Embed Size (px)
Citation preview

Gene 521 (2013) 303–310
Contents lists available at SciVerse ScienceDirect
Gene
j ourna l homepage: www.e lsev ie r .com/ locate /gene
The alpaca agouti gene: Genomic locus, transcripts and causative mutations ofeumelanic and pheomelanic coat color
Bathrachalam Chandramohan, Carlo Renieri, Vincenzo La Manna, Antonietta La Terza ⁎School of Environmental Sciences, University of Camerino, Via Gentile III da Varano, 62032, Camerino, MC, Italy
Abbreviations: RNA, ribonucleic acid; DNA, deoxyriboRNA; bp, base pair; kb, kilobase; nt, nucleotide; cDNA, comerase chain reaction; RT, reverse transcription; BLASTTool; NCBI, National Center for Biotechnology Information⁎ Corresponding author. Tel.: +39 0737 403272; fax:
E-mail address: [email protected] (A. La T
0378-1119/$ – see front matter © 2013 Elsevier B.V. Allhttp://dx.doi.org/10.1016/j.gene.2013.03.060
a b s t r a c t
a r t i c l e i n f oArticle history:Accepted 16 March 2013Available online 1 April 2013
Keywords:PigmentationGene duplicationTranscriptionUTRBiological resourcesSkin
The agouti gene encodes the agouti signaling protein (ASIP) which regulates pheomelanin and eumelaninsynthesis in mammals. To investigate the role of agouti in coat color variation of alpaca, we characterizedthe agouti gene and identified three mutations potentially involved with the determinism of eumelanicand pheomelanic phenotypes. The exon-4 hosts the mutations g.3836C>T, g.3896G>A and g.3866_3923del57.Further analysis of these mutations revealed two genotypes for black animals. The reverse transcription analysisof mRNA purified from skin biopsies of alpaca revealed the presence of three transcripts with different 5′untranslated regions (UTRs) and color specific expression. The white specific transcript, possibly originatingfrom a duplication event (intra-chromosomal recombination) of the agouti gene is characterise by a 5′UTRcontaining 142 bp of the NCOA6 gene sequence. Furthermore, the relative level expression analysis of mRNAdemonstrates that the agouti gene has up-regulated expression in white skin, suggesting a pleiotropic effect ofagouti in the white phenotype. Our findings refine the structure of the agouti locus and transcripts and provideadditional information in order to understand the role of agouti in the pigmentation of alpaca.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
Pigmentation in mammals is known to be influenced by morethan 350 genes (Montoliu et al., 2011). Among these, agouti is awell studied gene which encodes for a small secreted factor (agoutisignaling protein) that normally functions as a paracrine regulatorof hair pigmentation in mice and other mammals (Lamoreux et al.,2010; Vage et al., 1997; Wang et al., 1998). Mutually exclusivebinding of themelanocortin-1-receptor (MC1R) by the agouti signalingprotein and the α-melanocyte stimulating hormone (α-MSH) signalsthe hair-bulb melanocytes to synthesize pheomelanin (yellow–redpigments) and eumelanin (dark pigments), respectively (Jackson,1994). In all the analyzed mammalian species the agouti transcriptsare approximately 800 bp long and encode a protein of 131–133amino acids. So far more than 50 agouti alleles have been identified inmice (Montoliu et al., 2011) and several studies have investigated themolecular genetics of agouti in other mammals and its associationwith pigmentation phenotypes. Recessive mutations at the agoutilocus, which impair either agouti protein activity or alter the level ofagouti mRNA synthesis, resulting in a darker coat color have beendocumented in many mammals, such as rodents (Kingsley et al.,
nucleic acid; mRNA, messengermplementary DNA; PCR, poly-, Basic Local Alignment Search.+39 0737 403290.erza).
rights reserved.
2009; Linnen et al., 2009; Steiner et al., 2007), horses (Rieder et al.,2001), cats (Eizirik et al., 2003) foxes (Vage et al., 1997), and rabbits(Fontanesi et al., 2010). Moreover, other agouti alleles such as Ay (lethalyellow), which displays constitutive and ubiquitous expression ofagouti, increases yellow pigmentation in mice (Miller et al., 1993).Apart from the yellow coat, mice that carry Ay display obesity, insulinresistance, premature infertility, increased body length and tumorsusceptibility (Duhl et al., 1994).
In alpaca (Vicugna pacos) a wide variety of colors exists and it istherefore preferred in the fiber industry for the manufacturing ofecologically sustainable and organic textile products. The Peruviansalways sought to protect the alpaca industry. Peru was the lastSouth American country to open its borders to exportation and onlyapproximately 3000 alpacas have actually left Peruvian soil. At themoment, Peru has four million alpacas representing approximately90% of the global population. Therefore, this South-American countryhosts the largest reserve of alpaca biological resources in the world.An estimated 20% of colored wool is presently produced in Peru andthe alpaca fiber exportation represents one of the major economiesfor Peru, thus there is an increasing interest in better understandingthe genetic basis of coat color in the Peruvian alpaca population.
So far, the main body of studies on alpaca has mainly regardedthe analysis of the inheritance of phenotypes (Gandarillas, 1971;Valbonesi et al., 2009, 2011; Velasco et al., 1978a, 1978b), the bio-chemical properties of melanin (Cecchi et al, 2011; Fan et al., 2010;Renieri et al., 1991, 1995), and the morphology of melanosomes(Cozzali et al., 1998, 2001), and only few studies have focused on

304 B. Chandramohan et al. / Gene 521 (2013) 303–310
the molecular characterization of candidate genes responsible forcoat colors, e.g. MC1R from American (Powell et al., 2008) andAustralian (Feeley and Munyard, 2009) alpaca populations. A prelim-inary characterization of full length agouti transcripts from skinbiopsies from an experimental trial for color segregation in Peruvianalpacas (Bathrachalam et al., 2012) and on the coding sequences ofthe same gene from genomic DNA isolated from blood samples fromrandom Australian alpaca populations have recently been carriedout (Feeley et al. 2011). Both studies have identified two missensemutations (g.3836C>T, g.3896G>A) and Feeley et al. (2011) alsoreported a 57 bp deletion (g.3866_3923del57), all located in exon 4of agouti gene which codes for the last 40 amino acid residues consti-tuting the Cys-rich C-terminal portion of ASIP, a domain of crucialimportance for the functionality of the protein itself (McNulty et al.,2005). All these are probable loss-of-function mutations and theyseem to be strongly associated with black color even if they neitherexplain all black phenotypes in alpaca nor all the phenotypic variationpossible in ASIP (Feeley et al., 2011). Many other factors individuallyor in combination such as, mutations in regulatory region, genomicreorganization events (duplications and/or inversion), effects dueto dilution genes (MATP, TYR, TYRP1) or contribution of MC1Rgenotypes (for black phenotypes) might be involved in the determi-nation of the large variety of coat color observed in alpaca.
