Embed Size (px)
Citation preview

0
UNIVERSIDADE FEDERAL DE PELOTAS
Programa de Pós-Graduação em Fisiologia Vegetal
TESE
Cultivo in vitro de Pyrus sp.,
cultivares Carrick, Cascatense e Ya-Li.
ILDA MARICLEI DE CASTRO DA SILVA
Pelotas, 2013

1
ILDA MARICLEI DE CASTRO DA SILVA
Cultivo in vitro de Pyrus sp.,
cultivares Carrick, Cascatense e Ya-Li.
Orientador: Prof. Dr. José Antonio Peters
Co-orientadores: Dr. Leonardo Ferreira Dutra
Prof. Dr. Valmor João Bianchi
Profa. Dra. Eugenia Jacira Bolacel Braga
Pelotas, 2013
Tese apresentada à Universidade Federal de Pelotas, sob a orientação do Prof. Dr. José Antonio Peters, como parte das exigências do Programa de Pós-Graduação em Fisiologia Vegetal, para a obtenção do título de Doutor em Fisiologia Vegetal.

Dados de catalogação na fonte: Ubirajara Buddin Cruz - CRB 10/901 Biblioteca de Ciência & Tecnologia - UFPel
S586c Silva, Ilda Mariclei de Castro da
Cultivo in vitro de Pyrus sp., cultivares Carrick, Cascatense e Ya-Li / lda Mariclei de Castro da Silva. – 85f. ; il. – Tese (Doutorado). Programa de Pós-Graduação em Fisiologia Ve-getal. Universidade Federal de Pelotas. Instituto de Biologia. Pelotas, 2013. – Orientador José Antonio Peters; co-orientador Leonardo Ferreira Dutra, Valmor João Bianchi e Eugenia Jacira Bolacel Braga.
1.Fisiologia vegetal. 2.Pereiras. 3. Micropropagação.
4.Reguladores de crescimento. 5.Radiação gama. 6.Mutação. I.Peters, José Antonio. II.Dutra, Leonardo Ferreira. III.Bianchi, Valmor João. IV.Braga, Eugenia Jacira Bolacel. V.Título.
CDD: 634.13

2
ILDA MARICLEI DE CASTRO DA SILVA
Banca examinadora:
Prof. Dr. José Antonio Peters – Orientador
Dr. Paulo Celso de Mello Farias – Universidade Federal de Pelotas
Dra. Daiane Peixoto Vargas – Embrapa Clima Temperado
Dra. Ilisandra Zanandrea – Universidade Federal de Pelotas

“...Tenha fé em Deus
Tenha fé na vida
Tente outra vez!...
...Queira!
Basta ser sincero e desejar profundo
Você será capaz de sacudir o mundo
Tente outra vez!...
...Tente
E não diga que a vitória está perdida
Se é de batalhas que se vive a vida
Tente outra vez!...” Raul Seixas
Tente Outra vez

A Deus,
Aos meus avós Marcos (In Memoriam) e Ilda, A meu sobrinho Lorenzo
Ofereço
A meus pais Nelson e Maria José Com carinho,
Dedico

5
AGRADECIMENTOS
A Deus, por tudo! Por iluminar meu caminho, dar forças e coragem para lutar
todos os dias.
À Universidade Federal de Pelotas, em particular, ao Programa de Pós-
Graduação em Fisiologia Vegetal, pela oportunidade de realizar o curso.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES),
pela concessão da bolsa de estudos.
À Embrapa Clima Temperado, pela possibilidade de realização do projeto,
disponibilização do material vegetal e recursos para a realização dos experimentos.
Ao Centro de Oncologia do Departamento de Radiologia da Faculdade de
Medicina da Universidade Federal de Pelotas, pela irradiação do material vegetal.
Ao Professor Dr. José Antonio Peters, pela orientação, ensinamentos e
disposição, e acima de tudo, pelo apoio e carinho, sendo sempre um „paizão‟.
Ao Professor Dr. Valmor João Bianchi, pela co-orientação, ensinamentos,
sugestões e amizade.
À Professora Dra. Eugenia Jacira Bolacel Braga, pela co-orientação,
sugestões, incentivo e simpatia.
Ao Dr. Leonardo Ferreira Dutra, pela co-orientação, carinho, incentivo e
confiança.
Ao Professor Dr. Willian Barros, pelo apoio estatístico.
A minha amiga, colega de curso e de trabalho, Cristina, pela amizade, parceria,
convivência, conversas, ajuda e „indiadas‟.
A minha amiga Aline, pela grande amizade, conversas, risadas, ajuda, estudos,
momentos que ficarão para sempre guardados.
A minha amiga Janieli, que, mesmo não estando junto, está sempre presente
em meu coração e nas lembranças dos bons momentos durante o mestrado.
Aos estagiários Mara Cintia Winhelmann e Maurício Flores, pela preciosa ajuda
durante a realização dos experimentos.
Às colegas Alítcia e Elisia, pelos momentos de distração, risadas, „papos
cabeça‟ e estudos.
Aos colegas do Laboratório de Cultura de Tecidos de Plantas, no período
desses seis anos, pela convivência e amizades criadas.

Ao pessoal do Laboratório de Cultura de Tecidos da Embrapa, em especial
Nino, Osmi, Falcão, Daiane e Fernanda Thiel.
Ao Pesquisador da Embrapa Clima Temperado José Francisco Pereira, pela
atenção, simpatia, sugestões e auxílio.
Ao Professor Mário Ariel Posada, pela motivação em seguir em frente e buscar,
em Pelotas, os cursos de mestrado e doutorado.
À amiga Cristina Larré, pelo auxílio, troca de ideias e estímulo.
À amiga Claudia Lima, pelo incentivo e conversas.
À secretária do curso, Sandra Sacco da Silva, pela simpatia e disposição de
sempre.
A minha família, meu porto seguro, pra onde eu sei que posso voltar sempre e
terei amor, carinho e incentivo, em especial minha vó Ilda, pelas „rezas‟ e chás,
carinho e ensinamentos; e meus pais, Nelson e Maria José, por terem sempre uma
palavra de apoio, incentivo, coragem; pelo amor imensurável, confiança, orgulho e
por acreditarem em mim.
A minha irmã Juliana, pelo amor, cumplicidade, amizade e incentivo; meu
cunhado, Jiuliano pelo carinho, estímulo e por ser um „irmão‟; e meu sobrinho
Lorenzo, por trazer muita alegria a minha vida.
À amiga e irmã de coração, Sandra, pelo estímulo, força e amizade em todas
as horas.
À amiga Gizele, pela presença constante, tornando meus dias menos solitários,
dividindo despesas, angústias e alegrias.
A minha amiga e comadre Tais, pela amizade e por ter me presenteado com
um afilhado tão mimoso como o João Victor.
A todos, citados aqui ou não, de perto ou de longe, que, de alguma forma,
colaboraram e apoiaram a conclusão desta etapa.

7
RESUMO
SILVA, I. M. de C. da. Cultivo in vitro de Pyrus sp., cultivares Carrick,
Cascatense e Ya-Li. 2013. 85f. Tese (Doutorado) – Programa de Pós-Graduação
em Fisiologia Vegetal. Universidade Federal de Pelotas/UFPel, Pelotas/RS.
No Brasil, entre as frutas de clima temperado, a pera (Pyrus sp.) é a terceira mais
consumida, porém, a maior parte é importada (95%), pois seu cultivo ainda é
pequeno. A falta de cultivares adaptadas às condições edafoclimáticas das regiões
potencialmente produtoras são os principais obstáculos à sua produção sustentável,
em vista que estas cultivares necessitam estar aptas a produzir em quantidade e
qualidade e com regularidade para suprir a demanda do mercado interno. Técnicas
de indução de mutação, como o uso de radiação gama, têm se mostrado eficientes
na produção de variabilidade genética e, por isso, muito utilizadas em programas de
melhoramento de várias espécies de plantas. Em pereira, devido a pouca
variabilidade genética, ciclos longos e alogamia, características estas que dificultam
a obtenção de novas cultivares através do melhoramento convencional, a utilização
da radiação gama pode suprir estas dificuldades. Esta técnica, acoplada à cultura de
tecidos é uma alternativa que permite o desenvolvimento de cultivares específicas,
em um curto espaço de tempo, em larga escala e com alta qualidade sanitária.
Porém, para viabilizar a utilização da propagação in vitro se faz necessária a
obtenção de protocolos de propagação. Neste contexto, o trabalho teve como
objetivo otimizar o cultivo in vitro de três cultivares de pereira (Pyrus sp.): Carrick,
Ya-Li e Cascatense (cultivar lançada recentemente pelo Programa de Melhoramento
Genético da Embrapa Clima Temperado), visando potencializar sua produção e a
indução de mutações, almejando selecionar novos genótipos com características de
interesse, a partir dos genótipos originais. O estudo foi conduzido em experimentos
que buscaram multiplicar in vitro as cultivares acima citadas, bem como irradiar e, a
partir disso, micropropagar a cv. Ya-Li. Para a realização dos experimentos foram
utilizados segmentos nodais (1,0 cm) contendo de duas a três gemas axilares.
Visando a obtenção de um protocolo eficiente de multiplicação in vitro, os explantes
foram inoculados em diferentes meios de cultivo e concentrações de sais, BAP e
sacarose. Para verificar os efeitos de diferentes doses de radiação gama (0, 10, 20,
30 e 40 Gy) durante a micropropagação, foram realizados experimentos de
multiplicação e enraizamento in vitro e aclimatização. Os resultados obtidos
evidenciaram que a multiplicação in vitro da cultivar Carrick pode ser potencializada

com o uso do meio MS Himédia comercial, com suplementação de 1,2 mg L-1 de
BAP e 30 g L-1 de sacarose, enquanto que o meio mais adequado para a
multiplicação in vitro da cultivar Ya-Li é o MS, com 1,2 mg L-1 de BAP e 45 g L-1 de
sacarose. Para a „Cascatense‟, o MS Himédia, suplementado de 0,8 mg L-1 de BAP
e 30 g L-1 de sacarose foi o que apresentou melhores respostas. Em relação ao uso
de irradiação, a radiação gama induziu alterações morfofisiológicas nos explantes da
cultivar Ya-Li, durante a micropropagação, sendo que estas podem ser devido a
variações epigenéticas e/ou genéticas (herdáveis). Desta forma, baseando-se nos
resultados do presente estudo, devido à obtenção de altas taxas de multiplicação in
vitro a partir da otimização dos protocolos, verifica-se que é possível multiplicar as
três cultivares em larga escala, bem como obter possíveis mutantes da cv. Ya-Li via
irradiação com raios gama. Porém, para determinar se as alterações
morfofisiológicas encontradas são positivas ou negativas e se transmitem pela
progênie, ainda se fazem necessárias pesquisas futuras nos materiais obtidos.
Palavras-chave: Pereiras. Micropropagação. Reguladores de crescimento.
Radiação gama. Mutação.

9
ABSTRACT
SILVA, I. M. de C. da. In vitro Cultivation of Pyrus sp., cultivars Carrick,
Cascatense and Ya-Li. 2013. 85f. Thesis (Doctorate) – Plant Physiology Post
Graduation Program. Federal University of Pelotas/UFPel, Pelotas/RS.
In Brazil, among the temperate climate fruit, the pear (Pyrus sp.) is the third more
commonly consumed although most of it is imported (95%) as its cultivation is still in
small-scale. The lack of cultivars adapted to the soil and climate conditions of
potentially productive regions is the main obstacle for its sustainable production, as
these cultivars need to be apt to produce in regularly, with a considerable quality and
amount to supply the demand of the of the internal market. Mutation induction
techniques, such as the use of gamma radiation, have been quite efficient in the
production of genetic variability and, thus, are very used in improvement programs of
several species of plants. In pear trees, due to the little genetic variability, long cycles
and cross-pollination, characteristics which make it more difficult to obtain new
cultivars through the conventional improvement, the usage of the gamma radiation
may fulfill these difficulties. This technique, along the tissue culture is an alternative
which enables the development of specific cultivars, in a short term, in large scale
and with a high health quality. However, to help the usage of in vitro propagation it is
necessary to obtain propagation protocols. In this context, the present work aimed to
optimize the in vitro and ex vitro cultivation of three pear cultivars (Pyrus sp.): Carrick,
Ya-Li and Cascatense (a cultivar recently launched by the Genetic Improvement
Program from Embrapa Temperate Climate), in order to foster its production and the
mutations induction, aiming to select new genotypes with interest characteristics,
from the original genotypes. The study was conducted in experiments which focused
on multiplying in vitro the cultivar mentioned above, as well as radiate and, from this,
micropropagation the cv. Ya-Li. For the carrying out of the experiments, nodal
segments were used (1,0 cm) with two to three axilliary gems. To reach an efficient
protocol of multiplication in vitro, the explants were inoculated in different cultivation
means and salt concentrations, BAP and sucrose. To check the effects of different
dosage of gamma radiation (0, 10, 20, 30 and 40 Gy) during the micropropagation,
were carried experiments out of multiplication and rooting in vitro and
aclimmatization. The results obtained showed that the multiplication in vitro of the
cultivar Carrick can be fostered by using MS commercial Himédia, with

supplementation of 1,2 mg L-1 of BAP and 30 g L-1 of sucrose, while the most
appropriate mean for the multiplication in vitro of the cultivar Ya-Li is the MS, with 1,2
mg L-1 of BAP and 45 g L-1 of sucrose. For the „Cascatense‟, the MS Himédia,
supplemented by 0,8 mg L-1 of BAP and 30 g L-1 of sucrose presented the best
responses. Concerning the use of irradiation, the gamma radiation led to
morphological changes in the explants of the cultivar Ya-Li, during the
micropropagation, being that these might be due to epigenetic and/or genetic
(inheritable) changes. Thus, based on the results of this study, due to obtaining high
rates of in vitro multiplication from the optimization of protocols, it appears that is
possible to multiply the three cultivars on a large scale, as well as possible to obtain
mutants of cv. Ya-Li via gamma radiation. However, to determine whether the
morphological changes found are positive or negative and is transmitted by the
progeny, although future research is needed in the materials obtained.
Key words: Pear trees. Micropropagation. Plant growth regulators. Gamma
radiation. Mutation.

11
LISTA DE FIGURAS
CAPÍTULO 1
Figura 1 Número de brotações por explante (A), comprimento das brotações (B), número de gemas axilares por brotação (C), massa fresca e massa seca das brotações (g) (D e E), respectivamente, de pereira, cvs. Ya-Li e Carrick, submetidos a diferentes meios de cultivo e concentrações de sais...................................................................................
27
Figura 2 Número de brotações por explante (A), comprimento das
brotações (B), número de gemas axilares por brotação (C), massa fresca e massa seca das brotações (g) (D e E), respectivamente, de pereira, cvs. Ya-Li e Carrick, submetidos a diferentes concentrações de BAP (mg L-1)....
29
Figura 3 Número de brotações por explante (A), comprimento das
brotações (B), número de gemas axilares por brotação (C), massa fresca e massa seca das brotações (g) (D e E), respectivamente, de pereira, cvs. Ya-Li e Carrick, submetidos a diferentes concentrações de sacarose (g L-
1)............................................................................................
31
Figura 4 Número de brotações por explante (A), comprimento das
brotações (B), número de gemas axilares por brotação (C), massa fresca e massa seca das brotações (g) (D e E), respectivamente, de pereira, cvs. Ya-Li e Carrick, submetidos a diferentes concentrações de BAP (mg L-1)....
34
Figura 5 Brotações de Pyrus sp., cvs. Ya-Li e Carrick, obtidas após
serem submetidas a diferentes meios de cultivo e concentrações de sais (A e B), concentrações de BAP (C, D, E e F) e concentrações de sacarose (G e H), respectivamente....................................................................
35

CAPÍTULO 2
Figura 1 Número de brotações por explante (A), comprimento das brotações (B), número de gemas axilares por brotação (C), massa fresca e seca das brotações (g) (D e E), respectivamente, de pereira, cv. Cascatense, submetidos a diferentes meios de cultivo e concentrações de sais........
50
Figura 2 Número de brotações por explante (A), comprimento das
brotações (B), número de gemas axilares por brotação (C), massa fresca e seca das brotações (g) (D e E), respectivamente, de pereira, cv. Cascatense, submetidos a diferentes concentrações de BAP (mg L-1)........................
52
Figura 3 Número de brotações por explante (A), comprimento das
brotações (B), número de gemas axilares por brotação (C), massa fresca e seca das brotações (g) (D e E), respectivamente, de pereira, cv. Cascatense, submetidos a diferentes concentrações de sacarose (g L-1)....................
54
Figura 4 Brotações de Pyrus sp., cv. Cascatense, obtidas após
serem submetidas a diferentes meios de cultivo e concentrações de sais (A), concentrações de BAP (B) e concentrações de sacarose (C)............................................
56
CAPÍTULO 3
Figura 1 Percentagem de explantes sobreviventes (A) e regenerantes (B), número de brotações por explante (C), número de gemas axilares por brotação (D) e comprimento das brotações (E) de pereira, cv. Ya-Li, submetidos a diferentes doses de radiação gama......................................
69
Figura 2 Percentagem de enraizamento (A), número de raízes por
planta (B), comprimento da maior raiz (cm) (C e D), percentagem de calos formados e de plantas sobreviventes após aclimatização (%) (E e F), em pereira, cv. Ya-Li, submetidos a diferentes doses de radiação gama.....................................................................................
71
Figura 3 Plantas irradiadas de Pyrus sp., cv. Ya-Li, após avaliações
de regeneração/multiplicação in vitro [com aspecto normal (A) e presença de caules espessos (B)], após enraizamento in vitro [com uso de ágar como substrato (C) e com vermiculita (D)] e no período de aclimatização [em bandejas com tampas (E), em bandejas de polipropileno (F), em potes plásticos de 3 L após transplantadas(G) e em potes plásticos de 3 L antes de ir para o campo]...........
73

13
SUMÁRIO
RESUMO................................................................................................... 07 ABSTRACT............................................................................................... 09 INTRODUÇÃO GERAL............................................................................. 14 REFERÊNCIAS BIBLIOGRÁFICAS....................................................... 18 CAPÍTULO 1 – Multiplicação in vitro de Pyrus sp., cvs. Ya-Li e Carrick
INTRODUÇÃO........................................................................................ 22 MATERIAL E MÉTODOS....................................................................... 23 RESULTADOS....................................................................................... 25 DISCUSSÃO.......................................................................................... 36 CONCLUSÕES...................................................................................... 40 REFERÊNCIAS BIBLIOGRÁFICAS....................................................... 40 CAPÍTULO 2 – Multiplicação in vitro de pereira, cv. Cascatense INTRODUÇÃO........................................................................................ 45 MATERIAL E MÉTODOS....................................................................... 47 RESULTADOS....................................................................................... 49 DISCUSSÃO.......................................................................................... 55 CONCLUSÕES...................................................................................... 59 REFERÊNCIAS BIBLIOGRÁFICAS....................................................... 60 CAPÍTULO 3 – Radiosensibilidade em micropropagação de Pyrus sp., cv. Ya-Li: efeitos morfofisiológicos
INTRODUÇÃO........................................................................................ 64 MATERIAL E MÉTODOS....................................................................... 66 RESULTADOS....................................................................................... 68 DISCUSSÃO.......................................................................................... 72 CONCLUSÕES...................................................................................... 78 REFERÊNCIAS BIBLIOGRÁFICAS....................................................... 78 CONSIDERAÇÕES FINAIS...................................................................... 84

14
INTRODUÇÃO GERAL
A pereira pertence à família Rosaceae, subfamília Pomoideae, gênero Pyrus
e compreende mais de 20 espécies, todas nativas da Ásia e da Europa. As espécies
mais importantes são: Pyrus communis L.; P. pyrifolia (Burm) Nak., P. ussuriensis
Max, P. bretschneideri Rehd, P. betaluefolia Bge., P. pashia P. Don., P. nivalis Jacq.
e P. calleryana Dcne (PERAZOLLO, 2006).
Devido à grande aceitação no consumo in natura, a produção de pera é
realizada em todo o mundo, o que a torna de grande importância nos mercados
internacionais (FACHINELLO et al., 2011). Segundo os dados de produção de pera
no ano de 2011, o Brasil produz 20 532 toneladas de peras por ano (IBGE, 2013).
Porém, esta quantidade não é suficiente para atender o mercado interno, pois
importa mais de 90% da pera de alta qualidade que consome (aproximadamente
140 mil toneladas) (NAKASU et al, 2007; FACHINELLO et al., 2011), apesar do
grande mercado interno para seus frutos (PETRI, 2008). Tal situação tem
posicionado o Brasil como o segundo maior importador de pera no mercado mundial
(JUNQUEIRA; PEETZ, 2003; NAKASU, 2003; PETRI, 2008; GUTIERREZ, 2009).
Entre as frutíferas de clima temperado, a pera é a terceira mais consumida,
entretanto é a que apresenta menor expressão em termos de produção e área
cultivada (FACHINELLO; FRANCESCATTO, 2009). Dada esta situação, o plantio da
pereira, no Brasil, é de fundamental importância, sendo uma alternativa de
diversificação econômica importante (JUNQUEIRA; PEETZ, 2003; NAKASU et al.,
2007).
No entanto, problemas relacionados com o vigor das plantas, tipo de porta-
enxerto, abortamento floral, insuficiência de frio hibernal, falta de cultivares
adaptadas às condições edafoclimáticas nacional, propriedades físicas, químicas e
biológicas do solo, dentre outros, impedem a expansão da cultura no País

15
(FACHINELLO et al., 2011). Dessa forma, o principal entrave para o bom
desenvolvimento da cultura é a limitação de cultivares adaptadas às diferentes
regiões potencialmente produtoras e também a falta de pesquisas em relação à
definição de portaenxertos (BIANCHI et al., 2002; TOMAZ et al., 2009) e de
cultivares copa utilizadas como pé franco, bem como à adaptação destes a solos
mais ácidos, como os do Rio Grande do Sul.
Para tanto, os estados do Rio Grande do Sul e Santa Catarina apresentam-se
como regiões favoráveis à produção da pereira, uma vez que se aproveita toda a
infra-estrutura de processamento e armazenagem já instaladas para a cultura da
macieira (RIBEIRO et al., 1991), além de apresentarem condições climáticas
adequadas (JUNQUEIRA; PEETZ, 2003). Isso faz com que esta região tenha
84,65% da área total nacional cultivada com peras (IBGE, 2009), porém essa área
de cultivo ainda é pequena para atender a demanda do mercado interno, devido à
baixa produtividade e qualidade inferior das frutas produzidas (WREGE et al., 2006).
Segundo Oliveira et al. (2004), a implantação de pomares com mudas de qualidade
é uma etapa decisiva para o sucesso do sistema produtivo, pois a muda de
qualidade potencializa o nível de reposta a toda tecnologia empregada no pomar,
sendo um componente decisivo na redução de custos e para a produção de frutas
com qualidade e alta produtividade.
Neste contexto, a micropropagação, ou propagação in vitro, tem sido
recomendada como uma alternativa viável para a propagação de mudas de grande
interesse econômico, devido à rapidez da propagação em larga escala, manutenção
das características genéticas (COUTO, 2003) e produção de material vegetal com
melhor qualidade sanitária, em relação àquele obtido por técnicas convencionais
como estaquia e mergulhia de cepa (BIANCHI et al., 2003).
A micropropagação é dividida em fases, que vão desde o estabelecimento da
cultura in vitro, passando pela multiplicação in vitro, enraizamento in vitro ou ex vitro
e aclimatização. A multiplicação visa produzir o maior número possível de plantas,
no menor espaço de tempo, sendo que para obter altas taxas de multiplicação dos
explantes é necessário o ajuste do protocolo para cada espécie/cultivar em estudo
(SCHUCH; ERIG, 2005). Dentre os fatores mais importantes que influenciam a taxa
de multiplicação destacam-se o tipo de meio de cultura utilizado, bem como a
concentração de citocinina e carboidratos adicionados ao meio (SILVEIRA et al.,
2001). Durante o enraizamento de várias espécies frutíferas lenhosas, o uso de

16
auxinas e substratos adequados são necessários para um bom processo de
rizogênese (RADMANN et al., 2002; TIBOLA et al., 2004). Neste sentido, o sucesso
do enraizamento dos explantes é pré-requisito para qualquer protocolo de
micropropagação, pois é importante para facilitar o estabelecimento da muda no solo
(PATI et al., 2006). Além disso, é imprescindível para sua aclimatização, uma vez
que o material micropropagado passa de uma condição heterotrófica e/ou
mixotrófica para autotrófica (BANDEIRA et al., 2007).
Sendo assim, diante do exposto acima, o sistema de micropropagação é um
método de reprodução assexuada importante, que pode ser usado para a produção
de plantas clonais, mas também forma a base para a introdução de variação
genética por transformação genética ou mutagênese (PÉREZ-TORNERO et al.,
2010), pois permite a multiplicação e/ou melhoramento de cultivares específicas e de
interesse econômico, como a pereira.
A radiação gama é considerada um dos principais indutores de mutações e de
aberrações cromossômicas estruturais, e as mudanças morfofisiológicas que
beneficiam a espécie em questão é uma das principais utilidades desta técnica no
melhoramento genético (HUNG; JOHNSON, 2008). Estudos sobre os efeitos das
radiações ionizantes em plantas são conduzidos mediante a irradiação de sementes,
explantes, plântulas e plantas, em diversos estádios de desenvolvimento (FU et al.,
2008; HUNG; JOHNSON, 2008). O melhoramento genético através do uso de
mutações induzidas em diversas culturas tem provado ser um método eficaz na
melhoria da qualidade, produtividade e resistência a estresses bióticos e abióticos
(NICHTERLAIN et al., 2000; BIBI et al., 2009). Dessa forma, técnicas de indução de
mutação têm sido muito utilizadas em programas de melhoramento de várias
espécies de plantas, como a pereira, em vista que o melhoramento genético
convencional não supre estas carências (JOSEPH et al., 2004). Em geral, as
espécies frutíferas são perenes, de longo ciclo vegetativo, de porte relativamente
grande e com alto nível de heterozigosidade, o que dificulta e aumenta o tempo
necessário para um programa de melhoramento genético através do método de
hibridização (CABONI et al., 2000).
Neste contexto, a produção de mudas sadias, a introdução de novas espécies
e o melhoramento genético contribuem para o aumento da eficiência do sistema
produtivo. Dessa forma, a Embrapa Clima Temperado, em parceria com a UFPel,

17
têm investido em pesquisas com a finalidade de melhorar os sistemas de produção
de pereira, realizando pesquisas com as cultivares „Carrick‟, „Cascatense‟ e „Ya-Li‟.
A cultivar Carrick é oriunda do cruzamento entre as cultivares Seckel x
Garber, obtida nos EUA. A planta é grande, vigorosa e produtiva. A plena floração
ocorre, em geral, na segunda semana de setembro. A fruta é de tamanho médio a
grande, de formato oblongo-piriforme e de epiderme bronzeada com manchas
avermelhadas. A polpa é branco-amarelada, medianamente macia, moderadamente
suculenta, doce, com pouca acidez, leve aroma e adstringente. A colheita ocorre em
fins de janeiro (NAKASU; FAORO, 2003; NAKASU et al., 2007).
A cultivar Cascatense foi criada pelo Programa de Melhoramento Genético da
Embrapa Clima Temperado, resultante do cruzamento de „Packham‟s Triumph‟ x „Le
Conte‟, realizado em 1968, selecionada em 1982 como Pirus 9 e lançada como
cultivar em 1992. „Packham‟s Triumph‟ é de origem australiana, resultante do
cruzamento entre „Uvedale St. Germain‟ (Bell) x „William‟s Bom Chrétien‟. „Le Conte‟
é americana, obtida do cruzamento entre pêra chinesa (arenosa) e européia
(manteigosa). A planta é de vigor médio a semi-vigorosa e semi-aberta. Floresce na
segunda quinzena de agosto, apresentando brotações mais precoces e uniformes,
enquanto que a fruta é piriforme e de tamanho médio. A epiderme é fina, de
coloração amarelo-esverdeada a amarela, com pouco de russeting na área
peduncular, de aparência regular. A polpa é branca, de textura parcialmente
manteigosa, suculenta, moderadamente aromática e de bom sabor com 12° Brix a
14° Brix. Além disso, sua exigência em frio hibernal situa-se ao redor de 400 h
(temperatura ≤ 7,2 °C) (NAKASU; LEITE, 1992; NAKASU; FAORO, 2003; NAKASU
et al., 2007).
Já a cultivar Ya-Li é oriental. A planta é de tamanho médio a grande, vigorosa,
aberta e levemente pendente. A floração é precoce, com antese ocorrendo na
primeira semana de setembro. Está entre as três principais variedades de pereiras
plantadas no Estado. É muito produtiva, apresentando frutas de tamanho médio a
grande, ovalado-piriforme, com epiderme amarelo-esverdeado-clara e lenticelas
grandes, de coloração marrom, principalmente junto ao pedicelo. A qualidade da
fruta melhora com atraso na colheita, a qual apresenta bom potencial de
armazenamento. A polpa é branca, crocante, pouco áspera e suculenta, doce.
Amadurece de março a abril. Tem necessidade de frio moderada (NAKASU;
FAORO, 2003; NAKASU et al., 2007).

