Embed Size (px)
Citation preview

The Biophysical Property of A549 Cells Transferred by VEGF‐D
ZHEN WANG,1 XIU‐LI WU,2 XU WANG,3 HONG‐XIA TIAN,1 ZHI‐HONG CHEN,1 AND YANG‐QIU LI2,3
1Guangdong Lung Cancer Institute, Guangdong General Hospital, Guangdong Academy of Medical Sciences,Guangzhou, PR China2Institute of Hematology, Medical College, Jinan University, Guangzhou, PR China3Key Laboratory for Regenerative Medicine of Ministry of Education, Jinan University, Guangzhou, PR China
Summary: Vascular endothelial growth factor‐D(VEGF‐D) together with VEGF‐C is considered to beassociated with lymphangiogenesis and angiogenesis andinvolve in tumorization. This study aims to investigatethe influence of exogenous VEGF‐D gene on thebiophysical property of cell surface of lung adenocarci-noma cell line. A panel of lung adenocarcinoma cell lineswere examined the expression of VEGF‐D and VEGF‐Cby real‐time PCR. The VEGF‐D recombinant plasmidcontaining enhanced green fluorescence protein (EGFP)was constructed and transfected to the cell line with noexpression of VEGF‐D and confirmed by real‐time PCRand Western blot analysis. Topographic images of cellswereobtainedbyusingatomic forcemicroscope (AFM) incontact mode. Unlike VEGF‐C, VEGF‐D was found tohave a very low expression or undetectable expression inlung adenocarcinoma cell lines. The VEGF‐D recombi-nant plasmid had been constructed successfully and wastransferred into the human lung adenocarcinoma cell lineA549 cells which had no endogenous expression ofVEGF‐D, and exogenous VEGF‐D could be detected inmRNA and protein expression levels in the genemodifiedcells, while the VEGF‐C gene expression had no changeafter VEGF‐D transfection. After transfection, the irregu-lar microspikes or nano clusters could observe on the
surfaceofA549cells, andVEGF‐DtransfectedA549cellsbecame more rigid. The exogenous VEGF‐D gene mightcause the remarkable biophysical architectural changes inthe A549 cells, which might as a novel biomarker forevaluation of its biological function. SCANNING 36:202–208, 2014. © 2013 Wiley Periodicals, Inc
Key words: VEGF‐D, lung cancer, atomic forcemicroscope, gene transfer
Introduction
It is well known, during the tumoral processes, thelymphatic system is diverted by the tumor whichrequires endothelial microvasculature development fora nutrient supply and metastatic dissemination to distaltissues and organs (Karkkainen and Alitalo, 2002).Vascular endothelial growth factor (VEGF), a memberof the platelet‐derived growth factor family, consists offive glycoproteins as VEGF‐A, VEGF‐B, VEGF‐C,VEGF‐D, VEGF‐E, and placenta growth factor (PlGF)(Berse et al., ’92; Ferrara et al., ’92; 2003; Veikkolaand Alitalo, ’99). The VEGF‐C and VEGF‐D areligands for VEGF receptor (VEGFR)‐3 (Flt‐4), atyrosine kinase receptor that is expressed predomi-nantly in lymphatic endothelial cells (Kaipainen et al.,’95). Moreover, VEGF‐C and VEGF‐D also bind toVEGFR‐2 (KDR/FLK‐1), which is mainly expressedin vascular endothelial cells. The expression of VEGF‐C and VEGF‐D is considered to be associated withlymphangiogenesis and angiogenesis (Liu et al., 2008;Cao et al., 2012).
VEGF‐D, also known as c‐fos induced growth factor(FIGF), is a member of the platelet‐derived growthfactor/vascular endothelial growth factor (PDGF/VEGF)family and is active in angiogenesis, lymphangiogenesis,and endothelial cell growth. This secreted proteinundergoes a complex proteolytic maturation, generatingmultiple processed forms, which bind and activateVEGFR‐2 and VEGFR‐3 receptors. This protein isstructurally and functionally similar to vascular endothe-lial growth factor C. However, the relationship between
Contract grant sponsor: Medical Scientific Research Foundation of Guang-dong Province; contract grant number: A2010010; contract grant sponsor:Guangdong Natural Science Foundation; contract grant number:10451008004004158.
Conflicts of interest: None.
Zhen Wang and Xiu‐Li Wu contributed equally.
