Embed Size (px)
Citation preview

Telomerase and Neuronal Marker Statusof Differentiated NT2 and SK-N-SHHuman Neuronal Cells and PrimaryHuman Neurons
Pooja Jain,1 Maria A. Cerone,1,2 Andrea C. LeBlanc,1,3 and Chantal Autexier1,2,4*1Bloomfield Center for Research in Aging, Lady Davis Institute for Medical Research,Sir Mortimer B. Jewish General Hospital, Montreal, Canada2Department of Anatomy and Cell Biology, McGill University, Montreal, Canada3Department of Neurology and Neurosurgery, McGill University, Montreal, Canada4Department of Medicine, McGill University, Montreal, Canada
Upon treatment with retinoic acid, NTera-2 (NT2) humanteratocarcinoma and SK-N-SH neuroblastoma cells canbe induced to terminally differentiate into postmitotic neu-ronal cells. The neuronal cell yield obtained from the NT-2cells is partially dependent on the time of differentiation(24–55 days). SK-N-SH cells differentiate into a mixedpopulation of neuronal and epithelium-like cells. Here wereport modified protocols that increase the number of dif-ferentiated NT-2 and SK-N-SH cells and that establish anenrichedneuronal SK-N-SH-derived cell population essen-tially devoid of nonneuronal cells. Differentiated cellsexpress the cytoskeleton-associated protein tau andother typical neuronal markers, such as Map2, Ngn1,NeuroD, Mash1, and GluR which are also expressed inprimary human fetal neurons. Telomerase activity isdown-regulated in differentiated cells, which is consistentwith the telomerase status of primary fetal human neu-rons. Thus, differentiated NT2 and SK-N-SH cells mayrepresent an excellent source for studies investigating therole of telomerase or other survival-promoting activities inprotecting human neuronal cells from cell death-media-ting stresses associated with neurodegenerative dis-eases. VVC 2006 Wiley-Liss, Inc.
Key words: NT2 cells; SK-N-SH cells; telomerase
Molecular mechanisms that mediate both normal andabnormal changes that occur during aging have been investi-gated (Troen, 2003; LeBlanc, 2005; Lombard et al., 2005;Mattson and Magnus, 2006). Certain premature aging syn-dromes, such as ataxia telangiectasia, exhibit symptoms ofneurodegeneration; are mediated by defects in genes that reg-ulate DNA repair, genome surveillance, and the cell cycle;and are characterized by genomic instability, including telo-meric impairment (Callen and Surralles, 2004; Barlow andTreuner, 2005). The increased chromosomal instability isone of the causes of the neuronal cell degeneration and deaththat are also present in neurological disorders such as Alzhei-mer’s disease (Barlow and Treuner, 2005; LeBlanc, 2005).
Much research has focused on the identification ofmechanisms that promote neuronal cell function. Althoughneurons are postmitotic, they are transcriptionally activeand require maintenance of genomic stability to survive(Nouspikel and Hanawalt, 2002; Becker and Bonni, 2005).Mechanisms that maintain chromosome integrity, includ-ing telomerase, could ultimately be developed into thera-peutic strategies to improve brain cellular survival (Mattsonand Klapper, 2001; Blasco, 2005; Dong et al., 2005).
Several studies have implicated telomerase in thecontrol of cell survival and proliferation and resistance togrowth inhibition and cell death (Smogorzewska and deLange, 2004; Calado and Chen, 2006). These effects oftelomerase are sometimes observed in the absence of telo-mere length changes or cell division. For example,expression of telomerase in rodent cell lines and mousehippocampal neurons increases resistance to DNA-dam-aging agents and amyloid b-peptide, a neurotoxic proteinlinked to Alzheimer’s disease (Zhu et al., 2000; Lu et al.,2001). Because the regulation of telomerase and telomerehomeostasis in rodents and humans is different (Wrightand Shay, 2000), it is important to understand the role oftelomerase in human neuronal cell survival.
The first two authors contributed equally to this work.
Contract grant sponsor: Canadian Institutes of Health (CIHR); Contract
grant number: IAO-64655 (to C.A.); Contract grant number: MOP-
15118 (to A.C.L.); Contract grant sponsor: Valorisation Recherche Que-
bec (to C.A., A.C.L.); Contract grant sponsor: U.S. Army Department of
Defense Breast Cancer Research Program (to M.A.C.).
*Correspondence to: Chantal Autexier, Bloomfield Center for Research in
Aging, Lady Davis Institute for Medical Research, Sir Mortimer B. Jewish
General Hospital, Montreal , Canada H2T 1E2.
E-mail: [email protected]
Received 17 May 2006; Revised 23 August 2006; Accepted 25 August
2006
Published online 30 October 2006 in Wiley InterScience (www.
interscience.wiley.com). DOI: 10.1002/jnr.21094
Journal of Neuroscience Research 85:83–89 (2007)
' 2006 Wiley-Liss, Inc.