18
Assim, esses atributos tornam as três cultivares alvo de pesquisas mais
detalhadas, pois são de grande interesse econômico. Contudo, apresentam alguns
problemas que dificultam uma melhor produção e qualidade das frutas, sendo
necessárias, desta forma, maiores pesquisas que visem introduzir novos genótipos
e/ou melhorar os atuais. Estes problemas estão relacionados ao fato de ambas
cultivares serem suscetíveis a entomosporiose, sendo que a Ya-Li também é
suscetível à sarna; a cv. Carrick apresenta frutas com qualidade média; as frutas
da „Cascatense‟ apresentam qualidade semelhante à Carrick, além de ser uma
cultivar lançada recentemente e ainda não existirem muitos trabalhos desenvolvidos.
E, por fim, as frutas da „Ya-Li‟ são de qualidade fraca a regular, com pouco teor de
açúcar, delicadas e exigem cuidados especiais na colheita e as plantas demoram a
entrar em produção (NAKASU; FAORO, 2003; FIORAVANÇO et al., 2007; NAKASU
et al., 2007).
Considerando o exposto acima, o objetivo deste trabalho foi otimizar o cultivo
in vitro de três cultivares de pereira (Pyrus sp.): Carrick, Cascatense e Ya-Li, visando
potencializar sua produção e a indução de mutações, almejando selecionar novos
genótipos com características de interesse, a partir dos genótipos originais.
Referências bibliográficas
BANDEIRA, F. S.; XAVIER, A.; OTONI, W. C.; LANI, E. R. G. Aclimatação ex vitro de plantas propagadas pela enxertia in vitro de Eucalyptus urophylla x E. grandis. Revista Árvore, v.3, n.5, p.773-781, 2007. BIANCHI, V. J.; CHAVES, A. da C.; SCHUCH, M. W.; FACHINELLO, J. C. Estabelecimento in vitro de marmeleiro: efeito do tipo de explante e tempo de imersão em hipoclorito de sódio. Revista Brasileira de Agrociência, v.9, n.2, p.177-179, 2003. BIANCHI, V. J.; VICENZI, M.; FACHINELLO, J. C. Resposta do crescimento de quatro cultivares de pereira, em viveiro, enxertadas sobre diferentes cultivares de marmeleiro na região sul do Brasil. In: CONGRESSO BRASILEIRO DE FRUTICULTURA, 2002, Florianópolis. Anais... Florianópolis: Tecnologia, Competitividade e Sustentabilidade, 2002. BIBI, S.; KHAN, I. A.; BUGHIO, H. R.; ODHANO, I. A.; ASAD, M. A.; KHATRI, A. Genetic differentiation of rice mutants based on morphological traits and molecular marker (RAPD). Pakistan Journal of Botany, v.41, p.737-743, 2009. CABONI, E.; LAURI, P.; D‟ANGELI, S. In vitro plant regeneration from callus of shoot apices in apple shoot culture. Plant Cell Reports, v.19, p.755.760, 2000.

19
COUTO, M. Propagação in vitro dos portaenxertos híbridos de pessegueiro ‘Barrier’ e ‘Cadman’ (Prunus sp.) 2003. 77f. Dissertação (Mestrado em Agronomia – Fruticultura de Clima Temperado) – Faculdade de Agronomia Eliseu Maciel, Universidade Federal de Pelotas, Pelotas. FACHINELLO, J. C.; FRANCESCATTO, P. Compatibilidade de enxerto e porta-enxerto de pereira. In: XI ENFRUTE (Encontro Nacional sobre Fruticultura de Clima Temperado), v.1, 2009, Fraiburgo, SC. Anais... Caçador: EPAGRI, 2009. p.33-42. FACHINELLO, J. C.; PASA, M. S.; SCHMTIZ, J. D.; BETEMPS, D. L. Situação e perspectivas da fruticultura de clima temperado no Brasil. Revista Brasileira de Fruticultura, v.33, n.1, p.109-120, 2011. FIORAVANÇO, J.C. A cultura da pereira no Brasil: situação econômica e entraves para o seu crescimento. Informações Econômicas, SP, v.37, n.3, 2007. FU, H. W.; LI, Y. F; SHU, Q. Y. A revisit of mutation induction by gamma rays in rice (Oryza sativa L.): implications of microsatellite markers for quality control. Molecullar Breeding, v.22, p.281-288, 2008. GUTIERREZ, A. Guia de variedades da pêra européia. 2009. Disponível em: http://www.jornalentreposto.com.br/qualidade/cqh/269-guia-de-variedades-da-peraeuropeia. Acesso em: 24 dez. 2009. HUNG, D. C.; JOHNSON, K. Effects of ionizing radiation on the growth and allyl isothiocyanate accumulation of Wasabia japonica in vitro and ex vitro. In Vitro Cellular Developmental Biology-Plant, v.44, p.51-58, 2008. IBGE – Instituto Brasileiro de Geografia e Estatística, Levantamento Sistemático da Produção Agrícola 2009. Disponível em: http:www.ibge.gov.br, 2011. IBGE – Instituto Brasileiro de Geografia e Estatística, Anuário Estatístico do Brasil 2012. Disponível em: http:www.ibge.gov.br, 2013. JOSEPH, R.; YEOH, H.; LOH, C. Induced mutations in cassava using somatic embryos and the identification of mutant plants with altered starch yield and composition. Plant Cell Report, v.23, p.91–98, 2004. JUNQUEIRA, A. H.; PEETZ, M. S. Mercados interno e externo. In: NAKASU, B.H.; CENTELLAS-QUEZADA, A.; HERTER, F.G. Pêra. Produção. Brasília: Embrapa Informação Tecnológica, 2003. p.10-19. (Frutas do Brasil, 46). NAKASU, B. H. Introdução. In: NAKASU, B. H.; CENTELLAS-QUEZADA, A.; HERTER, F. G. Pêra. Produção. Brasília: Embrapa Informação Tecnológica, 2003. p.9 (Frutas do Brasil, 46). NAKASU, B. H.; FAORO, I. D. Cultivares. In: NAKASU, B. H.; CENTELLAS-QUEZADA, A.; HERTER, F. G. Pêra. Produção. Brasília: Embrapa Informação Tecnológica, 2003. p.29-36 (Frutas do Brasil, 46).

20
NAKASU, B. H.; HERTER, F. G.; CAMELATTO, D.; JÚNIOR, C. R.; FORTES, J. F.; CASTRO, L. A. S. de; RASEIRA, A.; FREIRE, C. J. da S.; BASSO, C.; FAORO, I.; PETRI, J. L.; LEITE, G. B.; PEREIRA, J. F. M.; CANTILLANO, R. F. F.; VERÍSSIMO, V.; SIMÕES, F. A cultura da pêra. Brasília: Embrapa Informação Tecnológica, 2007. 58p. (Coleção Plantar, 58) NAKASU, B. H.; LEITE, D. L. Pirus 9: seleção de pereira para o Sul do Brasil. Hortisul, v.2, n.3, p.19-20, 1992. NICHTERLAIN, K.; BOHLMAN, H.; NICLEN, S.; MALUSZYNSKI, M. Achievements and trends of using induced mutations in crop improvement. In: DAEBRNS Symposium on the use of Nuclear and Molecular Techniques in Crop Improvement, p.27-35, 2000. OLIVEIRA, R. P. de; NINO, A. F. P.; NICKEL, O. Limpeza de patógenos e propagação in vitro de Cultivares de Pereira. Pelotas: Embrapa Clima Temperado, 2004. 5p. (Comunicado Técnico 105). PATI, P. K.; RATH, S. P.; SHARMA, M.; SOOD, A.; AHUJA; P. S. In vitro propagation of rose: a review. Biotechnology Advances, v.24, p.94-114, 2006. PERAZZOLO, G. Tecnologia para a produção de pêras européias. In: IX ENFRUTE (Encontro Nacional sobre Fruticultura de Clima Temperado), 1, 2006, Fraiburgo, SC. Anais... Caçador: EPAGRI, 2006. p.109-115. PÉREZ-TORNERO, O; TALLÓN, C. I; PORRAS, I. An efficient protocol for micropropagation of lemon (Citrus limon) from mature nodal segments. Plant Cell Tissue Organ Culture, v.100, n.3, p.263-271, 2010. PETRI, J. L. Problemática da cultura da pereira no Brasil. In: II Reunião Técnica da Cultura da Pereira, 2, 2008, Lages. Anais... Lages, SC, 2008. p.17-19. RADMANN, E. B.; FACHINELLO, J. C; PETERS, J. A. Efeito de auxinas e condições de cultivo no enraizamento in vitro de portaenxertos de macieira „M-9‟. Revista Brasileira de Fruticultura, v.24, n.3, p.624-628, 2002. RIBEIRO, P. A.; BRIGUENTI, E.; BERNARDI, J. Comportamento de algumas cultivares de pereira Pyrus communis L. e suas características nas condições do Planalto catarinense. Boletim Técnico, n.56, Florianópolis: EMPASC, 1991. 53p. SCHUCH, M. W.; ERIG, A. C. Micropropagação de plantas frutíferas. In. FACHINELO, J. C.; HOFFMANN, A.; NACHTIGAL, J. C. Propagação de plantas frutíferas. Brasília: Embrapa Informações Tecnológicas, 2005. p.155-173. SILVEIRA, C. A. P.; FACHINELLO, J. C.; FORTES, G. R. de L.; CITADIN, I.; RODRIGUES, A. C.; QUEZADA, A. C.; SILVA, J. B. Multiplicação in vitro de porta-enxertos do gênero Prunus sob diferentes concentrações de BAP em dois meios de cultura. Revista Brasileira de Fruticultura, v.23, n.3, p.488-492, 2001.

21
TIBOLA, C. S.; RADMANN, E. B.; RODRIGUES, A. C.; FORTES, G. R de L.; FACHINELLO, J. C. Diferentes meios de cultivo no enraizamento in vitro de porta-enxertos de Prunus sp. Revista Brasileira de Agrociência, v.10, n.2, p.191-195, 2004. TOMAZ, Z. F. P.; RODRIGUES, A. C.; VERÍSSIMO, V.; MARAFON, A. C.; HERTER, F. G.; RUFATO, A. DE R. Compatibilidade de enxertia de cultivares de marmeleiros com pereiras. Revista Brasileira de Fruticultura, 2009, v.31, n.4, p. 1211-1217. WREGE, M. S.; HERTER, F. G.; CAMELATTO, D.; STEINMETZ, S.; JÚNIOR, C. R.; GARRASTAZU, M. C.; FLORES, C. A.; IUCHI, T.; BERNARDI, J.; VERÍSSIMO, V.; MATZENAUER, R. Zoneamento agroclimático para pereira no Rio Grande do Sul. Pelotas: Embrapa Clima Temperado, 2006. 29p. (Embrapa Clima Temperado. Documentos, 182).

22
CAPÍTULO 1 – Multiplicação in vitro de Pyrus sp., cvs. Ya-Li e Carrick.
1 Introdução
A cultura da pereira (Pyrus sp.) tem grande potencial de expansão na Região
Sul do Brasil, devido à existência de condições favoráveis de clima e solo
adequados para seu desenvolvimento (WREGE et al., 2006). Esta região apresenta
uma área aproximada de 1180 hectares plantados de pera, com produção em torno
de 12.464 toneladas de frutos, equivalendo a 84,65% da área total cultivada (IBGE,
2009). No entanto, seu cultivo ainda é pequeno para atender a demanda do
mercado interno, pois a maior parte da fruta consumida no País é importada (mais
de 90%) (FACHINELLO; FRANCESCATTO, 2009; MUNIZ, et al., 2012).
Dentre os fatores que contribuem para a estagnação dos plantios e pela
pequena área cultivada com pereira pode ser citado a falta de cultivares adaptadas
(NAKASU; FAORO, 2003), gerando, assim, baixa produtividade e qualidade inferior
das frutas (WREGE et al., 2006).
Neste contexto, a implantação de pomares com mudas de qualidade é uma
etapa decisiva para o sucesso do sistema produtivo, pois a muda de qualidade
potencializa o nível de reposta a toda tecnologia empregada no pomar, sendo um
componente decisivo na redução de custos e para a produção de frutas com
qualidade e alta produtividade (OLIVEIRA et al., 2004).
Assim, visando à produção de mudas com alta qualidade fitossanitária,
técnicas de propagação in vitro apresentam-se como uma alternativa viável e
eficiente, pois possibilita a multiplicação do material vegetal e sua obtenção em larga
escala, num curto período de tempo e durante o ano todo (BOTELHO et al., 2005;
MACHADO et al., 2006; CARVALHO et al., 2011).
Entretanto, para que a taxa de reprodutibilidade vegetal seja elevada, é
necessário ajustar alguns fatores que interferem no desenvolvimento de brotações in

23
vitro, como o tipo de meio de cultura, seguido do suplemento de reguladores de
crescimento, concentração de sacarose, iluminação, tipo de explante, entre outros
(SILVEIRA et al., 2001; ZHANG et al., 2003; CAMPOS et al., 2007), sendo
necessário encontrar as condições mais adequadas para cada genótipo que se
pretende propagar (MACHADO et al., 2013).
Em relação aos meios de cultura, diversas formulações de meios básicos têm
sido utilizadas no cultivo in vitro, como o meio MS (MURASHIGE & SKOOG, 1962),
com suas modificações e diluições, o qual tem sido utilizado com sucesso para
diversas espécies (CENTOFANTE et al., 2009), e o meio WPM (LLOYD; MCCOWN,
1980), amplamente utilizado para a micropropagação de espécies lenhosas
(PASQUAL, 2001), QL (QUOIRIN; LEPOIVRE, 1977), entre outros.
A concentração adequada de sacarose no meio de cultivo é de grande
importância, pois a mesma serve como fonte de esqueletos de carbono, fornecendo
energia para a planta (PATI et al., 2006; VILLA et al., 2009).
As citocininas são reguladores de crescimento que desempenham um papel
fundamental na morfogênese in vitro, estimulando a divisão celular, assim como a
indução e a proliferação de brotações adventícias (PEREZ-TORNERO; BURGOS,
2000; DZAZIO et al., 2002), sendo a 6-benzilaminopurina (BAP) a mais utilizada na
multiplicação in vitro de pereiras, numa ampla faixa de concentração que varia de
0,5 a 2,0 mg L-1, de acordo com a espécie ou cultivar (DANTAS et al., 2002).
Desta forma, o trabalho teve como objetivo avaliar o efeito dos diferentes
meios de cultivo, concentrações de sacarose e BAP na multiplicação in vitro das cvs.
Ya-Li e Carrick, almejando estabelecer as condições adequadas para otimizar sua
propagação.
2 Material e Métodos
Os explantes utilizados para o desenvolvimento do trabalho foram
provenientes de brotações de pereira, cvs. Ya-Li e Carrick, pré-estabelecidas e
multiplicadas in vitro, durante 40 dias, em meio de cultivo MS pronto Himédia® (meio
comercial com formulação igual ao MS), acrescido de 0,8 mg L-1 de BAP, 30 g L-1 de
sacarose e 7 g L-1 de ágar, sendo o pH ajustado para 5,8 (Comunicação Pessoal).
Os estudos foram conduzidos em quatro experimentos sequenciais, conforme
descrito abaixo, sendo que o terceiro e quarto experimentos foram realizados
concomitantemente. Cada experimento foi repetido duas vezes. Para a realização

24
dos experimentos, como explante inicial, foram utilizados segmentos nodais de
aproximadamente 1,0 cm contendo de duas a três gemas axilares. As gemas
axilares presentes nos segmentos nodais, bem como o comprimento destes foram
incluídos nas avaliações realizadas. Gemas e brotações incipientes, menores que
três milímetros, não foram consideradas por serem muito pequenas e de difícil
manuseio.
Experimento 1 – Efeito de diferentes meios de cultivo e concentrações de sais
Os explantes iniciais foram cultivados em cinco meios diferentes: MS
completo e MS modificado (¾ da concentração normal de NH4NO3 e KNO3,
identificado nos resultados como MS¾), MS Himédia, WPM e QL; todos contendo
0,8 mg L-1 de BAP, 100 mg L-1 de mio-inositol, 30 g L- 1 de sacarose, 7 g L-1 de ágar
e pH 5,8. Os meios MS completo, MS¾ e QL foram preparados com EDTA-Férrico,
enquanto o MS Himédia e WPM com EDTA-Ferroso, conforme formulação original
do meio MS.
Os frascos foram mantidos em câmara de crescimento com temperatura de
23±2ºC, fotoperíodo de 16 horas e 48 μmol m-2 s-1 de fluxo de fótons por 40 dias.
Após este período, avaliou-se o número de brotações por explante, o comprimento
das brotações (cm), o número de gemas axilares por brotação, massa fresca e
massa seca das brotações (g), sendo a secagem do material realizada em estufa de
ventilação forçada a ±50 ºC até massa constante.
Experimento 2 – Influência das concentrações de 6-benzilaminopurina (BAP)
O meio de cultura usado foi o que apresentou melhores resultados após a
realização do primeiro experimento (MS Himédia para Carrick e MS para Ya-Li),
suplementado com diferentes concentrações de BAP (0; 0,4; 0,8; 1,2 e 1,6 mg L-1),
30 g L-1 de sacarose, 7 g L-1 de ágar e pH 5,8. As condições de cultivo e as variáveis
analisadas foram as mesmas do experimento anterior.
Experimento 3 – Efeito de diferentes concentrações de sacarose
O meio de cultivo e a concentração de BAP utilizados foram os que
apresentaram melhores resultados após a realização do primeiro e segundo
experimentos (MS + 1,2 mg L-1 de BAP para Ya-Li e MS Himédia + 1,2 mg L-1 de
BAP para Carrick), variando-se a concentração de sacarose (0, 15, 30, 45 e 60 g L-

25
1), com 7 g L-1 de ágar e pH 5,8. As condições de cultivo e as variáveis analisadas
foram as mesmas dos experimentos anteriores.
Experimento 4 – Efeito do aumento das concentrações de BAP
Quando o segundo experimento foi avaliado observou-se, em alguns
parâmetros, respostas crescentes até a última concentração de citocinina testada.
Em vista disso, com a finalidade de verificar o efeito do aumento das concentrações
de BAP na proliferação in vitro das cultivares em estudo, os explantes iniciais foram
inoculados em meios de cultura contendo concentrações maiores de BAP (0; 1; 2; 3
e 4 mg L-1). A composição básica do meio, bem como as condições de cultivo e as
variáveis analisadas foram as mesmas do segundo experimento.
Delineamento experimental e análise estatística
O delineamento experimental adotado para todos os experimentos foi
inteiramente casualizado, com nove repetições por tratamento, representadas, cada
uma, por um frasco contendo cinco explantes. Para as avaliações de massa fresca
e massa seca das brotações foram selecionadas, ao acaso, três plantas de cinco
repetições, totalizando quinze plantas por tratamento.
Os experimentos seguiram um arranjo bifatorial (2x5), composto por dois
níveis do fator cultivar (Ya-Li e Carrick) em todos os experimentos e cinco níveis dos
fatores: meio de cultura (MS, MS¾, MS Himédia, WPM e QL) para o primeiro
experimento, concentração de BAP (0; 0,4; 0,8; 1,2 e 1,6 mg L-1) para o segundo
experimento, concentração de sacarose (0, 15, 30, 45 e 60 g L-1) para o terceiro
experimento e concentração de BAP (0; 1; 2; 3 e 4 mg L-1) para o quarto
experimento.
Os resultados obtidos foram submetidos à análise de variância e as médias
comparadas pelo teste de Tukey, ao nível de 5% de probabilidade de erro, ou
analisadas por regressão polinomial, utilizando o software WINSTAT 1.0
(MACHADO; CONCEIÇÃO, 2007).
3 Resultados
Segundo a análise de variância e/ou regressão polinomial, observou-se
diferença significativa entre os tratamentos e interação entre os fatores para todas
as variáveis analisadas, nos quatro experimentos.

