Embed Size (px)
Citation preview

The carbon balance of European croplands: a cross-site comparison of
simulation models
Martin Wattenbach1/*+, Oliver Sus2, Nicolas Vuichard3, Simon Lehuger4, Pia
Gottschalk1, Longhui Li3, Adrian Leip5, Mathew Williams2, Enrico Tormellieri6,
Werner Leo Kutsch6, Nina Buchmann7, Werner Eugster7, Dominique Dietiker7, Marc
Aubinet8, Eric Ceschia9, Pierre Béziat9, Thomas Gruenwald10, Astley Hastings1, Bruce
Osborne11, Philippe Ciais3, Pierre Cellier4, Pete Smith1
1/*University of Aberdeen, Institute of Biological and Environmental Sciences' St.
Machar Drive 23, Aberdeen, AB24 3UU, UK, email: [email protected]
2The University of Edinburgh, Crew Building, The King's Buildings, West Mains
Road, Edinburgh, EH9 3JN, UK32Laboratoire des Sciences du Climat et de
l’Environnement (LSCE/IPSL) CEA-CNRS-UVSQ (UMR 1572) CE Saclay L’Orme
des merisiers, Bât 712 91191 Gif sur Yvette Cedex France
4Agroscope Reckenholz-Tänikon Research Station ART, Air Pollution/Climate Group,
Reckenholzstrasse, 8046 Zurich, Switzerland
5Joint Research Centre, Institute for Environment and Sustainability, Climate Change
Unit (TP 050), I - 21020 Ispra (VA), Italy
6Max-Planck-Institute for Biogeochemistry, P.O. Box 10 01 64, 07701 Jena, Germany
Phone: +49 3641 576140, Fax: +49 3641 577100
7ETH Zurich, Institute of Plant Sciences LFW C56, Universitaetsstrasse 2, 8092
Zurich, Switzerland
9CESBIO - Unite mixte CNES-CNRS-UPS-IRD- UMR 5126, 18 Avenue Edouard
Belin, 31401 Toulouse Cedex 9 - France.
1

10Technical University Dresden, Institute of Hydrology and Meteorology, Department
of Meteorology, Pienner Strasse 23, D-01737 Tharandt, Germany
11UCD School of Biology and Environmental Science, University College Dublin,
Belfield, Dublin 4, Ireland
+current affiliation: Freie Universität Berlin, Institute of Meteorology, Carl-Heinrich-
Becker Weg 6-10, D-12165 Berlin, Germany
*corresponding author
2

Abstract
Croplands cover approximately 45% of Europe and play a significant role in the
overall carbon budget of the continent. However, the simulation of their carbon
balance is still uncertain due to the strong effects of human interference. Here, we
present a multi-site model comparison for four cropland ecosystem models namely
the DNDC, ORCHIDEE-STICS, CERES-EGC and SPA model. We compare the
accuracy of the models in predicting net ecosystem exchange (NEE), gross primary
production (GPP), ecosystem respiration (Reco) as well as actual evapo-transpiration
(ETa) for winter wheat (Triticum aestivum L.), winter barley (Hordeum vulgare L.) and
maize (Zea mays L.) derived from eddy covariance measurements on five sites of the
CarboEurope IP network. The models are all able to simulate mean daily GPP. The
simulation results for mean daily ETa and Reco are, however, less accurate. The
resulting simulation of daily NEE is adequate beside some cases where models fail
due to a lack in phase and amplitude alignment. ORCHIDEE-STICS and the SPA
demonstrate the best performance, nevertheless, they are not able to simulate full crop
rotations under consideration of multiple management. CERES-EGC and especially
DNDC although exhibiting a lower level of model accuracy are able to simulate such
conditions resulting in more accurate annual cumulative NEE.
keywords: croplands, eddy flux, Carbon, CO2, modelling
1. Introduction
3

Croplands are an important component of the European carbon balance {Janssens,
2003 #414; Janssens, 2005 #836; Ciais, in press #863; Kutsch, this issue #862}.They
cover a large area of between 1.10 (EPA - Corine2000) to 1.24 Mkm-2 {Gervois, 2008
#795}, within the EU27 plus Switzerland and there have been a number of integrated
studies that attempt to quantify, at the continental scale, the carbon and GHG balance,
each using different data sources from regional statistics, through remote sensing to
modelling approaches {Janssens, 2003 #414; Smith, 2005 #742; Smith, 2004 #440;
Leip, 2008 #716; Vleeshouwers, 2002 #763; Bondeau, 2007 #658; Gervois, 2008
#795; Ciais, in press #863}. However, greenhouse gas (GHG) emissions are largely
determined by the temporal and spatial sequence of human activity and there remains
a considerable degree of uncertainty {Smith, 2005 #762; Osborne, this issue #865}.
Firstly, regional or continental scale statistics are not consistently available for the
entire area of Europe {Ramankutty, 2008 #764} and available experimental data are
scare and come from heterogeneous sources. Secondly, remote sensing products lack
the accuracy and precision to reflect the degree of temporal and/or spatial
heterogeneity of croplands {Reeves, 2005 #694}. Thirdly, in the case of modelling,
the data now available through the CarboEurope network are the first comprehensive
high resolution flux data in order to parameterise process based agro-ecosystem
models (Smith et al., this issue). Croplands, however, play an important role in the
process of climate change mitigation {Smith, 2008 #735; IPCC, 2007 #761}. It is
therefore imperative to establish a better understanding of processes in order to
reproduce the current pattern of cropland carbon and GHG dynamics. In the
framework of the CarboEurope integrated project, detailed information about soil,
vegetation and carbon dynamics from eddy covariance systems in connection with
comprehensive crop management data covering entire crop rotations are available.
4

The network of sites covers all main regions of EU25 and Switzerland reflecting the
regional specifics of crops and their management (see Ceschia et al., this issue;
Eugster et al. this issue; Kutsch et al., this issue). Here, we present a multi-site model
comparison for four ecosystem models, namely the DeNitrification DeComposition
model (DNDC) {Li, 2005 #464; Li, 1994 #449; Li, 1992 #789}, the coupled
vegetation-crop model "Organising Carbon and Hydrology In Dynamic EcosystEms -
Simulateur mulTIdisciplinaire pour les Cultures Standard" (ORCHIDEE-STICS)
{Gervois, 2008 #795; De Noblet-Ducoudré, 2004 #796}, the "Crop Environment
REsource Synthesis Environnement et Grandes Cultures" (CERES-EGC) {Gabrielle,
2006 #813; Lehuger, 2009 #860} model and the Soil Plant Atmosphere model (SPA)
{Williams, 1996 #802}. These models represent a crosscut of widely applied model
species that are currently used to analyse the carbon dynamics of croplands. These
include site scale semi-empirical models, biogeochemical regional scale process
models, soil-vegetation-atmosphere transfer models (SVAT), and coupled global
vegetation models {Smith, 1997 #452; Brown, 2002 #466; Li, 1997 #791; Zhang,
2006 #794; Vuichard, 2008 #817; Gervois, 2008 #818, Lehuger, 2009 #819; Lehuger,
2007 #820; Law, 2000 #821; Williams, 1999 #822}. We compare the models in terms
of their performance to simulate the cycling of carbon and water between vegetation
and the atmosphere on a daily time scale. This study does not include other
greenhouse gasses due to the lack of high resolution measurements and the limitation
of ORCHIDEE-STICS and SPA to only simulate the carbon cycle.
The key elements of the carbon cycle are the fixation of atmospheric carbon dioxide
(CO2) by photosynthesis and its release by autotrophic and heterotrophic respiration.
The net flux as the sum of these three components is the net ecosystem exchange
5

(NEE) which can be measured by eddy covariance systems {Baldocchi, 2003 #831;
Black, 1996 #832; Moncrieff, 1997 #833; Baldocchi, 1996 #834; Reichstein, 2005
#816; Aubinet, 2008 #808; Smith, this issue #866}. NEE is the net uptake or release
of carbon by terrestrial ecosystems influenced by climatic and by non-climatic factors
{Morales, 2005 #830} like the plant water supply, leaf area index and soil carbon
dynamics, which are again influenced by crop type and associated management.
The terrestrial water cycle includes the precipitation that reaches the vegetation
surface from the atmosphere, which is subsequently partitioned into rain intercepted
by the canopy, surface and sub-surface runoff and water infiltrating the soil profile.
The intercepted water on the vegetation surface and the water which enters the soil
profile are subjected to the moisture gradient between the surface and atmosphere
causing it to evaporate. Another part of the water which enters the soil profile is taken
up by plant roots and transported to the leaves where it is released to the atmosphere
trough the stomata of the plant as transpiration. The sum of the two is the process of
evapotranspiration {Bosch, 1982 #436; Farley, 2005 #475; Zhang, 2001 #437;
Falloon #853; Morales, 2005 #830}}.
The two processes of water release and carbon uptake are closely interlinked by the
plant stomatal conductance for water and CO2 {Beer, 2007 #758}. Wide open stomata
maintain a high CO2 concentration for efficient photosynthesis, but also lose a great
amount of water that needs to be re-supplied by the rooting system {Bosch, 1982
#436; Jarvis, 1998 #637; Jarvis, 1985 #638; Monteith, 1965 #396; Monteith, 1977
#294; Beer, 2007 #758}.
6

Besides being a key factor in the exchange of carbon and water between the plant and
atmosphere, the soil water status also influences the microbial decay of carbon which
is strongly constrained by soil moisture conditions, as both too much and too little
water reduce microbial activity. Too little reduces the size of the aqueous environment
in which the microorganisms live and too much reduces the diffusion of CO2 and O2
through the soil {Pastor, 1986 #839; Davidson, 2006 #840, Morales, 2005 #830}.
There are studies looking at carbon fluxes from croplands to evaluate SVAT models
{Wang, 2007 #846; Adiku, 2006 #847; Wang, 2005 #848; Huang, 2009 #849}. In this
issue three studies conduct a detailed evaluation of the SPA (Sus et al, this issue),
CERES-EGC (Lehuger et al., this issue) and the DNDC (Dietiker et al., this issue)
model. However, the combined evaluation of water and carbon fluxes is relatively
rare in the literature {Adiku, 2006 #847}. Two studies are comparable to the aims of
this paper {Grant, 2007 #850; Morales, 2005 #830}. The study by {Grant, 2007
#850} only evaluates one model against cropland eddy covariance data for latent heat
and net biome productivity (NBP) measured over an irrigated and rain fed Maize-
Soybean rotation in the US. {Morales, 2005 #830}, on the other hand, compares a
number of biogeochemical and coupled global vegetation models including
ORCHIDEE against global EUROFLUX data but in that case for global forest biomes
and not cropland ecosystems.
Here we focus on the accurate representation of main components of the carbon cycle,
net ecosystem exchange (NEE), ecosystem respiration (Reco) and gross primary
production (GPP) in connection with the actual evapo-transpiration (ETa). This model
evaluation is conducted on a daily time scale and over a gradient of environmental
7

conditions in Europe from the eastern part of Germany (mean annual temperature
(T)=7.3oC; precipitation (P)=850mm) over a central mountainous alpine region in
Switzerland (T=9.0°C; P=1100 mm) to the central and southern part of France
(T=12.9°C; P=700mm). This multi-criterion, multi-model, multi-site evaluation
enables insights into the applicability of the models to simulate the carbon balance of
cropland ecosystems within the European Union.
2 Material and Methods
2.1 The cropland sites
The four models were run at four sites (Oensingen, Grignon, Aurade, Klingenberg)
for one year of winter wheat (Triticum aestivum) at each site, one year for winter
barley (Hordeum vulgare) at Gebesee, and one year for maize (Zea mays) at Grignon.
However, the extent of our comparison is limited by model differences in the number
of crop types simulated and the kind of output data produced. For example, SPA has
no maize and ORCHIDEE-STICS no winter barley implementation yet, and CERES-
EGC produces no estimate of Reco. The combinations of sites, models, crops and years
are given in Table 2. Model results were compared accordingly.
A more detailed description of the sites is given in other papers in this special issue
(Kutsch et al. this issue, Ceschia et al. this issue).
2.2 Models
8

2.2.1 The DNDC model
The DNDC model (in this study version 9.2), is a general model of C and N
biogeochemistry in agricultural ecosystems {Li, 2005 #464; Li, 1994 #449; Li, 1992
#789}. It is a process-oriented simulation model, which contains four interacting sub-
models for soil climate, de-nitrification, decomposition and plant growth. The model
has been tested against numerous field data sets of nitrous oxide (N2O) emissions
{Tonitto, 2007 #788; Frolking, 1998 #790; David, 2009 #792; Abdalla #783} and soil
carbon dynamics {Smith, 1997 #452; Brown, 2002 #466; Li, 1997 #791; Zhang, 2006
#794}. In the application of DNDC presented here, we focus on analysing the
implementation of the carbon cycle in the decomposition and plant growth part of the
model, together with the section of the model to calculate the latent heat flux.
plant growth
DNDC simulates plant growth using an empirical approach calculating
photosynthesis, respiration, water and N demand, C allocation, crop yield, and litter
production on a daily time step for about sixty different crops. Photosynthesis is
calculated using the radiation use efficiency approach {Monteith, 1977 #294}, with
interception depending on leaf areas index based on Beer’s law. Phenology is
simulated using accumulative thermal degree days approach. A user defined amount
of litter either from roots or aboveground residue after harvest is assumed to enter the
carbon cycle of the model {Qiu, 2009 #793}.
soil carbon dynamics
9

The soil organic carbon (SOC) dynamics are simulated by assuming four main pools:
plant residue, microbial biomes, active humus, and passive humus. Each of the main
pools is subdivided into one or more sub-pools with different properties. The daily
decomposition rate is calculated depending on the relative size of each sub-pool and is
regulated by each pool size, its decomposition rate, the soil clay content, N
availability, soil temperature and moisture, and its depth in the soil profile. In the
process of the decomposition simulation, carbon is transferred to the soil pool with the
next lower decomposition rate, partially assimilated into microbial biomes, and
partially converted into CO2 {Qiu, 2009 #793}.
latent heat
Potential evapo-transpiration (ET) in DNDC is calculated using a daily average value
from the Thornthwaite formula {Thornthwaite, 1965 #503}. Subsequently, potential
ET is separated into potential evaporation and transpiration. To calculate the potential
transpiration, the water demand of plants is calculated based on the daily biomass
increment using the water/biomass ratio of the crops. The actual plant transpiration is
then calculated by taking the actual soil water content of the soil profile into account
{Li, 2006 #812}.
PLEASE INSERT TABLE 1
2.2.2 The ORCHIDEE-STICS model
ORCHIDEE-STICS is a coupled model {Gervois, 2008 #795; De Noblet-Ducoudré,
2004 #796} consisting of a dynamic global vegetation model ORCHIDEE {Krinner,
10

2005 #797}, and a process-oriented crop model STICS {Brisson, 2002 #798; Brisson,
1998 #799; Brisson, 2003 #800}.
The ORCHIDEE model calculates, for diverse vegetation types (plant functional
types), surface CO2, water vapour and heat fluxes driven by varying weather, and the
soil water and C pools dynamics. It contains a biophysical module, dealing with
photosynthesis and energy balance calculations each 30 min, and a carbon dynamics
module, dealing with phenology, growth, allocation, mortality and SOM
decomposition, on a daily time step.
For better representing cultivated plants, their phenology and management related
growth is calculated by the STICS model which is coupled to ORCHIDEE providing
daily foliar index, root density profiles, nitrogen stress, vegetation height, and
irrigation requirements. These variables are then sequentially assimilated into
ORCHIDEE each day to further calculate accurately gross primary production (GPP).
Currently, the ORCHIDEE-STICS model has been used for simulating Wheat,
Soybean and Maize although STICS has sufficiently generic parameterizations to
allow simulation of other crop species. The processes of the STICS sub model are:
plant growth
Crop growth is driven by the plant carbon accumulation {de Witt, 1978 #899}, solar
radiation intercepted by the foliage. According to the plant type, crop development is
driven either by a thermal index (degree-days), a photothermal index or a
photothermal index taking into account vernalisation. The vernalisation factor is the
ratio between the sum of vernalising days since planting and plant vernalisation
requirements {Weir, 1984 #900}. Water stress and nitrogen stress, if any, reduce leaf
growth and biomass accumulation, based on stress indices that are calculated in water
11

and nitrogen balance modules {Brisson, 2002 #798; Brisson, 1998 #799; Brisson,
2003 #800}.
soil carbon dynamics
latent heat flux
2.2.3 The SPA model
The Soil Plant Atmosphere model (SPA; {Williams, 1996 #802}) is a process-based
model that simulates ecosystem photosynthesis and water balance at fine temporal
and spatial scales (up to 30 minute time step, ten canopy and twenty soil layers). The
scale of parameterization (leaf-level) and prediction (canopy-level) have been
designed to allow the model to diagnose eddy flux data and to provide a tool for
scaling up leaf level processes to canopy and landscape scales {Williams, 2000
#803}.
plant growth
The SPA model employs the Farquhar approach of leaf-level photosynthesis
{Farquhar, 1980 #62} to calculate the amount of carbohydrates synthesised at each
time step. The carbohydrates are then allocated to one root and four aboveground
carbon pools (labile, foliage, stem and storage organ carbon pools), and accounts for
autotrophic and heterotrophic respiratory processes {Vertregt, 1987 #804}. The C
allocation pattern itself is dependent on the developmental stage (DS) of the crop
plant. DS is calculated as the sum of daily developmental rates, which are a function
12

