Embed Size (px)
Citation preview

173
The actions of enzymes on lampbrush chromosomes
By H. C. MACGREGOR and H. G. CALLAN
(From the Department of Natural History, The University, St. Andrews, Fife)
With 6 plates (figs. 2 to 5, 9 and 10)
SummaryThe chromomeres of lampbrush chromosomes of Triturus cristatus are Feulgen-positive; they therefore contain DNA. After removal of their DNA in boiling tri-chloracetic acid, the chromomeres stain with fast green at alkaline pH; they thereforecontain basic protein. The lateral loops are Feulgen-negative; they stain with toluidineblue at acid pH, but much less intensely following RNase digestion; they thereforecontain RNA. The spheres of chromosomes V and VIII do not contain RNA.
Unfixed lampbrush chromosomes retain a life-like appearance in 0-07 M K/NaClat pH 6-2; in this medium the nuclear sap disperses. As pH is raised to 8rs the matricesof lateral loops dissolve but chromosome axes remain unbroken. Above pH 8-5 lamp-brush chromosomes dissolve. As pH is lowered from 6-2, at between 5-8 and 5-4coagulation occurs. If pH is rapidly reduced still further, a persistent relaxed condi-tion sets in between 2-5 and 2.
In concentrations of K/NaCl above 0-5 M lampbrush chromosomes dissolve.Lateral loop matrices dissolve in 0-25 M K/NaCl but chromosome axes remain un-broken. In concentrations of K/NaCl below 0-05 M lateral loop matrices dissolve,but even in distilled water chromosome axes remain unbroken.
Trypsin at pH 6-2 and at pH 7-8 strips the matrices from lateral loops and occasion-ally breaks matrix fusions. It causes chromomeres to swell and coalesce, but fails tobreak chromosome axes. The action of 'pan-protease' resembles that of trypsin inall respects.
Pepsin at pH 6-2 strips the matrices from lateral loops, but does not destroychromomeres. At low pH peptic digestion is slow: the enzyme is attacking coagulatedchromosomes; but if peptic digestion precedes a lowering of pH the overall outcome isa rapid solution of loop matrix, and under these conditions matrix and sphere fusionsare broken. If trypsin or 'pan-protease' digestion precedes a lowering of pH there isa similarly rapid solution of loop matrix; thus the action is not specifically referableto pepsin. Under no conditions does pepsin break the axes of lampbrush chromo-somes.
RNase at pH 6 2 strips the matrices from lateral loops; this action is detectable atextreme dilution. RNase does not destroy chromomeres, nor does it break chromo-some axes. If tryptic digestion follows RNase digestion this too fails to break chromo-some axes.
Unlike the proteolytic enzymes and RNase, DNase at pH 6-2 breaks the fibrilbetween adjacent chromomeres, and it also breaks the axes of lateral loops.
Contrary to Mazia's experience with salivary gland chromosomes, versene does notbreak the axes of lampbrush chromosomes even when applied in media of low electro-lyte concentration.
These results indicate that uninterrupted fibres of DNA run throughout the lengthsof lampbrush chromosomes.
[Quarterly Journal of Microscopical Science, Vol. 103, part 2, pp. 173-203, June 1962.]

174 Macgregor and Callan—Actions of enzymes
Contentspage
I n t r o d u c t i o n . . . . . . . . . . . . 1 7 4M a t e r i a l s a n d m e t h o d s . . . . . . . . . . . 1 7 6S e l e c t e d c h r o m o s o m e r e g i o n s . . . . . . . . . . 1 7 8
T r i t u r u s c r i s t a t u s c a r n i f e x : m i d d l e of chromosome I I . . . . . . 1 8 0T. c. carnifex: ends of left arms of chromosomes V and VIII • • . . 1 8 0T . c. c a r n i f e x : giant loops of chromosome XI . . . . . . . 180T . c. c r i s t a t u s : end of left arm of chromosome XII . . . . . . 1 8 1T . c. k a r e l i n i i : centric regions . . . . . . . . . . 1 8 1
T h e s t a i n i n g of fixed c h r o m o s o m e s . . . . . . . . . 1 8 2Feulgen's reagent and light green . . . . . . . . . 1 8 2Toluidine blue, with and without previous digestion by ribonuclease . . . 183
Fast green FCF 184Observations on unfixed chromosomes . . . . . . . . 1 8 4
Effects of varying pH . . . . . . . . . . . 1 8 5Effects of varying saline concentration . . . . . . . . 1 8 6Trypsin 188Pan-protease . . . . . . . . . . . . . 1 8 9Pepsin . . . . . . . . . . . . . 1 9 0Ribonuclease . . . . . . . . . . . . 1 9 4Deoxyribonuclease . . . . . . . . . . . . 1 9 5Versene . . . . . . . . . . . . . 1 9 6
D i s c u s s i o n . . . . . . . . . . . . . 1 9 7R e f e r e n c e s . . . . . . . . . . . . . 2 0 2
IntroductionT H E dissection of organic matter by means of enzymes has become a poten-tially refined operation now that several reasonably pure enzymes are available.The present paper describes effects produced by trypsin, 'pan-protease',pepsin, ribonuclease (RNase) and deoxyribonuclease (DNase) on lampbrushchromosomes isolated from newt oocytes. It also describes the actions ofsalines of various pH values and ionic strengths on these chromosomes,actions which must be taken into account when we attempt to interpret theeffects of enzymes. Furthermore, the lack of action of the chelating agentversene on isolated lampbrush chromosomes is recorded.
Because of the key role played by chromosomes in determining schemesand details of biological organization, there has for long been a widespreadinterest in their chemical make-up, and many analyses have been attempted.Indeed, if we equate chromosomes with sperm-head nuclei, we may date theearliest of these analyses back to the classical research published by Miescherin 1871. Chemical analyses of sperm-head nuclei in bulk have repeatedlyshown that the two main constituents are deoxyribonucleic acid (DNA) andarginine-rich protamine (Felix, 1952). Analyses of interphase nuclei in bulk,and of the so-called 'chromosomes' prepared by centrifugation of disruptedinterphase nuclei, have demonstrated that the main constituents of suchmaterial are DNA, histone, tryptophane-containing protein, and ribonucleicacid (RNA) (Mirsky and Ris, 1947; Mirsky, 1947). If allowance be madefor polyploidy, constancy in the quantity of DNA per interphase nucleusfrom a given organism, irrespective of the tissue of origin, has been established;

on lampbrush chromosomes 175
and this quantity has been demonstrated as twice that present in sperm-headnuclei from the same organism (Vendrely and Vendrely, 1948; Swift, 1950;Mirsky and Ris, 1951; Leuchtenberger, 1952; Frazer and Davidson, 1953).There is, furthermore, some evidence for the assumption that there isa quantitative parallel between the DNA and histone contents of interphasenuclei (Vendrely and Vendrely, 1953; Alfert, 1955; Bloch and Godman,1955; Mirsky and Ris, 1951).
Inferences concerning the chemical make-up of chromosomes have beendrawn from these bulk analyses, and some of these inferences are contra-dictory (Mirsky and Ris, 1947; Mirsky, 1947; Polli, 1951). Since inter-phase nuclei are not just chromosomes, and since there are doubts as towhether Mirsky's interphase 'chromosomes' are genuine (Lamb, 1950), thecontradictions are hardly surprising. If compelling evidence for the chemicalmake-up of chromosomes is sought, then clearly the observations must bemade on chromosomes as such.
No one has yet succeeded in collecting mitotic or meiotic chromosomes,uncontaminated by other cellular components, for bulk analysis; neither hasthis been achieved for the giant polytene chromosomes of larval dipteransalivary glands. On the other hand, there have been several qualitativeanalyses of giant chromosomes, both from dipteran salivary glands and fromamphibian oocyte nuclei, and some of these studies have included observa-tions of the effects of enzymatic digestion. We can at least be sure that suchstudies relate to genuine chromosomes, but nevertheless they too have givenconflicting results.
The particular qualities of lampbrush chromosomes, which enable theirenzymatic degradation to be critically observed, were emphasized by Gall(1954) in his paper on Triturus viridescens. Whereas D'Angelo (1946) andother workers have tried but have failed to separate unfixed and undamagedpolytene chromosomes from the cytoplasm and nuclear sap of dipteran salivarygland cells, the isolation of lampbrush chromosomes in lifelike condition insaline is a simple procedure. The isolated chromosomes may then be observedwith an inverted phase-contrast microscope, the system first advocated and usedby Gall (1954), and provided they lie in a suitable saline their morphologyremains unchanged for several hours. Enzymes or other agents can now beapplied to such preparations, and any morphological changes which ensuecan be observed directly. Because one can dispense with fixatives and stainswhen studying lampbrush chromosomes, artifacts introduced by these aidsto observation can be eliminated. The use of conventional cytologicaltechniques has been at least in part responsible for the conflicting resultsobtained by previous workers in this field.
Newts have larger lampbrush chromosomes than other organisms, andhence were chosen for the present study. Lampbrush chromosomes aremorphologically heterogeneous, and we wished to take account of this hetero-geneity when observing the effects of enzymes. Callan and Lloyd (19605)have described the morphologies of the lampbrush chromosomes of four

176 Macgregor and Callan—Actions of enzymes
geographical races (subspecies) of T. cristatus (Laurenti) in some detail, andin the present study we have concentrated attention on selected chromosomeregions from newts belonging to three of these races. To avoid a repetitionhere of much background information, the reader is referred to Callan andLloyd's (1960&) paper. We now follow the terminology of that paper, andhold to the conventions there established when defining particular sites onthe chromosomes.
As well as the lampbrush chromosomes, there are other objects in newtoocyte nuclei which can be seen with a light microscope and which in life,like the chromosomes, are embedded in nuclear sap. These objects are un-avoidable contaminants of preparations of lampbrush chromosomes. Theyfall into two classes: detached chromosome products and 'nucleoli'. Observa-tions on the 'nucleoli' are deliberately omitted from the present paper; the'nucleoli' of amphibian oocytes are almost certainly not homologous to thenucleoli of somatic cells, and it is uncertain whether they are detached chromo-some products. Their peculiarities will be discussed elsewhere.
Nuclear sap diluted in saline is also a contaminant of preparations oflampbrush chromosomes. Brown, Callan, and Leaf (1950) have demonstratedthat the nuclear sap of newt oocytes consists in the main of protein or proteinswhich, on hydrolysis, yield an extensive range of amino-acids. These findingshave been confirmed by Gall (1952). The sap protein is soluble in certainsalines which do not dissolve components of the lampbrush chromosomes,and hence contamination with nuclear sap is of little consequence whenstructural components of the chromosomes are being qualitatively analysed.
Materials and methodsThe three subspecies of the crested newt T. cristatus which have been
used in this study are: T. c. cristatus (Laurenti), collected in the south ofEngland and supplied by the dealer L. Haig of Newdigate, Surrey; T. c.carnifex (Laurenti), collected near Naples, Italy, and supplied by Dr. P. Dohrnof the Stazione Zoologica; and T. c. karelinii (Strauch), collected nearIstanbul and supplied by Dr. A. Sengiin.
Portions of ovaries were removed from anaesthetized newts and stored' dry' in covered glass containers, the covers being sealed in place with a paraffinoil / vaseline mixture. Although portions of ovaries stored at about 2° Cremain in good condition for several days, in this study we have workedexclusively with freshly excised ovary fragments. We have as far as possiblestandardized our material by choosing for study oocytes from within a re-stricted size range. Such oocytes are casually referred to as 'half-grown', andmeasure about i-o mm diameter in cristatus, 0-9 mm diameter in carnifex,and i-1 mm diameter in karelinii. In oocytes of these sizes the great majorityof the lateral loops of the lampbrush chromosomes are not at their longest(they are longer in smaller oocytes), but 'landmark' structures are welldeveloped and permit the ready identification of chromosomes and parts ofchromosomes.