Studies of human neuronal function and dysfunctionare limited by the lack of an adequate source of humanbrain cells. Ultimately, a source of readily available humanneurons could be used for neural transplantation and brainrepair and to screen neuroactive compounds for the imple-mentation of neuroprotective and neuroregenerative strat-egies (Trojanowski et al., 1997; Paquet-Durand et al.,2003; Villa et al., 2004; Wright et al., 2006). Neuronalfunction has typically been investigated with rodent celllines such as PC12 cells, primary hippocampal and corticalrodent cultures, and human cell types such as the humanteratocarcinoma NTera2 (NT2) and SK-N-SH neuroblas-toma cell lines (Greene and Tischler, 1976; Sidell et al.,1983; Andrews, 1984; Ciccarone et al., 1989; Mattsonet al., 1995). Alternative approaches involve the use of pri-mary human fetal neurons or human stem cells (LeBlanc,1995; Ostenfeld et al., 2000; Reali et al., 2006). Of interestin our studies on telomerase were the NT2 and SK-N-SHcells and primary human fetal neurons. Upon treatmentwith retinoic acid, NT2 and SK-N-SH cells can beinduced to terminally differentiate into postmitotic neuro-nal cells (Andrews, 1984; Ciccarone et al., 1989; Pleasureet al., 1992; Przyborski et al., 2000; Pizzi et al., 2002).
To assess the differentiated NT2, SK-N-SH, and pri-mary human fetal neurons as possible models systems forinvestigating telomerase function in human neurons, wecharacterized the telomerase and neuronal marker status inthese cells. We developed modified differentiation meth-ods for NT2 and SK-N-SH cells, which increase the num-ber of differentiated NT-2 and SK-N-SH cells comparedwith previous protocols and which establish an enrichedneuronal SK-N-SH-derived cell population essentiallydevoid of nonneuronal cells. We determined that the telo-merase and neuronal marker statuses of differentiated NT2and SK-N-SH cells and of primary human fetal neuronsare similar. Thus these cells may constitute excellent sour-ces for studies to investigate the role of telomerase in pro-tecting human neuronal cells from cell death-mediatingstresses that can lead to neurodegenerative diseases.
MATERIALS AND METHODS
Cell Lines and Cell Adhesion Differentiation Protocols
The NT2 (Ntera2/D1) cells obtained from Stratagene (LaJolla, CA) were differentiated by using the manufacturer’s celladhesion protocol from Pleasure and colleagues (1992). Briefly,cells were grown in DMEM/F-12 (Invitrogen, San Diego, CA)supplemented with 2 mM L-glutamine and 1% penicillin andstreptomycin (complete media) and 10% (v/v) fetal bovine se-rum (FBS) in the presence of 10 lM retinoic acid [all-trans-reti-noic acid (RA); Sigma, St. Louis, MO] for a period of 5–6 weeks. Medium was replaced every 2–3 days. The cells werethen expanded, replated in complete medium with 10% FBS inthe absence of RA, and allowed to attach for 2–3 days, duringwhich time the cells begin to differentiate into postmitotic neu-ronal cells (replate I). To prevent undifferentiated cells fromovergrowing the neuronal cell population, the cultures weretreated with mitotic inhibitors [MI; 10 lM 5-fluoro-20-deoxyur-idine (FUdR), 10 lM 1-b-D-ribofuranosyluracil (Urd), 10 lM
cytosine b-D-arabinofuranoside (araC)] for 7–10 days in 50%conditioned media (obtained from cells grown for 2–3 days afterthe first replate) and 50% complete medium containing low FBS(10% replaced by 5%). The neuronal cells were then selectivelytrypsinized and maintained on poly-D-lysine- and Matrigel ma-trix-coated plates in 50% conditioned medium and 50% com-plete medium with 5% FBS to promote neurite outgrowth(replate II). The SK-N-SH cells were obtained from ATCC anddifferentiated by using the cell adhesion method of differentia-tion, which consisted essentially of maintaining the cells in MEM(Invitrogen) supplemented with 2 mM L-glutamine, 1% penicil-lin/streptomycin and antimycotic, 1 mM sodium pyruvate, and10% FBS plus 10 lM RA for 2 weeks (Kraveka et al., 2003).
Modified NT2 Cell Aggregation Differentiation Protocol
This protocol was modified from the previously reportedcell aggregation method (Paquet-Durand et al., 2003). Cells(1–1.5 3 106) were grown in DMEM/F-12 medium (Gibco,Grand Island, NY) supplemented with 2 mM L-glutamine, 1%penicillin/streptomycin, and 10% FBS on bacteriological platesto maintain cells in suspension to induce neurosphere forma-tion. On day 2, 10 lM RA was added, and differentiation me-dium with RA was changed every 2 days, for 2–3 weeks, toprevent cells from adhering to the plates. Cells were transferredto cell culture dishes for adhesion, and differentiation mediumwas changed every 2–3 days until cells reached 100% confluence(7–10 days). Cells were trypsinized and plated onto poly-D-ly-sine plates in medium containing 5% FBS and mitotic inhibitors(10 lM FUdR, 10 lM Urd, 10 lM araC) for 1–2 weeks(replate I). Low FBS and mitotic inhibitors reduce nonneuronalcell proliferation, and poly-D-lysine allows the neuronal cells toadhere to the cell culture plates without relying on the non-neuronal cells. To enrich further for pure neuronal cells, the cellswere selectively trypsinized and replated on poly-D-lysine- andMatrigel matrix-coated plates (replate II). Neurons were main-tained in medium with 5% FBS and mitotic inhibitors, and me-dium was replaced twice per week. Neurons could be main-tained for approximately 2 months.