Address for reprints: Yang‐Qiu Li, Institute of Hematology, MedicalCollege and Key Laboratory for Regenerative Medicine of Ministry ofEducation, Jinan University, Guangzhou 510632, PR ChinaE‐mail: [email protected]
Received 16 January 2013; revised 2 February 2013; Accepted withrevision 11 February 2013
DOI: 10.1002/sca.21087Published online 22 March 2013 in Wiley Online Library(wileyonlinelibrary.com).
SCANNING VOL. 36, 202–208 (2014)© Wiley Periodicals, Inc.

VEGF‐C andVEGF‐D is controversial (Niki et al., 2000;O‐charoenrat et al., 2001; Koyama et al., 2003).
VEGF‐D was found to be expressed in gastriccancers, colon tissue, breast cancers and oral cancers(Hanrahan et al., 2003; Ishikawa et al., 2003; Nakamuraet al., 2003; Shintani et al., 2004). In non‐small cell lungcancer (NSCLC), the high expression of VEGF‐C andlow expression of VEGF‐D pattern seems to contributeto poor prognosis, however, this is controversial (Nikiet al., 2000; Kajita et al., 2001; Arinaga et al., 2003;Ogawa et al., 2004; Maekawa et al., 2007). As ourpreviously reported, VEGF‐D had a high expression innormal lung, low expression in lung adenocarcinomaincluding those with acquired resistance to gefitinib(epidermal growth factor receptor tyrosine kinaseinhibitors, EGFR‐TKIs) (Wang et al., 2007). VEGF‐Chas also higher expression in tumor marginal region thanin tumor center as VEGF‐D (Feng and Wang, 2010).
Rapid developments in biological atomic forcemicroscope (AFM) have allowed researchers to investi-gate the behavior of single living cells (Lekka andLaidler, 2009; Lekka, 2012). Using AFM to understandthe correlation between intracellular chemical processesand cellular physical properties will significantlyimprove our understanding of physiological events.All the changes in cell ultrastructure help us betterunderstand the structure–function relationships. TheAFM applications have the possibility to detect theearly subtle changes before significant pathology occurand to give the decision which time to terminate thetreatment. In the present study, we constructed theVEGF‐D recombinant plasmid, transfected the humanlung adenocarcinoma cell line, observed the exogenousVEGF‐D gene expression, and investigated the influenceof exogenous VEGF‐D gene on the expression pattern ofVEGF‐C and VEGF‐D, and detected the biophysicalarchitectural changes of lung adenocarcinoma cell lineafter transfection using AFM.
Materials and Methods
Human Lung Adenocarcinoma Cell Lines
The human lung adenocarcinoma cell lines Hcc827,PC‐9 (As previously described, Hcc827 and PC‐9 arederived from adenocarcinoma, harboring an EGFR exon19 in‐frame deletion that is highly sensitive to EGFR‐TKIs (Cheng et al., 2011), H460, H1299, H2228, SPAC‐1, L78, G1082, H1975, and A549, were cultured incomplete RPMI 1640 medium (Invitrogen, New York,USA) supplemented with 10% heat‐inactivated fetal calfserum (FCS; HyClone, Logan, UT), 100 U/ml penicillin,100 µg/ml streptomycin, 2 mM L‐glutamine, and 50 µM2‐mercaptoethanol and maintained at 37°C in a 5% CO2
incubator and harvested with trypsin‐EDTA when thecells were in exponential growth.
RNA Extraction and cDNA Synthesis
Total RNAwas extracted from the normal lung tissuesand cell lines with Trizol reagent according to themanufacturer’s recommendations (Trizol reagent; Invi-trogen). The RNA was reverse‐transcribed into firstsingle‐strand cDNA using random hexamer primers,reverse transcriptase, and the Superscript II kit (Power-Script™ Reverse, BD, San Jose, CA, USA) according tothe manufacturer’s instructions. The quality of cDNAwas confirmed by reverse transcriptase polymerase chainreaction (RT‐PCR) for b2‐microglubin gene amplifica-tion (Yin et al., 2011).
