnakota et
al , 1997). The DNA was 2163 base pairs long and coded for a 621-amino
acid protein. It exhibited >93% homology with the mouse brain fransporter.
The protein was expressed in frog oocytes and its funcdon was found to be
sodium- and chloride-dependent, and sfrongly inhibited by P-alanine,
hypotaurine and guanidinoethanesulfonic acid (GES), a taurine analogue.
Surprisingly, when in situ hybridization studies were done, the highest
expression of the mTAUT mRNA was not found in the retina but in the
ciliary body, specifically in the outer layer (Vinnakota et al , 1997). The
iris, conjunctiva and the cornea showed little signal. The retina per se
showed only a modest level of silver grains. Cells in the ganglion cell layer,
the inner nuclear layer, inner plexiform layer, inner segment layer and the
RPE cells were labeled. In the rat retina, the mRNA for the taurine
fransporter was also found in the same areas (Morimura et al , 1997). The
transporter mRNA in the rat retina was also found to increase in chronic, but
not in acute, hypemafremia (Morimura et al , 1997). In condast, antibodies
directed against two cloned high-affinity fransporters (TAUTl and TAUT2)
revealed that these fransporters were expressed in photoreceptor and bipolar
10

cells in the retina (Pow et al , 2002), Regardless of the localizadon and
regulation of taurine fransporters, deletion of the mTAUT fransporter gene
results in vision loss due to severe retinal degeneration, suggesting that the
function of taurine fransporters to take up taurine into the cell is crucial in
the retina (Heller-Sdlb et al , 2002),
Taurine binding to rednal issue
Similar to taurine uptake, binding studies with retina membrane
reveal at least two sites of varying affinities for taurine (Huxtable, 1989), It
was assumed that taurine binding receptors corresponded, at least in part, to
fransporter proteins responsible for taurine uptake, Ligand affinity is
characterized by the ligand concenfration at which half-maximal binding is
observed, a value defined as the dissociadon constant (Kd), Sodium-
dependent high affinity sites have been studied in the retina of rat and
chicken, and have been found to have a Kd value of > 10 |iM (Lopez-
Colome and Pasantes-Morales, 1980; Lombardini and Prien, 1983),
Binding sites that are sodium-independent and of lower affinity have also
been demonsfrated (Salceda and Pasantes-Morales, 1982), A low-affinity
sodium-dependent binding site has also been idendfied which co-exists with
the high-affinity site (Lombardini and Prien, 1983).
Lopez-Colome et al, (1991) used osmodcally shocked membrane
preparadons from chick RPE cells in culdire for taurine binding
experiments and found a single saturable binding site (Kd = -237 nM
taurine). In fresh membranes, taurine binding increased with sodium and
with higher incubation temperature. Freezing and thawing of the membrane
11

appeared to eliminate sodium- and temperature-dependent increases in
binding. Taurine uptake was also sdidied in fresh and frozen samples, and
uptake was not compromised by the freeze-thaw procedure. Thus, there
appears to be distinction between uptake and binding sites, at least in RPE
cells. The distinction was verified by studies with GES, which is a taurine
uptake inhibitor, GES was to found to have very weak effects on taurine
binding. The data suggested that in some cases, taurine binding sites do not
correlate with taurine uptake sites.
Taurine and calcium uptake in the retina
It is well-established that taurine is a biphasic modulator of rednal
calcium uptake in in vitro experiments (reviewed in Lombardini, 1991),
These in vitro experiments, in brief, entail the incubation of tissue with
radiolabeled calcium in the presence of varying concenfrations of unlabelled
calcium. Calcium uptake increases as calcium levels are increased in the
buffer (Liebowitz et al, 1989). The exact nature of this uptake is unknown,
although it is reasonable to assume that it involves more than one
biochemical mechanism. As with any experimental model, there are
limitations to its use as a physiological system and conclusions drawn from
its use must be taken in the proper context.
At low concenfradons of calcium in the reacdon buffer (10-500 |iM),
both ATP and taurine act as stimulatory agents. However, taurine is
effecdve only in the presence of ATP. It is unclear if taurine potentiates the
effect of ATP to stimulate calcium uptake, or if taurine and ATP together
sdmulate uptake through an altogether different mechanism. The effect of
12

taurine is also dependent on the presence of bicarbonate (Pasantes-Morales
and Ordonez, 1982), hence the use of at least 25 mM bicarbonate in most
calcium uptake experiments. Half-maximal stimulation by taurine has been
estimated at anywhere from -8 to -30 mM concentradon (Liebowitz et al,
1987; Liebowitz et a l , 1988; Lombardini et al, 1989).
ATP and taurine singly are inhibitory at higher calcium
concenfradons (> 1,2 mM) (Liebowitz et al, 1989). At these
concenfrations, basal calcium uptake increases in propordon to the calcium
included in the buffer. The data suggest that in the presence of higher
calcium concenfrations, uptake is increased through additional mechanisms
that are modulated by taurine in a manner disdnct from its effects on
calcium uptake at lower calcium concenfrations.
Within the intermediate calcium concenfration range, the ATP-
dependent stimulatory effect of taurine on calcium uptake diminishes. It is
probably more accurate to say that ATP and taurine stabilize calcium uptake
in the face of changing calcium levels, sdmulatory when there is less
calcium and inhibitory when there is more (Militante and Lombardini,
2000), The phenomenon may be physiologically relevant as calcium levels
change within the cell as a mechanism for cellular signaling.
Taurine characterisdcally acts at the millimolar level which is
indiitive if not predictable given that, in nature, animal cells act to maintain
millimolar quandties of taurine infracellulariy. Taurine, thus, is considered
to be an osmotic agent, first and foremost. However, the effects of taurine
on calcium uptake in the retina are mimicked by some taurine analogues and
not by others, and the opposite effect is seen with yet others (reviewed in
13

Militante and Lombardini, 2002), The data strongly suggest that the effects
of taurine are not mediated by osmotic mechanisms. As of yet, the
mechanism behind these effects of taurine is not yet fully understood.
Taurine depletion in the rat with both GES and P-alanine did not
result in any change in the effect of taurine to stimulate calcium uptake at
low calcium concenfration (Militante and Lombardini, 2002), The data are
interesting in that one might expect some form of cellular adaptation to
occur in response to taurine depledon which would render the retina more
sensidve to the effects of taurine. However, this is not the case, at least with
this pardcular effect of taurine on the redna.
Calcium uptake and calcium binding in the retina
Care must be taken in the analysis of the effects of taurine as the
modulation of both calcium fransport and binding to membranes may
explain the specific effects of taurine on calcium uptake. In fact, eariy data
from rat retina revealed the reladve lack of effect of A23187, a divalent
ionophore, in increasing calcium uptake in the retina at high buffer
concenfradons of calcium, and thus, calcium uptake as such was atfributed
to membrane surface components rather than acdial influx of calcium
(Lombardini and Liebowitz, 1990). However, these sdidies were preceded
by sdidies that demonstrated increases in calcium uptake in PI and P2
fracdons from chick retina with A23187 freatment (Pasantes-Morales and
Quesada, 1980), All the preceding uptake assays were done at 36-37°C for
5 minutes or less to select for uptake acdvity over binding acdvity.
However, the "uptake" measured may still have included some level of
14

calcium to membranes. In whole rat retinal membranes, two putative
"uptake" sites have been described, exhibidng Km values of 35 and 2076
\XM (Lombardini, 1983), The high affinity system has been logically
identified with specific calcium transporter proteins. The lower affinity
"uptake" system may actually be membrane binding, as fransporter systems
usually work at subsfrate levels in the low micromolar range. Suffice to say,
the calcium activity measured should be attributed to more than one specific
mechanism and may not involve the acdial flux of calcium through the
plasma membrane. Thus, the term calcium accumulation is probably more
appropriate and precise than is calcium uptake, the more commonly used
term in the scientific literature.
Calcium flux and phototransduction
The biphasic effects of taurine on calcium uptake are observed both
in whole retinal samples and in isolated ROS (Liebowitz et al , 1989;
Militante and Lombardini, 1998a, b, 2000), It is important to note that the
ROS is a very specialized segment of the photoreceptor cell which is
responsible for the conversion of photonic signals to neural signals which
the brain can understand (Baylor, 1996), The whole retinal sample, on the
other hand, would contain the whole photoreceptor cell plus all the other
types of neuronal and glial cells which are found in the retina. Clearly, the
nature and the physiologic significance of the calcium uptake assayed in the
ROS and the whole rednal samples would be radically different.
The precise regulation of the inward flow of calcium through cGMP-
gated channels is crucial in the light-sensing function of the ROS (Baylor,
15

1996), In fact, the photofransducdon process and the reguladon of the
cGMP-gated calcium channel may provide a physiologic significance for
the biphasic effects of taurine on calcium uptake, in the ROS at least. In the
absence of light stimulus, the cGMP-gated channels are acdvated in the
ROS, allowing for the flow of calcium and sodium into the ROS. At this
point, taurine would be inhibitory to the uptake of calcium and presumably
is not acting on channel function. With light stimulus, cGMP levels drop
drastically, inactivating the channel. Calcium levels then drop as active
extrusion proceeds with the NaVCa^-K^ exchanger. The lower calcium
levels signal an increase in both the synthesis of cGMP and the affinity of
the channel for cGMP, resulting in the reopening of the channel, an event
necessary for continued photofransduction in the ROS, At this stage when
calcium levels have dropped, taurine may then assist in the process of
recovery by increasing calcium uptake through the cGMP-gated channels.
Taurine and protein phosphorylation
The most important funcdonal effect of taurine is most probably
associated with the reversible elecfroretinogram deficits observed after
taurine depledon in the cat, monkey, and rat (Lombardini, 1991), This
deficit is coincident with an increase in the phosphoryladon of a specific
-20 K protein in the mitochondrial fracdon of the rat retina. Conversely,
the phosphoryladon of this protein has been demonsfrated to be inhibited by
taurine in vitro (Lombardini, 1992). However, the idendty of the protein and
its link to the fiancdonal deficit observed are unknown.
16

Chelerythrine (CHT) is a benzophenanthridine alkaloid that exhibits
several biological effects including the inhibidon of protein kinase C (PKC)
(Herbert et al , 1990) and adenosinefriphosphatase (ATPase) (Cohen et al,
1978). Previously, it was demonsfrated that CHT causes sdmulation, both
in the presence and absence of taurine, of the in vitro phosphorylation of the
same -20 K mitochondrial protein in the rat retina that taurine depledon
stimulates (Lombardini, 1995; Lombardini, 1996; Lombardini et al, 1996),
Half-maximal stimuladon was estimated at approximately 37 |iM. PKC
acdvation was shown to have no effect on the phosphorylation of the -20 K
protein (Lombardini, 1993), and so it was assumed that CHT was acdng
through some mechanism other than PKC inhibition to cause the increase in
phosphorylation. Because of these data, CHT was then considered as a
possible pharmacologic tool in studying the effects of taurine on calcium
uptake.
Hypotheses
Main Interest
The mechanism of action of taurine is the primary interest of these
experiments. CHT was used as a pharmacologic tool as CHT exhibited
modulatory effects of the phosphorylation of the -20K protein in the retina,
the same protein which taurine appears to also modulate. In preliminary
experiments, CHT was found to inhibit the sdmulatory effects of taurine on
calcium uptake in the retina, and was thus used to study the mechanism of
acdon of taurine. The data presented in this thesis all revolve around a
blanket hypothesis: The stimulatory effect of taurine on ATP-dependent
17

calcium uptake in the retina is dependent on the moduladon of ATPase
acdvity, on the uptake of taurine into retina tissue, and on the activation of
calcium channels, and not on the modulation of calcium binding to
membranes. This hypothesis can be divided into several sub-hypotheses
under which all the experiments are organized.
ATPase activity in the retina
The first sub-hypothesis is: Taurine modulates ATPase activity in the
retina. The data pertinent to this sub-hypothesis were previously published
in the Biochemical Pharmacology joumal (Militante and Lombardini,
1998a), The sdmulatory effects of taurine are profoundly dependent on the
presence of ATP, One of the earliest studies which described this specific
dependence suggested that the hydrolysis of ATP was necessary in the
potentiating effects of taurine (Pasantes-Morales, 1982), implying the need
for ATPase activity. In addidon, the effects of CHT, an inhibitor of PKC
and ATPase activity, were tested, and CHT was found to antagonize the
effects of taurine. Thus, it was surmised that the ATPase acdvity of the
retinal dssue is involved in the effects of taurine. As the activity of the
different forms of ATPase is dependent on the specific ions present in the
buffer, ATPase acdvity was measured in the rednal dssue under buffer
conditions that were identical to that under which calcium uptake
experiments were conducted, ATPase activity was then studied reladve to
the effects of taurine in the retina.
18

Moduladon of taurine uptake
The second sub-hypothesis is: The specific inhibidon of taurine
uptake abolishes or attenuates the sdmulatory effects of taurine on calcium
uptake. Most of the data presented in this secdon were previously
published in the Brain Research iowmdX (Militante and Lombardini, 1999a)
and in the Journal of Pharmacology and Experimental Therapeutics
(1999b). In all of the calcium uptake experiments, the retinal dssue was
exposed to taurine in solution and, accordingly, two possibilities exist: the
dssue may actively take up taurine and taurine may bind to dssue
membrane. Similarly, calcium uptake was studied in bovine ROS, and
specifically the sdmulatory effect of ATP was characterized (Hemminki,
1975), The data suggested that the uptake of ATP into the dssue was
necessary in ATP-dependent stimulation. The same question could be asked
of taurine: Is taurine uptake necessary in taurine-stimulated calcium uptake?
Taurine is thought to bind to membranes and to alter calcium binding by
altering the membrane environment (Huxtable and Sebring, 1986), but
taurine uptake into the dssue most certainly occurs, too. Taurine uptake
kinetics were characterized in the whole retina and in isolated ROS, and the
inhibidon of uptake was studied. The inhibition of uptake was then
associated with the effects of taurine on calcium uptake.
Possible moduladon of calcium channels
The third sub-hypothesis is: The specific inhibition of calcium
channels abolishes or adenuates the stimulatory effects of taurine. Most of
the data presented in this section come from a research ardcle published in
19

the Amino Acids journal (Militante and Lombardini, 1998b), One
possibility is that the stimulatory effects of taurine on calcium uptake may
be due to the opening of calcium channels. It is strange that the effects of
taurine on calcium channels in the retina have not been studied extensively
given that the modulatory effects of taurine on ion channels in the heart are
well-documented (Satoh and Sperelakis, 1998). In the retina, the most
important calcium channel is the cGMP-gated calcium channel.
Experiments were performed to sdidy the involvement of cGMP-gated
channel activation in the effects of taurine. Modulators of the channel were
used in conjuncdon with taurine freatment in calcium uptake experiments.
Calcium uptake and calcium binding
The last subhypothesis is: Taurine does not affect calcium binding to
retinal membranes. Experiments were done to study the nadire of the
calcium uptake that taurine modulates, specifically to differendate between
calcium uptake through retinal membrane and calcium binding to retinal
membrane. Calcium uptake was measured in retinal tissue by incubating
retinal samples with radiolabeled calcium and measuring the amount that
remained after filtering the sample through a glass fiber filter. In theory, the
acdvity measured would be comprised of calcium taken up into the dssue
and calcium that bound to the membrane, and is more correcdy described as
calcium accumulation. It is possible that taurine binds to the membrane and
increases the binding of calcium to the membrane in addidon to modulating
calcium uptake. In these experiments, retinal dssue was osmodcally lysed
to denadire acdve calcium fransport. Calcium uptake was also measured
20

with and without treatment with A23187, a calcium ionophore that makes
membranes freely permeable to calcium. A23187 would allow the calcium
gradient produced by uptake systems to dissipate when the tissue samples
are washed and filtered, leaving only calcium bound to membranes. The
effects of taurine on calcium binding can thus be studied. Calcium binding
was also measured at 0°C in the presence of ATP and taurine. The use of
lower incubadon temperatures has been fraditionally used to study binding
as acdve uptake systems usually require a physiologic 37°C temperature to
funcdon adequately.
21

CHAPTER II
METHODS
Preparation of dssue samples
Adult Sprague-Dawley or Wistar rats were anesthetized with CO2 and
killed by cervical dislocation or decapitation, after which the eyes were
removed and either used immediately or frozen at -80°C to be thawed later
for experimental use (Militante and Lombardini, 1998ab), Whole retinal
dssue samples were isolated by cutting the comea open and by gently
teasing the tissue out of the eye cup into a 0.32 mM sucrose soludon while
on ice. All subsequent procedures were done on ice. The dssue was then
cenfrifliged for 15 minutes at 16,000 x g (4°C), washed in 20 mM sodium-
bicarbonate, recentrifuged as before and then washed in sodium-bicarbonate
buffer [50 mM NaHCOj, 50 mM NaCl, 50 mM KCl, 1.2 mM KH2PO4, 2
mM MgCl2, pH 7.4 (Kuo and Miki, 1980)] with CaCl2 added in the desired
concenfrations. The tissue was recentrifuged, resuspended in the
aforementioned sodium-bicarbonate buffer and gendy homogenized with a
glass-to-glass homogenizer. For some experiments, Krebs-Ringer-
bicarbonate (KRB) buffer [118 mM NaCl, 25 mM NaHCOj, 5 mM glucose,
1,2 KH2PO4, 4.7 KCl, 1.17 mM MgS04, with the desired amount of CaCl2
added] was used in place of the sodium-bicarbonate buffer (Militante and
Lombardini, 1999ab). KRB buffer was aerated with 5%I95% oxygen for 15
minutes and the pH of the soludon adjusted to 7.4 with concenfrated HCl
For the isoladon of ROS, 0,3 mM mannitol was used instead of 0,32
mM sucrose, Rednal tissue was dissected out as before and the ROS were
22

removed by vortex-mixing the tissue for 10-20 seconds, allowing the tissue
to setde, and the decanting the supernatant which contained the ROS in
suspension. The procedure was sometimes repeated with the pellet to
maximize ROS yield. The supernatant was then centrifiiged for 15 minutes
at 16,000 X g (4°C) and the pellet was then suspended in sodium-
bicarbonate buffer. The remaining tissue components were discarded. For
some experiments, KRB buffer was used instead of sodium-bicarbonate
buffer.
Protein assay
The amount of protein used was assayed using the bicinchoninic acid
(BCA) method (Militante and Lombardini, 1998ab), Briefly, standards and
samples were mixed with a soludon of BCA protein assay reagent and 4%
copper II sulfate (50:1). The mixdire was incubated in a 37°C water bath for
30 minutes and absorbance was read at 560 nm. Protein content was used to
correct and standardize all the data measured. Commonly, 100-300 fig for
the whole retinal homogenate or 50-150 |ig for the ROS was used for each
reaction.
Calcium uptake assav
The assay was performed with either the sodium-bicarbonate or KRB
buffer (Militante and Lombardini, 1998a, 1999b), Reagents such as taurine
and ATP were added to the reaction dibe in the appropriate amounts and
kept on ice until the start of the reacdon, Idendcal amounts of'^CaCl2
(400,000-500,000 dpm) were added to the dibes in the presence of varying
23

amounts of CaCl2. The reaction tubes were then preincubated in a 37°C
water bath for 2 minutes. Whole retinal homogenate or ROS samples were
added in equal amounts to start the reacdon. The reacdon was terminated
by adding 3 ml of the chilled buffer and by immediate filtering through a
glass-fiber filter in a Millipore apparadis. The filter was washed three dmes
with 3 ml chilled buffer and then counted for radioactivity in a scintillation
counter. The amount of' ^Ca' taken up by the tissue sample was
determined by subfracting the counts retained on the filter after a zero-time
incubation with the retinal preparation,
ATPase assay
The assay was adapted from commonly used procedures (Ottlecz et
al , 1993) and activity was measured in whole retinal homogenate in
sodium-bicarbonate buffer only. Briefly, equal amounts of [y-^^PjATP
(400,000-500,000 dpm) were added to each incubadon mixture and the
reaction tubes were preincubated in a 37°C water bath for 2 minutes. The
retinal dssue sample was added to start the reaction with a final total volume
of 250 |il, and the tubes were incubated for an additional 2 minutes. The
reacdon was terminated by adding 250 |il of perchloric acid, and the
radioactivity was exfracted into «-butanol/benzene (1:1) from which an
aliquot was taken and counted in a scindllation counter, Confrol dibes were
incubated for the same length of time, and radioacdvity was counted the
same way, but the perchloric acid was added prior to preincubadon and the
addidon of the rednal dssue, ATPase acdvity was determined by
subfracdng experimental values from confrol counts.
24

Taurine uptake assay
The assay used in measuring taurine uptake was modified from
previous procedures (Lake and Cocker, 1983; Quesada et al, 1984; Salceda,
1980). Briefly, the reacdon was done with whole retinal homogenate or
with isolated ROS in KRB buffer. The appropriate reagents was added to
the buffered solution in the reaction dibes with varying concenfrations of
unlabeled taurine (10 |iM - 10 mM) or CaCl2 (0-1000 |iM) present. Equal
amounts of [^Hjtaurine were added to each test dibe (-1 |iCi) and incubated
in a 37°C water bath for 2 minutes. The reacdon was initiated by the
addition of equal amounts of tissue sample (200-300 |ig for the whole
retinal homogenate and 50-150 |ig for the ROS) to each dibe in a final
volume of 250 |il. The reaction was allowed to proceed for 7 minutes
before the reaction was terminated with the addidon of 3 ml ice-cold buffer
and filfration through a Millipore apparatus. Each filter paper was then
washed three times with 3 ml ice-cold buffer and the remaining radioactivity
was measured in a scintillation counter. Blanks were determined by letting
the reaction proceed exactly as the experimental tubes except the reaction
was kept on ice.
Calcium binding assav
The calcium binding assay was adapted from previous studies
(Lombardini and Prien, 1983; Sebring and Huxtable, 1985), and
experiments were done with either lysed whole retinal homogenate or lysed
ROS in KRB buffer. Reagents such as taurine and ATP were added to the
reaction tube in the appropriate amounts and kept on ice until the end of the
25

reacdon. Idendcal amounts of ^ CaCl2 (400,000-500,000 dpm) were added
to the dibes in the presence of 10 |iM CaCl2, In preliminary binding
experiments, calcium binding increased slowly and equilibrium was reached
at around 60 minutes and maintained for an additional 60 minutes. Tissue
samples were added in equal amounts to start the reaction, and the reacdon
was terminated after 90 minutes for whole rednal samples. The reacdon
was terminated by adding 3 ml of the chilled buffer and by immediate
filtering through a glass-fiber filter in a Millipore apparatus. The filter was
washed three times with 3 ml of the chilled buffer and then counted for
radioactivity in a scintillation counter. The amount of " Ca" taken up by the
tissue sample was determined by subfracting the counts from confrol
reactions that were freated with 6N HCl,
Statisdcal analvsis
Each datum point was a measurement derived from an independent
experiment. Analyses were performed on the means ofdata pooled from
several independent experiments unless otherwise stated. Statisdcal
analyses were performed using the GraphPad Prism software (GraphPad
Software, Inc., San Diego, CA). Data were analyzed using the one-way
analysis of variance (ANOVA), Linear and non-linear regression analysis
were also performed for some of the data to esdmate various kinedc
constants like IC50 (antagonist concenfradon which produces half-maximal
inhibidon). Km (subsfrate concenfrations which produces half-maximal
activity), and Vmax (maximal activity). Post-hoc analysis was
accomplished using Dunnett's multiple comparison test, a test that
26

specifically compares experimental groups only with the confrol group.
Otherwise, the Bonferroni multiple comparison test was performed to
compare all pairs of groups. On certain experiments, unpaired t-test was
performed to determine significance between the means of specific
experimental groups.
For the taurine uptake experiments, raw data were corrected for
diffusion by graphically calculating a diffusion constant (K^) at high taurine
concenfradons. This factor was then subfracted from each measure of
taurine uptake concenfrations (K^S; S = total taurine). Non-linear
regression analysis was then performed on uptake data following the
method described by Neal and White (1978). Mathemadcal equadons for a
single and a double saturable hyperbola were used to test for a one- and a
two-component uptake system, respecdvely. Iterative optimization and best
fit analysis were done with GraphPad Prism software. Because the two
equations have different numbers of variables, the best equadon fitdng the
data was determined by the program as follows: the simpler equadon (single
saturable hyperbola) was deemed best if the corresponding sum-of-squares
were lower, but if the more complicated equadon (double saturable
hyperbola) had the lower sum-of-squares, then a F-test was calculated. The
simpler equadon was deemed best if the P value is > 0,05, and the more
complication best if the P value is < 0,05, Maximum velocity (Vmax) and
Michaelis-Menten constants (Km) were then calculated using the
appropriate equations. Linear regression analyses were also performed on
transformed data (Eadie-Hofstee plots) to provide graphic representation
and qualitadve descriptions of the uptake system or systems observed. The
27