on lampbrush chromosomes 177
Details of the methods used for isolation of oocyte nuclei and for removalof nuclear membranes have already been given by Callan and Lloyd (io,6o£),where Gall's observation chamber for use with an inverted microscope isillustrated. Only the bare essentials of our routine procedure will be statedhere. Each observation chamber consists of a 3 in. X1 in. microscope slidethrough which a \ in. diameter hole has been bored; a number o coverslip issealed to one side of the slide with paraffin wax, across the hole, and this formsthe floor of the chamber. A nucleus is isolated and freed from cytoplasm andyolk in a 5:1 mixture of o-i M potassium and sodium chlorides (the K/Naratio of 5:1 being in accordance with unpublished data of Dr. W. T. W.Potts) and is then transferred to Callan and Lloyd's medium C in an observa-tion chamber for removal of its membrane and for the sap to disperse.Medium C consists of 7 parts of the 5:1 o-i M K/NaCl mixture, togetherwith 3 parts of o-ooi M KH2PO4. This saline, after sterilization by boiling,has a pH of 6-2.
The observations which we describe in this paper were all made on freshlyisolated nuclei, and the changes induced by the various agents were allsufficiently rapid to be ascribable to these agents, not to natural deterioration.
When about 50 mm3 of medium C is placed in an observation chamber itforms a meniscus convex upwards. Once the nuclear membrane has beenremoved from a nucleus lying in the chamber, a small square coverslip isdropped in place over the saline, care being taken to avoid trapping airbubbles in the chamber. The preparation is now observed with an invertedphase-contrast microscope (Cooke, Troughton, and Simms Ltd., York)giving 'dark contrast' (alternatively known as 'bright field'), and furtheraction is delayed for a few minutes until the nuclear sap has sufficientlydispersed to leave the chromosomes spread out in one plane over the lowercoverslip. Nuclei from oocytes in the size range which we have chosen havediameters of the order 0-35 mm, and hence are of volume approximately0-022 mm3. Thus when the sap has fully dispersed its dilution is more thantwo thousandfold.
Each agent whose effects we have studied has been applied by pipette asa drop of solution beside the top coverslip; and we have endeavoured to addthe same quantity of fluid (50 mm3) as that previously present in thechamber. The coverslip rises to accommodate the extra fluid, which thenmixes with the original medium C. Description of the control over pH, andfurther technical details, is deferred to the several chapters devoted to parti-cular agents which we have used. In these chapters the term 'concentration'of an agent, unless otherwise qualified, refers to the ultimate concentrationreached after mixing with medium C in the observation chamber.
To record morphological changes in the lampbrush chromosomes, photo-graphs of the primary image formed by the objective lens have been takenwith a single exposure camera loaded with Ilford micro-neg pan film. Forcertain series of photographs a xenon flash served as the light source, and thissuccessfully 'stopped' Brownian movement. However, for series attempting

178 Macgregor and Callan—Actions of enzymes
to record rapid changes the time-lag between successsive exposures, neces-sitated by recharging of the condenser unit which serves the flash, provedtoo great; and a conventional light source with a 'compur' shutter of3 5-mm aperture on a vibration-free mount interposed between the light andthe microscope condenser was therefore used instead, the exposures givenbeing one-tenth or one twenty-fifth sec. Although such exposures fail toeliminate blurring due to Brownian movement, they proved adequate for ourneeds.
Series of negatives were processed identically, and so were prints taken fromnegatives. In this way an impression of changes in the refractility of compon-ents of the lampbrush chromosomes, albeit imperfect because of the limita-tions of the phase-contrast system, has been recorded.
The enzymes pepsin and trypsin were purchased in crystalline form fromArmour Pharmaceutical Co. Ltd. (Eastbourne, England). Worthington's'pan-protease' was provided by Dr. D. Mazia, who suggested that we com-pare its action with that of trypsin. Ribonuclease (RNase) and deoxyribonu-clease (DNase) were first obtained from Nutritional Biochemicals (Cleveland,U.S.A.), later from L. Light (Colnbrook, England). The chelating agentversene (diamino-ethane-tetra-acetic acid, disodium salt) was obtained fromBritish Drug Houses (Poole, England).
Measurements of pH were taken with a Cambridge bench-type meterusing a glass electrode.
Selected chromosome regionsThe structural organization which is common to all lampbrush chromo-
somes has been discussed in several recent papers (Callan, 1955, 1957;Callan and Lloyd, 1960a; Duryee, 1950; Gall, 1954, 1956) and, moreparticularly related to this paper, the lampbrush chromosomes of T. cristatushave been described by Callan and Lloyd (19606). In the present paperattention will be confined to those aspects of the structural organization onwhich the enzyme work directly bears.
Each oocyte nucleus of T. cristatus contains 12 pairs of lampbrush chromo-somes, homologous chromosomes being associated at one or more points sothat they form bivalents. There is a series of granules, chromomeres, in eachlampbrush chromosome axis, neighbouring granules being connected to oneanother by a thin, extensible and elastic fibre. Pairs or multiples of pairs ofloops project laterally from the overwhelming majority of chromomeres, andmost of these lateral loops conform to the type described by Callan and Lloydas 'normal'. Such loops have a fine fibrous texture, the fibres appearing toproject radially from a denser loop axis. Throughout the present paper wehave applied the term 'matrix' to this fibrous material. There is a gradationin the amount of matrix surrounding loop axis as one follows a loop out fromits 'parent' chromomere and through its length back to the same chromomere,one end of the loop being bare of matrix.
When not impeded by nuclear sap and when not coagulated, normal lateral

on lampbnish chromosomes 179
loops in saline show Brownian movement. Brownian movement is dis-advantageous for photographic recording, but in other respects it is an asset:whereas undamaged normal lateral loops retain their linear continuity despitethe stresses to which they are evidently exposed, breaks produced by anyagent are immediately apparent; and the independent movements of the
V
VI
VII
VIII
9 9
it' n
t i $ i* i
XII
FIG. 1. Working map of the 12 lampbrush chromosomes of T. c. carnifex. Centromerepositions are indicated by the vertically alined arrows. Structures which often show 're-
flected fusion' are linked together by brackets.
matrix fibres show that these fibres are each attached to loop axis, not to oneanother.
Some normal lateral loops are present in all the regions which we haveselected for special study, but also included in these regions are other objectshaving distinctive morphologies, objects which have been used by Callan andLloyd as 'landmarks' for chromosome identification. A working map of thelampbrush chromosomes of T. c. carnifex, showing the relative lengths of the

180 Macgregor and Callan—Actions of enzymes
12 members of the complement, the positions of their centromeres, and of themajor landmarks is given in fig. 1. Whenever the positions of landmarks aredescribed in terms of units, a unit is defined as being 'one-hundredth part ofthe length of chromosome V, or its equivalent in length on another chromo-some' (Callan and Lloyd, 19606). On the working map the longer arm ofeach chromosome is drawn projecting to the left, so defining the 'left-handend' of each chromosome. The chromosome regions which we have selectedfor special attention will now be detailed.
T. c. carnifex: middle of chromosome II
The left end of this region is defined by 'marker' loops at 56 units, the rightend by a large granule, not associated with conspicuous lateral loops, whichlies in the chromosome axis at 78-5 units. The marker loops are stiff andgranular in texture, and there is a particularly large granule at the point wherethey are attached to the chromosome axis. Through the mid-region of chromo-some II dense 'lumpy' structures project at intervals laterally from the axis,the largest of these being situated at 61-5 and 69^5 units. The lumpy objectsmay be fused together in various ways: within the length of one and the samechromosome, producing reflections of its axis; or between partner chromo-somes, not necessarily at homologous loci. The centromere lies between thelargest of the lumpy objects, at 65 units, and in carnifex (as in cristatus) it isa smooth, round, dense granule which lacks lateral loops.
T. c. carnifex: ends of left arms of chromosomes V and VIII
' Spheres' are the striking features of these two regions, there being spheresat 2 and 11 units on chromosome V and a single sphere at 3-5 units on chromo-some VIII. These objects are smooth and round with firm outlines, they mayreach up to 15 /A in diameter, they may be homogeneous in texture or they maycontain one or more vacuoles (fig. 2, H). During their formation spheres aretightly attached to chromosome axes, but detached spheres are often presentin newt oocyte nuclei and they are the most conspicuous and easily recognizedof freed chromosome products. Two chromosome sites concerned with sphereformation may both be connected to a single sphere, and such 'sphere-fusions' may associate homologous or non-homologous sites. The left armsof chromosomes V and VIII end in telomeres: smooth, round, loop-freestructures which are characteristic of the ends of all lampbrush chromosomes.Telomere fusions are common: usually they associate homologous ends, butoccasionally the fusions are non-homologous.
T. c. carnifex: giant loops of chromosome XI
These massive structures lie in the left arm of chromosome XI at 24 units.They have well-defined but irregular outlines. They are formed of stiff, highlyrefractile and often vacuolated material (figs. 3, A; 9, E). The degree to whichthey preserve the form of loops is variable from oocyte to oocyte; the loopform may be partially or totally obliterated by fusion, fusions occurring within

on lampbrush chromosomes 181
single loops, between sister loops or between homologous loops. Fusions canalso take place between these loops and structures of similar texture which incertain individual newts are also present on chromosomes X and XII. Inlarger oocyte nuclei big, roughly spherical bodies may detach from the giantloops and for a time whilst free in the nuclear sap they conserve the texture ofthe parent structures; but oocyte nuclei of the size with which we are hereconcerned generally lack free products from this source.
T. c. cristatus: end of left arm of chromosome XII
The landmark to which we have given particular attention in this chromo-some region is a pair of giant granular loops at 9 units. These loops, each ofwhich may exceed 100 /x in overall extended length (fig. 9, A) are a feature ofsubspecies cristatus; they are absent from the other subspecies. Each giantgranular loop shows a striking gradation in form along its length. At oneend a thin strand leaves the chromomere of origin; this becomes thicker andtwisted in a spiral; then follows a zone of variable extent where refractile rodsare wrapped around, or project radially from, the loop axis; the rest of theloop, two-thirds or so of its length, consists of granules or strings of granulesattached by very thin fibres to the loop axis, the granules progressivelydiminishing in size as the other end of the loop is approached. The axis ofthe giant granular loop is usually not visible in a freshly made preparation,but with continuing dispersal of nuclear sap (or possibly due to the solutionof some constituent of the loop itself) the axis becomes evident after severalhours. The granules or strings of granules in this loop show independentBrownian movement, but they do not scatter; this is our main justificationfor claiming that they are attached by fine fibres to the loop axis.
When describing experiments involving the granular loops we have usedthe term 'matrix' as referring to the fibrous material attaching granules to theloop axis, and to any other diffuse material which may be accumulated in thespaces between fibres and granules. Although the giant granular loops areexceptional and readily identifiable lampbrush loops, they are exceptional inone respect only, their great size. As Gall (1954) found also in T. viridescens,many smaller lateral loops of all subspecies of T. cristatus carry refractilegranules which show independent Brownian movement, and the thinnerinsertions of lateral loops of diverse morphologies are not infrequently woundin tight spirals.
The left arm of chromosome XII terminates in a typical telomere; all thetelomeres of cristatus chromosomes are larger than the comparable structuresof other subspecies, and telomere fusions are more frequent.
T. c. karelinii: centric regions
The centromeres of all karelinii lampbrush chromosomes are flanked bydense 'axial bars' which lack lateral loops. This is a feature which kareliniidoes not share with other subspecies of T. cristatus, though Gall (1954) hasdescribed similar structures in the lampbrush chromosomes of T. viridescens