Construction of Recombinant Plasmids
Normal lung tissues were obtained from the thoracicsurgery of complete resection of early staged lung cancer.And the written informed consent forms were provided bythe patients. Briefly, the full‐length VEGF‐D gene ofnormal lung tissue was amplified by PCR using forwardprimer (VEGF‐D‐XhoI: 5' CCCTCGAGATGTACAGA-GAGTGGGTAG 3') and reverse primer (VEGF‐D‐EcoRI:5' CGGAATTCTCAAGGATTCTTTCGGCT 3'), andsubsequently cloned into the eukaryotic expression vectorpIRES2‐EGFP (Clontech, Palo Alto, CA, USA). The full‐length VEGF‐D gene was ligated via XhoI and EcoRIrestriction sites into the pIRES2‐EGFP vector with T4DNA ligase. The VEGF‐D recombinant plasmid wasverified by restriction analysis and sequencing.
Transfection of VEGF‐D Recombinant Plasmid
The human lung adenocarcinoma cell line with noVEGF‐D expression was selected and cultured for 5–6days to achieve high transfection efficiency, and thentransfected with VEGF‐D recombinant plasmid byLipofectamine™ 2000 transfection reagent (Invitrogen)according to the manufacturer’s recommendations. Thetransfection of pIRES2‐EGFP empty vector used asnegative control. Stably gene‐expressing cells wereobtained by G418 selection.
Immunophenotyping analysis of enhanced greenfluorescence protein (EGFP) expression was performedfor the determination of transfection efficiency by laserconfocal microscopy (LCM; 510 META DuoScan, CarlZeiss,Germany) andflowcytometry (FCM)at48and72 hafter transfection. The transfected cell linewas collected at48 and 72 h after transfection for RNA extraction, cDNAsynthesis and VEGF‐D gene expression analysis.
Real‐Time PCR
The mRNA expressions of VEGF‐C and VEGF‐Dgenes were detected by real‐time PCR using SYBR
Z. Wang et al.: VEGF‐D alters biophysical property of A549 cells 203

Green I with the Real Master Mix kit (Tiangen, Beijing,China). The primers for real‐time PCR were VEGF‐Dforward primer (VEGF‐D‐F: 5'‐ATGGACCAGTGA-AGCGATCAT‐3') VEGF‐D reverse primer (VEGF‐D‐R: 5'‐CAGCTTCCAGTCCTCAGAGTGA‐3') (VEGF‐Dproducts length 116 bp), VEGF‐C forward primer(VEGF‐C‐F: 5'‐ACAGGCCAACCTCAACTCAAG‐3')and VEGF‐C reverse primer (VEGF‐C‐R: 5'‐CCCCAC-ATCTATACACACCTCC‐3') (VEGF‐C products length139 bp). The b2M gene was used as an endogenousreference andwasused the followingprimer:b2Mforwardprimer (5'‐TACACTGAATTCACCCCCAC‐3') and b2Mreverse primer (5'‐CATCCAATCCAAATGCGGCA‐3')(products length 145 bp). The total volume of PCRreaction mixture was 25 µl containing 1 µl cDNA,0.4 mmol/l sense and antisense primers respectively,2.5 � Real Master Mix 11.25 µl (Tiangen). Reactionswere run in triplicate and repeated in three independentexperiments using theMJResearch real‐time PCR system(Bio‐Rad, Philadelphia, USA) under the followingconditions: 95°C for 2 min, followed by 40 cycles of95°C for 15 s and 60°C for 1 min. After reading theplates at 82°C, the melting curve could be obtainedfrom 55°C to 95°C (0.17°C/s). The relative mRNAexpression level of VEGF‐D or VEGF‐C gene wasused by the 2�DCt � 100% method (DCt ¼ Ct(VEGF‐D or
VEGF‐C) � Ct(b2M). For the �DCt calculation to be valid,the amplification efficiencies of target and reference mustbe approximately equal.
Western Blot Analysis
Soluble protein was extracted with the additionof RIPA lysis solution (including 1 � PBS, 1% NonidetP‐40, 0.5% sodium deoxycholate, 0.1% sodium dodecylsulfate (SDS), 10 mmol/L phenylmethylsulfonyl fluo-ride, 1 µg/ml aprotinin, and 100 mmol/L sodiumorthovanadate), Lysates were cleared by centrifugation(12,000 rpm, 4°C, 15 min), and soluble protein extractswere stored frozen at �80°C. Protein concentrationswere quantitated using the bicinchoninic acid assay.Proteins (100 µg) were separated by 7.5% SDS–PAGEand transferred to nitrocellulose membranes (Invitrogen)using a damp‐dry transfer device (Bio‐rad). Afterblocking for 1 h in 5% (w/v) defatted milk diluted inTrisbuffered saline Tween 20 (TBST), the membranewas washed and then probed with 1:500 mouse‐anti‐human VEGF‐D monoclonal antibody (abcam). Similarprocedures were performed with 1:800 mouse–anti‐human b actin (BOSTER, Wuhan, China). The 1:1,000horseradish peroxidase (HRP)‐conjugated rabbit–anti‐mouse IgG (eBioscience, San Diego, CA, USA) servedas the secondary antibodies. An Ultra ECL Kit(Enhanced Chemiluminescence Detection Kit forHRP, Multiscience, China) was used to visualize thebands corresponding to each antibody.