Therefore, the present investigations have been conducted toelucidate the genetic mechanisms behind alpaca pigmentation witha special emphasis on the characterization of the genomic and tran-script structures, the relative levels of mRNA expression as well as,the description of mutations that probably affect eumelanin andpheomelanin synthesis from the skin biopsies of the native Peruvianalpaca population. These mutations have been analyzed in black,brown and white Peruvian alpacas involved in an experimental trialfor color segregation analysis as described by Valbonesi et al. (2011).
2. Materials and methods
2.1. Collection and storage of skin biopsies
Skin biopsies from white and colored (brown and black) alpacaswere collected by disposable biopsy punch (8 mm diameter) inRNAlater (SIGMA, Germany) from ILPA-Puno, Quimsachata Experi-mental Station, Instituto Nacional de Innovacion Agraria (INIA),Peru which is located at 4300 m above sea level. The alpacas analyzedin the present experiment were part of a previous phenotypic segre-gation study on coat color inheritance (Valbonesi et al., 2011).It should be noted that, due to the difficulties in sampling at thishigher altitude, the total number of skin biopsies was limited to 80samples. The biopsies were transferred to the School of Environmen-tal Sciences, University of Camerino, Italy. Subsequently, they wereremoved from RNAlater, blotted with sterile blotting paper andstored at −196 °C (liquid nitrogen) for further analysis. All experi-ments were approved and performed according to the guidelinesof the Animal Ethics Committee of the University of Camerino.
2.2. Nucleic acid extraction and cDNA synthesis
Total RNA from stored skin biopsies was extracted using anRNAeasy fibrous tissue mini kit (Qiagen S.A., Courtaboeuf, France).Simultaneously, genomic DNA was also isolated using a DNAeasytissue kit (Qiagen S.A., Courtaboeuf, France) according to themanufacturer's instruction. The isolated RNA and DNA samples werestored at −80 °C for further analysis. The first strand cDNA was syn-thesized with 2 μg of total RNA using 10 pmol OdTm primer (Supple-mentary Table 1 and Supplementary Fig. S1), 0.5 mM dNTPs (Roche),1× RT buffer, 20 U RNase inhibitor and 200 U PrimScript™ ReverseTranscriptase (Takara Biotech, Japan) in 20 μl total reaction volumeaccording to the manufacturer's instructions. The reaction mixture
was incubated for 45 min at 50 °C and then at 70 °C for 15 min andthe resulting cDNA was used in coding sequence and 3′ end amplifi-cation. All the reverse transcription reactions were carried out usinga Perkin-Elmer Thermal Cycler (Perkin-Elmer Corporation, Norwalk,CT, USA).
2.3. Primer design and PCR amplification of full length agouti transcriptsby rapid amplification of cDNA ends (RACE)
Orthologous sequences of the agouti gene from mammals wereretrieved from NCBI GenBank (http://www.ncbi.nlm.nih.gov/) andaligned with EMBL ClustalW (http://www.ebi.ac.uk//Tools/clustalw/)to identify conserved regions for the design of primers to amplify thecoding region. PCR amplification of complete coding sequence fromcDNA was done with the primers AF1/AR1 (Supplementary TableS1) in 50 μl reaction volume containing 1× Expand Long Range buffer(1.25 mM MgCl2), 0.3 mM dNTP, 0.3 μmol of each primer, and 3.5 UExpand Long Range enzymemix (Roche, Germany)with the followingcycling condition: initial denaturation at 95 °C for 3 min, followed by35 cycles of 95 °C for 30 s, 66 °C for 30 s and 72 °C for 25 s, with afinal extension at 72 °C for 7 min. The PCR products were clonedinto the pGEM-T easy vector system (Promega, USA) and sequencedat BMR Genomics, Italy. The coding sequence was determined andused to design gene specific primers AF2, AR2, and AR3 (Supplemen-tary Table S1 & Supplementary Fig. S1) for 3′ and 5′ RACE, a PCRapproach specifically devised to characterize untranslated regulativeregion of the transcripts. The 3′ RACEwas carried out with the primersAF2/nOdTm followed by amplification with AF1/OdTm primers(Supplementary Table S1 & Supplementary Fig. S1). Subsequently, 5′RACE was carried out according to the method reported in Shi andJarvis (2006). The first strand cDNAwas synthesized with gene specif-ic primer AR3, followed by amplification with UPML/AR1 primers andnested amplification was performed by using the UPMs/AR2 primerpair (Supplementary Table S1 & Supplementary Fig. S1). The ampli-fications were carried out as described above except for the annealingtemperature (Supplementary Table S1) and the PCR products werecloned into the pGEM-T easy vector system and sequenced at STARseq,Germany. At least 5 independent clones were sequenced for eachamplicon category.
2.4. Long range PCR
Expand Long-Template PCR (Roche Diagnostics, Switzerland) wasperformed to amplify the whole (ATG-TGA) alpaca agouti gene byusing the AF1/AR1 primers (Supplementary Table S1) according tothe manufacturer's instruction. Amplification of alpaca DNA withthe agouti primers produced a 3945 bp fragment; the product wascloned into the InsTAclone PCR Cloning Kit (Fermentas GMBH,Germany) and sequenced at STARseq, Germany.
2.5. Sequence analysis
Nucleic acid and protein database searches were performed usingBLAST at the NCBI server and visualized with Circoletto (Darzentas,2010). The cDNA and DNA sequence data were analyzed usingDNASTAR 5.0 software (Dayhoff et al., 1978). The alignment of theamino acid sequence of agouti proteins were performed usingClustalW (Thompson et al., 1994). The mRNA secondary structurepredictions were performed with m-fold (Zuker, 2003).
2.6. Semi-quantitative RT-PCR
Expression of the agouti gene in white and colored skin (brownand black) biopsies were analyzed on cDNA obtained as describedabove using AF3/AR4 primers (Supplementary Table 1). PCR wascarried out in a 25 μl of reaction volume containing 1× Dream Taq

305B. Chandramohan et al. / Gene 521 (2013) 303–310
buffer, 0.2 mM dNTPs, 10 pmol of each primer and 1.25 U Dream Taqpolymerase (Fermentas, GMBH) with the following cycling condition;95 °C for 3 min, 30 cycles at 95 °C for 30 s, 66 °C for 30 s and 72 °Cfor 15 s, followed by a 7 min incubation at 72 °C. A pair of primersGAPF/GAPR (Supplementary Table S1) was used to amplify thecDNA encoding Glyceraldehyde 3-phosphate dehydrogenase (GAPDH)as reference gene with the above mentioned PCR condition exceptfor the annealing temperature (Supplementary Table S1). The samevolumes of the PCR products were applied to 1.5% agarose gel, stainedwith ethidium bromide and evaluated by band densitometry withQscan 3.0. The relative level of gene expression was analyzed withone-way ANOVA (analysis of variance) using SPSS. Individual meancomparisons were performed using Duncan's test. Differences ofp b 0.05 were considered significant.