26
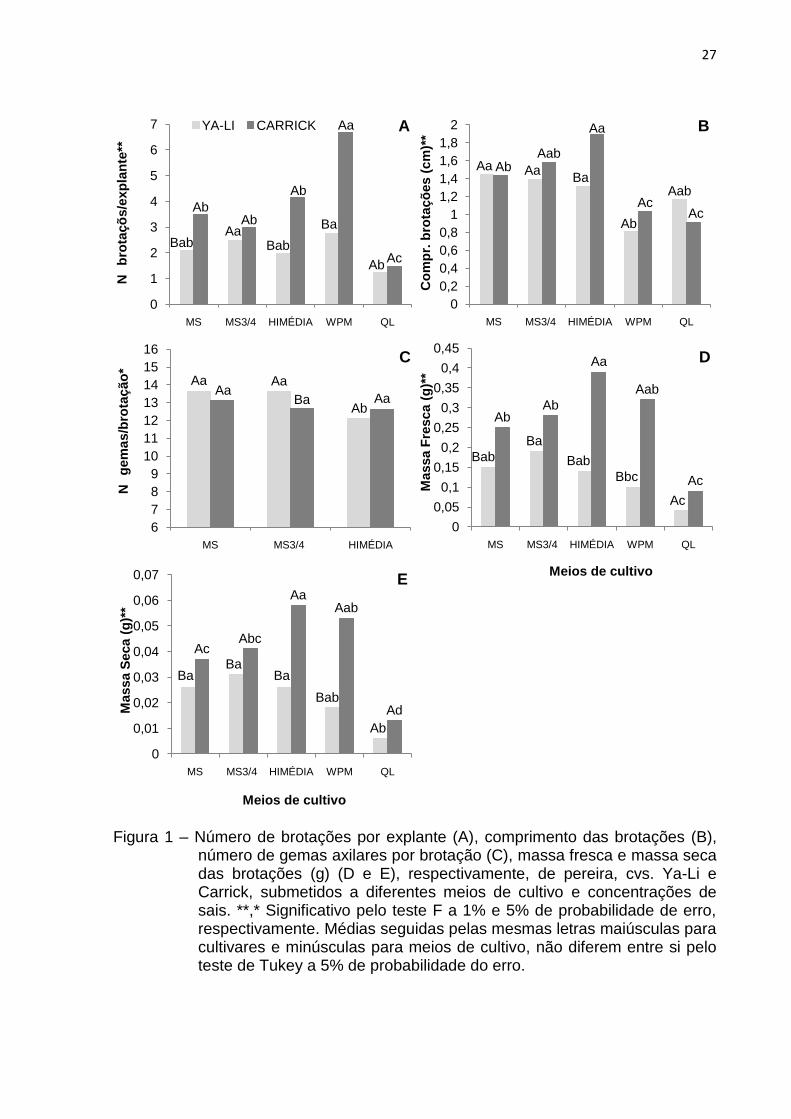
Experimento 1 – Efeito de diferentes meios de cultivo e concentrações de sais
Nas cultivares CarricK e Ya-Li, o maior número de brotações por explante
(6,67 e 2,75), respectivamente, foi obtido com o uso do meio de cultivo WPM,
porém, na „Ya-Li‟, este não diferiu estatisticamente do meio MS¾, com 2,48
brotações formadas (Fig. 1A).
Em relação ao comprimento das brotações, os maiores valores foram obtidos
com a utilização dos meios Himédia (1,89 cm) na cv. Carrick e MS (1,44 cm) na „Ya-
Li‟. Este último não diferiu significativamente dos meios MS ¾ (1,39 cm) e Himédia
(1,31 cm) (Fig. 1B).
No que se refere ao número de gemas axilares por brotação, não foi possível
ser realizada a avaliação dos meios WPM e QL, pois em ambas cultivares, houve
entufamento das brotações, ocasionado pela redução no comprimento das mesmas,
dificultando seu manuseio. Desta forma, para a cv. Carrick não foi verificada
diferença estatística entre os meios analisados, com uma média de 12,79 gemas
formadas, enquanto que para „Ya-Li‟, os meios MS e MS¾, foram os que
apresentaram os melhores resultados, propiciando a obtenção de 13,65 e 13,62
gemas, consecutivamente (Fig. 1C).
Os resultados obtidos para massa fresca e massa seca (Fig. 1D e 1E)
mostraram que, na cultivar Ya-Li, ocorreu maior incremento de massa fresca com o
meio MS¾ (0,19 g), e maior acúmulo de massa seca com os meios MS (0,026 g),
MS¾ (0,031 g) e Himédia (0,026 g), os quais não diferiram entre si. Já para a cv.
Carrick, os maiores valores de massa fresca (0,39 g) e seca (0,058 g) foram obtidos
com o uso de Himédia.
Analisando todas as variáveis avaliadas, verificou-se que, para a cultivar Ya-
Li, o meio de cultivo MS foi o que propiciou a obtenção de respostas mais uniformes,
com formação de 2,09 brotações por explante, com comprimento médio de 1,44 cm,
13,65 gemas axilares por brotação, 0,15 g de massa fresca e 0,026 g de massa
seca. Na cv. Carrick, o meio Himédia foi o mais responsivo, obtendo-se 4,16
brotações por explante, com 1,89 cm de comprimento, 12,64 gemas axilares por
brotação, 0,39 g de massa fresca e 0,058 g de massa seca.
27
Bab Aa
Bab
Ba
Ab
AbAb
Ab
Aa
Ac
0
1
2
3
4
5
6
7
MS MS3/4 HIMÉDIA WPM QL
Nb
rota
çõ
s/e
xp
lan
te**
YA-LI CARRICK
Aa AaBa
Ab
Aab
AbAab
Aa
AcAc
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
2
MS MS3/4 HIMÉDIA WPM QL
Co
mp
r. b
rota
çõ
es (
cm
)**
Aa Aa
Ab
AaBa Aa
6
7
8
9
10
11
12
13
14
15
16
MS MS3/4 HIMÉDIA
Ng
em
as/b
rota
ção
*
Bab
Ba
Bab
Bbc
Ac
AbAb
Aa
Aab
Ac
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0,45
MS MS3/4 HIMÉDIA WPM QL
Massa F
resca (
g)*
*
BaBa
Ba
Bab
Ab
AcAbc
AaAab
Ad
0
0,01
0,02
0,03
0,04
0,05
0,06
0,07
MS MS3/4 HIMÉDIA WPM QL
Massa S
eca (
g)*
*
Figura 1 – Número de brotações por explante (A), comprimento das brotações (B), número de gemas axilares por brotação (C), massa fresca e massa seca das brotações (g) (D e E), respectivamente, de pereira, cvs. Ya-Li e Carrick, submetidos a diferentes meios de cultivo e concentrações de sais. **,* Significativo pelo teste F a 1% e 5% de probabilidade de erro, respectivamente. Médias seguidas pelas mesmas letras maiúsculas para cultivares e minúsculas para meios de cultivo, não diferem entre si pelo teste de Tukey a 5% de probabilidade do erro.
Meios de cultivo
Meios de cultivo
A B
C D
E

28
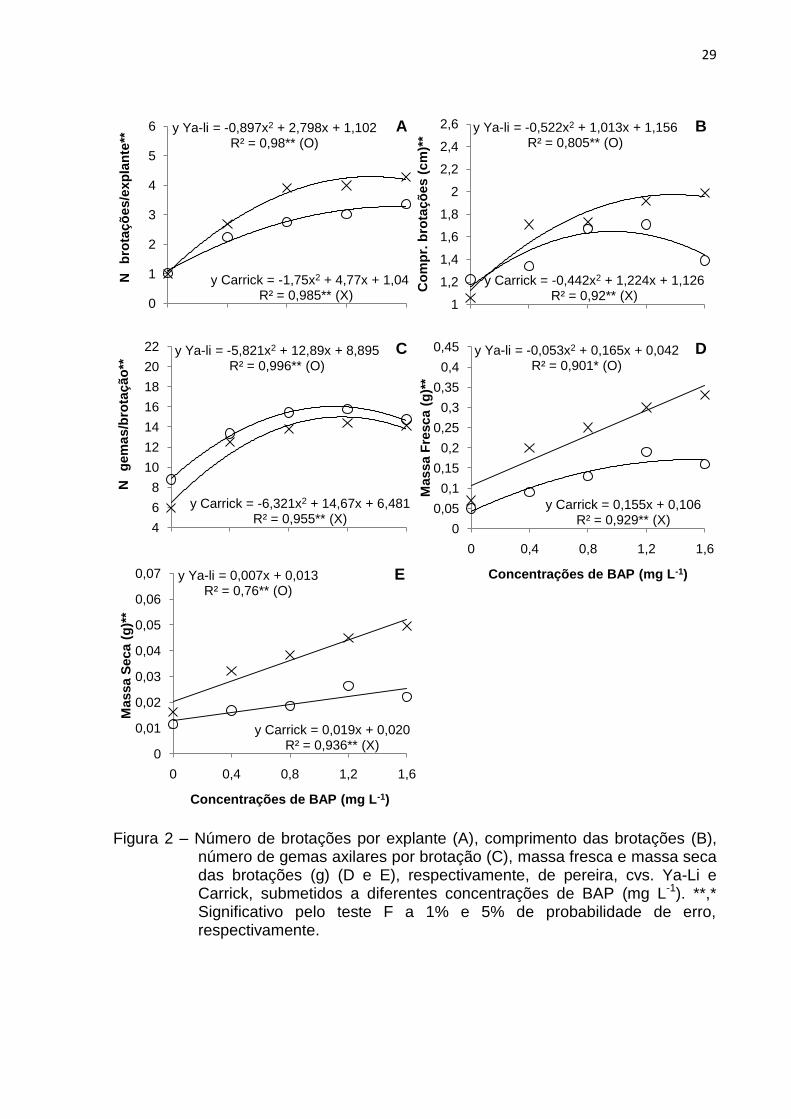
Experimento 2 – Influência das concentrações de 6-benzilaminopurina (BAP)
Em ambas cultivares, para as variáveis número e comprimento de brotações
por explante e número de gemas axilares por brotação, verificou-se uma tendência
quadrática, com ponto de máxima, para a cultivar Ya-Li, nas concentrações 1,56,
0,97 e 1,1 mg L-1 de BAP, com a formação de 3,28 brotações, de aproximadamente
1,65 cm de comprimento e 16,03 gemas axilares por brotação, respectivamente. Em
contrapartida, na cultivar Carrick, a adição de 1,36, 1,38 e 1,16 mg L-1 de BAP ao
meio de cultivo, ocasionou a formação de 4,29 brotações, de aproximadamente
1,97 cm de comprimento e 14,99 gemas axilares por brotação, consecutivamente
(Fig. 2A, 2B e 2C).
Nas cultivares em estudo, observou-se um incremento na massa fresca com o
aumento de BAP, sendo que, para a „Ya-Li‟, pela estimativa da equação de
regressão, a resposta foi positiva até 1,56 mg L-1, com obtenção de 0,17 g. Já para
„Carrick‟, os maiores valores foram obtidos na concentração mais elevada (1,6 mg L-
1), apresentando uma média 0,33 g (Fig. 2D).
Em relação à massa seca, verificou-se uma resposta linear crescente, com o
acúmulo de 0,022 e 0,049 g na concentração de 1,6 mg L-1 da citocinina testada,
para as cvs. Ya-Li e Carrick, respectivamente (Fig. 2E).
Embora os resultados obtidos para todas estas variáveis, em ambas
cultivares, sejam estimados conforme a equação de regressão, os valores absolutos
encontrados mostram que a concentração de 1,2 mg L-1 de BAP foi a que
desencadeou as melhores e mais uniformes respostas, pois apresentou a formação
de 3,02 brotações, com 1,71 cm de comprimento, 15,78 gemas axilares por
brotação, 0,19 g de massa fresca e 0,026 g de massa seca, para „Ya-Li‟. Para a cv.
Carrick, obteve-se 3,99 brotações por explante, com 1,92 cm de comprimento cada,
14,4 gemas axilares por brotação e incremento de 0,3 g de massa fresca e 0,045 g
de massa fresca.
29
y Ya-li = -0,897x2 + 2,798x + 1,102 R² = 0,98** (O)
y Carrick = -1,75x2 + 4,77x + 1,04R² = 0,985** (X)
0
1
2
3
4
5
6N
bro
taçõ
es/e
xp
lan
te**
y Ya-li = -0,522x2 + 1,013x + 1,156 R² = 0,805** (O)
y Carrick = -0,442x2 + 1,224x + 1,126 R² = 0,92** (X)
1
1,2
1,4
1,6
1,8
2
2,2
2,4
2,6
Co
mp
r. b
rota
çõ
es (
cm
)**
y Ya-li = -5,821x2 + 12,89x + 8,895 R² = 0,996** (O)
y Carrick = -6,321x2 + 14,67x + 6,481 R² = 0,955** (X)
4
6
8
10
12
14
16
18
20
22
Ng
em
as/b
rota
ção
**
y Ya-li = -0,053x2 + 0,165x + 0,042 R² = 0,901* (O)
y Carrick = 0,155x + 0,106 R² = 0,929** (X)
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0,45
0 0,4 0,8 1,2 1,6
Massa F
resca (
g)*
*
Concentrações de BAP (mg L-1)y Ya-li = 0,007x + 0,013 R² = 0,76** (O)
y Carrick = 0,019x + 0,020 R² = 0,936** (X)
0
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0 0,4 0,8 1,2 1,6
Massa S
eca (
g)*
*
Concentrações de BAP (mg L-1)
Figura 2 – Número de brotações por explante (A), comprimento das brotações (B),
número de gemas axilares por brotação (C), massa fresca e massa seca das brotações (g) (D e E), respectivamente, de pereira, cvs. Ya-Li e Carrick, submetidos a diferentes concentrações de BAP (mg L-1). **,* Significativo pelo teste F a 1% e 5% de probabilidade de erro, respectivamente.
A B
C D
E

30
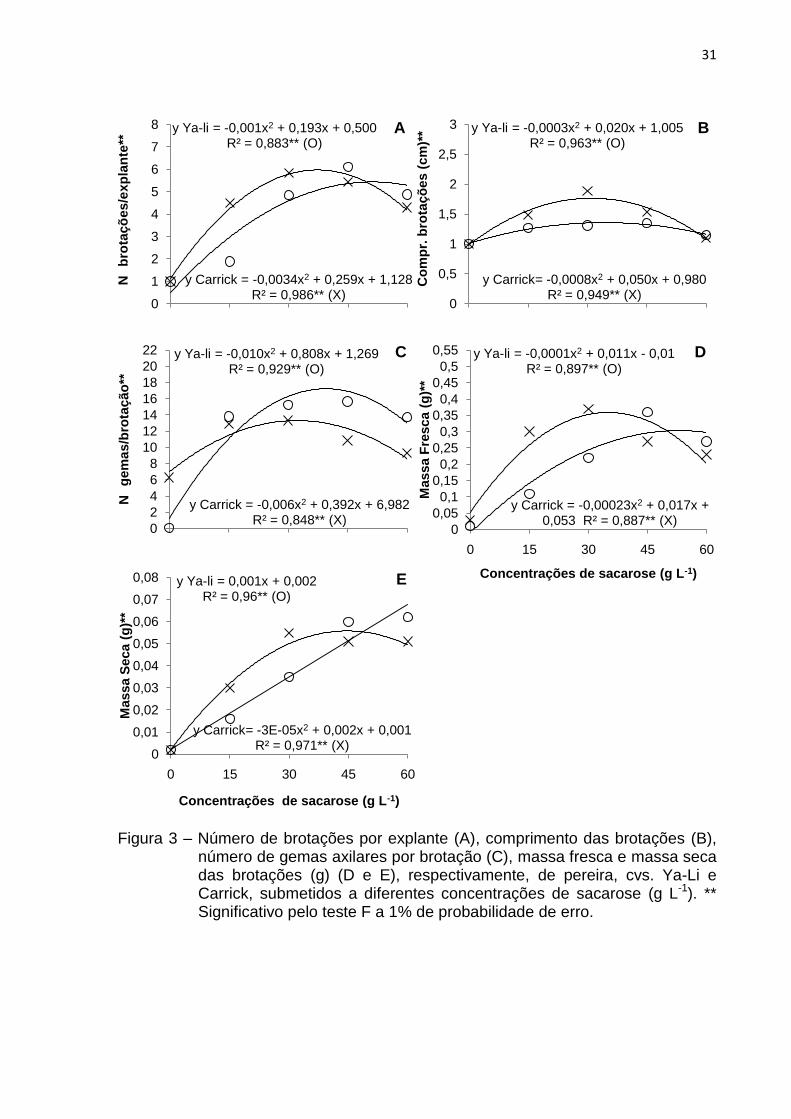
Experimento 3 – Efeito de diferentes concentrações de sacarose
Quanto às diferentes concentrações de sacarose adicionadas aos meios de
cultivo verificou-se uma tendência quadrática para todas as variáveis analisadas,
nas duas cultivares, exceto no que diz respeito à massa seca da pereira Ya-Li, na
qual houve uma resposta linear crescente, com maiores valores obtidos na
concentração de 60 g L-1, apresentando um acúmulo de 0,062 g (Fig. 3E).
Segundo a equação de regressão, maiores valores para número de brotações
por explante (5,67) (Fig. 3A), comprimento das brotações (1,33 cm) (Fig. 3B), gemas
axilares por brotação (17,59) (Fig. 3C) e massa fresca (0,312 g) (Fig. 3D) foram
obtidos nas concentrações de 53,61; 33,33; 40,4 e 55 g L-1 de sacarose,
respectivamente, na cultivar Ya-Li. Em contrapartida, para a cultivar Carrick, o ponto
de máxima inferiu as maiores médias de brotações por explante (6,06) (Fig. 3A),
comprimento das brotações (1,76 cm) (Fig. 3B), gemas axilares por brotação (13,38)
(Fig. 3C), massa fresca (0,367 g) (Fig. 3D) e massa seca (0,056 g) (Fig. 3E) quando
utilizadas 38,09; 31,25; 32,67; 36,96 e 45,4 g L-1 deste carboidrato,
consecutivamente.
Ainda que as avaliações estimadas segundo a tendência quadrática tenham
propiciado respostas diversas, fazendo-se uma análise dos resultados obtidos com
os valores absolutos, pode-se inferir que, para a cv. Ya-Li, a concentração de 45 g L-
1 de sacarose corrobora para a obtenção das melhores e mais uniformes respostas,
pois nela ocorreu a formação de 6,11 brotações, com comprimento médio de 1,35
cm, 15,63 gemas axilares e incremento 0,36 g de massa fresca e 0,06 g massa
seca; enquanto que, para a cv. Carrick, os maiores valores de número de brotações
(5,84), comprimento de brotações (1,88 cm), gemas axilares por brotação (13,29),
massa fresca (0,37 g) e massa seca (0,055 g) foram observados na concentração de
30 g L-1 deste açúcar.
31
y Ya-li = -0,001x2 + 0,193x + 0,500 R² = 0,883** (O)
y Carrick = -0,0034x2 + 0,259x + 1,128 R² = 0,986** (X)
0
1
2
3
4
5
6
7
8N
bro
taçõ
es/e
xp
lan
te**
y Ya-li = -0,0003x2 + 0,020x + 1,005 R² = 0,963** (O)
y Carrick= -0,0008x2 + 0,050x + 0,980 R² = 0,949** (X)
0
0,5
1
1,5
2
2,5
3
Co
mp
r. b
rota
çõ
es (
cm
)**
y Ya-li = -0,010x2 + 0,808x + 1,269 R² = 0,929** (O)
y Carrick = -0,006x2 + 0,392x + 6,982 R² = 0,848** (X)
0
2
4
6
8
10
12
14
16
18
20
22
Ng
em
as/b
rota
ção
**
y Ya-li = -0,0001x2 + 0,011x - 0,01 R² = 0,897** (O)
y Carrick = -0,00023x2 + 0,017x + 0,053 R² = 0,887** (X)
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0,45
0,5
0,55
0 15 30 45 60
Massa F
resca (
g)*
*
Concentrações de sacarose (g L-1)y Ya-li = 0,001x + 0,002
R² = 0,96** (O)
y Carrick= -3E-05x2 + 0,002x + 0,001 R² = 0,971** (X)
0
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0 15 30 45 60
Massa S
eca (
g)*
*
Concentrações de sacarose (g L-1)
Figura 3 – Número de brotações por explante (A), comprimento das brotações (B),
número de gemas axilares por brotação (C), massa fresca e massa seca das brotações (g) (D e E), respectivamente, de pereira, cvs. Ya-Li e Carrick, submetidos a diferentes concentrações de sacarose (g L-1). ** Significativo pelo teste F a 1% de probabilidade de erro.
A B
C D
E

32
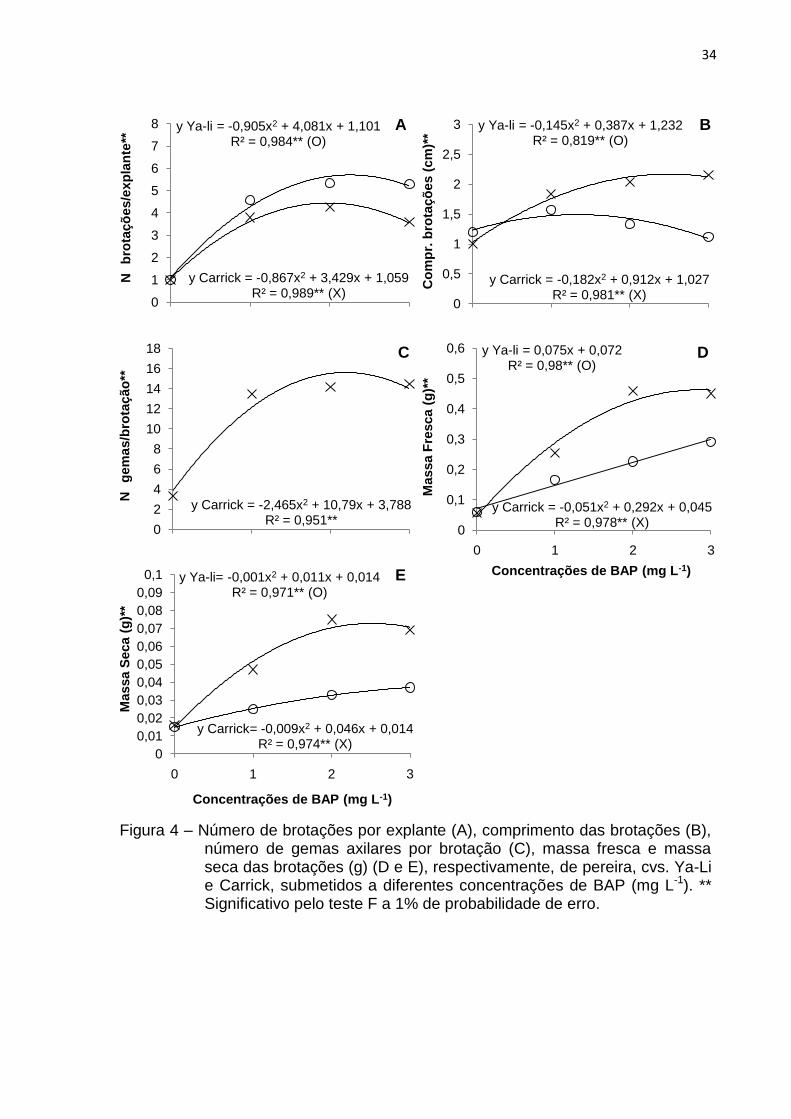
Experimento 4 – Efeito do aumento das concentrações de BAP
Mediante os resultados obtidos no segundo experimento, realizou-se esta
avaliação complementar, com a finalidade de verificar as respostas observadas com
o incremento das concentrações de BAP.
Pela equação de regressão, nas cvs. Ya-Li e Carrick, os maiores valores
estimados para número (5,7 e 4,45) (Fig. 4A) e comprimento das brotações (1,49 e
2,17 cm) (Fig. 4B) e massa seca (0,044 e 0,073 g) (Fig. 4E) foram observados nas
concentrações de 2,25 e 1,73 mg L-1; 1,33 e 2,5 mg L-1 e 5,5 e 2,55 mg L-1 de BAP,
respectivamente.
No que se refere às variáveis número de gemas axilares por brotação e
massa fresca, na cv. Carrick, observou-se uma resposta quadrática, com pontos de
máxima nas concentrações de 2,19 e 2,86 mg L-1 de BAP, com 15,59 gemas
formadas (Fig. 4C) e incremento de 0,463 g de massa fresca (Fig. 4D),
consecutivamente.
Entretanto, para a cultivar Ya-Li, ocorreu um efeito linear crescente à massa
fresca com a adição de BAP ao meio de cultivo, obtendo-se valores máximos de
0,291 g quando se utilizou 3 mg L-1 deste regulador de crescimento (Fig. 4D). O
número de gemas axilares da „Ya-Li‟ não foi avaliado nas concentrações de 2 e 3 mg
L-1 de BAP, em vista que houve dificuldade na sua contagem, sendo que nas
concentrações inferiores (0 e 1 mg L-1) verificou-se a formação de 9,35 e 14,61
gemas, respectivamente.
Embora as respostas encontradas sejam estimadas conforme a equação de
regressão, em ambas cultivares, os valores absolutos obtidos para a cv. Ya-Li são
diversos, não permitindo identificar uma concentração em que as respostas sejam
uniformes. Já para a cv. Carrick a concentração de 2 mg L-1 desencadeou bons
resultados, com a formação de 4,27 brotações, com 2,04 cm de comprimento, 14,14
gemas axilares por brotação, 0,458 g de massa fresca e 0,075 g de massa seca.
Porém, fazendo-se uma avaliação conjunta dos resultados obtidos e das
observações visuais (Fig. 5E e 5F), é possível verificar que concentrações de 2 e 3
mg L-1 de BAP são danosas às cultivares em estudo, pois na „Ya-Li‟, ocorreu
entufamento das brotações, ocasionada pela grande proliferação de brotações, as
quais apresentaram redução e variação no comprimento (Fig. 5E). Em contrapartida,
para „Carrick‟ houve diferenças no comprimento das brotações formadas na
concentração de 2 mg L-1 e formação de brotações vitrificadas com o uso de 3 mg L-

33
1 deste fitoregulador (Fig. 5F). Neste contexto, nas cultivares em estudo, o uso de
1,2 mg L-1 de BAP é adequado, em vista que, no segundo experimento apresentou-
se favorável à proliferação de brotações, levando à obtenção dos melhores
resultados, e neste experimento a concentração de 1 mg L-1 deste regulador de
crescimento propiciou respostas positivas e crescentes.
34
y Ya-li = -0,905x2 + 4,081x + 1,101 R² = 0,984** (O)
y Carrick = -0,867x2 + 3,429x + 1,059 R² = 0,989** (X)
0
1
2
3
4
5
6
7
8N
bro
taçõ
es/e
xp
lan
te**
y Ya-li = -0,145x2 + 0,387x + 1,232 R² = 0,819** (O)
y Carrick = -0,182x2 + 0,912x + 1,027 R² = 0,981** (X)
0
0,5
1
1,5
2
2,5
3
Co
mp
r. b
rota
çõ
es (
cm
)**
y Carrick = -2,465x2 + 10,79x + 3,788 R² = 0,951**
0
2
4
6
8
10
12
14
16
18
Ng
em
as/b
rota
ção
**
y Ya-li = 0,075x + 0,072 R² = 0,98** (O)
y Carrick = -0,051x2 + 0,292x + 0,045 R² = 0,978** (X)
0
0,1
0,2
0,3
0,4
0,5
0,6
0 1 2 3
Massa F
resca (
g)*
*
Concentrações de BAP (mg L-1)y Ya-li= -0,001x2 + 0,011x + 0,014
R² = 0,971** (O)
y Carrick= -0,009x2 + 0,046x + 0,014 R² = 0,974** (X)
0
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
0,1
0 1 2 3
Massa S
eca (
g)*
*
Concentrações de BAP (mg L-1)
Figura 4 – Número de brotações por explante (A), comprimento das brotações (B), número de gemas axilares por brotação (C), massa fresca e massa seca das brotações (g) (D e E), respectivamente, de pereira, cvs. Ya-Li e Carrick, submetidos a diferentes concentrações de BAP (mg L-1). ** Significativo pelo teste F a 1% de probabilidade de erro.
A B
C D
E

35
Figura 5 – Brotações de Pyrus sp., cvs. Ya-Li e Carrick, obtidas após serem submetidas a diferentes meios de cultivo e concentrações de sais (A e B), concentrações de BAP (C, D, E e F) e concentrações de sacarose (G e H), respectivamente.