Atomic Force Microscope Measurement
The cell suspension was dropped on glass cover slipsand fixed in 4% paraformaldehyde and air dried prior toatomic force microscope (AFM) scanning and phase-contrast micrograph. Topographic images of cells wereobtained using an AFM (Autoprobe CP Research,Veeco) in contact mode. In all AFM experiments,gold‐coated silicon nitride tips (UL20B; Park ScientificInstruments) with a spring constant of 2.5 N/m and a tipdiameter of 20 nm were used (Jin et al., 2011; Zhanget al., 2012). An optical microscope was used to helpselect the desired cells and direct the position of the AFMtip. Single‐cell imaging was performed for five cells ofeach condition, and each cell was scanned three times. Togain information on the topography of the cell, all imageswere analyzed using the instrument‐equipped IP2.1software (Veeco Metrology Group, Santa Barbara, CA).The area analyzed with AFM was a 10 µm � 10 µmsquare. The length (L) and width (W) defined themaximum and minimum values of the cell diameter,respectively. The valley‐to‐peak value (Rp�v) representsthe difference between the maximum and minimumvalues of the z coordinate on the surface in the analyticalarea. The peak (H) and valley (h) value means themaximum and minimum height values of the cellmembrane surface, respectively. The r defines the ratioof L/W. The surface roughness value Ra was analyzed foreach sample. This value defined the mean value of thesurface roughness in the area being analyzed. All theseparameters were directly determined using the IP2.1software.
Roughness: Ra ¼ ð1=NÞPNn¼1 Zn � Z
�� ��. Z: givenby the average height within the included areas definedby a region group Z ¼ ð1=NÞPN
n¼1 Zn.
Statistical Analysis
Differences between numerical variables were calcu-lated by means of the independent‐sample t tests. The Pvalues were two tailed, and P < 0.05 was consideredstatistically significant. The SPSS software package 13.0(SPSS, Chicago, IL) was used for all data analysis.
Results
VEGF‐C and VEGF‐D Gene Expression in HumanLung Adenocarcinoma Cell Lines
The VEGF‐C and VEGF‐D expression pattern inhuman lung adenocarcinoma cell lines remainsunclear. We first selected 10 common human lungadenocarcinoma cell lines for analyzing the expressionpattern. The relative mRNA expression levels ofVEGF‐C and VEGF‐D gene in human lung
204 SCANNING VOL. 36, 2 (2014)

adenocarcinoma cell lines were determined by real‐time PCR with SYBR Green I technique and relativequantification method. The expression levels of VEGF‐Cgene could detected in the PC‐9 (0.4093 � 0.0369%),H460 (0.5300 � 0.0110%), L78 (0.2160 � 0.0137%),H1975 (0.0002 � 0.0001), Hcc827 (0.6187 �0.0131%), H1299 (0.5813 � 0.0851%), H2228(1.0196 � 0.1264%), SPAC‐1 (0.0319 � 0.0027%),G1082 (0.1925 � 0.0215%), and A549 (0.1968 �0.0245) cell lines. The expression levels of VEGF‐D genewere maintained the baseline in the Hcc827, H1299,H2228, SPAC‐1, G1082, and A549 cell lines, and theresults of electrophoresis confirmed the VEGF‐D genewere undetectable in these cell lines. A very low expres-sion level of VEGF‐D gene was observed as0.00008 � 0.00007%, 0.00009 � 0.00002%, 0.00044� 0.00001%, and 0.00002 � 0.00002% in the PC‐9,H460, L78, and H1975 cell line, respectively.
VEGF‐D Recombinant Plasmid Construction
The full‐length VEGF‐D gene of normal lung tissuehad been amplified for the construction of recombinantplasmids. After the construction of VEGF‐D recombi-nant plasmid, full‐length VEGF‐D gene sequence and
reading frame were confirmed by restriction enzymedigestion analysis, RT‐PCR and sequencing (data notshown).