2.7. Genotyping
The two non synonymous SNPs (g.3836C>T & g.3896G>A)observed in the exon-4 were genotyped by direct sequencing andthe in frame 57 bp deletion (g.3866_3923del57) in the exon-4 wasanalyzed by direct electrophoresis on 1.8% agarose gel in 1× TAEbuffer, stained with ethidium bromide and visualized by a UV
Fig. 1. Agouti full length transcripts from the skin. A. A representative banding pattern of 5′Rblack alpacas. C— common transcript observed in all three phenotypes; A— black specific tratranscripts. The three different 5′UTR nucleotide sequences (transcripts-C, A and D) and thheadings. The uORF start codons (atg) in the 5′UTR sequences are underlined and namedthe same frames are colored identically. In all transcripts the main ORF is indicated in uppUTR sequence.
transilluminator, followed by the amplification of sequence spanningthe mutations with the primers designed on Exon-4 (AF4) and 3′UTR(AR4) region (Supplementary Table S1). The PCR reactions wereperformed with an initial denaturation at 95 °C for 3 min followedby 30 cycles of denaturation at 95 °C for 30 s, annealing at 65 °C for30 s, extension at 72 °C for 20 s and with a final extension at 72 °Cfor 5 min. The amplifications were carried out in 25 μl of reactionvolume containing 1× Dream Taq buffer, 0.2 mM dNTPs, 10 pmol ofeach primer and 1.25 U Dream Taq polymerase (Fermentas, GMBH).
2.8. In silico functional analysis and homology modeling of agouti
In silico functional analysis of missense mutations were obtainedusing PANTHER (Thomas et al., 2003) and SNAP tool (Brombergand Rost, 2007). Homology modeling was carried out using theSWISSMODEL (Kiefer et al., 2009). The original sequence withmutated structure was also submitted to the SWISSMODEL serverto observe the change caused by a single mutation. The torsionangles (psi, phi) between the original sequence and the mutatedsequence (R98C) were compared using the tool RamachandranPlot 2.0 (http://dicsoft1.physics.iisc.ernet.in/rp/).
ACE, coding sequence and 3′RACE products of agouti transcripts from white, brown andnscript; and D—white specific transcript. B. 5′and 3′UTR nucleotide sequences of agoutie common 3′UTR of agouti transcripts are presented separately under the appropriateimmediately above them. In transcript-C the various uORFs and STOP codons sharinger-case and bold letters. The poly-A signaling sequence is underlined in bold in the 3′

306 B. Chandramohan et al. / Gene 521 (2013) 303–310
3. Results
3.1. Characterization of the agouti full length transcript and genomiclocus
The combined transcript sequence initially obtained from totalRNA purified from lateral skin of brown alpaca is 822 bp long(excluding poly A tail). It is composed by a 402 bp open readingframe (ORF), a 196 bp 5′UTR and a 224 bp 3′UTR (Fig. 1). The ORF(GenBank accession no: FJ847229, FJ847230, FJ847232) is 88% and89% identical at the nucleotide level to the ovine and bovinesequences, respectively. It encodes a putative 133 amino acid protein,which is 83% and 85% identical to the ovine and bovine proteins,respectively. The coded amino acid sequence has the characteristicsignature for mammalian agouti protein (Bultman et al., 1992, 1994;Fontanesi et al., 2010; Girardot et al., 2005; Kerns et al., 2004; Milleret al., 1993; Wilson et al., 1995). A signal sequence of 22 aminoacids (Kyte and Doolittle, 1982) is followed by a basic amino-terminal domain and a cysteine-rich carboxy-terminal domain(Bultman et al., 1992), whose pattern of spacing is highly conservedamong vertebrates (Supplementary Fig. S2).
The amplification of genomic DNA using AF1/AR1 primersdisplayed a 3945 bp fragment (GenBank accession no: HQ645014),which contains three coding exons (2, 3 and 4), in addition to intronicsequences (Fig. 2A). The genomic organization of the alpaca agouticoding exons 2, 3 and 4 and introns 2 and 3 is similar to that reportedfor all other mammals (Bultman et al., 1992, 1994; Fontanesi et al.,2010; Girardot et al., 2005; Kerns et al., 2004; Miller et al., 1993;Wilson et al., 1995). The coding exons 2, 3 and 4 are separated by1262 and 2281 bp intronic sequences, respectively. Each exon isflanked by consensus splice donor and acceptor sites, with the excep-tion of exon-4, which shows only a splice acceptor site (Fig. 2A), aspreviously reported in the bovine agouti gene (Girardot et al., 2005).
Fig. 2. The structure of alpaca agouti genomic locus, polymorphisms and its transcripts. A. Tindicate protein coding sequences (exons 2, 3 and 4) and non-coding (1A, 1C, 2A, 3A and 3(TGA) are indicated by the green circle ( ) and red sparkle ( ) symbols, respectively; 5′splicthe gene structure indicate the length of exons and introns in bp. Polymorphisms identifiedagouti locus, deduced from this study. Non-agoutimutations identified in other mammalian sThe causative mutations involved in the determination of the black and brown phenotypespecific (D) alpaca agouti transcripts. Transcript C, A and D are represented under the corr3′UTR were deduced from this study. The locations of non coding exons (2A, 3A and 1C) werexon 1A is not known ( ).