36
4 Discussão
A partir da cultura de tecidos é possível a obtenção de plantas geneticamente
idênticas à planta matriz, livre de pragas e doenças e em número elevado
(CARVALHO et al., 2006). Contudo, essa alta produção de mudas está relacionada
ao sucesso da etapa de multiplicação in vitro, a qual é influenciada por diversos
fatores, dentre eles a composição do meio de cultivo, a utilização de citocininas
como reguladores de crescimento e condições ambientais a que os tecidos são
submetidos (SOTIROPOULOS et al., 2006).
Convém ressaltar, que é necessário otimizar as condições de cultivo para
cada espécie e/ou cultivar que se pretende propagar (ROGALSKI et al., 2003), em
vista que cada genótipo responde de forma diferenciada aos tratamentos testados,
podendo ocorrer diferenças no padrão de desenvolvimento das brotações
(BORGHEZAN et al., 2003; CAMPOS et al., 2007), como observado neste trabalho,
em que as cultivares Ya-Li e Carrick apresentaram variação na morfogênese in vitro
(Fig. 5). Assim, existe uma série de estudos para determinadas cultivares de pereira,
mas muitas vezes estes resultados não podem ser transferidos para genótipos
individuais, porque estes podem diferir muito em suas necessidades nutricionais,
energéticas e hormonais, durante o cultivo in vitro (REED et al., 2013a)
Considerando o exposto acima e visando obter um protocolo apropriado de
propagação do material vegetal em estudo, em que o objetivo é conseguir altas
taxas de multiplicação in vitro, os parâmetros de crescimento avaliados
considerados mais importantes são o número e comprimento das brotações e o
número de gemas axilares por brotação. Esses, em conjunto, vão determinar uma
maior ou menor eficiência do processo de multiplicação e propiciar que, a partir do
protocolo criado, as brotações obtidas tenham comprimento e qualidade adequados
para o sucesso das fases de enraizamento e aclimatização.
Em relação à composição do meio de cultura, segundo Torres et al. (2001),
os principais componentes são água, sais inorgânicos, carboidratos, vitaminas e
reguladores de crescimento, em vista que os explantes estão em ambiente
heterotrófico e/ou mixotrófico e requerem meio nutritivo para suplementar suas
necessidades exógenas, em termos de elementos essenciais, constituintes
orgânicos e energia.
Neste sentido, muitos experimentos com diluições nas concentrações de
macro e micronutrientes têm sido realizados. As grandes diferenças nos

37
macronutrientes para os meios MS, WPM e QL são nas concentrações dos sais
nitrato de amônio e nitrato de potássio e a concentração total de íons (BELL et al.,
2009). E, embora o WPM tenha gerado resultados positivos em alguns parâmetros
avaliados (ex: número de brotações, em ambas cultivares; e no incremento de
massa fresca e seca na cultivar Ya-Li) (Fig. 5A e 5B) e seja muito utilizado na
multiplicação de brotações de várias espécies lenhosas, em razão da sua menor
força iônica total em relação a outros meios empregados no cultivo in vitro
(PASQUAL, 2001; JHA; GHOSH, 2006; RADMANN et al., 2009), o MS (tanto na
forma comercial como a preparada no laboratório) e suas diluições foram os que, no
geral, desencadearam a obtenção das melhores respostas (Fig. 5A e 5B). Com o
uso de WPM houve um acréscimo no número de brotações, porém estas tiveram um
comprimento inadequado, levando ao entufamento das mesmas (principalmente na
cultivar Ya-Li), o que torna difícil o manuseio e separação dos explantes, bem como
sua utilização nas etapas posteriores de micropropagação. Esta redução no
comprimento das brotações e, consequentemente, impossibilidade de obtenção de
altas taxas de multiplicação também pode ser visualizada no meio QL (Fig. 5A e 5B),
o qual apresenta força iônica total menor que o WPM.
De acordo com Reed et al. (2013b), as espécies e cultivares de pereira são
cultivadas em vários meios de cultura conhecidos, entretanto o MS é o mais
comumente usado devido à qualidade superior das brotações formadas. O melhor
crescimento de células e tecidos de plantas em meio MS tem sido atribuído, em
grande parte, à alta concentração de amônio e nitrato. E, além do nitrogênio, o
potássio encontra-se também em dosagens altas (FADEL et al., 2010). Estes
macronutrientes estão envolvidos em muitos processos metabólicos da planta,
relacionados ao crescimento e vigor vegetal, sendo que o nitrogênio é constituinte
de aminoácidos, amidas, proteínas, ácidos nucléicos, nucleotídeos, coenzimas e
outros (XU et al., 2012) e o potássio é requerido como cofator de mais de 40
enzimas, além de ser o principal cátion no estabelecimento do turgor celular e
manutenção da eletroneutralidade celular (WANG; WU, 2013).
Outro aspecto a ser considerado, em relação aos resultados obtidos durante a
multiplicação das duas cultivares, nos diferentes meios de cultivo, é no que se refere
às diferenças no desenvolvimento das brotações observadas entre Himédia e MS,
uma vez que ambos são preparados com as mesmas concentrações de sais. Esta
resposta pode estar relacionada ao fato do meio Himédia ter sido preparado com

38
Ferro na forma ferrosa – reduzida, enquanto o MS na forma de EDTA-Férrico –
oxidada. Ambos são absorvidos normalmente pelas plantas, porém algumas (a
maioria) absorvem melhor o Ferro como Fe2+ (forma mais solúvel), e outras como
Fe3+-quelato (SCHMIDT, 2003). Além disso, o Himédia é comercial e já vem pronto,
enquanto que o MS é preparado em laboratório, assim, a qualidade dos sais
minerais, bem como possíveis alterações na quantidade destes, podem variar entre
eles.
No que se refere ao uso de citocininas no meio de cultivo (Fig. 5C à 5F),
verificou-se que ambas cultivares necessitam de adição exógena deste regulador de
crescimento, pois nos explantes em que não houve suplemento de BAP observou-se
ausência de multiplicação. Isso ocorreu devido às citocininas serem indispensáveis
nesta fase, uma vez que promovem a superação da dominância apical e induzem a
proliferação de gemas axilares, favorecendo, assim, a emissão de novas brotações
(SOUZA; JUNGHANS, 2006; GRIMALDI et al., 2008).
Em contrapartida, o excesso de citocinina é tóxico e caracteriza-se,
principalmente, pela formação de tufos de brotações e alterações no seu
comprimento, encurtamento dos entrenós, engrossamento exagerado dos caules e
hiperidricidade generalizada (GEORGE et al., 2008), sendo que essas
características foram observadas nas concentrações mais altas de BAP (2 e 3 mg L-
1) (Fig. 5E e 5F). Brotações hiperídricas (como as formadas na concentração de 3
mg L-1, na cv. Carrick) (Fig. 5F), apresentam caules espessos, entrenós curtos, com
aparência túrgida, hipolignificados, contendo água nos espaços intercelulares e
sobre sua superfície; as folhas são malformadas, com alterações na filotaxia,
frequentemente alongadas, esfaceláveis e encarquilhadas ou curvadas; múltiplas
brotações evidenciando mau funcionamento dos meristemas primários, que podem
degenerar rapidamente por necrose simultânea dos ápices meristemáticos
(GASPAR et al., 2000). Assim, o uso de brotações formadas nessas condições é
indesejável, em vista que geram sérios problemas para os cultivos subsequentes e
demais etapas da micropropagação, comprometendo o desenvolvimento das
culturas (RAMAGE; WILLIAMS, 2004). Diante do exposto, almejando obter uma taxa
média satisfatória de brotações em que sejam priorizadas qualidade e
homogeneidade, levando à produção de mudas em larga escala, a concentração de
BAP que desencadeou os melhores resultados, para as duas cultivares, foi a de 1,2
mg L-1 (Fig. 5C e 5D).

39
Em relação ao uso de carboidratos, a adição de sacarose ao meio de cultura
pode atuar como substrato para a respiração celular, fornecendo energia também
para o processo de rizogênese (PIO et al., 2002). Neste sentido, diversas pesquisas
são realizadas buscando adequar uma concentração em que exista eficiência no uso
desta para cada espécie/cultivar, pois a redução de sacarose no meio de cultivo tem
sido testada em busca de melhorar a capacidade fotossintética dos tecidos
cultivados in vitro e reduzir perdas atribuídas à contaminação microbiana. No
entanto, muitos autores são contrários à ideia de redução de sacarose durante as
etapas da micropropagação e afirmam que os mecanismos pelos quais a
concentração de carboidrato influencia na aclimatização ainda não são muito claros
(ARAÚJO et al., 2007). Em contrapartida, o seu excesso pode ser prejudicial, pois
inibe a síntese de clorofila e, portanto, reduz a capacidade fotossintética das culturas
(HAZARIKA, 2006).
Assim, estudos de multiplicação de pereiras são realizados com o uso de
diferentes concentrações de sacarose no meio de cultivo, como com os porta-
enxertos de Pyrus betulaefolia (HASSANEN; GABR et al., 2012) e Pyrus communis
'Pyrodwarf' (RUZIC et al., 2011), nos quais utiliza-se 20 g L-1, enquanto que em
Pyrus sp (REED et al., 2013b), Pyrus calleryana D-6 (MORAES et al., 2004), Pyrus
pyrofolia „Século XX‟ e „Housui‟ e Pyrus communis ‘Red Bartlet‟ (DANTAS et al.,
2002) usa-se 30 g L-1 (concentração de sacarose normalmente utilizada no cultivo in
vitro). E, neste trabalho, os melhores resultados obtidos foram com o uso de 30 g L-1
de sacarose para „Carrick‟ e 45 g L-1 para Ya-Li‟ (Fig. 5G e 5H), onde foram
verificadas as maiores taxas de multiplicação das brotações, mostrando que a
concentração apropriada é dependente do genótipo.
Durante a realização dos experimentos (os quais foram realizados em
sequência, sendo que o terceiro e o quarto foram realizados concomitantemente), foi
possível aferir um aumento no número das brotações, em ambas cultivares, entre o
segundo e terceiro experimentos (Fig. 2A e 3A), já que no segundo experimento os
valores estavam condizentes com os encontrados no primeiro (Fig. 1A). As
cultivares Ya-Li e Carrick, que produziam 3 e 3,9 brotações por explante, passaram
a produzir 4,8 e 5,8, respectivamente, sendo que isso se manteve no quarto
experimento (Fig. 4A). Esse fato pode estar relacionado ao estado fisiológico do
explante e juvenilidade do tecido após os subcultivos, bem como às condições de
luminosidade, pois ocorre diminuição na intensidade de fluxo de fótons conforme as

40
lâmpadas vão sendo utilizadas na sala de crescimento. A luz é um fator importante
para o desenvolvimento das plantas, podendo influenciar a taxa de multiplicação in
vitro (RADMANN et al., 2001), regulando os processos fotossintéticos e a
fotomorfogênese (KOZAI et al., 1997). Poucos trabalhos têm sido realizados com o
objetivo de verificar os efeitos da qualidade de luz ou, mesmo, de diferentes níveis
de irradiância no crescimento e no desenvolvimento de espécies cultivadas in vitro
(BRAGA et al., 2009). No entanto, Zanandrea et al. (2009), estudando a influência
de diferentes densidades de fluxo de fótons no cultivo in vitro de macieira, obteve as
melhores respostas com o uso de 14 mmol m-2 s-1, o que é considerado baixo,
comparado aos 48 μmol m-2 s-1 comumente utilizados na sala de crescimento.
Assim, isso justifica a possibilidade de que, uma redução na luminosidade possa ter
causado o aumento no número das brotações verificado no presente estudo.
5 Conclusões
De acordo com os resultados obtidos neste trabalho, pode-se concluir que:
– O meio mais apropriado para a multiplicação in vitro das cultivares é o MS, tanto
na forma comercial como o preparado em laboratório, mesmo quando utilizando
ferro na forma férrica ou ferrosa;
– Ambas cultivares necessitam de suplementação de citocinina exógena para a
proliferação de brotações (1,2 mg L-1 de BAP);
– As cultivares em estudo respondem diferentemente quanto à concentração de
sacarose utilizada no processo de multiplicação in vitro, sendo a cultivar Ya-Li mais
exigente (45 g L-1) que a cv. Carrick (30 g L-1).
6 Referências bibliográficas
ARAÚJO, A. G. de; PASQUAL, M.; CASTRO, E. M. de; RODRIGUES, F. A.; SANTOS, D. N. dos; DUTRA, L. F. Efeito da concentração de sacarose e qualidade de luz na propagação in vitro de plântulas de orquídea. Plant Cell Culture Micropropagation, v.3, n.2, p.96-102, 2007. BELL, R. L., SRINIVASAN, C., LOMBERK, D. Effect of nutrient media on axillary shoot proliferation and preconditioning for adventitious shoot regeneration of pears. In Vitro Cell Dev Biol-Plant, v.45, p.708–714, 2009. BORGHEZAN, M.; MORAES, L. K. A.; MOREIRA, F. M.; SILVA, A. L. Propagação in vitro e avaliação de parâmetros morfofisiológicos de portaenxertos de videira. Pesquisa Agropecuária Brasileira, v.38, n.7, p.783-789, 2003.

41
BOTELHO, R. V.; MAIA, A. J.; PIRES, E. J. P.; TERRA, M. M.; SCHUCK, E. Efeitos de reguladores vegetais na propagação vegetativado portaenxerto de videira VR043-43 (Vitis vinifera x Vitis rotundifolia). Revista Brasileira de Fruticultura, v.27, p.6-8, 2005. BRAGA, F. T; PASQUAL, M.; CASTRO, E. M. de; DIGNART, S. L.; BIAGIOTTI, G.; PORTO, J. M. P. Qualidade de luz no cultivo in vitro de Dendranthema grandiflorum cv. Rage: características morfofisiológicas. Ciência Agrotécnica, v.33, n.2, p.502-508, 2009. CAMPOS, R. V.; BIANCHI, V. J.; ROCHA, P. S. G. da; SCHUCH, M. W.; FACHINELLO, J. C. BAP na multiplicação in vitro de portaenxertos de Prunus spp. Plant Cell Culture Micropropagation, v.3, n.2, p.55-60, 2007. CARVALHO, A. C. P. P. de; TORRES, A. C.; BRAGA, E. J. B.; LEMOS, E. P. de; SOUZA, F. V. D.; PETERS, J. A.; WILLADINO, L.; CÂMARA, T.R. Glossário de Cultura de Tecidos de Plantas. Plant Cell Culture & Micropropagation, v.07, p.30-60, 2011. CARVALHO, J. M. F. C.; SILVA, M. M. A.; MEDEIROS, M. J. L. Fatores inerentes à micropropagação. Campina Grande: Embrapa Algodão, 2006. 5p. (Documentos 148). CENTOFANTE, A. R.; CASTRO, E. M. de; ABBADE, L. C.; PAIVA, R.; CENTOFANTE, E. Germinação in vitro de sementes de Merremia tomentosa Hallier F.: Influência de meios de cultura e GA3 Plant Cell Culture Micropropagation, v.5, n.2, p. 135-139, 2009. DANTAS, A. C. de M.; NESI, A. N.; MACHADO, L. B.; HAERTER, J.; FORTES, G. R. de L. Estabelecimento e multiplicação in vitro de cultivares de Pyrus spp. Revista Brasileira de Agrociência, v.8, n.1, p.19-23, 2002. DZAZIO, P. M.; BIASI, L. A.; ZANETTE, F. Micropropagação do portaenxerto de videira „420A‟. Revista Brasileira de Fruticultura, v.24, n.3, p.759-764, 2002. FACHINELLO, J. C.; FRANCESCATTO, P. Compatibilidade de enxerto e porta-enxerto de pereira. In: XI ENFRUTE (Encontro Nacional sobre Fruticultura de Clima Temperado), 1, 2009, Fraiburgo, SC. Anais... Caçador: EPAGRI, 2009. p.33-42. FADEL, D.; KINTZIOS, S.; ECONOMOU, A. S.; MOSCHOPOULOU, G.; CONSTANTINIDOU, H-I. A. Effect of different strength of medium on organogenesis, phenolic accumulation and antioxidant activity of Spearmint (Mentha spicata L.). The Open Horticulture Journal, v.3, p.31-35, 2010. GASPAR, T.; KEVERS, C.; BISBIS, B.; FRANCK, T. Special Symposium: in vitro plant recalcitrance loss of plant organogenic totipotency in the course of in vitro neoplastic progression. In Vitro Cellular and Developmental Biology-Plant, v.36, p.171-181, 2000.

42
GEORGE, F. E.; HALL, M. A.; DE KLERK, G. J. Plant propagation by tissue culture. 3.ed. The Background, v.1, 2008. 473 p. GRIMALDI, F.; GROHSKOPF, M. A.; MUNIZ, A. W.; GUIDOLIN, A. F. Enraizamento in vitro de frutíferas da família Rosaceae. Revista de Ciências Agroveterinárias, v.7, n.2, p.160-168, 2008. HASSANEN, S. A.; GABR, M. F. In vitro propagation of pear Pyrus betulaefolia rootstock. American-Eurasian Journal Agriculture e Environment Science, v.7, n.12, p484-489, 2012. HAZARIKA, B. N. Morpho-physiological disorders in in vitro culture of plants. Scientia Horticulturae, v.108, n.2, p.105-120, 2006. IBGE – Instituto Brasileiro de Geografia e Estatística, Levantamento Sistemático da Produção Agrícola 2009, http:www.ibge.gov.br, 2011. JHA, T. B.; GHOSH, B. Plant Tissue Culture: Basic and Applied. India: Universities Press (India) Private Limited. 2006. 206p. KOZAI, T.; KUBOTA, C.; JEONG, B. R. Environmental control for the large-scale production of plants through in vitro techniques. Plant Cell, Tissue and Organ Culture, v.51, 49-56, 1997. LLOYD, G.; McCOWN, B. Commercially-feasible micropropagation of mountain laurel, Kalmia latifólia, by use of shoot-tip culture. Combined Proceedings International Plant Propagators Society, v.30, p.421-427, 1980. MACHADO, A. A.; CONCEIÇÃO, A. R. Sistema de análise estatística para Windows. Winstat. Versão 1.0. UFPel, 2007. MACHADO, M. P.; CIOTTA, M. N.; DESCHAMPS, C.; ZANETTE, F.; COCCO, L. C.; BIASI, L. A. Propagação in vitro e caracterização química do óleo essencial de Lavandula angustifólia cultivada no Sul do Brasil. Ciência Rural, v.43, n.2, p.283-289, 2013. MACHADO, M. P.; BIASI, L. A.; RITTER, M.; RIBAS, L. L. F.; KOEHLER, H. S. Multiplicação in vitro do portaenxerto de videira „VR043-43‟ (Vitis vinifera x Vitis rotundifolia). Ciência e Agrotecnologia, v.30, n.4, p.648-655, 2006. MORAES, L. K. A. de; FELISBINO, C.; CRESTANI, L.; SILVA, A. L. da. Estabelecimento e multiplicação in vitro de Pyrus calleryana D-6 em sistema de cultura dupla-fase‟ . Revista Brasileira de Fruticultura, v.26, n.3, p. 403-405, 2004. MUNIZ, J. N.; KRETZSCHMAR, A. A.; RUFATO, L. Importância econômica da cultura. In: RUFATO, L.; KRETZSCHMAR, A. A.; BOGO, A. A cultura da pereira. Florianópolis: DIOESC, 2012. 247p. (Série Fruticultura) MURASHIGE, T.; SKOOG, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiologia Plantarum, v.15, p.473-479, 1962.

43
NAKASU, B. H.; FAORO, I. D. Cultivares. In: NAKASU, B. H.; CENTELLAS-QUEZADA, A.; HERTER, F. G. Pêra. Produção. Brasília: Embrapa Informação Tecnológica, 2003. p.29-36 (Frutas do Brasil, 46). OLIVEIRA, R.P.de; NINO, A.F.P.; NICKEL, O. Limpeza de patógenos e propagação in vitro de Cultivares de Pereira. Pelotas: Embrapa Clima Temperado, 2004. 5p. (Comunicado Técnico 105). PASQUAL, M. Introdução: fundamentos básicos. Lavras: FAEPE, 2001. 97 p. PATI, P. K.; RATH, S. P.; SHARMA, M.; SOOD, A.; AHUJA; P. S. In vitro propagation of rose: a review. Biotechnology Advances, v.24, p.94-114, 2006. PÉREZ-TORNERO, O.; BURGOS, L. Different media requirements for micropropagation of apricot cultivars. Plant Cell, Tissue and Organ Culture, v.63, p.133-141, 2000. PIO, R.; RAMOS, J. D.; MENDONÇA, V.; SILVA, A. B.; PASQUAL, M. Enraizamento in vitro de brotações do portaenxerto de citros Tangerina sunki x Trifoliata english 63-256 como uso de sacarose e ácido indolbutírico. Ciência e Agrotecnologia, v.26, n.1, p.66-70, 2002. QUOIRIN, M.; LEPOIVRE, P. Etude de milieux adaptes aux cultures in vitro de Prunus. Acta Horticulturae, v.78, p.437-442, 1977. RADMANN, E. B.; BIANCHI, V. J.; SOUZA, T. M.; FACHINELLO, J. C.; OLIVEIRA, R. P. Influência da composição do meio de cultivo e do tipo de explante na micropropagação do portaenxerto de Prunus sp. „GxN-9‟. Scientia Agraria, v.10, n.2, p.95-101, 2009. RADMANN, E. B.; BRAGA, E. J. B.; KARAN, M. A. L.; POSADA, M. A. C.; PETERS, J. A. Influência da densidade de fluxo luminoso na qualidade de plantas micropropagadas de Gypsophila paniculata L. Revista Brasileira de Agrociência, v.7, 171-175, 2001. RAMAGE, C. M.; WILLIAMS, R. R. Cytokinin-induced abnormal shoot organogenesis is associated with elevated Knotted1-type homeobox gene expression in tobacco. Plant Cell Reports, v.22, p.919-924, 2004. REED, B. M.; WADA, S.; DENOMA, J.; NIEDZ, R. P. Improving in vitro mineral nutrition for diverse pear germplasm. In Vitro Cellular Development Biology Plant. v.49, p.343–355, 2013a. REED, B. M.; DENOMA, J.; WADA, S.; POSTMAN, J. Micropropagation of pear (Pyrus sp.). Methods in Molecular Biology, v.11013, p.3-18, 2013b. ROGALSKI, M.; MORAES, L. K. A.; FELISBINO, R. C.; CRESTANI, L.; GUERRA, M. P.; SILVA, A. L. Enraizamento in vitro de portaenxertos de Prunus sp. Revista Brasileira de Fruticultura, v.25, n.2, p.293-296, 2003.

44
RUSIC, D. J.; VUJOVIC, T.; NIKOLIC, D.; CEROVIC, R. In vitro growth responses of the „Pyrodwarf‟ pear rootstock to cytokinin types. Romanian Biotechnological Lette, n.5, v.16, p.6631-6637, 2011. SCHMIDT, W. Iron solutions: acquisition strategies and signaling pathways in plants. TRENDS in Plant Science, v.8, p.188-193, 2003. SILVEIRA, C. A. P.; FACHINELLO, J. C.; FORTES, G. R. de L.; CITADIN, I.; RODRIGUES, A. C.; QUEZADA, A. C.; SILVA, J. B. Multiplicação in vitro de porta-enxertos do gênero Prunus sob diferentes concentrações de BAP em dois meios de cultura. Revista Brasileira de Fruticultura, v.23, n.3, p.488-492, 2001. SOTIROPOULOS, T. E.; FOTOPOULOS, S.; DIMASSI, K. N.; TSIRAKOGLOU, V.; THERIOS, I. N. Response of the pear rootstock to boron and salinity in vitro. Biologia Plantarum, v.50, n.4, p779-781, 2006. SOUZA, S. da S.; JUNGHANS, T. G. Introdução à micropropagação de Plantas. Cruz das Almas, BA: EMBRAPA, v.1, 2006. p. 79-96. TORRES, A.C.; REIS, N. V. B. dos; WILLADINO, L.; GUERRA, M. P.; FERREIRA, C. F.; PAIVA, S. A. V. de. Meios e condições de incubação para a cultura de tecidos de plantas: formulações de meio de cultura de tecidos de plantas. Brasília: EMBRAPA, 2001. 19p. (Circular técnica, 24). WANG,Y. ; WU, W. H. Potassium Transport and Signaling in Higher Plants. Annual Review Plant Biology, v.64, p.451–76, 2013. WREGE, M. S.; HERTER, F. G.; CAMELATTO, D.; STEINMETZ, S.; JÚNIOR, C. R.; GARRASTAZU, M. C.; FLORES, C. A.; IUCHI, T.; BERNARDI, J.; VERÍSSIMO, V.; MATZENAUER, R. Zoneamento agroclimático para pereira no Rio Grande do Sul. Pelotas: Embrapa Clima Temperado, 2006. 29p. (Embrapa Clima Temperado. Documentos, 182). VILLA, F.; PASQUAL, M.; ASSIS, F. A.; PIO, L. A. S. Sacarose e um aditivo orgânico complexo na micropropagação de amoreira-preta (Rubus sp.). Plant Cell Culture and Micropropagation, v.5, n.1, p.1- 8, 2009. XU, G.; FAN,X.; MILLER, A.J. Plant nitrogen assimilation and use efficiency. Annual Reviews Plant Biology, n.63, p.153–182, 2012. ZANANDREA, I.; BACARIN, M. A.; BRAGA, E. J. B.; BIANCHI, V. J.; PETERS, J. A. Morphological and Physiological photon flux influence under in vitro culture of apple shoots. Brazilian Archives of Biology andTechnology, v.52 n.5, p.1091-1098, 2009. ZHANG, L.; XU, T.; SUN, X.; ZHANG, H.; TANG, T.; TANG, K. Factors influencing shoot regeneration from cotyledons of tetraploid Isatis indigotica F. In Vitro Cellular & Developmental Biology - Plant, v.39, n.05, p.459-462, 2003.