VEGF‐D Gene‐Transduced A549 Cells
A549 cells (lung adenocarcinoma cell line, endoge-nous VEGF‐D gene expression undetectable) weretransfected with VEGF‐D recombinant plasmid byLipofectamine™ 2000 transfection. Immunophenotyp-ing analysis at 48 and 72 h after transfection confirmedthe high transfection efficiency by analyzing the EGFPexpression; Higher fluorescence intensity of transducedcells with VEGF‐D recombinant plasmid were found,compared with that transfected with empty vector byflow cytometry after transfection (Fig. 1(A, B)); Westernblot analysis confirmed the VEGF‐D protein expressionof transduced cells (Fig. 1(C)); High expression levels ofVEGF‐D gene after the VEGF‐D recombinant plasmidtransfection were confirmed by RT‐PCR and real‐timePCR, respectively (data not shown). These resultsindicated that the VEGF‐D recombinant plasmids hadbeen constructed successfully and the expression ofexogenous VEGF‐D gene could be detected in thetransduced cells. The expression of VEGF‐Cgene had no
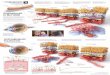
Fig 1. Exogenous gene expression was detected in the A549 cells after transfection. (A) Fluorescence intensity of EGFP expression inA549 cells transfected with empty vector detected by flow cytometry after transfection; (B) Fluorescence intensity of EGFP expression inA549 cells transfected with VEGF‐D recombinant plasmid detected by flow cytometry after transfection; (C) Exogenous VEGF‐Dexpression was confirmed in the A549 cells after transfection byWestern blot analysis. (a) A549 cells transfected with VEGF‐D recombinantplasmid; (b) A549 cells transfected with empty vector.
Z. Wang et al.: VEGF‐D alters biophysical property of A549 cells 205

significant change after VEGF‐D recombinant plasmidstransfection, which was detected by real‐time PCR.
Biophysical Changes in the A549 Cells AfterTransfection in the AFM
The AFM was used to study the biophysicalproperties (the parameters including morphology, mem-brane nanostructure, membrane pore, adhesion, distribu-tion, and so on) of A549 cells before and aftertransfection. After transfection, the AFM morphologicalimages of the A549 cells were shown in Figure 2. TheA549 cells transfected with VEGF‐D plasmid (Fig. 2(A–C)) formed a bigger and long fusiform shape andabundant pseudopodia compared with the A549 cellstransfected with empty vector (Fig. 2(D)). The cellsurface architecture in the A549 cells transfected withVEGF‐D plasmid showed more irregular microspikesespecially in the cell surface center. The irregularmicrospikes or nano clusters might be the ultrastructuremembrane proteins, which were reorganized aftertransfection of VEGF‐D plasmid and corresponding tothe large change in the structure of the cells surface.
The biophysical property data of A549 cells trans-fected with VEGF‐D plasmid, A549 cells transfected
with empty vector and A549 cells before transfectionmeasured by AFM were shown in Table I. The statisticalresults showed that the average surface fluctuation Ra ofthe A549 cells transfected with VEGF‐D plasmid wereincreased compared with that of the A549 cells trans-fected with empty vector (P ¼ 0.002) and the A549 cellsbefore transfection (P ¼ 0.001). The A549 cells trans-fected with empty vector had similar surface roughnessand mean particles high with the A549 cells beforetransfection (P ¼ 0.193). By measuring the adhesionforce of three groups of A549 cells, it was found that thestiffness of the VEGF‐D plasmid transfection group, theempty vector transfection group, and no transfectiongroup were 15.99 � 1.35, 11.56 � 0.95, and10.73 � 0.86 pN/nm, respectively. The comparison ofthe stiffness among these three groups demonstrated thatA549 cells transfected with VEGF‐D plasmid showedsignificantly more rigid than the A549 cells transfectedwith empty vector and A549 cells with no transfection(P ¼ 0.002 andP ¼ 0.001), and there was no significantdifference between the A549 cells transfected withempty vector and A549 cells with no transfection(P ¼ 0.243).