of temperature, photoperiod, and vernalisation (Sus et al., this issue). Senescence is
calculated as a function of either mutual shading effects of canopies with a LAI > 4,
or developmental rate in the reproductive phase, whichever is dominant. Senescent
carbon is either remobilised and subsequently reallocated to the growing storage
organ, or added to a standing dead leaf biomass carbon pool.
soil carbon dynamics
At harvest, the fraction of the aboveground biomass exported from the field is
estimated by the storage organ C content plus non-crop residue leaf and stem C mass.
The residual crop biomass gradually enters the litter carbon pool. The fraction of crop
residue entering either the litter or soil carbon pool further depends on land
management and can be adjusted accordingly. Following this approach, SPA models
the carbon mass balance for winter/spring barley and wheat (Sus et al. this issue)
latent heat
SPA uses the Penman-Monteith equation to determine leaf-level transpiration
{Penman, 1948 #497; Monteith, 1965 #396; Monteith, 1977 #294}. It is linked to the
photosynthesis module by a novel model of stomatal conductance that optimizes daily
carbon gain per unit leaf nitrogen, within the limitations of canopy water storage and
soil to canopy water transport.
2.2.4 The CERES-EGC model
The original CERES model is a soil-crop model {Jones, 1986 #814}. It was extended
to be CERES-EGC by {Gabrielle, 2006 #813; Lehuger, 2009 #860} by moving the
13

focus towards the simulation of nitrogen cycle related processes such as nitrate
leaching, emissions of N2O and nitrogen oxides. CERES-EGC runs on a daily time
step, and requires daily rain, mean air temperature and Penman potential evapo-
transpiration {Penman, 1948 #497} as forcing variables to calculate actual evapo-
transpiration.
CERES-EGC simulates water, carbon and nitrogen in the soil-crop systems in a
number of sub modules. A physical sub-model simulates heat, water and nitrate
movement in the soil. It is also responsible for the calculation of soil evaporation,
plant water uptake and transpiration. A biological sub-model simulates the growth and
phenology of the crops.
plant growth
The model calculates net photosynthesis as a linear function of intercepted radiation
according to the Monteith approach {Monteith, 1977 #294}, with light interception
depending on leaf area index based on Beer’s law. The key species specific parameter
in this calculation is the radiation use efficiency (RUE) defined as the dry biomass
produced per unit of radiation intercepted. Photosynthates are partitioned on a daily
basis to currently growing organs (roots, leaves, stems, fruits) according to crop
development stage. The latter is driven by the accumulation of growing degree days,
as well as cold temperature and day-length for crops sensitive to vernalisation and
photoperiod. Lastly, crop N uptake is computed through a supply/demand scheme,
with soil supply depending on soil nitrate and ammonium concentrations and root
length density.
14

soil carbon
A micro-biological sub-model simulates the turnover of organic matter in the plough
layer. Decomposition, mineralisation and N-immobilisation are modelled with three
pools of organic matter (OM): the labile OM, the microbial biomass and the humads.
Kinetic rate constants de ne the C and N ows between the different pools. Directfi fl
eld emissions of COfi 2, N2O, NO and NH3 into the atmosphere are simulated with
different trace gas modules (Lehuger et al. this issue).
latent heat
The Penman potential evapo-transpiration {Penman, 1948 #497} as forcing variables
to calculate actual evapo-transpiration based on the water status of the soil and crop
respectively.
2.2.5 Simulation set-up
Input data
Models are driven by meteorological variables derived from half hourly
measurements at each site. Simulation time steps differ also among models (and
consequently time resolution of input meteorological variables): from daily (for
DNDC) and CERES-EGC) to half-hourly (for SPA and ORCHIDEE-STICS). The
number of meteorological variables used also differs among the models: from only
two for DNDC (temperature and precipitation) to up to six for ORCHIDEE-STICS
(temperature, precipitation, incident long and short-wave radiation, relative humidity
and wind speed).
15

In case of gaps in the on-site daily time series the data from the nearest climate station
out of the ECAD dataset was used to gap-fill with daily values for models with daily
time steps {Klein Tank, 2002 #801}. In case of gaps in the half hourly data reuired by
the ORCHIDEE-STICS and SPA, data were gap filled with long term site specific
half hourly average.
The soil texture is determined from measurements made on each site (Kutsch et al.
this issue, Sus et al., this issue) and prescribed accordingly for all four models.
Management events, such as fertilisation, irrigation, planting, harvest or ploughing are
also defined by on-site observed values (Kutsch et al. this issue). In the SPA model,
the effects of fertilization are not taken into account, but reported harvest dates and
crop residue management are considered in the model runs.
The three simulated crops, winter wheat (Triticum aestivum L.), Winter Barley winter
barley (Hordeum vulgare) and maize (Zea mays L.), were parameterised for each
model using ‘standard’ published values {Mueller, 2005 #539; Kätterer, 1997 #765;
Van Den Boogaard, 1997 #767; McMaster, 2003 #774 ; Lohila, 2003 #641; Juskiw,
2001 #645; López, 1997 #775} or data provided by the site. For the DNDC model,
values given in table 1 are for the time of harvest as prescribed in the DNDC user
manual. For the estimation of plant C/N ratios in DNDC, site data for biomass and
carbon and nitrogen content were used. Where carbon and nitrogen content data were
not available, fertiliser application data, provided by the site managers, were used to
derive site specific C/N ratios under the assumption of optimum nitrogen supply
during growth.
16

Initialisation procedure
For the initialisation of the soil carbon and nitrogen pools, the DNDC model was run
for ten years using daily ECAD weather data from the nearest weather station {Klein
Tank, 2002 #801}. The litter input for the initialisation period was manually adjusted
so that modelled matched measured total soil organic carbon at the beginning of the
simulation period. The fertilizer input for this initialisation period was assumed to be
in the same order of magnitude as the mineral fertilizer input during the simulation
period. For simulations performed by ORCHIDEE-STICS, the soil carbon pools were
initialized to their steady state equilibrium values after a thousand-years spin-up
during which climate and management practice of the simulated year were repeatedly
cycled. For SPA simulations, the initial soil organic matter as well as labile carbon
contents are estimated, based on field observations reported in the literature {Anthoni,
2004 #807; Aubinet, 2008 #808; Halley, 1988 #805}. The initial labile carbon content
is equal to the seed carbon content at sowing. The CERES-EGC model was run for
two rotations in all sites before the measurement period to stabilize the C and N soil
pools and dampen the initial conditions. The same meteorological data were
repeatedly used in case the historical data were not available.
DNDC was set up to run for the full crop rotation using litter inputs based on the
calculated crop growth. The optimum yield parameter was adjusted to match the site
specific yields. The DNDC model does not incorporate the concept of crop
germination and emergence, it assumes an initial biomass at the day of sowing with
the immediate start of photosynthesis. In order to simulate a realistic crop growth we
assume 20 days for the crop to emerge from the day of seeding to the start of
photosynthesis {McMaster, 2003 #774; Stone, 1999 #770}. For the ORCHIDEE-
17

STICS runs, one simulation was performed for each crop season. The SPA runs have
been initiated at sowing (date as reported from the site) and terminated by the end of
the following year. It is important to note that the SPA outputs are only truly
representative of the actual growth period from sowing to harvest, as no post-harvest
voluntary re-growth, ploughing or sowing has been considered. However, reported
fractions of crop residual biomass were considered for the simulation of post-harvest
heterotrophic respiration fluxes (Sus et al. this issue). The simulations of CERES-
EGC were set up for full crop rotations. The sowing date of each crop was initiated in
the management file and harvest time was simulated when crops attained the
physiological maturity. Grain and straw were exported out of the field while crop
residues and roots were incorporated into the SOM pools at the date of the post-
harvest tillage. Catch crops (at Grignon) and volunteers from the previous crop (at
Auradé) were simulated between the crop seasons of a winter crop (barley and wheat)
and a spring crop (maize and sunflower).
PLEASE INSERT TABLE 1
Model comparison
We use the site eddy covariance derived data for GPP, NEE, Reco {Reichstein, 2005
#827} and latent heat aggregated on a daily time step for the model comparison. The
data are provided in gap filled format which is checked for errors and outliers and
aggregated for different time intervals from half hourly to weekly in the CarboEurope
IP ecosystem database {Papale, 2006 #824; Papale, 2006 #825; Moffat, 2007 #815;
Reichstein, 2005 #816}.
18

The statistical methods of the model comparison are based on {Smith, 1997 #784;
Morales, 2005 #531}. The data analysis itself was performed by the statistical
package R {R, 2009 #809}. To determine if the daily measured NEE, GPP and Reco
data are normally distributed we used the Shapiro-Wilk implementation in R
{Royston, 1992 #835} independently for each site and each year of the comparison.
The highly significant result of the test (p<0.01) indicates a very high probability that
the data are not from a normally distributed population. Subsequently we use the non-
parametric Kendall tau rank correlation coefficient or r, to assess if simulations and$
measurements are associated:
Eq.1
where r is the correlation coefficient, n the number of observations, nc is the number
of concordant pairs, and nd is the number of discordant pairs. We estimate the
significance based on {Best, 1974 #861}.
To analyse the association between measured and simulated data we calculate a
locally weighted polynomial regression {Cleveland, 1979 #811; Cleveland, 1981
#810}, the "lowess" smoother, within 1:1 scatter plots of measured against simulated
values. The lowess fit function is a continuous fit to subsets of the data without
requiring a predefined fitting model. Thus, the fitted line describes the best model fit
for a subsection of data by a moving window indicator. A change in the slope of the
fitted line indicates a change in the correlation between measured and modelled
values due to shifting from low to high values. In all pair plots, the right hand side
19

panel shows the r values in black for significant correlation coefficient results
(p<0.05), and in light grey for non-significant associations (p>0.05). The size of the
text indicates the strength of the association. We use a 95% confidence interval as
suggested by {Smith, 1997 #784: Morales, 2005 #531}.
The model efficiency factor E {Nash, 1979 #298} is another measure of model
performance. It compares the squared sum of the absolute error with the squared sum
of the difference between the observations and their mean value. It compares the
ability of the model to reproduce the daily data variability with a much simpler model
that is based on the arithmetic mean of the measurements:
Eq. 2
where Oi are the observed values, Pi are the simulated values, n are the total number
of observations and i the current observation. It ranges from 1 to - . Any model!
giving a negative value cannot be recommended, a value of 0 indicates that the model
does not perform better than using the mean of the observations, and values close to 1
indicate a ‘near-perfect’ fit.
To compare the annual carbon balance of the sites, the total annual flux is calculated
for observed and simulated NEE:
Eq. 3
20

Eq 4
where mo,i is the cumulative sum of the daily (i) observed fluxes and mp,i the
cumulative sum of the simulated flux. The difference of the two is the absolute error
x presented in table 3. We tested the median of the two cumulative flux distributions#
for significant differences on the 95% confidence level by using the R implementation
of the Wilcoxon rank sum test {Corder, 2009 #854; Hollander, 1973 #855}.
2. Results
The statistical results of r and E for all sites are summarised in table 3 and in the
figures that show the site to site performance in terms of r. There are common patterns
for all sites and for all models which are:
o inconsistency between the models to reproduce low carbon fluxes ranging
between -2 gC m2 day-1 and 2 g C m2 day-1 (odd numbered figures 1-11).
o all models have problems with capturing the crop phenology, which is indirectly
indicated by either an overestimation of the amplitude of growth in the later
stage of crop development (mainly the case for ORCHIDEE-STICS, SPA and
CERES model) or a phase shift of growth as seen for DNDC (especially for
winter wheat and barley, by simulating a too early onset of growth) (even
numbered figures 2-12)
o a good to very-good fit for GPP and Reco at fluxes below respectively above the
-2 gC m2 day-1 and 2 g C m2 day-1 flux rates, but a relatively poorer fit for NEE.
21