182 Macgregor and Callan—Actions of enzymes
and Amblystoma tigrinum. The axial bars increase in length as oocytes grow insize, and Callan and Lloyd (19606) have demonstrated that this results fromprogressive incorporation of the substance of neighbouring chromomeres,which withdraw their lateral loops as they amalgamate with the axial bars.Most chromomeres are so tiny that their reactions to agents are difficult toobserve. For our purposes the axial bars of karelinii are conveniently largeaggregates of chromomere material, and, moreover, they betray the positionsof the centromere granules, which in other subspecies are often difficult tofind. Though we have not observed centromere granule fusions in cristatusor carnifex, centromere fusions are commonly present in karelinii bivalentsand the nature of these attachments is of some interest. Furthermore, inregions near to the centromeres of karelinii chromosomes II, III, IV, V, VI,and VIII, fusions between what look, at first glance, to be unusually largechromomeres are frequently encountered. These structures, called 'axialgranules' by Callan and Lloyd, may fuse between homologues at homologoussites; but they may also fuse non-homologously, and when two axial granuleson the same chromosome are fused to one another a 'reflected' portion of thechromosome axis results. We have studied the actions of enzymes on suchfusions in an attempt to discriminate between those which do and those whichdo not include chiasmata.
The staining of fixed chromosomesAlthough the fixation and staining of lampbrush chromosomes does not
fall within the title limits of this paper, we include a chapter on the subjectbecause it is complementary to the enzyme study.
Gall (1952) has shown that formaldehyde vapour and osmic acid vapourfix isolated lampbrush chromosomes in a reasonably adequate way, thoughboth these agents cause chromosome axes and lateral loops to contract, stiffen,and increase in refractility. We have fixed exclusively with formaldehydevapour. Fig. 2, A shows bivalent XI of T. c. karelinii unfixed, and fig. 2, Bshows the same bivalent after 15 min exposure to formaldehyde vapour.
Fixation in the literal sense, i.e. attachment to a glass surface, is necessaryif isolated lampbrush chromosomes are to be studied as stained preparations.To ensure attachment we have allowed oocyte nuclear sap to disperse forat least 30 min in medium C before placing slides in a formaldehyde vapourchamber. During fixation the slides must not be disturbed, otherwise thechromosomes fail to attach to the coverslip below. After 1 h in formaldehydevapour all fixed chromosome preparations were placed in 4% neutralizedformaldehyde solution for 12 h or more.
Feulgen's reagent and light greenAfter fixation, each observation chamber was washed in water, and then
placed in N HC1 at 6o° C for 6 min hydrolysis. During hydrolysis the cover-slip drops off the bored slide and bears the lampbrush chromosomes providedthey were firmly attached during fixation. Each coverslip was recovered and

on lampbrush chromosomes 183
placed for 2 h in Feulgen's reagent (prepared according to Coleman's 1938recipe), rinsed twice in'SO2 water', rinsed in water, taken up through alcoholsto 70%, counter-stained for 3 min in 0-2% light green in 70% alcohol,dehydrated, cleared, and mounted in DePeX (G. T. Gurr, London).
We find that the chromomeres of lampbrush chromosomes stain withFeulgen's reagent, and thus confirm the observations of Brachet (1940),Dodson (1948), Guyenot and Danon (1953), and Gall (1954). Furthermore,we confirm Gall's (1954) finding (made on T. viridescens) that centromeregranules and the 'axial bars' adjacent to the centromeres of T. c. karelinii arealso Feulgen-positive. None of the structures attached laterally to chromo-some axes appear Feulgen-positive, though these structures survive the mildhydrolysis necessary for the Feulgen reaction and stain with light green.Feulgen / light green staining reveals that telomeres and axial granules arecompound structures, despite the fact that when observed by phase contrastunfixed they simply appear to be large chromomeres. Each telomere and axialgranule consists of a crescent-shaped Feulgen-positive part closely applied toa spherical or nearly spherical mass staining with light green. Fig. 2, C, Dshows part of bivalent XI of karelinii (the same bivalent as that illustratedunfixed in fig. 2, A) from a preparation stained in Feulgen's reagent andlight green. Fig. 2, c was photographed using as a filter Feulgen's reagentdeveloped with formaldehyde, and it shows the distribution of material-stained by light green. Fig. 2, D was photographed using a solution of lightgreen as a filter; although the light green-stained lateral loops are visible inthe photograph, the Feulgen-positive axial structures (particularly the barsadjacent to the two centromeres) can be distinguished.
Toluidine blue, with and without previous digestion by ribonuclease
Formalin-fixed slides were washed in water and the coverslips bearinglampbrush chromosomes were then prized off. Some of these preparationswere stained for 40 min in 0-2% solution of toluidine blue (G. T. Gurr,London) in o-oi M phosphate buffer at pH 5-4, then washed in similarbuffer, run quickly through an alcohol series, cleared, and mounted in DePeX.In these circumstances all components of the chromosomes, both axial andlateral, stained with toluidine blue, staining intensities being roughly inaccordance with the refractilities of the various structures as observed un-fixed by phase contrast.
Other preparations prior to staining were incubated for 2 h at 370 C inribonuclease at 0-2 mg/ml dissolved in o-oi M phosphate buffer at pH 6, withcontrols incubated in buffer only. The staining of control preparationsresembled in all particulars that of unincubated preparations. Preparationsincubated with ribonuclease showed normal staining of chromomeres and,surprisingly enough, of the spheres of chromosomes V and VIII, but markedlyreduced staining of all other lateral structures.
Gall (1954) found that ribonuclease pre-treatment eliminated rather thanmerely reduced the toluidine blue-staining of lateral loops; this discrepancy

184 Macgregor and Callan—Actions of enzymes
between our results is probably due to the use of stains from different sources.Our findings are otherwise in accordance; Gall found that the stainability ofthe 'knobs' of T. viridescens is unaffected by ribonuclease, just as we havefound for the spheres of T. cristatus; and they demonstrate that all lateralcomponents of lampbrush chromosomes, spheres ( = knobs) excluded, con-tain RNA.
Fast green FCF
We have used the method of Alfert and Geschwind(i953) for detecting basicproteins by means of fast green FCF at a pH between 8 and 8-5. Formalin-fixed slides were immersed in boiling trichloracetic acid for 30 min so asto remove both DNA and RNA from the chromosomes. During this treat-ment the coverslips drop off the bored slides. Each coverslip was recovered,washed in 70% alcohol, rinsed in water, and stained for 30 min in o-i% solu-tion of fast green FCF (National Aniline, U.S.A.) in 0-2 M Na2HPO4 at pH8*2. Preparations were subsequently washed in buffer, transferred direct to95% alcohol, dehydrated, cleared, and mounted in DePeX. In such prepara-tions only the chromomeres are stained; these axial components of lampbrushchromosomes therefore contain basic proteins.
Observations on unfixed chromosomesBefore considering the actions of special reagents on unfixed lampbrush
chromosomes, we must take note of how such chromosomes react to variationsin saline pH and concentration. Ideally, one would wish to start with know-ledge of the appearance of lampbrush chromosomes in their natural surround-ings, inside intact, living oocytes. Unfortunately, lampbrush chromosomesare not visible in living oocytes; they become visible when the nuclear sap isdiluted with saline, but we now face the problem of assessing whether theirstate is lifelike. The problem is not so intractable as it at first appears. Wecan recognize degrees of leaching out of components of lampbrush chromo-somes, when the nuclear sap has been greatly diluted, by reduction in thedensity of their phase-contrast images and by enhanced Brownian movementof the lateral loops; moreover, we can recognize when lampbrush chromo-
FIG. 2 (plate). A, bivalent XI of T. c. karelinii lying in medium C at pH 62, unfixed.B, same as A, after 30-min exposure to formaldehyde vapour.c, part of the bivalent shown in A and B, stained by Feulgen's reagent and light green, and
photographed using 'developed Feulgen' as a filter.D, same as c, but photographed using light green in 70% alcohol as a filter. Arrows point
to the axial bars by the centromeres.E, left end of bivalent V of T. c. carnifex, with fused telomeres and fused spheres. In medium
C at pH 6-2.F, same as E, but at pH 7.G, same as E and F, but at pH 8.H, end of left arm of chromosome V of T. c. carnifex, showing two attached spheres and
one free sphere (top left). In medium C, at pH 6'2.J, same as H, but at pH 5 and coagulated.K, same as H and J, but at pH 2 and 'relaxed'.
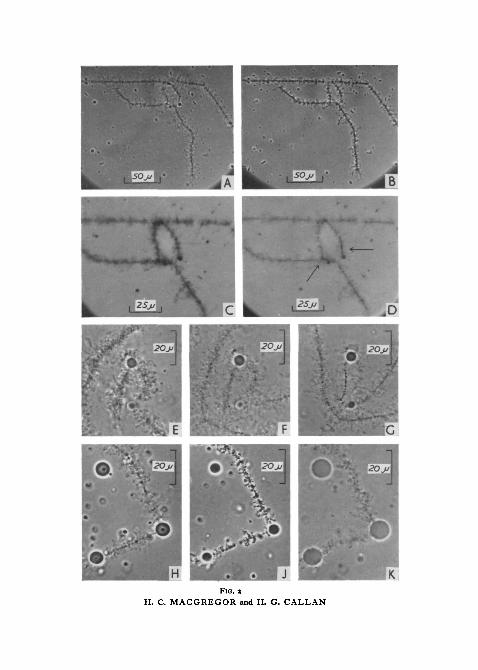
FIG. aH. C. MACGREGOR and H. G. CALLAN