Modified SK-N-SH Cell Adhesion DifferentiationProtocol
This protocol was modified from the previously reportedcell aggregation method (Paquet-Durand et al., 2003). Cells (4–5 3 105) per 10-cm cell culture dish were plated in MEM sup-plemented with 2 mM L-glutamine, 1% penicillin/streptomycinand antimycotic, 1 mM sodium pyruvate, and 10% FBS. On day2, 10 lM RA was added, and cells were kept in this differentia-tion medium for 1–2 weeks until they reached 100% confluenceor until the end of the 2-week differentiation period. Differen-tiation medium was changed every 2–3 days. Once cells areconfluent, they must be replated because neuronal loss may oc-cur as a result of the continous proliferation of nonneuronalcells. Two weeks is sufficient for the neuronal differentiation ofSK-N-SH cells (Kraveka et al., 2003). The neuronal cells wereselectively trypsinized, replated on poly-D-lysine-coated plates,and grown in medium containing 5% FBS, 10 lM RA, and mi-totic inhibitors for 2 weeks to prevent the proliferation of undif-ferentiated nonneuronal cells (replate I). To maintain the neuro-
84 Jain et al.
Journal of Neuroscience Research DOI 10.1002/jnr

nal cells in culture for extended periods and obtain a highly purepopulation of neuronal cells, the cells were selectively trypsi-nized and replated on poly-D-lysine- and Matrigel matrix-coated cell culture plates in medium containing 5% FBS and mi-totic inhibitors in the absence of RA (replate II).
Neuronal Cultures
Human primary neurons were cultured as described previ-ously (LeBlanc et al., 1998). The McGill Institutional ReviewBoard approved the protocol. Briefly, 12–16-week-old fetalbrains were dispersed with trypsin, treated with deoxyribonucle-ase I, filtered through 130- and 70-mm nylon mesh, and platedon poly-D-lysine-coated cell culture plates at a density of 3 3106 cells/ml. The cultures contain approximately 90% neurons and10% astrocytes. Microglial cells were eliminated with FudR. Theexperiments were performed after 10 days of neuronal culture.
Reverse Transcriptase-Polymerase Chain Reaction
RNA was isolated with Trizol (Invitrogen) according to themanufacturer’s instructions. Reverse transcription of the RNAswas followed by PCR. The following primers were used: forhTR, F3B 50-TCTAACCCTAACTGAGAAGGGCGTAG-30and R3C 50-GTTTGCTCTAGAATGAACGGTGGA-30 (Fenget al., 1995); for hTERT, TERT-1748S 50-CGGAAGAGT-GTCTGGAGCAA-30 and TERT-1928A 50-GGATGAAGC-GGAGTCTGGA-30 (Ulaner et al., 1998); for human GAPDH,RT11 50-CGGAGTCAACGGATTTGGTCGTAT-30 and RT1250TGCTAAGCAGTTGGTGGTGCAGGA-30.
For the neuronal markers Ngn1, NeuroD, GluR, andMash1, we used previously described primers (Megiorni et al.,2005). PCR was performed by using standard conditions withthe following annealing temperatures: for Ngn1 548C, for Neu-roD and GluR 588C, for Mash1 658C. For tau and Map2 weused previously described primers (McLaughlin et al., 2006).PCR was performed by using standard conditions with the fol-lowing annealing temperatures: for tau 548C and for MAP2568C.
Telomerase Assays
Whole-cells extracts were prepared by detergent lysis andassayed by the PCR-based telomeric repeat amplification pro-tocol (TRAP; Kim et al., 1994) using 0.5–1 lg protein extract.
RESULTS AND DISCUSSION
The identification and characterization of cultured humancell lines that can be induced to differentiate into neuronal cellsare essential to validate them as relevant experimental systems ormodels and also to establish these differentiated cells as a conven-ient and readily available neuronal source.
Establishment of NT2 and SK-N-SH CellDifferentiation
Several protocols for the differentiation of NT-2 cellshave been established. The time required for differentiationusing cell adhesion-based protocols is between 44 and 56 days(Pleasure et al., 1992; Fig. 1A). Neural stem cells can proliferatein suspension as cell clusters termed neurospheres, also called con-
glomerates (Reynolds and Weiss, 1992). Therefore, cell aggrega-tion protocols have been reported for NT2 cells. One such cellaggregation method has the advantage of reducing the time ofdifferentiation, but it also reduces the yield of neurons (Cheunget al., 1999). Most recently, a modified differentiation cellaggregation protocol using floating sphere or conglomerate cul-ture was reported that reduces the time needed for differentia-tion to 24 days, without compromising the quantity of theneurons obtained (Paquet-Durand et al., 2003; Fig. 1B). Weused a cell adhesion protocol provided by the manufacturerfrom which we obtained the NT2 cells (Stratagene; Pleasureet al., 1992; Fig. 1A) and developed a modified version of the
Fig. 1. Differentiation methods. A: The original NT2 cell adhesionmethod (Pleasure et al., 1992) is compared with the NT2 cell aggrega-tion method (B; Paquet-Durand et al., 2003) and with the NT2 modi-fied cell aggregation method (C; this paper). D: The SK-N-SH celladhesion method (Kraveka et al. 2003) is compared with the SK-N-SH modified cell adhesion method (E; this paper). RA, 10 lM reti-noic acid; FBS, fetal bovine serum; CM, conditioned media; MI, mi-totic inhibitors. See text for details.
Telomerase and Neuronal Markers in Human Neuronal Cells 85
Journal of Neuroscience Research DOI 10.1002/jnr

most recently reported cell aggregation protocol (Paquet-Durand et al., 2003; Fig. 1C).