o DNDC and CERES-EGC: a relatively poor performance in reproducing the
latent heat flux in contrast to a better performance for NEE, GPP and Reco,
suggesting problems in the coupling of the water and carbon flux in these
models.
PLEASE INSERT TABLE 3
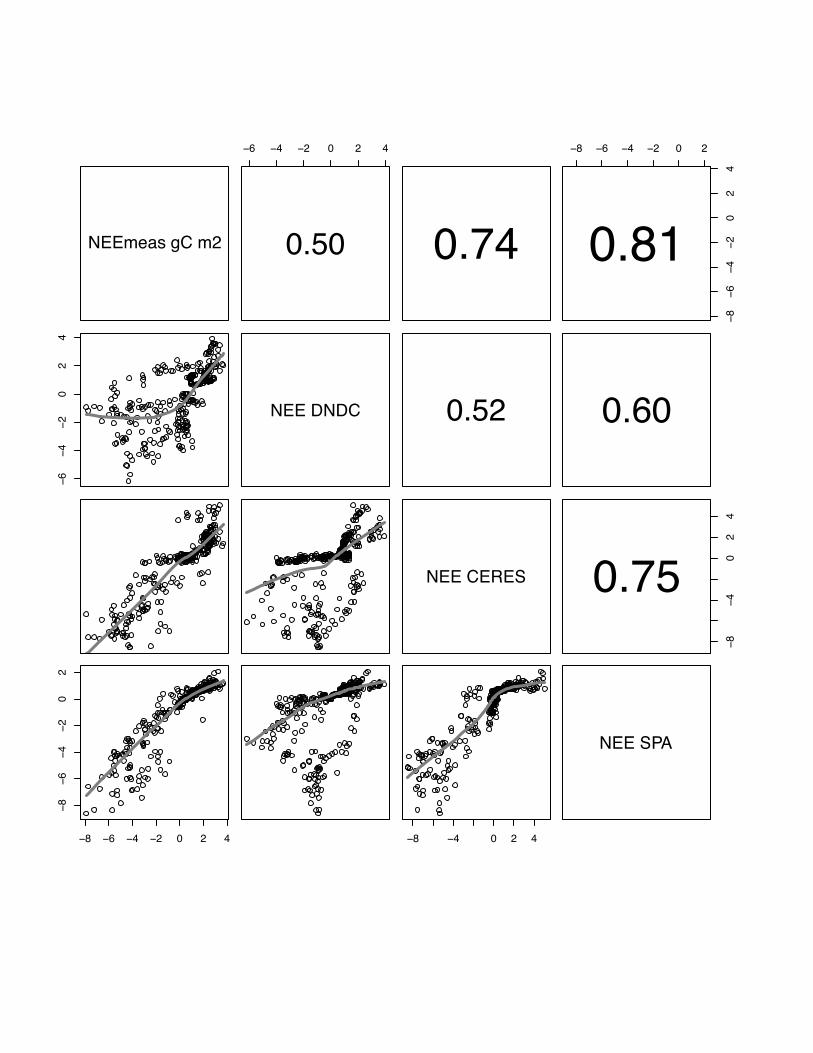
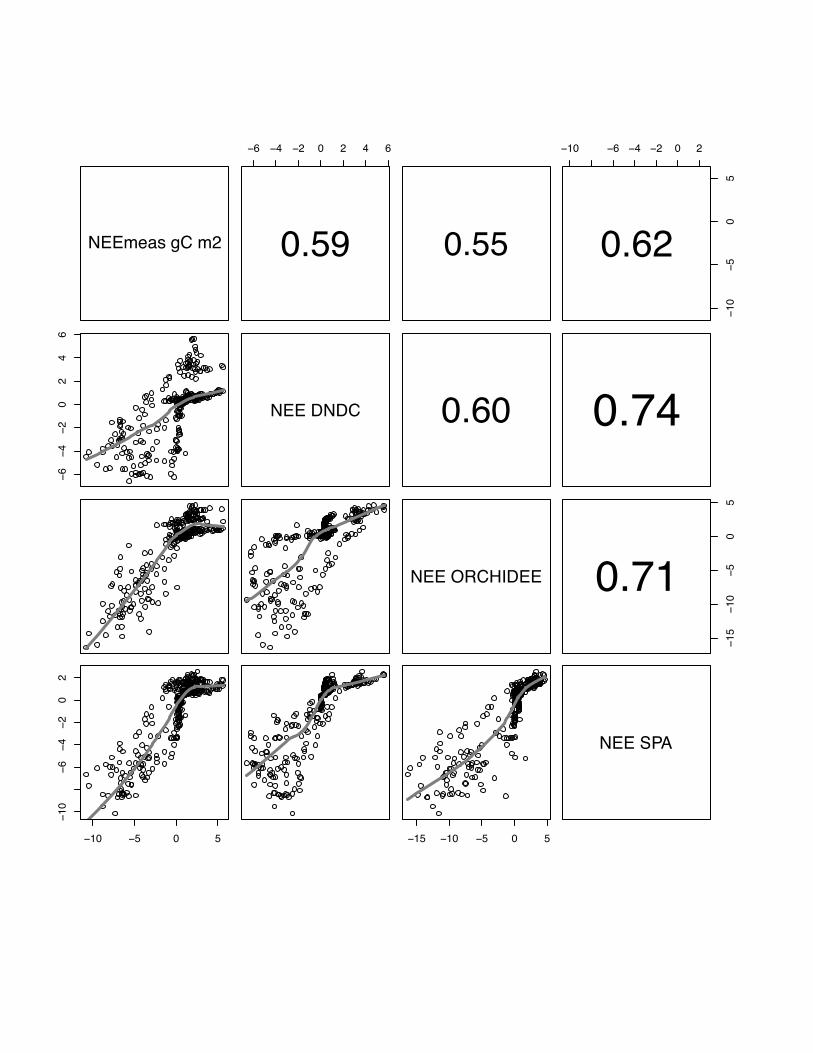
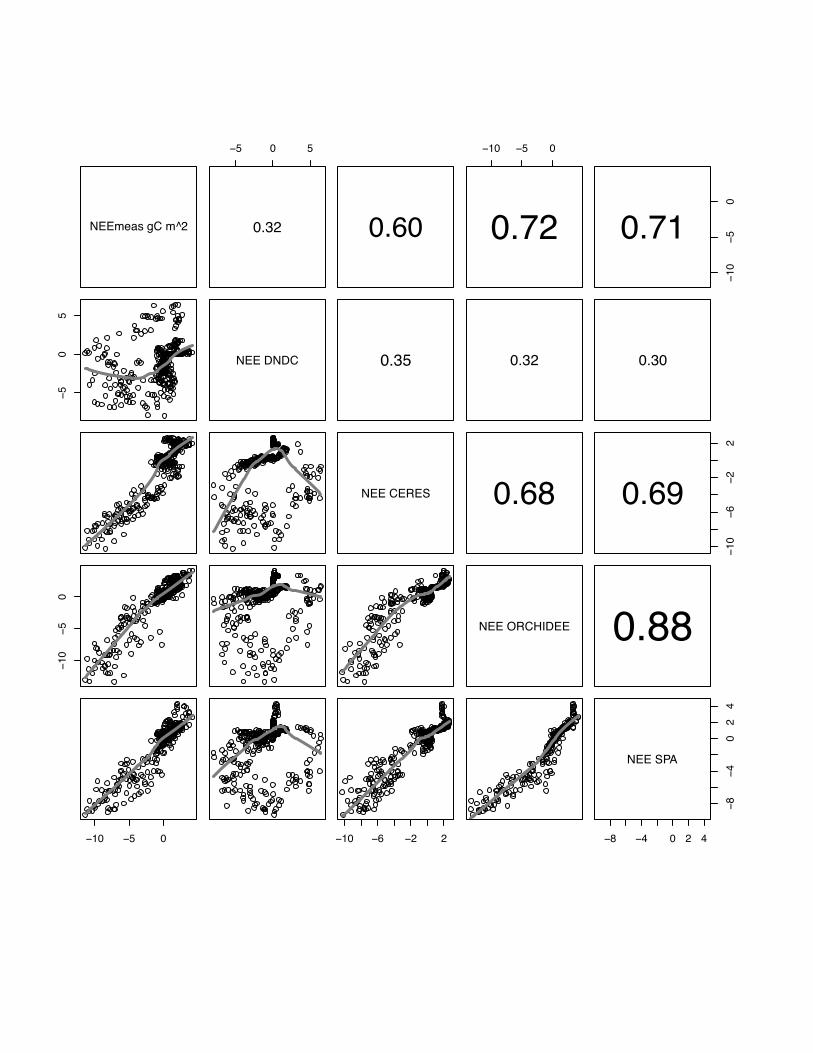
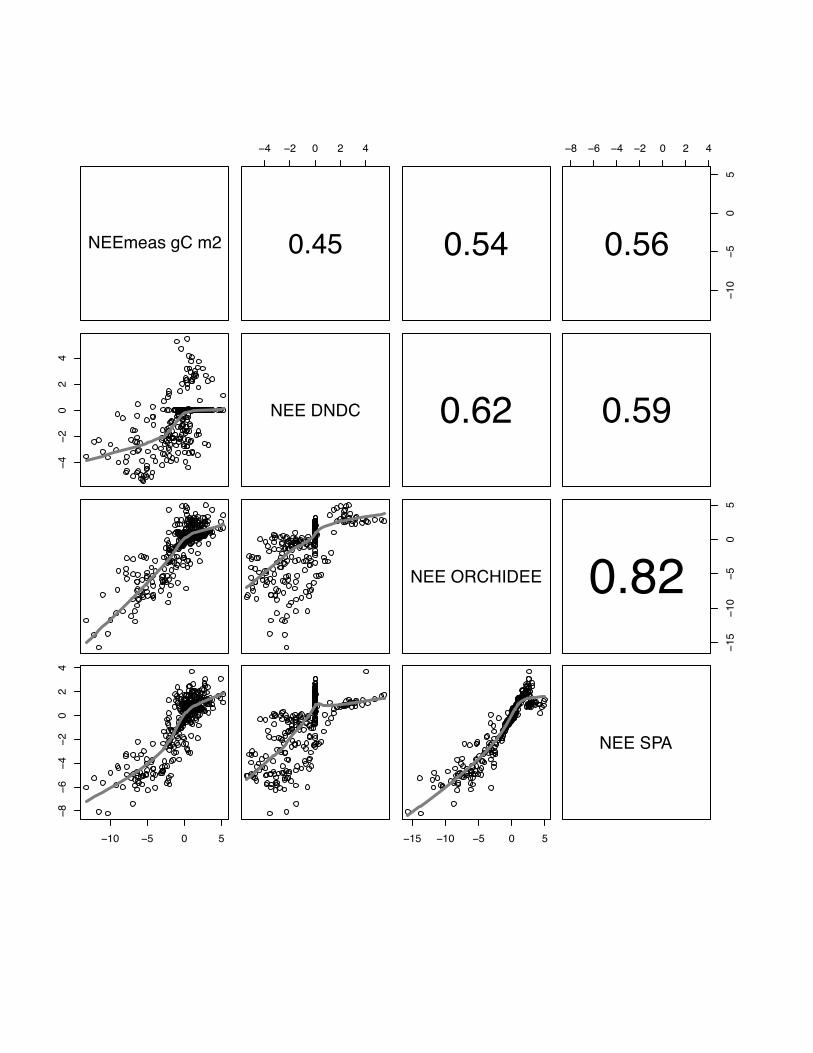
3.1 NEE
There is a wide range in the performance of the various models to reproduce the
measured NEE patterns at different sites and in different years, and also between
models at one site in the same year. Correlations range from r=0.28, p<0.05 for
DNDC simulating maize in year 2005 at Grignon to r=0.81, p<0.05 for the SPA model
simulating winter barley at Gebesee in 2007 (see table 3). In general, all models
perform better for simulating winter crops (site mean: E=56, r=0.61) than the summer
maize crop at Grignon (site mean: E=0.46, r=0.42).
In general, there is a better inter-model agreement between CERES-EGC,
ORCHIDEE-STICS and SPA than between any of these models and DNDC. The only
exception is the Klingenberg site where we observe a slightly better agreement
between SPA and DNDC than between ORCHIDEE-STICS and DNDC. If we test
average correlation of all models per site, models perform on average best at Gebesee
with winter barley in 2007 (r=0.68), followed by Aurade (r=0.65), Klingenberg
(r=0.61), Girgnon 2006 (r=0.58), Oensingen (r=0.52) and maize crop in Grignon in
2005 (r=0.42). Even though these results are relatively satisfying, all models expose a
22

weak and inconsistent performance at all sites for low fluxes in the range of -2 gC m2
day-1 to 2 g C m2 day-1 .
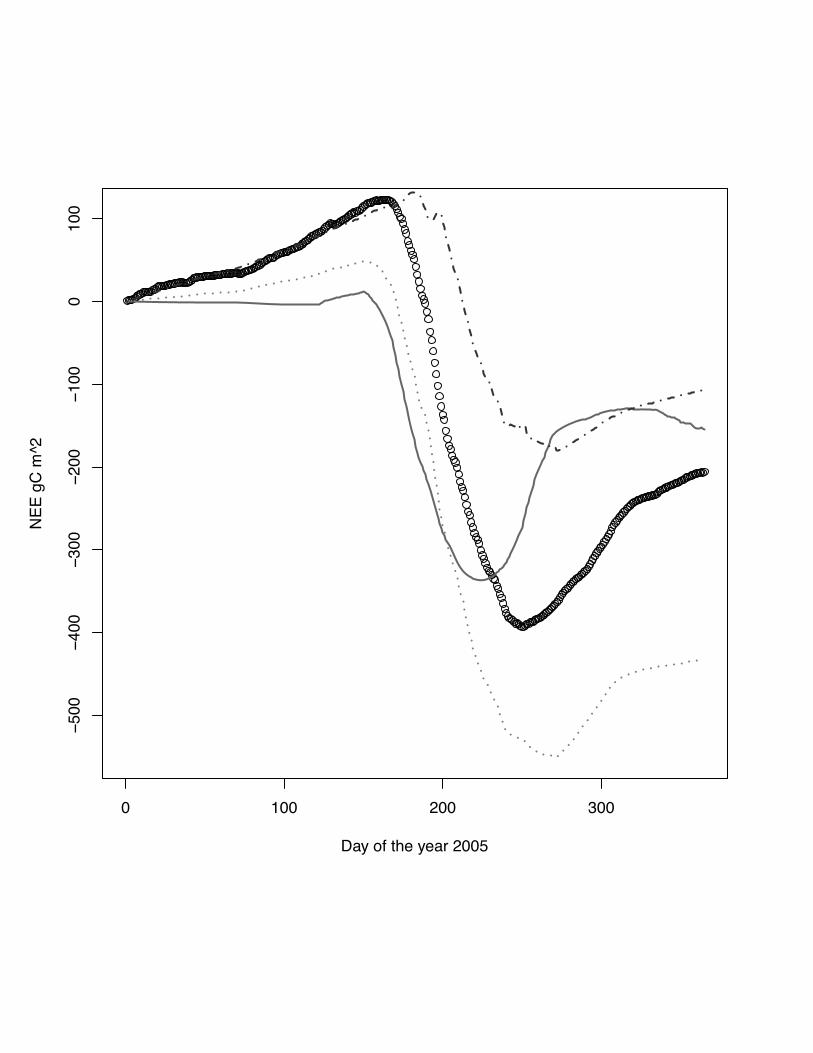
If we compare cumulative NEE fluxes (even-numbered fig. 2-12), we observe a
mismatch for all models in the early stage of the growing season, when low fluxes are
predominate. The DNDC model shows a stronger divergence from measurements
compared to the other models in the first 100 days of the year (DOY). The
ORCHIDEE-STICS, SPA and CERES-ECG models start with very similar trajectories
but begin to diverge between DOY 100 and 200 at most of the sites, except for maize
at Grignon in 2005. A common pattern for the three models is to overestimate the
NEE peak and the failure to reproduce the senescence and post harvest fluxes. This
leads to a mismatch of the cumulative NEE for the year. In general, the SPA model
shows the best performance in terms of NEE for the sites even though deficiencies
remain in reproducing peak and post-harvest NEE fluxes.
PLEASE INSERT FIG. 1 to 12
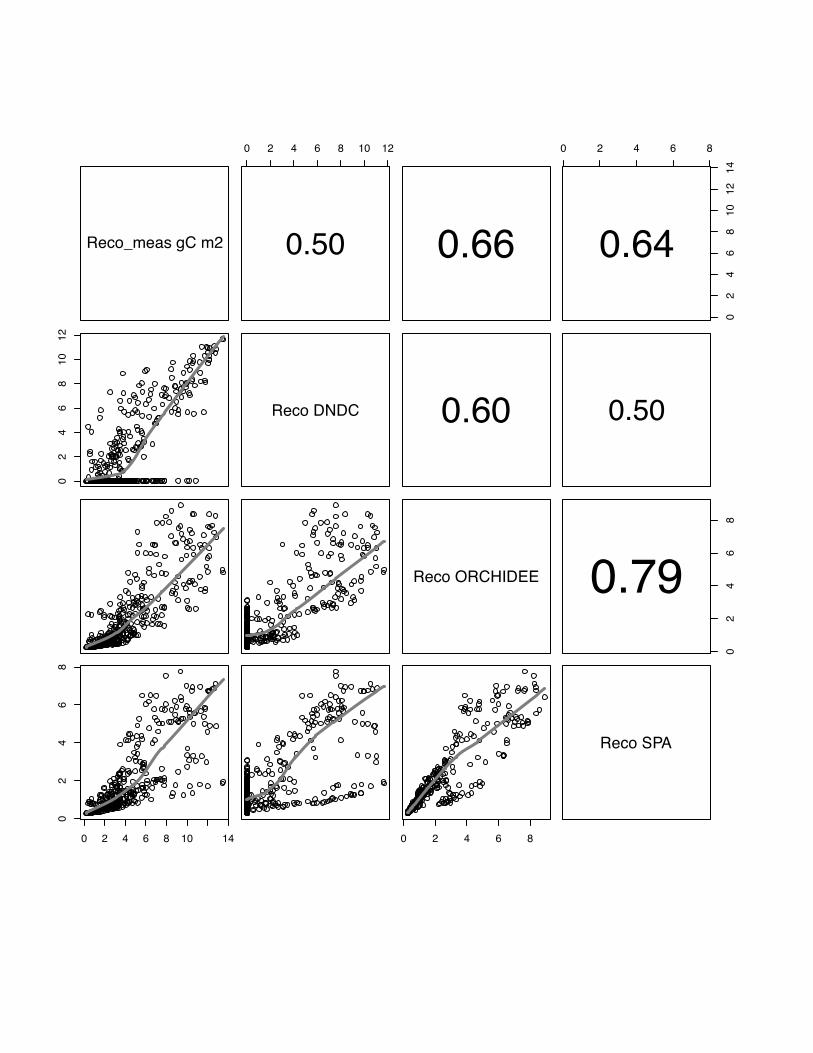
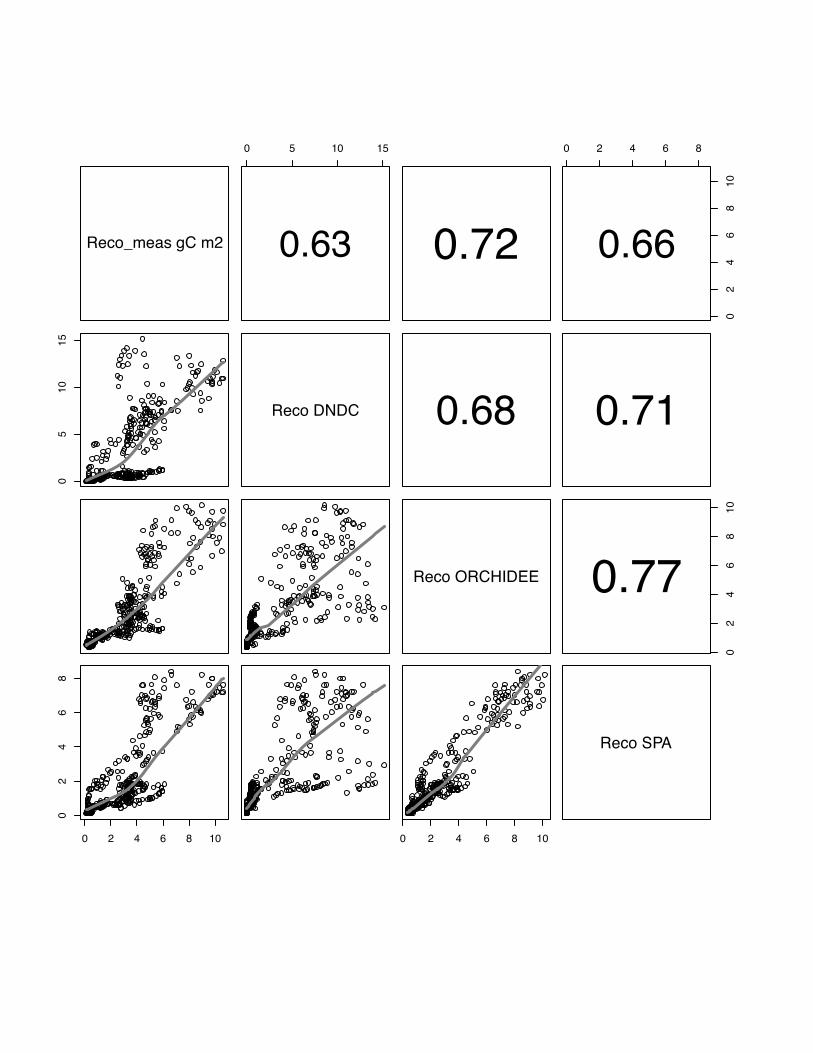
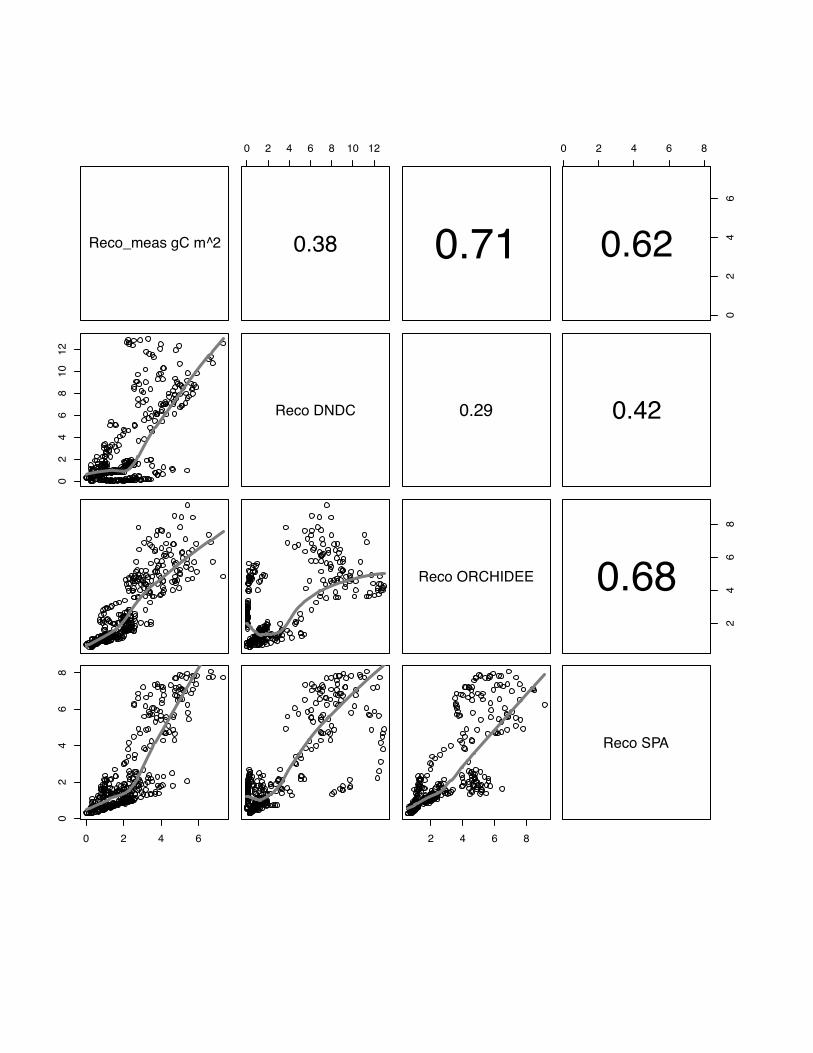
3.3 Reco
The ranking the models according to their association with the data shows
ORCHIDEE-STICS with the best performance on average over all sites (model mean:
r=0.72, p<0.05). This model is also consistent in its performance over all sites with
comparable r values (table 3). Between models the r values vary strongly from r=0.38
for DNDC in Aurade to r=0.79 (p<0.05) for ORCHIDEE-STICS at Klingenberg in
2006. Similar as for the simulation of NEE, we can identify the highest significant
agreement between SPA and ORCHIDEE-STICS (Note: CERES-EGC does not
23