FIG. 3H. C. MACGREGOR and H. G. CALLAN

on lampbrush chromosomes 185
somes have coagulated (see later). We have taken as our criterion of a lifelikestate that, with the nuclear sap dispersed, the chromosomes should showmaximum optical contrast consonant with the maintenance by normal lateralloops of a supple and relaxed form. Though this criterion is not entirelyobjective, it is useful.
Effects of varying pH
When studying the effects produced by pH alteration, medium C, itselfof pH 6-2, was used as the initial sap-dispersing saline in each observationchamber. In this medium lampbrush chromosomes maintain a lifelikestate for many hours. The pH was subsequently altered by pipetting a'modified' C medium around the top coverglass of the preparation; mixingfollows rapidly. Modified C media ranging in pH from 6-2 to a little above8 were prepared by replacing o-ooi M KH2PO4 with an equal fraction of o-ooiM KH2PO4/Na2HPO4 buffer. Modified C media ranging in pH from 8 to10 were obtained by adding small quantities of NaOH to ordinary C medium,whilst for pH values from 6-2 to i-8 small quantities of HC1 were similarlyadded. The quantities of NaOH or of HC1 so added were too small to havesignificantly altered the ionic strengths of such media. The pH reached aftermixing of media was not measured directly within each observation chamberbut was found by testing larger volumes of media mixed on an aliquotbasis.
As pH is increased within the range 6-2 to 8, lampbrush chromosomesshow progressively less and less contrast (fig. 2, E-G) but no other physicalchange. Once the pH has risen above 7, back adjustment to pH 6-2 fails torestore the original contrast, though in other respects the chromosomesremain lifelike. Thus although no visible aggregates detach from lamp-brush chromosomes between pH 7 and 8 we may assume that some loopmatrix constituents pass into solution.
FIG. 3 (plate). A, part of bivalent XI of T. c. carnifex, including two homologous pairs ofgiant loops. The arrow points to a pair of giant loops which have formed a 'double bridge'.In medium C at pH 6-2.
B, same as A, 3 min after addition of trypsin at effective concentration of 0-25 mp'ml andpH 6'2.
c, same as B, 1 min later.D, same as c, 1 min later. The arrow points to a 'double bridge' formed by unbroken axes
of the giant loops exposed as matrix has dissolved.E, middle region of bivalent II of T. c. carnifex. Arrows point to the centromeres. The
lumpy objects near the top of the photograph lie in the chromosomes' left arms. In mediumC at pH 6-2.
F, same as E, 1 min after addition of trypsin at effective concentration of 025 mg/ml andpH 7-8.
G, same as F, I min later.H, same as G, 13 min later.J, same as H, 40 min later.K, same as J, fixed by adding 4% formaldehyde to the preparation after 65 min of tryptic
digestion.All these photographs are at uniform magnification.

186 Macgregor and Callan—Actions of enzymes
Above pH 8 there is more evident solution of normal loop matrix. Thewidths of normal loops are reduced, previously discernible gradations in widthare largely obliterated, and increased violence of Brownian movement showsthat such loops have lost substance. As their matrix dissolves, normal loopscontract in length and instead of projecting from chromosome axes in smooth,sweeping curves they become irregularly bent and twisted, an observationwhich suggests that in their natural state the axes of normal lateral loopsare stretched out by accumulating matrix. The denser landmarks are lessstrikingly altered, though by pH 8-5 they too show significant loss of contrast.
At some pH between 8-5 and 9 there is a sudden fall in the contrast oflampbrush chromosomes, followed by their disappearance. Once havingdisappeared from view even fixation in formalin vapour fails to make thechromosomes visible again. We must conclude that they have dissolved.
Let us now turn to media more acid than pH 6-2. Between pH 6-2 and 5-8lampbrush chromosomes do not alter in appearance, but at some pH between5-8 and 5-4 the chromosomes stiffen, contract, and increase sharply incontrast (fig. 2, j). This is the easily recognizable coagulated condition, lamp-brush chromosomes taking on the same appearance when exposed to formal-dehyde or osmic acid vapours, or to calcium ions at molarities above io"3.At this same pH the nuclear sap, previously dispersed in saline, precipitates.Once coagulated by low pH, back adjustment to pH d-2 fails to restorelampbrush chromosomes to a lifelike state. The coagulated state is maintainedin media down to pH 2-5, and to still lower values if the rate of fall of pH isslow. If, on the other hand, pH is rapidly lowered, then at some valuebetween 2-5 and 2 there is a striking change in the appearance of lampbrushchromosomes. From a previously coagulated state, all chromosome com-ponents become relaxed, swollen, and of reduced contrast (fig. 2, K). Therelaxed condition persists provided pH is held below 2, but if pH is raisedabove 2-5 the coagulated state reappears and is now irreversible.
Effects of varying saline concentrationWhen an oocyte nucleus is isolated in an unbuffered 5:1 mixture of M KC1
and NaCl it swells, the nuclear sap clarifies and hydrates, whilst a componentof the sap investing the chromosomes, called by Callan (1952) the 'structuralcolloid' and by Duryee and Doherty (1954) the 'chromogel', sinks to thebottom of the nucleus. After the nuclear membrane has been removed thisstructural colloid may take up to 3 h to disperse. The lampbrush chromo-somes first become visible as ragged strands of low contrast: they lack lateralloops and all landmarks, and do not display chromomeric patterns. Asdispersal of the structural colloid continues, the chromosomes' axes fragmentand 3 h after isolation in M saline a cloud of tiny granules in violent Brownianmovement is all that remains of the original chromosome complement.
0-5 M K/NaCl is slightly less destructive to lampbrush chromosomes.About 2 h after isolation of a nucleus, when the structural colloid has dis-persed, occasional short lengths of axis show signs of chromomeric pattern,

on lampbrush chromosomes 187
but lack all traces of lateral loops. The spheres of chromosomes V and VIIIare the only landmarks remaining recognizable in saline of this strength.
In 0-25 M K/NaCl the structural colloid disperses within 30 min of isola-tion, revealing unbroken lampbrush chromosomes of low contrast. Thechromosomes carry lateral loops, but these have suffered so much loss ofsubstance that the various kinds of loops, including landmarks, are indistin-guishable from one another. The spheres of chromosomes V and VIII,alone amongst the landmarks, are recognizable. Chromomeres are visible, butswollen. The appearance of lampbrush chromosomes in 0-25 M salineremains unaltered for several hours after isolation; chromosome axes do notbreak in this medium.
In 01 M K/NaCl the nuclear sap remains stiff and slightly opaque forseveral minutes after isolation. The nuclear membrane can be peeled off andthereafter the sap disperses, allowing the chromosomes to spread out over thelower coverslip. Sap dispersal is well advanced within 20 min of removal ofthe nuclear membrane, but saline of this strength, in other respects a satis-factory isolation medium, usually fails to bring about complete dispersal ofthe structural colloid. A layer of this material persists between the chromo-somes and the coverslip forming the bottom of the observation chamber,interfering with critical observation and preventing the chromosomes fromspreading in one focal plane. Ino- i M K/NaCl the normal loops of lamp-brush chromosomes are of high contrast and the chromomeres dense andhighly refractile. Lampbrush chromosomes are well preserved in this medium;days may elapse before there is perceptible loss of substance from normal loops.
Medium C has an approximate molarity of 0-07, but unlike other salinesdiscussed in this section it contains a low concentration of KH2PO4 and hasa pH of 6-2 after sterilization by boiling. Medium C produces convenientlyrapid dispersal of the sap, including the structural colloid, after removal ofthe nuclear membrane. The chromosomes spread and come to lie flat on thebottom of the observation chamber. All their features are well preserved formany hours and we have noticed only slight deterioration in preparations keptin medium C at z° C for up to 2 weeks.
We have isolated oocyte nuclei and removed their membranes in unbufferedK/NaCl solutions of 0-05, 0-005, a n ^ o-ooi M, and we have examined thereactions of nuclei and chromosomes to isolation in distilled water. As saltconcentration of the medium is reduced the nuclear sap hydrates more andmore quickly and components of the lateral loops go into solution. Thematrix of normal loops is most sensitive to low saline concentration, dissolvingrapidly in media which are less than 0-05 M. The spheres of chromosomesV and VIII and the giant loops of chromosomes X, XI, and XII are the mostresistant extra-axial structures. In distilled water normal loop matrix dis-solves almost instantaneously, and within a few minutes of isolation onlychromosome axes, still structurally continuous but of very low optical con-trast, remain. Chromomeres swell but remain recognizable, and chromosomeaxes are still unbroken more than 24 h after isolation.

188 Macgregor and Callan—Actions of enzymes
Trypsin
The optimum pH for tryptic digestion is between 7-5 and 8-5. We haveworked with trypsin initially dissolved in ordinary C medium at 0-5 mg/ml,the pH remaining at 6-2; and dissolved at a like concentration in modified Cmedium, with pH adjusted to 8-05 by replacement of the KH2PO4 fractionwith KH2PO4/Na2HPO4 buffer at o-ooi M. After application to a preparationof chromosomes in ordinary C medium, both solutions give an effectiveenzyme concentration of 0-25 mg/ml, the former at pH 6-2, the latter atP H 7 - 8 .
At pH 6-2 the first sign of tryptic action is a breakdown of the matrix ofnormal and granular loops, a process which we have been able to follow indetail on the giant granular loop of chromosome XII of cristatus. During thefirst 3 min of enzyme action the matrix of this loop is progressively dissolvedand the granules released and scattered by Brownian motion. Solution ofmatrix and release of granules starts at the thick end of the loop and progressestowards the thin end, until an extremely slender but unbroken loop axis,surrounded by a mass of rapidly dispersing granules, is all that remains of theoriginal structure. The fate of individual granules is uncertain; we havewatched them for periods of up to 2 h after detachment and although theyare reduced in size, they are not totally destroyed. All dense landmarkssimilarly undergo a slow reduction in size and are rarely completely dissolved.As a rule, no fusions of any sort are broken by the prolonged action of trypsinat pH 6-2. On four occasions, however, we have witnessed the complete andrapid destruction of the giant loops of chromosome XI of carnifex (fig. 3, A-D).On one of these occasions all 4 giant loops of one bivalent were fused togetherprior to application of the enzyme. Trypsin totally dissolved the materialinvolved in this fusion and the 2 chromosome arms drifted apart where theyhad originally been joined; only shrunken loop axes remained to mark thesites of the original loops.
After about 30 min of tryptic digestion at pH 6-2 the chromosomes' axeslose some optical contrast. The chromomeres swell and coalesce, givingto the axis the appearance of a more or less uniform cylinder. This effect isconsidered in more detail in the chapter on pan-protease.
The effect of trypsin at pH 7-8 is similar to its action at pH 6-2, but morerapid and more drastic. Application of trypsin solution is followed in a matterof seconds by disruption of the matrix of normal and granular loops, accom-panied by an equally rapid disappearance of all granules. For a time loopaxes remain visible. Dense landmark structures are slowly reduced in sizebut are not completely dissolved (figs. 3, E-K; 4, A-D). After an hour thechromosomes are almost invisible (figs. 3, j ; 4, D) but their linear continuitycan be demonstrated by adding a drop of 4% formaldehyde to the preparation,so increasing their refractility (fig. 3, K). We have not observed breakage of anysort of fusion by the action of trypsin at pH 7-8.
Roth (i960), in a study of the interaction of proteolytic enzymes and