45
CAPÍTULO 2 – Multiplicação in vitro de pereira, cv. Cascatense
1 Introdução
Por ser uma fruta de grande aceitação para o consumo in natura, a produção
de pera (Pyrus sp.) é realizada em todo o mundo, o que a torna de grande
importância nos mercados internacionais (FAO, 2011). No Brasil, entre as frutas de
clima temperado, é a terceira mais consumida, embora seja a fruta fresca importada
em maior quantidade, aproximadamente 140.000 ton., o que representa mais de
90% do total consumido, devido a uma menor expressão em termos de produção e
área cultivada (NAKASU; FAORO, 2003; NAKASU et al., 2007; FACHINELLO;
FRANCESCATTO, 2009; FACHINELLO et al., 2011). Segundo Nakasu; Faoro
(2003) e Tomaz et al. (2009), o principal entrave para o bom desenvolvimento e
expansão da cultura é a falta de cultivares adaptadas às condições edafoclimáticas
das regiões potencialmente produtoras.
Os estados do Rio Grande do Sul e Santa Catarina, além da capacidade de
aproveitamento das estruturas de pós-colheita, conservação e canais de
comercialização utilizados na cadeia produtiva da maçã, são os que apresentam
condições climáticas favoráveis à produção da pereira (JUNQUEIRA; PEETZ, 2003).
Entretanto, em muitos locais e em determinados anos, a quantidade de horas de frio
hibernal é insuficiente para que ocorra uma boa e uniforme brotação e floração,
gerando, com isso, baixa produtividade e qualidade das frutas. Assim, cultivares
recomendadas para essas regiões devem apresentar média a baixa exigência em
frio (FAORO, 2002; SATO; ASSUMPÇÃO, 2003).
Neste contexto, o Programa de Melhoramento Genético da Embrapa Clima
Temperado lançou a cv. Cascatense, resultante do cruzamento de „Packham‟s
Triumph‟ x „Le Conte‟, realizado em 1968, selecionada em 1982 como Pirus 9 e
lançada como cultivar em 1992. A planta é de vigor médio a semi-vigorosa, semi-

46
aberta e altamente produtiva, apresentando brotações mais precoces e uniformes,
enquanto que a fruta é piriforme e de tamanho médio. Além disso, sua exigência em
frio hibernal situa-se ao redor de 400 h (temperatura ≤ 7,2 °C) (NAKASU et al., 2007;
NAKASU; FAORO, 2003; NAKASU; LEITE, 1992).
A propagação do material vegetal, almejando à produção de mudas com
qualidade e uniformidade pode ser realizada através da micropropagação ou
propagação in vitro, em vista de ser um método biotecnológico que se constitui na
produção de um elevado número de plantas, num curto espaço de tempo, ocupando
uma área física bastante reduzida, sendo realizado durante o ano todo e com alta
qualidade sanitária (SERAFINI, et al. 2001; CARVALHO et al., 2011).
Protocolos de micropropagação vêm sendo desenvolvidos para que as
condições de cultivo in vitro sejam otimizadas, possibilitando a obtenção de taxas
elevadas de multiplicação, e consequentemente, qualidade nas mudas formadas
(ROGALSKI et al., 2003; GUERRA; NODARI, 2006). Para que ocorra um adequado
crescimento e desenvolvimento das plantas durante a fase de multiplicação do
material vegetal, é necessário o controle da morfogênese, a qual é influenciada por
vários fatores como espécie, cultivar, tipo de explante, componentes do meio
nutritivo, reguladores de crescimento adicionados ao meio e ambiente de cultivo
(CARVALHO et al., 2006).
Os meios de cultivo são compostos por sais minerais (macro e
micronutrientes), carboidratos, vitaminas e reguladores de crescimento, podendo ser
líquidos ou semi-sólidos (solidificados com ágar ou outro geleificante) (GUERRA;
NODARI, 2006). Existem diversos meios de cultura, variando as concentrações de
sais e vitaminas de acordo com as necessidades da espécie ou cultivar, porém os
mais utilizados são o clássico MS (MURASHIGE; SKOOG, 1962) e suas
modificações e diluições; assim como o WPM [Wood Plant Medium (LLOYD;
MCCOWN, 1980)] e o QL (QUOIRIN; LEPOIVRE, 1977), cujas formulações são
consideradas mais fracas.
A concentração da fonte de carbono utilizada no meio é outro fator importante
durante a multiplicação dos explantes, pois os carboidratos fornecem energia e
contribuem para o equilíbrio do potencial osmótico do meio de cultura (PATI et al.,
2006). Em geral, a sacarose é utilizada preferencialmente, porque é o carboidrato
predominante na seiva do floema da grande maioria das plantas e também a mais
barata (LEITE et al., 2000).

47
Em relação aos reguladores de crescimento, a aplicação de uma fonte de
citocinina no meio de multiplicação é indispensável para a superação da dominância
apical do explante e induzir a proliferação de gemas axilares. Dentre as citocininas,
a 6-benzilaminopurina (BAP) é a mais utilizada, sendo considerada um fitorregulador
responsável pela divisão celular e crescimento da parte aérea, além de se destacar
das demais por ter um menor custo (PEREZ-TORNERO; BURGOS, 2000).
Diante do exposto, buscou-se neste trabalho verificar a influência dos
diferentes meios de cultivo, concentrações de sacarose e BAP na multiplicação in
vitro da cv. Cascatense, visando obter um protocolo eficiente para potencializar sua
propagação/reprodutibilidade.
2 Material e Métodos
Os explantes utilizados para o desenvolvimento do trabalho foram
provenientes de brotações de pereira, cv. Cascatense, pré-estabelecidas e
multiplicadas in vitro, durante 40 dias, em meio de cultivo MS pronto Himédia® (meio
comercial com formulação igual ao MS), acrescido de 0,8 mg L-1 de BAP), 30 g L-1
de sacarose e 7 g L-1 de ágar, sendo o pH ajustado para 5,8.
Os estudos foram conduzidos em três experimentos sequenciais, conforme
descrito abaixo, sendo que cada experimento foi repetido duas vezes. Para a
realização dos experimentos, como explante inicial, foram utilizados segmentos
nodais de aproximadamente 1,0 cm contendo de duas a três gemas axilares. As
gemas axilares presentes nos segmentos nodais, bem como o comprimento destes
foram incluídos nas avaliações realizadas. Gemas e brotações incipientes, menores
que três milímetros, não foram consideradas por serem muito pequenas e de difícil
manuseio.
Experimento 1 – Influência de diferentes meios de cultivo e concentrações de
sais
Os explantes iniciais foram cultivados em cinco meios diferentes: MS
completo e MS modificado (¾ da concentração normal de NH4NO3 e KNO3,
identificado nos resultados como MS¾), MS Himédia, WPM e QL; todos contendo
0,8 mg L-1 de BAP, 100 mg L-1 de mio-inositol, 30 g L- 1 de sacarose, 7 g L-1 de ágar
e pH 5,8. Os meios MS completo, MS¾ e QL foram preparados com EDTA-Férrico,

48
enquanto o MS Himédia e WPM com EDTA-Ferroso, conforme formulação original
do meio MS.
Os frascos foram mantidos em câmara de crescimento com temperatura de
23±2ºC, fotoperíodo de 16 horas e 48 μmol m-2 s-1 de fluxo de fótons por 40 dias.
Após este período, avaliou-se o número de brotações por explante, o comprimento
das brotações (cm), o número de gemas axilares por brotação, massa fresca e
massa seca das brotações (g), sendo a secagem do material realizada em estufa de
ventilação forçada a ±50 ºC até massa constante.
Experimento 2 – Efeito das concentrações de 6-benzilaminopurina (BAP)
O meio de cultura usado foi o que apresentou melhores resultados após a
realização do primeiro experimento (MS Himédia), suplementado com diferentes
concentrações de BAP (0; 0,4; 0,8; 1,2 e 1,6 mg L-1), 30 g L-1 de sacarose, 7 g L-1 de
ágar e pH 5,8. As condições de cultivo e as variáveis analisadas foram as mesmas
do experimento anterior.
Experimento 3 – Efeito de diferentes concentrações de sacarose
O meio de cultivo e a concentração de BAP utilizados foram os que
apresentaram melhores resultados após a realização do primeiro e segundo
experimentos (MS Himédia e 0,8 mg L-1 de BAP), variando-se a concentração de
sacarose (0, 15, 30, 45 e 60 g L-1), com 7 g L-1 de ágar e pH 5,8. As condições de
cultivo e as variáveis analisadas foram as mesmas dos experimentos anteriores.
Delineamento experimental e análise estatística
O delineamento experimental adotado para todos os experimentos foi
inteiramente casualizado, com nove repetições por tratamento, representadas, cada
uma, por um frasco contendo cinco explantes. Para as avaliações de massa fresca e
massa seca das brotações foram selecionadas, ao acaso, três plantas de cinco
repetições, totalizando quinze plantas por tratamento.
Os experimentos foram compostos por apenas um fator de tratamento, com
cinco níveis: meio de cultura (MS, MS¾, MS Himédia, WPM e QL) para o primeiro
experimento, concentração de BAP (0; 0,4; 0,8; 1,2 e 1,6 mg L-1) para o segundo
experimento e concentração de sacarose (0, 15, 30, 45 e 60 g L-1) para o terceiro
experimento.

49
Os resultados obtidos foram submetidos à análise de variância e as médias
comparadas pelo teste de Tukey, ao nível de 5% de probabilidade de erro, ou
analisadas por regressão polinomial, utilizando o software WINSTAT 1.0
(MACHADO; CONCEIÇÃO, 2007).
3 Resultados
Segundo a análise de variância e/ou regressão polinomial, observou-se
diferenças significativas entre os tratamentos para todas as variáveis analisadas,
nos três experimentos.
Experimento 1 – Influência de diferentes meios de cultivo e concentrações de
sais
O maior número de brotações por explante (6,18) foi observado no meio WPM
(Fig. 1A). Em contrapartida, os maiores valores para comprimento (2,5 cm), massa
fresca (0,39 g), massa seca (0,06 g) e número de gemas axilares por brotação
(13,17) foi obtido no meio Himédia (Fig. 1B, C, D e E), o qual não diferiu
estatisticamente dos meios MS (12,1) e MS¾ (11,83) para esta última variável. O
meio de cultivo QL não pode ser avaliado quanto ao número de gemas axilares por
brotação devido ao entufamento das brotações ocasionada pela redução no número
e comprimento das mesmas, não permitindo sua contagem.
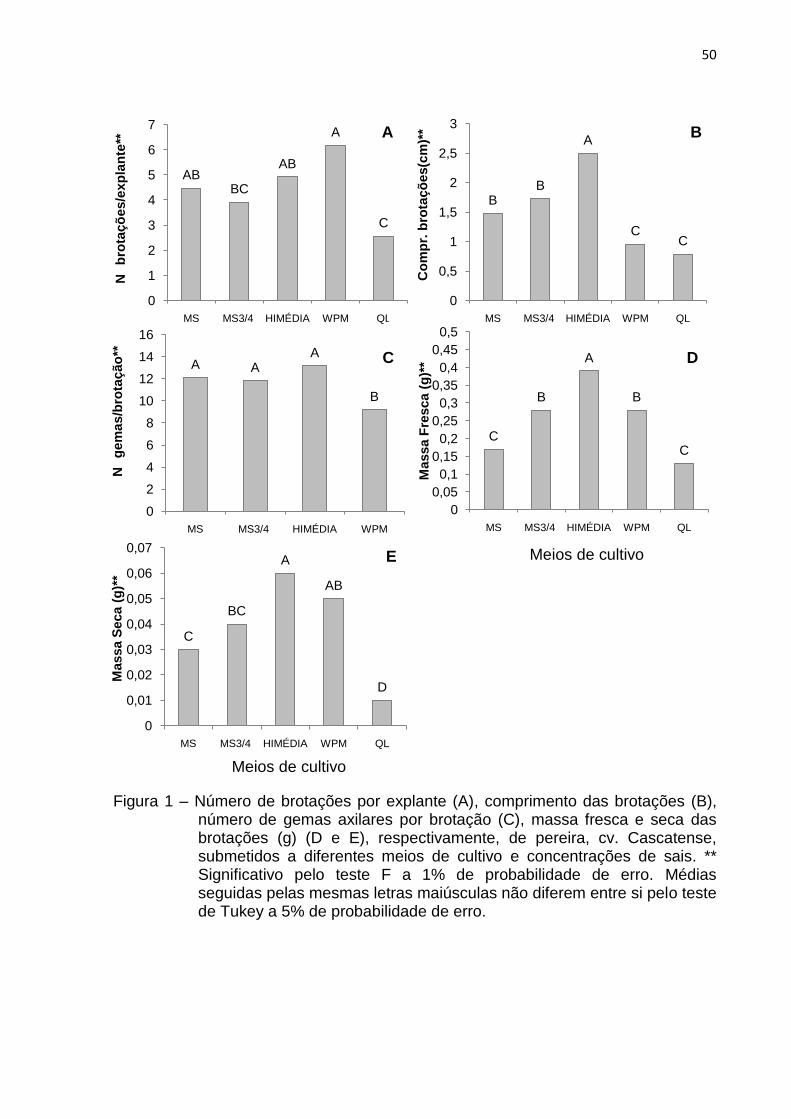
50
ABBC
AB
A
C
0
1
2
3
4
5
6
7
MS MS3/4 HIMÉDIA WPM QL
Nb
rota
çõ
es/e
xp
lan
te**
C
BC
A
AB
D
0
0,01
0,02
0,03
0,04
0,05
0,06
0,07
MS MS3/4 HIMÉDIA WPM QL
Massa S
eca (
g)*
*
C
B
A
B
C
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0,45
0,5
MS MS3/4 HIMÉDIA WPM QL
Massa F
resca (
g)*
*
Figura 1 – Número de brotações por explante (A), comprimento das brotações (B), número de gemas axilares por brotação (C), massa fresca e seca das brotações (g) (D e E), respectivamente, de pereira, cv. Cascatense, submetidos a diferentes meios de cultivo e concentrações de sais. ** Significativo pelo teste F a 1% de probabilidade de erro. Médias seguidas pelas mesmas letras maiúsculas não diferem entre si pelo teste de Tukey a 5% de probabilidade de erro.
BB
A
CC
0
0,5
1
1,5
2
2,5
3
MS MS3/4 HIMÉDIA WPM QL
Co
mp
r. b
rota
çõ
es(c
m)*
*
A AA
B
0
2
4
6
8
10
12
14
16
MS MS3/4 HIMÉDIA WPM
Ng
em
as/b
rota
ção
** C D
E
A B
Meios de cultivo
Meios de cultivo

51
Experimento 2 – Efeito das concentrações de 6-benzilaminopurina (BAP)
A adição de BAP ao meio de cultivo resultou em uma resposta quadrática
para as variáveis número de brotações por explante, comprimento, massa fresca e
massa seca das brotações.
Para as variáveis número de brotações por explante e comprimento das
brotações, embora o ponto de máxima tenha ocorrido nas concentrações de 1,02 e
1,26 mg L-1 de BAP, com 4,95 brotações formadas e 2,28 cm de comprimento,
respectivamente, o maior número de brotações por explante (5,2) foi obtido na
concentração de 0,8 mg L-1 (Fig. 2A), enquanto que o maior comprimento das
mesmas foi verificado quando o meio foi suplementado com 1,6 mg L-1 desta
citocinina (2,35 cm), sendo que este resultado não é tão superior ao encontrado
quando utilizada a concentração de 0,8 mg L-1 de BAP (2,09 cm) (Fig. 2B).
Em relação à massa fresca e massa seca, pela estimativa da equação de
regressão, os maiores valores (0,4 e 0,06 g) ocorreram nas concentrações de 1,28 e
1,18 mg L-1 de BAP, consecutivamente (Fig. 2D e 2E). Porém, para massa seca
verificou-se que a partir de 0,8 mg L-1 de BAP já era obtido esse valor (0,06 g), não
variando nas concentrações mais elevadas.
No que se refere ao número de gemas axilares por brotação (Fig.2C), houve
uma resposta linear crescente, com maiores valores obtidos na concentração de 1,6
mg L-1, apresentando uma média 16,68 gemas formadas.
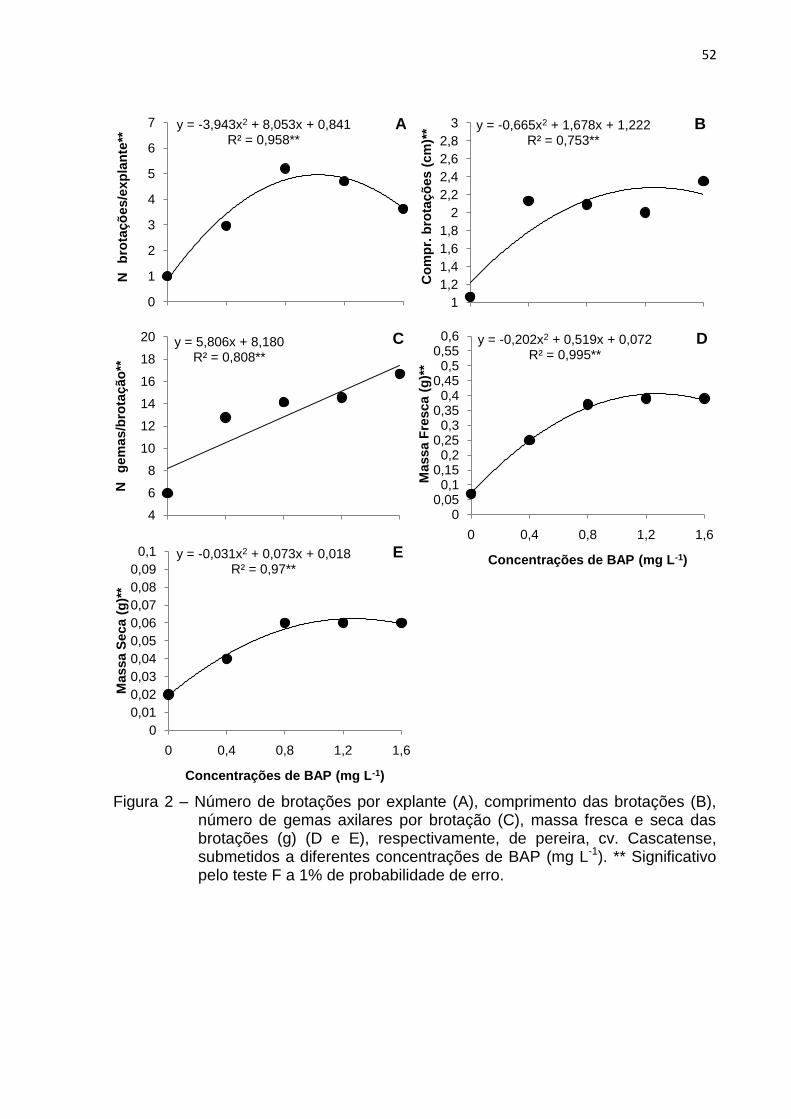
52
y = -3,943x2 + 8,053x + 0,841R² = 0,958**
0
1
2
3
4
5
6
7N
bro
taçõ
es/e
xp
lan
te**
y = 5,806x + 8,180R² = 0,808**
4
6
8
10
12
14
16
18
20
Ng
em
as/b
rota
ção
**
y = -0,202x2 + 0,519x + 0,072R² = 0,995**
00,05
0,10,15
0,20,25
0,30,35
0,40,45
0,50,55
0,6
0 0,4 0,8 1,2 1,6
Massa F
resca (
g)*
*
Concentrações de BAP (mg L-1)y = -0,031x2 + 0,073x + 0,018R² = 0,97**
0
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,09
0,1
0 0,4 0,8 1,2 1,6
Massa S
eca (
g)*
*
Concentrações de BAP (mg L-1)
Figura 2 – Número de brotações por explante (A), comprimento das brotações (B),
número de gemas axilares por brotação (C), massa fresca e seca das brotações (g) (D e E), respectivamente, de pereira, cv. Cascatense, submetidos a diferentes concentrações de BAP (mg L-1). ** Significativo pelo teste F a 1% de probabilidade de erro.
y = -0,665x2 + 1,678x + 1,222R² = 0,753**
1
1,2
1,4
1,6
1,8
2
2,2
2,4
2,6
2,8
3
Co
mp
r. b
rota
çõ
es (
cm
)** A B
C D
E

53
Experimento 3 – Efeito de diferentes concentrações de sacarose
Quando adicionada sacarose ao meio de cultivo, verificou-se uma tendência
quadrática para todas as variáveis analisadas. Os maiores valores estimados para
número de brotações por explante (4,95) e comprimento das brotações (2,06 cm)
ocorreram nas concentrações de 32,13 e 29 g L-1 de sacarose, respectivamente.
Entretanto, em valores numéricos, observou-se que o uso de 30 g L-1 deste
carboidrato induziu as melhores respostas, com a formação de 5,1 brotações (Fig.
3A) de 2,2 cm de comprimento cada (Fig. 3B).
Segundo a equação de regressão, maiores valores para número de gemas
axilares por brotação (13,8) (Fig. 3C) e massa fresca (0,368 g) (Fig. 3D) foram
obtidos nas concentrações de 29,03 e 33,93 g L-1 de sacarose, consecutivamente.
Porém, estes valores não são tão superiores quando utilizada a concentração de 30
g L-1 de sacarose, com 13,52 gemas axilares formadas e obtenção de 0,36 g massa
fresca.
Com relação à massa seca, embora o ponto de máxima tenha inferido que a
concentração ideal seja 36,83 g L-1 de sacarose, com obtenção de 0,048 g (Fig. 3E),
verificou-se, em valores absolutos, que as médias foram maiores quando
suplementados 30 e 45 g L-1 deste açúcar ao meio de cultivo (0,05 g).
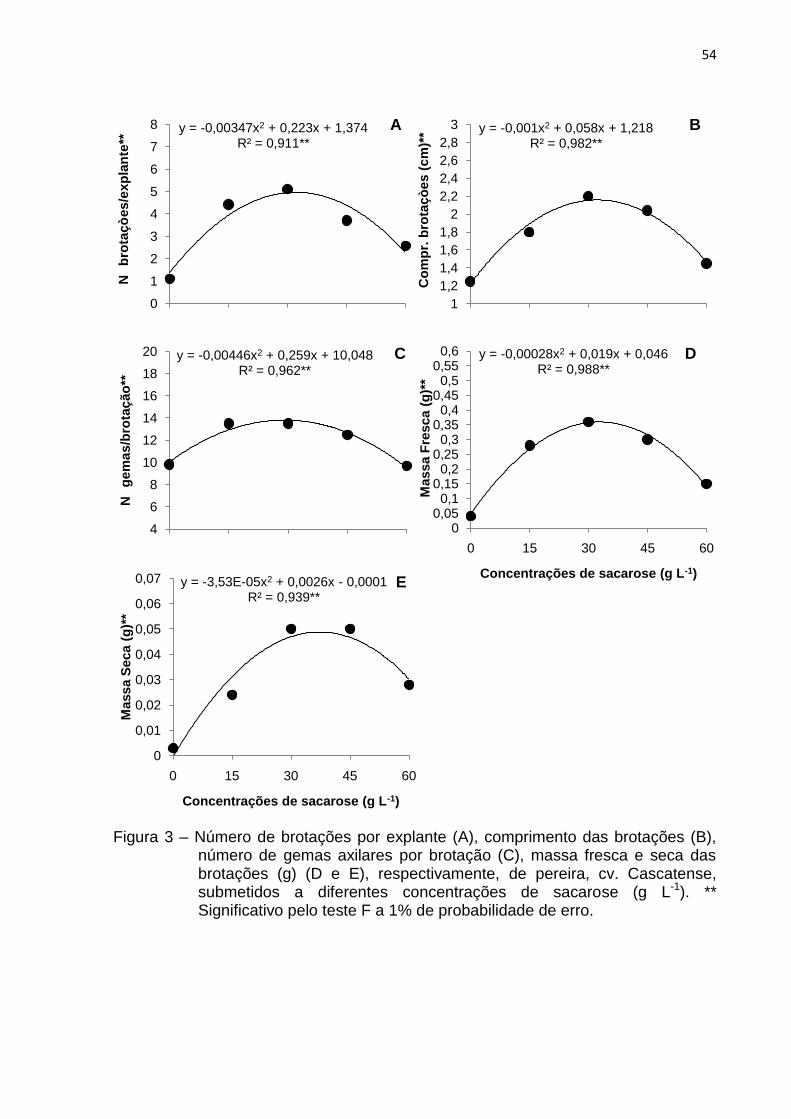
54
y = -3,53E-05x2 + 0,0026x - 0,0001 R² = 0,939**
0
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0 15 30 45 60
Massa S
eca (
g)*
*
Concentrações de sacarose (g L-1)
y = -0,001x2 + 0,058x + 1,218R² = 0,982**
1
1,2
1,4
1,6
1,8
2
2,2
2,4
2,6
2,8
3
Co
mp
r. b
rota
çò
es (
cm
)**
y = -0,00446x2 + 0,259x + 10,048R² = 0,962**
4
6
8
10
12
14
16
18
20
Ng
em
as/b
rota
ção
**y = -0,00347x2 + 0,223x + 1,374
R² = 0,911**
0
1
2
3
4
5
6
7
8N
bro
taçò
es/e
xp
lan
te**
Figura 3 – Número de brotações por explante (A), comprimento das brotações (B), número de gemas axilares por brotação (C), massa fresca e seca das brotações (g) (D e E), respectivamente, de pereira, cv. Cascatense, submetidos a diferentes concentrações de sacarose (g L-1). ** Significativo pelo teste F a 1% de probabilidade de erro.
y = -0,00028x2 + 0,019x + 0,046R² = 0,988**
00,05
0,10,15
0,20,25
0,30,35
0,40,45
0,50,55
0,6
0 15 30 45 60
Massa F
resca (
g)*
*
Concentrações de sacarose (g L-1)
A B
C D
E

55
4 Discussão
As taxas de crescimento e desenvolvimento de plantas cultivadas in vitro são
geneticamente determinadas, embora sejam limitadas pelo microambiente físico e
químico dos recipientes de cultivo, como excesso de nutrientes e alta concentração
de reguladores de crescimento, que podem comprometer a formação das plantas
(CASANOVA et al., 2008). Dessa forma, as quantidades fornecidas e o balanço de
nutrientes e reguladores de crescimento, bem como de carboidratos, são fatores
determinantes para o sucesso da multiplicação do material vegetal in vitro (ZHANG
et al., 2003; MORAES et al., 2004), como visualizado neste trabalho, a partir dos
resultados obtidos nos experimentos.
Conforme Guerra e Nodari (2006), a constituição de cada meio de cultivo
deve ser baseada nas exigências das plantas quanto aos nutrientes minerais para
atender necessidades específicas, em vista que os minerais exercem importante
papel não apenas no crescimento, mas também na regulação da morfogênese. Seu
fornecimento, absorção, transporte e metabolismo parecem variar entre os estágios
de desenvolvimento dos órgãos, iniciação dos meristemas e durante o crescimento
(RAMAGE; WILLIAMS, 2002). Baseado nisso, analisando-se a influência de
diferentes meios de cultivo e concentrações de sais durante a multiplicação da
cultivar em estudo (Fig. 4A), verificou-se que, em geral, os melhores resultados
foram obtidos em MS Himédia. Por se tratar de um meio pronto, comercial, essa
resposta positiva pode ser devido à qualidade dos sais minerais utilizados e/ou a
alterações na quantidade dos reagentes quando as soluções são preparadas no
laboratório.