simulate Reco), and a lower association between these two models and DNDC.
However, the pattern with contrasting behaviour of all models for fluxes below 2 g C
m2 is again clear (Fig. 13, 14, 15), and a considerable scatter is seen for high flux
rates. This is also reflected in low mean model efficiency values: E=-1.02, E=0.44 and
E=0.33 for DNDC, ORCHIDEE-STICS and SPA, respectively.
PLEASE INSERT FIG 13,14,15
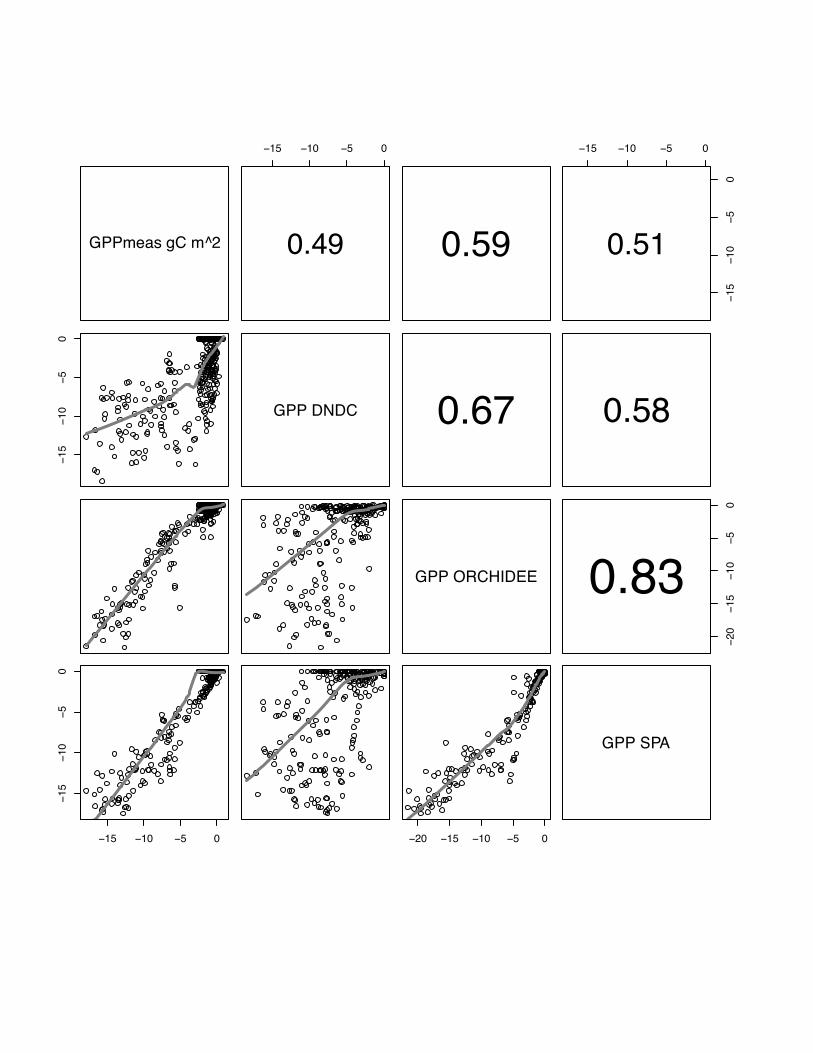
3.2 GPP
All models simulate GPP with association r comparable to performance (mean
r=0.85) when simulating NEE, with correlation coefficient that varies from r=0.43
(ORCHIDEE at Oensingen) to r=0.75 (SPA at Gebesee) (p<0.05) (see fig 16, 17 and
18).
Overall the models agree better with each other (mean between models all sites
r=0.68) than they associate to the measurements (mean over all sites measurement
model r=0.58). In this respect, ORCHIDEE-STICS and SPA are the two models that
relate the most (mean r = X), followed by SPA and DNDC (mean r=X). Last, DNDC
and ORCHIDEE-STICS models disagree the most (mean r=X). The good agreement
between ORCHIDEE-STICS and SPA. In this case we again observe consistent
behaviour with a correlation of r=0.83 between the ORCHIDEE-STICS and SPA
model. Thus, the scatter between the two models is lower than in their respective
reproduction of the measured values which suggests a common reason for the misfit
with the data.
24

Model-data comparisons exhibit a strong mismatch for low fluxes as indicated by a
similar and almost ‘flat’ shape of the regression line for the comparison with low
measured values (see Fig. 16-18). As already observed r score, there is a better
agreement for these low fluxes between ORCHIDEE-STICS and SPA than between
each of the two models and the measurements.
Overall model-data agreement as evaluated by the Nash-Sutcliffe E is relatively high,
being generally greater than 0.55. It is minimal for DNDC at Grignon in 2006
(E=0.31) and goes up to 0.80 for three model-site combinations (ORCHIDEE-SITCS
and SPA at Grignon 2006 and SPA at Gebesee).
PLEASE INSERT FIG 16, 17, 18
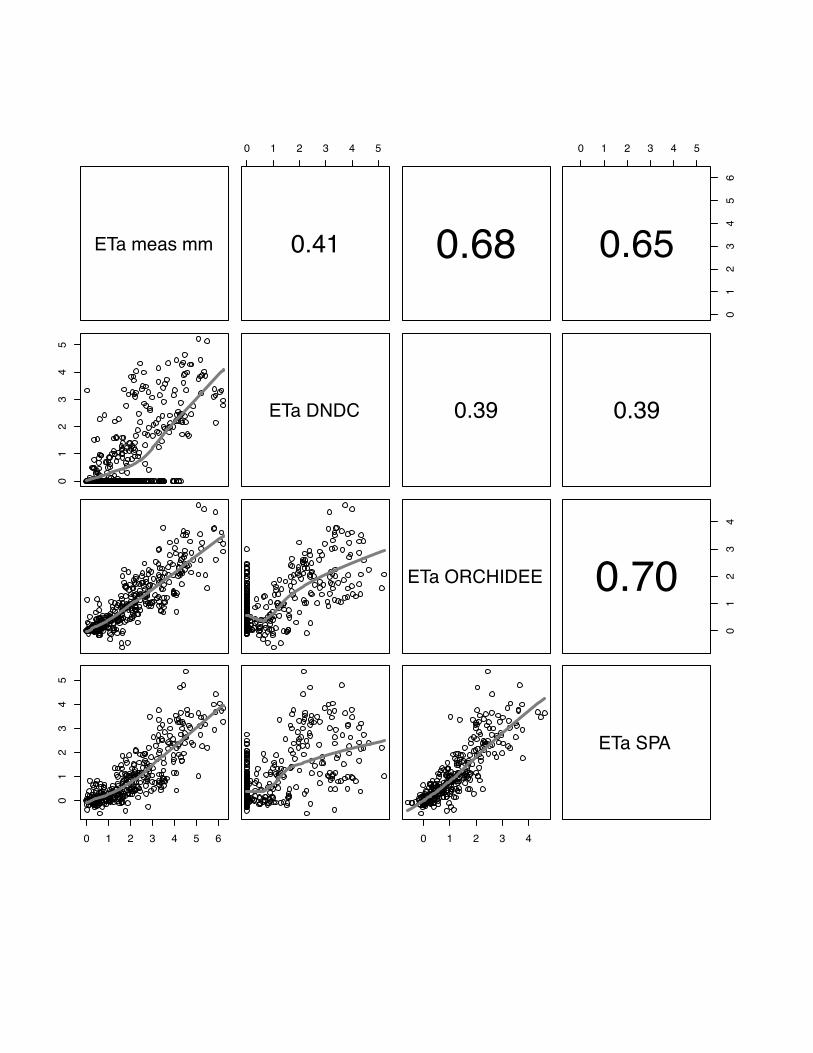
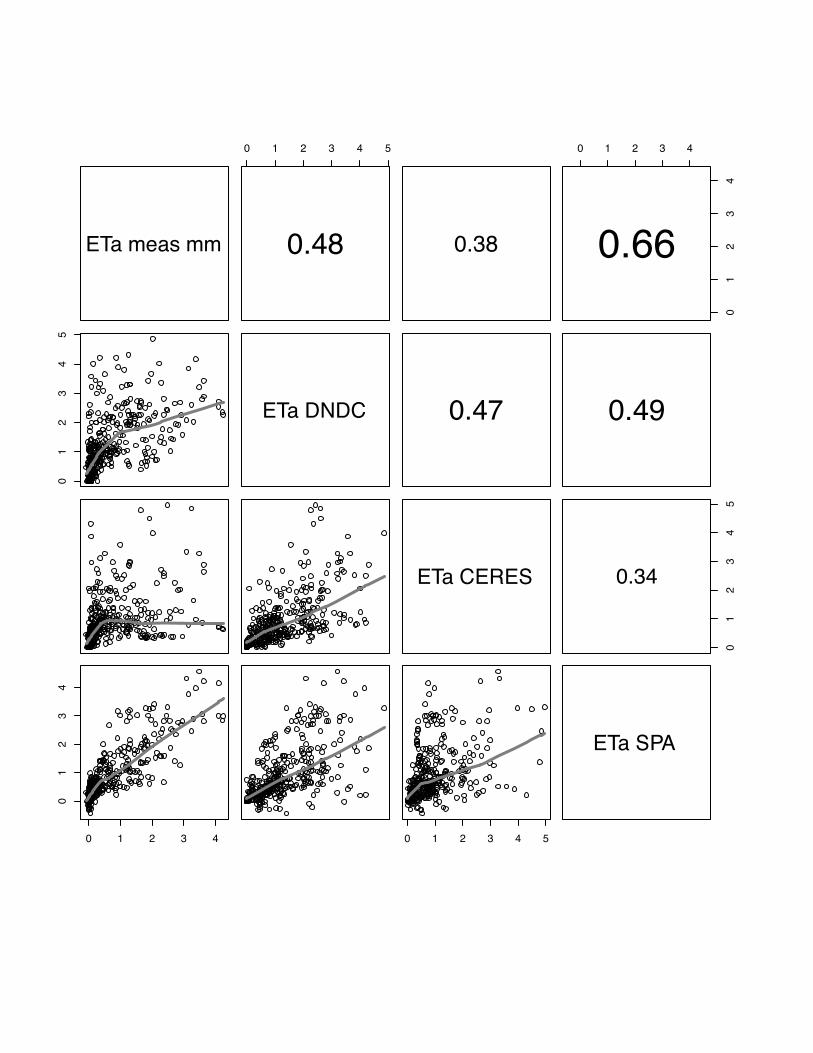
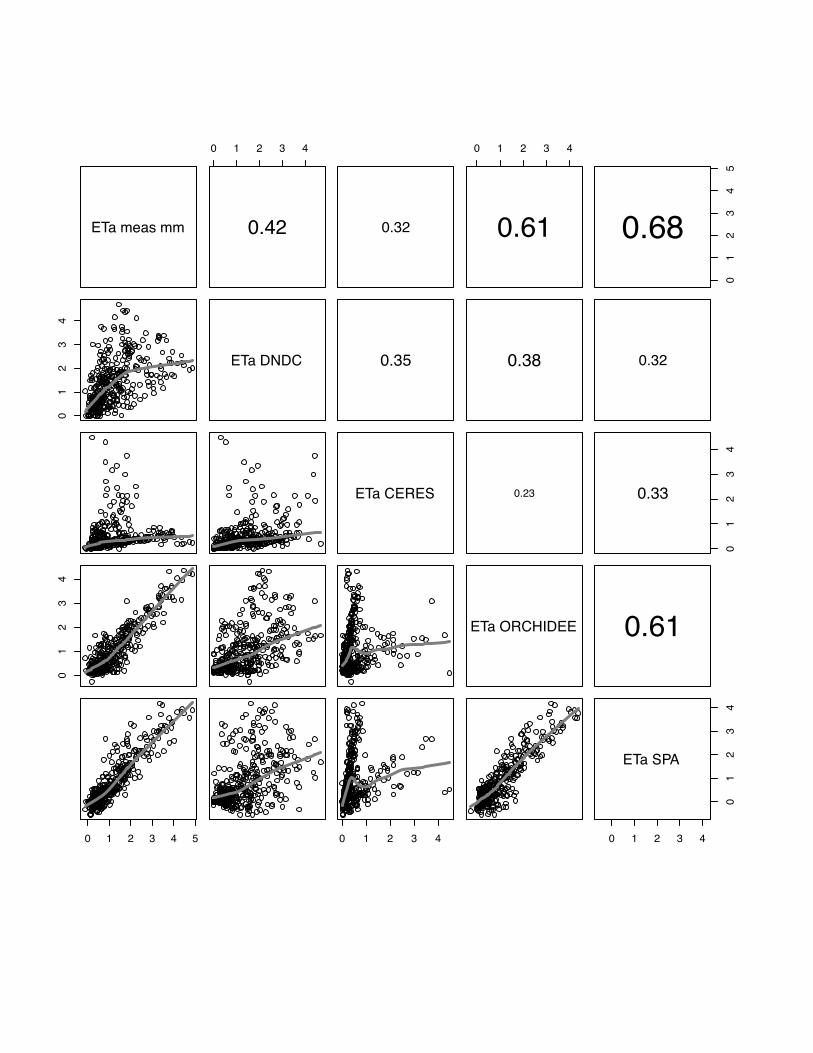
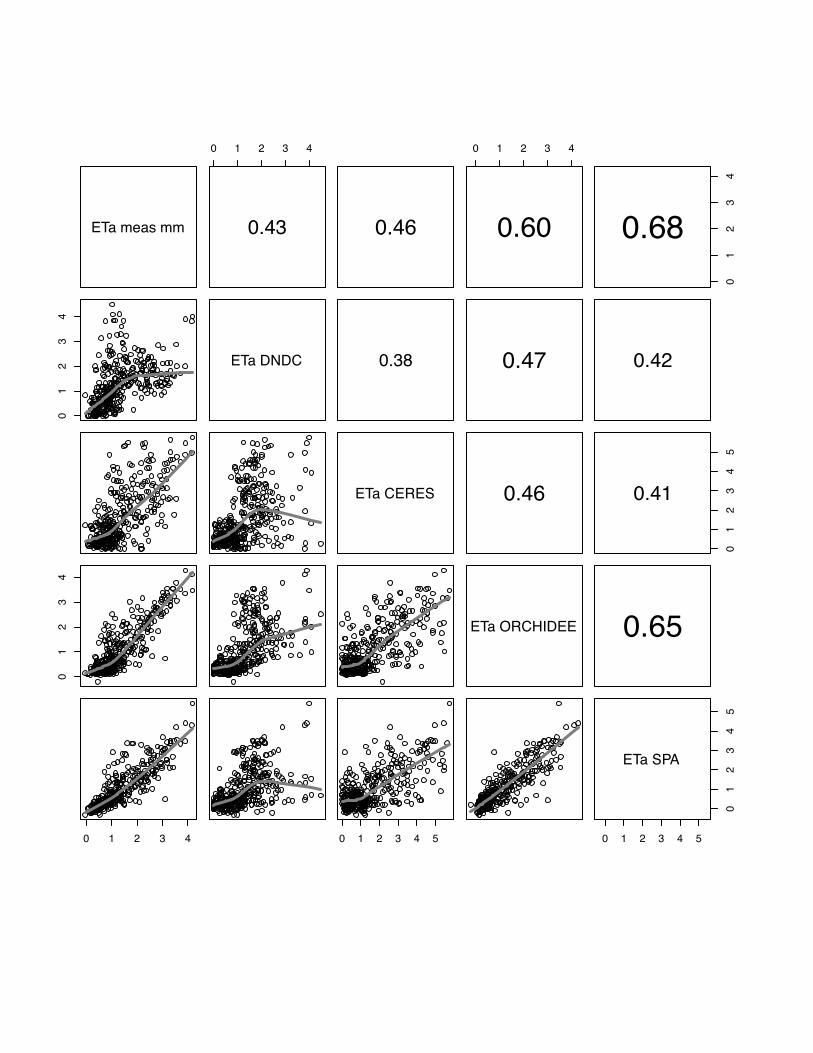
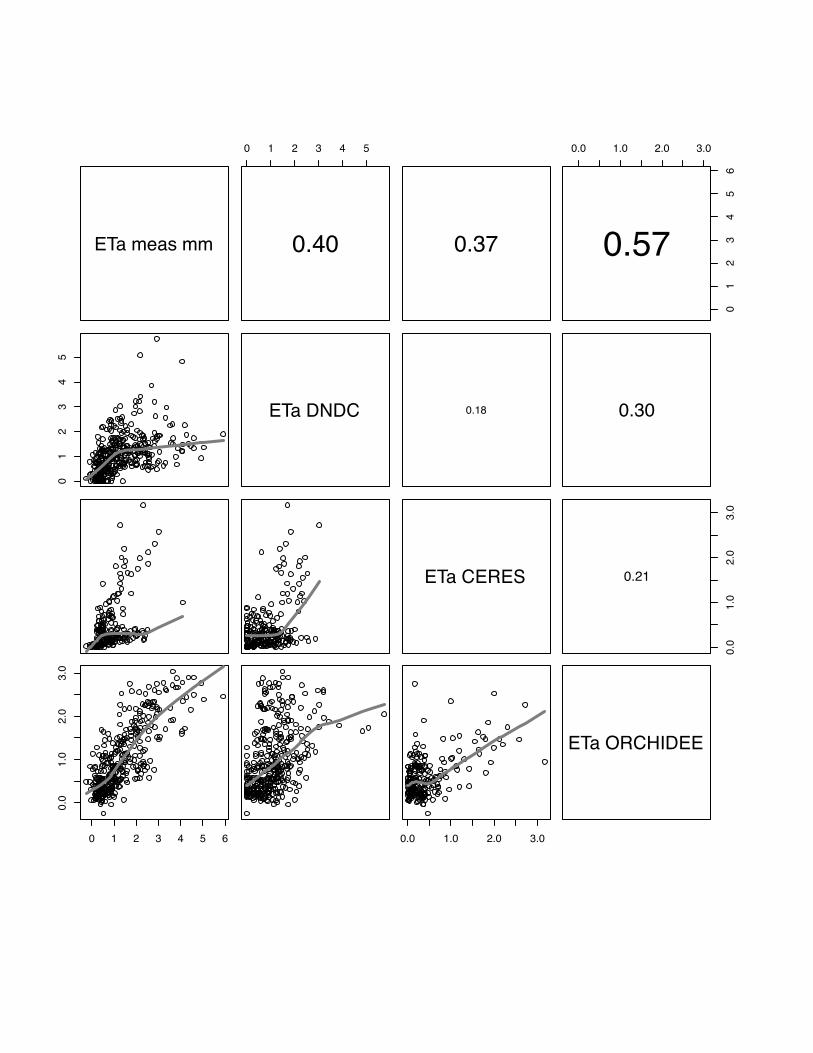
3.4 ETa
The comparison of observed and modelled evapotranspiration fluxes generally shows
that ORCHIDEE-STICS and SPA show highest correlations with measurements
(mean r for all sites: 0.60 for ORCHIDEE-STICS, 0.69 for SPA). Again, these models
also exhibit a considerable degree of cross-correlation, indicating a general agreement
in the response to driving variables. In relation to that, the accuracy of modelled ETa
as simulated by CERES-EGC (mean r: 0.38) and DNDC (mean r: 0.45) is
considerably lower. For CERES-EGC and DNDC, the relationship between modelled
and observed ETa, expressed by the slope of the lowess fit curve, is changing around
a value of 1 mm d-1. Beyond this value, the fit curve gradually takes over a quasi-
logarithmic behaviour for most scatter plots. For certain sites (e.g. both Grignon
25