Discussion
VEGF‐C and VEGF‐D are two known VEGFhomologues, which bind VEGFR‐3 on lymphaticendothelial cells (McColl et al., 2003; Alitaloet al., 2005; Tammela et al., 2005). Stimulation orinhibition of lymphangiogenesis, especially via theVEGF‐C/VEGFD/VEGFR‐3 pathway, has proven ben-eficial in preclinical models of lymphedema and tumormetastasis, respectively, but their application to thetreatment of human disease requires more detailedunderstanding of the molecular basis of lymphangio-genesis. We quantified VEGF‐C and VEGF‐D genesexpression at mRNA levels in different lung adenocarci-noma cell lines. Unlike VEGF‐C mRNA, VEGF‐DmRNA expression was not detected in all cell lungadenocarcinoma cell lines in this study. The VEGF‐Dhasa very low expression in lung cancer cell lines, as theprevious results we found in lung cancer tissues,compared with that in the normal lung tissues (Wanget al., 2007). VEGF‐D potently stimulates lymphangio-genesis in adults and promotes metastasis of both humanand experimental mouse tumors (Rissanen et al., 2003;Kopfstein et al., 2007). However, studies in gene‐deficient mice have suggested that it is dispensable forembryonic lymphatic vascular development (Baldwinet al., 2005). VEGF‐D might function as a modifier ofVEGF‐C–driven early sprouting and migration oflymphatic endothelial cells in developing frog embryosand demonstrate the critical role of their receptorVEGFR‐3 in this process (Ny et al., 2008). In thisstudy, VEGF‐D was found to keep inactive in lung
Fig 2. AFM images of A549 cells. (A, B) AFM three‐dimensional images of A549 cells transfected with VEGF‐Drecombinant plasmid; (C, D) AFM three‐dimensional images ofA549 cells transfected with empty vector; (E, F) AFM three‐dimensional images of A549 cells before transfection.
206 SCANNING VOL. 36, 2 (2014)

adenocarcinoma cell lines. The inactivation of VEGF‐Dmight play an important role in tumorization. Therefore,it is worthily to investigate the difference of biologicalbehaviour in tumor cells with or without VEGF‐Dexpression.
In this study, we successfully constructed the VEGF‐D recombinant plasmid, and transfected to the A549 cellline (endogenous VEGF‐D negative), and observed theexogenous VEGF‐D gene expression. After we inten-tionally promote the expression level of VEGF‐D, theVEGF‐C expression level showed no change. Thissuggested that VEGF‐D might have no function ofregulating the expression of VEGF‐C in lung adenocar-cinoma cell lines and that might cause the VEGF‐C andVEGF‐D function differently in lung cancer.
Alteration of the cell membrane structure coulddirectly influence the cell function including tumor cell.AFM can detect cell membrane proteins and study cellsurface structures. By comparing the topography detailsof the cell surface before and after transfection of VEGF‐D plasmid, we found that more irregular microspikescould observe on the surface of A549 cells transfectedwith VEGF‐D. The irregular microspikes or nanoclusters might be the ultrastructure membrane proteins.These results indicate that the transfection of VEGF‐Dgene might cause the remarkable biophysical architec-tural changes in the A549 cells. Change in cell stiffness isa new characteristic of cancer cells that affects the waythey spread (Cross et al., 2007; Lekka and Laidler, 2009;Lekka, 2012). We found the A549 cells transfected withVEGF‐D plasmid became more rigid than the A549 cellstransfected with empty vector. That means the A549 cellswith exogenous VEGF‐Dmight be less metastatic or lessaggressive. In our previous study, VEGF‐D had a lowexpression in lung adenocarcinoma with acquiredresistance to EGFR‐TKIs (Wang et al., 2007). Thebiophysical architectural changes in the A549 cells afterVEGF‐D gene transfection suggested that the inactiva-tion of VEGF‐Dmight be involved in acquired resistanceto EGFR‐TKIs. However, further investigation is needed
to characterize the biological function as well as thesensitivity to TKIs in VEGF‐D modified A549 cells.
In conclusion, this is first attempt to observe thebiophysical architectural changes in the lung adenocar-cinoma cell line A549 cells after upregulating VEGF‐D.Upregulation of VEGF‐D might change the characteris-tic inmetastatic or aggressive function of A549 cells. Theunderlying complicated mechanism needs furtherresearch.
References
Alitalo K, Tammela T, Petrova TV. 2005. Lymphangiogenesis indevelopment and human disease. Nature 438:946–953.