3.2. Characterization of 5′UTR in black, brown and white alpaca
In mice and rabbit, the agouti gene produces four transcripts thatdiffer solely in their 5′UTRs (Bultman et al., 1992; Fontanesi et al.,2010; Miller et al., 1993). In order to assess such feature in alpaca,we performed 5′RACE experiments with total RNA isolated fromwhite and pigmented (brown and black) alpaca. We identified threedifferent 5′UTRs, 136, 142 and 196 bp long (Fig. 1A). Interestingly,among the identified transcripts, one is observed to be present inall three phenotypes (5′UTR of 196 bp; GenBank accession no:HQ645017) and found to have 85% identity with the bovine exon 1Ccounterpart, hence the name transcript C. Nucleotide analysis of the5′UTR revealed the presence of 4 upstream open reading frames(uORF), of which the first 3 appear to be in the same reading frameas the start codon of the agouti ORF, while codon 4 is not (Fig. 1B).Translation starting at these upstream start codons could potentiallyresult in 4 short peptides, which range in size from 14 to 27 aminoacids terminating before the main agouti ORF, none of which showedany identity with previously known proteins. The BLAST analysis ofcommon 5′UTR with 2× genome of alpaca (Ensembl) showed thatthe sequences are derived from a single exon (1C), which is locatedapproximately 19.5 kb upstream from the main ORF (Figs. 2A & B).The second transcript which is specific to the black phenotype has a5′UTR of 136 bp (GenBank accession no: HQ645015) and predictedto have only one uORF (Fig. 1B). The predicted uORF is not in-framewith the agouti ORF and it could produce a short peptide of ca. 52amino acids and does not show any identity with any previouslyreported protein. Sequence comparison of black specific 5′UTR withthe genomic sequence of alpaca revealed this 5′UTR results fromthree non-coding exons (1A, 2A and 3A) splicing as in the bovine(Girardot et al., 2005) and thus designated as transcript A (Figs. 2A& B). We are unable to locate the non-coding exon-1A on the avail-able alpaca genome; non-coding exons 2A and 3A are separated by
he structure of alpaca agouti genomic locus and polymorphisms. Black and gray boxes′UTRs) sequences, respectively. The agouti main start codon (ATG) and the stop codone donor (gt) and splice acceptor (ag) sites are also indicated. Numbers above and belowin the white, brown and black alpacas are indicated on the genomic organization of thepecies are reported above the corresponding position related to the alpaca agouti locus.s are highlighted in red. B. Structures of the common (C), black specific (A) and whiteesponding genomic locus. The position of coding exons (2, 3 & 4), introns (2 & 3) ande deduced based on Ensembl 2× alpaca genome release. The location of the non coding
White Brown Black0,00
0,30
0,60
0,90
1,20
Rel
ativ
e es
pres
sion
of
agou
ti m
RN
AA
B B
A B
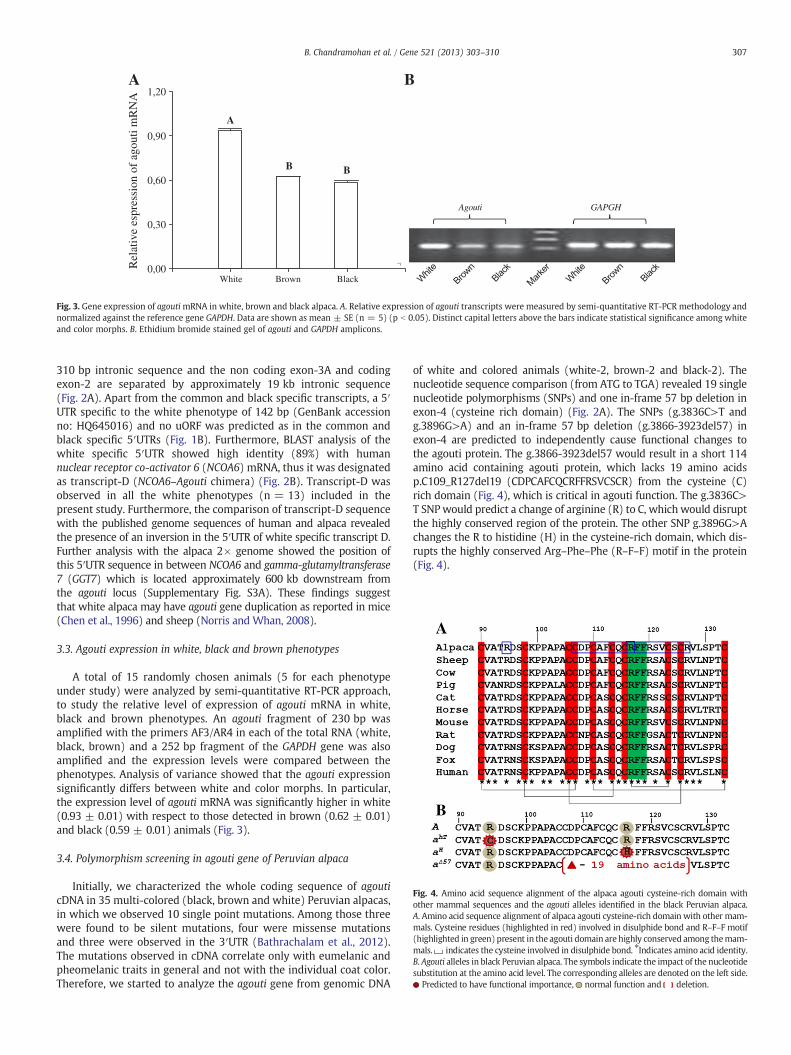
Fig. 3. Gene expression of agoutimRNA in white, brown and black alpaca. A. Relative expression of agouti transcripts were measured by semi-quantitative RT-PCR methodology andnormalized against the reference gene GAPDH. Data are shown as mean ± SE (n = 5) (p b 0.05). Distinct capital letters above the bars indicate statistical significance among whiteand color morphs. B. Ethidium bromide stained gel of agouti and GAPDH amplicons.
Fig. 4. Amino acid sequence alignment of the alpaca agouti cysteine-rich domain withother mammal sequences and the agouti alleles identified in the black Peruvian alpaca.A. Amino acid sequence alignment of alpaca agouti cysteine-rich domain with other mam-mals. Cysteine residues (highlighted in red) involved in disulphide bond and R–F–F motif(highlighted in green) present in the agouti domain are highly conserved among themam-mals. indicates the cysteine involved in disulphide bond. ⁎Indicates amino acid identity.B. Agouti alleles in black Peruvian alpaca. The symbols indicate the impact of the nucleotidesubstitution at the amino acid level. The corresponding alleles are denoted on the left side.
Predicted to have functional importance, normal function and deletion.
307B. Chandramohan et al. / Gene 521 (2013) 303–310
310 bp intronic sequence and the non coding exon-3A and codingexon-2 are separated by approximately 19 kb intronic sequence(Fig. 2A). Apart from the common and black specific transcripts, a 5′UTR specific to the white phenotype of 142 bp (GenBank accessionno: HQ645016) and no uORF was predicted as in the common andblack specific 5′UTRs (Fig. 1B). Furthermore, BLAST analysis of thewhite specific 5′UTR showed high identity (89%) with humannuclear receptor co-activator 6 (NCOA6) mRNA, thus it was designatedas transcript-D (NCOA6–Agouti chimera) (Fig. 2B). Transcript-D wasobserved in all the white phenotypes (n = 13) included in thepresent study. Furthermore, the comparison of transcript-D sequencewith the published genome sequences of human and alpaca revealedthe presence of an inversion in the 5′UTR of white specific transcript D.Further analysis with the alpaca 2× genome showed the position ofthis 5′UTR sequence in between NCOA6 and gamma-glutamyltransferase7 (GGT7) which is located approximately 600 kb downstream fromthe agouti locus (Supplementary Fig. S3A). These findings suggestthat white alpaca may have agouti gene duplication as reported in mice(Chen et al., 1996) and sheep (Norris and Whan, 2008).