56
Figura 4 – Brotações de Pyrus sp., cv. Cascatense, obtidas após serem submetidas a diferentes meios de cultivo e concentrações de sais (A), concentrações de BAP (B) e concentrações de sacarose (C).

57
Outro aspecto a ser considerado é que, tanto o MS Himédia como o WPM
foram preparados com Ferro na forma reduzida e mais solúvel, enquanto o MS
completo, MS¾ e QL, na forma de EDTA-Férrico – oxidada. Segundo Schmidt
(2003), o ferro é considerado um micronutriente essencial para o crescimento e
desenvolvimentos das plantas, estando envolvido em diversos processos, como na
respiração mitocondrial e fotossíntese, sendo que a forma absorvida pela maioria
das plantas é Fe2+ (forma mais solúvel), embora possa também ser absorvido como
Fe3+-quelato. Sendo assim, isto também pode explicar o aumento no número de
brotações por explante quando utilizado o WPM, no qual, em contrapartida,
observou-se menor comprimento das brotações. O aumento exacerbado no número
de brotações promove uma competição por sais e vitaminas presentes no meio de
cultivo, o que pode acarretar em redução do crescimento (OLIVEIRA et al., 2001).
Com o uso do meio nutritivo QL observou-se redução em todos os parâmetros
analisados, além da impossibilidade de avaliar o número de gemas por brotação, o
que pode estar relacionado ao fato deste apresentar apenas 25% da concentração
de nitrogênio (NH4NO3) em comparação ao MS, mostrando que composições muito
diluídas deste macronutriente não são favoráveis à multiplicação da cultivar
Cascatense. O nitrogênio é um dos principais nutrientes essenciais e ativos, sendo
absorvido, principalmente, na forma de nitrato (NO3-) e amônio (NH4
+) e é
constituinte de várias biomoléculas essenciais, como aminoácidos, ácidos nucléicos,
proteínas, enzimas e outros. Em razão disso, possui diferentes efeitos no
crescimento, no vigor do vegetal e na produção de biomassa (XU et al., 2012).
Neste contexto, analisando os diferentes meios de cultivo e concentrações de
sais utilizadas em diversos trabalhos, apesar de o WPM ter sido desenvolvido para o
cultivo de brotações de plantas lenhosas (BRUM, 2001), e apresente resultados
positivos na micropropagação de espécies como o mirtilo (SILVA et al., 2006) e em
Prunus sp. (RADMANN et al., 2009), e, embora as espécies e cultivares de pera
sejam cultivadas em vários meios de cultura conhecidos, segundo Reed et al.
(2013), o MS é o mais comumente usado. Assim, na ausência de MS pronto
Himédia, esse seria recomendado para a cv. Cascatense, mesmo com diluições na
concentração de NH4NO3 e KNO3, como pode ser visualizado na Fig. 4A.
Os resultados obtidos em relação à utilização de citocininas no meio de
cultivo (Fig. 4B) demonstraram que estas são indispensáveis na fase de
multiplicação in vitro, pois são responsáveis pela divisão celular, quebra da

58
dominância apical e indução da proliferação de gemas axilares (MOK et al., 2000).
Estes resultados são similares aos verificados por Moraes et al. (2004) em Pyrus
calleryana D-6, bem como Radmann et al. (2011), em Prunus sp., portaenxerto
„Flordaguard‟, os quais constataram que a ausência de BAP não estimula a emissão
de novos brotos, evidenciando assim uma dependência pela adição de citocinina
externa para a indução de brotações.
De acordo com o observado neste trabalho, a concentração adequada de
BAP para a multiplicação in vitro de „Cascatense‟ foi 0,8 mg L-1, pois, analisando
todas as variáveis, foi a que desencadeou melhores resultados, uma vez que é
importante obter brotações com comprimento adequado, visando facilitar seu
manuseio, separação e posterior utilização no enraizamento. Neste sentido, de
acordo com Dantas et al. (2002), para que a multiplicação in vitro seja considerada
eficiente, é necessário obter uma taxa média satisfatória de brotações em que sejam
priorizadas qualidade e homogeneidade, pois isso vai contribuir para o sucesso da
fase de enraizamento e, por conseguinte, para a produção de mudas.
Enquanto o uso de citocinina estimula maior produção de parte aérea, o
excesso é tóxico e compromete o desenvolvimento das culturas, podendo levar à
hiperidricidade (RAMAGE; WILLIAMS, 2004), que, segundo Santos-Serejo et al.
(2006), é um evento que pode ocorrer na cultura de tecidos, gerando anormalidades
fisiológicas e morfológicas no tecido vegetal, afetando os processos de fotossíntese
e trocas gasosas da planta. Os explantes vitrificados caracterizam-se por
apresentarem folhas translúcidas, túrgidas, rígidas, com aspecto encharcado e
facilmente quebráveis, apresentando brotações mal formadas e alongadas
(HAZARIKA, 2006), o que pode ser verificado no presente estudo, quando utilizada a
concentração mais elevada de BAP (1,6 mg L-1) (Fig. 4B).
No que tange ao uso de carboidratos, é clara sua função como fonte de
energia para a produção de novos metabólitos, além de atuar como substrato para o
processo de respiração celular (BORTOLINI, 2006; PIO et al., 2002). A concentração
destes no meio de cultivo é um dos parâmetros que reflete no
crescimento/desenvolvimento das brotações e pode apresentar correlação com a
habilidade das plantas ao enraizamento (BORTOLINI, 2006), uma vez que, para a
formação de raízes, a auxina requer fonte de carbono para a biossíntese de ácidos
nucléicos e proteínas. Assim, há necessidade de energia e carbono para a formação
das raízes (NORBERTO et al., 2001). A adição de carboidratos no meio de cultivo

59
também pode contribuir para o equilíbrio do potencial osmótico do meio, modificando
o potencial hídrico e, assim, regulando a absorção de água e nutrientes pelo
explante (LÉDO et al., 2007). Além disso, seu excesso pode ser prejudicial, pois
inibe a síntese de clorofila e, portanto, reduz a capacidade fotossintética das culturas
(HAZARIKA, 2006).
Diante do exposto, e conforme os resultados obtidos em relação ao efeito das
diferentes concentrações de sacarose durante a multiplicação in vitro da pereira
„Cascatense‟, verificou-se que a concentração de 30 g L-1 foi a mais efetiva, pois
desencadeou as melhores respostas, em vista que concentrações inferiores e
superiores acarretaram em redução no crescimento e desenvolvimento das
brotações, como pode ser visualizado na Fig. 4C. Embora a concentração de 3% de
sacarose seja a comumente suplementada in vitro na maioria das espécies, avaliar a
concentração adequada para cada espécie/cultivar é de suma importância devido a
sua função regulatória em muitos processos vitais da planta. Ainda, segundo De
Riek et al. (1997), 75% a 85% do aumento da biomassa se devem à incorporação de
carbono pela adição de sacarose ao meio de cultivo.
O sucesso na obtenção de um protocolo apropriado de propagação de
plantas em larga escala está intimamente relacionado às altas taxas de multiplicação
dos explantes. Assim, os parâmetros avaliados no decorrer dos experimentos
servem como ferramentas úteis para a verificação dessas taxas. Contudo, as
variáveis mais importantes são número e comprimento das brotações, bem como o
número de gemas axilares por brotação, pois estas, em conjunto, vão determinar
uma maior ou menor eficiência do processo de multiplicação. Além disso, conforme
preconizado por Radmann et al. (2009), o crescimento das brotações formadas
durante a fase de multiplicação é uma variável determinante na qualidade do
material destinado à fase de enraizamento.
5 Conclusões
Nas condições que o trabalho foi conduzido, pode-se concluir que o protocolo
mais eficiente para a multiplicação in vitro da cultivar em estudo é constituído pelo
MS Himédia comercial, com suplementação de 0,8 mg L-1 de BAP e 30 g L-1 de
sacarose.

60
6 Referências bibliográficas
BORTOLINI, M. F. Uso de ácido indol butírico na estaquia de Tibouchina sellowiana (Cham.) Cogn. 2006. 72f. Dissertação (Mestrado em Agronomia). Universidade Federal do Paraná, Curitiba. BRUM, G. R. Micropropagação da figueira (Ficus carica L.) Roxo de Valinhos. 2001. 41f. Dissertação (Mestrado em Fitotecnia) – Universidade Federal de Lavras, Lavras. CARVALHO, A. C. P. P. de; TORRES, A. C.; BRAGA, E. J. B.; LEMOS, E. P. de; SOUZA, F. V. D.; PETERS, J. A.; WILLADINO, L.; CÂMARA, T. R. Glossário de Cultura de Tecidos de Plantas. Plant Cell Culture & Micropropagation, v.07, p.30-60, 2011. CARVALHO, J. M. F. C.; SILVA, M. M. A.; MEDEIROS, M. J. L. Fatores inerentes à micropropagação. Campina Grande: Embrapa Algodão, 2006. 5p. (Documentos 148). CASANOVA, E.; MOYSSET, L.; TRILLAS, M. I. Effects of Agar concentration and vessel closure on the organogenesis and hyperhydricity of adventitious carnation shoots. Biologia Plantarum, v.52, p.1-8, 2008. DANTAS, A. C. de M.; NESI, A. N.; MACHADO, L. B.; HAERTER, J.; FORTES, G. R. de L. Estabelecimento e multiplicação in vitro de cultivares de Pyrus spp. Revista Brasileira de Agrociência, v.8, n.1, p.19-23, 2002. DE RIEK, J.; PIQUERAS, A.; DEBERGH, P. C. Sucrose uptake and metabolism in a double layer system for Micropropagation of Rosa multiflora. Plant Cell, Tissue and Organ Culture, v.47, n.03, p.269-278, 1997. FACHINELLO, J. C.; FRANCESCATTO, P. Compatibilidade de enxerto e porta-enxerto de pereira. In: XI ENFRUTE (Encontro Nacional sobre Fruticultura de Clima Temperado), 1, 2009, Fraiburgo, SC. Anais... Caçador: EPAGRI, 2009. p.33-42. FACHINELLO, J. C.; PASA, M. S.; SCHMTIZ, J. D.; BETEMPS, D. L. Situação e perspectivas da fruticultura de clima temperado no Brasil. Revista Brasileira de Fruticultura, v.33, n.1, p.109-120, 2011. FAO. Faostat Database. Prodstat. Disponível em: <http://www.fao.org>. Online. Acesso em 10 jul. 2011. FAORO, I. D.; NAKASU, B. H. Japanese pear growing in Brazil. Acta Horticulturae Wageningen, v.567, p.97-105, 2002. GUERRA, M. P.; NODARI, R. O. Apostila de biotecnologia – CCA/UFSC. Edição da Steinmacher, 2006. p1-48. HAZARIKA, B. N. Morpho-physiological disorders in in vitro culture of plants. Scientia Horticulturae, v.108, n.2, p.105-120, 2006.

61
JUNQUEIRA, A. H., PEETZ, M. S. Mercados interno e externo. Introdução. In: QUEZADA, A. C.; NAKASU, B. H; HERTER, F. G. (Ed.). Pêra Produção. Brasília: Embrapa Informação Tecnológica, 2003. p.10-19. (Frutas do Brasil, 46) LÉDO, A. S.; CUNHA, A. O.; ARAGÃO, W. M.; TUPINAMBÁ, E. A. Efeito da sacarose e do manitol na conservação in vitro por crescimento lento de coqueiro anão. Magistra, v.19, n.04, p.346-351, 2007. LEITE, G. B. U.; FINARDI, N.; FORTES, G. R. Efeitos de concentrações de sacarose no meio de cultura e da intensidade luminosa no enraizamento “in vitro” do porta-enxerto de Pereira OHxF971. Ciência agrotécnica, v.24, n.2, p.353-357, 2000. LLOYD, G.; McCOWN, B. Commercially-feasible micropropagation of mountain laurel, Kalmia latifólia, by use of shoot-tip culture. Combined Proceedings International Plant Propagators Society, v.30, p.421-427, 1980. MACHADO, A. A.; CONCEIÇÃO, A. R. Sistema de análise estatística para Windows. Winstat. Versão 1.0. UFPel, 2007. MORAES, L. K. A. de; FELISBINO, C.; CRESTANI, L.; SILVA, A. L. da. Estabelecimento e multiplicação in vitro de Pyrus calleryana D-6 em sistema de cultura dupla-fase‟ . Revista Brasileira de Fruticultura, v.26, n.3, p.403-405, 2004. MOK, M. C.; MARTIN, R. C.; MOK, D. W. S. Cytokinins: biosynthesis metabolism and percepcion. In Vitro Cellular & Developmental Biology Plant, v.2, n.36, p.102-107, 2000. MURASHIGE, T.; SKOOG, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiologia Plantarum, v.15, p.473-479, 1962. NAKASU, B. H.; FAORO, I. D. Cultivares. In: NAKASU, B. H.; CENTELLAS-QUEZADA, A.; HERTER, F. G. Pêra. Produção. Brasília: Embrapa Informação Tecnológica, 2003. p.29-36 (Frutas do Brasil, 46). NAKASU, B. H.; HERTER, F. G.; CAMELATTO, D.; JÚNIOR, C. R.; FORTES, J. F.; CASTRO, L. A. S. de; RASEIRA, A.; FREIRE, C. J. da S.; BASSO, C.; FAORO, I.; PETRI, J. L.; LEITE, G. B.; PEREIRA, J. F. M.; CANTILLANO, R. F. F.; VERÍSSIMO, V.; SIMÕES, F. A cultura da pêra. Brasília: Embrapa Informação Tecnológica, 2007. 58p. (Coleção Plantar, 58) NAKASU, B. H.; LEITE, D. L. Pirus 9: seleção de pereira para o Sul do Brasil. Hortisul, v.2, n.3, p.19-20, 1992. NORBERTO, P. M.; CHALFUN, N. N. J.; PASQUAL, M.; VEIGA, R. D.; PEREIRA, G. E.; MOTA, J. H. Efeito da época de estaquia e do AIB no enraizamento de estacas de figueira (Ficus carica L.). Ciência Agrotécnica, v. 25, n. 3, p. 533-541, 2001. OLIVEIRA, R. P.; SILVEIRA, D. G.; SILVA, S. O. Concentração de BAP e a eficiência de microcpropagação de bananeira tetraplóide (grupo AAAB). Scientia Agrícola, v.58, p.73-78, 2001.

62
PATI, P. K.; RATH, S. P.; SHARMA, M.; SOOD, A.; AHUJA; P. S. In vitro propagation of rose: a review. Biotechnology Advances, v.24, p.94-114, 2006. PIO ,R.; RAMOS, J. D.; PIO, L. A. S.; MENDONÇA, V.; SILVA, A. B.; PASQUAL, M. Enraizamento in vitro de brotações do portaenxerto de citros Tangerina sunki x Trifoliata english com o uso de sacarose e ácido indolbutírico. Ciência Agrotécnica, v.26, n.1, p.66-70, 2002. PÉREZ-TORNERO, O.; BURGOS, L. Different media requirements for micropropagation of apricot cultivars. Plant Cell, Tissue and Organ Culture, v.63, p.133-141, 2000. QUOIRIN, M.; LEPOIVRE, P. Etude de milieux adaptes aux cultures in vitro de Prunus. Acta Horticulturae, v.78, p.437-442, 1977. RADMANN, E. B.; BIANCHI, V. J.; SOUZA, T. M.; FACHINELLO, J. C.; OLIVEIRA, R. P. Influência da composição do meio de cultivo e do tipo de explante na micropropagação do portaenxerto de Prunus sp. „GxN-9‟. Scientia Agraria, v.10, n.2, p.095-101, 2009. RADMANN, E. B.; BIANCHI, V.J.; FACHINELLO, J.C.; FERREIRA, L.V.; OLIVEIRA, R.P. In vitro multiplication of „Flordaguard‟ rootstock: cytokinin source and concentration effects, explants orientation and period of permanence in the cultive medium. Brazilian Archives of biology and Technology, v.54, n.1, p.25-34, 2011. RAMAGE, C.M.; WILLIAMS, R.R. Cytokinin-induced abnormal shoot organogenesis is associated with elevated Knotted1-type homeobox gene expression in tobacco. Plant Cell Reports, v.22 p.919–924, 2004. RAMAGE, C. M.; WILLIAMS, R. R. Mineral nutrition and plant morphogenesis. In vitro Cellular and Developmental Biology-Plant, v.38, p.116-124, 2002. REED, B.M.; DENOMA, J.; WADA, S.; POSTMAN, J. Micropropagation of pear (Pyrus sp.). Methods in Molecular Biology, v.11013, p.3-18, 2013. ROGALSKI, M.; MORAES, L. K. A.; FELISBINO, R. C.; CRESTANI, L.; GUERRA, M. P.; SILVA, A. L. Enraizamento in vitro de portaenxertos de Prunus sp. Revista Brasileira de Fruticultura, v.25, n.2, p.293-296, 2003. SANTOS-SEREJO, J. A.; JUNGHANS, T. G.; SOARES, T. L.; SILVA, K. M. Meios nutritivos para micropropagação de plantas. In: SOUZA, A. S.; JUNGHANS, T. G. Introdução à micropropagação de plantas. Cruz das Almas: Embrapa Mandioca e Fruticultura Tropical, 2006. p.80-98. SATO, G.S.; ASSUMPÇÃO, R. Cultivo da pêra no Brasil e no Estado de São Paulo. Informações Econômicas, SP, v.33, n.7, jul.2003. SCHMIDT, W. Iron solutions: acquisition strategies and signaling pathways in plants. TRENDS in Plant Science, v.8, p.188-193, 2003.

63
SERAFINI, L. A.; BARROS, N. M.; AZEVEDO, J. L. Biotecnologia na agricultura e na agroindústria. Guaíba: Agropecuária, 2001. 463p. SILVA, L. C.; SCHUCH, M. W.; SOUZA, J. A.; ERIG, A. C.; ANTUNES, L. E. C. Meio nutritivo, reguladores de crescimento e frio no estabelecimento in vitro de mirtilo (Vaccinium ashei Reade) cv. „Delite‟. Revista Brasileira de Agrociência, v.12, n.04, p.405-408, 2006. TOMAZ, Z. F. P.; RODRIGUES, A. C.; VERÍSSIMO, V.; MARAFON, A. C.; HERTER, F. G.; RUFATO, A. DE R. Compatibilidade de enxertia de cultivares de marmeleiros com pereiras. Revista Brasileira de Fruticultura, v.31, n.4, p.1211-1217, 2009. XU, G.; FAN, X.; MILLER, A. J. Plant nitrogen assimilation and use efficiency. Annual Reviews Plant Biology, n.63, p.153–182, 2012. ZHANG, L.; XU, T. F.; SUN, X. F.; ZHANG, H. M.; TANG, T. L.; TANG, K. X. Factors influencing shoot regeneration from cotyledons of tetraploid Isatis indigotica F. In Vitro Cellular & Developmental Biology - Plant, v.39, n.05, p.459-462, 2003.

64
CAPÍTULO 3 – Radiosensibilidade em micropropagação de Pyrus sp., cv.
Ya-Li: efeitos morfofisiológicos
1 Introdução
Na fruticultura moderna buscam-se, cada vez mais, tecnologias que
possibilitem a produção de frutas de alta qualidade, com menor investimento e alto
retorno econômico. Isso se deve ao nível crescente das exigências do mercado,
cada vez mais competitivo (SOUZA, 2010). O Brasil é o segundo maior produtor
mundial de frutas, entretanto possui uma pequena participação no mercado
internacional. A exportação de frutas de clima temperado está aos poucos ganhando
espaço no mercado, e nos últimos anos, a quantidade de frutas exportadas, como a
maçã, ameixa, pêssego e morango aumentaram (REETZ et al., 2007). Entre as
frutas de clima temperado, a pera (Pyrus sp.) é a terceira mais consumida, porém, a
maior parte é importada (95%), pois seu cultivo ainda é pequeno (NAKASU et al.,
2007; FACHINELLO; FRANCESCATTO, 2009). A falta de cultivares adaptadas,
aptas a produzir com regularidade, em quantidade e qualidade, é um dos maiores
obstáculos à sua produção sustentável (NAKASU; FAORO, 2003; NAKASU, 2003).
Técnicas de indução de mutação têm se mostrado eficientes na produção de
variabilidade genética e, por isso, muito utilizadas em programas de melhoramento
de várias espécies de plantas, como a pereira, em vista que o melhoramento
genético convencional não supre estas carências, devido a pouca variabilidade
genética, ciclos longos e alogamia (JOSEPH et al., 2004; DE BONDT et al., 1996).
Com o uso de radiações ionizantes já foram obtidos vários mutantes com
características de interesse como maior produtividade, precocidade, menor porte,
maior resistência às doenças e pragas em diferentes espécies (KLEINHANS, 2009),
sendo que mais de 2500 variedades mutantes melhoradas já foram liberadas para o
cultivo comercial, demonstrando o valor econômico da mutação induzida para o

65
melhoramento das mesmas (PATADE; SUPRASANNA, 2008). Dessa forma, o
número de cultivares derivados de mutação induzida aumenta constantemente,
existindo catalogados no banco da FAO com cerca de 50 cultivares de fruteiras
pertencentes a mais de 20 diferentes espécies (FAO, 2009).
Segundo Predieri e Zimmermann (2001), a irradiação com raios gama é a via
mais utilizada para induzir mutação em plantas, pois possui dosimetria precisa, alta
e uniforme penetrabilidade nos tecidos e células, podendo ser realizada em várias
fases de desenvolvimento da planta. Neste contexto, esta técnica, acoplada à
cultura de tecidos, é uma alternativa promissora que permite a multiplicação e/ou
melhoramento de cultivares específicas e de interesse econômico, como a pereira
(MORAES et al., 2004).
O cultivo in vitro destaca-se também pela produção de plantas em larga
escala e com alta qualidade sanitária (BELL; REED, 2002; SILVA et al., 2006;
SOUZA et al., 2006; ZANANDREA et al., 2006). Entretanto, o sucesso desta prática
depende do crescimento e desenvolvimento adequado dos explantes durante as
fases de indução e multiplicação das brotações, aliados à capacidade de
enraizamento das brotações e das plantas obtidas resistirem à aclimatização
(HAZAKIRA, 2006).
Nas etapas de indução, multiplicação e enraizamento in vitro das brotações,
vários fatores podem influenciar, sendo os principais a composição do meio de
cultivo, como os tipos e concentrações de reguladores de crescimento utilizados
(principalmente citocininas e auxinas) e as condições ambientais às quais os tecidos
foram submetidos (SOTIROPOULOS et al., 2006; ZANANDREA et al., 2006). Além
desses fatores, o uso de radiações ionizantes também pode acarretar variações,
sejam elas epigenéticas ou genéticas, como observado por Silva (2009), Silva
(2013) e Hung e Johnson (2008).
Conforme Bandeira et al. (2012), o processo de aclimatização é considerado
um dos gargalos do cultivo in vitro, principalmente em espécies lenhosas, devido à
alta taxa de mortalidade das plantas, ocasionada, muitas vezes, por um sistema
radicular deficiente. Sendo assim, visando a formação de raízes com qualidade,
muitos autores apontam para a necessidade de inclusão de uma auxina na fase de
indução do enraizamento em espécies frutíferas, promovendo acréscimo na taxa de
sobrevivência das plantas após a transferência para as condições ex vitro
(RADMANN et al., 2002; ROGALSKI, et al., 2003; BARBOSA et al., 2008). Convém

66
ressaltar, ainda, que durante o processo de rizogênese in vitro, o uso de substratos
inertes embebidos no meio nutritivo, como a vermiculita, tem demonstrado
vantagens em relação ao ágar, pois além da propriedade de melhorar a aeração do
substrato e favorecer o desenvolvimento das raízes adventícias, são mais
econômicos (TIBOLA et al., 2004).
Diante do exposto, o presente trabalho tem como objetivo verificar os efeitos
de diferentes doses de radiação gama (60Co) durante a micropropagação de pereira,
cv. Ya-Li, almejando selecionar novos genótipos com características de interesse, a
partir dos genótipos originais, os quais serão utilizados em futuros programas de
melhoramento da espécie.
2 Material e Métodos
Os explantes utilizados para o desenvolvimento do trabalho foram
provenientes de brotações de pereira, cv. Ya-Li, pré-estabelecidas e multiplicadas in
vitro em meio de cultivo MS (MURASHIGE; SKOOG, 1962), suplementado com 0,8
mg L-1 de BAP (6-benzilaminopurina), 100 mg L-1 de mio-inositol, 30 g L-1 de
sacarose, pH 5,8 e 7,0 g L-1 de ágar.
Para a irradiação do material vegetal, segmentos nodais de aproximadamente
1,0 cm, contendo de duas a três gemas axilares, foram inoculados em placas de
Petri com 20 mL de meio MS, acrescido de 7,0 g L-1 de ágar, sem reguladores de
crescimento e expostos a diferentes doses de radiação gama (0, 10, 20, 30 e 40
Gy), sendo esta obtida de uma fonte de 60Co com taxa de rendimento de 0,8211
Gy/min para um campo de30x30 cm e uma distância foco-objeto irradiado de 80 cm,
pertencente ao Centro de Irradiação da Faculdade de Medicina da Universidade
Federal de Pelotas.
Durante as fases de indução e multiplicação das brotações irradiadas e não
irradiadas, os explantes foram cultivados em meio MS contendo 0,8 mg L-1 de BAP,
100 mg L-1 de mio-inositol, 30 g L-1 de sacarose, pH 5,8 e 7 g L-1 de ágar e mantidos
em câmara de crescimento com temperatura de 23±2ºC, fotoperíodo de 16 horas e
48 μmol m-2 s-1 de densidade de fluxo de fótons. Após 45 dias (período de indução
das brotações), avaliou-se a percentagem de explantes sobreviventes e
regenerantes (%), o número de brotações por explante, o comprimento das
brotações (cm) e o número de gemas axilares por brotação.