lack of a single line fit was taken as an indication that more than one uptake
system was present.
To estimate the antagonist concenfration that produces half-maximal
inhibition (IC50), the Dixon plot methods was also employed, specifically in
cases where the use of non-linear regression analysis was ruled out by
insufficient numbers of inhibitor concentrations (Dixon and Webb, 1964).
The procedure calls for the fransformadon of enzyme or uptake activity data
into fraction of confrol values (experimental/confrol). The reciprocal of the
fraction of confrol values were then plotted against the inhibitor
concenfration and linear regression analysis was performed. The inhibitor
concenfration corresponding to the value of 2, which denotes 50%)
inhibidon, was taken as the IC50. Only data points found between the
threshold concenfrations producing minimal and maximal inhibition were
used as exfreme concenfradons producing 100%) or 0%) inhibidon can skew
the linear regression analysis of the Dixon plot, IC50 values were taken from
independent experiments and the mean ± SEM was calculated.
28
CHAPTER III
RESULTS
Preliminary data
Calcium uptake was measured in the presence of 10 \xM CaCl2 in
whole retinal homogenate by measuring the uptake of radiolabeled ' ^CaClj
in tissue that was filtered over glass fiber filters in a Millipore filfration
apparatus. The appropriate reagents were added to the reaction tube before
the reaction was started. The reaction was carried out in a 37°C water bath
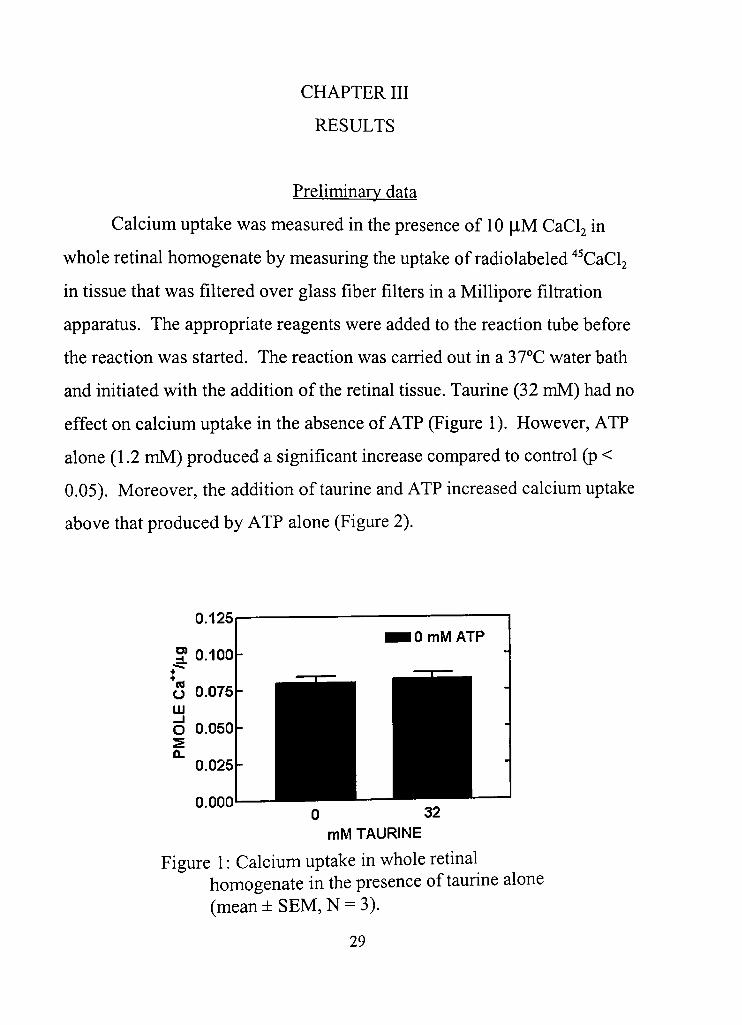
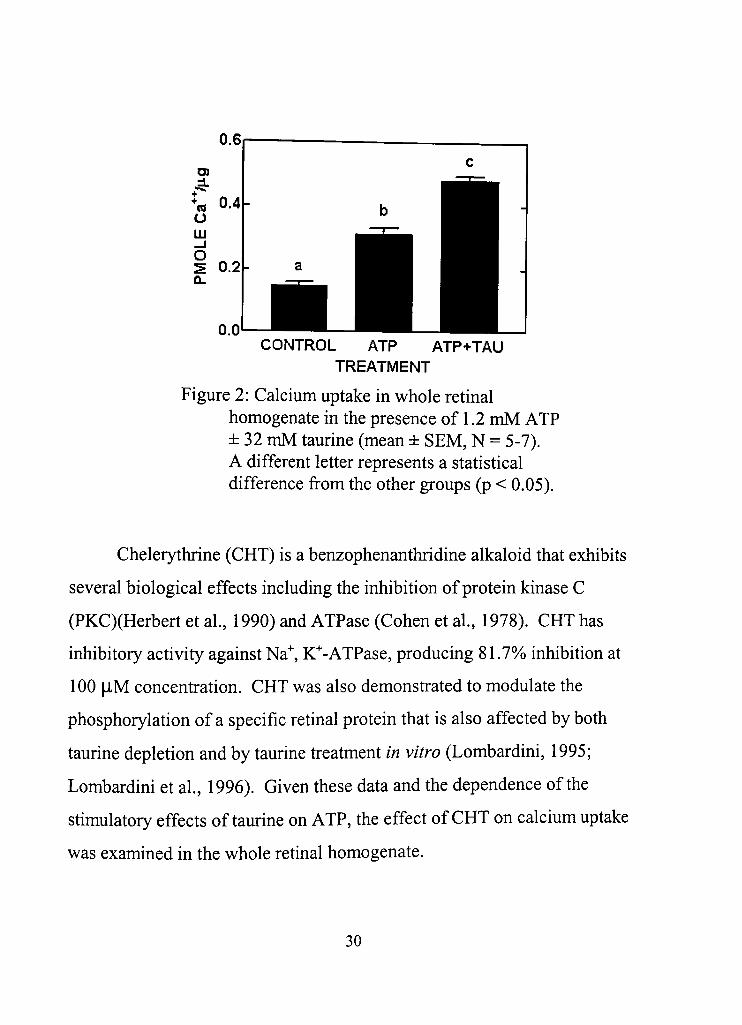
and initiated with the addition of the retinal tissue. Taurine (32 mM) had no
effect on calcium uptake in the absence of ATP (Figure 1), However, ATP
alone (1,2 mM) produced a significant increase compared to confrol (p <
0.05). Moreover, the addition of taurine and ATP increased calcium uptake
above that produced by ATP alone (Figure 2),
0,125
0.000 0 32 mM TAURINE
Figure 1: Calcium uptake in whole retinal homogenate in the presence of taurine alone (mean ± SEM, N = 3),
29
O)
O
CONTROL ATP ATP+TAU TREATMENT
Figure 2: Calcium uptake in whole retinal homogenate in the presence of 1.2 mM ATP ± 32 mM taurine (mean ± SEM, N = 5-7). A different letter represents a statisdcal difference from the other groups (p < 0,05).
Chelerythrine (CHT) is a benzophenanthridine alkaloid that exhibits
several biological effects including the inhibition of protein kinase C
(PKC)(Herbert et al , 1990) and ATPase (Cohen et al , 1978). CHT has
inhibitory activity against Na" , K" -ATPase, producing 81.7% inhibition at
100 |lM concentration. CHT was also demonsfrated to modulate the
phosphorylation of a specific retinal protein that is also affected by both
taurine depledon and by taurine freatment in vitro (Lombardini, 1995;
Lombardini et al , 1996). Given these data and the dependence of the
stimulatory effects of taurine on ATP, the effect of CHT on calcium uptake
was examined in the whole retinal homogenate.
30
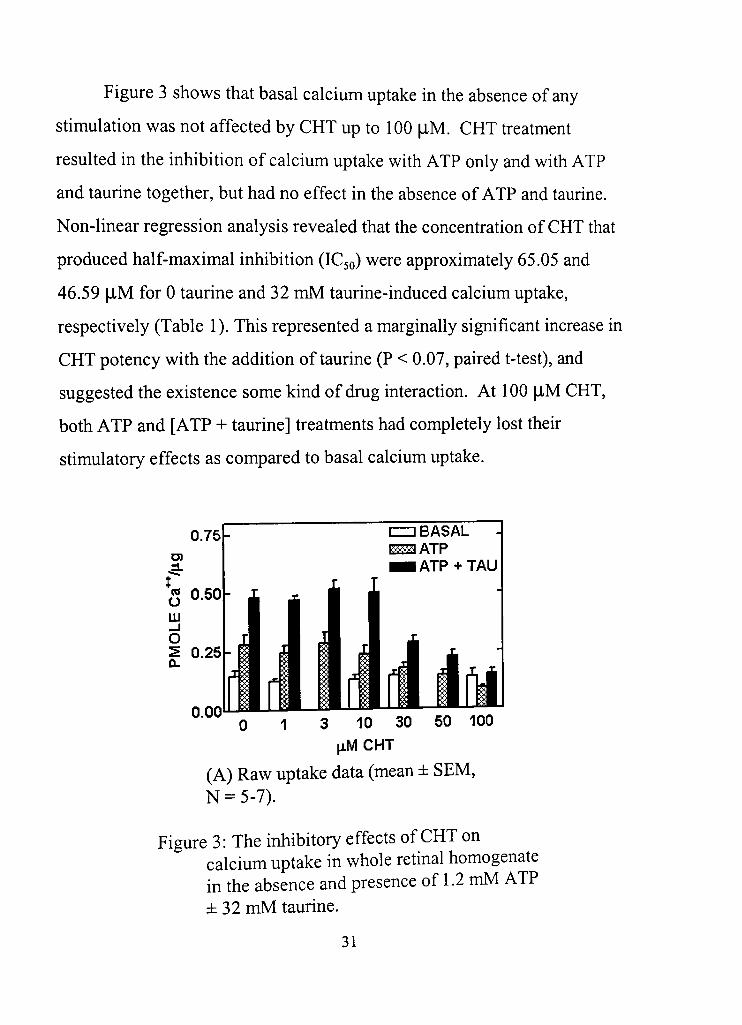
Figure 3 shows that basal calcium uptake in the absence of any
sdmulation was not affected by CHT up to 100 flM. CHT freatment
resulted in the inhibition of calcium uptake with ATP only and with ATP
and taurine together, but had no effect in the absence of ATP and taurine.
Non-linear regression analysis revealed that the concenfradon of CHT that
produced half-maximal inhibition (IC50) were approximately 65.05 and
46.59 |iM for 0 taurine and 32 mM taurine-induced calcium uptake,
respectively (Table 1). This represented a marginally significant increase in
CHT potency with the addidon of taurine (P < 0,07, paired t-test), and
suggested the existence some kind of drug interacdon. At 100 [J-M CHT,
both ATP and [ATP + taurine] treatments had completely lost their
sdmulatory effects as compared to basal calcium uptake.
0.75
+
*« 0,50 LJJ _ l
o S 0.25
° ° ° 0 1 3 10 30 50 100
jiMCHT
(A) Raw uptake data (mean ± SEM, N = 5-7),
Figure 3: The inhibitory effects of CHT on calcium uptake in whole retinal homogenate in the absence and presence of 1.2 mM ATP ± 32 mM taurine.
31
UJ ^ ^ 1 0 0
%S 76 s z =) o « 3 o 50 - 1 ^ < 2-O 25
0
I
i ^ B' i
1
1 0
'
1
1
T
1
2
'
• ATP
^
-
3 4
LOG \M CHT
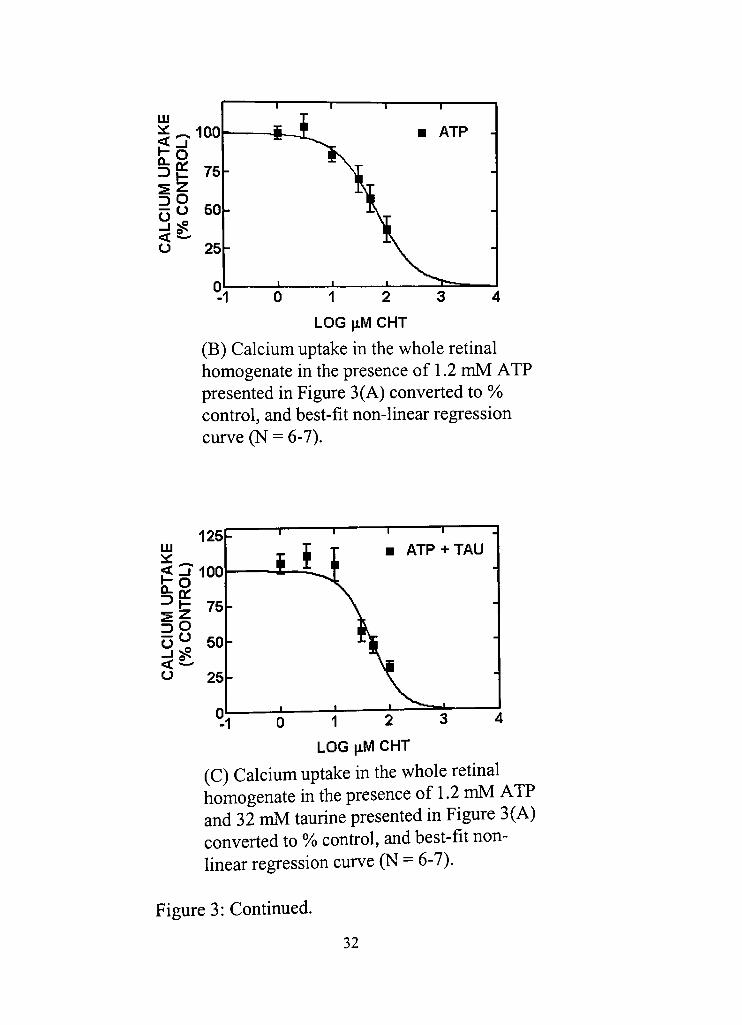
(B) Calcium uptake in the whole retinal homogenate in the presence of 1,2 mM ATP presented in Figure 3(A) converted to %> confrol, and best-fit non-linear regression curve (N = 6-7),
1 2
LOG jiM CHT
(C) Calcium uptake in the whole retinal homogenate in the presence of 1,2 mM ATP and 32 mM taurine presented in Figure 3(A) converted to Vo confrol, and best-fit nonlinear regression curve (N = 6-7),
Figure 3: Continued,
32

Table 1: Inhibitory potency (IC50) of CHT against calcium uptake and ATPase acdvity in rat retinal homogenate ±32 mM taurine, as shown in Figures 3B and C, and in Figure 8B, respecdvely. Experiments were performed in the presence of 1,2 mM ATP, and the means of esdmated IC50 values are presented with the Hill coefficient (n^) and the 95%) confidence values in parentheses. For these experiments, IC50 values were calculated from non-linear regression analysis of data converted to Vo confrol and combined from several independent experiments.
Assay
calcium uptake (N = 6-7)
ATPase activity (N = 3)
0 mM taurine
65,05 \1M CHT nH = -1.16
(50,61-83.62)
88.29 |iM CHT nH = -1.58
(75,96 - 102,6)
32 mM taurine
46,59 jlM CHT nH = -1,56
(27.98 - 77.56)
165.6 flM CHT nH = -2,13
(128,9-212,9)
33
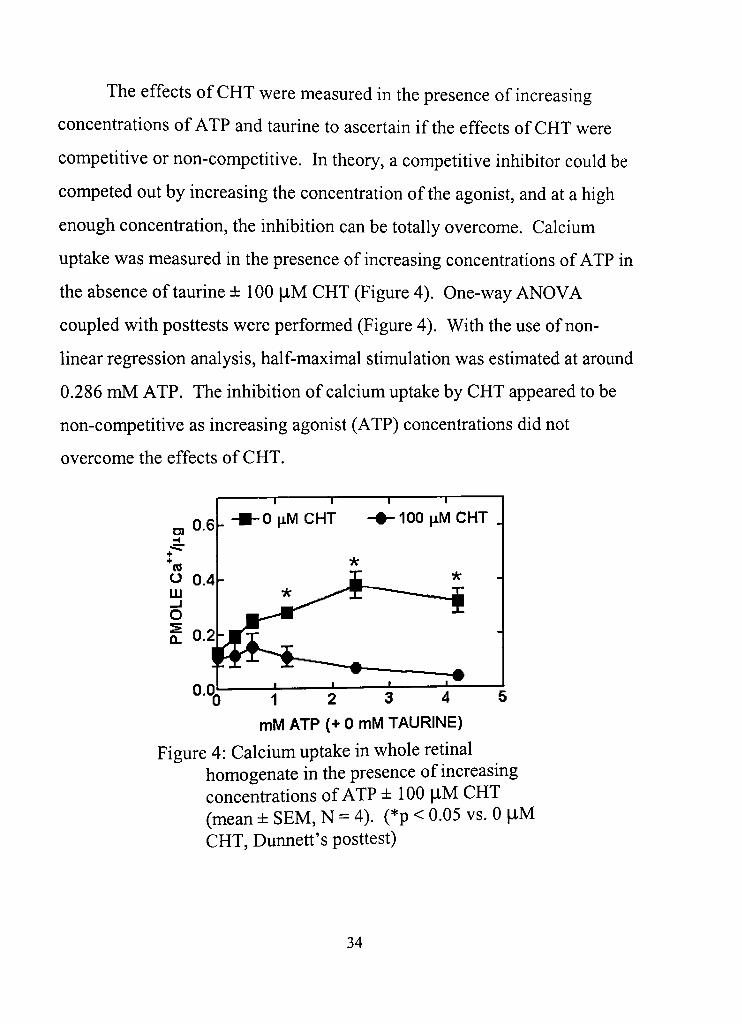
The effects of CHT were measured in the presence of increasing
concentrations of ATP and taurine to ascertain if the effects of CHT were
competitive or non-competitive. In theory, a competidve inhibitor could be
competed out by increasing the concentradon of the agonist, and at a high
enough concenfradon, the inhibition can be totally overcome. Calcium
uptake was measured in the presence of increasing concenfrations of ATP in
the absence of taurine ± 100 flM CHT (Figure 4). One-way ANOVA
coupled with posttests were performed (Figure 4), With the use of non
linear regression analysis, half-maximal stimulation was estimated at around
0.286 mM ATP. The inhibidon of calcium uptake by CHT appeared to be
non-competitive as increasing agonist (ATP) concenfradons did not
overcome the effects of CHT,
OfiM CHT T 1
-lOOjiMCHT
1 2 3 4 5
mM ATP (+ 0 mM TAURINE)
Figure 4: Calcium uptake in whole retinal homogenate in the presence of increasing concentradons of ATP ± 100 |iM CHT (mean ± SEM, N = 4). (*p < 0,05 vs, 0 |iM CHT, Dunned's posdest)
34
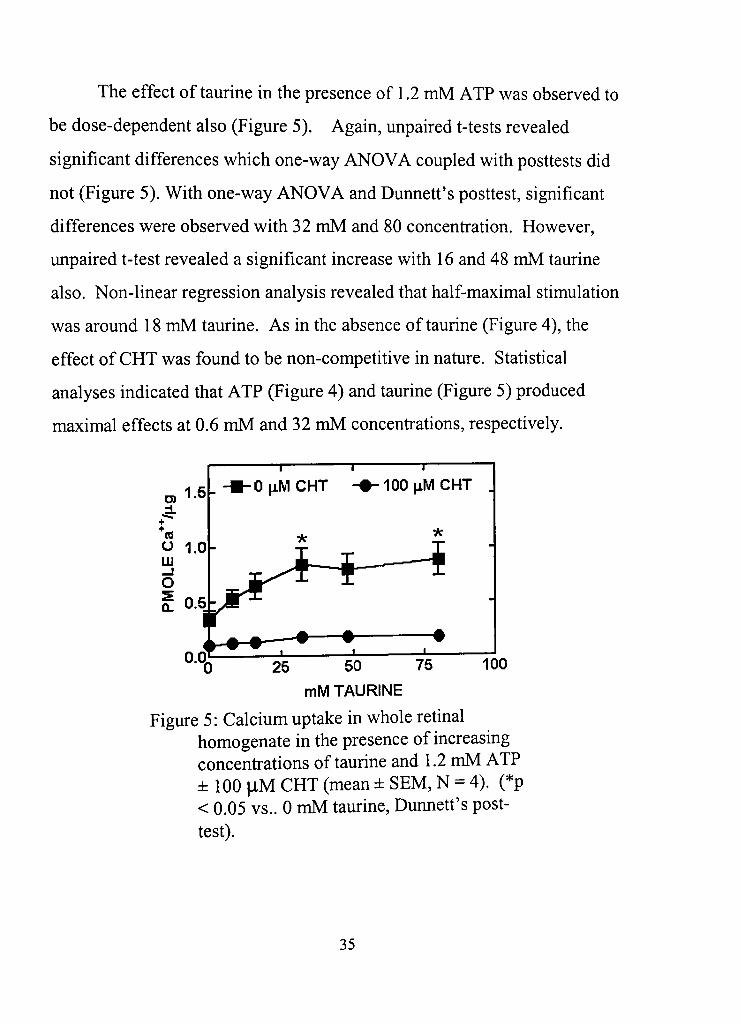
The effect of taurine in the presence of 1,2 mM ATP was observed to
be dose-dependent also (Figure 5), Again, unpaired t-tests revealed
significant differences which one-way ANOVA coupled with posttests did
not (Figure 5). With one-way ANOVA and Dunned's posttest, significant
differences were observed with 32 mM and 80 concenfration. However,
unpaired t-test revealed a significant increase with 16 and 48 mM taurine
also. Non-linear regression analysis revealed that half-maximal stimulation
was around 18 mM taurine. As in the absence of taurine (Figure 4), the
effect of CHT was found to be non-compedtive in nature. Stadstical
analyses indicated that ATP (Figure 4) and taurine (Figure 5) produced
maximal effects at 0.6 mM and 32 mM concenfrations, respecdvely.
25 50 75 100
mM TAURINE
Figure 5: Calcium uptake in whole rednal homogenate in the presence of increasing concenfradons of taurine and 1,2 mM ATP ± 100 |iM CHT (mean ± SEM, N = 4), (*p < 0,05 vs,. 0 mM taurine, Dunned's post-test).
35
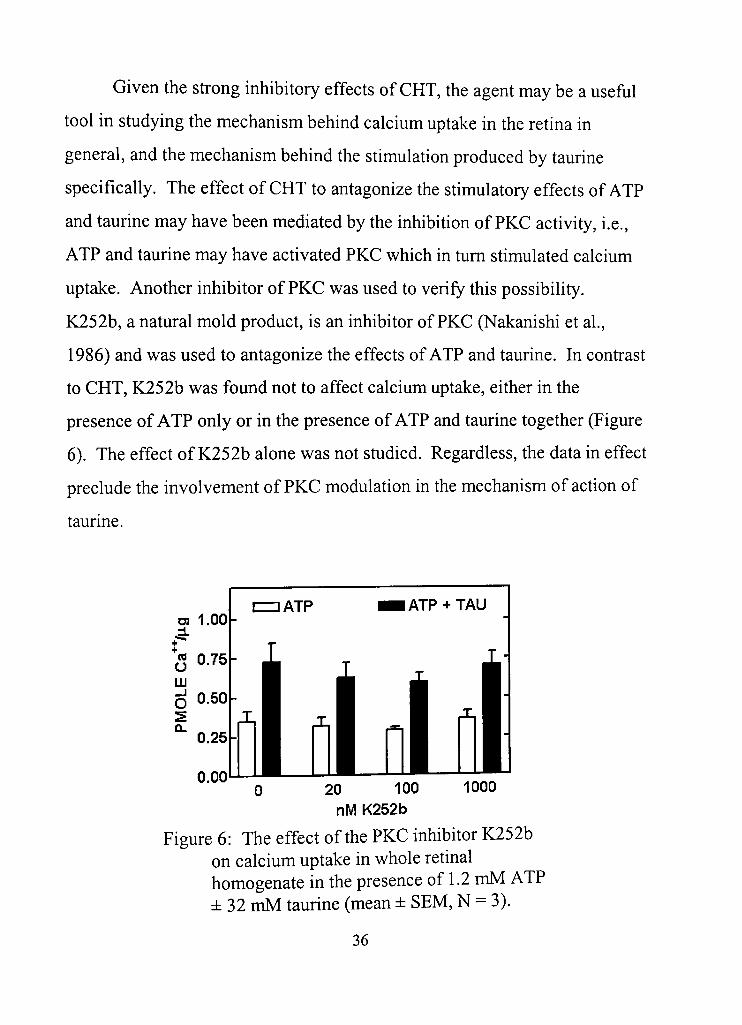
Given the sfrong inhibitory effects of CHT, the agent may be a usefiil
tool in studying the mechanism behind calcium uptake in the retina in
general, and the mechanism behind the stimulation produced by taurine
specifically. The effect of CHT to antagonize the stimulatory effects of ATP
and taurine may have been mediated by the inhibition of PKC activity, i,e.,
ATP and taurine may have activated PKC which in dim sdmulated calcium
uptake. Another inhibitor of PKC was used to verify this possibility,
K252b, a natural mold product, is an inhibitor of PKC (Nakanishi et al,
1986) and was used to antagonize the effects of ATP and taurine. In condast
to CHT, K252b was found not to affect calcium uptake, either in the
presence of ATP only or in the presence of ATP and taurine together (Figure
6), The effect of K252b alone was not studied. Regardless, the data in effect
preclude the involvement of PKC modulation in the mechanism of acdon of
taurine.
0.00 1000 0 20 100
nM K252b
Figure 6: The effect of the PKC inhibitor K252b on calcium uptake in whole retinal homogenate in the presence of 1.2 mM ATP ± 32 mM taurine (mean ± SEM, N = 3).
36
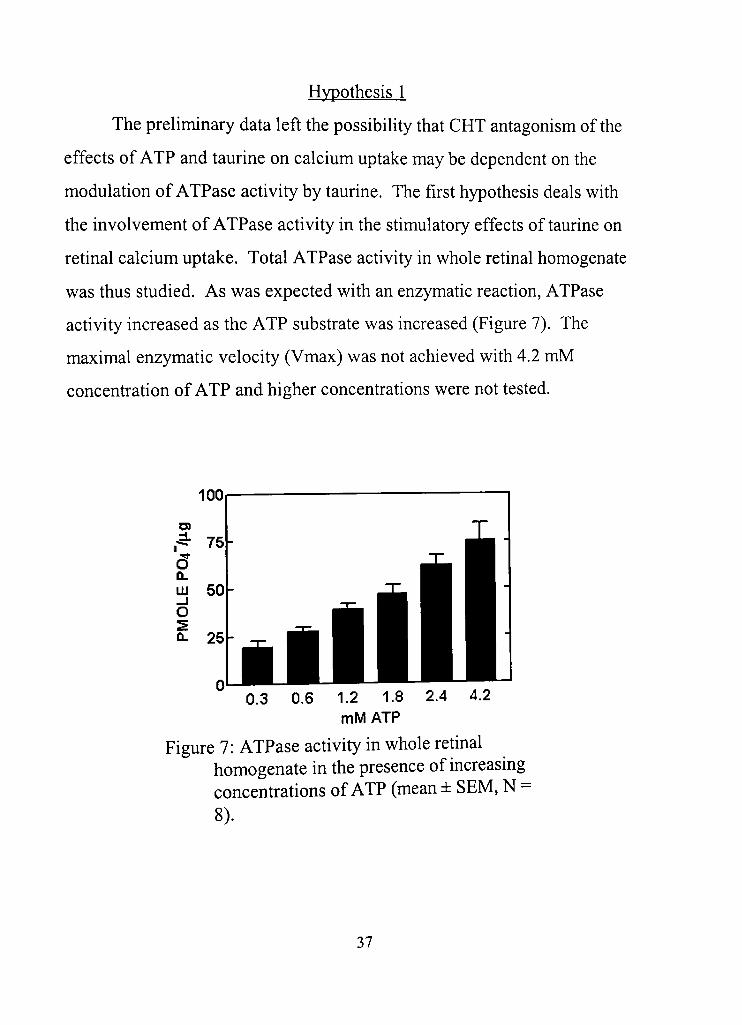
Hypothesis 1
The preliminary data left the possibility that CHT antagonism of the
effects of ATP and taurine on calcium uptake may be dependent on the
modulation of ATPase activity by taurine. The first hypothesis deals with
the involvement of ATPase activity in the stimulatory effects of taurine on
retinal calcium uptake. Total ATPase activity in whole retinal homogenate
was thus studied. As was expected with an enzymatic reaction, ATPase
acdvity increased as the ATP substrate was increased (Figure 7). The
maximal enzymatic velocity (Vmax) was not achieved with 4.2 mM
concenfradon of ATP and higher concentrations were not tested.
100
UJ 50
Q. 25
0,3 0,6 1.2 1.8 2.4 4.2 mMATP
Figure 7: ATPase activity in whole retinal homogenate in the presence of increasing concenfrations of ATP (mean ± SEM, N = 8),
37