We modified several steps in the cell aggregation proto-col (Paquet-Durand et al., 2003; compare Fig. 1B and C). Inthe first step, we increased the time of incubation in mediumcontaining RA from 7–10 days to 2–3 weeks. We found thatthe appearance of neuronal-like cells increased when the float-ing cell conglomerate culture period was prolonged. The dura-tion of the second step, which includes seeding the conglomer-ates onto cell culture plates, was lengthened from 6–8 days to7–10 days. Step 3 (replate I) in which conditioned medium isobtained [to be used in step 5 (replate III)] was omitted,because we found that the cells do not remain healthy in con-ditioned medium. The next step (replate I) was similar toreplate II in the cell aggregation protocol in which cells aregrown in media with low FBS (5% instead of 10%) and mitoticinhibitors (FUdR, Urd, araC) for 1–2 weeks. However, at thisstep, we also grew the cells on plates coated with poly-D-ly-sine, because the neuronal cells adhere more efficiently tocoated plates (used only at replate III in the cell aggregationprotocol). Finally, the cells were transferred to plates coatedwith poly-D-lysine and Matrigel matrix and maintained in mi-totic inhibitor (replate II), whereas, in the cell aggregation pro-tocol, the mitotic inhibitors were removed upon the last platingon poly-D-lysine and Matrigel matrix. We maintained the cellsin mitotic inhibitors, because we found that it prevented anyremaining undifferentiated cells from overgrowing the neuronalcell population. When grown on poly-D-lysine and Matrigelmatrix, the differentiated cells aggregated to form ganglion-likecell clusters (Fig. 2Ac). Cells showed neuronal morphologywith neuritic extensions. As previously reported for the cellaggregation protocol, we obtained approximately 95% NT2neuronal cells (Paquet-Durand et al., 2003; data not shown).We maintained these neurons for approximately 2 months incell culture (Fig. 2Ab,c).
Few studies have reported the use of differentiated SK-N-SH neuronal cells. We were also interested in investigatingthese cells as a readily available source of neuronal cells. Whenexposed to RA, SK-N-SH cells can differentiate to a neuronalphenotype (Gaitonde et al., 2001; Wainwright et al., 2001; Pizziet al., 2002; Kraveka et al., 2003). However, RA treatment hasbeen reported to lead either exclusively to differentiation intoepithelial-like cells or typically to a mixture of neuronal type (N-type) and substrate-adherent fibroblast or epithelial type (S-type)cells (Sidell et al., 1983; Ciccarone et al., 1989). Differentiationof SK-N-SH cells is commonly performed by growth for 2weeks in medium containing RA (Fig. 1D). Using this method,we obtained few neuronal cells, mixed with a population of epi-thelial-like cells. We investigated the possibility of using the NT2cell aggregation protocol to differentiate SK-N-SH cells (Fig.1C). However, unlike NT2 cells and neural stem cells, whichproliferate in suspension, SK-N-SH cells are dependent on adhe-sion for proliferation. Nonetheless, we successfully adapted cer-tain steps of the cell aggregation protocol into a modified cell ad-hesion protocol for the differentiation of SK-N-SH cells. SK-N-SH cells were initially maintained in medium containing RA for1–2 weeks (Fig. 1E). Neuronal cells were selectively trypsinizedand subsequently maintained on poly-D-lysine-coated plateswith RA in medium with 5% FBS and mitotic inhibitors for
2 weeks (replate I). This step allows the neuronal cells to growand form a network without relying on the secondary populationof cells that typically proliferates in the absence of mitotic inhibi-tors. Finally, neuronal cells were grown on poly-D-lysine- andMatrigel matrix-coated plates without RA in the presence of mi-totic inhibitors (replate II). This protocol led to neuronal cellsessentially devoid of the nonneuronal cell type (Fig. 2Bb,c; datanot shown). SK-N-SH cells showed neuronal morphology withneuritic extensions (Fig. 2Bc). These differentiated neuronal SK-N-SH cells could be maintained in culture for 2 months.
Differentiated NT2 and SK-N-SH Neuronal Cells andPrimary Human Fetal Neurons Express Neural BasicHelix-Loop-Helix Genes and the Neuronal Markers Tau,Glu, and Map2
To ascertain the relevance of the differentiated NT2 andSK-N-SH neuronal cells as models of human neurons, we eval-uated the expression of several basic helix-loop-helix (bHLH)transcription factors that are expressed specifically during the
Fig. 2. Induction of neuronal morphology in NT2 and SK-N-SH celllines. A: NT2 cells. a: Proliferating NT2 cells. b: NT2 cells differenti-ated by using the cell adhesion method (Pleasure et al., 1992). c: NT2cells differentiated by using the modified cell aggregation method (thispaper). The differentiated cells aggregated to form ganglion-like cellclusters and showed neuronal morphology with neuritic extensions.B: SK-N-SH cells. a: Proliferating SK-N-SH cells. b: SK-N-SH cellsdifferentiated by using the cell adhesion method (Kraveka et al., 2003).c: SK-N-SH cells differentiated by using the modified cell adhesionmethod (this paper). Neuronal cells were essentially devoid of thenonneuronal cell type and showed neuronal morphology with neuriticextensions.