3.3. Agouti expression in white, black and brown phenotypes
A total of 15 randomly chosen animals (5 for each phenotypeunder study) were analyzed by semi-quantitative RT-PCR approach,to study the relative level of expression of agouti mRNA in white,black and brown phenotypes. An agouti fragment of 230 bp wasamplified with the primers AF3/AR4 in each of the total RNA (white,black, brown) and a 252 bp fragment of the GAPDH gene was alsoamplified and the expression levels were compared between thephenotypes. Analysis of variance showed that the agouti expressionsignificantly differs between white and color morphs. In particular,the expression level of agouti mRNA was significantly higher in white(0.93 ± 0.01) with respect to those detected in brown (0.62 ± 0.01)and black (0.59 ± 0.01) animals (Fig. 3).
3.4. Polymorphism screening in agouti gene of Peruvian alpaca
Initially, we characterized the whole coding sequence of agouticDNA in 35 multi-colored (black, brown and white) Peruvian alpacas,in which we observed 10 single point mutations. Among those threewere found to be silent mutations, four were missense mutationsand three were observed in the 3′UTR (Bathrachalam et al., 2012).The mutations observed in cDNA correlate only with eumelanic andpheomelanic traits in general and not with the individual coat color.Therefore, we started to analyze the agouti gene from genomic DNA
of white and colored animals (white-2, brown-2 and black-2). Thenucleotide sequence comparison (from ATG to TGA) revealed 19 singlenucleotide polymorphisms (SNPs) and one in-frame 57 bp deletion inexon-4 (cysteine rich domain) (Fig. 2A). The SNPs (g.3836C>T andg.3896G>A) and an in-frame 57 bp deletion (g.3866-3923del57) inexon-4 are predicted to independently cause functional changes tothe agouti protein. The g.3866-3923del57 would result in a short 114amino acid containing agouti protein, which lacks 19 amino acidsp.C109_R127del19 (CDPCAFCQCRFFRSVCSCR) from the cysteine (C)rich domain (Fig. 4), which is critical in agouti function. The g.3836C>T SNPwould predict a change of arginine (R) to C, which would disruptthe highly conserved region of the protein. The other SNP g.3896G>Achanges the R to histidine (H) in the cysteine-rich domain, which dis-rupts the highly conserved Arg–Phe–Phe (R–F–F) motif in the protein(Fig. 4).

Table 1The PHI, PSI angle values of normal (p.R98) and mutated (p.C98) structures. Data wereobtained with software Ramachandran Plot 2.0.
Agouti with R98 Agouti with C98
Amino acid PHI PSI Amino acid PHI PSITHR −73.68 −1.45 THR −74.20 −4.01ARG −97.03 −12.87 CYS −93.70 −10.64ASP −83.04 120.84 ASP −83.76 116.97
Table 2Non-agouti mutations (in exon-4), alleles and genotypes identified in the black, brownand white alpacas in the present study.
ObservedGenotype
g.3836C>Tp.R98C
g.3896G>Ap.R118H
g.3866-3923del57p.C109_R127del
Phenotype Number ofanimals
aH/aΔ57 C/C A/– Yes Black 17aH/aht C/T A/G – Black 10A/A C/C G/G – Brown 2A/aht C/T G/G – Brown 10aH/A C/C A/G – White 11A/A C/C G/G – White 2
308 B. Chandramohan et al. / Gene 521 (2013) 303–310
3.5. In silico functional analysis and molecular modeling of agouti
In silico functional analysis by PANTHER, cSNP of the p.R98C andp.R118H supports the putative functional role of these mutations(p.R98C, subSPEC = −3.67256; Pdeleterious = 0.66208 and p.R118H,subSPEC = −6.36458; Pdeleterious = 0.96658). This was furtherevidenced by the SNAP tool (p.R98C, non-neutral; RI, 3; EA, 78% andp.R118H, non-neutral; RI, 6; EA, 93%). These predictions suggestthat the mutation at the position g.3896G>A may produce a nonfunctional agouti and the g.3836C>T variant may produce agouti pro-tein with minimal/partial activity. The protein sequences of alpacaagouti with mutation at the position C98 and agouti sequence (pdbstructure 2kza) were submitted to CPHmodels 3.0 Server (Lundet al., 2002; Nielsen et al., 2010). The phi, psi angles at this mutatedposition represented in Table 1 were checked using softwareRamachandran Plot 2.0 (http://dicsoft1.physics.iisc.ernet.in/rp/).There is not much difference in these torsion angles between the mu-tated and normal sequences and could possibly have less impact onthe structure and function of the protein. The quality of the modelwas further checked with WHATIF (Vriend, 1990) which reaffirmedthe quality of the model obtained from the CPHmodels server(Nielsen et al., 2010). The original amino acid R97 in 2kza showshydrogen bonding with C125 (Fig. 5A) but the mutated residue atthis position from the model shows simulated hydrogen bondingwith two amino acids, such as C125 and R126 (Fig. 5B). Therefore,the p.R98C variant seems to have minimal/partial effect on the func-tional property. As a result, it could be important to confirm the puta-tive predicted functional effect of the latter mutation with specificexperiments evaluating the pharmacological activity of this variant.
Fig. 5. Homology modeling of agouti mutation p.R98C. A. Normal structure showing the ARARG97 with CYS and its three hydrogen bonds with CYS125 and ARG126. The hydrogenlines with values.
3.6. Association analysis of g.3836C>T, g.3896G>A and g.3866_3923del57with phenotypes
A coat color panel comprising 52 DNA samples from white (13),brown (12) and black (27) Peruvian alpacas was genotyped forg.3836C>T and g.3896G>A SNPs and g.3866_3923del57 as reportedin Materials and methods. Table 2 summarizes the results of ouranalysis. As shown in the table, in black animals we found 2 geno-types: (i) black heterozygous for the g.3866_3923del57 agouti alleleand (ii) black homozygous to the undeleted agouti allele (see alsoFig. 4B & Supplementary Fig. S4). We further investigated this varia-tion in the black individuals to gain more insight, from which weidentified that the agouti allele without in-frame deletion presentin the g.3866_3923del57 animals has disrupted the R–F–F motif(g.3836C and g.3896A). This condition supported that the other alleleobserved in the g.3866_3923del57 (aΔ57) black could be non func-tional. The black animals homozygous to the undeleted agouti alleleare heterozygous to both the SNPs (g.3836C>T and g.3896G>A).
Furthermore, we performed a Fisher's exact test to better explore thepotential association between aht allele and black and non-black pheno-types. aΔ57 and aH alleles were, in fact, excluded from the test becauseaΔ57 segregates only in black alpaca and aH is present in all the blackalpacas. The aH allele is associated with aht and aΔ57 in 37% and in 63%of the black animals, respectively. The Fisher's test for aht resulted to benot significant (p-value 0.256). The aht allele is associated in non-blackalpaca with the wild dominant A allele in carrier heterozygous alpacas.