The effect of increasing concenfradons of CHT was sdidied in the
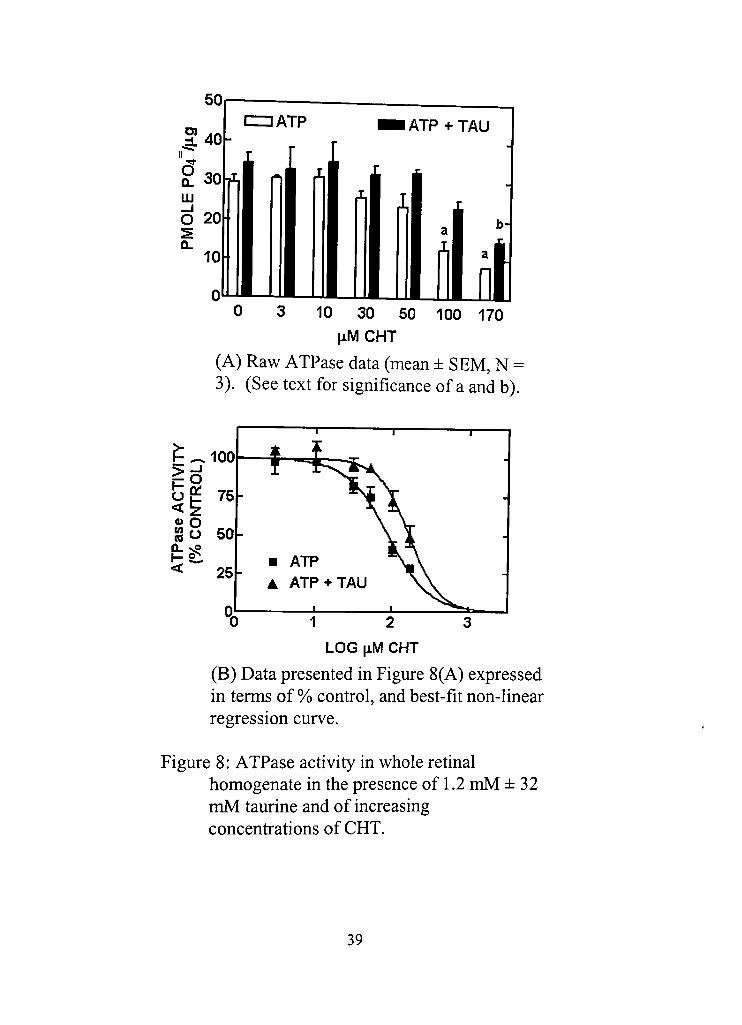
presence of 1.2 mM ATP ± 32 mM taurine. It was found that 100 |iM
concenfration or greater of CHT decreased ATPase activity significandy in
the absence of taurine (Figure 8 A, a: p < 0.05 vs. corresponding control at 0
jiM CHT). In the presence of taurine, only 170 |J-M CHT produced
significant inhibidon (Figure 8A, b: p < 0.05 vs. corresponding control at 0
|iM CHT), Taurine in the presence of ATP produced no effect on ATPase
activity (ATP vs, ATP+TAU, 0 \XU CHT), suggesting that the modulation of
ATPase activity may not be a mechanism of action for taurine. However, the
inhibitory effect of CHT was found to be slighdy diminished with taurine
(Figure 8A, a: p < 0.05 vs. corresponding ATP+TAU ) which suggested an
interacdon between the drugs. Non-linear regression revealed that in the
absence of taurine, IC50 was esdmated at around 88.29 |iM CHT
concenfradon, while in the presence of 32 mM taurine the value was
produced was significandy higher at around 165.6 |iM CHT (Table 1, page
33).
38
3 10 30 50 100 170 [iMCHT
(A) Raw ATPase data (mean ± SEM, N = 3), (See text for significance of a and b).
1 2 3 LOG \iM CHT
(B) Data presented in Figure 8(A) expressed in terms of % confrol, and best-fit non-linear regression curve.
Figure 8: ATPase activity in whole rednal homogenate in the presence of 1,2 mM ± 32 mM taurine and of increasing concenfrations of CHT.
39

Characterization of the total ATPase activity was performed in an
effort to identify the specific ATPase involved. ATPase activity was sdidied
using pharmacological agents instead of alteradon of the ionic composidon
of the buffer, as is usually done (Berman et al, 1977), as this would result in
acdvity that might not be present in the original buffer condidons under
which calcium uptake had been measured. Ouabain and thapsigargin were
used to inhibit Na^ K^-ATPase and SERCA (sarcoplasmic/endoplasmic
redculum calcium ATPase), respecdvely (Kijima et al, 1991; Lytton et al,
1991;Ottclezetal, 1993).
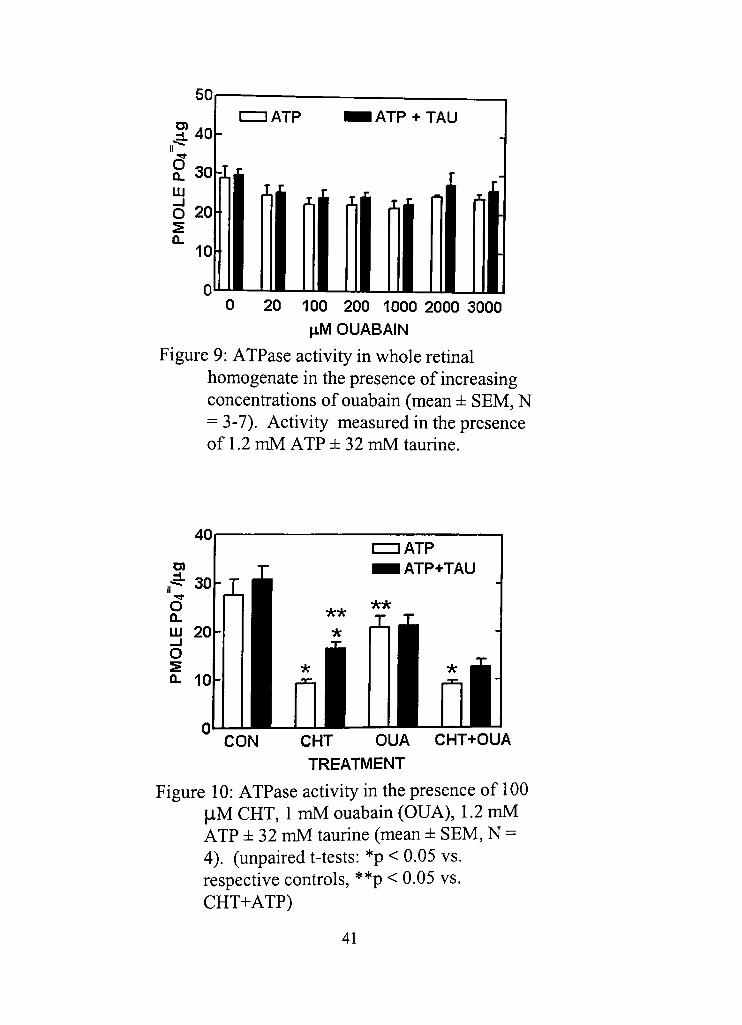
Na^, K^-ATPase was first studied because of the known effects of
CHT to inhibit its acdvity. Ouabain had been used to inhibit Na" ,
K" -ATPase in the rat retina using a bicarbonate-free system and was found to
cause half-maximal inhibition at around 20 jiM (Otdecz et al , 1993). In our
system, ouabain at concenfrations ranging from 20-3000 \XM did not cause a
decrease in total ATPase activity in the presence of 1.2 mM ATP, with or
without 32 mM taurine present (Figure 9). CHT (in the absence of taurine)
consistently produced inhibidon greater than 1 mM ouabain (Figure 10) and
so is shown, for the first time, to inhibit rednal ATPase activity other than
ouabain-sensidve Na^, K^-ATPase. Because all of the components of the
whole cell were present in the sample, it can be assumed that the ouabain-
sensidve Na^, K^-ATPase enzyme was present. Ouabain caused a slight but
insignificant decrease in total ATPase, suggesting that the enzyme may be
minimally funcdonal, if at all, under the ionic condidons of the bicarbonate
buffer used.
40
I ATP I ATP + TAU
h i r nl ^i ^i ^i x nl
10
0 20 100 200 1000 2000 3000 ^M OUABAIN
Figure 9: ATPase acdvity in whole retinal homogenate in the presence of increasing concenfradons of ouabain (mean ± SEM, N = 3-7). Activity measured in the presence of 1,2 mM ATP ±32 mM taurine.
CON CHT OUA CHT+OUA TREATMENT
Figure 10: ATPase activity in the presence of 100 |iM CHT, 1 mM ouabain (OUA), 1.2 mM ATP ± 32 mM taurine (mean ± SEM, N = 4). (unpaired t-tests: *p < 0.05 vs, respective confrols, **p < 0,05 vs, CHT+ATP)
41
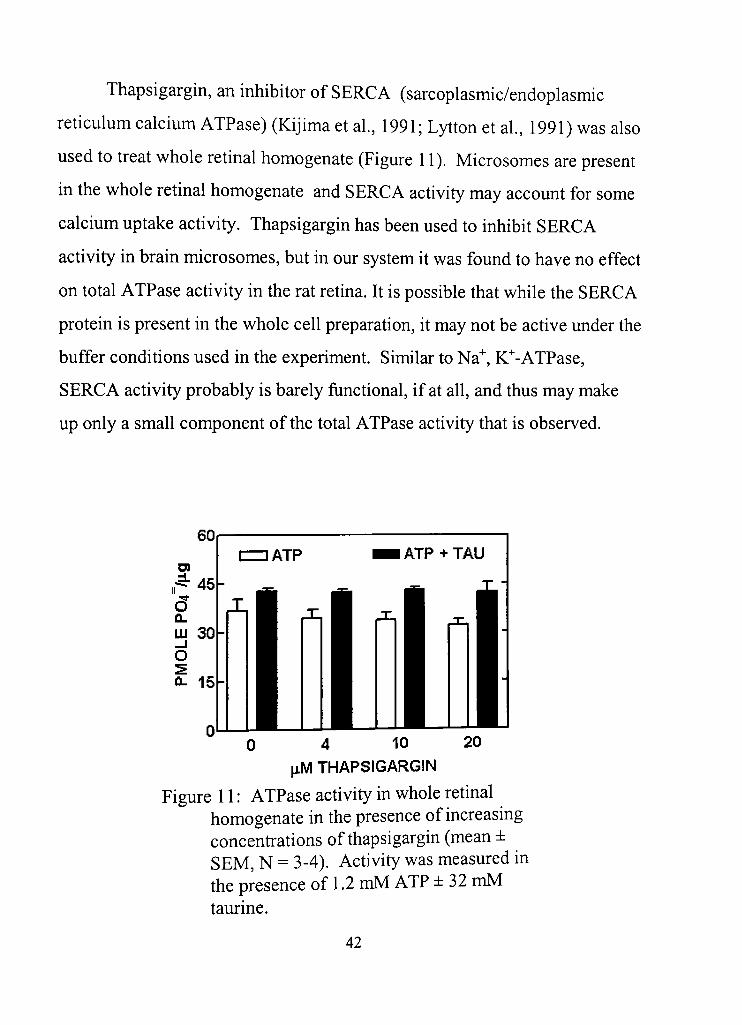
Thapsigargin, an inhibitor of SERCA (sarcoplasmic/endoplasmic
redculum calcium ATPase) (Kijima et al , 1991; Lytton et al, 1991) was also
used to treat whole retinal homogenate (Figure 11). Microsomes are present
in the whole rednal homogenate and SERCA activity may account for some
calcium uptake acdvity, Thapsigargin has been used to inhibit SERCA
activity in brain microsomes, but in our system it was found to have no effect
on total ATPase acdvity in the rat retina. It is possible that while the SERCA
protein is present in the whole cell preparation, it may not be active under the
buffer condidons used in the experiment. Similar to Na" , K""-ATPase,
SERCA activity probably is barely funcdonal, if at all, and thus may make
up only a small component of the total ATPase activity that is observed.
0 4 10 jiM THAPSIGARGIN
Figure 11: ATPase activity in whole retinal homogenate in the presence of increasing concentrations of thapsigargin (mean ± SEM, N = 3-4). Acdvity was measured in the presence of 1.2 mM ATP ± 32 mM taurine.
42
Hvpothesis 2
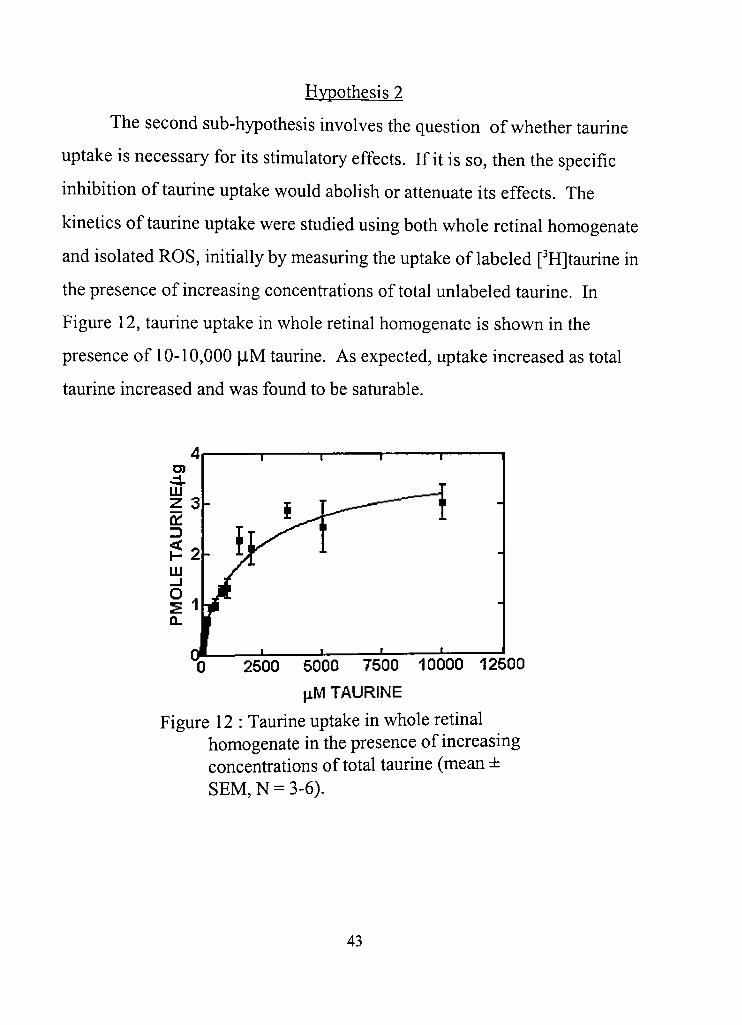
The second sub-hypothesis involves the question of whether taurine
uptake is necessary for its stimulatory effects. If it is so, then the specific
inhibition of taurine uptake would abolish or attenuate its effects. The
kinedcs of taurine uptake were sdidied using both whole retinal homogenate
and isolated ROS, initially by measuring the uptake of labeled [^Hjtaurine in
the presence of increasing concenfradons of total unlabeled taurine. In
Figure 12, taurine uptake in whole retinal homogenate is shown in the
presence of 10-10,000 |iM taurine. As expected, uptake increased as total
taurine increased and was found to be saturable.
2500 5000 7500 10000 12500 \i.M TAURINE
Figure 12 : Taurine uptake in whole retinal homogenate in the presence of increasing concenfrations of total taurine (mean ± SEM, N = 3-6),
43
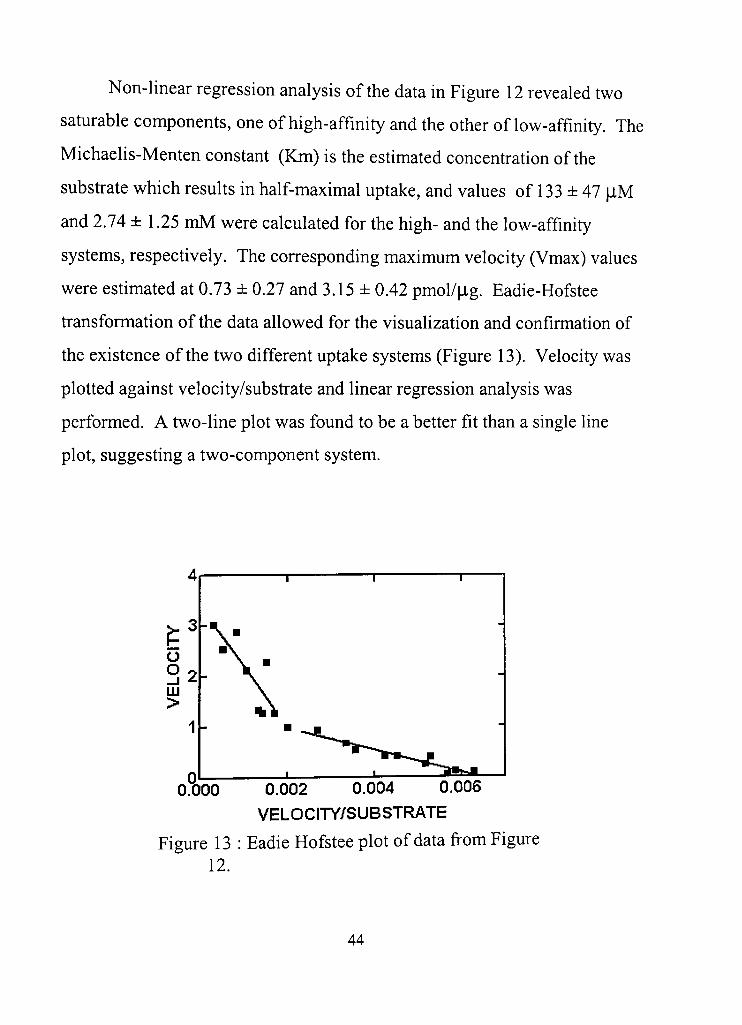
Non-linear regression analysis of the data in Figure 12 revealed two
saturable components, one of high-affinity and the other of low-affinity. The
Michaelis-Menten constant (Km) is the estimated concenfradon of the
subsfrate which results in half-maximal uptake, and values of 133 ± 47 |J,M
and 2.74 ± 1.25 mM were calculated for the high- and the low-affinity
systems, respectively. The corresponding maximum velocity (Vmax) values
were estimated at 0,73 ± 0.27 and 3.15 ± 0.42 pmol/|ig. Eadie-Hofstee
fransformation of the data allowed for the visualization and confirmation of
the existence of the two different uptake systems (Figure 13). Velocity was
plotted against velocity/subsfrate and linear regression analysis was
performed. A two-line plot was found to be a better fit than a single line
plot, suggesting a two-component system.
o o _ l m >
O.'OOO 0,002 0,004 0.006
VELOCITY/SUBSTRATE
Figure 13 : Eadie Hofstee plot ofdata from Figure 12,
44
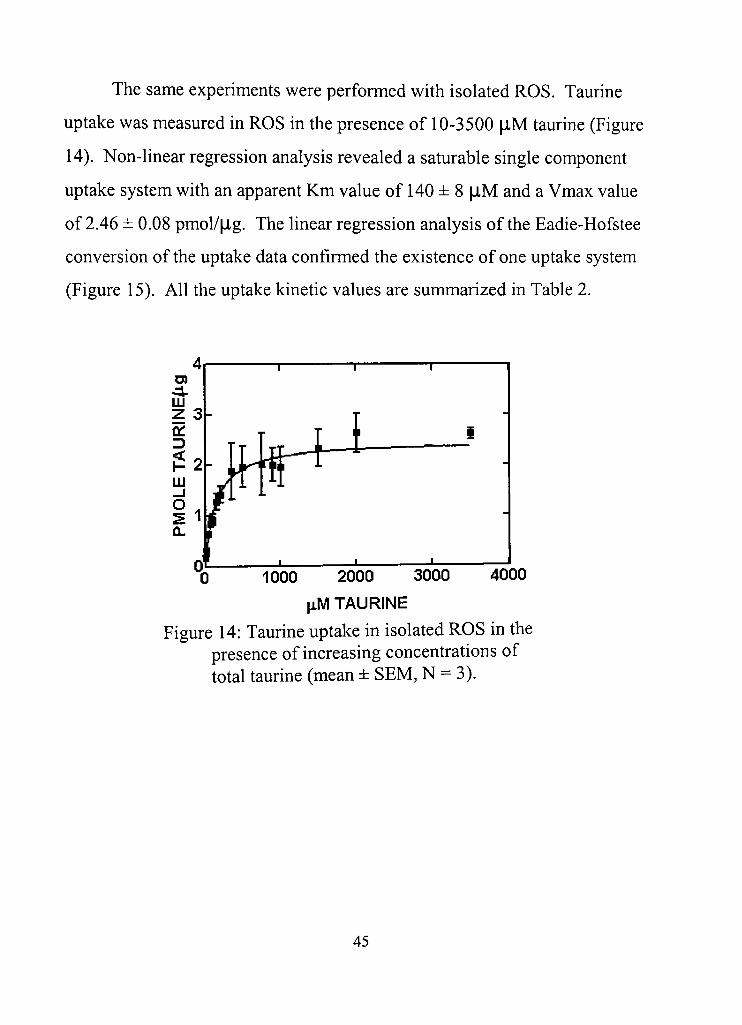
The same experiments were performed with isolated ROS, Taurine
uptake was measured in ROS in the presence of 10-3500 jlM taurine (Figure
14), Non-linear regression analysis revealed a saturable single component
uptake system with an apparent Km value of 140 ± 8 ^iM and a Vmax value
of 2,46 ± 0.08 pmol/|ig. The linear regression analysis of the Eadie-Hofstee
conversion of the uptake data confirmed the existence of one uptake system
(Figure 15). All the uptake kinetic values are summarized in Table 2.
"0 1000 2000 3000 4000 [iM TAURINE
Figure 14: Taurine uptake in isolated ROS in the presence of increasing concenfrations of total taurine (mean ± SEM, N = 3),
45
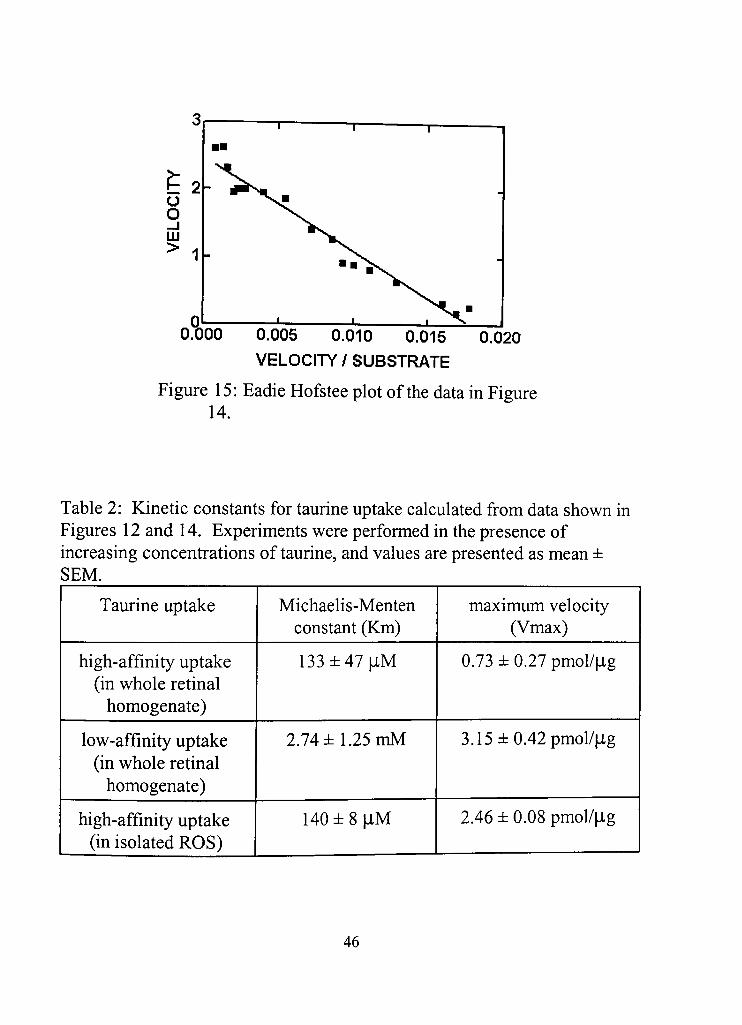
0.005 0,010 0.015
VELOCITY / SUBSTRATE 0,020
Figure 15: Eadie Hofstee plot of the data in Figure 14,
Table 2: Kinetic constants for taurine uptake calculated from data shown in Figures 12 and 14. Experiments were performed in the presence of increasing concenfrations of taurine, and values are presented as mean ± SEM,
Taurine uptake
high-affinity uptake (in whole retinal
homogenate)
low-affinity uptake (in whole retinal
homogenate)
high-affinity uptake (in isolated ROS)
Michaelis-Menten constant (Km)
133±47|iM
2.74 ± 1,25 mM
140±8| iM
maximum velocity (Vmax)
0.73 ± 0.27 pmol/|ag
3.15 ± 0.42 pmol/|ig
2.46 ± 0.08 pmol/|ig
46