86 Jain et al.
Journal of Neuroscience Research DOI 10.1002/jnr
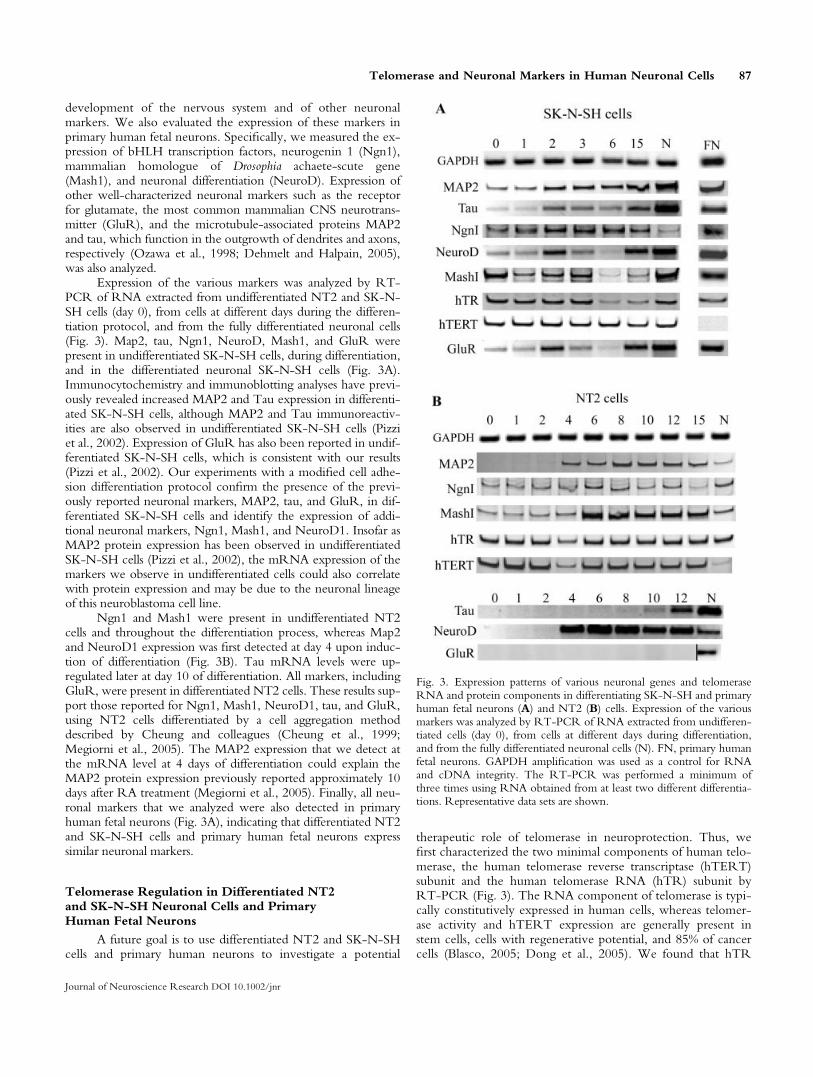
development of the nervous system and of other neuronalmarkers. We also evaluated the expression of these markers inprimary human fetal neurons. Specifically, we measured the ex-pression of bHLH transcription factors, neurogenin 1 (Ngn1),mammalian homologue of Drosophia achaete-scute gene(Mash1), and neuronal differentiation (NeuroD). Expression ofother well-characterized neuronal markers such as the receptorfor glutamate, the most common mammalian CNS neurotrans-mitter (GluR), and the microtubule-associated proteins MAP2and tau, which function in the outgrowth of dendrites and axons,respectively (Ozawa et al., 1998; Dehmelt and Halpain, 2005),was also analyzed.
Expression of the various markers was analyzed by RT-PCR of RNA extracted from undifferentiated NT2 and SK-N-SH cells (day 0), from cells at different days during the differen-tiation protocol, and from the fully differentiated neuronal cells(Fig. 3). Map2, tau, Ngn1, NeuroD, Mash1, and GluR werepresent in undifferentiated SK-N-SH cells, during differentiation,and in the differentiated neuronal SK-N-SH cells (Fig. 3A).Immunocytochemistry and immunoblotting analyses have previ-ously revealed increased MAP2 and Tau expression in differenti-ated SK-N-SH cells, although MAP2 and Tau immunoreactiv-ities are also observed in undifferentiated SK-N-SH cells (Pizziet al., 2002). Expression of GluR has also been reported in undif-ferentiated SK-N-SH cells, which is consistent with our results(Pizzi et al., 2002). Our experiments with a modified cell adhe-sion differentiation protocol confirm the presence of the previ-ously reported neuronal markers, MAP2, tau, and GluR, in dif-ferentiated SK-N-SH cells and identify the expression of addi-tional neuronal markers, Ngn1, Mash1, and NeuroD1. Insofar asMAP2 protein expression has been observed in undifferentiatedSK-N-SH cells (Pizzi et al., 2002), the mRNA expression of themarkers we observe in undifferentiated cells could also correlatewith protein expression and may be due to the neuronal lineageof this neuroblastoma cell line.
Ngn1 and Mash1 were present in undifferentiated NT2cells and throughout the differentiation process, whereas Map2and NeuroD1 expression was first detected at day 4 upon induc-tion of differentiation (Fig. 3B). Tau mRNA levels were up-regulated later at day 10 of differentiation. All markers, includingGluR, were present in differentiated NT2 cells. These results sup-port those reported for Ngn1, Mash1, NeuroD1, tau, and GluR,using NT2 cells differentiated by a cell aggregation methoddescribed by Cheung and colleagues (Cheung et al., 1999;Megiorni et al., 2005). The MAP2 expression that we detect atthe mRNA level at 4 days of differentiation could explain theMAP2 protein expression previously reported approximately 10days after RA treatment (Megiorni et al., 2005). Finally, all neu-ronal markers that we analyzed were also detected in primaryhuman fetal neurons (Fig. 3A), indicating that differentiated NT2and SK-N-SH cells and primary human fetal neurons expresssimilar neuronal markers.