years), DNDC and CERES-EGC fail to reproduce high flux rates, indicated by a high
degree of scattering around the fit curve. The correlation of modelled values is high
between DNDC and CERE-EGC, and even higher than with the observations for the
Gebesee site.
We found considerably low values of model efficiency for DNDC and CERES-EGC,
which are negative for Grignon 2006, Gebesee and Klingenberg. For SPA and
ORCHIDEE-STICS, E ranges from 0.2 (SPA at Oensingen) to 0.78 (ORCHIDEE-
STICS at Grignon 2006). Thus, the overall efficiency of these two models is higher
and broadly satisfying.
In general, lower evapotranspiration fluxes were captured with higher accuracy by all
models, which is indicated by an increasing data spread beyond 1 mm d-1 in most of
the scattering plots (fig 19-23).
PLEASE INSERT FIG 19 to 23
4. Discussion
The aim of the paper was to test four models for accuracy in simulating the main
components of the carbon cycle, net ecosystem exchange (NEE), ecosystem
respiration (Reco) and gross primary production (GPP) in connection with the actual
evapo-transpiration (ETa) on a daily time scale over a gradient of environmental
conditions in Europe. The results show a heterogeneous picture, with strong
differences between models and between sites. The two models with the highest
accuracy over all sites when simulating daily NEE and latent heat are SPA and
ORCHIDEE-STICS. Both models run on a half hourly time-step with a high process
26

resolution, so they are able to capture the diurnal variability of processes leading to a
high level of agreement when measurements are aggregated to daily fluxes. They are
strong, in particular, in simulating water fluxes where they clearly out-perform the
two other models. However, the models are not able to simulate crop rotations due to
the limited number crops parameterised, and their current limitation to the growing
period of the crop. Consequently they cannot produce accurate annual cumulative
NEE fluxes in Grignon, Klingenberg, Gebesee, and in case of ORCHIDEE-STICS in
Oensingen. However, even though the model are limited to winter wheat, winter
barley for SPA, and winter wheat and maize for ORCHIDEE-STICS, they are already
able to capture 43% and 37%, respectively, of the crop area of the EU27 (Kutsch et al.
this issue). Through their ability to simulate these three crops between them, the two
models cover more than 50% of the EU27. However, for the purposes of examining a
wide range of mitigation options envisaged for agriculture in the context of climate
change {IPCC, 2007 #761; Smith, 2008 #735}, the models need to be extended to
simulate a number of post-harvest activities, like catch and cover crops, and crop
management options like low tillage and non-tillage systems etc.
On the other hand, the CERES and DNDC models are both less accurate in their
representation of daily NEE. In the model comparison presented here DNDC fails to
reproduce NEE in Grignon, and exhibits a poor performance at Oensingen. The main
factor leading to the failure of DNDC at the Grignon site in 2006 was the particularly
mild winter, with temperatures rising above +10°C in the first 100 days of the year.
The DNDC model seems to be more sensitive to these temperatures than the other
models. DNDC responds with immediate growth which leads to a strong
overestimation of early GPP and to early senescence because of its, in contrast to the
27

other models, temperature sum based phenology, which causes the strong phase
mismatch with the measurement and with the other models. Another important factor
determining the accuracy of the DNDC model is the lack of consideration of
germination/emergence in the model, which leads to unrealistic growth in the early
stage of the growing season. Both factors lead to a relatively poor performance of the
model when simulating NEE in particular.
The lack-of-fit for the simulation of water fluxes for DNDC can be explained by the
models use of the Thornthwaite formula {Thornthwaite, 1965 #503} to calculate
evapo-transpiration. The Thornthwaite formula is known to underestimate potential
latent heat flux, especially under arid and semiarid conditions {Amatya, 1995 #857,
McKenney, 1993 #858}. This explains the lack-of-fit for the all sites except
Oensingen, which has the highest precipitation combined with a relatively low mean
annual temperature. There are also reported problems with the hydrological
component in DNDC, especially regarding the calculation of water filled pore space
in the soil {Tonitto, 2007 #787; Tonitto, 2007 #788}.
However, there are a number of factors that limited the performance of DNDC in
comparison to the other models. The first and most important point when compared to
the best performing models, SPA and ORCHIDEE-STICS, is the simulation of the full
crop rotations at all sites. Where SPA and ORCHIDEE-STICS were initialised to
determine the initial conditions only for the year of the simulation, the DNDC model
had to use the conditions related to the previous crops. The model also considers all
management options applied at the sites including manure applications, tillage and the
growth of cover crops. The level of model complexity for this part has also a profound
28

impact on the uncertainties associated with the simulations which also increases the
chance of poorer model fit to measurements.
In the case of CERES, we see a comparable performance to ORCHIDEE-STICS and
SPA when we compare the simulated to measured NEE. However, the model was
calibrated for the sites in Grignon and Aurade, (Lehuger et al., this issue), limiting the
degrees of freedom when simulating fluxes at these sites. If we compare the
performance for the un-calibrated site in Gebesee though, the performance is still
good for the simulation of NEE.
In general the results are in agreement with other studies examining carbon fluxes. A
direct model comparison of simulation of carbon and water fluxes at forest sites of the
EUROFLUX network, including the two model classes present in this study
(biogeochemical and dynamic global vegetation models), showed very similar results
{Morales, 2005 #531}. In that study, great variance of model accuracy across all sites
was also found. A main problem was the representation of water fluxes in the
Mediterranean region, where models performed poorly. We observe that especially
DNDC has a low performance at all sites, except for Oensingen, but also for CERES-
EGC exhibits problems at the site Grignon in 2005 and 2006 as well as Gebesee, and
in terms of model efficiency at Aurade in 2006.
Why do models fail to simulate low fluxes?
The lack of accuracy when simulating carbon fluxes close to zero is the most
prevalent effect observed in this model comparison across sites and across models.
29

There are two possible explanations. The first is that the concepts implemented in the
models under comparison are not adequate to simulate the underlying processes. The
second is that the measured fluxes are unrealistic due to the inherent properties of
uncertainty of eddy covariance measurements, such as u* correction, gap filling,
outlier filtering, advection and flux partitioning etc. {Lasslop, 2008 #748; Hollinger,
2005 #828; Richardson, 2005 #829; Osborne, this issue #865; Smith, this issue #866;
Reichstein, 2005 #816}. The results show that the models fail to reproduce low fluxes
in all flux components namely NEE, GPP and Reco. It is also very important to note
that the models are inconsistent in how they stimulate the flux, but still show partial
consistency when directly compared to each other (this is especially true for
ORCHIDEE-STICS and SPA). The measurements and models approach the same
quantity from completely different directions. The entity measured in eddy covariance
systems is NEE. The values for GPP and Reco are derived from NEE using flux
partitioning {Reichstein, 2005 #816} which uses measured weather data as drivers.
The models, on the other hand, use the same weather data as drivers to simulate GPP
and Reco to calculate NEE. So given the same driving variables, we arrive at different
results for these low fluxes. If we look at the results of GPP and Reco simulations, we
find a generally higher agreement for fluxes below -2 gC m-2 day-1 and above 2 gC m-2
day-1, respectively, which indicates that the underlying assumptions for these fluxes
are consistent. The time when these low fluxes appear are the early growing stages of
the crop or the intercropping period, which might hold one key to answer the question
about the inconsistency. In this part of the growing season, NEE fluxes are dominated
by heterotrophic respiration of soil organic matter and litter carbon. This respiration
flux is clearly higher than photosynthesis by the small and slowly growing crop
biomass. As a consequence, initial values of soil organic matter, but more importantly
30

litter carbon, largely determine the overall match of modelled vs observed NEE in the
early growth phase. For example, we found clearly improved SPA results for early
season low NEE fluxes when increasing initial litter carbon content (from 0 gC m-2,
the initial litter carbon content for all runs shown here, to 200 gC m-2, results not
shown). ORCHIDEE-STICS and SPA, the models with the highest agreement
between each other for the low fluxes, are the two models that only simulate the
cropping season with a limited implementation of management action, which can
explain the mismatch. On the other hand DNDC shows a good performance in
simulating GPP, and in some cases Reco, but was not able to adequately reproduce the
temporal development of the crop, which in turn caused a lack of alignment between
the peaks in GPP and Reco. Because NEE is calculated as the difference of the two, the
errors amplify, especially at the end of the growing season when the model simulates
senescence, reducing GPP, which in parallel increases the ecosystem respiration due
to a higher carbon input into the soil. However, the inherent properties of eddy
covariance measurements also have effects on the model comparison. There are a
number of papers discussing the influence of different assumptions on aggregated
NEE fluxes at agricultural sites {Anthoni, 2004 #807; Aubinet, 2008 #808;
Moureaux, 2008 #859}, which show a considerable systematic uncertainly in the flux
estimates varying from 10 g C m-2 y-1 to 40 g C m-2 y-1. Estimates of NEE at Gebesee
in 2003 ranged from 185 g C m" -2 y-1 to 245 g C m" -2 y-1 , based on different
assumptions in the data processing {Anthoni, 2004 #807}.
Especially for low fluxes, as discussed here, with low photosynthetic activity in the
early, late or inter growing season, the flux measurements at night, when turbulent
mixing is insufficient are becoming over proportional important, because they are also
used to derive daytime Reco . These data need also to be gap filled in order to be used
31

to calculate daily means. The error introduced by gap filling is as follows: at daytime,
maximum observed errors were ±0.20 g C m 2" , and at night time the maximum was
±0.14 g C m 2 " per percentage of day filled {Falge, 2001 #549}. The percentage of
gap-filled half hourly data range from as low as 29.2% for 302 days at Gebesee 2005
up to 58.9% for 347 days at Klingenberg 2005 which could explain the inconsistent
reproduction of these fluxes by the models.
5. Conclusions
Overall, the models tested in this study show an acceptable to very good performance
when simulating NEE with significant associations and efficiencies above zero,
beside DNDC in Grignon in 2006. All models have problems in reproducing fluxes
between 2 and -2 g C m2 day-1, most probably due to a combination of a lack of
accuracy in simulating the correct temporal sequence of development stages,
problems to capture the ecosystem respiration flux and not considering all
management actions.
For European scale application, models likes SPA and ORCHIDEE-STICS are highly
accurate in simulating net carbon fluxes (NEE) and water fluxes. However, they are
only able to simulate the seasonal carbon balance of a limited number of crops with a
limited consideration of management. Thus they are not yet capable of evaluating the
wide range of mitigation options envisaged for agriculture in the context of climate
change {IPCC, 2007 #761; Smith, 2008 #735}. They are also not able to simulate
accurately annual carbon budgets because they do not consider post-harvest effects
(such as intercrop or re-growth).
32

On the other hand models like CERES-EGC and DNDC are less accurate in
simulating NEE and especially poor in reproducing latent heat fluxes. They are
however capable of simulating mitigation options because they can simulate full crop
rotations and associated management effects. CERES-EGC and DNDC also consider
other greenhouse gases like N2O, and in case of DNDC and CH4, that might affect the
GHG balance. DNDC has, in this case, the widest range of crop/management options
under irrigated and non-irrigated conditions {Leip, 2008 #716} but shows the lowest
accuracy in the carbon daily flux simulations. Both models, however, fail to
accurately present the associated water fluxes, and this might limit their ability to
simulate possible drought effects, which will have an increasing importance in future
due to climate change {Seneviratne, 2002 #851; Salinger, 2005 #852; Falloon #853}.
Finally, this model comparison shows that multiply constrained model evaluation is
clearly improved with high temporal resolution data sources, such as those from eddy
flux data, in combination with detailed management information, such as that
provided at the CarboEurope IP sites.
6. Acknowledgements
Pete Smith is a Royal Society-Wolfson Research Merit Award holder. The
CarboEurope IP project was funded by the EU’s Sixth Framework Programme for
Research and Technological Development. The work of the paper contributed to the
CC-Tame, Carbo-Extreme and NitroEurope IP project.
33