on lampbrush chromosomes 189
ribonucleoprotein, found that all samples of trypsin which he tested werecontaminated with RNase. The action of RNase on lampbrush chromosomescan be distinguished from the action of trypsin and since RNase, unliketrypsin, retains its activity after boiling, we were able to test our sample oftrypsin for RNase contamination. C medium containing trypsin at 0-5 mg/mlwas boiled for 15 min, allowed to cool, and its pH checked (it remained at6-2), and was then applied to preparations of lampbrush chromosomes. Thetest gave unequivocal evidence of RNase contamination, which we judge tobe of the order 1 to 5% by comparison with the rates of action of knownconcentrations of RNase on similar material.
Pan-protease
This enzyme preparation is listed in pre-1959 Worthington catalogues as'crude protease'. More recently it has been given the name 'pan-protease'and has been assayed to contain 4,000 chymotryptic units and 7,000 trypticunits per milligram (using benzoyl-L-arginine-ethyl ester as substrate). Aswith trypsin, we have studied the action of pan-protease at an effective con-centration of 0-25 mg/ml, dissolved in ordinary C medium at pH 6-2 and inmodified C medium to give a pH of 7-8.
At pH 6-2 the action of pan-protease on lampbrush chromosomes is justlike that of trypsin. During the first 10 min of enzyme action the matrix ofnormal and granular loops is dissolved, loop granules are scattered by Brown-ian motion, whilst dense landmark structures are slowly reduced in size.Each chromosome takes on a 'fuzzy' appearance. After about 50 min thechromomeres start to lose refractility; they swell and coalesce with a con-sequent loss of the normal chromomeric pattern.
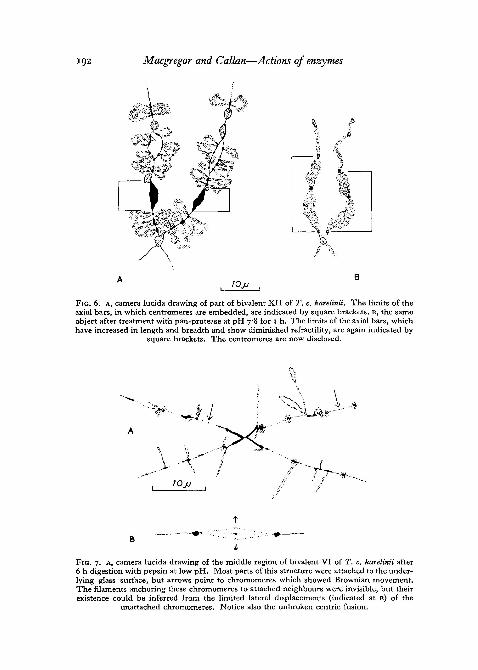
Pan-protease works more rapidly at pH 7-8 and after 1 h lampbrush chromo-somes reach a stable, almost invisible state. Their chromomeres hydrate, likethose of salivary gland chromosomes digested by trypsin in the presence ofelectrolytes (Kaufmann 1952), and as though in life they owe their compact-ness to some bonding material which is removed by pan-protease. The axialbars flanking the centromeres of karelinii lampbrush chromosomes provideconvenient material for studying this process, and fig. 4, E-K show the mainfeatures of pan-protease action at pH 7-8 on the centric regions of bivalentXII. When all but the last two photographs of this series were taken,the chromosomes were floating free in the medium and were showingBrownian movement. During pan-protease digestion the axial bars showsome elongation as well as transverse swelling (figs. 4, G, H; 6, A, B),whilst the centromere granules remain relatively unaffected. In the laterstages of digestion all that remains of the axial bars is a fine fibrousmaterial, which nevertheless still maintains its lengthwise coherence (fig.4> j)- When fixed in formaldehyde vapour after 45 min of pan-proteasedigestion (fig. 4, K) this fibrous material and the included centromere granulegave on testing a faint but positive Feulgen reaction. Centromere fusions inkarelinii persist even after prolonged treatment with pan-protease. Fusions

190 Macgregor and Callan—Actions of enzymes
between giant loops, spheres, axial granules, and telomeres also persist,though these bodies may be reduced to less than 5% of their original volumes.No interchromomeric breaks are produced by pan-protease. Moreover, wehave noted that in chromosome regions which were accidentally stretchedduring preparation, and which remained under tension due to occasionalattachments to the glass surface below, this enzyme failed to weaken intra-chromomeric bonding, the mechanical disruption of which normally leadsto the production of 'double bridges' (Callan, 1955). As with trypsin, testswith boiled pan-protease showed that this enzyme preparation is con-taminated with RNase.
Pepsin
The optimum pH for peptic action is between 1-5 and 2'5. Our test solu-tions were made up to give an effective enzyme concentration of 0-25 mg/mlin ordinary C medium at pH 6-2 and in modified C medium adjusted to givea pH of i-8.
The action of pepsin at pH 6-2 on lampbrush chromosomes is mild incomparison with that of trypsin or pan-protease. A rapid disintegration ofnormal and granular loop matrix, with dispersal of loop granules by Brownianmotion, is followed by a slow reduction in size of dense landmarks. At thispH pepsin appears to have no effect upon chromomeres or on the axial barsof karelinii.
Our observations on the effects of pH variation alone need to be borne inmind whilst assessing the action of pepsin at pH i-8. To recapitulate, a dropin pH much below 6-2 produces an initial contraction and sharp increase inthe optical contrast of lampbrush chromosomes followed, at pH 2, by anextension and loss of refractility. The latter state persists provided the pHremains low. For convenience we have used the word 'coagulated' to describethe contracted, refractile state and the word 'relaxed' to describe the state ofextension with low optical contrast. The sequence of events accompanyinga drop in pH from 6-2 to 2 is different if pepsin is present. The relaxedcondition sets in as expected, but it lasts for only a few seconds. Thereafter
FIG. 4 (plate), A, left end of bivalent V of T. c. carnifex. The spheres at the proximal sitesare separate from one another, whilst the distal sites together bear a single fused sphere(arrow). In medium C at pH 6-2.
D, same as A, 5 min after addition of trypsin at effective concentration of 0̂ 25 mg/ml andpH 7-8.
C, same as B, 10 min later.D, same as c, 45 min later.E, part of bivalent XII of T. c. karelinii. The arrow points to a centromere, which is flanked
by axial bars. The other centric region lies to the right, out of focus. In medium C at pH 62.F, same as E, 10 min after addition of pan-protease at effective concentration of 0-25 mg/ml
and pH 78.G, same as F, 5 min later.H, same as G, 15 min later.j , same as H, 10 min later. The arrow points to a centromere.K, same as J, fixed by adding 4% formaldehyde to the preparation after 45 min of digestion
by pan-protease.

FIG. 4H. C. MACGREGOR and H. G. CALLAN
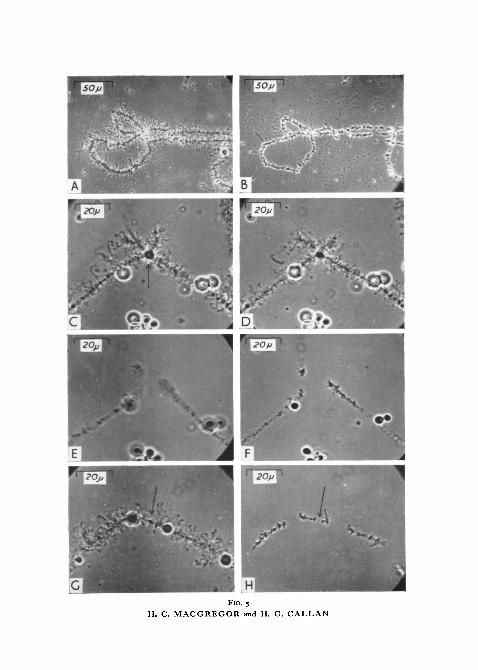
FIG. S
H. C. MACGREGOR and H. G. CALLAN

on lampbrush chromosomes 191
the chromosomes contract again, stiffen, stick to the glass bottom of thechamber, and become highly refractile. Peptic digestion begins at this pointand all structures attached laterally to lampbrush chromosomes are slowlyreduced in volume (fig. 5, A, B). After 6 h of peptic digestion at pH i-8 theremay be long stretches of chromosome consisting only of chromomeres anda tenuous interchromomeric thread. The thread is not always visible butits presence can be inferred when two chromomeres are attached to the glassand have one or more unattached chromomeres between them. The un-attached chromomeres show Brownian motion but remain anchored to theirneighbours (fig. 7).
Under these experimental conditions our primary aim is being frustrated, andwe are forced to witness the action of pepsin on coagulated chromosomes. Inan attempt to circumvent this difficulty we have adopted a two-stage approach.Pepsin in C medium at pH 6*2 is allowed to act for 10 min on a preparation oflampbrush chromosomes, and only thereafter is pH dropped to i-8. Duringa relaxed phase, which lasts for about 5 sec after the drop in pH, there isa spectacular solution of gene products. Spheres, axial granules, and thematrices of all lateral loops are stripped from the chromosomes. Yet in spiteof the violence of peptic digestion when allowed to proceed in this way, theaxes of lampbrush chromosomes remain unbroken. Moreover, intrachromo-meric breaks are not produced.
Under these conditions the fate of fusions between lampbrush chromo-somes is of particular interest. Sphere fusions on chromosomes V and VIIIof carnifex are always broken (fig. 5, C-F), each site where a sphere wasoriginally located being later recognizable as a gap in the chromosome axistraversed by a thin fibre (fig. 5, H).
Fusions between axial granules fall into two categories which can oftenbe discriminated in fresh undigested preparations. In one type (fig. 8, A) thetwo chromosomes are associated by a granule which they share in common,
FIG. 5 (plate), A, part of bivalent VII of T. c. carnifex. In medium C at pH 6-2.D, same as A, 10 min after addition of pepsin at effective concentration of 025 mg/mlandpH
1'8. The irregular particles scattered over the field are of precipitated nuclear sap.C, left end of bivalent VIII of T. c. carnifex. The arrow points to a single fused sphere; the
other round objects are 'nucleoli'. In medium C at pH 6-2.D, same as c, 10 min after addition of pepsin at effective concentration of 0-25 mg/ml and
pH 62.E, same as D, £ min later and after lowering pH to 1 -8. The sphere has dissolved, allowing
the chromosome arms to drift apart during the very brief 'relaxed' phase.F, same as E, 10 min later. The chromosomes are now 'coagulated' and stuck to the glass
surface below.G, left end of bivalent V of T. c. carnifex. The arrow points to fused telomeres; none of
the spheres are fused, and thus 4 are visible. In medium C at pH 62.H, same as G, 20 min after addition of pepsin at effective concentration of C25 mg/ml and
pH 6-2, and 10 min after lowering pH to i*8. The arrow points to an apparent gap in thechromosomes' axes where telomere material has dissolved away, but the chromosomes arestill terminally united and a tiny particle lies at their junction. The other apparent gaps inthe chromosomes' axes are the sites of sphere production, the spheres having dissolved. Eachof these gaps is traversed by an unbroken fibril.
192 Macgregor and Callan—Actions of enzymes
1OjJ
FIG. 6. A, camera lucida drawing of part of bivalent XII of T. c. karelinii. The limits of theaxial bars, in which centromeres are embedded, are indicated by square brackets. B, the sameobject after treatment with pan-protease at pH 7-8 for 1 h. The limits of the axial bars, whichhave increased in length and breadth and show diminished refractility, are again indicated by
square brackets. The centromeres are now disclosed.
FIG. 7. A, camera lucida drawing of the middle region of bivalent VI of T. c. karelinii after6 h digestion with pepsin at low pH. Most parts of this structure were attached to the under-lying glass surface, but arrows point to chromomeres which showed Brownian movement.The filaments anchoring these chromomeres to attached neighbours were invisible, but theirexistence could be inferred from the limited lateral displacements (indicated at B) of the
unattached chromomeres. Notice also the unbroken centric fusion.