Arinaga M, Noguchi T, Takeno S, et al. 2003. Clinical significanceof vascular endothelial growth factor C and vascularendothelial growth factor receptor 3 in patients with nonsmallcell lung carcinoma. Cancer 97:457–464.
Baldwin ME, Halford MM, Roufail S, et al. 2005. Vascularendothelial growth factor D is dispensable for development ofthe lymphatic system. Mol Cell Biol 25:2441–2449.
Berse B, Brown LF, Van deWater L, Dvorak HF, Senger DR. 1992.Vascular permeability factor (vascular endothelial growthfactor) gene is expressed differentially in normal tissues,macrophages, and tumors. Mol Biol Cell 3:211–220.
Cao R, Ji H, Feng N, et al. 2012. Collaborative interplay betweenFGF‐2 and VEGF‐C promotes lymphangiogenesis andmetastasis. Proc Natl Acad Sci USA 109:15894–15899.
Cheng H, An SJ, Dong S, et al. 2011. Molecular mechanism of theschedule‐dependent synergistic interaction in EGFR‐mutantnon‐small cell lung cancer cell lines treated with paclitaxel andgefitinib. J Hematol Oncol 4:5.
Cross SE, Jin YS, Rao J, Gimzewski JK. 2007. Nanomechanicalanalysis of cells from cancer patients. Nat Nanotechnol 2:780–783.
Feng Y, Wang W, Hu J, et al. 2010. Expression of VEGF‐C andVEGF‐D as significantmarkers for assessment of lymphangio-genesis and lymph node metastasis in non‐small cell lungcancer. Anat Rec 293:802–812.
Ferrara N, Gerber HP, LeCouter J. 2003. The biology of VEGF andits receptors. Nat Med 9:669–676.
Ferrara N, Houck K, Jakeman L, Leung DW. 1992. Molecular andbiological properties of the vascular endothelial growth factorfamily of proteins. Endocr Rev 13:18–32.
Hanrahan V, Currie MJ, Gunningham SP, et al. 2003. Theangiogenic switch for vascular endothelial growth factor
TABLE I Biophysical property of A549 cells by detection of AFM
A549 cells Width (µm) Length (µm) Height (nm) Ra (nm) Rq (nm)
Group 1 25.537�4.089 83.775�1.115 550.966�29.948 132.501�50.130 167.500�53.370Group 2 17.749�1.433 72.233�1.464 517.263�6.069 87.633�1.097 107.333�2.082Group 3 17.500�1.212 71.910�0.781 515.090�2.151 86.967�1.595 105.667�3.056a <0.0001 <0.0001 0.0002 0.0017 0.0002b <0.0001 <0.0001 0.0001 0.0015 0.0001c 0.6114 0.4572 0.2018 0.1935 0.0920
Group 1, transfected with VEGF‐D recombinant plasmid (constructed in pIRES2‐EGFP vector); group 2, transfected with empty vector; group 3, notransfection.a, b and c are P values of comparison. a, Group 1 compared with group 2; b, group 1 compared with group 3; c, group 2 compared with group 3.Ra, average roughness Ra¼(1/N) Ra ¼ ð1=NÞPN
n¼1 Zn � Z�� ��;
Z, Given by the average height within the included areas defined by a region group Z ¼ ð1=NÞPNn¼1 Zn;
Rq, root‐mean‐squared roughness Rq ¼ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi½PN
n¼1 ðZn � ZÞ2�ðN � 1Þq
.
Z. Wang et al.: VEGF‐D alters biophysical property of A549 cells 207

(VEGF)‐A, VEGF‐B, VEGF‐C, and VEGF‐D in the adeno-ma–carcinoma sequence during colorectal cancer progression.J Pathol 200:183–194.
Ishikawa M, Kitayama J, Kazama S, Nagawa H. 2003. Expressionof vascular endothelial growth factor C and D (VEGF‐C and ‐D) is an important risk factor for lymphatic metastasis inundifferentiated early gastric carcinoma. Jpn J Clin Oncol33:21–27.
Jin H, Ma S, Song B, et al. 2011. Liposome impaired the adhesionand spreading of HEK293 cells: an AFM study. Scanning33:413–418.
Kaipainen A, Korhonen J, Mustonen T, et al. 1995. Expression ofthe fms‐like tyrosine kinase 4 gene becomes restricted tolymphatic endothelium during development. Proc Natl AcadSci USA 92:3566–3570.