G97 and its hydrogen bond with CYS125. B. Mutated structure showing the replacedbond and its length between the interacting amino acids are shown in green dotted

309B. Chandramohan et al. / Gene 521 (2013) 303–310
4. Discussion
In this paper, we report the genomic structure of the agouti locus,its full length transcripts produced in skin and mutations that affectcoat color in native Peruvian alpaca populations. Our results revealedthat alpaca have the shortest agouti gene when compared to othermammals (Supplementary Table S2). The comparative sequenceanalysis with the human and alpaca genomes shows that the alpacaagouti gene is embedded in a conserved syntenic block, which con-tains RALY, IF2S2, XPOTP1, Agouti, AHCY, ITCH, PIGU, NCOA6 andGGT7. It has been previously reported in mice that the agouti geneproduces four different transcripts in the skin, among which a0.8 kb hair cycle specific transcript and its abundance correlateswell with the presence of pheomelanin (Bultman et al., 1992;Michaud et al., 1993). However, RT-PCR and RACE experimentspresented here clearly show that in alpaca, the agouti gene producesthree (A, D and C) similar sized transcripts in skin that differ as in the5′UTRs, as observed in mice. One class of transcript, here referred toas transcript-C consists of a sequence derived from the first, second,third, and fourth agouti exons, as described originally by Bultmanet al. (1992) and Miller et al. (1993). The transcript-C is common toall three phenotypes and specific to hair cycle (Vrieling et al., 1994).Moreover, the nested PCR amplification (data not shown) revealedthat the other two transcripts (A and D) observed are black andwhite specific. It is necessary to clarify the role of the two phenotypespecific transcripts with different 5'UTR sequences in alpaca.
Analysis of the 3 identified alpaca agouti full length transcriptswith m-fold using the Zucker algorithm (Zuker, 2003) predictedvery stable secondary structures and the dG values were observed tobe −285.59 kcal/mol (transcript-A), −312.49 kcal/mol (transcript-D)and dG = −296.56 kcal/mol (transcript-C). All 3 transcripts possessa common 224 bp 3′UTR bearing the canonical polyadenylation signalindicating that this region may play a role in the regulation of agoutiexpression at the transcriptional level. However, bioinformatic surveysof this region do not reveal any other known regulatory motifs such asAU rich element, miRNA seeds etc. On the contrary, characterization ofthe three identified 5′UTRs revealed several interesting sequence fea-tures. The main characteristic of transcript-C resides in its 4 uORFs. Thisis an uncommon situation, as less than 10% of known mammalian geneshave uORFs in their 5′UTR (Geballe and Morris, 1994; Hinnebusch,1990). The putative peptides encoded by the 4 uORFs have lengths rang-ing from 14 to 27 amino acids and show no homology with any knownprotein sequences. The black specific 5′UTR (transcript-A) has a singleuORF that can produce a short peptide of 52 amino acids and in thewhite specific 5′UTR none was predicted. This condition suggests thatthe different transcripts identified may play critical roles in regulatingthe expression of the gene at the translational level. Moreover, theuORFs found in the 5′UTR of transcripts support the alternative utilizationofmultiple promoterswhichmay serve tofine-tune agoutimRNAactivity.Numerous studies have been conducted regarding the optimal sequencestructure in which the AUG codon is embedded and have revealed thatthe optimal context for initiation of translation in vertebrates is represent-ed by the motif gccaccAUGg (Kozak, 1986, 1991, 1994). Experimentallydetermined optimal complexes have suggested the strong contributionof an adenine in position −3 and a guanine in position +4. None of theuAUGs present in the identified agouti 5′UTR perfectly matches thisstructure, except the uAUG present in the transcript-A (SupplementaryTable S3). Nevertheless, uAUG and the main start codon have an ade-nine in −3, placing the AUG of the main ORF in the optimal conditionfor translation initiation. Therefore, we performed a software-basedprediction of the secondary structures of transcript C, A, and D, focusingour attention on the 5′UTR and especially on the AUG sequence con-texts. Once again, the main AUG appears to be located in the bestcontext for translation initiation (Supplementary Fig. S6). The threedifferent 5′UTR sequences which belong to three potential promotersmay be physiologically significant. However, further investigations are
needed to evaluate the potential role of the secondary structuresand/or uORFs on the initiation efficiency of protein synthesis fromthe major ORF of alpaca agouti transcripts. Moreover, the coat colordependent expression of agouti mRNA indicates its potential role incoat color determination.
In agouti protein, the cysteine-rich domain alone is sufficient forhigh-affinity binding and activity at the respective melanocortin re-ceptors (MCRs) (Jackson et al., 2006; McNulty et al., 2001, 2005;Patel et al., 2010). The amino acid deletion/insertion/substitutionin this region may result in significant functional alteration. In ourpopulation the identified polymorphisms/deletions at the C-rich do-main may be of functional importance. The deletion at the positionp.C109_Rdel19 eliminates the two beta sheets and the R–F–F motiffrom the agouti functional domain, which are essential againstα-MSH (McNulty et al., 2005). Therefore, the deleted allele appearsto lose function. The other SNPs observed at the amino acid positions98 & 118 change the conserved R to C and the R–F–F motif into H–F–F.The R–F–F motif is important for functioning at MCRs; the disruptionin this motif may result in a non functional agouti protein since the al-teration of residues in and around R–F–F causes a decrease in agoutiprotein inhibition of α-MSH binding to MCRs (Kiefer et al., 1998)during signal transduction.
Three different alleles at the Asip locus have been identified in thisstudy in the native Peruvian population. They are all consistent withthe hypothesis of a recessive black mode of inheritance becauseall three mutations cause loss of function of the Asip protein. Thethree alleles are randomly distributed among black animals thereforesuggesting that this phenotype is subject to strong allelic heterogeneity.
In mice, the opposite orientation of an inverted duplication of agoutiand its association with difference in the ventral pigmentation has beenreported (Chen et al., 1996). A duplication encompassing the ovineagouti and AHCY coding sequence and the ITCH promoter causes thewhite phenotype in sheep (Norris and Whan, 2008). In our study, theinversion observed in 5′UTR of the white specific transcript (NCOA6–Agouti chimera) and its sequence localization in the alpaca genome,suggests that white phenotype may be caused by an allele at the ASIPlocus and the causativemutationmay be a chromosomal rearrangement(Supplementary Fig. S3). The genomic location of the white specific 5′UTR lies approximately 600 kb downstream from the agouti gene(Supplementary Fig. S3A) and, this condition supports the fact thatwhite alpaca seem to have duplication of the agouti gene locus bymeans of an intra-chromosomal homologous recombination event(Supplementary Fig. S3B). However, further research is needed to con-firm the possible association between the white phenotype and theidentified chromosomal rearrangement in alpaca.