The Km values in Table 2 were used to determine the taurine
concentrations to use in the inhibition experiments. Taurine uptake was
measured in the presence of 50 [IM and 1,5 mM taurine to study high- and
low-affinity uptake, respectively, CHT effecdvely inhibited both uptake
systems in whole retina (Figures 16A and 17A) and high-affinity uptake in
the ROS (Figure 18A), CHT may, thus, inhibit the effects of taurine on
calcium uptake by blocking taurine uptake. Half-maximal inhibition
constants were calculated by normalizing the data to % control which would
correspond to uptake in the absence of CHT (Figures 16B, 17B and 18B).
The means normalized values were calculated and non-linear regression
analyses were done to esdmate the concenfradon of CHT that produces half-
maximal inhibition (IC50).
47
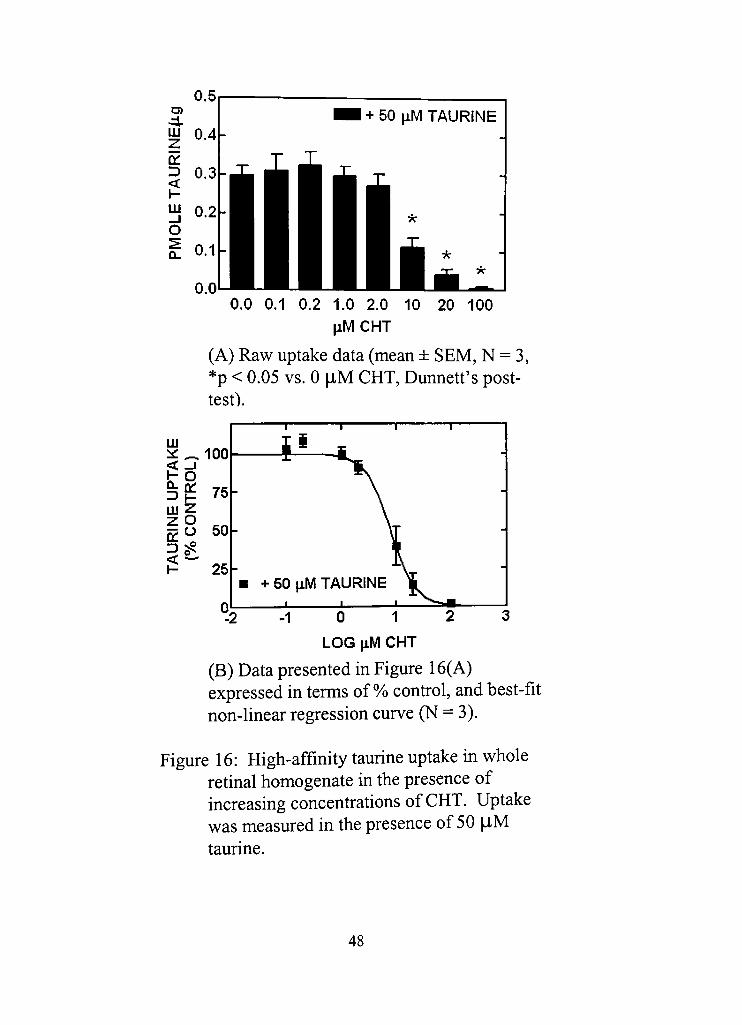
D) •4. LU Z
a: D < 1 -UJ _ l o Q-
0.5
04
0.3
0.2
0.1
0,0 0.0 0,1 0.2 1,0 2.0 10 20 100
jiMCHT
(A) Raw uptake data (mean ± SEM, N = 3, *p < 0,05 vs. 0 iM CHT, Dunned's post-test).
UJ ii:_100
a a: ^ I - 75-
50-
25-
i
+ 50 ^M TAURINE
-1 0 1 2 LOG jiM CHT
(B) Data presented in Figure 16(A) expressed in terms of %) confrol, and best-fit non-linear regression curve (N = 3),
Figure 16: High-affinity taurine uptake in whole retinal homogenate in the presence of increasing concenfrations of CHT, Uptake was measured in the presence of 50 |iM taurine.
48
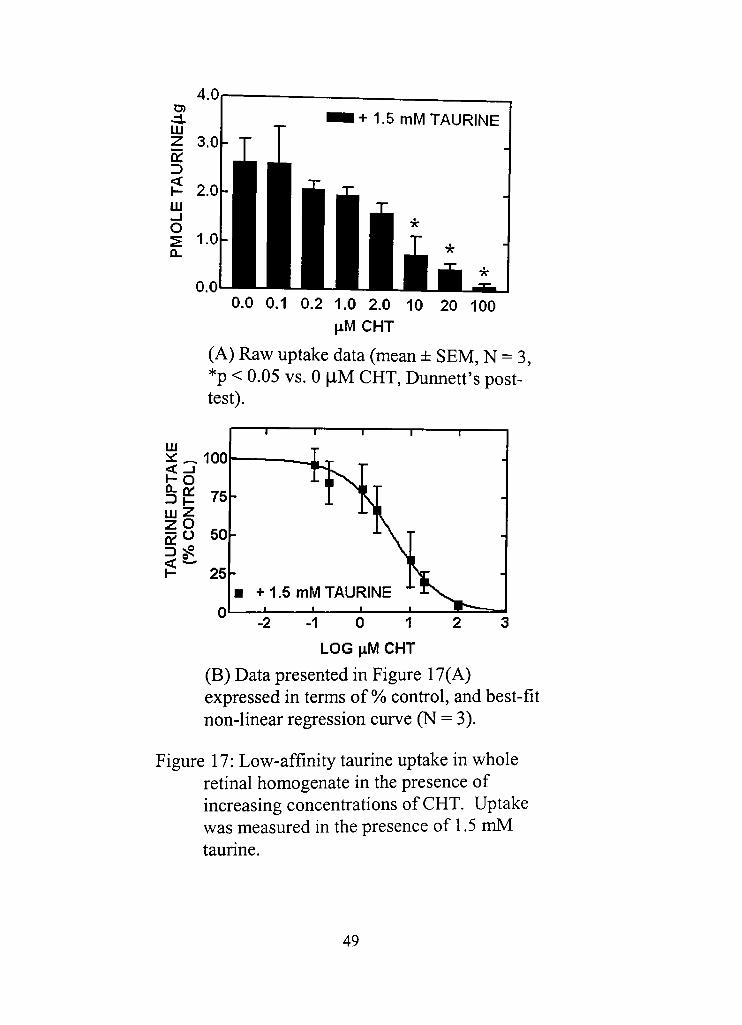
0,0 0,1 0,2 1.0 2.0 10 20 100 HMCHT
(A) Raw uptake data (mean ± SEM, N = 3, *p < 0.05 vs. 0 [XM CHT, Dunned's post-test).
- 2 - 1 0 1 2 3 LOG \iM CHT
(B) Data presented in Figure 17(A) expressed in terms of % confrol, and best-fit non-linear regression curve (N = 3).
Figure 17: Low-affinity taurine uptake in whole retinal homogenate in the presence of increasing concenfrations of CHT, Uptake was measured in the presence of 1.5 mM taurine.
49
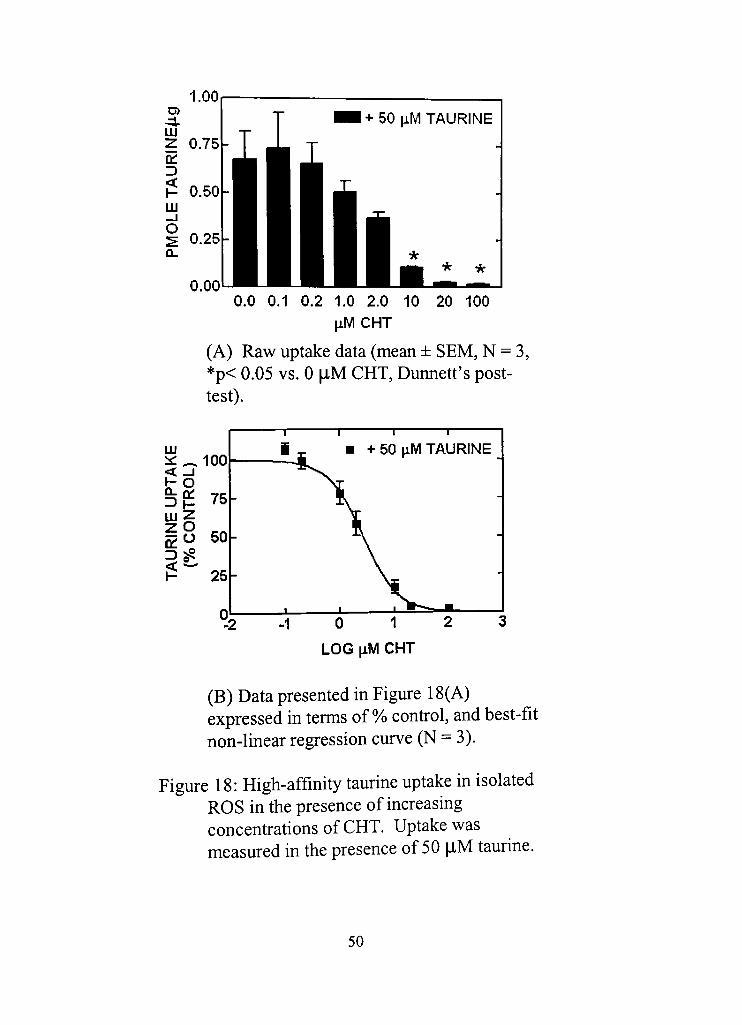
0,0 0.1 0.2 1.0 2.0 10 20 100 jiMCHT
(A) Raw uptake data (mean ± SEM, N = 3, *p< 0.05 vs. 0 |iM CHT, Dunned's post-test).
LU is: _ 100 < -1 t -o ^f^ 75 U j Z z o ^ g O 50 = ) ^ < ^ ^^ P 25
n
1
-
I
— 1 1 1
• + 50 iiM TAURINE 1 _
( ,
• » » _
" 2 - 1 0 1 2 3
LOG \LM CHT
(B) Data presented in Figure 18(A) expressed in terms of %> confrol, and best-fit non-linear regression curve (N = 3),
Figure 18: High-affinity taurine uptake in isolated ROS in the presence of increasing concenfradons of CHT. Uptake was measured in the presence of 50 [iM taurine.
50

Table 3: Inhibitory potency (IC50) of CHT against high- and low-affinity uptake in whole rednal homogenate, and of high-affinity taurine uptake in isolated ROS, as shown in Figures 16B and 17B, and in Figure 18B, respectively. The means of estimated IC50 values are presented with the Hill coefficient (nH) and 95% confidence values in parentheses. For these experiments, IC50 values were calculated from non-linear regression analysis of data converted to %> confrol and combined from several independent experiments.
Final taurine concenfration
50| iM (for high-affinity uptake)
1,5 mM (for low-affinity uptake)
Whole rednal homogenate
7,92 flM nH = -1.88
(6.16-10.20)
4.37 |iM nH = -0.84
(3.17-6.03)
ROS
2.66 [IM nH = -1.35
(2.04 - 3.47)
51
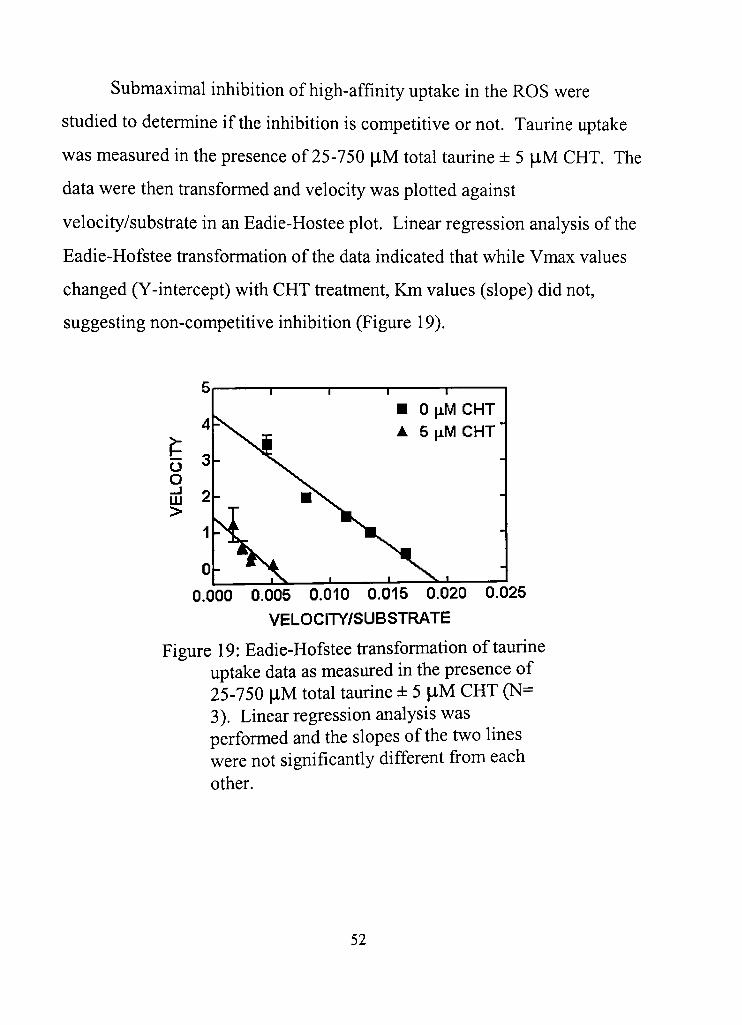
Submaximal inhibition of high-affinity uptake in the ROS were
studied to determine if the inhibition is competitive or not. Taurine uptake
was measured in the presence of 25-750 \xM total taurine ± 5 jiM CHT. The
data were then transformed and velocity was plotted against
velocity/subsfrate in an Eadie-Hostee plot. Linear regression analysis of the
Eadie-Hofstee fransformation of the data indicated that while Vmax values
changed (Y-intercept) with CHT freatment. Km values (slope) did not,
suggesting non-competidve inhibition (Figure 19).
• 0 [iM CHT A 5 iiM CHT
0,000 0.005 0.010 0.015 0.020 0.025
VELOCITY/SUBSTRATE
Figure 19: Eadie-Hofstee fransformadon of taurine uptake data as measured in the presence of 25-750 |iM total taurine ± 5 |lM CHT (N= 3). Linear regression analysis was performed and the slopes of the two lines were not significandy different from each other.
52
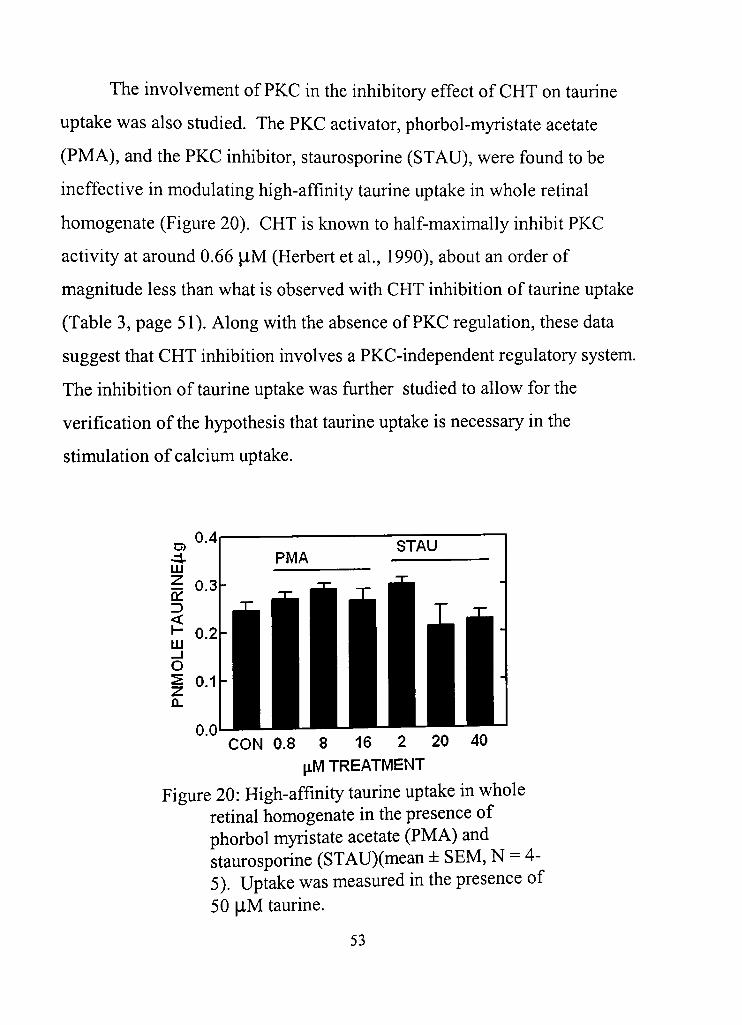
The involvement of PKC in the inhibitory effect of CHT on taurine
uptake was also sdidied. The PKC activator, phorbol-myristate acetate
(PMA), and the PKC inhibitor, staurosporine (STAU), were found to be
ineffective in modulating high-affinity taurine uptake in whole retinal
homogenate (Figure 20). CHT is known to half-maximally inhibit PKC
acdvity at around 0.66 [XM (Herbert et al , 1990), about an order of
magnitude less than what is observed with CHT inhibition of taurine uptake
(Table 3, page 51). Along with the absence of PKC reguladon, these data
suggest that CHT inhibition involves a PKC-independent regulatory system.
The inhibition of taurine uptake was further studied to allow for the
verificadon of the hypothesis that taurine uptake is necessary in the
stimulation of calcium uptake.
CON 0.8 8 16 2 20 40 [iM TREATMENT
Figure 20: High-affinity taurine uptake in whole retinal homogenate in the presence of phorbol myristate acetate (PMA) and staurosporine (STAU)(mean ± SEM, N = 4-5). Uptake was measured in the presence of 50 l-lM taurine.
53
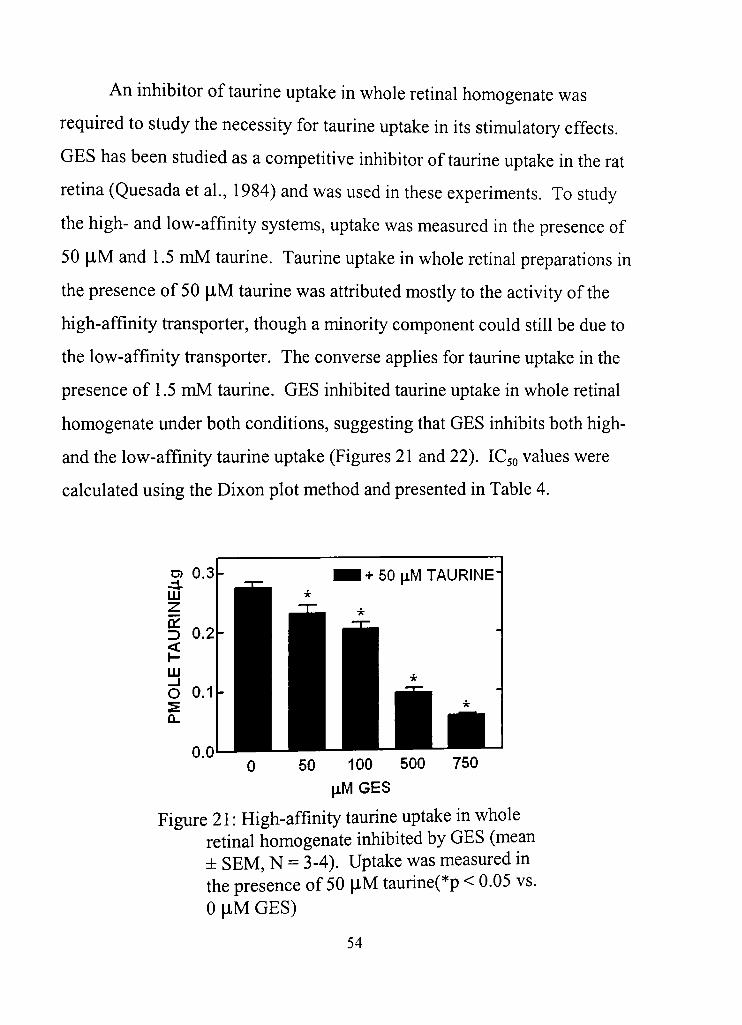
An inhibitor of taurine uptake in whole retinal homogenate was
required to study the necessity for taurine uptake in its stimulatory effects.
GES has been sdidied as a compeddve inhibitor of taurine uptake in the rat
retina (Quesada et al , 1984) and was used in these experiments. To study
the high- and low-affinity systems, uptake was measured in the presence of
50 |iM and 1.5 mM taurine. Taurine uptake in whole retinal preparations in
the presence of 50 |iM taurine was atfributed mostly to the activity of the
high-affinity fransporter, though a minority component could still be due to
the low-affinity fransporter. The converse applies for taurine uptake in the
presence of 1.5 mM taurine. GES inhibited taurine uptake in whole retinal
homogenate under both condidons, suggesting that GES inhibits both high-
and the low-affinity taurine uptake (Figures 21 and 22). IC50 values were
calculated using the Dixon plot method and presented in Table 4.
50 100 500
[iMGES
Figure 21: High-affinity taurine uptake in whole retinal homogenate inhibited by GES (mean ± SEM, N = 3-4). Uptake was measured in the presence of 50 flM taurine(*p < 0,05 vs. 0 |iM GES)
54
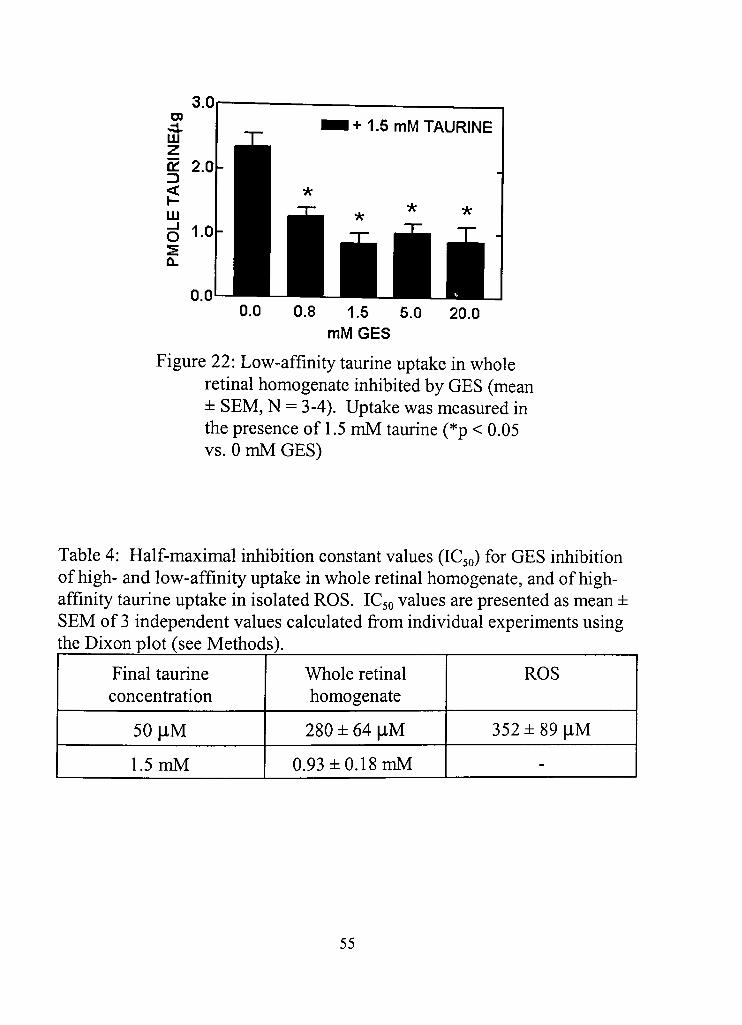
+ 1,5 mM TAURINE
0.8 1,5 5.0 mMGES
20.0
Figure 22: Low-affinity taurine uptake in whole retinal homogenate inhibited by GES (mean ± SEM, N = 3-4), Uptake was measured in the presence of 1.5 mM taurine (*p < 0.05 vs. 0 mM GES)
Table 4: Half-maximal inhibidon constant values (IC50) for GES inhibidon of high- and low-affinity uptake in whole retinal homogenate, and of high-affinity taurine uptake in isolated ROS. IC50 values are presented as mean ± SEM of 3 independent values calculated from individual experiments using the Dixon plot (see Methods).
Final taurine concenfration
50| iM
1.5 mM
Whole retinal homogenate
280 ± 64 |iM
0.93 ± 0.18 mM
ROS
352±89| iM
-
55
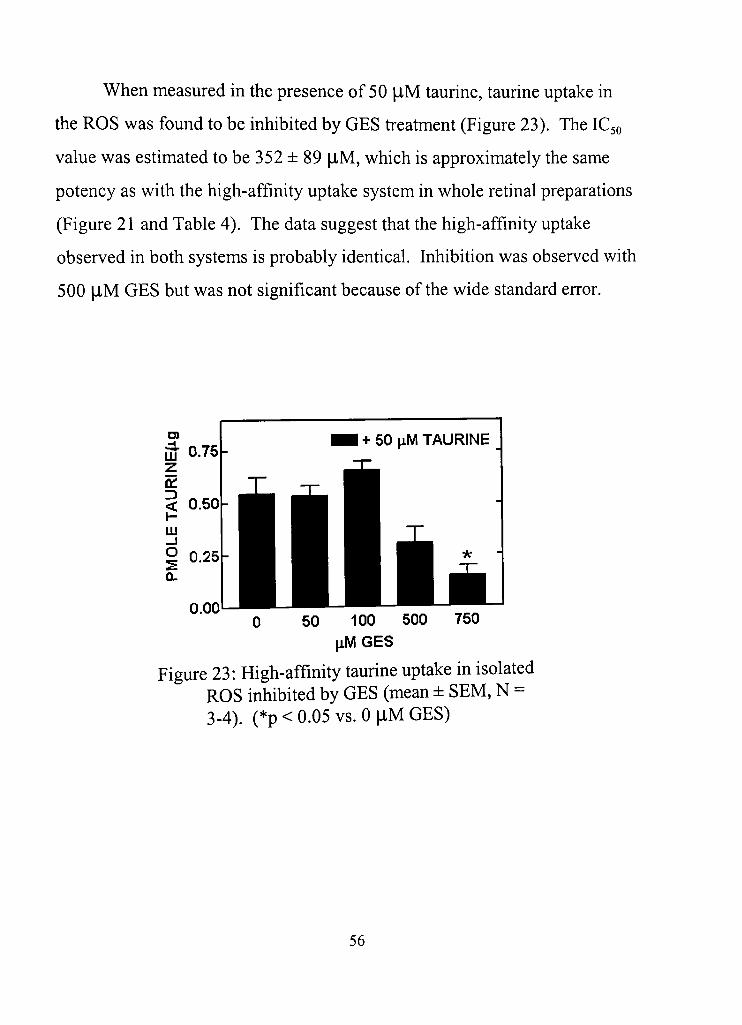
When measured in the presence of 50 jiM taurine, taurine uptake in
the ROS was found to be inhibited by GES freatment (Figure 23). The IC50
value was estimated to be 352 ± 89 |iM, which is approximately the same
potency as with the high-affinity uptake system in whole retinal preparations
(Figure 21 and Table 4). The data suggest that the high-affinity uptake
observed in both systems is probably identical. Inhibition was observed with
500 [IM GES but was not significant because of the wide standard error.
iJ 0.75 z Of < 0.50 liJ _i
I 0.25
0.00
1 + 50 ^M TAURINE _
0 50 100 500 750 tiMGES
Figure 23: High-affinity taurine uptake in isolated ROS inhibited by GES (mean ± SEM, N = 3-4). (*p < 0.05 vs. 0 jlM GES)
56