Kajita T, Ohta Y, Kimura K, et al. 2001. The expression of vascularendothelial growth factor C and its receptors in non‐small celllung cancer. Br J Cancer 85:255–260.
Karkkainen MJ, Alitalo K. 2002. Lymphatic endothelial regula-tion, lymphoedema, and lymph node metastasis. Semin CellDev Biol 13:9–18.
Kopfstein L, Veikkola T, Djonov VG, et al. 2007. Distinct roles ofvascular endothelial growth factor‐D in lymphangiogenesisand metastasis. Am J Pathol 170:1348–1361.
Koyama Y, Kaneko K, Akazawa K, et al. 2003. Vascularendothelial growth factor‐C and vascular endothelial growthfactor‐d messenger RNA expression in breast cancer:association with lymph node metastasis. Clin Breast Cancer4:354–360.
Lekka M. 2012. Atomic force microscopy: a tip for diagnosingcancer. Nat Nanotechnol 7:691–692.
Lekka M, Laidler P. 2009. Applicability of AFM in cancerdetection. Nat Nanotechnol 4:72 author reply 72–73.
Liu B, Ma J, Wang X, et al. 2008. Lymphangiogenesis and itsrelationship with lymphatic metastasis and prognosis inmalignant melanoma. Anat Rec 291:1227–1235.
Maekawa S, Iwasaki A, Shirakusa T, et al. 2007. Correlationbetween lymph node metastasis and the expression of VEGF‐C, VEGF‐D and VEGFR‐3 in T1 lung adenocarcinoma.Anticancer Res 27:3735–3741.
McColl BK, Baldwin ME, Roufail S, et al. 2003. Plasmin activatesthe lymphangiogenic growth factors VEGF‐C and VEGF‐D. JExp Med 198:863–868.
Nakamura Y, Yasuoka H, Tsujimoto M, et al. 2003. Flt‐4‐positivevessel density correlates with vascular endothelial growthfactor‐d expression, nodal status, and prognosis in breastcancer. Clin Cancer Res 9:5313–5317.
Niki T, Iba S, Tokunou M, et al. 2000. Expression of vascularendothelial growth factors A, B, C, and D and theirrelationships to lymph node status in lung adenocarcinoma.Clin Cancer Res 6:2431–2439.
Ny A, Koch M, Vandevelde W, et al. 2008. Role of VEGF‐Dand VEGFR‐3 in developmental lymphangiogenesis, achemicogenetic study in Xenopus tadpoles. Blood 112:1740–1749.
O‐charoenrat P, Rhys‐Evans P, Eccles SA. 2001. Expression ofvascular endothelial growth factor family members in headand neck squamous cell carcinoma correlates with lymph nodemetastasis. Cancer 92:556–568.
Ogawa E, Takenaka K, Yanagihara K, et al. 2004. Clinicalsignificance of VEGF‐C status in tumour cells and stromalmacrophages in non‐small cell lung cancer patients. Br JCancer 91:498–503.
Rissanen TT, Markkanen JE, Gruchala M, et al. 2003. VEGF‐D isthe strongest angiogenic and lymphangiogenic effector amongVEGFs delivered into skeletal muscle via adenoviruses. CircRes 92:1098–1106.
Shintani S, Li C, Ishikawa T, et al. 2004. Expression of vascularendothelial growth factor A, B, C, and D in oral squamous cellcarcinoma. Oral Oncol 40:13–20.
Tammela T, Enholm B, Alitalo K, Paavonen K. 2005. The biologyof vascular endothelial growth factors. Cardiovasc Res65:550–563.
Veikkola T, Alitalo K. 1999. VEGFs, receptors and angiogenesis.Semin Cancer Biol 9:211–220.
Wang Z, Wu Y, Zhang G, et al. 2007. VEGF‐D expression in lungadenocarcinoma with or without acquired resistance togefitinib and normal lung tissue. Zhongguo Fei Ai Za Zhi10:477–480.
Yin Q, Zha X, Yang L, et al. 2011. Generation of diffuse largeB cell lymphoma‐associated antigen‐specific Valpha6/Vbeta13 þ T cells by TCR gene transfer. J Hematol Oncol4:2.
Zhang Y, Zhang W, Wang S, et al. 2012. Detection of erythrocytesin patient with iron deficiency anemia using atomic forcemicroscopy. Scanning 34:215–220.
208 SCANNING VOL. 36, 2 (2014)