7. References
34
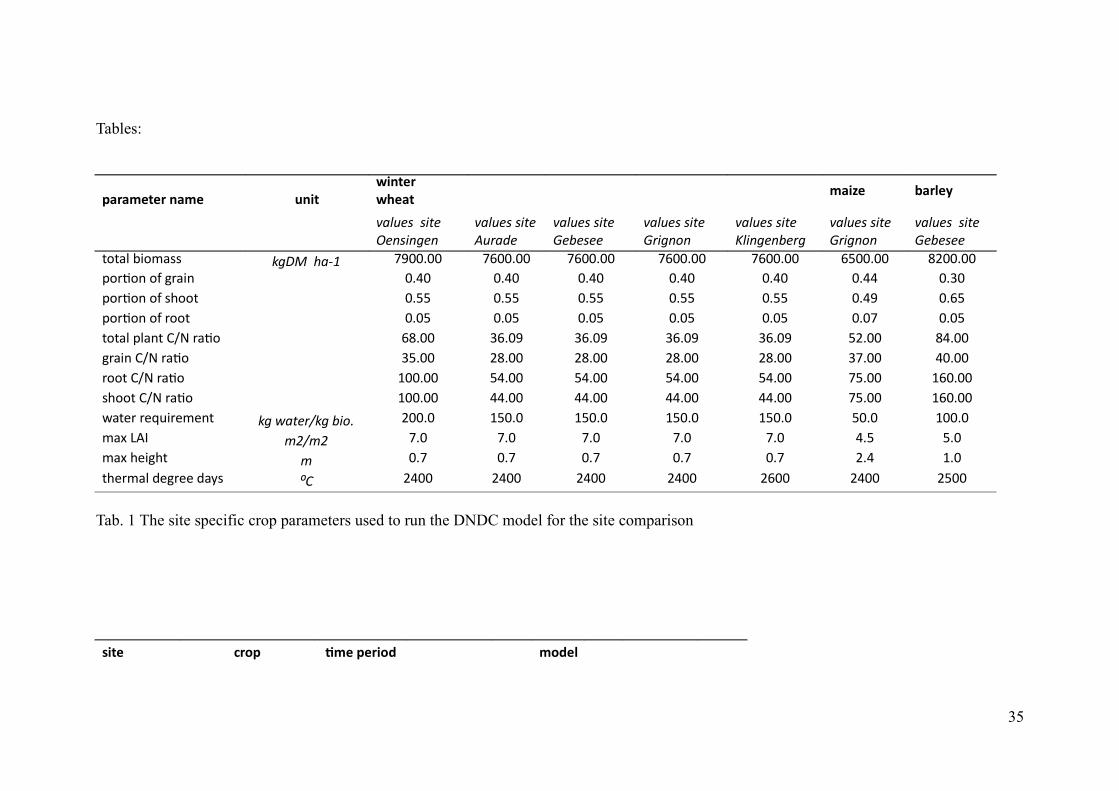
Tables:
."/"+&1&/!,"+& 3,)14),1&/!4(&"1 ! ! ! !
+")7& #"/*&6
! !B1:A5?!!?8@5 !
,5<?8<65<!
B1:A5?!?8@5 !
"A>145
B1:A5?!?8@5 !
&525?55
B1:A5?!?8@5 !
&>86<=<
B1:A5?!?8@5 !
):8<65<25>6
B1:A5?!?8@5 !
&>86<=<
B1:A5?!!?8@5 !
&525?55
C>C2;!3:><2BB! 96$*!!71HI UWNNKNN UTNNKNN UTNNKNN UTNNKNN UTNNKNN TSNNKNN VPNNKNN?>AD>=!>7!8A2:=! NKRN NKRN NKRN NKRN NKRN NKRR NKQN?>AD>=!>7!B9>>C NKSS NKSS NKSS NKSS NKSS NKRW NKTS?>AD>=!>7!A>>C NKNS NKNS NKNS NKNS NKNS NKNU NKNSC>C2;!?;2=C!#L,!A2D> TVKNN QTKNW QTKNW QTKNW QTKNW SPKNN VRKNN8A2:=!#L,!A2D> QSKNN PVKNN PVKNN PVKNN PVKNN QUKNN RNKNNA>>C!#L,!A2D> ONNKNN SRKNN SRKNN SRKNN SRKNN USKNN OTNKNNB9>>C!#L,!A2D> ONNKNN RRKNN RRKNN RRKNN RRKNN USKNN OTNKNNF2C6A!A6@E:A6<6=C 96!C1@5>G96!28=F PNNKN OSNKN OSNKN OSNKN OSNKN SNKN ONNKN<2G!*"( ;JG;J UKN UKN UKN UKN UKN RKS SKN<2G!96:89C ; NKU NKU NKU NKU NKU PKR OKNC96A<2;!568A66!52HB K# PRNN PRNN PRNN PRNN PTNN PRNN PSNN
Tab. 1 The site specific crop parameters used to run the DNDC model for the site comparison
0)1&! $/-. 2+&!.&/)-% +-%&*
35
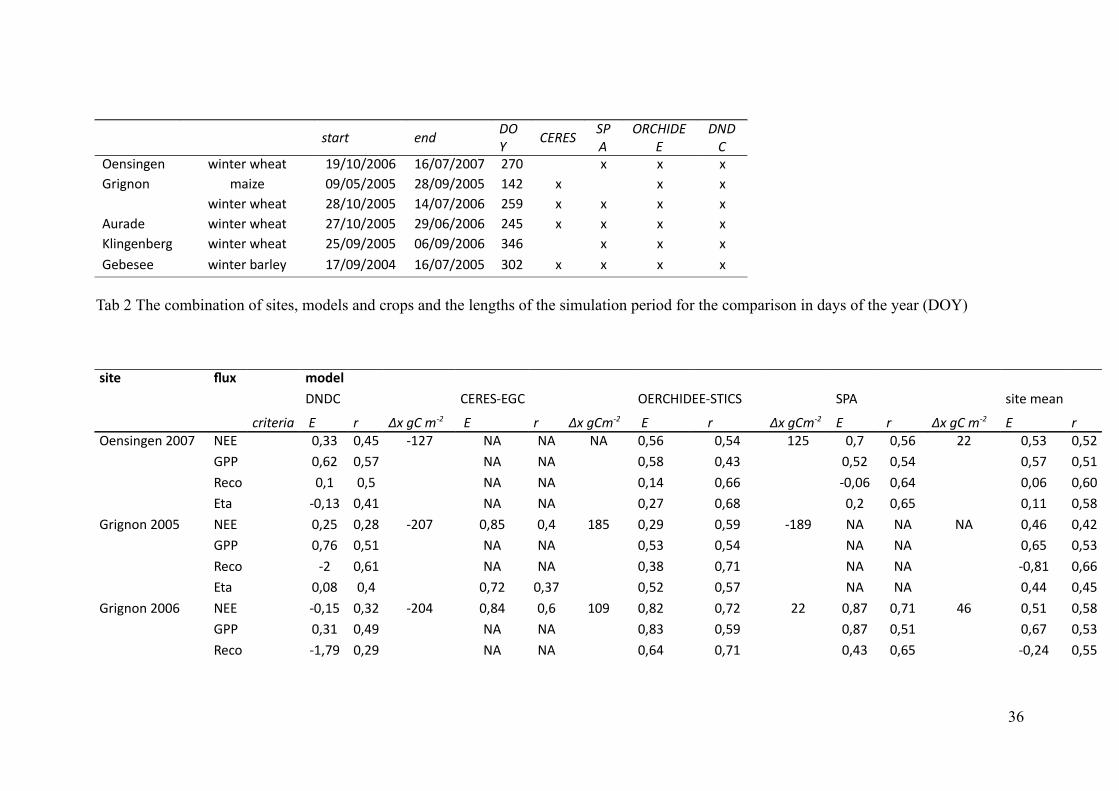
! ! ?@1>@ 5<4$,
0#%.%/
/-
"
,.#'($%
%
$+$
#
-6=B:=86= F:=C6A!F962C OWLONLPNNT OTLNULPNNU PUN G G G&A:8=>= <2:I6 NWLNSLPNNS PVLNWLPNNS ORP G G G
F:=C6A!F962C PVLONLPNNS ORLNULPNNT PSW G G G G"EA256 F:=C6A!F962C PULONLPNNS PWLNTLPNNT PRS G G G G);:=86=36A8 F:=C6A!F962C PSLNWLPNNS NTLNWLPNNT QRT G G G&636B66 F:=C6A!32A;6H OULNWLPNNR OTLNULPNNS QNP G G G G
Tab 2 The combination of sites, models and crops and the lengths of the simulation period for the comparison in days of the year (DOY)
0)1& '35 ! +-%&* ! ! ! ! ! ! ! ! ! ! ! ! !
$,$# #%/%0M%&# -%/#'($%%M01(#0 0." B:C6!<62=! ! 3>8@5>81 !% >! ED!6#!;HJ !% > ED!6#;HJ !% !!!!> ED!6#;HJ % !> ED!6#!;HJ % >
-6=B:=86=!PNNU ,%% NJQQ NJRS MOPU ," ," ," NJST NJSR OPS NJU NJST PP NJSQ NJSP&.. NJTP NJSU ," ," NJSV NJRQ NJSP NJSR NJSU NJSO
/64> NJO NJS ," ," NJOR NJTT MNJNT NJTR NJNT NJTN%C2 MNJOQ NJRO ," ," NJPU NJTV NJP NJTS NJOO NJSV
&A:8=>=!PNNS ,%% NJPS NJPV MPNU NJVS NJR OVS NJPW NJSW MOVW ," ," ," NJRT NJRP&.. NJUT NJSO ," ," NJSQ NJSR ," ," NJTS NJSQ
/64> MP NJTO ," ," NJQV NJUO ," ," MNJVO NJTT%C2 NJNV NJR NJUP NJQU NJSP NJSU ," ," NJRR NJRS
&A:8=>=!PNNT ,%% MNJOS NJQP MPNR NJVR NJT ONW NJVP NJUP PP NJVU NJUO RT NJSO NJSV&.. NJQO NJRW ," ," NJVQ NJSW NJVU NJSO NJTU NJSQ
/64> MOJUW NJPW ," ," NJTR NJUO NJRQ NJTS MNJPR NJSS
36
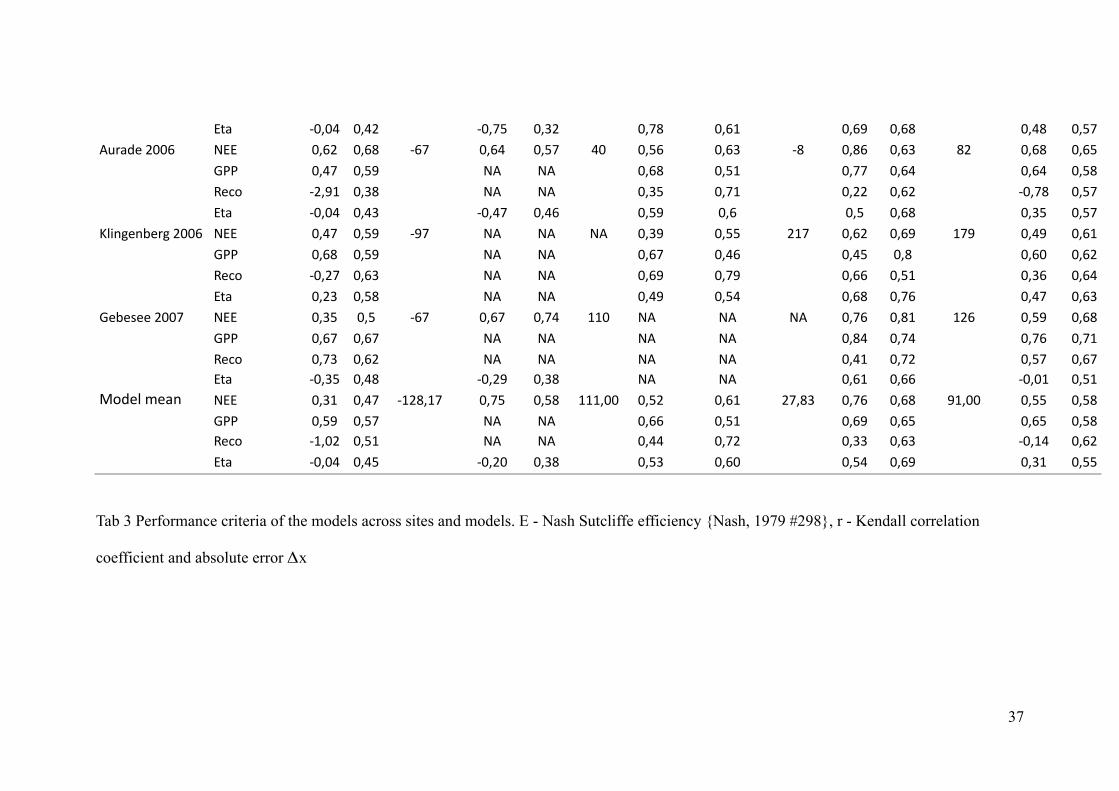
%C2 MNJNR NJRP MNJUS NJQP NJUV NJTO NJTW NJTV NJRV NJSU"EA256!PNNT! ,%% NJTP NJTV MTU NJTR NJSU RN NJST NJTQ MV NJVT NJTQ VP NJTV NJTS
&.. NJRU NJSW ," ," NJTV NJSO NJUU NJTR NJTR NJSV/64> MPJWO NJQV ," ," NJQS NJUO NJPP NJTP MNJUV NJSU
%C2 MNJNR NJRQ MNJRU NJRT NJSW NJT NJS NJTV NJQS NJSU);:=86=36A8!PNNT ,%% NJRU NJSW MWU ," ," ," NJQW NJSS POU NJTP NJTW OUW NJRW NJTO
&.. NJTV NJSW ," ," NJTU NJRT NJRS NJV NJTN NJTP/64> MNJPU NJTQ ," ," NJTW NJUW NJTT NJSO NJQT NJTR
%C2 NJPQ NJSV ," ," NJRW NJSR NJTV NJUT NJRU NJTQ&636B66!PNNU ,%% NJQS NJS MTU NJTU NJUR OON ," ," ," NJUT NJVO OPT NJSW NJTV
&.. NJTU NJTU ," ," ," ," NJVR NJUR NJUT NJUO/64> NJUQ NJTP ," ," ," ," NJRO NJUP NJSU NJTU%C2 MNJQS NJRV MNJPW NJQV ," ," NJTO NJTT MNJNO NJSO
+>56;!<62= ,%% NJQO NJRU MOPVJOU NJUS NJSV OOOJNN NJSP NJTO PUJVQ NJUT NJTV WOJNN NJSS NJSV&.. NJSW NJSU ," ," NJTT NJSO NJTW NJTS NJTS NJSV/64> MOJNP NJSO ," ," NJRR NJUP NJQQ NJTQ MNJOR NJTP
! %C2 ! MNJNR NJRS ! MNJPN NJQV ! NJSQ NJTN ! NJSR NJTW ! NJQO NJSS
Tab 3 Performance criteria of the models across sites and models. E - Nash Sutcliffe efficiency {Nash, 1979 #298}, r - Kendall correlation
coefficient and absolute error x#
37