Telomerase Regulation in Differentiated NT2and SK-N-SH Neuronal Cells and PrimaryHuman Fetal Neurons
A future goal is to use differentiated NT2 and SK-N-SHcells and primary human neurons to investigate a potential
therapeutic role of telomerase in neuroprotection. Thus, wefirst characterized the two minimal components of human telo-merase, the human telomerase reverse transcriptase (hTERT)subunit and the human telomerase RNA (hTR) subunit byRT-PCR (Fig. 3). The RNA component of telomerase is typi-cally constitutively expressed in human cells, whereas telomer-ase activity and hTERT expression are generally present instem cells, cells with regenerative potential, and 85% of cancercells (Blasco, 2005; Dong et al., 2005). We found that hTR
Fig. 3. Expression patterns of various neuronal genes and telomeraseRNA and protein components in differentiating SK-N-SH and primaryhuman fetal neurons (A) and NT2 (B) cells. Expression of the variousmarkers was analyzed by RT-PCR of RNA extracted from undifferen-tiated cells (day 0), from cells at different days during differentiation,and from the fully differentiated neuronal cells (N). FN, primary humanfetal neurons. GAPDH amplification was used as a control for RNAand cDNA integrity. The RT-PCR was performed a minimum ofthree times using RNA obtained from at least two different differentia-tions. Representative data sets are shown.
Telomerase and Neuronal Markers in Human Neuronal Cells 87
Journal of Neuroscience Research DOI 10.1002/jnr

was expressed in proliferating and differentiated SK-N-SH andNT2 cells and in primary human fetal neurons (Fig. 3). Theconstitutive presence of hTR in differentiated cells is consistentwith previously published reports (Dong et al., 2005).
Expression of hTERT was present in SK-N-SH andNT2 cells and subsequently down-regulated during differentia-tion of NT2 cells (Fig. 3). hTERT levels were present in pro-liferating, diffentiating, and differentiated SK-N-SH cells andabsent in primary human fetal neurons (Fig. 3). This is consist-ent with previous findings that most human somatic cells con-tain no or low hTERT levels (Masutomi et al., 2003); how-ever, this is the first description of the hTERT status of primaryhuman fetal neurons. We found telomerase activity to bedown-regulated during SK-N-SH and NT2 cell differentiationand absent in primary human fetal neurons (Fig. 4). Completedown-regulation of the telomerase activity of NT2 cell differ-entiated by the cell adhesion protocol has previously beenreported, although proliferating NT2 cells appeared to haveinitially low telomerase activity (Kruk et al., 1996). Interest-ingly, by using the modified cell adhesion protocol, we foundthat telomerase activity in SK-N-SH cells can be more com-pletely repressed than that previously reported after 10 days oftreatment with 10 lM RA (Kraveka et al., 2003), although wealso occasionally observed low levels of telomerase remainingin the differentiated neuronal SK-N-SH cells. Low levels oftelomerase activity in differentiated cell populations could bedue to the low numbers of undifferentiated cells still present in
cultures. The down-regulation of telomerase activity in differ-entiated NT2 cells correlates with the low levels of hTERTmRNA in these cells (Fig. 3). Because differentiated NT2 andSK-N-SH cells and primary human fetal neurons exhibit no orlow levels of telomerase activity, they could be utilized toinvestigate the protective effects of overexpressed or up-regu-lated telomerase (Fu et al., 2000; Haik et al., 2000; Misiti et al.,2000; Mattson and Klapper, 2001).
The goal of our study was to develop modified NT2 andSK-N-SH cell differentiation methods and investigate theexpression of telomerase activity, core telomerase componentshTERT and hTR, and neuronal markers in these cells and pri-mary human fetal neurons. We report modified protocols thatincrease the number of differentiated NT-2 and SK-N-SH cellsand establish an enriched neuronal SK-N-SH-derived cell popu-lation essentially devoid of nonneuronal cells. Our results indi-cate that the telomerase and neuronal marker statuses of differen-tiated NT2 and SK-N-SH cells and of primary human fetal neu-rons are similar. Thus these differentiated cell lines, in parallelwith primary human fetal neurons, may constitute excellentsources for studies to investigate the role of telomerase or othersurvival-promoting activities in protecting human neuronal cellsfrom cell death-mediating stresses that can lead to neurodegener-ative diseases.
ACKNOWLEDGMENTS
We thank Jennifer Hammond for the technical prepara-tion of the primary human neuron cultures. C.A. and A.C.L.are Chercheur Boursier and Chercheur National of Le Fondsde la Recherche on Sante du Quebec, respectively.
REFERENCES
Andrews PW. 1984. Retinoic acid induces neuronal differentiation of a
cloned human embryonal carcinoma cell line in vitro. Dev Biol 103:
285–293.
Barlow C, Treuner K. 2005. DNA instability in the brain: survival of the
‘‘fittest.’’ Nat Med 11:474–475.
Becker EBE, Bonni A. 2005. Beyond proliferation—cell cycle control of
neuronal survival and differentiation in the developing mammalian brain.
Semin Cell Dev Biol 16:439–448.
Blasco MA. 2005. Telomeres and human disease: ageing, cancer and
beyond. Nat Rev Genet 6:611–622.
Calado RT, Chen J. 2006. Telomerase: not just for the elongation of telo-
meres. Bioessays 28:109–112.
Callen E, Surralles J. 2004. Telomere dysfunction in genome instability
syndromes. Mutat Res 567:85–104.