on lampbrush chromosomes 193
but their axes are not otherwise in contact. Such fusions are disrupted bypepsin at low pH. In the other type of fusion (fig. 8, B, C) the 2 chromosomesmay or may not share a common axial granule but their axes are intimatelyassociated. These latter are not disrupted by pepsin even though the granulesare dissolved: in all probability chiasmata are involved in such fusions.
, 20JJ |
FIG. 8. Camera lucida drawings of parts of the shorter (right) arms of bivalentsVIII of T. c. carnifex extending from the centromeres, which are marked byarrows. A, shows an axial granule fusion of the kind which can be disrupted by
pepsin at low pH. B and c show fusions which cannot be so disrupted.
Telomere fusions are not disrupted by peptic digestion, whether theassociated telomeres be homologous or not. This is a rather surprisingobservation since in fresh undigested preparations it is often possible to see2 terminal chromomeres apparently separate from one another but bothembedded in a round mass of telomere material. The telomere material isdissolved by pepsin, but a thin strand joining the terminal chromomeres andbearing a tiny thickening midway along its length is left exposed (fig. 5, H).
Centromere fusions in karelinii are not broken by pepsin. In the case ofbivalents from relatively large oocytes the fused centromere granules arethemselves large and are usually so embedded in axial bar material that thewhole association is a stiff and compact structure. It remains so throughoutthe duration of peptic digestion. In the case of bivalents from small oocytes,where constrictions generally separate the axial bars from the centromeregranule, the constrictions may lengthen during peptic digestion (fig. 7, A),yet they do not break. When the medium containing such a bivalent is

194 Macgregor and Callan—Actions of enzymes
disturbed, all 4 chromosome arms may move relatively to one another but theyremain firmly attached at the fused centromeres.
Reflected fusions, such as occur very frequently on chromosomes III, IV,and VI of carnifex, likewise resist peptic digestion. This is another surprisingobservation, since such fusions are necessarily between genetically non-homologous structures and thus presumably of gene product rather than ofprimary genetic material.
The role played by pepsin, when this enzyme is applied before lowering pH,can be gauged by comparing its action with that of trypsin similarly applied.After tryptic digestion for a short time at pH 6-2, lowering of pH to 2 likewiseresults in stripping of the matrices of all lateral loops and the breakage of spherefusions, but the chromosomes remain relaxed. It is unlikely that trypsin hasmuch enzymatic activity at pH 2, hence we must assume that the effect oflowering pH is direct, i.e. that both peptic and tryptic digestion of lampbrushchromosomes at pH 6-2 renders loop matrices and other gene products acid-soluble.
Unlike trypsin and pan-protease, boiled pepsin proved to be inactive, andhence we can infer that our sample was not contaminated with RNase.
Ribonuclease
The optimum pH for RNase action is 7-7 (Kunitz, 1940), but as spontaneousdegradation of the enzyme takes place at this pH, and as RNase is also activeat pH 6-2, we have used this enzyme in ordinary C medium at an effectiveconcentration of 0-25 mg/ml.
The first observable effect of RNase at pH 6-2 on the giant granular loopof chromosome XII of cristatus is similar to that of trypsin, pepsin, or pan-protease at the same pH. Fig. 9, A-D are part of a photographic record toshow the first 7 min of RNase action on this loop. The series would equallyserve to demonstrate the early action of a proteolytic enzyme on this structure.Fig. 9, B was taken 3 min after the application of RNase solution. The enzymehas begun to act upon the loop matrix and 2 granules, attached to one anotherby a thin fibril, have come adrift. The loop axis can be seen at the thick end ofone loop. In fig. 9, c, taken 2 min later, more loop matrix has gone intosolution, granules are being scattered by Brownian motion, and the loop axis
FIG. 9 (plate), A, end of left arm of chromosome XII of T. c. cristatus, including the giantgranular loops. In medium C at pH 6-2.
B, same as A, 3 min after addition of RNase at effective concentration of C25 mg/ml andpH 62. The arrow points to 2 granules which have become detached.
c, same as D, 2 min later.D, same as c, 2 min later. The arrow points to loop axis exposed as the loop granules have
scattered.E, part of bivalent XI of T. c. carnifex, including 2 homologous pairs of giant loops. In
medium C at pH 6-2.F, same as E, 30 min after addition of RNase at effective concentration of C25 mg/ml
and pH 62.All these photographs are at uniform magnification.

FIG. 9H. C. MACGREGOR and H. G. CALLAN
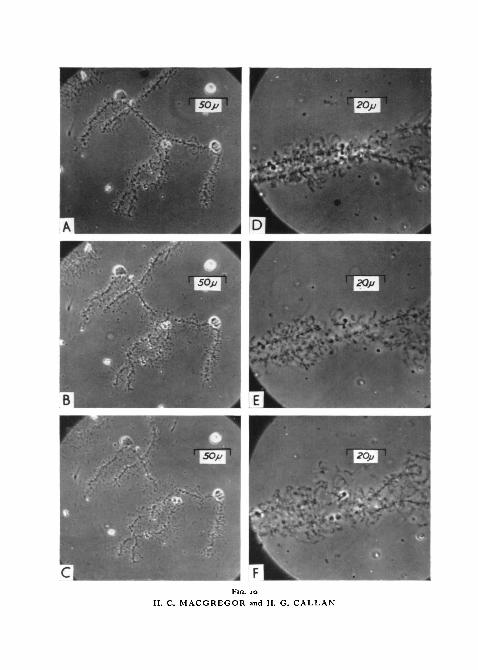
FIG. IO
H. C. MACGREGOR and H. G. CALLAN

on lampbrush chromosomes 195
is clearly visible. Fig. 9, D was taken 7 min after application of RNase. Theloop axis has been stripped of its matrix and granules for over half its length.Stripping progresses towards the thin end of the loop, leaving the axis bareand limp, but unbroken. In the long run, RNase seems to remove less materialfrom normal loops and from landmarks than does trypsin, pepsin, or pan-protease. The giant loops of chromosomes X, XI, and XII of carnifex firstbecome increasingly vacuolated; later they slowly shrink to about half theiroriginal volumes (fig. 9, E, F). AS normal loop matrices are dissolved byRNase, loop axes are exposed; these contract and increase in refractility.There is also some contraction of the main axes of chromosomes, but no lossof chromomeric pattern and no disruption of fusions even if pH is sub-sequently lowered to 2. The linear continuity of lampbrush chromosomesremains undisturbed by RNase, and is equally maintained if tryptic digestionfollows RNase.
The rate of RNase action on lampbrush chromosomes appears roughlyproportional to the concentration at which the enzyme is used. The finalstate reached by the chromosomes is the same over a range of RNase con-centrations from 0-25 mg to 0-000025 rng/ml. Some evidence of RNase actioncan be seen even at a concentration as low as io~6 mg/ml.
In accordance with expectation, tests made with RNase which had beenboiled for 15 min showed that enzymatic activity is retained.
Deoxyribonuclease
In the preceding chapters we have described how trypsin, pan-protease,pepsin, and ribonuclease affect lateral loop matrices, spheres, and other land-marks. We have also demonstrated that none of these enzymes break the axesof lampbrush chromosomes, no matter how the enzymes are applied. Takenby itself, this is evidence that the linear continuity of lampbrush chromosomesdoes not depend either on protein, or RNA, or on a combination of thesesubstances. The other known constituent of lampbrush chromosomes isDNA, and we will now present the complementary evidence which demon-strates that it is this substance which holds lampbrush chromosomes together.
DNase was used at effective concentrations of 0-125 o r 0-0125 mg/ml dis-solved in C medium at pH 6-2. In some experiments MgSO4 was dissolvedin the enzyme solution at 0-005% to function as activator, but its presencemade no significant difference to the results. DNase was used at lower con-centrations than the other enzymes in order to slow down its action and thusallow its dramatic effects to be recorded photographically.
FIG. 10 (plate), A, bivalent X of T. c. carnifex. In medium C at pH 6-2.B, same as A, i min after addition of DNase at effective concentration of 0-015 rng/ml and
pH 6-2.c, same as B, 3 min later.D, middle region of bivalent VII of T. c. carnifex. In medium C at pH 6-2.E, same as D, 1 min after addition of DNase at effective concentration of 0-015 mg/ml and
pH 6-2.F, same as E, 3 min later.

196 Macgregor and Callan—Actions of enzymes
A few minutes after application of DNase solution lampbrush chromosomesbreak at several places along their axes, each break occurring across the slenderfibril connecting adjacent chromomeres (fig. 10, E). The first breaks generallyoccur in accidentally stretched regions. As time passes more and more axialbreaks occur, but soon breaks in the lateral loops also become evident. Lateralloops are rapidly broken into smaller and smaller fragments, each fragmentretaining, for a time at least, its original fine morphology (fig. 10, B, c, F).Brownian motion so scatters the products of this continuing disruption thatafter 15 to 20 min the chromosome group is no longer recognizable.
For purposes of comparison we have followed the action of DNase on thegiant granular loop of chromosome XII of cristatus. Besides breaking theaxis of this loop, DNase mimics the action of ribonuclease or a proteolyticenzyme in that the matrix of the loop is dissolved and the granules released.This led us to wonder whether our sample of DNase was perhaps contamin-ated with other enzymes. DNase solution after being boiled for 15 min provedentirely inactive, hence we can rule out the possibility of RNase contamina-tion. We cannot absolutely rule out the possibility of contamination bya proteolytic enzyme, hence the 'stripping' action of our sample of DNase onthe giant granular loop of chromosome XII of cristatus remains an incon-clusive observation.
The crucial result of our tests with DNase, however, is the discovery thatthis enzyme breaks the main axes and the lateral loops of lampbrush chromo-somes. Possible contamination with proteolytic enzymes is here inconsequen-tial, since we have already shown that proteolytic enzymes are unable toproduce such breaks.
Versene
In a paper published in 1954, Mazia described the results of certain experi-ments on grasshopper mitotic and meiotic chromosomes and on salivary glandchromosomes of Drosophila, from which he concluded that these chromo-somes may consist of particulate 'macromolecular complexes of nucleic acidand proteins . . . linked together by bridges of divalent ions'. The basis forMazia's claim was that in material frozen by solid CO2, extracted in distilledwater, and then either treated with sodium citrate followed by furtherextraction in distilled water or treated with the chelating agent versene, thechromosomes were totally dissolved or at least partly fragmented. In Mazia'sopinion 'chromosomes may be dispersed without splitting peptide bonds iftwo conditions are met: (1) treatment with an agent capable of binding Ca orMg ions, and (2) provision of a medium of sufficiently low ionic strength'.In view of Mazia's findings we decided to extend the scope of our researchand study whatever action versene might have on lampbrush chromosomes.
First, a 0-005 M solution of versene was prepared in ordinary C medium,and pH readjusted to 6-2 by addition of a trace of NaOH. When this solutionwas applied to a fresh preparation of carnifex lampbrush chromosomes lying inC medium it had no discernible effect.