Figures:
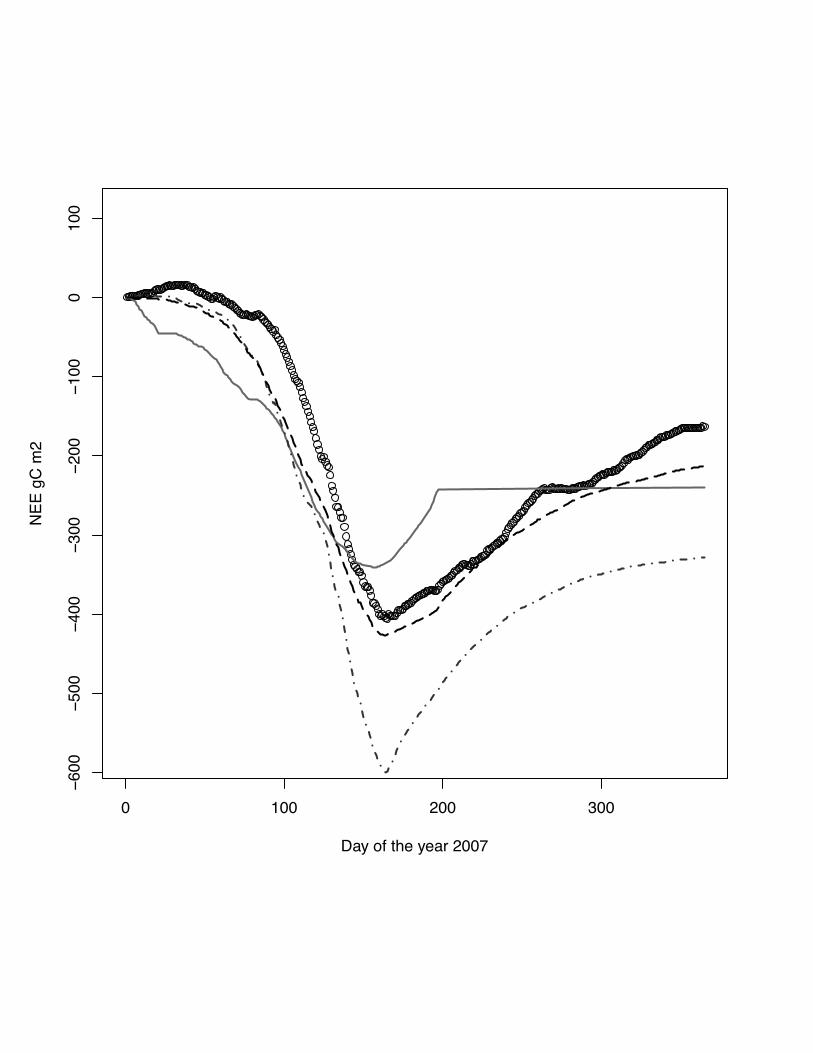
Fig. 1 The scatter plots of simulated versus measured NEE for the site Oensingen,
winter wheat, for the year 2007. The grey line indicates the lowess regression. The
numbers in right hand site panel are Kendall correlations coefficients. The size of the
number indicates the strength of the association. Noticeable is the change in the
regression for low fluxes between -2 gC m2 day-1 - 2 gC m-2day-1
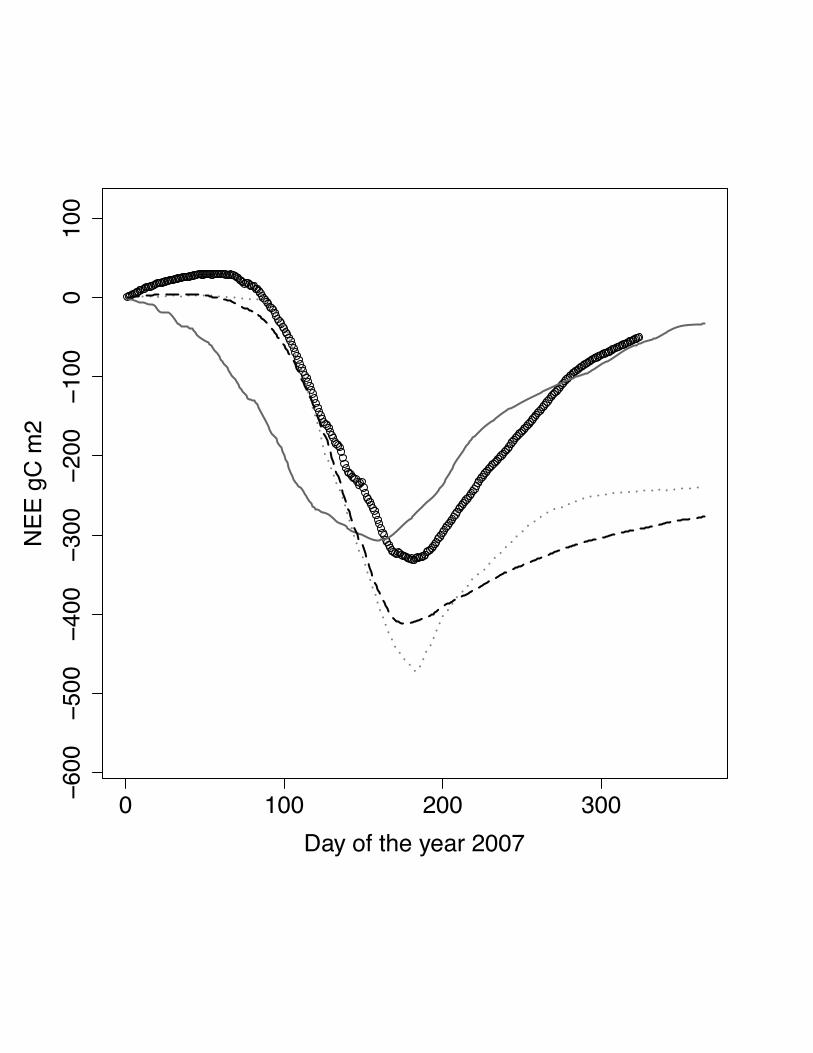
Fig. 2 Cumulative NEE for the year 2007 at the site Oensingen, winter wheat. DNDC
model: grey solid line, ORCHIDEE-STICS model: dark grey dash dot line, SPA -
black dash line, measurements indicated by open circles. In this case ORCHIDEE-
STICS is well below the measured NEE even before the end of the cropping season
(DOY 190).
Fig 3 The scatter plots of simulated versus measured NEE for the site Grignon, maize,
2005. The grey line indicates the lowess regression. The numbers in right hand site
panel are Kendall correlations coefficients. The size of the number indicates the
strength of the association. Noticeable is the change in the regression for low fluxes
between -2 gC m-2day-1 - 2 gC m-2day-1. There is no significant (p<0.05) correlation
for DNDC and ORCHIDEE-STICS.
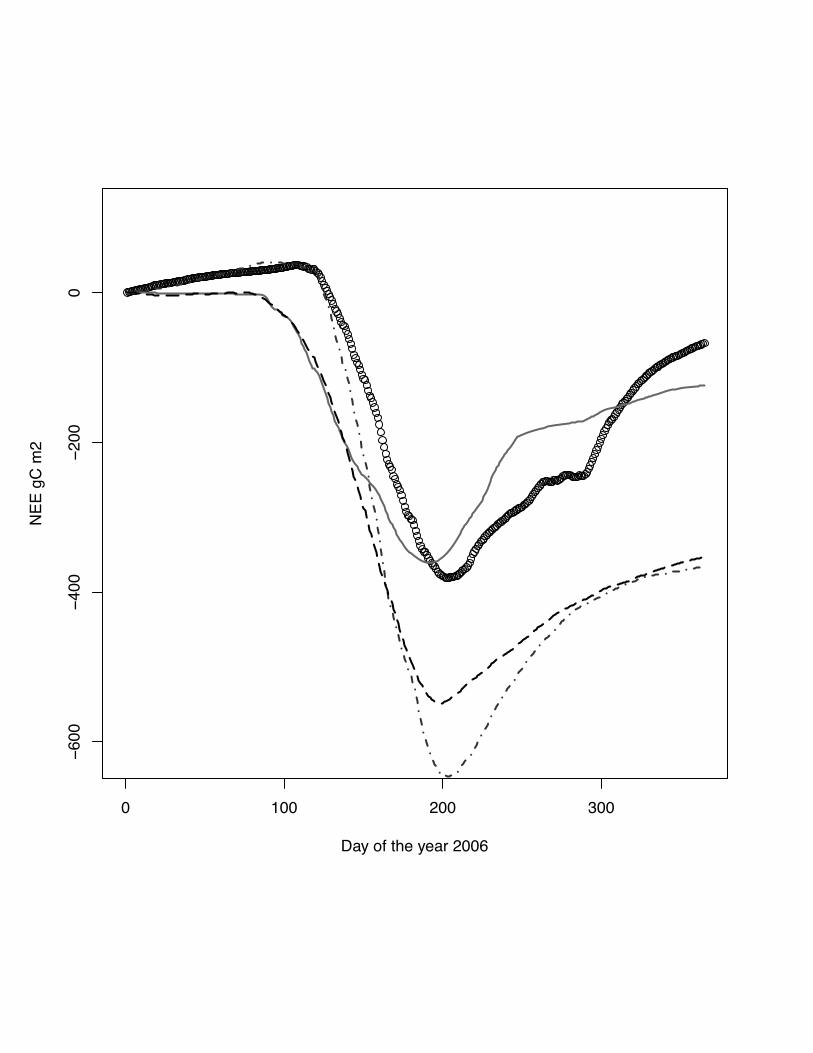
Fig. 4 Cumulative NEE for the year 2005 at the site Grignon, maize. DNDC model:
grey solid line, ORCHIDEE-STICS model: dark grey dash dot line, CERES-EGC -
38

grey dots, measurements indicated by open circles. All model fail to capture the
cumulative flux for the year.
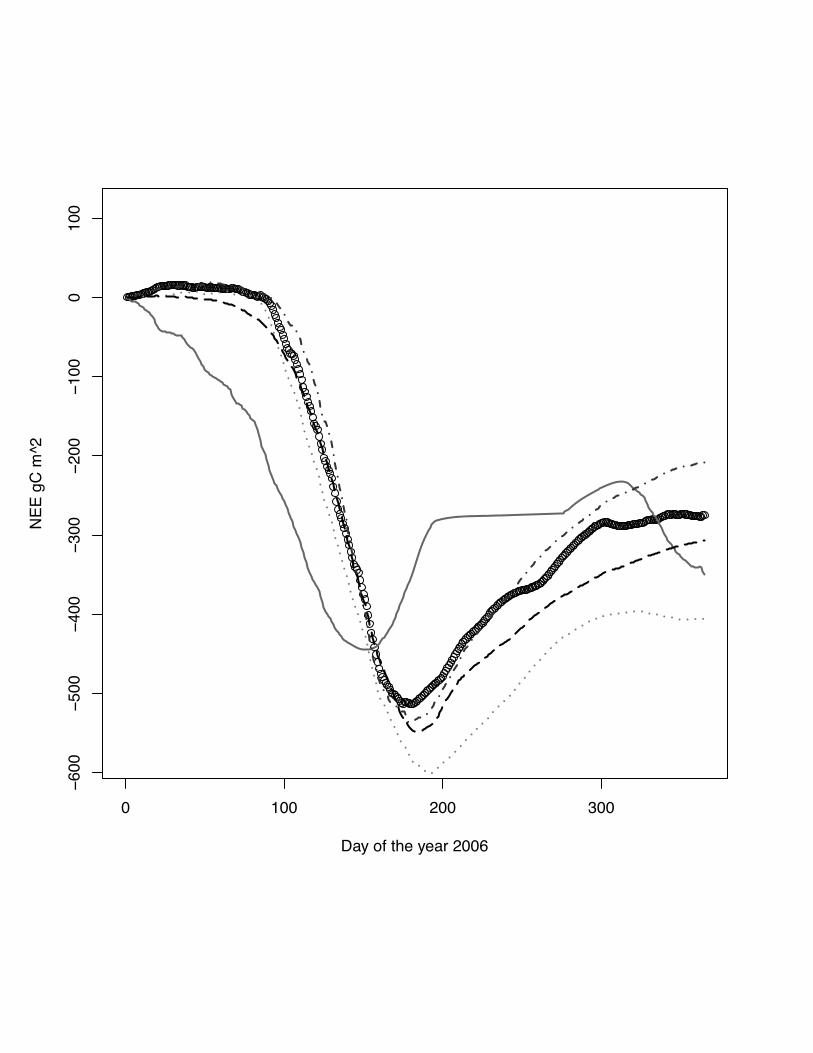
Fig 5 The scatter plots of simulated versus measured NEE for the site Grignon, winter
wheat, 2006. The grey line indicates the lowess regression. The numbers in right hand
site panel are Kendall correlations coefficients. The size of the number indicates the
strength of the association. Noticeable is the change in the regression for low
agreement for DNDC with measurements as well as the other models. The high
degree of association between ORCHIDEE-STICS and SPA indicates a similar
response to driving variables.
Fig. 6 Cumulative NEE for the year 2005 at the site Grignon, winter wheat. DNDC
model: grey solid line, ORCHIDEE-STICS model: dark grey dash dot line, CERES-
EGC - grey dots, SPA - black dash line, measurements indicated by open circles. Very
pronounced in the case the missing phase alignment between DNDC simulations and
observations. All models beside CERES-EGC are close to the measured cumulative
NEE.
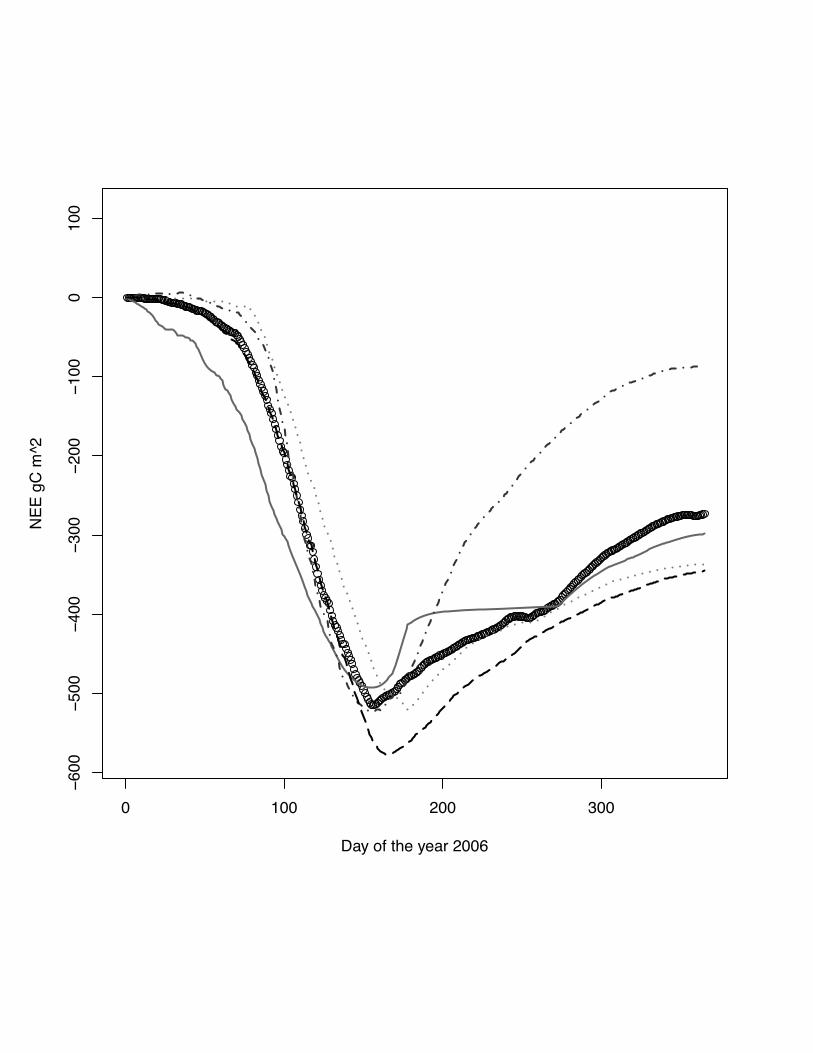
Fig 7 The scatter plots of simulated versus measured NEE for the site Aurade, winter
wheat, 2006. The grey line indicates the lowess regression. The numbers in right hand
site panel are Kendall correlations coefficients. The size of the number indicates the
strength of the association. IN this case all model capture the NEE with good levels of
association.
39

Fig. 8 Cumulative NEE for the year 2005 at the site Aurade, winter wheat. DNDC
model: grey solid line, ORCHIDEE-STICS model: dark grey dash dot line, CERES-
EGC - grey dots, SPA - black dash line, measurements indicated by open circles.
Beside capturing the daily dynamic of fluxes (fig 7) ORCHIDEE-STICS fails to
capture the fluxes after the growing season.
Fig 9 The scatter plots of simulated versus measured NEE for the site Klingenberg,
winter wheat, 2006. The grey line indicates the lowess regression. The numbers in
right hand site panel are Kendall correlations coefficients. The size of the number
indicates the strength of the association. Again apparent, the different interpretation of
small fluxes between models and measurements as well as between models.
Fig. 10 Cumulative NEE for the year 2005 at the site Klingenberg, winter wheat.
DNDC model: grey solid line, ORCHIDEE-STICS model: dark grey dash dot line,
SPA - black dash line, measurements indicated by open circles. Here DNDC captures
again the cumulative flux better than the two other models.
Fig 11 The scatter plots of simulated versus measured NEE for the site Gebesee,
winter barley, 2006. The grey line indicates the lowess regression. The numbers in
right hand site panel are Kendall correlations coefficients. The size of the number
indicates the strength of the association.
Fig. 12 Cumulative NEE for the year 2005 at the site Gebesee, winter barley. DNDC
model: grey solid line, CERES-EGC - grey dots, SPA - black dash line, measurements
40

indicated by open circles. The DNDC model is again able to capture the cumulative
flux even so it shows a strong temporal misalignment.
Fig 13 The scatter plots of simulated versus measured Reco for the Oensingen, winter
wheat, for the year 2007. The grey line indicates the lowess regression. The numbers
in right hand site panel are Kendall correlations coefficients. The size of the number
indicates the strength of the association. We observe a strong mismatch for the low
fluxes especially for DNDC but also for the other models indicated by the change in
the lowess fit function.
Fig 14 The scatter plots of simulated versus measured Reco for the Aurade, winter
wheat, for the year 2005. The grey line indicates the lowess regression. The numbers
in right hand site panel are Kendall correlations coefficients. The size of the number
indicates the strength of the association. The pattern of lack of fit for small fluxes
reappears now more pronounced for ORCHIDEE-STICS and SPA as in fig 13.
Fig 15 The scatter plots of simulated versus measured Reco for the Klingenberg, winter
wheat, for the year 2005. The grey line indicates the lowess regression. The numbers
in right hand site panel are Kendall correlations coefficients. The size of the number
indicates the strength of the association. The misfit of small fluxes observed for the
two other sites is apparent but less pronounced in this case.
Fig 16 The scatter plots of simulated versus measured GPP for the Oensingen, winter
wheat, for the year 2007. The grey line indicates the lowess regression. The numbers
in right hand site panel are Kendall correlations coefficients. The size of the number
41

indicates the strength of the association. There is a good reproduction of GPP by the
models and a less pronounced small flux disagreement. Interesting here the high
agreement between ORCHIDEEE-STICS and SPA indicating, again, the high
similarity of the model response to the external drivers.
Fig 17 The scatter plots of simulated versus measured GPP for the Grignon, winter
wheat, for the year 2006. The grey line indicates the lowess regression. The numbers
in right hand site panel are Kendall correlations coefficients. The size of the number
indicates the strength of the association. At this site we now also observe a small flux
inconsistency between measurements and models as DNDC and the two other models.
Very pronounces again the agreement between SPA and ORCHIDEE-STICS.
Fig 18 The scatter plots of simulated versus measured GPP for the Klingenberg,
winter wheat, for the year 2006. The grey line indicates the lowess regression. The
numbers in right hand site panel are Kendall correlations coefficients. The size of the
number indicates the strength of the association. The models exhibit a similar
behaviour as we saw in the other plots with an overall moderate to good agreement.
Fig 19 The scatter plots of simulated versus measured ETa for the Oensingen, winter
wheat, for the year 2007. The grey line indicates the lowess regression. The numbers
in right hand site panel are Kendall correlations coefficients. The size of the number
indicates the strength of the association. Here the two SVAT models have the best
association with the measures latent heat flux. DNDC has again problems to
reproduce low fluxes but is still able to simulate high fluxes.
42