In conclusion, the characterization of the agouti gene providesnew knowledge about the role of this gene in coat color phenotypicvariation in alpaca. The identification of non-agoutimutation in alpacarepresents an important step in the development of marker assistedbreeding program for coat colors. The characterization of transcriptsprovides another level of information about agouti gene regulationin alpaca pigmentation. Moreover, camelids are the only group ofmammals left out of research in the field of pigmentation biology.Therefore, our findings could greatly improve the knowledge aboutthe role that the agouti gene plays in pigmentation of alpaca.
Acknowledgment
The authors would like to greatly thank the Alpaca Research Foun-dation (ARF) for their financial support. We would also like to thankthe INIA, Peru and the people who helped in the sampling.
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.gene.2013.03.060.

310 B. Chandramohan et al. / Gene 521 (2013) 303–310
References
Bathrachalam, C., La Manna, V., Renieri, C., La Terza, A., 2012. Asip and MC1R cDNApolymorphism in alpaca. Fibre production in South American camelids and otherfibre animals, Sprinter Book. Wageningen Academic Publishers, pp. 93–96 (Isbn:978-90-8686-727-1).
Bromberg, Y., Rost, B., 2007. SNAP: predict effect of non-synonymous polymorphismson function. Nucleic Acids Res. 35, 3823–3835.
Bultman, S.J., Michaud, E.J., Woychik, R.P., 1992. Molecular characterization of themouse Agouti locus. Cell 71, 1195–1204.
Bultman, S.J., Klebig, M.L., Michaud, E.J., Sweet, H.O., Davisson, M.T., Woychik, R.P.,1994. Molecular analysis of reverse mutations from nonagouti (a) to black-and-tan(at) and white-bellied agouti (Aw) reveals alternative forms of agouti transcripts.Gene Dev. 8, 481–490.
Cecchi, T., Passamonti, P., Frank, E.N., Gonzales, M., Pucciarelli, F., Renieri, C., 2011.SUPREME-Project: pigmentation in South American Camelids: I. Quantificationand variation of combined eumelanins and pheomelanins in various coat colours.In: Gerken, M., Renieri, C. (Eds.), Progress in South American Camelids Research.vol. 105, vol. 105. EAAP Publication, Wageningen, pp. 207–210.
Chen, Y., Duhl, D.M., Barsh, G.S., 1996. Opposite orientations of an inverted duplicationand allelic variation at the mouse agouti locus. Genetics 144, 265–277.
Cozzali, C., Lorvik, S., Ceccarelli, P., Renieri, C., 1998. Ultrastructural features of melano-cytes from South American domestic camelids. Proceedings of the Italian Society ofVeterinary Science (SISVet), (Brescia, Italy), 52, pp. 41–42.
Cozzali, C., Dell'Aglio, C., Gargiulo, A.M., Frank, E., Hick, M., Ceccarelli, P., 2001.SUPREME-Project: pigmentation in South American camelids: II. Morphologicalfeatures of follicular melanocytes. In: Gerken, M., Renieri, C. (Eds.), Progress inSouth American Camelids Research, 105. EAAP Publ., Wageningen, pp. 237–238.
Darzentas, N., 2010. Circoletto: visualizing sequence similarity with Circos. Bioinfor-matics 26, 2620–2621.
Dayhoff, M.O., Schwartz, R.M., Orcutt, B.C., 1978. A model of evolutionary change in pro-teins. Atlas of Protein Sequence and Structure. vol. 5, Suppl. 3, vol. 5, Suppl. 3. NationalBiomedical Research Foundation, Washington, DC, pp. 345–351.
Duhl, D.M., Vrieling, H., Miller, K.A., Wolff, G.L., Barsh, G.S., 1994. Neomorphic agoutimutations in obese yellow mice. Nat. Genet. 8, 59–65.
Eizirik, E., Yuhki, N., Johnson, W.E., Menotti-Raymond, M., Hannah, S.S., O'Brien, S.J.,2003. Molecular genetics and evolution of melanism in the cat family. Curr. Biol.13, 448–453.
Fan, R., Yang, G., Dong, C., 2010. Study of hair melanin in various hair color Alpaca(Lama pacos). Asian-Aust. J. Anim. Sci. 23, 444–449.
Feeley, N.L., Munyard, K.A., 2009. Characterisation of the melanocortin 1 receptor genein alpaca and identification of possible markers associated with phenotypic varia-tions. Anim. Prod. Sci. 49, 675–681.
Feeley, N.L., Bottomley, S., Munyard, K.A., 2011. Three novel mutations in ASIP associ-ated with black fibre in alpacas (Vicugna pacos). J. Agric. Sci. 1–10.
Fontanesi, L., et al., 2010. Characterization of the rabbit agouti signaling protein (ASIP)gene: transcripts and phylogenetic analyses and identification of the causativemutation of the non-agouti black coat colour. Genomics 95, 166–175.
Gandarillas, H., 1971. Identificacion preliminar de los genes involucrados en la herenciadel colour de las llamas y alpacas. Div. Invest. Agric. Estac. Exp. Patacamaya, Boliva,Bol. Exp. N° 19, La Paz, Bolivia, p. 29.
Geballe, A.P., Morris, D.R., 1994. Initiation codons within 5′-leaders of mRNAs as regu-lators of translation. Trends Biochem. Sci. 19, 159–164.
Girardot, M., Martin, J., Guibert, S., Leveziel, H., Julien, R., Oulmouden, A., 2005. Wide-spread expression of the bovine Agouti gene results from at least three alternativepromoters. Pigment Cell Res. 18, 34–41.
Hinnebusch, A.G., 1990. Involvement of an initiation factor and protein phosphoryla-tion in translational control of GCN4 mRNA. Trends Biochem. Sci. 19, 159–164.
Jackson, I.J., 1994. Molecular and developmental genetics of mouse coat color. Annu.Rev. Genet. 28, 189–217.
Jackson, P.J., et al., 2006. Structural and molecular evolutionary analysis of Agouti andAgouti-related proteins. Chem. Biol. 13, 1297–1305.
Kerns, J.A., et al., 2004. Characterization of the dog Agouti gene and a non-agouti muta-tion in German Shepherd Dogs. Mamm. Genome 15, 798–808.
Kiefer, F., Arnold, K., Künzli, M., Bordoli, L., Schwede, T., 2009. The SWISS-MODELRepository and associated resources. Nucleic Acids Res. 37, D387–D392.
Kingsley, E.P., Manceau, M., Wiley, C.D., Hoekstra, H.E., 2009. Melanism in Peromyscusis caused by independent mutation in Agouti. PLoS One 4, e6435.
Kozak, M., 1986. Point mutation define a sequence flanking the AUG initiator codonthat modulates translation by eukaryotic ribosomes. Cell 44, 283–292.
Kozak, M., 1991. An analysis of vertebrate mRNA sequences: intimations of translationalcontrol. J. Cell Biol. 115, 887–903.