Cheung WMW, Fu WY, Hui WS, Ip NY. 1999. Production of human
CNS neurons from embryonal carcinoma cells using a cell aggregation
method. Biotechniques 26:946–954.
Ciccarone V, Spengler BA, Meyers MB, Biedler JL, Ross RA. 1989. Phe-
notypic diversification in human neuroblastoma cells: expression of dis-
tinct neural crest lineages. Cancer Res 49:219–225.
Dehmelt L, Halpain S. 2005. The MAP2/Tau family of microtubule-asso-
ciated proteins. Genome Biol 6:204–214.
Dong CK, Masutomi K, Hahn WC. 2005. Telomerase: regulation, func-
tion and transformation. Crit Rev Oncol Hematol 54:85–93.
Feng J, Funk WD, Wang S-S, Weinrich SL, Avilion AA, Chiu C-P,
Adams RR, Chang E, Allsopp RC, Yu J, Le S, West MD, Harley CB,
Andrews WH, Greider CW, Villeponteau B. 1995. The RNA compo-
nent of human telomerase. Science 269:1236–1241.
Fig. 4. Telomerase activity in differentiating SK-N-SH and NT2 cellsand in primary human fetal neurons. Telomerase activity was analyzedby TRAP of cellular extracts prepared from undifferentiated cells (day0), from cells at different days during differentiation, and from the fullydifferentiated neuronal cells (N). For SK-N-SH and NT2 cells, 1 lgof protein extract was assayed. N1 and N2 represent two independentpreparations of primary human fetal neurons; 0.5 and 1 lg of proteinextract was assayed. IC, internal PCR control to ensure that absenceof telomerase activity is not due to inhibition of the PCR step of thetelomerase assay.
88 Jain et al.
Journal of Neuroscience Research DOI 10.1002/jnr

Fu W, Killen M, Culmsee C, Dhar S, Pandita TK, Mattson MP. 2000. The
catalytic subunit of telomerase is expressed in developing brain neurons
and serves a cell survival-promoting function. J Mol Neurol 14:3–15.
Gaitonde SV, Qi W, Falsey RR, Sidell N, Martinez JD. 2001. Morpho-
logic conversion of a neuroblastoma-derived cell line by E6-mediated
p53 degradation. Cell Growth Differ 12:19–27.
Greene LA, Tischler AS. 1976. Establishment of a noradrenergic clonal
line of rat adrenal pheochromocytoma cells which respond to nerve
growth factor. Proc Natl Acad Sci U S A 73:2424–2428.
Haik S, Gauthier LR, Granotier C, Peyrin J-M, Lages CS, Dormont D,
Boussin FD. 2000. Fibroblast growth factor 2 up regulates telomerase ac-
tivity in neural precursor cells. Oncogene 19:2957–2966.
Kim NW, Piatyszek MA, Prowse KR, Harley CB, West MD, Ho PLC,
Coviello GM, Wright WE, Weinrich SL, Shay JW. 1994. Specific asso-
ciation of human telomerase activity with immortal cells and cancer. Sci-
ence 266:2011–2015.
Kraveka JM, Li L, Bielawski J, Obeid LM, Ogretman B. 2003. Involve-
ment of endogenous ceramide in the inhibition of telomerase activity
and induction of morphologic differentiation in response to all-trans-
retinoic acid in human neuroblastoma cells. Arch Biochem Biophys 419:
110–119.
Kruk PA, Blajee AS, Rao KS, Bohr VA. 1996. Telomere reduction and
telomerase inactivation during neuronal cell differentiation. Biochem
Biophys Res Commun 224:487–492.
LeBlanc AC. 1995. Increased production of 4 kDa amyloid b peptide in
serum deprived human primary neuron cultures: possible involvement of
apoptosis. J Neurosci 15:7837–7846.
LeBlanc AC. 2005. The role of apoptotic pathways in Alzheimer’s disease
neurodegeneration and cell death. Curr Azheimer Res 2:389–402.
LeBlanc AC, Koutroumanis M, Goodyer C. 1998. Protein kinase C acti-
vation increases release of secreted amyloid precursor protein without
decreasing Ab production in human primary neuron cultures. J Neuro-
science 18:2907–2913.
Lombard DB, Chua KF, Mostoslavsky R, Franco S, Gostissa M, Alt FW.
2005. DNA repair, genome stability, and aging. Cell 120:497–512.
Lu C, Fu W, Mattson MP. 2001. Telomerase protects developing neurons
against DNA damage-induced cell death. Brain Res Dev Brain Res
131:167–171.
Masutomi K, Yu EY, Khurts S, Ben-Porath I, Currier JL, Metz GB,
Brooks MW, Kaneko S, Murakami S, DeCaprio JA, Weinberg RA,
Stewart SA, Hahn WC. 2003. Telomerase maintains telomere structure
in normal human cells. Cell 114:241–253.
Mattson MP, Klapper W. 2001. Emerging roles for telomerase in neuronal
development and apoptosis. J Neurosci Res 63:1–9.
Mattson MP, Magnus T. 2006. Ageing and neuronal vulnerability. Nat
Rev Neurosci 7:278–294.
Mattson MP, Barger SW, Begley JG, Mark RJ. 1995. Calcium, free radi-
cals, and excitotoxic neuronal death in primary cell culture. Methods
Cell Biol 46:187–216.
McLaughlin D, Tsirimonaki E, Vallianatos G, Sakellaridis N, Chatzistama-
tiou T, Stavropoulou-Gioka C, Tsezou A, Messinis I, Mangoura D.