Fig 20 The scatter plots of simulated versus measured ETa for the Grignon, maize, for
the year 2005. The grey line indicates the lowess regression. The numbers in right
hand site panel are Kendall correlations coefficients. The size of the number indicates
the strength of the association. In this case DNDC and CERES-EGC are not able to
simulate higher fluxes.
Fig 21 The scatter plots of simulated versus measured ETa for the Grignon, winter
wheat, for the year 2006. The grey line indicates the lowess regression. The numbers
in right hand site panel are Kendall correlations coefficients. The size of the number
indicates the strength of the association. CERES-EGC has together with DNDC
problems to reproduce high flux rates as observed before for this site.
Fig 22 The scatter plots of simulated versus measured Reco for the Aurade, winter
wheat, for the year 2005. The grey line indicates the lowess regression. The numbers
in right hand site panel are Kendall correlations coefficients. The size of the number
indicates the strength of the association. Apparent again, a lack of fit for DNDC for
higher flux rates.
Fig 23 The scatter plots of simulated versus measured ETa for the Gebesee, winter
barley, for the year 2007. The grey line indicates the lowess regression. The numbers
in right hand site panel are Kendall correlations coefficients. The size of the number
indicates the strength of the association. We observe the same pattern again, CERES-
EGC and DNDC fail to simulate the higher flux rates.
43
0 100 200 300
600
500
400
300
200
100
010
0
Day of the year 2007
NEE
gC m
2
0 100 200 300
600
500
400
300
200
100
010
0
Day of the year 2007
NEE
gC m
2
0 100 200 300
600
400
200
0
Day of the year 2006
NEE
gC m
2
0 100 200 300
600
500
400
300
200
100
010
0
Day of the year 2006
NEE
gC m
^2
0 100 200 300
600
500
400
300
200
100
010
0
Day of the year 2006
NEE
gC m
^2
0 100 200 300
500
400
300
200
100
010
0
Day of the year 2005
NEE
gC m
^2
Reco_meas gC m2
0 2 4 6 8 10 12
0.50 0.66
0 2 4 6 8
02
46
810
1214
0.64
02
46
810
12
Reco DNDC 0.60 0.50
Reco ORCHIDEE
02
46
8
0.79
0 2 4 6 8 10 14
02
46
8
0 2 4 6 8
Reco SPA
Reco_meas gC m2
0 5 10 15
0.63 0.72
0 2 4 6 8
02
46
810
0.66
05
1015
Reco DNDC 0.68 0.71
Reco ORCHIDEE
02
46
810
0.77
0 2 4 6 8 10
02
46
8
0 2 4 6 8 10
Reco SPA
Reco_meas gC m^2
0 2 4 6 8 10 12
0.38 0.71
0 2 4 6 8
02
46
0.62
02
46
810
12
Reco DNDC 0.29 0.42
Reco ORCHIDEE
24
68
0.68
0 2 4 6
02
46
8
2 4 6 8
Reco SPA
NEEmeas gC m2
6 4 2 0 2 4
0.50 0.74
8 6 4 2 0 2
86
42
02
4
0.81
64
20
24
NEE DNDC 0.52 0.60
NEE CERES
84
02
4
0.75
8 6 4 2 0 2 4
86
42
02
8 4 0 2 4
NEE SPA
NEEmeas gC m2
6 4 2 0 2 4 6
0.59 0.55
10 6 4 2 0 2
105
05
0.62
64
20
24
6
NEE DNDC 0.60 0.74
NEE ORCHIDEE
1510
50
5
0.71
10 5 0 5
106
42
02
15 10 5 0 5
NEE SPA
NEEmeas gC m^2
5 0 5
0.32 0.60
10 5 0
0.72
105
0
0.71
50
5
NEE DNDC 0.35 0.32 0.30
NEE CERES 0.68
106
22
0.69
105
0
NEE ORCHIDEE 0.88
10 5 0 10 6 2 2 8 4 0 2 4
84
02
4
NEE SPA
NEEmeas gC m2
4 2 0 2 4
0.45 0.54
8 6 4 2 0 2 4
105
05
0.56
42
02
4
NEE DNDC 0.62 0.59
NEE ORCHIDEE
1510
50
5
0.82
10 5 0 5
86
42
02
4
15 10 5 0 5
NEE SPA
NEEmeas gC m^2
10 5 0 5
0.28 0.59
10 5 0 5
1510
50
5
0.40
105
05
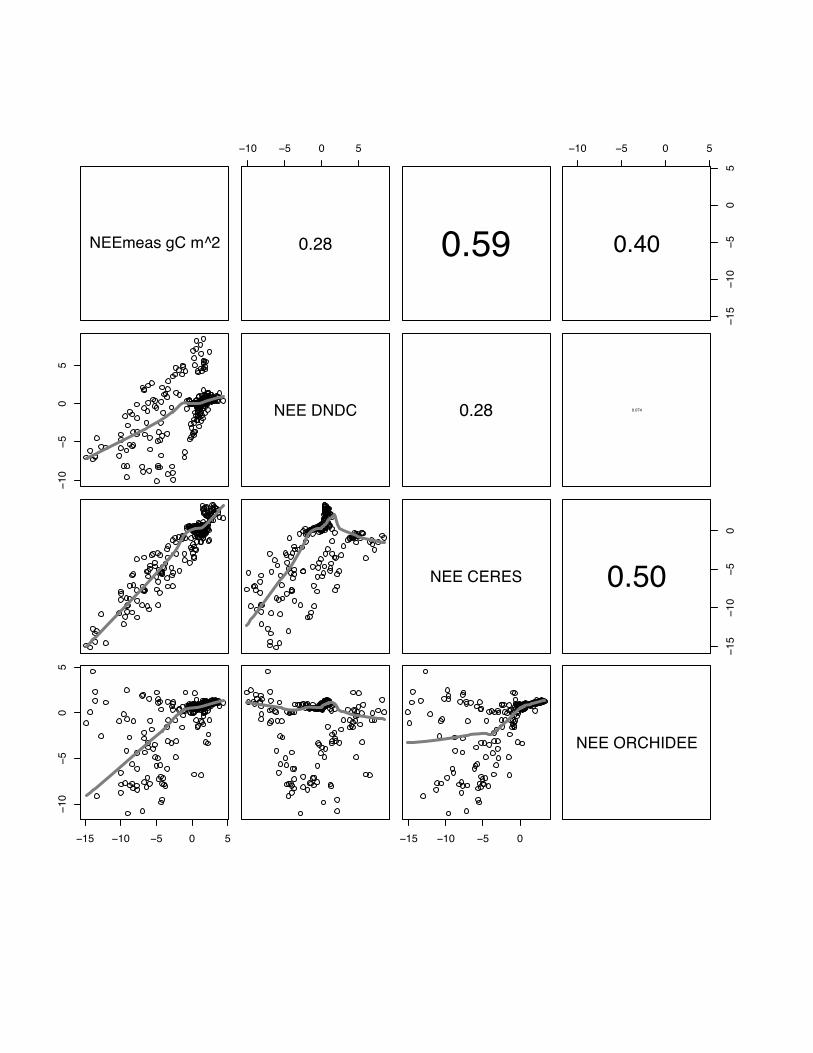
NEE DNDC 0.28 0.074
NEE CERES
1510
50
0.50
15 10 5 0 5
105
05
15 10 5 0
NEE ORCHIDEE
NEEmeas gC m^2
5 0 5
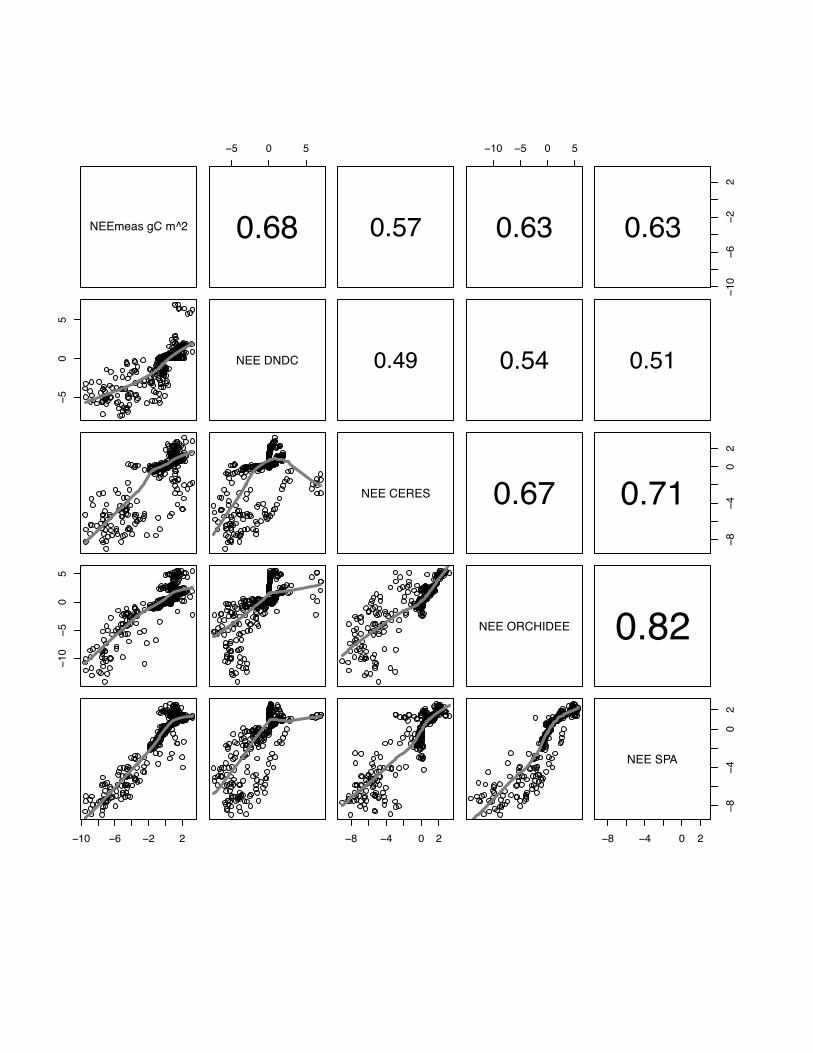
0.68 0.57
10 5 0 5
0.63
106
22
0.63
50
5
NEE DNDC 0.49 0.54 0.51
NEE CERES 0.67
84
02
0.71
105
05
NEE ORCHIDEE 0.82
10 6 2 2 8 4 0 2 8 4 0 2
84
02
NEE SPA
GPPmeas gC m2
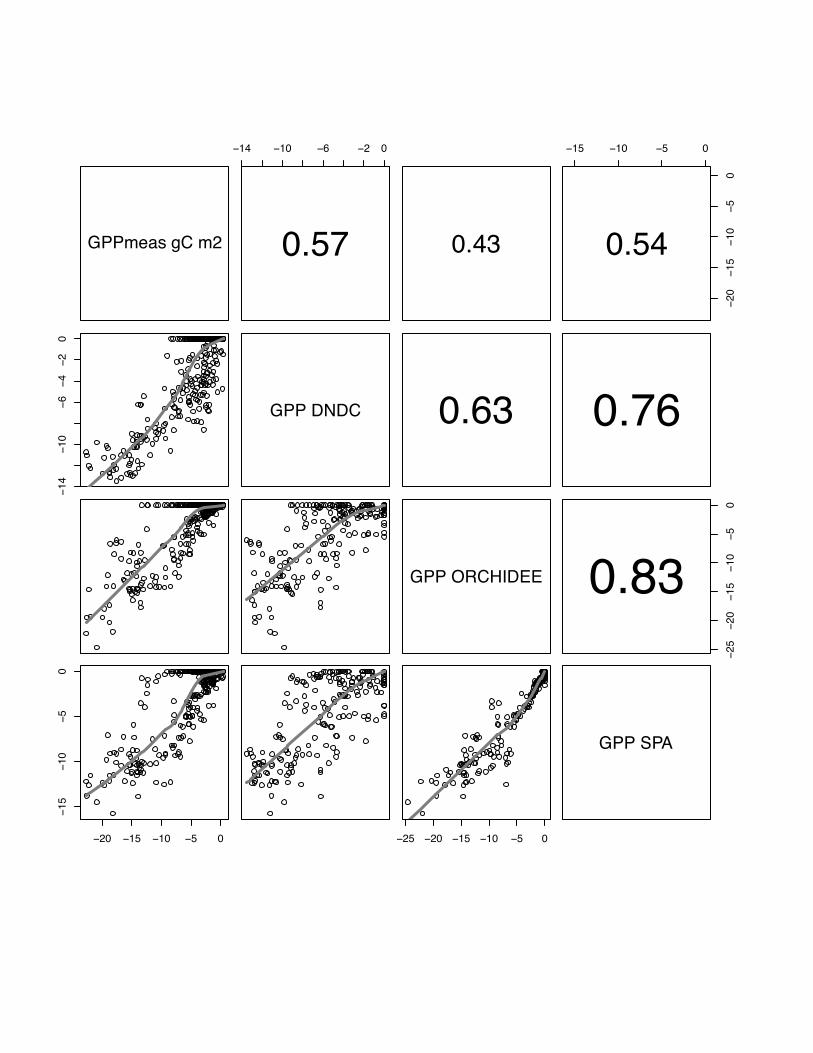
14 10 6 2 0
0.57 0.43
15 10 5 0
2015
105
0
0.54
1410
64
20
GPP DNDC 0.63 0.76
GPP ORCHIDEE
2520
1510
50
0.83
20 15 10 5 0
1510
50
25 20 15 10 5 0
GPP SPA
GPPmeas gC m2
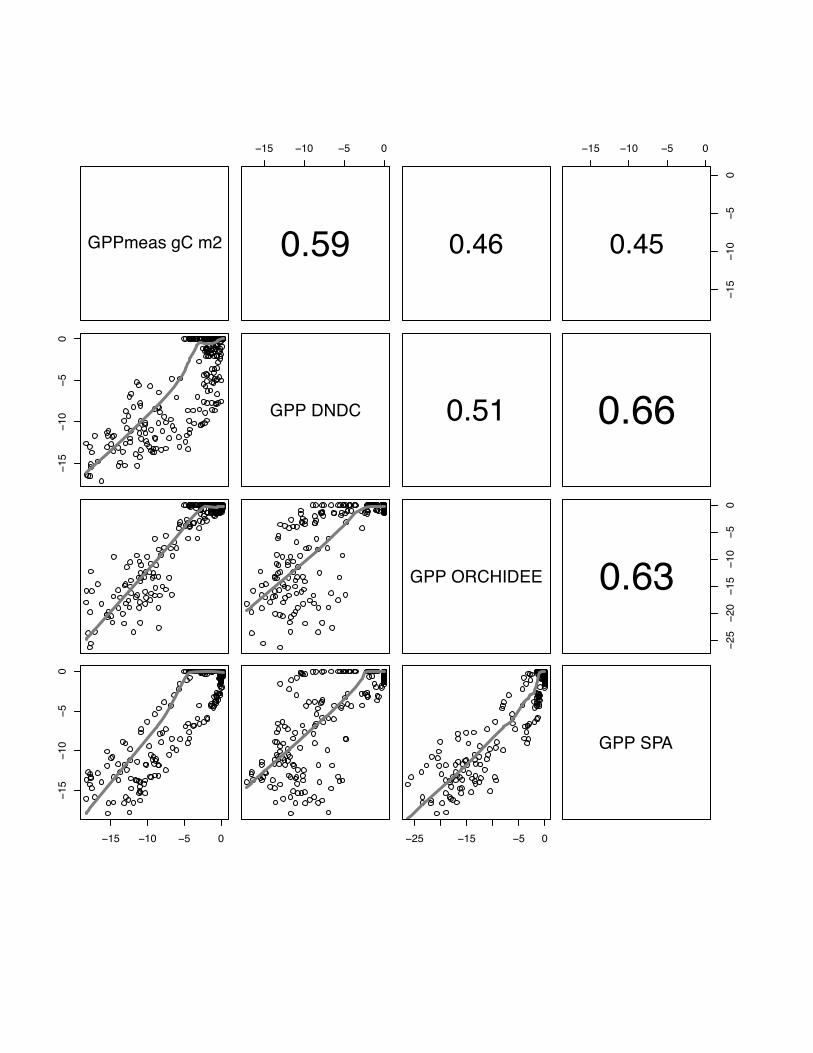
15 10 5 0
0.59 0.46
15 10 5 0
1510
50
0.45
1510
50
GPP DNDC 0.51 0.66
GPP ORCHIDEE
2520
1510
50
0.63
15 10 5 0
1510
50
25 15 5 0
GPP SPA
GPPmeas gC m^2
15 10 5 0
0.49 0.59
15 10 5 0
1510
50
0.51
1510
50
GPP DNDC 0.67 0.58
GPP ORCHIDEE
2015
105
0
0.83
15 10 5 0
1510
50
20 15 10 5 0
GPP SPA
ETa meas mm
0 1 2 3 4 5
0.41 0.68
0 1 2 3 4 5
01
23
45
6
0.65
01
23
45
ETa DNDC 0.39 0.39
ETa ORCHIDEE
01
23
4
0.70
0 1 2 3 4 5 6
01
23
45
0 1 2 3 4
ETa SPA
ETa meas mm
0 1 2 3 4 5
0.48 0.38
0 1 2 3 4
01
23
4
0.66
01
23
45
ETa DNDC 0.47 0.49
ETa CERES
01
23
45
0.34
0 1 2 3 4
01
23
4
0 1 2 3 4 5
ETa SPA
ETa meas mm
0 1 2 3 4
0.42 0.32
0 1 2 3 4
0.61
01
23
45
0.68
01
23
4
ETa DNDC 0.35 0.38 0.32
ETa CERES 0.23
01
23
4
0.33
01
23
4
ETa ORCHIDEE 0.61
0 1 2 3 4 5 0 1 2 3 4 0 1 2 3 4
01
23
4
ETa SPA
ETa meas mm
0 1 2 3 4
0.43 0.46
0 1 2 3 4
0.60
01
23
4
0.68
01
23
4
ETa DNDC 0.38 0.47 0.42
ETa CERES 0.46
01
23
45
0.41
01
23
4
ETa ORCHIDEE 0.65
0 1 2 3 4 0 1 2 3 4 5 0 1 2 3 4 5
01
23
45
ETa SPA
ETa meas mm
0 1 2 3 4 5
0.40 0.37
0.0 1.0 2.0 3.0
01
23
45
6
0.57
01
23
45
ETa DNDC 0.18 0.30
ETa CERES
0.0
1.0
2.0
3.0
0.21
0 1 2 3 4 5 6
0.0
1.0
2.0
3.0
0.0 1.0 2.0 3.0
ETa ORCHIDEE





