The effect of taurine on calcium uptake was sdidied in the presence of
GES to inhibit taurine uptake. Calcium uptake was measured in whole
retinal homogenate (Figure 24) and isolated ROS (Figure 25) in the presence
of increasing concenfradons of taurine and 32 mM GES. Given the
calculated IC50 values for GES (Table 4), the uptake of taurine should have
been inhibited significandy by the GES freatment. However, 32 mM GES
did not reverse the effects of taurine, suggesting that taurine uptake is not
essential in the stimulation of calcium uptake by taurine, either in the whole
retinal homogenate or in isolated ROS.
mM TAURINE (+ 1.2 mM ATP)
Figure 24: Calcium uptake in whole retinal homogenate in the presence of GES, a taurine uptake inhibitor (mean ± SEM, N = 3). (*p < 0.05 vs. 0 mM taurine)
57
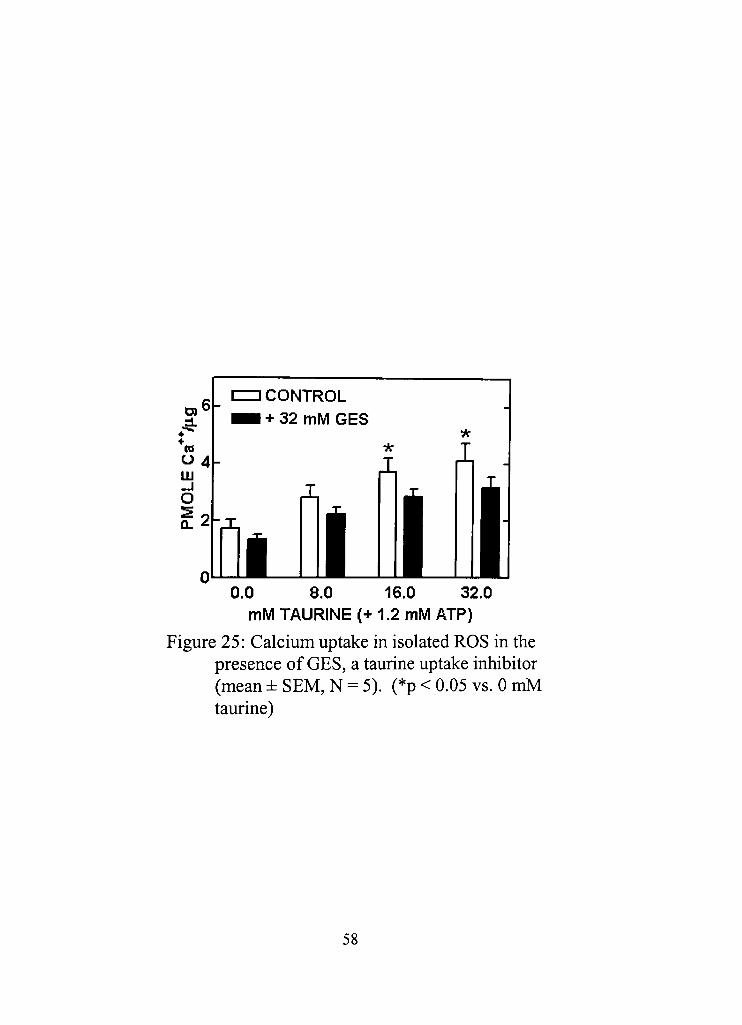
0,0 8,0 16,0 32.0 mM TAURINE (+ 1.2 mM ATP)
Figure 25: Calcium uptake in isolated ROS in the presence of GES, a taurine uptake inhibitor (mean ± SEM, N = 5). (*p < 0.05 vs. 0 mM taurine)
58
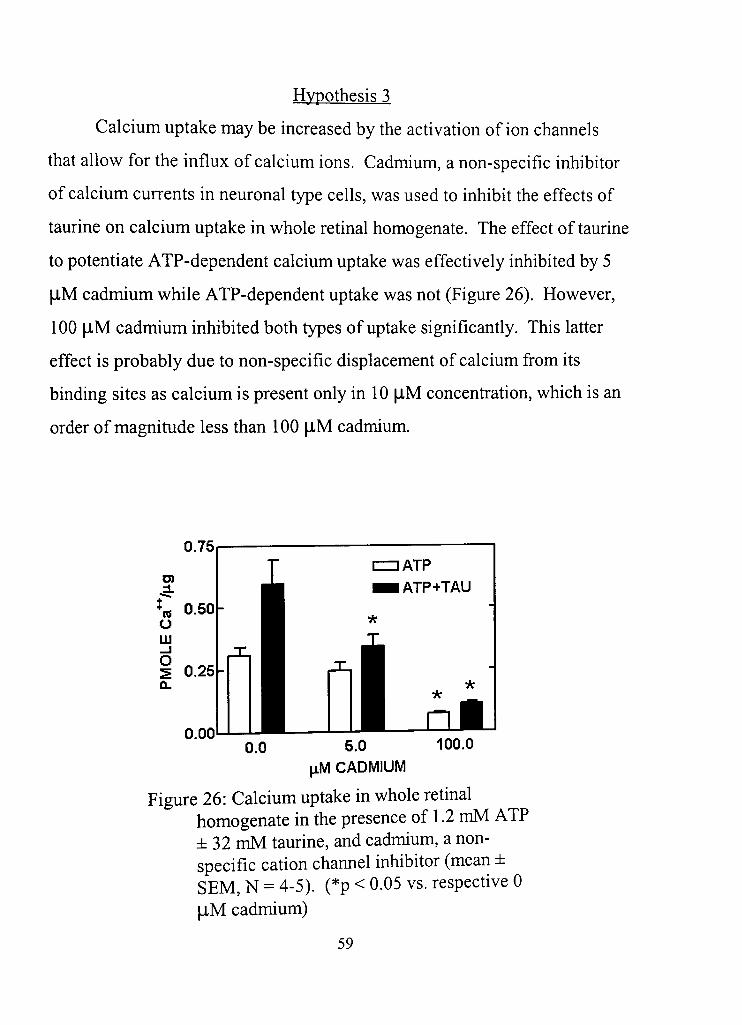
Hvpothesis 3
Calcium uptake may be increased by the acdvadon of ion channels
that allow for the influx of calcium ions. Cadmium, a non-specific inhibitor
of calcium currents in neuronal type cells, was used to inhibit the effects of
taurine on calcium uptake in whole retinal homogenate. The effect of taurine
to potentiate ATP-dependent calcium uptake was effectively inhibited by 5
|lM cadmium while ATP-dependent uptake was not (Figure 26). However,
100 |lM cadmium inhibited both types of uptake significantly. This latter
effect is probably due to non-specific displacement of calcium from its
binding sites as calcium is present only in 10 flM concenfration, which is an
order of magnitude less than 100 |iM cadmium.
0.75
0,00 0,0 100.0 5,0
^M CADMIUM
Figure 26: Calcium uptake in whole rednal homogenate in the presence of 1,2 mM ATP ±32 mM taurine, and cadmium, a nonspecific cation channel inhibitor (mean ± SEM, N = 4-5). (*p < 0-05 vs. respective 0 |iM cadmium)
59
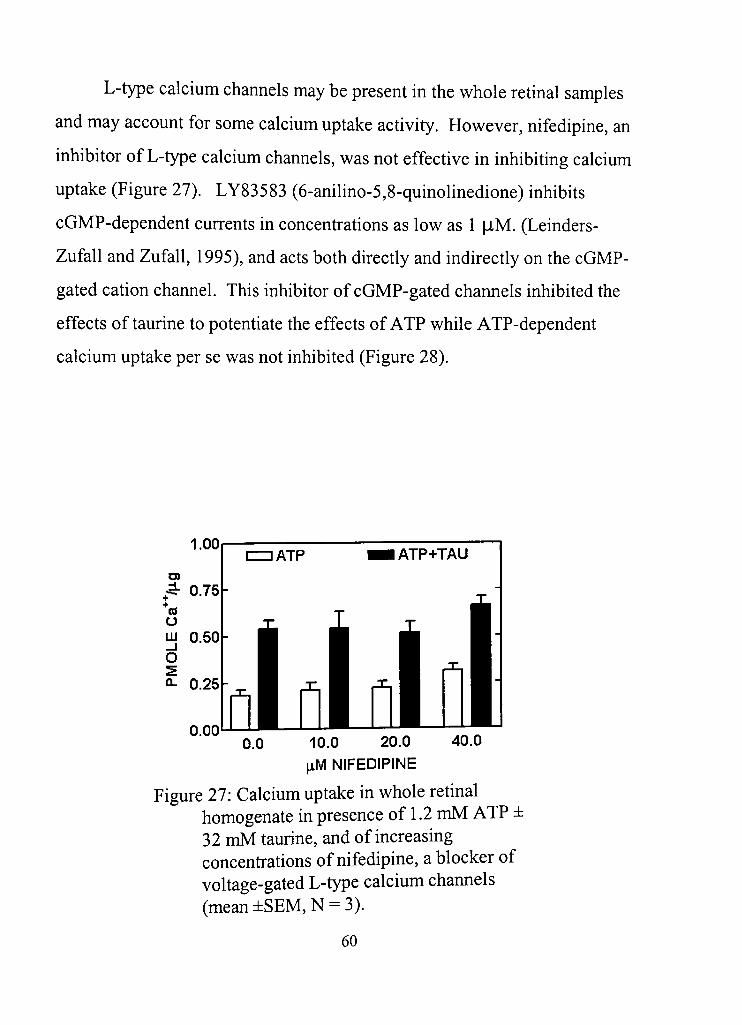
L-type calcium channels may be present in the whole retinal samples
and may account for some calcium uptake activity. However, nifedipine, an
inhibitor of L-type calcium channels, was not effecdve in inhibiting calcium
uptake (Figure 27). LY83583 (6-anilino-5,8-quinolinedione) inhibits
cGMP-dependent currents in concenfradons as low as 1 |iM. (Leinders-
Zufall and Zufall, 1995), and acts both directly and indirectly on the cGMP-
gated cation channel. This inhibitor of cGMP-gated channels inhibited the
effects of taurine to potentiate the effects of ATP while ATP-dependent
calcium uptake per se was not inhibited (Figure 28).
1.00
0.00 0.0 40,0 10,0 20,0
|xM NIFEDIPINE
Figure 27: Calcium uptake in whole rednal homogenate in presence of 1,2 mM ATP ± 32 mM taurine, and of increasing concenfrations of nifedipine, a blocker of voltage-gated L-type calcium channels (mean±SEM,N = 3).
60
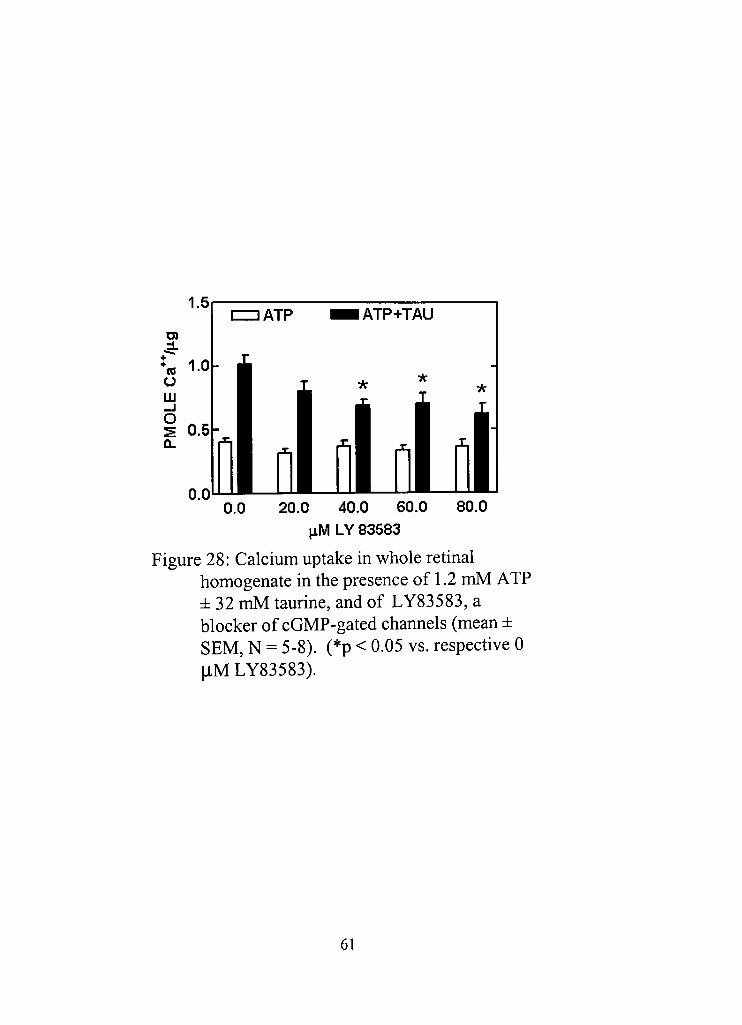
Ol
+
o UJ _J
o
1.5 IZIDATP IATP+TAU
1 oi i oi oil ^m nm r
0,0 80.0 20.0 40,0 60,0 IiM LY 83583
Figure 28: Calcium uptake in whole retinal homogenate in the presence of 1,2 mM ATP ± 32 mM taurine, and of LY83583, a blocker of cGMP-gated channels (mean ± SEM, N = 5-8). (*p < 0,05 vs, respective 0 [iU LY83583).
61
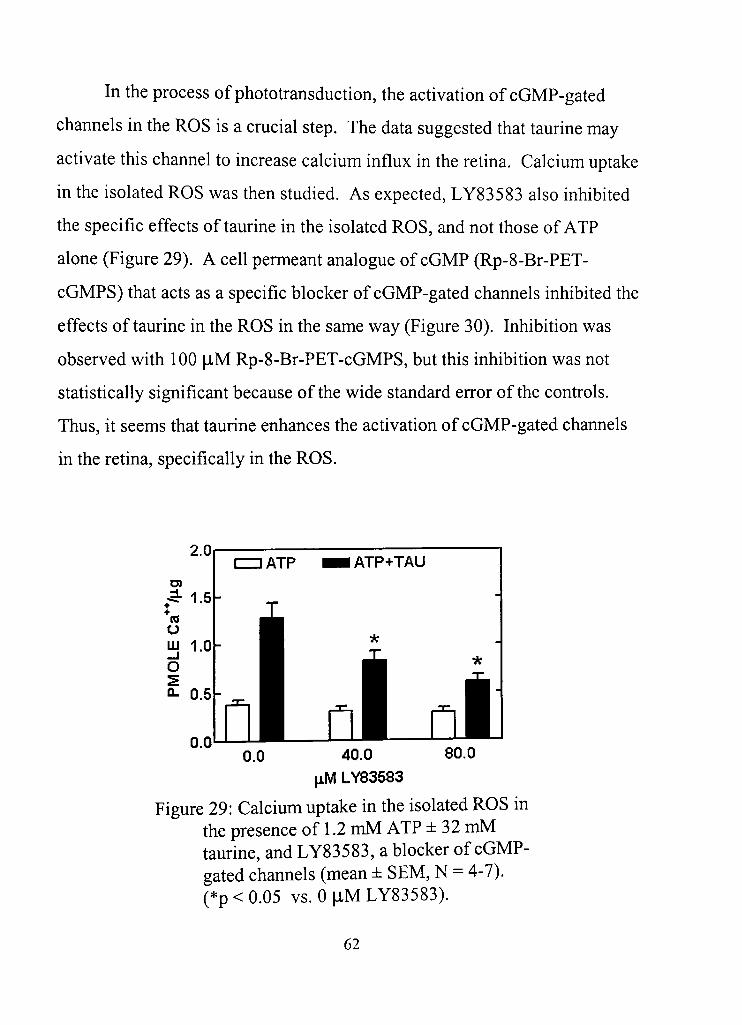
In the process of photofransducdon, the acdvadon of cGMP-gated
channels in the ROS is a crucial step. The data suggested that taurine may
activate this channel to increase calcium influx in the redna. Calcium uptake
in the isolated ROS was then studied. As expected, LY83583 also inhibited
the specific effects of taurine in the isolated ROS, and not those of ATP
alone (Figure 29), A cell permeant analogue of cGMP (Rp-8-Br-PET-
cGMPS) that acts as a specific blocker of cGMP-gated channels inhibited the
effects of taurine in the ROS in the same way (Figure 30), Inhibition was
observed with 100 |lM Rp-8-Br-PET-cGMPS, but this inhibition was not
statistically significant because of the wide standard error of the controls.
Thus, it seems that taurine enhances the activadon of cGMP-gated channels
in the retina, specifically in the ROS.
0.0 80.0 40,0 JiM LY83583
Figure 29: Calcium uptake in the isolated ROS in the presence of 1,2 mM ATP ± 32 mM taurine, and LY83583, a blocker of cGMP-gated channels (mean ± SEM, N = 4-7). (*p<0,05 vs. Op,MLY83583).
62
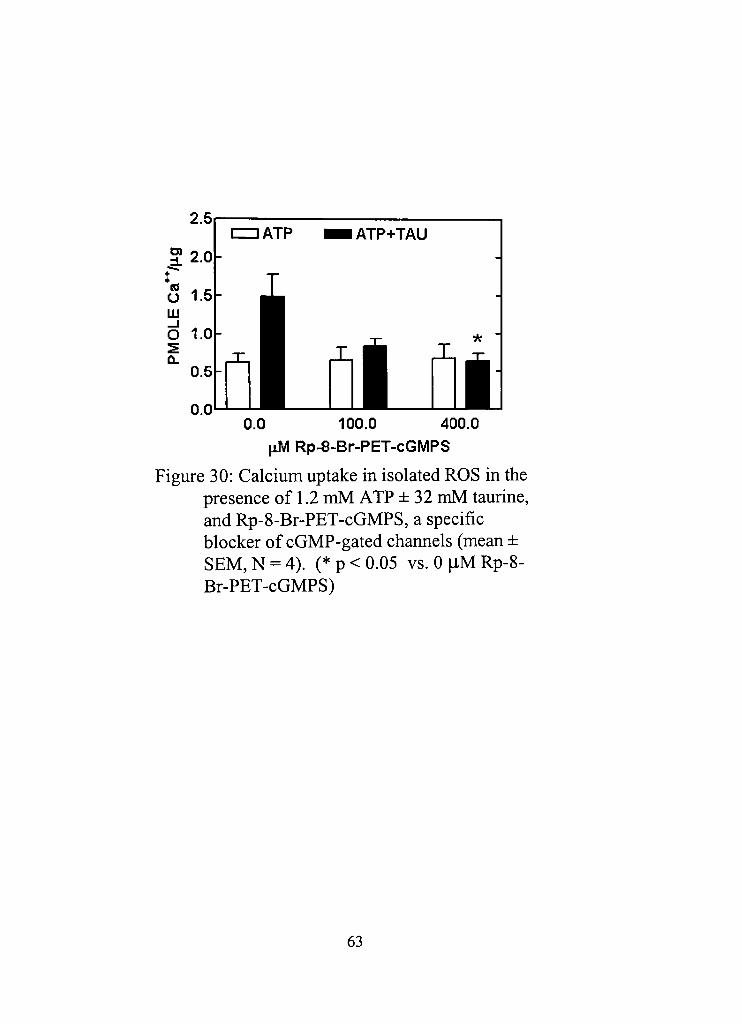
0.0 100.0 400.0 JiM Rp-8-Br-PET-cGMPS
Figure 30: Calcium uptake in isolated ROS in the presence of 1,2 mM ATP ± 32 mM taurine, and Rp-8-Br-PET-cGMPS, a specific blocker of cGMP-gated charmels (mean ± SEM,N = 4). (*p<0.05 vs, 0|iMRp-8-Br-PET-cGMPS)
63