67
As brotações obtidas na fase de multiplicação, desenvolvidas a partir dos
explantes inicialmente irradiados e não irradiados, com aproximadamente 2,0 cm de
comprimento, foram utilizadas para o enraizamento in vitro. Suas bases foram
excisadas em bisel e inoculadas em meio MS [com ½ da concentração dos macro e
micronutrientes (com exceção do Fe e vitaminas)], acrescido de 30 g L-1 de
sacarose, 100 mg L-1 de mio-inositol e 0,8 mg L-1 de AIB (ácido indolil-butírico), com
pH ajustado para 5,8 e utilizando-se dois tipos de substrato [vermiculita de grânulos
médios (3 g frasco-1) ou ágar (7 g L-1)]. Os frascos foram mantidos em câmara de
crescimento sob as mesmas condições de cultivo anteriormente descritas. Após 30
dias, foram analisados percentagem de enraizamento (%), número de raízes por
planta, comprimento da maior raiz (cm) e percentagem de calos formados (%).
Todas as brotações enraizadas foram transferidas para bandejas plásticas
com tampa, contendo vermiculita, permanecendo em estufa de vidro com
temperatura controlada (18±2ºC) por 30 dias. Após uma semana da transferência
para estufa, as tampas das bandejas começaram a ser abertas progressivamente.
Decorrido o período de aclimatização, foi avaliada a percentagem de plantas
sobreviventes (%), as quais foram transplantadas para bandejas alveoladas de
polipropileno contendo substrato Carolina®, onde foram mantidas por mais 60 dias e,
em seguida, transferidas para potes plásticos com capacidade de 3 L, onde
permaneceram por 60 dias. Posteriormente, foram levadas para o campo, para
posterior avaliação dos melhoristas da cultura (pesquisadores da Embrapa).
O experimento de indução das brotações in vitro foi conduzido com
delineamento experimental inteiramente casualizado, contendo apenas um fator de
tratamento composto por cinco doses de radiação, com nove repetições por
tratamento, representadas, cada uma, por um frasco contendo cinco explantes.
Para o experimento de enraizamento in vitro foi utilizado o delineamento
experimental inteiramente casualizado, seguindo um arranjo bifatorial 5x2 (cinco
doses de radiação e dois tipos de substrato), com cinco repetições por tratamento,
sendo cada repetição representada por um frasco contendo cinco explantes.
Os resultados obtidos foram submetidos à análise de variância e as médias
comparadas pelo teste de Tukey, ao nível de 5% de probabilidade de erro, ou
analisadas por regressão polinomial, utilizando o software WINSTAT 1.0
(MACHADO; CONCEIÇÃO, 2007). Porém, em relação à percentagem de plantas
sobreviventes após a aclimatização foi realizada apenas uma análise descritiva.

68
3 Resultados
Observou-se diferença significativa entre os tratamentos para todas as
variáveis analisadas durante a fase de indução de brotações in vitro. Com base nos
resultados obtidos, verificou-se que o incremento das doses de irradiação afetou
negativamente todas as variáveis analisadas, como pode ser observado na Fig. 1. A
dose de 40 Gy determinou uma redução de 53,33% nos parâmetros percentagem de
explantes sobreviventes e regenerantes, sendo que para as variáveis número de
brotações por explante, número de gemas axilares por brotação e comprimento das
brotações (cm) este decréscimo foi de 47,1; 51,37 e 80,55%, respectivamente, em
relação ao controle.
Porém, comparando-se a dose de 10 Gy com o tratamento não irradiado,
ambos os tratamentos apresentaram uma média de 80% de explantes regenerantes,
enquanto que nas demais variáveis a resposta foi semelhante, com redução de
apenas 6,67; 2,32; 9,65 e 16,67% para percentagem de explantes sobreviventes,
número de brotações por explante, número de gemas axilares por brotação e
comprimento das brotações (cm), respectivamente.
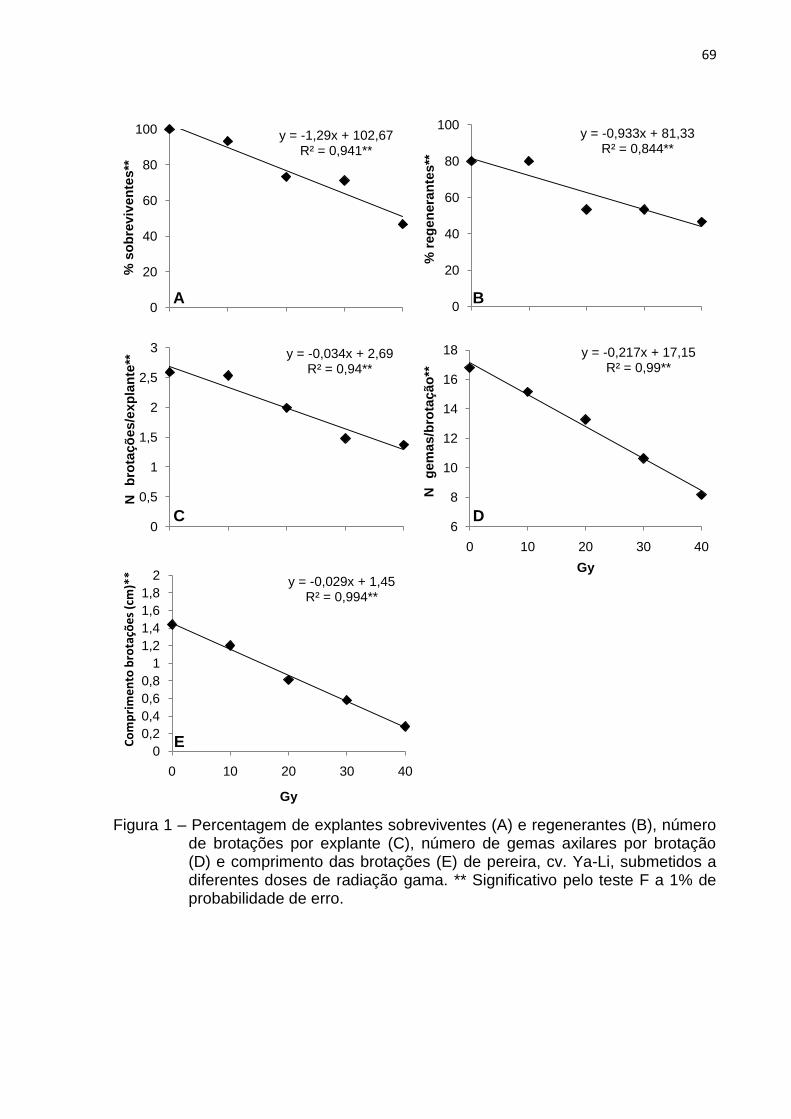
69
y = -0,933x + 81,33R² = 0,844**
0
20
40
60
80
100
% r
eg
en
era
nte
s**
y = -0,217x + 17,15R² = 0,99**
6
8
10
12
14
16
18
0 10 20 30 40
Ng
em
as/b
rota
ção
**
Gyy = -0,029x + 1,45
R² = 0,994**
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
2
0 10 20 30 40
Co
mp
rim
en
to b
rota
çõe
s (c
m)*
*
Gy
Figura 1 – Percentagem de explantes sobreviventes (A) e regenerantes (B), número
de brotações por explante (C), número de gemas axilares por brotação (D) e comprimento das brotações (E) de pereira, cv. Ya-Li, submetidos a diferentes doses de radiação gama. ** Significativo pelo teste F a 1% de probabilidade de erro.
y = -0,034x + 2,69R² = 0,94**
0
0,5
1
1,5
2
2,5
3
Nb
rota
çõ
es/e
xp
lan
te**
y = -1,29x + 102,67R² = 0,941**
0
20
40
60
80
100%
so
bre
viv
en
tes**
A B
C D
E

70
Na Fig. 2 são apresentados os resultados obtidos nos experimentos de
enraizamento in vitro e aclimatização. Para as variáveis percentagem de
enraizamento e comprimento da maior raiz não ocorreu interação entre os fatores.
Em relação ao parâmetro percentagem de enraizamento, verificou-se efeito somente
para o fator tipo de substrato utilizado (Fig. 2A), onde o meio contendo vermiculita
induziu o enraizamento de 98,4% dos explantes.
No que se refere ao comprimento da maior raiz (Fig. 2C e 2D), houve efeito
isolado para os fatores em estudo, obtendo-se, quanto ao tipo de substrato utilizado,
melhor resultado em meio geleificado com ágar, onde as raízes apresentaram cerca
de 1,86 cm. Entretanto, para o fator doses de irradiação, apenas as raízes expostas
à dose de 10 Gy mostraram menor comprimento (0,7 cm), significando uma redução
de 50% quando comparado ao controle.
Com relação às variáveis número de raízes por planta e percentagem de
calos formados, obteve-se interação entre os fatores, sendo que o maior número de
raízes foi obtido em brotações irradiadas com 10 Gy, cultivados tanto em meio com
vermiculita (13,64) como em meio com ágar (14,04) (Fig. 2B).
A presença de calos foi verificada em todos os tratamentos em que utilizou-se
ágar, sendo que o material irradiado diferiu significativamente em relação ao não
irradiado, apresentando maior percentagem de calos formados e que este tipo de
substrato é mais responsivo ao aparecimento de calos quando comparado à
vermiculita. Em contrapartida, em meio com vermiculita as doses mais elevadas (20,
30 e 40 Gy) diferiram significativamente da dose de 10 Gy e do controle, o qual não
apresentou formação dos mesmos (Fig. 2E).
Após o período de aclimatização, comparando-se os diferentes tratamentos,
pode-se observar que as brotações expostas às doses mais altas de irradiação (30 e
40 Gy), foram as que apresentaram maior percentagem de sobrevivência
(aproximadamente 90%), determinando um aumento de 20% em relação ao controle
(Fig. 2F).
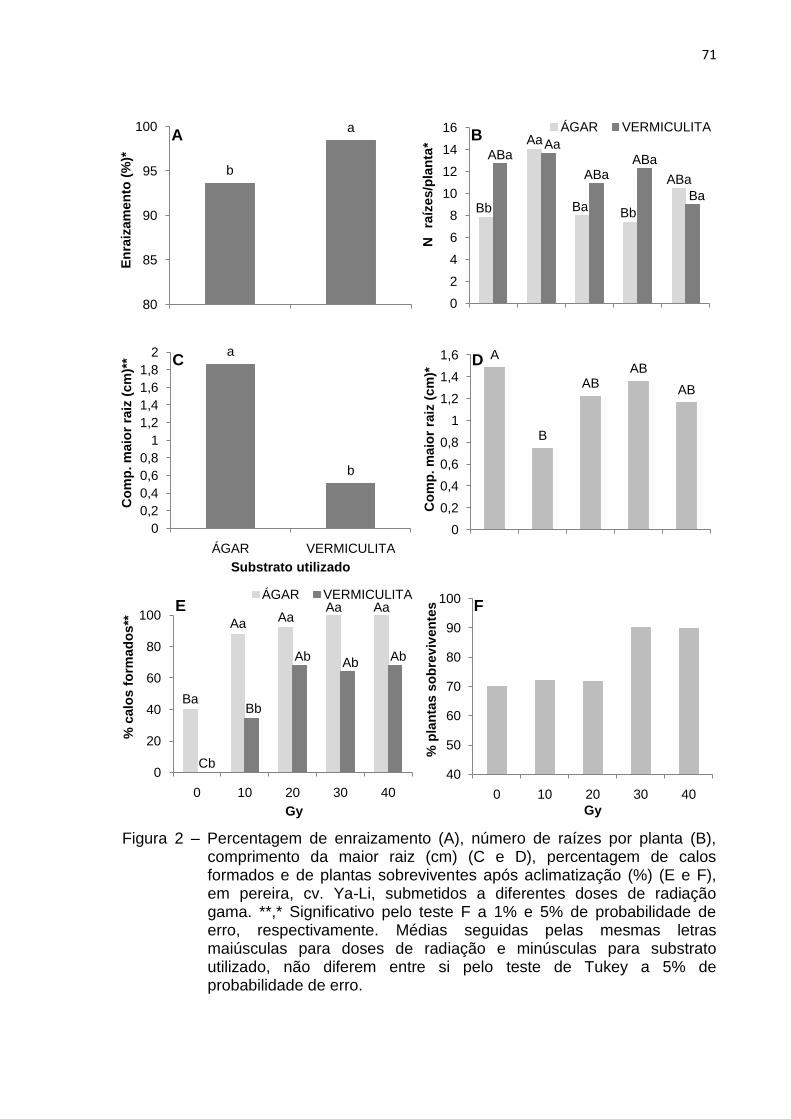
71
Bb
Aa
Ba Bb
ABa
ABaAa
ABaABa
Ba
0
2
4
6
8
10
12
14
16
Nra
ízes/p
lan
ta*
ÁGAR VERMICULITA
a
b
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
2
ÁGAR VERMICULITA
Co
mp
. m
aio
r ra
iz (
cm
)**
Substrato utilizado
A
B
ABAB
AB
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
Co
mp
. m
aio
r ra
iz (
cm
)*
Ba
AaAa
Aa Aa
Cb
Bb
AbAb
Ab
0
20
40
60
80
100
0 10 20 30 40
% c
alo
s f
orm
ad
os**
Gy
ÁGAR VERMICULITA
40
50
60
70
80
90
100
0 10 20 30 40
% p
lan
tas s
ob
rev
iven
tes
Gy
Figura 2 – Percentagem de enraizamento (A), número de raízes por planta (B),
comprimento da maior raiz (cm) (C e D), percentagem de calos formados e de plantas sobreviventes após aclimatização (%) (E e F), em pereira, cv. Ya-Li, submetidos a diferentes doses de radiação gama. **,* Significativo pelo teste F a 1% e 5% de probabilidade de erro, respectivamente. Médias seguidas pelas mesmas letras maiúsculas para doses de radiação e minúsculas para substrato utilizado, não diferem entre si pelo teste de Tukey a 5% de probabilidade de erro.
b
a
80
85
90
95
100E
nra
izam
en
to (
%)*
A B
C D
E F

72
4 Discussão
A utilização de irradiação gama em tecidos vegetais tem como alvo principal
as modificações das moléculas de DNA, sendo que a energia transferida das
radiações resulta em ionização no interior da célula, acentuando as chances de
obtenção de mutações de interesse (RODRIGUES; ANDO, 2003; FU et al., 2008).
Porém, segundo Patade e Suprasanna (2008), a dose ideal de radiação gama
que promova mutações sem a ocorrência de perdas no rendimento do organismo
irradiado é tecido-específica, variando a radiosensibilidade com a espécie, a cultivar,
as condições fisiológicas da planta e com a manipulação do material irradiado antes
e depois do tratamento. Portanto, diferentes tecidos respondem de forma própria,
sendo necessária a realização de testes de radiosensibilidade para determinar a
dose ideal (GAZZANEO et al., 2007). Neste contexto, este trabalho foi desenvolvido
baseando-se em estudo prévio realizado por Silva (2009), a qual, trabalhando com a
mesma cultivar, obteve polimorfismos entre os clones de pereira irradiados
(avaliados a partir de marcadores RAPD).
Conforme preconizado por Lu et al. (2007), o nível crítico de radiação gama
nos quais mutações são induzidas devem estar dentro da tolerância para a
regeneração de brotações (Fig. 3A). Doses elevadas podem gerar alta frequência de
mutações, mas baixa taxa de regeneração de culturas in vitro (AMJAD; AMJUN,
2007), o que pode ser evidenciado neste trabalho, durante a fase de indução das
brotações, pois o aumento das doses de radiação ocasionou um decréscimo
significativo nas variáveis relacionadas.

73
Figura 3 – Plantas irradiadas de Pyrus sp., cv. Ya-Li, após avaliações de
regeneração/multiplicação in vitro [com aspecto normal (A) e presença de caules espessos (B)], após enraizamento in vitro [com uso de ágar como substrato (C) e com vermiculita (D)] e no período de aclimatização [em bandejas com tampas (E), em bandejas de polipropileno (F), em potes plásticos de 3 L após transplantadas(G) e em potes plásticos de 3 L antes de ir para o campo].

74
O parâmetro mais afetado pelas doses mais altas de irradiação foi o
comprimento das brotações, dado este que corrobara com os encontrados por
diversos autores, tais como Coimbra et al. (2005), Hung e Johnson (2008),
Berenschot et al. (2008), Silva (2009) e Kleinhans (2009), confirmando que altas
doses de radiação gama podem produzir efeitos deletérios, tais como a diminuição
no crescimento (WATANABE et al., 2000), sendo que, neste trabalho também foram
constatadas alterações fenotípicas nos caules (mais espessos) das brotações
obtidas de explantes irradiados com doses mais altas (Fig. 3B). Diversos fatores
podem estar relacionados a esta redução no comprimento das brotações, ligados a
variações epigenéticas ou genéticas, dentre os quais podem ser citados alterações
de poucas bases até inserções e deleções de grandes fragmentos de DNA, como
também inibição de sua síntese, mudanças fenotípicas ou outros danos fisiológicos
ocorridos após os tratamentos mutagênicos (KLEINHANS, 2009; TABASUM et al.,
2011), uma vez que a radiação interfere nos processos de divisão celular,
culminando em crescimento alterado e variabilidade morfológica (AMJAD; AMJUN,
2007).
Por outro lado, em relação às doses de radiação testadas, observou-se que a
dose inferior (10 Gy) praticamente não apresentou diferenças em relação ao controle
nos parâmetros em estudo durante a fase de indução das brotações, sendo
verificados efeitos positivos também durante o enraizamento in vitro, no que se
refere ao número de raízes por planta. Com base nos dados obtidos nestas
avaliações e em estudos anteriores, como os realizados por Silva (2009), em
pereira, Scheer da Silva et al. (2012), em arroz e Silva (2013) em amoreira-preta, é
possível afirmar que a radiação gama possui efeito estimulatório quando ministrada
em baixas doses, em diversos parâmetros morfológicos e fisiológicos de crescimento
vegetal. Isto pode estar relacionado ao fato de que, em condições apropriadas de
sensibilidade e sob o efeito de determinadas doses de radiação ionizante, pode-se
romper o delicado equilíbrio hormonal e metabólico que regula o processo do
desenvolvimento vegetal levando muitas vezes a respostas do tipo estimulatórias
(BERENSCHOT et al., 2008). No entanto, os mecanismos pelos quais as plantas
reconhecem e respondem às doses a que foram expostas de radiação ainda não
são compreendidos (PRISTOV, 2013).
Estudos realizados têm demonstrado que o efeito da radiação depende do
estádio de desenvolvimento das plantas e das variáveis analisadas, ocorrendo

75
inclusive recuperação de alguns parâmetros que inicialmente apresentavam
alteração (HUNG; JOHNSON, 2008). Isto foi verificado neste trabalho, pois durante a
multiplicação das brotações, a partir do terceiro subcultivo houve um aumento no
número de brotações e de gemas axilares, nas brotações oriundas de explantes
irradiados com as doses mais elevadas (20, 30 e 40 Gy) (dados não mostrados).
Esse fato pode estar relacionado à ação das peroxidases, das enzimas reparadoras
e das catalases, as quais desempenham um papel fundamental na recuperação das
células e na decomposição de produtos da radiação (RODRIGUES; ANDO; 2003),
em vista que o alvo da radiação são as moléculas de água do organismo e estas,
quando irradiadas, sofrem radiólise, fenômeno ao qual se segue um rearranjo
eletrônico e a possibilidade de produção de radicais livres (SELVI et al., 2008).
No que se refere ao processo de enraizamento in vitro, embora o maior
número de raízes formadas tenha ocorrido na dose de 10 Gy, em ambos substratos
utilizados, verificou-se a presença das mesmas nos demais tratamentos, o que é de
grande valia, pois quando se aumenta a quantidade de raízes formadas in vitro,
aumenta-se também a área de contato raiz/substrato, refletindo em maior absorção
dos nutrientes (VILLA et al., 2008). Estes resultados, aliados à alta percentagem de
enraizamento obtida comprovam os relatos feitos por muitos autores, ou seja, da
importância da utilização de auxina no meio de cultura para induzir a formação e a
iniciação de raízes adventícias em determinadas espécies. Entretanto, é necessário
o uso de uma concentração adequada de auxina para obtenção de um sistema
radicular funcional e uniforme (SANTOS-SEREJO et al., 2006; COSTA et al., 2008;
SCHMILDT et al., 2010).
Verificou-se que a maior percentagem de enraizamento foi obtida quando
utilizada vermiculita como substrato. Segundo Vieira et al. (2007), o uso desses
substratos aerados possibilita ganhos substanciais na qualidade das raízes
formadas, por promoverem a formação de raízes ramificadas com presença
abundante de pelos absorventes, maior aeração do meio e menor transmissão de
luz, favorecendo, assim, o enraizamento e desenvolvimento das raízes adventícias
in vitro.
As raízes desenvolvidas em vermiculita emitiram pelos absorventes, porém o
comprimento médio da maior raiz foi menor, quando comparadas às cultivadas em
ágar (Fig. 3C e 3D). Hoffmann et al. (2001), em trabalhos realizados com macieira,
também constataram a formação de raízes mais curtas e com maior densidade de

76
pelos radiculares nos meios com vermiculita. De acordo com Tibola et al. (2004),
raízes mais curtas ou na forma de primórdios radiculares aceleram o pegamento da
planta e evitam o enovelamento. Além disso, estas são mais desejáveis, pois
facilitam a lavagem e retirada do meio de cultura aderido, bem como a introdução da
planta no substrato.
Nos meios semi-sólidos, o ágar, um polissacarídeo extraído de algas
marinhas, é o agente geleificante tradicionalmente usado e, em contrapartida, o
ingrediente mais caro do meio. Apesar de ser o mais utilizado, as raízes formadas
normalmente não possuem pelos radiculares devidamente desenvolvidos, pela falta
de oxigênio (LEITE, 1995), além de suas raízes serem mais fibrosas e quebradiças.
Observou-se, em todos os tratamentos, com exceção do material não
irradiado em meio contendo vermiculita, a ocorrência de calos na base dos
explantes. Segundo Carvalho et al. (2011) e Guerra e Nodari (2006), calos são
grupo ou massa de células em crescimento desordenado, as quais podem
apresentar certo grau de diferenciação e, em geral, têm forma e tamanho variado,
paredes celulares com diferentes graus de espessamento e se originam da
proliferação desordenada a partir de tecidos ou órgãos cultivados in vitro. Em razão
disso, sua presença pode afetar negativamente a rizogênese (GEORGE et al.,
2008), porém isto não foi evidenciado.
O que pode explicar esta produção de calos é a exposição do material vegetal
às doses de radiação, pois a radiação gama interfere nos processos de divisão
celular, ocasionando um crescimento alterado e variabilidade morfológica (AMJAD;
AMJUN, 2007), como já enfatizado.
Dessa maneira, isso pode ter ocorrido devido ao um desbalanço hormonal,
pois, segundo George e Sherrington (1984), a morfogênese in vitro é regulada pela
interação e balanço entre os reguladores de crescimento acrescidos ao meio de
cultura, principalmente citocininas e auxinas. As citocininas caracterizam-se por
promoverem a quebra da dominância apical, aumentando a taxa de multiplicação
por meio de brotações laterais relacionadas à atividade promovida nos meristemas
(BRUM et al., 2002; TOMBOLATO; COSTA, 1998). Porém, em concentrações
elevadas, podem incrementar a ação da citocinina-oxidase, impedir a atuação da
citocinina sobre a divisão celular e, consequentemente, a indução de brotações
(BARRUETO CID, 2000; SOUZA et al., 2006), sendo que em vários sistemas de
cultivo in vitro têm sido descritas anormalidades anatômicas e ultraestruturais,