2006. Stable expression of a neuronal dopaminergic progenitor pheno-
type in cell lines derived from human amniotic fluid cells. J Neurosci
Res 83:1190–1200.
Megiorni F, Mora B, Indovina P, Mazzilli MC. 2005. Expression of neu-
ronal markers during NTera2/cloneD1 differentiation by cell aggregation
method. Neurosci Lett 373:105–109.
Misiti S, Anni SN, Fontemaggi G, Cong YS, Wen J, Hirte HW, Piaggio
G, Sacchi A, Pontecorvi A, Bacchetti S, Farsetti A. 2000. Induction of
hTERT expression and telomerase activity by estrogens in human ovary
epithelium cells. Mol Cell Biol 20:3764–3771.
Nouspikel T, Hanawalt PC. 2002. DNA repair in terminally differentiated
cells. DNA Repair 1:59–75.
Ostenfeld T, Caldwell MA, Prowse KR, Linskens MH, Jauniaux E,
Svendsen CN. 2000. Human neural precursor cells express low levels of
telomerase in vitro and show diminishing cell proliferation with exten-
sive axonal outgrowth following transplantation. Exp Neurol 164:215–
226.
Ozawa S, Kamiya H, Tsuzuki K. 1998. Glutamate receptors in the mam-
malian central nervous system. Prog Neurobiol 54:581–618.
Paquet-Durand F, Tan S, Bicker G. 2003. Turning teratocarcinoma cells
into neurons: rapid differentiation of NT-2 cells in floating spheres.
Brain Res Dev Brain Res 142:161–167.
Pizzi M, Boroni F, Bianchetti A, Moraitis C, Sarnico I, Benarese M, Goffi F,
Valerio A, Spano P. 2002. Expression of functional NR1/NR2B-type
NMDA receptors in neuronally differentiated SK-N-SH human cell line.
Eur J Neurol 16:2342–2350.
Pleasure SJ, Page C, Lee VM-Y. 1992. Pure, postmitotic, polarized human
neurons derived from NTera 2 cells provide a system for expressing ex-
ogenous proteins in terminally differentiated neurons. J Neurosci 12:
1802–1815.
Przyborski SA, Morton IE, Wood A, Andrews PW. 2000. Developmental
regulation of neurogenesis in the pluripotent human embryonal carci-
noma cell line NTERA-2. Eur J Neurosci 12:3521–3528.
Reali C, Scintu F, Pillai R, Cabras S, Argiolu F, Ristaldi MS, Sanna MA,
Badiali M, Sogos V. 2006. Differentiation of human adult CD34+ stem
cells into cells with a neural phenotype: role of astrocytes. Exp Neurol
197:399–406.
Reynolds BA, Weiss S. 1992. Generation of neurons and astrocytes from
isolated cells of the adult mammalian central nervous system. Science
255:1707–1709.
Sidell N, Altman A, Haussler MR, Seeger RC. 1983. Effects of retinoic
acid (RA) on the growth and phenotypic expression of several human
neuroblastoma cell lines. Exp Cell Res 148:21–30.
Smogorzewska A, de Lange T. 2004. Regulation of telomerase by telo-
meric proteins. Annu Rev Biochem 73:177–208.
Troen BR. 2003. The biology of aging. Mount Sinai J Med 70:3–22.
Trojanowski JQ, Kleppner SR, Hartley RS, Miyazono M, Fraser NW,
Kesari S, Lee VM-Y. 1997. Transfectable and transplantable postmitotic
human neurons: a potential ‘‘platform’’ for gene therapy of nervous sys-
tem diseases. Exp Neurol 144:92–97.
Ulaner GA, Hu J-F, Vu TH, Giudice LC, Hoffman AR. 1998. Telomerase
activity in human development is regulated by human telomerase reverse
transcriptase (hTERT) and by alternate splicing of hTERT transcripts.
Cancer Res 58:4168–4172.
Villa A, Navarro-Galve B, Bueno C, Franco S, Blasco MA, Martinez-
Serrano A. 2004. Long-term molecular and cellular stability of human
neural stem cell lines. Exp Cell Res 294:559–570.
Wainwright LJ, Lasorella A, Iavarone A. 2001. Distinct mechanisms of cell
cycle arrest control the decision between differentiation and senescence
in human neuroblastoma cells. Proc Natl Acad Sci U S A 98:9396–9400.
Wright WE, Shay JW. 2000. Telomere dynamics in cancer progression
and prevention: fundamental differences in human and mouse biology.
Nat Med 6:849–851.
Wright WE, Prowse KR, Wallace K, Linskens MHK, Svendsen CN. 2006.
Human progenitor cells isolated from the developing cortex undergo
decreased neurogenesis and eventual senescence following expansion in
vitro. Exp Cell Res Apr 20 [E-pub ahead of print].
Zhu H, Fu W, Mattson MP. 2000. The catalytic subunit of telomerase
protects neurons against amyloid b-peptide-induced apoptosis. J Neuro-
chem 75:117–124.
Telomerase and Neuronal Markers in Human Neuronal Cells 89
Journal of Neuroscience Research DOI 10.1002/jnr


![Clinical feasibility for cell therapy using human neuronal ... · Development of Human hNT2.17 Cell Line We subcloned a human neuronal cell line from the parental NT2 cell line [33]](https://img.pdfslide.net/doc/110x75/5e85441a5f8b2528e12f70b2/clinical-feasibility-for-cell-therapy-using-human-neuronal-development-of-human.jpg)


