Kozak, M., 1994. Determinants of translational fidelity and efficiency in vertebratemRNAs. Biochimie 76, 815–821.
Kyte, J., Doolittle, R.F., 1982. A simple method for displaying the hydropathic characterof a protein. J. Mol. Biol. 157, 105–132.
Lamoreux, M.L., Delmas, V., Larue, L., Bennett, D.C., 2010. Pigment-Type Switching. TheColors of Mice. Wiley-Blackwell, pp. 177–206.
Linnen, C.R., Kingsley, E.P., Jensen, J.D., Hoekstra, H.E., 2009. On the origin and spread ofan adaptive allele in deer mice. Science 325, 1095–1098.
Lund, O., Nielsen, M., Lundegaard, C., Worning, P., 2002. CPHmodels 2.0: X3M aComputer Program to Extract 3D Models. Abstract at the CASP5 conference A102.
McNulty, J.C., Thompson, D.A., Bolin, K.A., Wilken, J., Barsh, G.S., Millhauser, G.L., 2001.High resolution NMR structure of the chemically-synthesized melanocortin recep-tor binding domain AGRP (87–132) of the agouti-related protein. Biochemistry 40,15520–15527.
McNulty, J.C., et al., 2005. Structures of the agouti signalling protein. J. Mol. Biol. 346,1059–1070.
Michaud, E.J., Bultman, S.J., Stubbs, L.J., Woychik, R.P., 1993. The embryonic lethality ofhomozygous lethal yellow mice (Ay/Ay) is associated with the disruption of anovel RNA-binding protein. Gene Dev. 7, 1203–1213.
Miller, M.W., et al., 1993. Cloning of the mouse agouti gene predicts a secreted proteinubiquitously expressed in mice carrying the lethal yellow mutation. Gene Dev. 7,454–467.
Montoliu, L., Oetting, W.S., Bennett, D.C., 2011. Color Genes. European Society forPigment Cell Research. (World Wide Web (URL: http://www.espcr.org/micemut/)(Last accessed August 18, 2011)).
Nielsen, M., Lundegaard, C., Lund, O., Petersen, T.N., 2010. CPHmodels-3.0 — remotehomology modeling using structure guided sequence profiles. Nucleic Acids Res.38, W576–W581.
Norris, B.J., Whan, V.A., 2008. A gene duplication affecting expression of the ovine ASIPgene is responsible for white and black sheep. Genome Res. 18, 1282–1293.
Patel,M.P., Cribb Fabersunne, C.S., Yang, Y.K., Kaelin, C.B., Barsh, G.S., Millhauser, G.L., 2010.Loop-swapped chimeras of the agouti-related protein and the agouti signaling proteinidentify contacts required for melanocortin 1 receptor selectivity and antagonism.J. Mol. Biol. 404, 45–55.
Powell, A.J., et al., 2008. Characterization of the effect of Melanocortin 1 Receptor, amember of the hair color genetic locus in alpaca (Lama pacos) fleece color differen-tiation. Small Rumin. Res. 79, 183–187.
Renieri, C., Trabalza Marinucci, M., Martino, G., Giordano, G., 1991. Preliminary analysison quality of fibres and coat colour in pigmented alpaca (Lama glama pacos PF).Proc. IX Italian Nat. Cong. ASPA, pp. 905–914.
Renieri, C., Lauvergne, J.J., Antonini, M., Lundie, R., 1995. Quantitative determinationof melanins in black, brown, grey and red phenotypes of sheep, goat and alpaca.Proc. 9th Int. Wool Textile Res. Conf., II, pp. 580–586.
Rieder, S., Taourit, S., Mariat, D., Langlois, B., Gue'rin,, G., 2001. Mutations in the agouti(ASIP), the extension (MC1R) and the brown (TYRP1) loci and their association tocoat colour phenotypes in horses. Mamm. Genome 12, 450–455.
Shi, X., Jarvis, D.L., 2006. A new rapid amplification of cDNA ends method for extremelyguanine plus cytosine-rich genes. Anal. Biochem. 356, 222–228.
Steiner, C.C., Weber, J.N., Hoekstra, H.E., 2007. Adaptive variation in beach miceproduced by two interacting pigmentation genes. PLoS Biol. 5, e219.
Thomas, P.D., et al., 2003. PANTHER: a library of protein families and subfamiliesindexed by function. Genome Res. 13, 2129–2141.
Thompson, J.D., Higgins, D.G., Gibson, T.J., 1994. CLUSTAL W: improving the sensitivityof progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22, 4673–4680.
Vage, D.I., Lu, D., Klungland, H., Lien, S., Adalsteinsson, S., Cone, R.D., 1997. A non-epistaticinteraction of agouti and extension in the fox, Vulpes vulpes. Nat. Genet. 15, 311–315.
Valbonesi, A., Apaza Castillo, N., La Manna, V., Gonzales Castilli, M.L., Huanca Mamani,T., Renieri, C., 2009. Inheritance of white, black and brown coat color in alpaca bysegregation analysis. EAAP Book of abstracts, p. 317.
Valbonesi, A., Apaza, N., La Manna, V., Gonzales, M.L., Huanca, T., Renieri, C., 2011.Inheritance of white, black and brown coat colors in alpaca (Vicuna pacos L.).Small Rumin. Res. 99, 16–19.
Velasco, J.M., Condorena, N.A., Novoa, C.M., Sumar, J.K., Franco, E., 1978a. Herencia decolores en alpacas. Centro de Investigacion IVITA, pp. 1–3.
Velasco, J.M., Condorena, N.A., Novoa, C.M., Sumar, J.K., Franco, E., 1978b. Herencia del colory tipo de fibra en alpacas. Resumenes de Proyectos de Investigacion (1975–1979),UNMSM, tomo II, pp. 1–127.
Vrieling, H., Duhl, D.M., Millar, S.E., Miller, K.A., Barsh, G.S., 1994. Differences in dorsaland ventral pigmentation result from regional expression of the mouse agoutigene. Proc. Natl. Acad. Sci. U. S. A. 91, 5667–5671.
Vriend, G., 1990. WHAT IF: a molecular modeling and drug design program. J. Mol.Graphics 8, 52–56.
Wang, Y., Westby, C.A., Johansen, M., Marshall, D.M., Granholm, N., 1998. Isolation, cloning,and sequencing of porcine agouti exon 2 (PorAex2). Pigment Cell Res. 11, 155–157.
Wilson, B.D., Ollmann, M.M., Kang, L., Stoffel, M., Bell, G.I., Barsh, G.S., 1995. Structureand function of ASP, the human homolog of the mouse agouti gene. Hum. Mol.Genet. 4, 223–230.
Zuker, M., 2003. Mfold web server for nucleic acid folding and hybridization prediction.Nucleic Acids Res. 31, 3406–3415.





