77
alterações metabólicas, nanismo, teratologia, albinismo e hiperidricidade,
decorrentes da exposição excessiva à citocininas (RAMAGE; WILLIAMS, 2004). As
auxinas promovem divisão, alongamento e diferenciação celular, além de serem
responsáveis pela dominância apical (CARVALHO et al., 2011). Altas concentrações
de auxinas induzem maior divisão celular e, por conseguinte, maior formação de
calo (RADMANN et al., 2002; SILVA, 2004), além de anular parte do efeito inibitório
que a citocinina promove sobre o crescimento das novas brotações (HWANG et al.,
2012).
Outro fator importante a ser considerado é que um dos possíveis efeitos
indiretos da radiação sobre a indução de variabilidade genética e epigenética está
relacionado à indução na formação de EROS, devido aos danos celulares, e estas
EROS podem causar danos no DNA ou alterar o padrão de expressão de muitos
genes e, consequentemente, em caracteres fenotípicos. Ou seja, há um efeito direto
e um indireto que pode levar à variabilidade pela mutação ou por variação
epigenética.
A aclimatização das plantas foi bem sucedida (Fig. 3E, 3F, 3G e 3H). Este
fato provavelmente está relacionado com a boa qualidade da parte aérea e do
sistema radicular, formado durante o enraizamento in vitro. Segundo Rogalski et al.
(2003) a adição de auxina no meio de enraizamento tem sido eficiente, em espécies
frutíferas lenhosas, promovendo acréscimo na sobrevivência de plantas na fase de
aclimatização.
Além disso, houve também influência das doses de radiação em relação à
percentagem de sobrevivência das plantas, em que maiores taxas de aclimatização
ocorreram nas doses mais elevadas (30 e 40 Gy), assim como verificado por Hung e
Johnson (2008), com Wasabia japonica, em que as plantas também apresentaram
incremento de 20% na sobrevivência, no tratamento de 40 Gy. Neste sentido,
conforme Amjad e Amjun (2007), doses elevadas de radiação podem gerar alta
frequência de mutações. E, em muitos casos, uma mutação pontual pode corrigir ou
melhorar alguns caracteres, permitindo a seleção de constituições genéticas
superiores (MARTINS et al., 2005), como já verificado em diversos trabalhos com
fruteiras, nos quais a mutagênese foi usada para introduzir muitas características
úteis que afetam o tamanho das plantas, tempo de florescimento, maturação de
frutos, coloração de frutos, auto-incompatibilidade, auto-raleio, resistência aos
agentes patogênicos, mudança na qualidade dos frutos, aumento no conteúdo de

78
ácido ascórbico, redução na acidez, maior consistência dos frutos e o aumento da
vida de prateleira (VISSER et al., 1971; LACEY, 1975; DECOURTYE, 1982;
BROERTJES; VAN HARTEN, 1988; MASUDA et al., 1997; VAN HARTEN, 1998;
BASARAN; KEPENEK, 2011).
5 Conclusões
Nas condições em que foi desenvolvido o trabalho pode-se concluir que:
– Altas doses de irradiação interferem em alguns parâmetros de crescimento na fase
de indução das brotações;
– Baixas doses de irradiação não afetam as variáveis estudadas durante a fase de
indução das brotações, bem como tendem a estimular um melhor enraizamento das
mesmas, juntamente com o uso de vermiculita;
– A vermiculita substitui com vantagens o ágar como substrato, no enraizamento in
vitro da cv. Ya-Li;
– As doses de irradiação utilizadas não afetam negativamente o processo de
aclimatização.
4 Referências bibliográficas
AMJAD, M.; ANJUM, M. A. Effect of post-irradiation ageing on onion seeds. Acta Physiologia Plantarum, v.29, p.63–69, 2007. BANDEIRA, J. DE M.; THUROW, L. B.; BRAGA, E. J. B.; PETERS, J. A.; BIANCHI, V. J. Rooting and acclimatization of the Japanese plum tree, cv. América. Revista Brasileira de Fruticultura, v.34, n.2, p. 597-603, 2012. BARBOSA, W.; PIO, R.; VEIGA, R. F. A.;CHAGAS, E. A.; FELDBERG, N. P.; CAMPAGNOLO, M. A.; DALASTRA, I. M. Efeito de concentrações do AIB no enraizamento in vitro de cultivares de figueira. Biosci. J., v.24, n.2, p.1-6, 2008. BARRUETO CID, L. P. Citocininas. In: BARRUETO CID, L. P. (Ed.). Introdução aos hormônios vegetais. Brasília, DF: Embrapa Recursos Genéticos e Biotecnologia, 2000. p.55-88. BASARAN, P.; KEPENEK, K. Fruit quality attributes of blackberry (Rubus sanctus) mutants obtained by 60Co gamma irradiation. Biotechnology and Bioprocess Engineering, v.16, p. 587-592, 2011. BELL, R. L.; REED, B. M. In vitro tissue culture of pear: Advances in techinques for micropropagation and germplasm preservation. Acta Horticulturae, n.596, p.412- 419, 2002.

79
BERENSCHOT, A. S.; TULMANN NETO, A. ; ZUCCHI, M. I. ; QUECINI, V. Mutagenesis in Petunia x hybrida Vilm. and isolation of a novel morphological mutant. Brazilian Journal of Plant Physiology, v.20, p.95-103, 2008. BROERTJES, C.; VAN HARTEN, A. M. Applied Mutation Breeding for Vegetatively Propagated Crops. Amsterdam: Elsevier Publishers, 1988. BRUM, G. R.; SILVA, A. B.; PASQUAL, M. Efeito de diferentes concentrações de BAP e ANA na propagação in vitro da figueira (Ficus carica L.). Ciência e Agrotecnologia, p.1403-1409, 2002. CARVALHO, A. C. P. P. de; TORRES, A. C.; BRAGA, E. J. B.; LEMOS, E. P. de; SOUZA, F. V. D.; PETERS, J. A.; WILLADINO, L.; CÂMARA, T. R. Glossário de Cultura de Tecidos de Plantas. Plant Cell Culture & Micropropagation, v.07, p. 30-60, 2011. COIMBRA, J. L. M.; CARVALHO, F. I. F. de; COIMBRA, S. M.; LORENCETTI, C.; CHOCOROSQUI, V. R. Doses de raios gama na cultura da aveia: estatura de planta gama-rays doses in aveia culture: stature of plant. Revista Brasileira de Agrociência, v.11, n.3, p.303-308, 2005. COSTA, F. H. S.; PASQUAL, M.; PEREIRA, J. E. S.; RODRIGUES, F. A.; MIYATA, L. Y. Relação entre o tempo de enraizamento in vitro e o crescimento de plantas de bananeira na aclimatização. Revista Brasileira de Fruticultura, v.30, n.1, p.031-037, 2008. DE BONDT, A.; EGGERMONT, K.; PENNINCKX, I.; GODERIS, I.; BROEKAERT, W. F. Agrobacterium-mediated transformation of apple (Malus domestica Borkh): and assessment of factors affecting regeneration of transgenic plants. Plant Cell Reports, v.15, p.549-554, 1996. DECOURTYE, L. Bilancio di 20 anni di miglioramento delle specie legnose da frutto mediante la mutagenesi e prospettive attuali. Atti della giornata di studio sull’uso di tecniche nucleari per il miglioramento genetico dei fruttiferi, p.21–40, 1982. FACHINELLO, J. C.; FRANCESCATTO, P. Compatibilidade de enxerto e porta-enxerto de pereira. In: XI ENFRUTE (Encontro Nacional sobre Fruticultura de Clima Temperado), 1, 2009, Fraiburgo. Anais... Caçador: EPAGRI, 2009. p.33-42. FAO. Faostat Database. Prodstat. Disponível em: <http://www.fao.org>. Online. Acesso em 10 jul. 2009. FU, H. W.; LI, Y. F; SHU, Q. Y. A revisit of mutation induction by gamma rays in rice (Oryza sativa L.): implications of microsatellite markers for quality control. Molecullar Breeding, v.22, p.281-288, 2008. GAZZANEO, L. R. S.; COLAÇO, W.; KIDO, E. A.; ISEPPON, A. M. B.; KIDO, L. M. H. Efeito da Radiação Gama Sobre o Desenvolvimento in vitro de Vigna unguiculata. Revista Brasileira de Biociências, v.5, n.2, p.27-29, 2007.

80
GEORGE, E. F.; HALL, M. A.; DE KLERK, G. J. Plant propagation by tissue culture. 3nd Edition, Dordrecht: Springer, v.1, 2008. GEORGE, E. F.; SHERINGTON, P. D. Factors affecting growth and morphogenesis. Plant propagation by issue culture, England: Exegeics, p.125-171, 1984. GUERRA, M.P.; NODARI, R.O. Apostila de biotecnologia – CCA/UFSC. Edição da Steinmacher, 2006. p1-48. HAZAKIRA, B.N. Morpho-physiological disorders in in vitro culture of plants. Scientia Horticulturae, v.108, p.105-120, 2006. HOFFMANN, A.; PASQUAL, M.; CHALFUN, N. N. J. Substratos na indução e desenvolvimento in vitro de raízes em dois portaenxertos de macieira. Pesquisa Agropecuária Brasileira, v.36, n.11, p.1371-1379, 2001. HWANG, I.; SHEEN, J.; MULLER, B. Cytokinin signaling networks. Reviews Plant Biology, v.63, p.353–80, 2012. HUNG, D. C.; JOHNSON, K. Effects of ionizing radiation on the growth and allyl isothiocyanate accumulation of Wasabia japonica in vitro and ex vitro. In Vitro Cellular Developmental Biology-Plant, v.44, p.51-58, 2008. JOSEPH, R.; YEOH, H.; LOH, C. Induced mutations in cassava using somatic embryos and the identification of mutant plants with altered starch yield and composition. Plant Cell Report, v.23, p.91–98, 2004. KLEINHANS, P. S. J. Radiação gama cobalto-60 na indução de variabilidade em mamona (Ricinus communis L.). 2009. 88f. Dissertação (Mestrado em Ciências) – Universidade Federal de Pelotas – UFPel, Pelotas. LACEY, C.N.D. Induction and selection of mutant form of fruit plants. Annual Report, v.1, p.22–24, 1975. LEITE, G. B. Efeito de reguladores de crescimento, substratos, sacarose e intensidade luminosa na micropropagação de pereira (Pyrus communis, L.) cv. Bartlett e do clone OH x F97. 1995, 50 f. Dissertação (Mestrado em Agronomia – Fruticultura de Clima Temperado) – Faculdade de Agronomia Eliseu Maciel, Universidade Federal de Pelotas – UFPel, Pelotas. LU, G.; ZHANG, X.; ZOU, Y.; ZOU, Q.; XIANG, X; CAO, J. Effect of radiation on regeneration of Chinese narcissus and analysis of genetic variation with AFLP and RAPD markers. Plant Cell, Tissue and Organ Culture, v.88, p.319–327, 2007. MACHADO, A. A.; CONCEIÇÃO, A. R. Sistema de análise estatística para Windows. Winstat. Versão 1.0. UFPel, 2003. MARTINS, A. F.; ZIMMER, P. D.; OLIVEIRA, A. da C.; CARVALHO, F. I. F. de; VIEIRA, E. A.; CARVALHO, M. F. de; MARTINS, L. F.; Fonseca, F. S. da

81
Variabilidade para caracteres morfológicos em mutantes de arroz. Ciência e Agrotecnologia, v.29, p.1215-1223, 2005. MASUDA, T.; OMIYA, I; YOSHIOKA, T.; INOUE, K.; MURATA, K.; KITAGAWA, K.; TABIRA, H.; YOSHIDA, A.; KOTOBUKI, K.; SANADA, T. Selection of mutants resistant to black spot disease by chronic irradiation of gamma rays in Japanese pear „Osanijisseiki‟. Journal of the Japanese Society Horticultural Science, v.66, p.85–92, 1997. MORAES, L.K.A. DE; FELISBINO, C.; CRESTANI, L.; SILVA, A.L. DA. Estabelecimento e multiplicação in vitro de Pyrus calleryana D-6 em sistema de cultura „dupla-fase‟ . Revista Brasileira de Fruticultura, v.26, n.3, p.403-405, 2004. MURASHIGE, T.; SKOOG, F. A revised medium for rapid growth and biossay with tobacco tissue cultures. Physiologia Plantarum, v.15, p.473-497, 1962. NAKASU, B. H. Introdução. In: NAKASU, B. H.; CENTELLAS-QUEZADA, A.; HERTER, F. G. Pêra. Produção. Brasília: Embrapa Informação Tecnológica, 2003. p.9 (Frutas do Brasil, 46). NAKASU, B. H.; FAORO, I. D. Cultivares. In: NAKASU, B. H.; CENTELLAS-QUEZADA, A.; HERTER, F. G. Pêra. Produção. Brasília: Embrapa Informação Tecnológica, 2003. p.29-36 (Frutas do Brasil, 46). NAKASU, B. H.; HERTER, F. G.; CAMELATTO, D.; JÚNIOR, C. R.; FORTES, J. F.; CASTRO, L. A. S. de; RASEIRA, A.; FREIRE, C. J. da S.; BASSO, C.; FAORO, I.; PETRI, J. L.; LEITE, G. B.; PEREIRA, J. F. M.; CANTILLANO, R. F. F.; VERÍSSIMO, V.; SIMÕES, F. A cultura da pêra. Brasília: Embrapa Informação Tecnológica, 2007. 58p. (Coleção Plantar, 58) PATADE, V. Y.; SUPRASANNA, P. Radiation induced in vitro mutagenesis for sugarcane improvement. Sugar Technology, v.10, p.14-19, 2008. PREDIERI, S.; ZIMMERMANN, R. H. Pear mutagenesis: In vitro treatment with gamma-rays and field selection for productivity and fruit traits. Euphytica, v.117, n.3, p.217-227, 2001. PRISTOV. J. B.; SPASIC, M.; SPASOJEVIC, I. Converting low dose radiation to redox signaling Plant signaling e behavior, v.8, 2013. REETZ, E. R., RIGON, L., VENCATO, A. Anuário brasileiro de fruticultura 2007. Santa Cruz do sul: ERNA, 2007. RADMANN, E. B.; FACHINELLO, J. C; PETERS, J. A. Efeito de auxinas e condições de cultivo no enraizamento in vitro de portaenxertos de macieira „M-9‟. Revista Brasileira de Fruticultura, v. 24, n. 3, p. 624-628, 2002. RAMAGE, C. M.; WILLIAMS, R. R. Cytokinin-induced abnormal shoot organogenesis is associated with elevated Knotted1-type homeobox gene expression in tobacco. Plant Cell Reports, v.22, p.919–924, 2004.

82
RODRIGUES, L. R. F.; ANDO, A. Uso da sensitividade à radiação gama na discriminação de variedades de arroz-de-sequeiro dos grupos índica e japônica. Bragantia, v.62, p.179-188, 2003. ROGALSKI, M.; MORAES, L. K. A.; FELISBINO, R. C.; CRESTANI, L.; GUERRA, M. P.; SILVA, A. L. Enraizamento in vitro de portaenxertos de Prunus sp. Revista Brasileira de Fruticultura, v.25, n.2, p.293-296, 2003. SANTOS-SEREJO, J. A.; JUNGHANS, T. G.; SOARES, T. L. SILVA, K. M. Meios nutritivos para micropropagação de plantas. In: SOUZA, A. S.; JUNGHANS, T. G. Introdução à micropropagação de plantas. Cruz das Almas: Embrapa Mandioca e Fruticultura Tropical, 2006. p.80-98. SCHEER DA SILVA, A.; DANIELOWSKI, R.; BRAGA, E. J. B.; DEUNER, S.; MAGALHÃES JUNIOR, A. M. de; PETERS, J. A. Radiossensibilidade em plântulas de arroz irrigado: potencial de crescimento e capacidade antioxidante. Revista de la Facultad de Agronomía, v.111, n.2, p.83-90, 2012. SCHMILDT, E. R.; AMARAL, J. A. T.; SCHMILDT, O.; COELHO, I. R.; RABELLO, W. S.; FILHO, S. M. Níveis de ácido indolbutírico (AIB) no enraizamento in vitro de microestacas de mamoeiro „Tainung 01‟. Acta Scientiarum Agronomy, v.32, n.1, p.125-129, 2010. SELVI, B. S.; PONNUSWAMI , V. ; SUMATHI, T. Identification of DNA Polymorphism induced by Gamma Ray Irradiation in Amla (Emblica officinalis Gaertn.) Grafts of V1 M1 and V2 M1 Generation. Journal of Applied Sciences Research, v.3, n.12, p.1933-1935, 2008. SILVA, C. P. Variablidade genética induzida por radiação gama em pereira cultivada in vitro. 2009. 74f. Dissertação (Mestrado em Fisiologia Vegetal) – Universidade Federal de Pelotas, Pelotas. SILVA, E. S. B. Propagação in vitro de Prunus spp.. 2004. 115f. Tese (Doutorado em Fruticultura de Clima Temperado) – Faculdade de Agronomia Eliseu Maciel. Universidade Federal de Pelotas, Pelotas. SILVA, L. C.; SCHUCH, M. W.; SOUZA, J. A.; ERIG, A. C.; ANTUNES, L. E. C. Meio nutritivo, reguladores de crescimento e frio no estabelecimento in vitro de mirtilo (vaccinium ashei reade) cv. delite. Revista Brasileira de Agrociência, v.12, p.405-408, 2006. SILVA, N. D. G. da. Radiação gama e conservação in vitro de amoreira-preta. 2013. 52f. Dissertação (Mestrado em Fisiologia Vegetal) – Universidade Federal de Pelotas, Pelotas. SOTIROPOULOS, T. E.; FOTOPOULOS, S.; DIMASSI, K. N.; TSIRAKOGLOU, V.; THERIOS, I. N. Response of the pear rootstock to boron and salinity in vitro. Biologia Plantarum, v.50, n.4, p779-781, 2006.

83
SOUZA, A.S.; COSTA, M.A.P.C.; SANTOS-SEREJO, J.A.; JUNGHANS, T.G.; SOUZA, F.V.D. Introdução à cultura de tecidos de plantas. In: SOUZA, A.S.; JUNGHANS, T.G. Introdução à Micropropagação de Plantas. Cruz das Almas: Embrapa Mandioca e Fruticultura Tropical, p.11-35, 2006. SOUZA, A. G. Produção de mudas enxertadas de pereira e pessegueiro em sistema hidropônico. 2010. 91f. Dissertação (Mestrado em Agronomia/Fitotecnia) – Universidade Federal de Lavras, Lavras. SOUZA, R. M.; NOGUEIRA, M. S.; LIMA, I. M.; MELARATO, M.; DOLINSKI, C. M. Management of the guava root-knot nematode in São João da Barra, Brazil, and report of new hosts. Nematologia Brasileira, v.30, p.165-169, 2006. TIBOLA, C. S.; RADMANN, E. B.; RODRIGUES, A. C.; FORTES, G. R de L.; FACHINELLO, J. C. Diferentes meios de cultivo no enraizamento in vitro de porta-enxertos de Prunus sp. Revista Brasileira de Agrociência, v.10, n.2, p.191-195, 2004. TOMBOLATO, A. F. C.; COSTA, A. M. M. Micropropagação de plantas ornamentais. Boletim técnico n.174. Campinas: Instituto Agronômico, 1998. 72p. WATANABE, Y.; YUKAWA, M.; HEE-SUN., K.; NISHIMURA, Y. Radiation effects on growth and seed germination of Arabidopsis. Annual Report 1999–2000. National Institute of Radiological Sciences, Anagawa, Chiba-shi, Japan, 2000. VAN HARTEN, A. M. Mutation Breeding: Theory and Practical Applications. Cambridge: Cambridge University Press, 1998. VIEIRA, R. L.; LEITE, G. B.; WAMSER, A. F. Efeito de substratos porosos no enraizamento in vitro do portaenxerto de macieira M-9 (Malus pumilla). Revista Brasileira de Fruticultura, v.29, n.1, p.128-132, 2007. VISSER, T.; VERHAEGH, J. J.; DE VRIES, D. P. Pre-selection of compact mutants induced by X-ray treatment in apple and pear. Euphytica, v.20, p.195–207, 1971. VILLA, F.; PASQUAL, M.; FRAGUAS, C.B.; REZENDE, J.C.de. Enraizamento in vitro do portaenxerto de videira „VR043-43‟: efeito do ANA e NaCl. Agrarian, v.1, n.2, p.103-111, 2008. ZANANDREA, I.; BACARIN, M. A.; SCHMITZ, D. D.; BRAGA, E. J. B.; PETERS, J. A. Chlorophyll Fluorescence in in vitro cultivated Apple. Revista Brasileira de Agrociência, v.12, p.305-308, 2006.

84
CONSIDERAÇÕES FINAIS
No decorrer das pesquisas realizadas, foi possível verificar que as cultivares
estudadas (Carrick, Cascatense e Ya-Li) respondem diferentemente quanto às
condições de cultivo testadas. A „Ya-Li‟ foi a cultivar que apresentou melhor
desempenho, tanto in vitro como ex vitro, o que possibilitou, assim, seu estudo mais
detalhado. Porém, os resultados obtidos nos experimentos de multiplicação in vitro
permitiram a obtenção de altas taxas de multiplicação do material vegetal,
possibilitando, desta forma, que as três cultivares sejam propagadas em larga
escala. Neste sentido, esta fase, acoplada às demais etapas do processo de
micropropagação potencializa sua produção, originando, assim, mudas com
qualidade. A multiplicação in vitro das cultivares pode ser potencializada com o uso
do meio MS Himédia comercial, com suplementação de 1,2 mg L-1 de BAP e 30 g L-1
de sacarose para „Carrick‟; MS com 1,2 mg L-1 de BAP e 45 g L-1 de sacarose para
„Ya-Li‟ e MS Himédia, suplementado de 0,8 mg L-1 de BAP e 30 g L-1 de sacarose
para „Cascatense‟.
Em relação aos experimentos que envolveram a irradiação do material
vegetal, embora o uso de doses mais elevadas de irradiação (40Gy) possam
interferir em alguns parâmetros de crescimento inicial (ex.: comprimento das
brotações), as doses testadas não acarretam efeitos severos na micropropagação
da cv. Ya-Li, sendo, por vezes, até estimulatórias, como no caso das doses mais
baixas (10Gy). Esta dose permite que as plantas irradiadas se assemelhem ao
material não irradiado em relação à responsividade em alguns parâmetros da fase
de indução das brotações e tende a estimular um melhor enraizamento das
mesmas, juntamente com o uso de vermiculita. Desta maneira, foi possível a
obtenção de mudas de material irradiado que servirão para estudos posteriores na
tentativa de obter genótipos com características de interesse, a partir de acréscimos
na variabilidade genética, em vista que há cerca de 750 plantas de „Ya-Li‟ no campo

85
da Embrapa Clima Temperado (controle + 46 diferentes plantas irradiadas), sendo
que as folhas já foram coletadas e acondicionadas em ultra-freezer, para serem
realizadas as análises moleculares e há continuidade do trabalho pelos melhoristas
da cultura.
Durante o primeiro ano de doutorado também foram realizados experimentos
de regeneração in vitro das três cultivares, variando-se as concentrações de TDZ
(Thidiazuron) (0, 1, 2, 3 e 4 mg L-1) e ANA (ácido naftaleno acético) (0,0; 0,2; e 0,3
mg L-1), no qual utilizou-se folhas inteiras ou seccionadas, com o lado adaxial ou
abaxial em contato com o meio de cultivo, com ou sem pecíolo, diferentes dias de
cultivo no claro e/ou no escuro, além de um período de 2h em meio líquido contendo
2,4-D (ácido 2,4 diclorofenoxiacético) (0,5 mg L-1). Os resultados obtidos foram
incipientes, o que tornou difícil descrevê-los, porém melhores taxas de regeneração
in vitro foram encontradas na „Ya-Li‟ (33,33%), com o uso de meio de indução MS,
suplementado com 1 mg L-1 de TDZ e 0,1 mg L-1 de ANA, cultivados durante 40 dias
no escuro, utilizando-se folhas inteiras, escarificadas no limbo e na nervura central,
sem pecíolo, com a face abaxial em contato com o meio. Após este período de
escuro os explantes permaneceram 30 dias em fotoperíodo de 16 horas e 48 μmol
m-2 s-1 de fluxo de fótons, em meio MS + 1 mg L-1 de TDZ.
Devido à falta de pesquisas em relação ao efeito do pH em plantas
autoenraizadas de pereira (pé franco), no terceiro ano de doutorado foi realizado um
experimento com diferentes pHs (4,5; 5,5; 6,5 e 7,5), em plantas oriundas de
sementes da cv. Ya-Li, as quais foram cultivadas em hidroponia, durante 60 dias. A
partir disso, foram realizadas avaliações de parâmetros de crescimento, conteúdo de
macro e micronutrientes, carboidratos e atividades das enzimas antioxidantes. De
acordo com os resultados obtidos, foram verificadas diferenças no crescimento
inicial das plantas, comprovando a variabilidade genética existente entre plantas
obtidas de sementes e que o pH pode influenciar no crescimento e desenvolvimento
destas. Dentre os pHs utilizados, os pHs 4,5 e 5,5 determinaram melhores respostas
quando todos os parâmetros foram analisados. Os dados não foram adicionados a
este trabalho, porém este estudo preliminar servirá de base para experimentos
posteriores, buscando selecionar plantas que sejam mais adaptadas a pHs mais
baixos.





