on lampbrush chromosomes 197
Second, carnifex oocyte nuclei were isolated in 5:1 K/NaCl at o-1 M, trans-ferred to observation chambers containing 0*005 M versene in C mediumat pH 6-2, and their membranes removed. The nuclear sap failed to disperse,but the chromosomes appeared lifelike. Identical results obtained whenversene at pH 6-7 was used.
Third, oocyte nuclei were isolated in 5:1 K/NaCl at o-1 M, rinsed indistilled water, transferred to observation chambers containing distilledwater, and their membranes removed. After 15 min the chromosomes werelocated and kept under observation whilst a o-ooi M solution of versenein distilled water (with pH adjusted to 7 by addition of a trace of NaOH)was added. A slight increase in the refractility of the chromosomesresulted, but their state was otherwise unaltered. The conditions of thisexperiment closely approach those used by Mazia—except for the omissionof freezing with solid CO2—and we observed no breakage of chromosomeswhatsoever.
Fourth, lampbrush chromosomes were isolated in C medium, and trypsin,at an initial concentration of 0-5 mg/ml in C medium adjusted to pH 8-05, wasadded. The chromosomes were kept under observation until they neared thelimits of visibility, and a 0-005 M solution of versene in C medium adjustedto pH 7-8 was then added. No change in the appearance of the chromosomesresulted. After 3 h the preparations were fixed with formaldehyde vapour;near the limits of visibility unbroken chromosomes could be seen.
Fifth, the above experiment was repeated after pretreatment for 30 minwith ribonuclease at 0-25 mg/ml in ordinary C medium. The result was thesame: unbroken chromosomes.
In passing, we may mention the result of one further experiment. To freshpreparations of lampbrush chromosomes lying in C medium, C mediumcontaining CaCl2 at o-oi M was added. The chromosomes became stiff,contracted, and highly refractile, showing their typical reaction to calciumions as first recorded by Duryee (1941). After 15 min a o-oi M solution ofversene in C medium at pH 6-2 was added in excess. There was no resultantchange in the appearance of the chromosomes.
DiscussionJorgensen (1913) was the first person to study the action of an enzyme on
giant chromosomes. In a long paper mostly concerned with the growth ofoocytes, Jorgensen recorded that whereas 'basichromatischen Chromosome'(chromosomes during mitosis) withstand peptic digestion, 'oxychromatischenChromosome' (lampbrush chromosomes) are rapidly and entirely dissolved.Jorgensen worked with alcohol-fixed sections of oocytes from various animals,including Salamandra maculosa, and concluded from his experience thatchromosomes during oocyte growth lack nucleic acids. More than 20 yearselapsed before giant chromosomes were again tested with enzymes, by Cas-persson (1936) working with dipteran salivary gland material. Casperssonsought for chemical corroboration of the inferences which he had drawn from

198 Macgregor and Callan—Actions of enzymes
studies of ultra violet light absorption. He squashed Drosophila salivaryglands in 45 % acetic acid, separated coverslip from slide, and fixed in Carnoy'sfluid, then exposed the squashed material to tryptic digestion. Casperssonfound that trypsin caused the stainable cross-bands to fall apart from oneanother; though they became greatly swollen they did not completely dis-solve.
Working with Chironomus Caspersson further observed that the structuraldegradation produced by trypsin could be largely inhibited if lanthanum ionswere present in the enzyme solution, an action which he ascribed to preci-pitation of nucleic acid as this substance became disengaged from protein.Caspersson concluded from these observations that salivary gland chromo-somes have throughout a structural framework of protein, with which nucleicacid is combined in the stainable cross-bands.
In 1939 Mazia and Jaeger described the actions of pepsin, trypsin, andnucleases on unfixed salivary gland chromosomes of Drosophila. They con-firmed Caspersson's observations that trypsin (in 0-05 M K2HPO4) dis-integrates salivary gland chromosomes, they found that pepsin reduceschromosome volume without causing disintegration, and that crude beefspleen nuclease digests away the Feulgen-staining material from cross-bands,again without disintegrating the chromosomes. Two years later Mazia (1941)recorded that, since even in moderately alkaline solutions, without enzymes,salivary gland chromosomes disperse into a fibrous mass, he had repeated hisexperiment with trypsin, this time at pH 5. Even at this pH, far from itsoptimum, trypsin caused the chromosomes to disintegrate. Mazia workedwith the salivary glands of Sciara, Drosophila, and Chironomus, and also withartificial fibres prepared from thymus nuclei. He again found that pepsinreduced salivary gland chromosome volume without destroying continuity,that pepsin failed to digest these chromosomes, and that crude protaminasefrom pancreas removed Feulgen-staining material from the cross-bands butleft the chromosomes otherwise intact. From these observations Maziaconcluded that, since trypsin splits peptide links near free basic groups(and hence would be effective against histone), whereas pepsin splits peptidelinks near free acidic groups (and hence would be ineffective against histone),and since trypsin disintegrates salivary gland chromosomes, whereas pepsindoes not, chromosomes must be held together lengthwise by histone. Indeed,Mazia contrasts 'matrix' non-histone protein with 'skeletal' histone whendiscussing salivary gland chromosome structure.
In 1944 Frolova, working with alcohol-fixed sections and aceto-carminesquash preparations of Chironomus salivary glands, confirmed that beef spleennuclease removed Feulgen-stainable material from cross-bands withoutcausing other structural degradation, but she further found that subsequenttreatment of nuclease-digested chromosomes with pepsin caused total dis-integration. Frolova also reported that RNase caused slight shrinkage ofsalivary gland chromosomes, but no other noticeable change in their appear-

Macgregor and Callan—Enzymes on lampbrush chromosomes 199
Three years later two further papers were published on this subject. Mazia,Hayashi, and Yudowitch (1947) reported once again the ability of trypsin todisintegrate salivary gland chromosomes, contrasting with the inability ofpepsin and cathepsin to act likewise. Catcheside and Holmes (1947), in apaper mainly concerned with the actions of various nuclease preparations,confirmed that trypsin destroyed salivary gland chromosomes of Drosophilawhich had been fixed in 45% acetic acid; they were unable to work withunfixed material, since the unfixed chromosomes 'lysed and lost their struc-ture' independently of enzyme treatment. They further reported that thymo-nuclease (DNase) prepared from pancreas removed Feulgen-stainablematerial from the cross-bands, but interpreted this result with caution becausethey found that several other preparations, inactive on their own, were alsocapable of removing Feulgen-stainable material if the chromosomes werefirst exposed to RNase. Catcheside and Holmes confirmed Frolova's obser-vation that RNase acting alone caused slight shrinkage of salivary glandchromosomes but no other recognizable change.
This generally consistent story was challenged by Kaufmann and variouscollaborators in a series of papers, the first of which appeared in 1946. Kauf-mann, Gay, and McDonald (1950) demonstrated that the apparent totaldigestion of onion root-tip and salivary gland chromosomes by trypsin dependson the presence of electrolytes in the enzyme solution. Salivary gland chromo-somes of Chironomus were fixed and squashed in 45% acetic acid, thendigested with salt-free trypsin solution. When examined by phase contrast,Kaufmann, Gay, and McDonald found that: ' Such treatment did not lead toany perceptible distortion of the pattern of banding of the chromosomes.When phosphate buffer replaced the trypsin solution, there was an immediateswelling of the chromosomes and lateral separation of the bands into com-ponent chromomeres.' Kaufmann, Gay, and McDonald stated further that:'The dissolution usually attributed to the specific action of trypsin has beenshown to be due to the removal by water of degradation products producedby the action of trypsin in combination with electrolytes.' Having observedthat chromosomes are destroyed by successive treatments with proteases andnuclease, or with proteases and hot trichloracetic acid, but not by either ofthese agents when used alone, Kaufmann, Gay, and McDonald concludedthat: ' . . . the chromosome represents an integrated fabric, in which no singleprotein or nucleic acid may be regarded as the primary structural component.'In later papers Kaufmann (1952, 1953), Kaufmann, Pennoyer, and Rowan(1953), and Kaufmann and McDonald (1956), the results of further ex-periments with enzymes are claimed to support this conclusion.
While these studies on salivary gland chromosomes were in progress,other cytologists had been experimenting with lampbrush chromosomes. Ina paper mainly concerned with the oocyte 'nucleoli' of Rana spp. Gersch(1940) observed that pepsin failed to destroy the lengthwise integrity of unfixedlampbrush chromosomes. Gersch also experimented with trypsin, but hisobservations with this enzyme were inconclusive.