To verify the specific involvement of cGMP-gated channels in the
calcium uptake measured in the retina and in the ROS, activating agents of
the channel were used to stimulate uptake. In these experiments, the isolated
ROS are supposed to be intact as no homogenization was done on the
samples, and the cGMP binding site is not readily accessed by compounds
added in the buffer. The acdvating agents were specifically chosen because
of their ability to cross the plasma membrane and bind to the agonist binding
site of the cGMP-gated channel. Dibutyryl cGMP is a membrane-permeant
analogue of cGMP, and it was found to stimulate ATP-dependent calcium
uptake, but not taurine potendated uptake (Figure 31), This effect was less
potent than what was expected as cGMP acdvates the receptor with a
dissociation constant of 17-30 |iM in patch clamp experiments (Pugh and
Lamb, 1990). The effects of another membrane-permeant analogue of cGMP
(Rp-cGMP) on ATP-dependent calcium uptake were studied, but no
significant effects were observed (Figure 32). The reason behind the
discrepancies with both potency and efficacy observed in these two
experiments may involve a slow rate of diffusion of the cGMP analogues
through the plasma membrane. Incubation was done for only 2 minutes,
during which time the cGMP analogue must diffiise through the plasma
membrane, bind to its receptor and acdvate the channel. A longer
incubadon time may have been necessary for the compounds to access the
appropriate binding site.
64
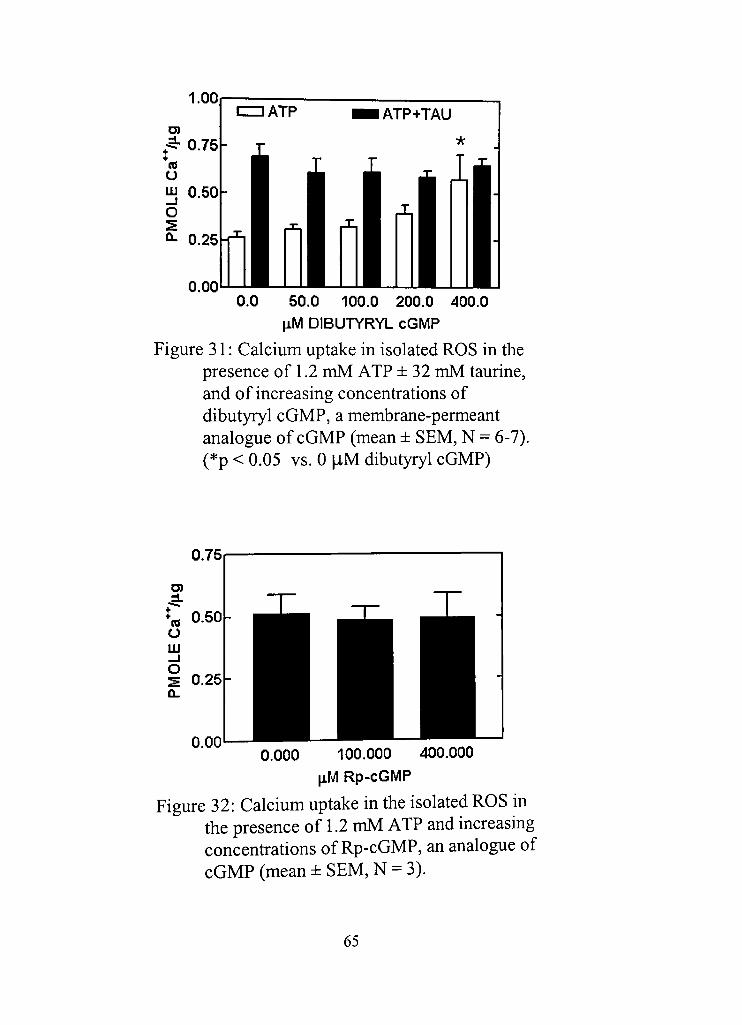
1.00
50.0 100.0 200,0 400.0
JiM DIBUTYRYL cGMP
Figure 31: Calcium uptake in isolated ROS in the presence of 1.2 mM ATP ±32 mM taurine, and of increasing concenfrations of dibutyryl cGMP, a membrane-permeant analogue of cGMP (mean ± SEM, N = 6-7). (*p < 0,05 vs, 0 |iM dibutyryl cGMP)
0.75
D)
*« 0,50 O UJ —I O S 0.25
0.00 0.000 100.000 400.000
JiM Rp-cGMP
Figure 32: Calcium uptake in the isolated ROS in the presence of 1.2 mM ATP and increasing concenfrations of Rp-cGMP, an analogue of cGMP (mean ± SEM, N = 3).
65

The cGMP-gated channels may be activated by increasing the
endogenous levels of cGMP inside the ROS. Zaprinast, otherwise known as
M&B 22,948, is a potent inhibitor (IC50 = 160 nM) of the cGMP-specific
phosphodiesterase (PDE) found in the rod photoreceptor (Gillespie and
Beavo, 1989), and it was used to prevent the degradadon of endogenous
cGMP. However, zaprinast did not have any significant effect on ATP-
dependent calcium uptake in the ROS either in the presence or absence of
taurine (Figure 33). The lack of effects in this experiment and in the
preceding experiment may be due to the fact that the total time of
preincubation and incubation was only 4 minutes which may not be
sufficient time for cGMP analogues and zaprinast to diffuse into the tissue
and produce their effects. It is entirely possible that longer incubation
periods would result in significant stimulatory effects for these. An
important consideration, however, was the maximization or saturation of
calcium uptake that occurs with longer incubation periods, at which point the
observadon of sdmulation would be impossible. Regardless, fiall dme course
studies need to be done to verify the results presented here.
66
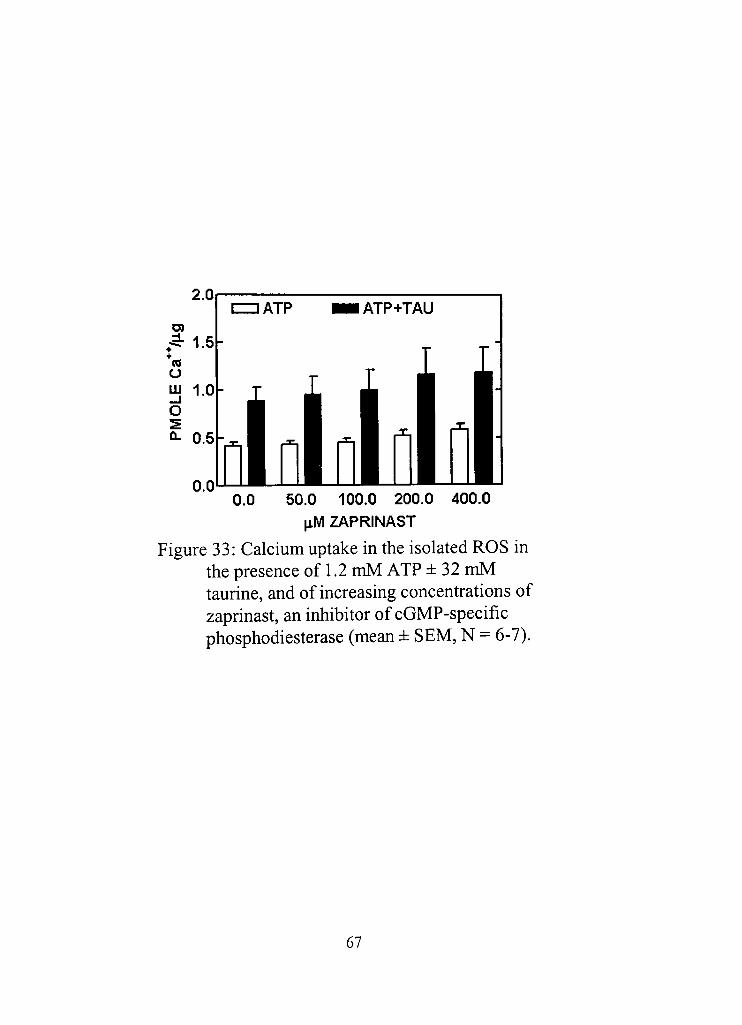
0,0 50,0 100,0 200.0 400,0 JiM ZAPRINAST
Figure 33: Calcium uptake in the isolated ROS in the presence of 1.2 mM ATP ±32 mM taurine, and of increasing concentradons of zaprinast, an inhibitor of cGMP-specific phosphodiesterase (mean ± SEM, N = 6-7).
67

Hvpothesis 4
The effects of taurine may involve calcium binding to membranes and
not calcium uptake exclusively. Experiments were performed to determine if
taurine affects calcium binding to retinal membranes. Calcium binding
experiments were performed in retinal tissue and the effects of taurine were
tested. The retinal tissue was osmodcally lysed with water to denadire the
membranes and incubated with radiolabeled calcium on ice. These
procedures were followed to inhibit active calcium uptake as much as
possible. In preliminary experiments, taurine (32 mM) and ATP (1.2 mM)
individually did not produce any significant effect, but together both
increased calcium binding (Figure 34), However, the effect could not be
replicated in succeeding experiments in which increasing concenfrations of
ATP and taurine were used to stimulate binding (Figure 35 and 36), There
appears to be a slight increase in calcium binding with ATP and taurine
freatment, but statistical significance could not be established. The
inconsistency may be due to the rather small magnitude of the effect
originally observed. Perhaps, increasing the N value for the experiments
would result in significance. Further studies were, thus, done to evaluate the
involvement of calcium binding in the stimulatory effects of taurine on
calcium uptake through the use of A23187, a calcium ionophore.
68
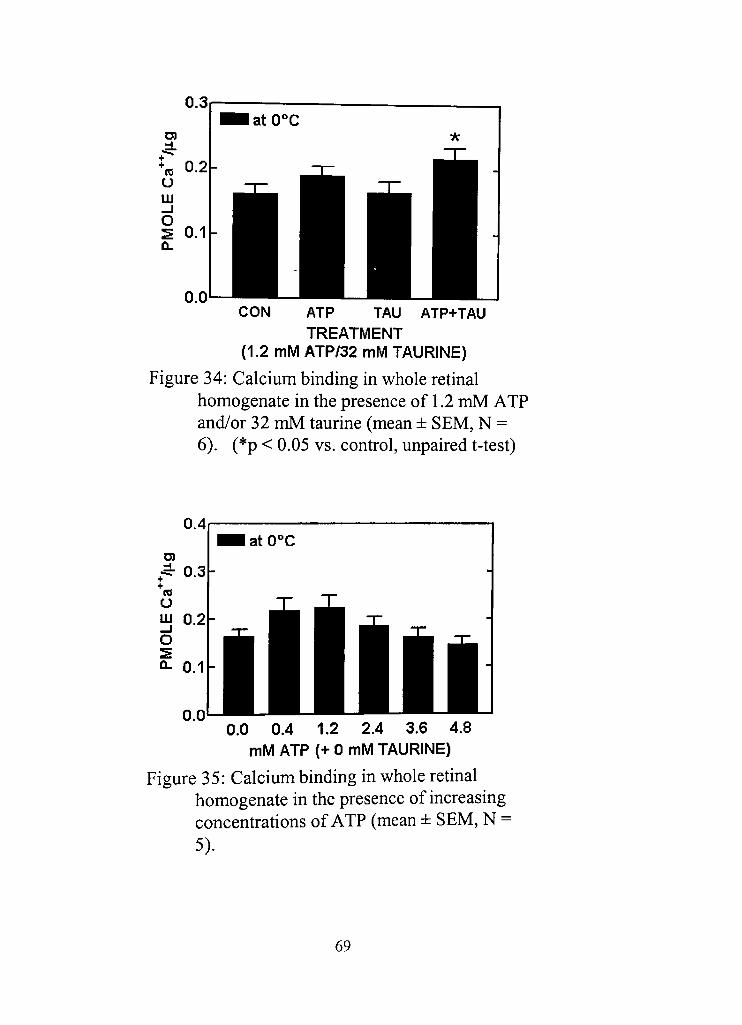
CON ATP TAU ATP+TAU TREATMENT
(1.2 mM ATP/32 mM TAURINE)
Figure 34: Calcium binding in whole retinal homogenate in the presence of 1.2 mM ATP and/or 32 mM taurine (mean ± SEM, N = 6), (*p < 0.05 vs. confrol, unpaired t-test)
0.4
;5- 0.3
o UJ 0.2
O
Q- 0.1
0.0
atO°C
llllU 0.0 0.4 1.2 2.4 3.6 4.8 mM ATP (+ 0 mM TAURINE)
Figure 35: Calcium binding in whole retinal homogenate in the presence of increasing concentradons of ATP (mean ± SEM, N =
5).
69
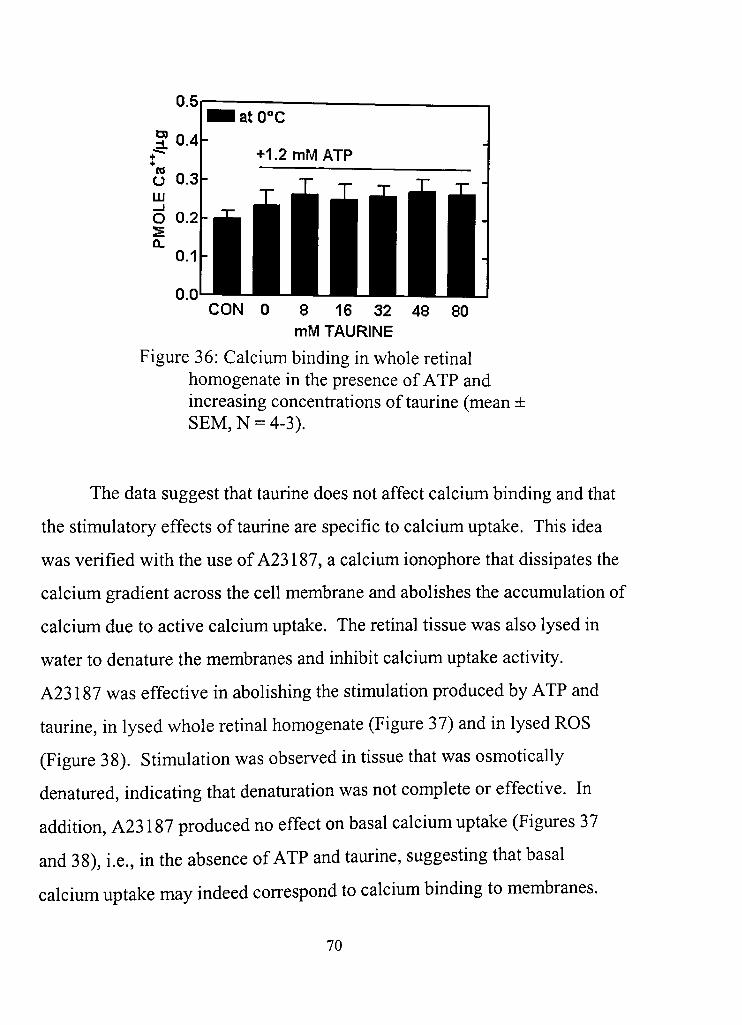
0.5
S 0.4 +
O 0.3 m d 0.2
0.1
0.0
atO°C
+1.2 mMATP
hll l l l l CON 0 8 16 32 48 80 mM TAURINE
Figure 36: Calcium binding in whole retinal homogenate in the presence of ATP and increasing concenfrations of taurine (mean ± SEM, N = 4-3).
The data suggest that taurine does not affect calcium binding and that
the stimulatory effects of taurine are specific to calcium uptake. This idea
was verified with the use of A23187, a calcium ionophore that dissipates the
calcium gradient across the cell membrane and abolishes the accumulation of
calcium due to acdve calcium uptake. The rednal tissue was also lysed in
water to denature the membranes and inhibit calcium uptake activity.
A23187 was effecdve in abolishing the sdmuladon produced by ATP and
taurine, in lysed whole retinal homogenate (Figure 37) and in lysed ROS
(Figure 38). Stimulation was observed in dssue that was osmodcally
denatured, indicadng that denaturadon was not complete or effective. In
addition, A23187 produced no effect on basal calcium uptake (Figures 37
and 38), i,e,, in the absence of ATP and taurine, suggesdng that basal
calcium uptake may indeed correspond to calcium binding to membranes.
70
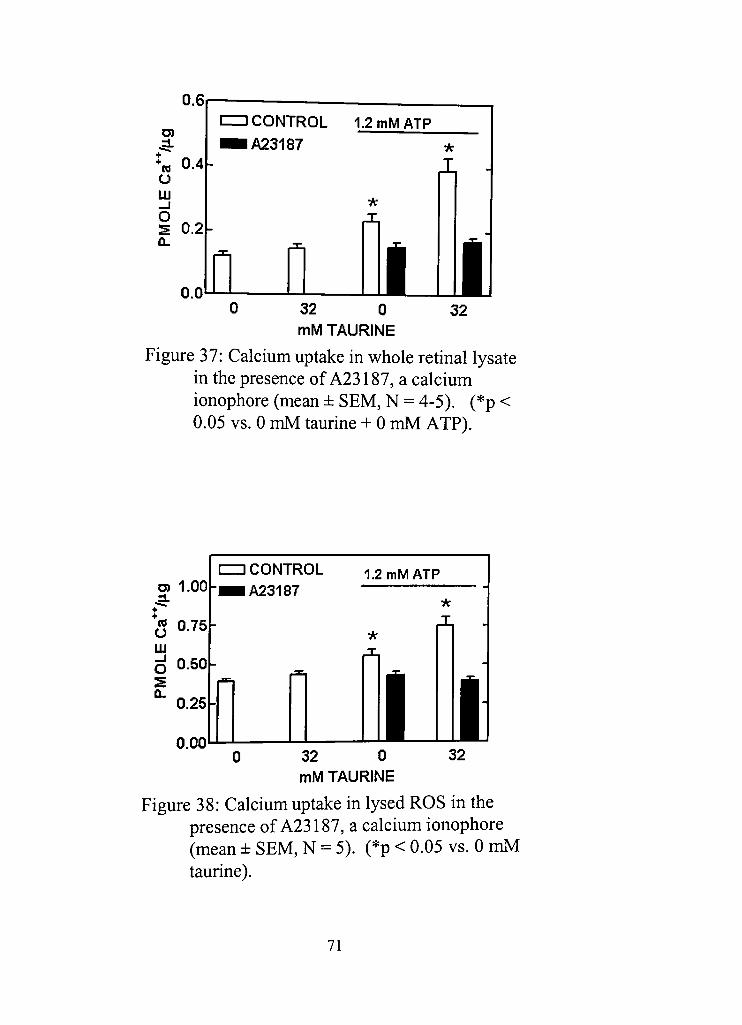
0.6
J . + o m _i o
0.4
0.2
0.0
IZZJ CONTROL 1.2 mMATP ^ A 2 3 1 8 7 *
i . -IL
n n n i l l l 32 0 32 0
mM TAURINE
Figure 37: Calcium uptake in whole retinal lysate in the presence of A23187, a calcium ionophore (mean ± SEM, N = 4-5). (*p 0.05 vs. 0 mM taurine + 0 mM ATP).
<
D) 1.00
5 0.75 UJ ^ 0.50-
^ 0.25
0.00
CONTROL A23187
1.2 mMATP
Ol 32 0 mM TAURINE
32
Figure 38: Calcium uptake in lysed ROS in the presence of A23187, a calcium ionophore (mean ± SEM, N = 5). (*p < 0.05 vs. 0 mM taurine).
71

CHAPTER IV
DISCUSSION
Taurine effects on calcium uptake
Taurine is abundant in the retina, comprising 40-50%o of the total free
amino acids (Macaione et al , 1974) and reaching concentrations as high as
79 mM (Voaden et al , 1977). Retinal cells, as well as other cell types,
possess rather well-regulated uptake systems for taurine, systems that
function to maintain a very steep concenfration gradient for taurine across
the plasma cell membrane. In comparison, only small osmotic ions like
chloride and potassium are handled as carefully and as efficiendy. Taurine is
not incorporated into protein molecules and therefore is not important in
building the cytoskeletal or organellar structure of the cell. Taurine exists
freely in solution, and so if we are to assume that physiologic form follows
physiologic function, then we can surmise that taurine can act as an
osmolyte, and even more significantly, as a ligand of some kind which binds
with low millimolar affinity to receptor molecules to produce specific
infracellular effects.
A good illusfration of the potendal of taurine to funcdon as a regulator
of infracellular events is in vitro phosphoryladon experiments that make use
of retinal subfractions (reviewed in Militante and Lombardini, 2002). In
these experiments, millimolar concenfradons of taurine produced what could
only be described as direct sdmulatory effects on the activity of kinases to
phosphorylate specific proteins. Similar concenfrations of taurine analogues
either did not produce any effects or produced effects opposite that of
72

taurine, suggesting that the effects of taurine are not osmotic in nature.
While the mechanism is unclear, we may assume that taurine binds with low
affinity to either the kinase or the protein substrate to be phosphorylated as a
necessary biochemical mechanism, ft is also reasonable to assume that
within the living cell, taurine is regulating kinase activity in much the same
way. The fact that taurine acts with millimolar potency, as opposed to
neurofransmitter or hormonal ligand molecules which usually act with
micromolar or even nanomolar potency to produce their effects, gives
physiologic relevance to the unusually high concenfradons of taurine
retained within the cell, at least in reladon to the funcdon of protein kinases.
The effects of taurine on calcium uptake present an even more
significant physiologic possibility for taurine. Dwarfing the signaling
possibilides of direct moduladon of specific proteins, moduladon of
intracellular calcium levels could potentially result in calcium-dependent
cascade events involving muldple proteins and multiple cellular systems.
Truly, taurine modulation of calcium flux in the redna could be
physiologically relevant in many ways. For example, in a model of
experimental regeneration of goldfish retina, taurine was demonsfrated to
sdmulate neuridc growth (Lima et al , 1988). This is a system commonly
used for the study of cenfral nervous system regeneration (Landreth and
Agranoff, 1979). In these experiments, taurine appeared to promote growth
by increasing calcium influx (Lima et al , 1993). The exact pathway for this
effect is unclear, but it is almost certain that the stimulation of growth is
dependent on a calcium signal spreading in multiple cellular compartments
and coordinating diverse biochemical processes in its wake.
73

The experiments in this thesis address a specific phenomenon long
observed with taurine and the retina. Taurine is known to produce
stimulatory effects on in vitro calcium uptake in whole retinal homogenate,
specifically, taurine potentiates ATP-enhanced calcium uptake increase
under condidons of low calcium (<500 |iM)(reviewed in Lombardini, 1991).
This effect was also observed with various subcellular fractions: PI fractions
(photoreceptor synaptosomes), P2 fractions (mitochondria and microsomes),
mitochondrial fracdons and ROS fractions (Lombardini, 1985a; Lombardini,
1988). Taurine requires the presence of ATP this stimulatory effect on
calcium uptake, and this phenomenological condngency is yet to be fully
understood.
In the experiments presented in this thesis, it is reported that taurine
produced half-maximal stimuladon at around -18 mM concenfradon, while
previous studies report this value at 8-10 mM (Liebowitz et al, 1987;
Liebowitz et al , 1988) or at -30 mM (Lombardini et al, 1989). While the
potency with which taurine acts may seem to be too low to be
physiologically relevant, the range of effecdve concenfrations, in fact,
coincides with reported levels of taurine in the retina. Additionally, the
active sequesfration of taurine inside the cells where the levels of calcium are
low, and the presence of ATP within the cell gives credence to the nodon
that this specific ATP-dependent sdmulation of calcium uptake in the retina
by taurine may be regarded as material to normal rednal function.
The experiments in this thesis studied the effects of taurine in the
presence of low calcium levels instead of high calcium levels which would
correspond to exfracellular levels of calcium. This condition was chosen
74

mainly to mimic the low intracellular levels of calcium so as to study the
effects of taurine under conditions comparable to conditions found inside the
cell Calcium uptake in the retinal tissue samples increases in almost direct
relation to the level of calcium in the buffer (Militante and Lombardini,
2000), and so it can be assumed that the use of high calcium levels would
result in tissue calcium levels that are higher than physiologic levels of
infracellular calcium. In all types of dssues, calcium flucdiadons occur
inside the cell and taurine may have different effects at different calcium
levels.
Isolated ROS, as opposed to whole retinal homogenate, were also used
in these experiments to discem more closely the site of action of taurine. In
many of the experiments, the stimulatory effects of taurine were
recapitulated in the ROS, mirroring previous reports on calcium uptake in
ROS (Lombardini, 1985a). The ROS is the focal point in the excitation and
recovery of photoreceptor cells with light stimuladon (Baylor, 1996). In
ROS, closure of cGMP-gated cation channels, net exfrusion of calcium, and
the decrease in infracellular calcium that occur during photoexcitadon lead to
the activation of a calcium-sensitive guanylyl cyclase and to an increase in
cGMP. Acdvation of cGMP-gated channels allows for the increased entry of
calcium. Increased calcium levels result in the inhibition of guanylyl cyclase
and in the breakdown of cGMP by phosphodiesterase, allowing for recovery
from the hyperpolarized state of the photoreceptor. The fast and efficient
uptake of calcium is, thus, key to the continued funcdon of the retina. The
data in this thesis suggest that taurine assists in this recovery, specifically in
light of the diminished levels of infracellular calcium that serve as the signal
75

for recovery and that at the same time satisfies the low calcium prerequisite
for stimulatory effects of taurine to occur. The involvement of taurine in
this recovery while unproven, makes kinetic sense as taurine would assist in
the faster uptake of calcium. Indeed, we can speculate that taurine is a vital
component in the immediate recovery of the depolarized state of the
photoreceptor cell during photofransduction. While this idea is largely
untested, it is supported by the report of electrical deficits found in the retina
after taurine depletion in the rat (Cocker and Lake, 1987) or with nufritional
taurine deficiency in human children (Geggel et al , 1985).
ATPase activity in the redna
A major requirement for the stimulatory effects of taurine is ATP.
Early studies reported that the stimulatory effects of taurine appear to be
dependent not only on the presence but also the hydrolysis of ATP
(Pasantes-Morales, 1982). This specific requirement has not been sdidied
much, owing perhaps to the default and seemingly obvious assumption that
adenosinefriphosphatase (ATPase) activity is necessary. Indeed, there have
been very few experiments reported on the interacdon between ATPase and
taurine in the retina.
In the retina, the total ATPase activity, as with other cell types, is
comprised of the activity of many different species of ATP-hydrolyzing
enzymes. Among the subtypes found in the retina are Na^ K^-ATPase, Ca'^-
acdvated, Mg^"-dependent ATPase (Ca^^ Mg^^-ATPase) and Mg^^-ATPase
(Berman et a l , 1977; Chambers et al , 1990). There is also evidence of
bicarbonate-stimulated ATPase activity (Winkler and Riley, 1977) but
76

almost certainly other types of ATPase activity are present. One eariy sdidy
done in this lab suggested that 20 mM taurine had no effect on 4 types of
retinal ATPase activity: NaVK" dependent, Mg^" dependent, Ca'^ dependent,
and Mg^VCa^^ dependent. However, the experiments required that different
buffers be used to delineate ATPase types. As the buffer conditions were
significandy different from that used with calcium uptake assays, it is
difficult to relate the findings to the ATP-dependence of the effects of
taurine. During experimentation, the activity of these enzymes and the other
ATPases that may be present may change depending on the specific ions
present in the assay buffer, e.g., without bicarbonate in the buffer,
bicarbonate-stimulated ATPase would be inactive, and vice versa.
In our experiments, total ATPase acdvity was measured under the
same buffer conditions as with the calcium uptake assay, to make certain that
the same types of ATPase activity are functional in both assays. Moreover,
ATPase activity was measured in the same time interval as the calcium
uptake assay was conducted. Taurine did not affect total ATPase activity,
sfrongly suggesting that the mechanism of action behind the sdmulatory
effects of taurine on calcium uptake does not involve the moduladon of
ATPase activity. However, there is a possibility that the assay is not
sensitive enough to measure the changes in ATPase activity taurine may
produce. Clearly, more accurate experimental techniques need to be used to
verify these findings in the future.
What type or types of ATPase activity acdially fiincdon in our system
and whether this activity is involved in calcium uptake is unknown and
constitutes another interesting field of sdidy. Ouabain did not inhibit the
77