200 Macgregor and Callan—Enzymes on lampbrush chromosomes
Duryee (1941) worked with pepsin, trypsin, and a variety of electrolytes;he found that both these enzymes, electrolytes at pH greater than 8-4 andacids at concentrations above 07 M, all 'removed' the lateral loops fromlampbrush chromosomes of Rana and Triturus spp. Although Duryee doesnot make the point distinctly, it would appear that none of the reagentswhich he tested broke the axes of lampbrush chromosomes, though thechromosomes were found to dissolve in o-oi M NaOH.
In 1948 Dodson gave an account of his studies on the lampbrush chromo-somes of Amphiuma means and Squalus suckleyi. Dodson worked throughouton fixed and sectioned oocytes, exposing 10 fi sections to enzymes and stainingthereafter to assess what actions had occurred. He reported that 3 daysdigestion with thymonuclease (DNase, possibly contaminated with RNase),though producing Feulgen-negative preparations, left chromosomes whichcould be stained by iron haematoxylin. He found that pepsin caused shrinkageof the lateral loops, but that chromosomes' axes survived peptic digestion andcould later be stained by Feulgen's reagent. Trypsin, on the contrary, dis-solved the lampbrush chromosomes entirely. After 12 h digestion even ironhaematoxylin staining failed to reveal any residual chromosome material.Dodson reached a conclusion substantially in agreement with Mazia andJaeger. He stated: 'The structural skeleton of the (lampbrush) chromosomeappears to be histone, while the nucleic acids are attached to this skeletonthrough protamine.'
In 1954 Gall, working with T. viridescens, demonstrated conclusively thatalthough the lateral loops of unfixed lampbrush chromosomes are 'removed'by trypsin this enzyme fails to destroy the lengthwise coherence of chromo-some axes. Gall claimed that Dodson's contrary observations were probablydue to the thin chromosome axis being mechanically removed, 'washed out',from sections after their loops had been digested. Gall further reported thatpepsin at low pH 'removes' the lateral loops from both fixed and unfixedlampbrush chromosomes, but that pepsin, like trypsin, fails to destroy chromo-some axes.
Having observed the 'double loop bridges' which are regularly producedwhen lampbrush chromosomes are stretched and their chromomeres broken,Callan (1955) predicted that each lateral loop must have a fibrillar core, com-parable in elasticity and tensile strength to the fibre running between adjacentchromomeres in the chromosome axes, and around which the products ofsynthesis accumulate. Gall (1956) put this theory to test by fixing lampbrushchromosomes to a glass slide by formaldehyde vapour; he then digested suchpreparations with pepsin, made shadow-cast formvar replicas, and observedthe replicas with an electron microscope. Gall showed that there is indeed aloop axis which resists peptic digestion whilst the bulk of the loop materialdissolves, and that its diameter is roughly equal to that of the fibre runningbetween adjacent chromomeres. Gall's observations shed new light on thestructure of lampbrush chromosomes; he inferred that the loop axis fibril maybe continuous with the interchromomeric fibril, and that both may contain

Macgregor and Callan—Enzymes on lampbrush chromosomes 201
DNA. With this background of information and speculation the enzymestudies described in the present paper were begun.
We have found that proteolytic enzymes and RNase dissolve matrix fromlateral loops. Since we have not worked with an electron microscope we donot know whether loop axes withstand trypsin and pan-protease digestion,but they certainly resist RNase. Neither of these enzymes break the axes oflampbrush chromosomes, nor does versene. Our critical observation is thatDNase, on the contrary, does break chromosome axes. This enzyme breaksnot only the fibril between adjacent chromomeres; it also breaks lateral loops,and the appearance of the loop fragments so produced indicates that DNaseacts directly upon loop axis. By breaking lampbrush chromosomes in thismanner DNase accomplishes something which is uncommonly hard to do,even mechanically. We conclude that DNA, not protein, is responsible forthe linear integrity of lampbrush chromosomes.
We now have to face the problem of reconciling this conclusion with thecontrary conclusion of Kaufmann and collaborators drawn from studies onsalivary gland chromosomes.
The action of DNase on lampbrush chromosomes was briefly reported byCallan and Macgregor in 1958. Dutt and Kaufmann (1959) have sincedescribed the action of DNase on unfixed salivary gland chromosomes ofDrosophila and on testicular chromosomes of 2 grasshopper species, and theyhave criticized our 1958 conclusions. Dutt and Kaufmann have shown thatDNase may lead to loss of DNA from salivary gland chromosomes withoutcausing marked deformation, or to structural alteration of the chromosomeswithout complete removal of DNA. They found testicular chromosomesmore refractory than salivary gland chromosomes, and conclude: 'that DNasedoes not cause deformation of chromosomes by removal of DNA that isessential for the maintenance of structural integrity. Deformation is attri-butable to modification of the properties of the complex nucleoprotein fabricof the chromosome.'
This may indeed be the appropriate negative conclusion to draw fromobservations on salivary and testicular chromosomes, but we have drawn apositive conclusion from work on different material. Dutt and Kaufmannclaim that in our 1958 paper we: 'emphasize the difference in response offixed and unfixed chromosomes to enzymatic hydrolysis' as though this mightexplain our different results. At least for lampbrush chromosomes theemphasis is justified. If a lampbrush chromosome, fixed and attached to aglass slide, is digested by DNase, it appears unbroken when viewed with alight microscope. This observation does not signify that interchromomericfibrils or lateral loop axes are intact; all it signifies is that in lampbrushchromosomes there are constituents other than DNA remaining attached tothe slide. Dutt and Kaufmann further claim that: '. . . dissociation in itself isnot a critical index to the selective action of a given agent on a specific chromo-somal component. . . . A more reliable criterion of the digestion of chromo-somal DNA by DNase is the loss of Feulgen stainability, whether this is

202 Macgregor and Callan—Enzymes on lampbrush chromosomes
associated with structural deformation or not.' Yet fixed lampbrush chromo-somes can be digested with DNase until all their Feulgen stainability has dis-appeared ; thereafter they still appear unbroken. As we have already describedin this paper, unfixed lampbrush chromosomes behave otherwise. They arebroken by DNase, not by proteolytic enzymes, RNase or versene, and becausethey are not attached to a slide, Brownian movement reveals that they havebeen broken.
Dutt and Kaufmann suggest and prefer an alternative explanation:'... thatthe meiotic lampbrush chromosomes differ from the animal and plant chromo-somes with which we [Dutt and Kaufmann] have worked in regard to thenature and distribution of their essential structural components'. Whilst wewould agree that lampbrush chromosomes differ in many respects fromdipteran salivary gland and grasshopper testicular chromosomes, such dif-ferences are more likely to concern the arrangement than the nature of theprimary genetic material, and the extent to which gene product is associatedwith this material.
The main conclusion which we have drawn from our observations is thatone constituent of a lampbrush chromosome is an uninterrupted DNAfibre. In those parts of lampbrush chromosomes where the DNA concen-tration per unit of length is high, and where histone is also demonstrable, i.e.in the chromomeres and, more strikingly, in the axial bars near the centromeresof karelinii, DNase fails to produce breaks. We do not know how DNA andhistone are arranged in such regions, but trypsin and pan-protease digestionreveals that when histone is withdrawn the remaining DNA preserves itslengthwise coherence though it fails to maintain a compact form. Such partsof lampbrush chromosomes may well resemble in structural organization thecross-bands of salivary gland chromosomes and the compact chromosomesof metaphase, which from our experience with lampbrush chromosomes wewould expect to withstand DNase digestion. We might, on the other hand,expect DNase to break the interband regions of salivary gland chromosomes.However, since salivary gland chromosomes are polytene, clean breaks wouldonly be demonstrable if several fibres were all broken at the same interband,and provided the cross-bands on either side separated to reveal the break.In our opinion these are the main reasons why Kaufmann and his co-workershave reached conclusions different from our own.
We are indebted to Dr. A. Sengun for supplying newts from Turkey; toDr. G. A. Horridge for constructing a xenon flash apparatus; to Dr. D. Maziafor supplying pan-protease; and to Dr. O. L. Miller for sending us a sample ofNational Aniline fast green FCF. We also wish to thank Dr. G. R. Tristramand Dr. J. G. Gall for helpful advice on several occasions.
ReferencesALFERT, M., 1955. Acta Anat., 23, 185.
and GESCHWIND, I., 1953. Proc. nat. Acad. Sci. Wash., 39, 991.D'ANGELO, E. G., 1946. Biol. Bull. Wood's Hole, 90, 71.

Macgregor and Callan—Enzymes on lampbrush chromosomes 203
BLOCH, D. P., and GODMAN, G. C, 1955. J. biophys. biochem. Cytol., 1, 17.BRACHET, J., 1940. Arch. Bio). Paris, 51, 151.BROWN, G. L., CALLAN, H. G., and LEAF, G., 1950. Nature Lond., 165, 600.CALLAN, H. G., 1952. Symp. Soc. exp. Biol., 6, 243.
1955- Symposium on fine structure of cells. I.U.B.S. publ. series B, 21, 89.1957. Pubbl. Staz. zool. Napoli, 29, 329.and LLOYD, L., 1960a. Symposium on new approaches in cell biology, 23. London and
New York (Academic Press).19606. Philos. Trans., ser. B, 243, 135.
and MACGREGOR, H. C, 1958. Nature Lond., 181, 1479.CASPERSSON, T., 1936. Skand. Arch. Physiol., suppl. 8.CATCHESIDE, D. G., and HOLMES, B., 1947. Symp. Soc. exp. Biol., 1, 225.COLEMAN, L. C , 1938. Stain Tech., 13, 123.DODSON, E. O., 1948. Univ. Calif. Publ. Zool., 53, 281.DURYEE, W. R., 1941. University of Pennsylvania bicentennial conference on cytology, genetics
and evolution, 129. Philadelphia (Univ. Pennsylvania Press).195°- Ann. N.Y. Acad. Sci., 50, 920.and DOHERTY, J. K., 1954. Ibid., 58, 1210.
DUTT, M. K., and KAUFMANN, B. P., 1959. Nucleus, 2, 85.FELIX, K., 1952. Experientia, 8, 312.FRAZER, S. C, and DAVIDSON, J. N., 1953. Exp. Cell Res., 4, 316.FROLOVA, S. L., 1944. J. Hered., 35, 235.GALL, J. G., 1952. Exp. Cell Res., suppl. 2, 95.
1954. J. Morph., 94, 283.1956. Brookhaven Symp. Biol. 8, 17.
GERSCH, M., 1940. Z. Zellforsch., 30, 483.GUYENOT, E., and DANON, M., 1953. Rev. suisse Zool., 60, 1.JORGENSEN, M., 1913. Arch. Zellforsch., 10, i.KAUFMANN, B. P., 1952. Proc. nat. Acad. Sci. Wash., 38, 464.
1953- Exp. Cell Res., 4, 408.GAY, H., and MCDONALD, M. R., 1950. Cold Spr. Harb. Symp. quant. Biol., 14, 85.MCDONALD, M. R., and GAY, H., 1946. Carneg. Instn. Wash. Year Book, 45, 158.
1956. Cold Spr. Harb. Symp. quant. Biol., 21, 233.KVNITZ, M., 1940. J. gen. Physiol., 24, 15.
PENNOYER, J. M., and ROWAN, M. E., 1953. J. cell. comp. Physiol., 41, 79.LAMB, W. G. P., 1950. Exp. Cell Res., 1, 571.LEUCHTENBERGER, C., 1952. Ibid., 3, 240.MAZIA, D., 1941. Cold Spr. Harb. Symp. quant. Biol., 9, 40.
1954. Proc. nat. Acad. Sci. Wash., 40, 521.HAYASHI, T., and YUDOWITCH, K., 1947. Cold Spr. Harb. Symp. quant. Biol., 12, 122.and JAEGER, L., 1939. Proc. nat. Acad. Sci. Wash., 25, 456.
MIESCHER, F., 1871. In Hoppe-Seyler, F., Medizinisch-chemische Untersuchungen, 4, 441.Berlin.
MIRSKY, A. E., 1947. Cold Spr. Harb. Symp. quant. Biol., 12, 143.and Ris, H., 1947. J. gen. Physiol., 31, 1.
1951. Ibid., 34, 451.POLLI, E. E., 1951. Chromosoma, 4, 621.ROTH, J. S., i960. J. biophys. biochem. Cytol., 7, 443.SWIFT, H., 1950. Physiol. Zool., 23, 169.VENDRELY, R., and VENDRELY, C, 1948. Experientia, 4, 434.
I9S3. Nature Lond., 172, 30.





