ATPase acdvity to a significant degree, suggesting that Na^, K^ATPase is not
active under the experimental conditions or that it consddites a very small
percentage of the total ATPase activity in the retina. Also, the preparation
of the membranes from the whole cell could have resulted in the formation of
sealed vesicles that do not allow for opdmal binding of sodium and
potassium to the ATPase enzyme, thereby decreasing ATPase activity. In
any case, these results differ from other reports that show measurable levels
of Na" , K"^ATPase activity in the rat redna, a difference probably home out
of the variation in the buffer composition or experimental procedure.
A non-mitochondrial Ca " , Mg^" -ATPase has also been examined in the
bovine retina (Chambers et al , 1990), and CHT might be inhibiting this type
of ATPase in the rat retina. SERCA probably comprises a part, if not all, of
this specific ATPase acdvity, though present literadire does not allow for any
certainty on the matter. SERCA is potentially an important component in the
mechanism of action of taurine in calcium uptake because it is involved in the
acdve uptake of calcium into the endoplasmic redculum. However,
experiments with thapsigargin suggest that SERCA is not a major component
of the total ATPase activity studied.
As mentioned before, some form of bicarbonate-acdvated ATPase is
fimctional with a 25 mM bicarbonate concenfration in the retina (Winkler and
Riley, 1977). Our experiments use 25-50 mM bicarbonate (see Methods),
which possibly activates the enzyme even more. It is possible that this is the
ouabain-insensidve ATPase that is inhibited by CHT. It is infriguing to
correlate the bicarbonate-dependency of this enzyme with the
78

bicarbonate-dependency of the potendadng effects of taurine on calcium
uptake, but our data are not extensive enough to support this concept.
An interesting set of experiments that could be done to verify the data
here reported would be to use subcellular retinal fractions in the ATPase
assays. The whole retinal homogenate samples used would almost certainly
possess several different ATPase types which would be pharmacologically
difficult to differentiate from each other. Perhaps, specific ATPase types
colocalize with different membrane fractions or subcellular organelles, and
the effect of taurine would be more easily demonsfrated. In this light,
ATPase activity in the isolated ROS should be studied as a priority.
Taurine uptake versus taurine binding
Taurine produces its effects on calcium uptake only in the millimolar
range. Thus, the relatively high infracellular concenfradon of taurine appears
to be a funcdonal necessity, at least with respect to its effects on calcium
uptake. Although some degree of endogenous biosynthesis exists, most of
the taurine present in mammalian dssues is exogenous in its source, requiring
a very efficient transport system to achieve high dssue concenfrations
(reviewed in Lombardini, 1991). In the redna of several different species,
taurine uptake is made up of two sadirable components, a high- and a
low-affinity system, and a non-sadirable diffiasion component (reviewed in
Huxtable, 1989). In experiments that use rats with hereditary retinal
degeneradon, the high-affinity system has been shown to be dependent on the
normal ftmcdon of the photoreceptor cells (Schmidt and Berson, 1978).
However, high-affinity taurine uptake may also occur in retinal pigment
79

epithelium (RPE) cells (Salceda and Saldana, 1993). Regardless, high-
affinity taurine uptake seems to disappear with the loss of photoreceptor cells
(Schmidt and Berson, 1978), suggesting that only high-affinity uptake occurs
in the photoreceptor cells and only low-affinity is found in the rest of the
retina.
It is of great interest that the inhibidon of taurine uptake does not
significantly diminish the stimulatory effects of taurine on calcium uptake.
Careful analysis of kinetic data would reveal that taurine uptake cannot be
correlated directed with taurine stimulation of calcium uptake. Half-maximal
low-affinity uptake (Km) in the retina occurs at -2,7 mM taurine, while high-
affinity taurine uptake is saturated at taurine concenfrations below 1 mM. In
confrast, previous studies report that taurine produced half-maximal
stimuladon (EC50) of calcium uptake at 8-10 mM concenfradons (Liebowitz
et al , 1987, 1988), In the experiments presented in this thesis, the
potendadng effect of taurine on ATP-dependent calcium uptake in whole
retinal homogenate was half-maximal at around 18 mM. Indeed, the half-
maximal values for Km for taurine uptake and EC50 for taurine stimulation of
calcium uptake do not correspond and so perhaps may not interact.
Theoredcally speaking, the mechanism behind the sdmulatory effects
of taurine on calcium uptake should exhibit affinity lower than what is
observed with taurine uptake systems. A mechanistic altemadve may be
taurine binding, i,e,, taurine may exert its effects by binding to and modifying
phospholipid membranes (Huxtable and Sebring, 1986), thereby moduladng
the ftinction of membrane-bound proteins. The biological consequences of
the binding of taurine to membranes is unclear. However, early sdidies in this
80

lab which measured calcium uptake while varying incubation temperadire
suggested that taurine acts to induce a conformadonal change within the lipid
membrane (Lombardini, 1985b), Calcium uptake in the retina is temperadire-
dependent, increasing as temperadire increases (Pasantes-Morales, 1982;
Lombardini, 1985b). Experiments were performed which correlated the
temperadire of incubation and retinal calcium uptake. Arrhenius plots for the
ATP-dependent calcium ion uptake in the presence and absence of 20 mM
taurine were constructed from the data and the activadon energy for calcium
uptake was calculated. Taurine produced a decrease in the activadon energy
for calcium uptake, an effect which is consistent with observed increase in
uptake seen with taurine. Moreover, the decrease in activation energy could
be interpreted as an alteration in the crystalline-liquid crystalline phase
fransition of the membrane. These studies suggest that the modificadon of
the cell membrane is a possible mechanism of action for the effects of taurine.
Taurine is known to increasingly bind to membranes in a dose-
dependent manner up to 30 mM concenfradon (for review, see Huxtable,
1989). Huxtable reviewed sodium-dependent taurine binding in neuronal-
type membranes (1989) and showed Kd measurements within the
corresponding range of taurine concenfrations used in experiments with rat
synaptosomal P2B fracdons, and rat glial, brain and synaptic membranes. In
these sdidies that resolved a low-affinity type of taurine binding, the Kd was
estimated to be higher than the highest concenfradon of taurine used. The
taurine binding sdidies of Lombardini and Prien (1983) observed that in the
redna, the calculated Kd of the low-affinity site (-2738 \iU) was higher than
the highest taurine concenfradon used (1000 |iM). By definidon, Kd values
81

determine the ligand concenfration that results in half-maximal binding, and
thus, in these experiments, the reported Kd values imply that maximal ligand
binding is achieved at taurine concentrations that are significantly higher than
taurine concentrations used and that, logically, maximal binding was not
achieved. As half-maximal binding cannot be determined if maximal binding
was not determined, these Kd values were probably significandy
underesdmated, suggesdng that ligand binding affinity is much lower than
expected. The apparent consistency may also represent a non-sadirable type
of binding that manifested itself as a dissociadon constant that would float
outside the range of total ligand used.
That the binding of taurine to neuronal type membrane is non-saturable
is not unreasonable, given that taurine is for the most part assumed to interact
with phospholipids in membranes (Huxtable and Sebring, 1986). High-
affinity binding interactions are associated with minute amounts of
endogenous ligands attaching to very specific protein receptor molecules. If
taurine were to act in this manner, then the vast amount of taurine actively
sequestered by the cell would be pharmacologically useless. The massive
binding of taurine to phospholipids was postulated to affect, among others,
the binding of calcium to membranes, the operadon of ion channels and
protein phosphoryladon processes. This nodon is of great importance in the
redna because of the high levels of taurine and the possible reguladon of ion
channels demonsfrated in the experiments in this thesis.
The binding of taurine to the retinal membrane should be sdidied in
relation to the effects of taurine to calcium uptake. Preliminary binding
sdidies showed that frifluoperazine (TFP), a calmodulin antagonist,
82

antagonized the effects of taurine on calcium uptake (Militante and
Lombardini, 1999b), TFP was also found to inhibit the binding of taurine to
retinal membrane, indicating that TFP, and other inhibitors of the effects of
taurine may interfere with taurine binding as a mechanism of action. The
experiments of this thesis suggest that taurine binding to membranes is a
major mechanism behind the effects of taurine, not only in the retina but also
in other cell types, A new approach to the study of taurine function may be to
relate modulators of taurine effects to the modulation of taurine binding to
membrane, i,e,, inhibitors of the effects of taurine may decrease taurine
binding. An interesting experiment would be to test the effect of CHT on
taurine binding. CHT may antagonize the effects of taurine by inhibidng the
binding of taurine to cell membranes.
The experimental use of chelervthrine (CHT)
The exact mechanism of acdon responsible for the PKC-independent
CHT inhibidon of calcium uptake is unknown. There are several interacdons
between CHT and taurine which are independent of PKC acdvity. For
example, CHT has been shown to exhibit taurine-related effects in other
rednal systems, namely, in the phosphorylation of a -20 kDa mitochondrial
protein (Lombardini, 1995). ft was reported that CHT stimulates
phosphorylation of a -20 kDa protein, a process that taurine inhibits. When
present together, it was found that taurine antagonized this specific effect of
CHT in a non-competitive manner. As shown in the experiments in this
thesis, CHT antagonized taurine uptake and the stimulatory effect of taurine
on ATP-dependent calcium uptake in a non-competitive manner. Both these
83

effects of CHT were also demonsfrated to be independent of PKC acdvity and
both effects exhibited similar potencies for CHT. A possible mechanism may
have CHT acting at one specific site to first affect the phosphorylation of the
-20 kDa protein which, in dim, affects ATP-dependent calcium uptake. It is,
thus, reasonable to speculate that the specific site of CHT action may also
regulate taurine uptake aside from regulating the phosphorylation of the -20
kDa protein and calcium uptake. As CHT and taurine produce effects on the
phosphorylation of the -20 kDa protein and on ATP-dependent calcium
uptake in the absence of the other agent, it is clear that the effects of CHT in
the redna are not totally dependent on its inhibitory effect on taurine uptake.
The data presented in this thesis suggest that taurine and CHT may
interact at the membrane sfructures of the cell. Both agents individually may
bind to the membrane to produce their effects, whether alone or together. This
notion would explain their effects on protein phosphorylation for this process
probably involve membrane-bound protein kinases or protein kinase
substrates. Even more relevant, the binding of taurine to membranes would
explain its effects on ion channels that by definition are always membrane-
bound. Regardless, understanding the effects of CHT will add gready to the
understanding of the effects of taurine.
Modulation of calcium channels
To test for the involvement of calcium channels in the observed taurine
effects, cadmium was used to inhibit calcium uptake. In neurons, cadmium
causes a non-selecdve block of calcium currents at micromolar concenfrations
(2-20 laM) (Carbone and Swandulla, 1990). In Figure 26, the increase in
84

ATP-dependent calcium uptake due to exogenous taurine is shown to be
inhibited by 5 xM cadmium while ATP-dependent calcium uptake (in the
absence of taurine) is not affected. At very high cadmium concentration (100
^M), both ATP-dependent and taurine-stimulated ATP-dependent calcium
uptake were inhibited. These data suggest that the effects of taurine on ATP-
dependent calcium uptake are dependent on the opening of a calcium channel
that cadmium blocks. Exacdy what this channel, or channels, may be is
uncertain as the dssue preparadon contains all the cell types found in the
redna.
In terms of the role taurine may play in the visual signaling process, the
possible effect of taurine on the cGMP-gated channel, among all the other
types of calcium channels, holds the most importance. LY83583 (6-anilino-5,
8-quinolinedione) has been shown to potently block cGMP-gated channels in
olfactory receptor neurons, causing inhibidon of cGMP-dependent currents at
concenfradons as low as 1 |iM (Leinders-Zufall and Zufall, 1995), LY83583
appears to act both directly on the channel and on soluble guanylyl cyclase,
the enzyme that produces cGMP in olfactory receptor neurons. This
compound was thus used to inhibit the increase of calcium uptake due to
taurine. Figure 28 shows the effect of LY83583 on calcium uptake in retinal
membrane preparations, LY83583 has no effect on ATP-dependent calcium
uptake but has a significant inhibitory effect on taurine-potentiated calcium
uptake, albeit much less potendy when compared to patch recording
experiments in olfactory receptor neurons that directly measured calcium
currents (Leinders-Zufall and Zufall, 1995). Though there is no direct way to
correlate patch recordings with the calcium uptake measured in these
85

experiments, the data suggest that the effect of taurine is at least partially
dependent on the open state of the cGMP-gated channel which is allowing
calcium flow into the cell.
In theory, the effect of LY83583 with retinal membrane preparations
should also be observed in isolated ROS. Figure 29 shows ATP-dependent
calcium uptake measured in isolated ROS and the inhibitory effect of
LY83583 on taurine-stimulated ATP-dependent calcium uptake, similar to the
effects seen in retinal membrane preparations. The inhibition is not complete,
indicadng the involvement of other uptake systems or, perhaps, inefficient
drug delivery or effect. As in the homogenate preparadon of the rednal
membranes, LY83583 had no effect on ATP-dependent calcium uptake in the
absence of taurine.
To verify the involvement of the cGMP-gated channel with the effects
of taurine, a competitive antagonist of the channel was used to inhibit ATP-
dependent calcium uptake in the ROS. Rp-8-Br-PET-cGMPS is a
cell-permeant cGMP derivative that has been found to inhibit cGMP-induced
current with an IC50 of 25 |iM in excised patches (Wei et al , 1996).
ATP-dependent calcium uptake was not affected by Rp-8-Br-PET-cGMPS
but taurine-stimulated uptake was inhibited (Figure 30), demonsfradng a
certain level of specificity in the interacdon of taurine and cGMP-gated
channels.
It is important to note that in the absence of taurine, ATP-dependent
calcium uptake in the retina, specifically in the ROS, does not seem to
involve the opening of the cGMP-gated channel, while in the presence of
taurine it does. Thus, it is reasonable to assume that while ATP-dependent
86

calcium uptake in the absence of taurine probably involves a variety of
different systems, taurine may specifically stimulate cGMP-gated channel
opening to induce calcium uptake in the ROS, Taurine could modulate
channel opening by increasing the levels of cGMP, thereby increasing
channel activation, or by increasing the affinity of the channel for its agonist.
In our experimental system, the cGMP-gated channels are assumed to
be preferentially closed as the retinal sample is exposed to ambient light. In
theory, the opening of these channels should result in increased ATP-
dependent calcium uptake, an effect best seen in isolated ROS, as the cGMP-
gated channels are found in the ROS. In patch clamp experiments, cGMP is
known to activate the channel with a dissociation constant Kd (for channel
opening) of 17-30 |iM (Pugh and Lamb, 1990), Dibutyryl-cGMP, a
cell-permeant analogue of cGMP, sdmulated ATP-dependent calcium uptake,
but the increase was minimal for all concenfradons of the agonist below 400
|iM (Figure 31), Another cGMP analogue, Rp-cGMP, did not produce any
effects at all, probably due to limited diffiision and disfribution into the dssue.
There is an obvious difference in potency when these effects of
dibutyryl-cGMP are compared to its effects in patch clamp sdidies, but this is
probably another manifestadon of the lack of direct correladon between
channel sdidies involving patch clamp techniques and acdial calcium uptake
measurements in ROS isolates. It is also possible that, within the
experimental dme period of 2 minutes, dibutyryl-cGMP did not diffiise
quickly enough through the cell membrane to raise the intemal cGMP level to
a level that would result in adequate channel opening and, in dim, stimulation
of ATP-dependent calcium uptake.
87

In confrast, no effect was observed when taurine-stimulated ATP-
dependent calcium uptake was measured in the presence of dibutyryl-cGMP,
However, it is known that without proper depletion protocols, some level of
endogenous cGMP remains in experimentally prepared dissociated ROS
(Cote and Brunnock, 1993), providing low levels of endogenous agonist for
channel activation. Thus, it is possible that cGMP-gated channels were
maximally opened in the presence of taurine, through a mechanism of action
that makes use of endogenously present cGMP, making ineffecdial the
addition of exogenous agonist.
Zaprinast, otherwise known as M&B 22,948, is a potent inhibitor (IC50
= 160 nM) of the cGMP-specific phosphodiesterase (PDE) found in the rod
photoreceptor (Gillespie and Beavo, 1989). This compound was used to
inhibit the degradation of endogenous cGMP in the ROS, potentially
elevadng, or at least maintaining, cGMP levels and theoredcally causing
greater acdvation of the cGMP-gated channels. Linear regression analyses of
the data indicated a significant increasing frend (P < O.Ol) in ATP-dependent
calcium uptake with zaprinast freatment (Figure 33), although the absolute
change above confrol (0 |iM zaprinast) was found to be not significant using
the one-way ANOVA, The data suggest that endogenous cGMP levels were
sufficiendy maintained to cause significant but not maximal channel opening.
Perhaps, there was not enough time for endogenous cGMP to accumulate or
that endogenous cGMP can only accumulate to a limited degree under these
experimental condidons. Similar to the effects of dibutyryl-cGMP, zaprinast
had no significant effect on taurine-sdmulated ATP-dependent calcium, also
suggesting that cGMP-gated channels have been maximally sdmulated
88

already in the presence of taurine. Though the effects of dibutyryl-cGMP and
zaprinast do not suggest an exact mechanism of acdon of taurine, the data still
suggest that the stimuladon of cGMP-gated channels is somehow involved in
calcium uptake measured in ROS,
Experiments that would verify the interaction between taurine and
cGMP-gated channels are primarily of the electrophysiological type. Patches
of ROS could be excised using the patch clamp pipede and cGMP-dependent
conductance through the patch sdidied. Then taurine could be applied to the
patch and any changes in conductance noted. How taurine may increase the
flow of calcium through the channel is unclear, but moduladon of cGMP-
gated channels could at least be verified. Another experiment which would
be more physiological in significance would be to use intact, dark-adapted
ROS with the calcium sensitive dye Indo-1 through a patch-clamp pipette
(Gray-Keller and Detwiler, 1994) and to sdidy light-dependent calcium flux.
Taurine can be dialyzed through the pipette also into the ROS and its effects
on the calcium response inside the intact ROS could be observed.
The idea that taurine modulates the opening of the cGMP-gated
channel is a novel one and may present a very important function for taurine
in the retina, specifically in the ROS. The decrease in calcium levels within
the ROS is a crucial step in the process of photoexcitation. During this period
of low calcium concenfration, the expected effect of taurine would be to
stimulate calcium uptake into the ROS, a potentially beneficial effect in the
process of reestablishing the standing dark current. Taurine, in theory,
stimulates the acdvation of the cGMP-gated channel by endogenous cGMP
during this recovery period. Afterwards, as infracellular calcium levels rise.
89

taurine would lose this stimulatory effect through some feedback mechanism
and would acdially inhibit calcium uptake, preventing calcium overioad. This
inhibitory effect of taurine may or may not involve the function of
cGMP-gated channels, and provides another interesting field for inquiry. In
any case, this type of biphasic modulation of calcium uptake by taurine could
become a very significant consideration in the understanding of
photofransduction in the ROS.
Calcium uptake versus calcium binding
Many similarities can be observed in cardiac sarcolemma calcium
binding and retinal calcium uptake. Taurine has long been considered as a
positive modulator of calcium binding in cardiac sarcolemma (Sebring and
Huxtable, 1985). Specifically, taurine increased calcium binding in
"infracellular" buffers (high sodium-low potassium) but did not affect low-
affinity calcium binding in "extracellular" (low sodium-high potassium)
buffers. When ATP was added, taurine increased calcium binding, regardless
of the sodium concenfration. The findings mirror the stimulatory effects of
ATP and taurine observed in retinal calcium uptake. Two calcium binding
sites were identified in the cardiac sarcolemma, one with dissociation
constant (Kd) about a hundredfold greater than the other (-3.94 mM vs. -0.03
mM). Similarly, in the whole rat retina, two putative "uptake" sites have been
described for calcium, exhibiting Km values of 2076 and 35 jiM
(Lombardini, 1983). Furthermore, previous sdidies with bovine ROS that
used A23187 suggested that more than 50%o of maximal calcium binding
capacity is adained after only one minute incubadon while active ATP uptake
90

condnues for up to 30 minutes after the start of the incubation (Hemminki,
1975). The current experiments measured uptake after only two minutes of
incubadon and may in fact be composed mosdy of calcium binding. It is thus
a valid suspicion that the calcium uptake measured in retinal membranes in
these experiments may have a significant binding component, and that the
stimulatory effects of taurine may also be directed toward this binding
component, as it is with the cardiac sarcolemma. The data in this thesis
which demonsfrated the effects of the calcium ionophore A23187 on calcium
uptake may support this notion. In theory, A23187 should not affect calcium
binding to membranes but only the flux of calcium through the membrane.
Differences should be noted between sarcolemmal calcium binding and
rednal calcium uptake. The calcium uptake components in the retina were
observed with experiments performed at 37''C, a temperature that allows for
the function of acdve calcium fransport systems, with a very short incubadon
time. In confrast, the calcium binding experiments with cardiac sarcolemma
were performed at 24°C for 45 minutes with a special equilibrium dialysis
system that displayed an equilibrium dme beginning at 30 minutes and lasting
for at least two hours. The low temperature and delayed equilibrium are
classic experimental parameters in binding experiments. In order to answer
the question of whether calcium binding or uptake is involved in the effects
of taurine in the retina, conditions were modified to allow for the inhibition of
active calcium uptake,
ATP sdmulated calcium uptake in frog ROS (Pasantes-Morales, 1982),
The ROS membranes were found to be osmodcally lysed, probably an ardfact
of dssue processing. While the osmodc lysis of bovine ROS membrane in
91

water inhibited ATP stimuladon of calcium uptake at 37°C temperadires,
some level of stimuladon could still be observed (Hemminki, 1975).
Similariy, the calcium uptake in both lysed whole retinal homogenate and
ROS (Figures 37 and 38), also performed at 37°C, exhibited some level of
ATP sdmulation and in tum taurine potentiation. It is most interesdng to note
that the osmotic lysis of cell membranes does not completely obliterate the
active fransport of calcium, although clearly ATP-dependent uptake is
compromised. The calcium ionophore A23187 was employed to distinguish
between calcium binding and calcium uptake. A23187 (0.9 |iM) obliterated
the sdmulatory effects of ATP on calcium uptake in both intact and lysed
bovine ROS (Hemminki, 1975). Calcium ionophores A23187 and X537A
also completely inhibited ATP-dependent calcium uptake in frog ROS
(Pasantes-Morales, 1982). In the same manner, A23187 (10 flM) inhibited
the effects of both ATP and taurine in rat retinal samples (Figure 37 and 38).
It can be assumed that calcium uptake measured in the presence of the
calcium ionophore corresponds to the calcium binding to membranes. Given
that A23187 did not decrease uptake below that of confrol levels, the data
suggest that in the absence of ATP and taurine, calcium uptake is almost
totally composed of calcium binding. It is clear that ATP and taurine do not
affect this calcium binding. Conversely, stimuladon by ATP and taurine
corresponds to calcium uptake and not to binding. Experiments on calcium
binding at 0°C support this nodon in that ATP and taurine did not appear to
have any stimulatory effect.
92

Conclusions
Taurine appears to bind to the retinal cell membrane and to increase the
activadon of calcium channels, allowing greater influx of calcium into the
cell. While the effects of taurine are dependent on the presence of ATP, the
activity of ATPase enzymes is probably not involved in the mechanism of
acdon of taurine. The specific inhibidon of taurine uptake did not affect the
stimulatory effects of taurine on calcium uptake, and it was concluded that
taurine uptake is also not involved in the mechanism of acdon of taurine. The
only other altemative was the binding of taurine to the cell membrane and the
moduladon of membrane-bound systems like ion channels. Indeed, the
sdmulatory effects of taurine on retinal calcium uptake was specifically
correlated to the activation of a specific calcium channel for the first time.
The identity of the calcium channels which taurine affects was established
through the use of specific channel inhibitors. Taurine appears to modulate
the activadon of cGMP-gated channels, the channels which confrol the flux of
calcium into the ROS and the function of which is crucial in the
photofransducdon process. The involvement of ion channel activadon was
complemented by the finding that the stimulatory effects of taurine on
calcium uptake are dependent on calcium flux and not on calcium binding to
membrane.
The set of data presented and interpreted in this thesis serves to clarify
some quesdons regarding the effects of taurine on calcium uptake in the
redna, but moreover, it serves to expand the same set of quesdons. If taurine
is acting on the membrane to produce its effects, in what way is it affecdng
the membrane? Does it change the fluidity of the membrane, as early studies
93

using Arrhenius plots seem to indicate (Lombardini, 1985b)? It is an
infriguing possibility as the mechanism would provide an almost blanket
explanadon for the various effects of taurine on cellular processes,
specifically those that can be considered membrane-limited or membrane-
dependent processes. For example, protein phosphorylation can be
modulated by taurine if the kinase and/or the substrate protein involved are
membrane-bound. The function of membrane-bound ion channels should be
easily affected by taurine with this theory in mind. The data truly expands on
the physiological possibilities of taurine beyond the realm of the retina, as the
data can be readily applied to any cell type that expresses funcdonal ion
channels on its cell membrane and that retains millimolar levels of taurine
infracellulariy. In fact, it is entirely possible that the physiological funcdon of
most ion channels is dependent on taurine because it is more the rule that high
levels of taurine are found within a cell than it is the excepdon. In the case of
the retina, the sdmulation of cGMP-gated channels by taurine would serve an
obvious role in the fact and efficient elecfrophysiological recovery of the
photoreceptor cell after light stimulus. This possibility warrants greater
study.
94

REFERENCES
Baylor, D, (1996) How photons start vision. Proceedings of the National Academy of Sciences of the United States of America, 93, 560-5.
Berman, A.L., Azimova, A.M., and Gribakin, F,G, (1977) Localization of Na , K^-ATPase and Ca^^-activated Mg^-dependent ATPase in retinal rods. Vision Research, 17:527-36.
Berson, E.L., Hayes, K.C, Rabin, A,R., Schmidt, S.Y., and Watson, G. (1976) Retinal degeneradon in cats fed casein. II. Supplementadon with methionine, cysteine, or taurine. Investigative Ophthalmology and Visual Science, 15, 52-8,
Carbone, E, and Swandulla, D, (1989) Neuronal calcium channels: kinetics, blockade and modulation. Progress in Biophysics and Molecular Biology, S4,'i\-5%.
Chambers, J,P,, Kumar, P,, Tsin, A,T,C., and Valdez, J,J, (1990) Pardal characterization of a high-affinity [Ca ^ + Mg^^]-dependent adenosinefriphosphatase from bovine retina. Experimental Eye Research, 50, 127-34.
Cocker, S.E. and Lake, N, (1987) Elecfrorednographic alteradons and their reversal in rats freated with guanidinoethyl sulfonate, a taurine depletor. Experimental Eye Research, 45, 977-87,
Cocker, S.E, and Lake, N, (1989) Effects of dark maintenance on retinal biochemistry and function during taurine depletion in the adult rat. Visual Neuroscience, 3, 33-8,
Cohen, H,G., Seifen, E.E., Sfraub, K.D., Tiefenback, C , and Stermitz, F,R. (1978) Strucdiral specificity of the NaK-ATPase inhibidon by sanquinarine, an isoquinolone benzophenanthridine alkaloid. Biochemical Toxicology, 27, 2555-8,
95

Cote, R,H„ and Brunnock, M,A. (1993) Intracellular cGMP concentradon in rod photoreceptors is regulated by binding to high and moderate affinity cGMP binding sites. Journal of Biological Chemistry, 268, 1/1 yu-o.
Fletcher, E,L„ and Kalloniatis, M. (1996) Neurochemical architecdire of the normal and degenerating rat retina. Journal of Comparative Neurology, 376, 343-60,
Gray-Keller, M,P., and Setwiler, P.B. (1994) Teh calcium feedback signal in the photofransducdon cascade of vertebrate rods. Neuron 13 849-61,
Geggel, H,S,, Ament, M,E,, Heckenlively, J,R,, Mardn, D,A., and Kopple, J,D, (1985) Nufridonal requirement for taurine in padents receiving long-term parenteral nutrition. New England Journal of Medicine, 312, 142-6.
Gillespie, P,G., and Beavo, J,A. (1989) Inhibidon and stimuladon of photoreceptor phosphodiesterases by dipyridamole and M&B 22,948. Molecular Pharmacology, 36, 773-81,
Gray-Keller, M.P,, and Detwiler, P,B, (1994) The calcium feedback signal in the phoofransduction cascade of vertebrate rods. Neuron, 13, 849-61,
Heller-Sdlb, B,, van Roeyen, C, Rascher, K., Hartwig, H,G,, Huth, A,, Seeliger, M,W,, Warskulat, U,, and Haussinger, D, (2002) Disrupdon of the taurine fransporter gene (taut) leads to rednal degeneradon in mice, FASEB Journal, 16: 231-3.
Hemminki, K. (1975) Accumuladon of calcium by retinal outer segments. Acta Physiologica Scandinavica, 95, 117-25.
Herbert, J,M,, Augereau, J,M,, Gleye, J,, and Maffrand, J.P, (1990) Chelerythrine is a potent and specific inhibitor of protein kinase C. Biochemical and Biophysical Research Communications, 173, 993-9.
96

Huxtable, R.J, (1989) Taurine in the central nervous system and the maimnalian actions of taurine. Progress in Neurobiology, 32,
Huxtable, R,J. (1992) Physiological actions of taurine. Physiological Reviews, 12, \0\-6?>.
Huxtable, R.J. and Sebring L.A. (1986) Towards a unifying theory for the acdons of taurine. Trends in Pharmacology, 1, 481-5.
Imaki, H„ Jacobson, S,G., Kemp, CM,, Knighton, R.W., Neuringer, M., and Sturman, J, (1993) Retinal morphology and visual pigment levels in 6- and 12-month-old rhesus monkeys fed a taurine-free human infant formula. Journal of Neuroscience Research, 36, 290-304.
Jacobson, S,G,, Kemp, CM,, Borruat, F,X,, Chaitin, M,H,, and Faulkner D.J, (1987) Rhodopsin topography and rod-mediated function in cats with the rednal degeneration of taurine deficiency. Experimental Eye Research, AS, A%\-9^.
Jhiang, S,M., Fithian, L., Smanik, P., McGill, J., Tong, Q,, and Mazzaferri, E,L, (1993) Cloning of the human taurine fransporter and characterizadon of taurine uptake in thyroid cells. FEBS Letters, 318, 139-44,
Kalloniads, M,, Marc, R,E,, and Murry, R.F. (1996) Amino acid signadires in the primate retina. Journal of Neuroscience, 16, 6807-29.
Kijima, Y,, Ogunbunmi, E,, and Fleischer, S. (1991) Dmg acdon of thapsigargin on the Ca^^ pump protein of sarcoplasmic reticulum. Journal of Biological Chemistry, 266, 22912-8.
Kuo, C-H., and Miki, N. (1980) Stimulatory effect of taurine on Ca-uptake by disk membranes from photoreceptor cell outer segments. Biochemical Biophysical Research Communications, 94, 646-51.
97

Lake, N. (1981) Depledon of retinal taurine by treatment with guanidinoethyl sulfonate. Life Sciences, 29, 445-8.
Lake, N. (1982) Depletion of taurine in the adult rat retina. Neurochemical Research,!, 1385-90.
Lake, N. (1986) Electroretinographic deficits in rats freated with guanidinoethyl sulfonate, a depletor of taurine. Experimental Eye Research, 42, S7-9\.
Lake, N. and Cocker, S.E. (1983) In vifro studies of guanidinoethyl sulfonate and taurine fransport in the rat retina. Neurochemical Research, 8, 1557-63,
Lake, N. and Malik, N, (1987) Retinal morphology in rats freated with a taurine fransport antagonist. Experimental Eye Research, 44, 331-46.
Lake, N. and De Marte, L. (1988) Effects of beta-alanine freatment on the taurine and DNA content of the rat heart and retina. Neurochemical Research, 13, 1003-6.
Lake, N,, Marshall, J, and Voaden, M.J. (1978) High affinity uptake sites for taurine in the retina. Experimental Eye Research, 27, 713-8,
Lake, N, and Verdone-Smith, C (1989) Immunocytochemical localization of taurine in the mammalian retina. Current Eye Research, 8, 163-73,
Landreth, G.E. and Agranoff, B,W, (1979) Explant culdire of adult goldfish retina: A model for the sdidy of CNS regeneradon. Brain Research, 161,39-55.
Leinders-Zufall, T. and Zufall, F. (1995) Block of cyclic nucleodde-gated channels in salamander olfactory receptor neurons by guanylyl cyclase inhibitor LY 83583. Journal of Neurophysiology, 74, 2759-62.
98

Leon, A., Levick, W.R., and Sarossy, M.G, (1995) Lesion topography and new histological features in feline taurine deficiency retinopathy. Experimental Eye Research, 61, 731-41.
Liebowitz, S,M., Lombardini, J,B,, and Salva, P, (1987) Cyclic taurine analogs: Synthesis and effects on ATP-dependent Ca2+ uptake in rat retina. Biochemical Pharmacology, 36, 2109-14.
Liebowitz, S.M., Lombardini, J.B., and Allen, CI. (1988) Effects of aminocycloalkanesulfonic acid analogs of taurine on ATP-dependent calcium ion uptake and protein phosphorylation. Biochemical Pharmacology, "M, 1303-9.
Liebowitz, S.M,, Lombardini, J.B,, and Allen, CI. (1989) Sulfone analogues of taurine as modifiers of calcium uptake and protein phosphorylation in rat retina. Biochemical Pharmacology, 38, 399-406.
Lima, L, (1999) Taurine and its frophic effects in the retina. Neurochemical Research, 24, 1333-8,
Lima, L,, Madis, P,, and Drujan, B, (1988) Taurine effects on neuridc growth from goldfish retinal explants. International Journal of Developmental Neuroscience, 6, 417-24,
Lima, L„ Madis, P,, and Drujan, B, (1993) Taurine-induced regeneradon of ' goldfish retina in culture may involve a calcium-mediated mechanism. Journal ofNeurochemistry, 60, 2153-8.
Liu, Q,_R„ Lopez-Corcuera, B., Nelson, H,, Mandiyan, S„ and Nelson, N, (1992) Cloning and expression of a cDNA encoding the fransporter of taurine and P-alanine in mouse brain. Biochemistry, 89, 12145-9,
Lombardini J B (1983) Effects of ATP and taurine on calcium uptake by
membrane preparadons of the rat redna. Journal ofNeurochemistry,
40, 402-6,
99

Lombardini, J,B. (1985a) Effects of taurine on calcium ion uptake and protein phosphorylation in rat retinal membrane preparations. Journal ofNeurochemistry, 45, 268-75.
Lombardini, J.B. (1985b) Taurine effects on the transition temperadire in Arrhenius plots of ATP-dependent calcium ion uptake in rat rednal membrane preparadons. Biochemical Pharmacology, 34,3741-5.
Lombardini, J.B. (1983) Effects of taurine and mitochondrial metabolic inhibitors on ATP-dependet Ca ^ uptake in synaptosomal and mitochondrial subcellular fractions of rat retina. Journal of Neurochemistry, 51, 200-5.
Lombardini, J.B. (1991) Taurine: retinal ftinction. Brain Research Brain Research Review, 16, 151-69. Review.
Lombardini, J.B. (1992) Effects of taurine on the phosphoryladon of specific proteins in subcellular fractions of the rat retina. Neurochemical Research, 17,821-4.
Lombardini, J.B. (1993) Partial characterizadon of an approximately 20 K M(r) retinal protein whose phosphorylation is inhibited by taurine. Biochemical Pharmacology, 46, 1445-51,
Lombardini, J,B, (1995) Paradoxical stimulatory effect of the kinase inhibitor chelerythrine on the phosphoryladon of a approximately 20 K M(r) protein present in the mitochondrial fracdon of rat redna. Brain Research, 673, 194-8,
Lombardini, J,B, (1996) Stimuladon by chelerythrine of the phosphorylation of the amino acid serine in an -20 kDa protein present in the mitochondrial fracdon of the rat retina. Biochemical Pharmacology, 52, 253-7.
Lombardini, J.B, and Liebowitz, S.M. (1990) Taurine analogues as modifiers of the accumulation of''^calcium ions in a rat rednal membrane preparadon. Current Eye Research, 9, 1147-56,
100

Lombardini, J,B, and Prien, S.D. (1983) Taurine binding by rat rednal membranes. Experimental Eye Research, 37, 239-50.
Lombardini, J.B., Liebowitz, S.M., and Chou, T.-C (1989) Analogues of taurine as sdmulators and inhibitors of ATP-dependem calcium ion uptake in rat retina: Combination kinetics. Molecular Pharmacology, 36, 256-264.
Lombardini, J.B., Young, R,S,L. and Props, CL. (1996) Taurine depledon increases phosphorylation of a specific protein in the rat retina. Amino Acids, 10, 153-165,
Lopez-Colome, A.M, and Pasantes-Morales, H. (1980) Taurine interacdons with chick retinal membranes. Journal of Neurochemistrv 34 1047-52.
Lopez-Colome, A.M,, Fragoso, G,, and Salceda, R, (1991) Taurine receptors in membranes from retinal pigment epithelium cells in culdire. Neuroscience, 41, 791-6.
Lytton, J,, Westin, M., and Hanley, M,R. (1991) Thapsigargin inhibits the sarcoplasmic or endoplasmic reticulum Ca-ATPase family of calcium ^umps. Journal of Biological Chemistry, 266, 17067-17071,
Macaione, S,, Ruggeri, P,, De Luca, F., and Tucci, G. (1974) Free amino acids in developing retina. Journal ofNeurochemistry, 22, 887-891.
Madsen, S,, Ottersen, 0,P,, and Storm-Mathisen, J. (1985) Immunocytochemical visualizadon of taurine: neuronal localization in the rat cerebellum. Neuroscience Letters, 60, 255-60.
Marc, R,E,, Murray, R.F,, and Basinger, S.F. (1995) Pattem recognidon of amino acids in retinal neurons. The Journal of Neuroscience, 15, 5106-29,
101

Marc, R,E., Murry, R.F., Fisher, S.K., Linberg, K.A., Lewis, G.P., and Kalloniatis, M. (1998) Amino acid signadires in the normal cat redna. Investigative Ophthalmology and Visual Science, 39, 1685-93.
Michel, S., Schoch, K., and Stevenson, P.A. (2000) Amine and amino acid fransmitters in the eye of the mollusc Bulla gouldiana: an immunocytochemical study. Journal of Comparative Neurolo2v 425, 244-56,
Militante, J.D, and Lombardini, J.B. (1998a) Effect of taurine on chelerythrine inhibition of calcium uptake and ATPase acdvity in the rat retina. Biochemical Pharmacology, 55, 557-65.
Militante, J.D. and Lombardini, J.B. (1998b) Pharmacological characterization of the effects of taurine on calcium uptake in the rat retina. Amino Acids, 15, 99-108.
Militante, J.D. and Lombardini, J.B. (1999a) Taurine uptake acdvity in the rat retina: protein kinase C-independent inhibidon by chelerythrine. Brain Research, 818, 368-74,
Militante, J,D, Lombardini, J.B, (1999b) Sdmulatory effect of taurine on calcium ion uptake in rod outer segments of rat retina is independent of taurine uptake. Journal of Pharmacology and Experimental Therapeutics, 291, 383-9,
Militante, J,D. and Lombardini, J.B, (2000) Stabilizadon of calcium uptake in rat rod outer segments by taurine and ATP. Amino Acids, 19, 561-70.
Militante, J.D. and Lombardini, J.B. (2002) Taurine: Evidence of physiological function in the retina. Nutritional Neuroscience, 5, 75-90.
Miyamoto, Y,, Liou, G,I., and Sprinkle, T,J. (1996) Isoladon of a cDNA encoding a taurine fransporter in the human retinal pigment epithelium. Current Eye Research, 15, 345-9,
102

Morimura, H„ Shimada, S., Otori, Y., Saishin, Y., Yamauchi, A., Minami, A., et al. (1997) The differendal osmoreguladon and localization of taurine fransorter mRNA and NaVmyo-inositol cofransporter mRNA in rat eyes. Brain Research Molecular Brain Research, 44, 245-252.
Nag, T.C, Jotwani, G., and Wadhwa, S. (1998) Immunohistochemical localization of taurine in the retina of developing and adult human and adult monkey. Neurochemistry International, 33, 195-200.
Nakahishi, S., Matsuda, Y., Iwahashi, K., and Kase, H. (1986) K-252b, c and d, potent inhibitors of protein kinase C from microbial origin. Journal of Antibiotics (Tokyo), 39, 1066-1071,
Neal, M,J. and White, R.D. (1978) Discriminadon between descriptive models of L-glutamate uptake by the retina using non-linear regression analysis. Journal of Physiology, 277, 387-394.
Neuringer, M, and Sturman, J. (1987) Visual acuity loss in rhesus monkey infants fed a taurine-free human infant formula. Journal of Neuroscience Research, 18, 597-601.
Okada, M,, Okuma, Y,, Osumi, Y., Nishihara, M,, Yokotani, K., and Ueno, H, (2000) Neurofransmitter contents in the redna of RCS rat. Graefe's Archive for Clinical and Experimental Ophthalmology, 238, 998-1001,
Omura, Y, and Inagaki, M. (2000) Immunocytochemical localization of taurine in the fish redna under light and dark adaptations. Amino Acids, 19, 593-604,
Ottclecz, A,, Garcia, C,A„ Eichberg, J., and Fox, D,A, (1993) Alteradons in retinal Na^, K^-ATPase in diabetes: Sfreptozotocin-induced and Zucker diabetic fatty rats. Current Eye Research, 12,1111-1121,
103

Ottersen, 0,P„ Madsen, S,, Meldmm, B.S., and Storm-Mathisen, J. (1985) Taurine m the hippocampal formation of the Senegalese baboon, Papio papio: an immunocytochemical sdidy with an antiserum against conjugated taurine. Experimental Brain Research, 59, 457-62.
Pasantes-Morales, H, (1982) Taurine-calcium interacdons in frog rod outer segments: Taurine effects on an ATP-dependent calcium franslocadon process. Vision Research, 22, 1487-1493.
Pasantes-Morales, H,, and Quesada, O, (1980) Effects of the ionophores X537A and A23187 on GABA, glycine, and taurine release from chick rednal subcellular fracdons. Neurochemical Research, 5, 1077-88,
Pasantes-Morales, H,, and Ordoiiez, A„ (1982) Taurine activadon of a bicarbonate-dependent, ATP-supported calcium uptake in frog rod outer segments. Neurochemical Research, 7, 317-28.
Pasantes-Morales, H., Quesada, O,, Carabez, A., and Huxtable, R.J, Effects of the taurine fransport antagonist, guanidinoethane sulfonate, and P-alanine on the morphology of rat retina, (1983) Journal of Neuroscience Research, 9, 135-43,
Pow, D.V. (1994) Taurine, amino acid fransmitters, and related molecules in the retina of the Ausfralian lungfish Neoceratodus forsteri: a light-microscopic immunocytochemical and elecfron-microscopic sdidy. Cell Tissue Research, 278, 311-26.
Pow, D,V., Sullivan, R., Reye, P., and Hermanussen, S. (2002) Localizadon of taurine fransporters, taurine, and ^H taurine accumulation in the rat retina, pituitary, and brain. Glia, 37, 153-168.
Pugh, E.N. Jr. and Lamb, T.D. (1990) Cyclic GMP and calcium: the intemal messengers of excitation and adaptadon in vertebrate photoreceptors. Vision Research, 30,1923-1948,
104

Quesada, O., Huxtable, R.J., and Pasantes-Morales, H. (1984) Effect of guanidinoethane sulfonate on taurine uptake by rat redna. Journal of Neuroscience Resarch, 11,179-86,
Quesada, O., Picones, A., and Pasantes-Morales, H. (1988) Effect of light deprivation on the ERG responses of taurine-deficient rats. Experimental Eye Research, 46, 13-20.
Qian, X,, Vinnakota, S,, Edwards, C, and Sarkar, H.K. (2000) Molecular characterization of taurine transport in bovine aortic endothelial cells. Biochimica et Biophysica Acta, 1509, 324-34.
Ramamoorthy, S., Leibach, F.H., Mahesh, V.B,, Han, H,, Yang-Feng, T., Blakely, R,D., et al, (1994) Functional characterizadon and chromosomal localizadon of a cloned taurine fransporter from human placenta. Biochemical Journal, 300, 893-900,
Rapp, L.M,, Thum, L.A., and Anderson, R.E. (1988) Synergism between environmental lighting and taurine depletion in causing photoreceptor cell degeneration. Experimental Eye Research, 46, 229-38.
Salceda, R. (1980) High-affinity taurine uptake in developing redna. Neurochemical Research, 5, 561-72,
Salceda, R, (1999) Insulin-sdmulated taurine uptake in rat retina and retinal pigment epithelium. Neurochemistry International, 35, 301-6.
Salceda, R, and Pasantes-Morales, H, (1982) Uptake, release, and binding of taurine in degenerated rat retinas. Journal of Neuroscience Research, S, 631-42.
Salceda, R. and Saldana, M,R. (1993) Glutamate and taurine uptake by retinal pigment epithelium during rat development. Comparative Biochemistry and Physiology C-Pharmacology, Toxicology, and Endocrinology, 104, 311-6,
105

Satoh, H. and Sperelakis, N, (1998) Review of some actions of taurine on ion channels of cardiac muscle cells and others. General Pharmacology, 30, 451-63.
Schafer, S., Bicker, G„ Ottersen, 0,P„ and Storm-Mathisen, J, (1988) Taurine-like immunoreacdvity in the brain of the honeybee. Journal of Comparative Neurology, 268, 60-70.
Schmidt, S.Y. and Berson, E.L. (1978) Taurine uptake in isolated retinas of normal rats and rats with hereditary rednal degeneration. Experimental Eye Research, 27, 191-8.
Schmidt, S.Y., Berson, E.L., and Hayes, K.C. (1976) Retinal degeneradon in cats fed casein. I. Taurine deficiency. Investigative Ophthalmology and Visual Science, 15, 47-52.
Sebring, L.A., and Huxtable, R.J, (1985) Taurine moduladon of calcium binding to cardiac sarcolemma. The Journal of Pharmacology and Experimental Therapeutics, 232, 445-51,
Shimada, C , Tanaka, S,, Hasegawa, M,, Kuroda, S., Isaka, K., Sano, M. et al, (1992) Beneficial effect of infravenous taurine infusion on elecfrorednographic disorder in taurine deficient rats, Japanese Journal of Pharmacology, 59, 43-50,
Smith, K,E., Borden, L,A,, Wang, CH., Hartig, P.R., Branchek, T.A., and Weinshank, R.L, (1992) Cloning and expression of a high affinity taurine fransporter from rat brain. Molecular Pharmacology, 42, 563-9,
Sturman, J.A. (1993) Taurine in development. Physiological Reviews, 73, 119-47,
Timbrell, J,A., Seabra, V., and Waterfield, CJ. (1995) The in vivo and in vitro protecdve properties of taurine. General Pharmacology, 26, 453-62. Review.
106

Uchida, S,, Kwon, H,M,, Yamauchi, A., Preston, A.S., Marumo, F,, and Handler, J,S, (1992) Molecular cloning of the cDNA for an MDCK cell Na(+)- and Cl(-)-dependent taurine fransporter that is regulated by hypertonicity. Proceedings of the National Academy of the Sciences of the United States of America, 89, 8230-4,
Uchida, S., Kwon, H.M,, Yamauchi, A,, Preston, A.S„ Marumo, F,, Handler, J,S, (1993) Molecular cloning of the cDNA for an MDCK cell Na(+)- and Cl(-)-dependent taurine transporter that is regulated by hypertonicity. Proceedings of the National Academy of the Sciences of the United States of America, 90, 7424.
Vinnakota, S., Qian, X., Egal, H,, Sarthy, V,, and Sarkar, H,K, (1997) Molecular characterizadon and in situ localizadon of a mouse rednal taurine fransporter. Journal ofNeurochemistry, 69, 2238-50.
Voaden, M.J,, Lake, N,, Marshall, J,, and Morjaria, B, (1977) Studies on the distribution of taurine and other neuroacdve amino acids in the redna. Experimental Eye Research, 25, 219-257.
Wei, J.-Y,, Cohen, E,D,, Tan, Y,-Y,, Genieser, H.-G., and Barnstable, C.J. (1996) Identification of competitive antagonists of the rod photoreceptor cGMP-gated cation channel: P-phenyl-1, N^-etheno-substituted cGMP analogues as probes of the cGMP-binding site. Biochemistry, 35, 16815-16823.
Winkler, B.S. and Riley, M.V. (1977) Na^-K^ and HCOj" ATPase activity in the redna: Dependence on calcium and sodium. Investigative Ophthalmology and Visual Science, 16, 1151-1154,
Winkler, B,S,, Riley, M,V, (1980) Influence of calcium on retinal ATPases, Investigative Ophthalmology and Visual Science, 19, 562-564.
Wright, CE,, Tallan, H,H., Lin, Y,Y., and Gaull, G.E, (1986) Taurine: biological update, Annuual Review of Biochemistry, 55,427-53,
107





























