Embed Size (px)
Citation preview
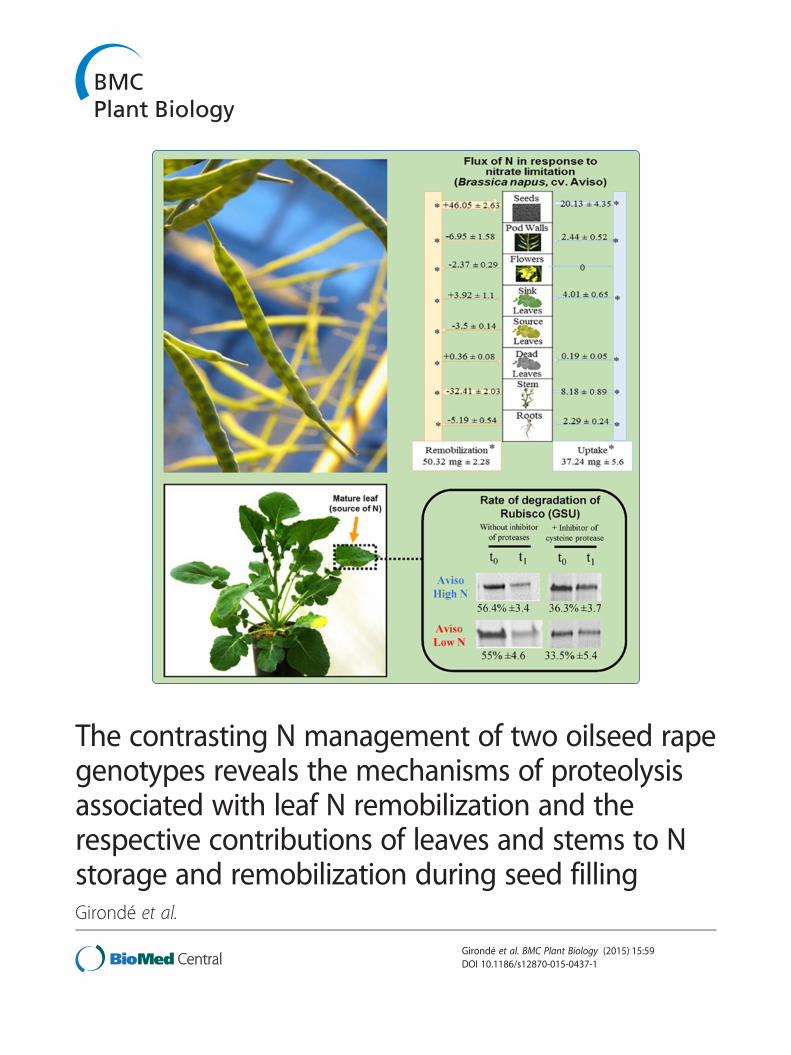
The contrasting N management of two oilseed rapegenotypes reveals the mechanisms of proteolysisassociated with leaf N remobilization and therespective contributions of leaves and stems to Nstorage and remobilization during seed fillingGirondé et al.
Girondé et al. BMC Plant Biology (2015) 15:59 DOI 10.1186/s12870-015-0437-1

Girondé et al. BMC Plant Biology (2015) 15:59 DOI 10.1186/s12870-015-0437-1
RESEARCH ARTICLE Open Access
The contrasting N management of two oilseedrape genotypes reveals the mechanisms ofproteolysis associated with leaf N remobilizationand the respective contributions of leaves andstems to N storage and remobilization duringseed fillingAlexandra Girondé1,2,3, Philippe Etienne1,2,3, Jacques Trouverie1,2,3, Alain Bouchereau4, Françoise Le Cahérec4,Laurent Leport4, Mathilde Orsel4,5,6, Marie-Françoise Niogret4, Nathalie Nesi4, Deleu Carole4, Fabienne Soulay7,Céline Masclaux-Daubresse7 and Jean-Christophe Avice1,2,3*
Abstract
Background: Oilseed rape is the third largest oleaginous crop in the world but requires high levels of N fertilizer ofwhich only 50% is recovered in seeds. This weak N use efficiency is associated with a low foliar N remobilization,leading to a significant return of N to the soil and a risk of pollution. Contrary to what is observed duringsenescence in the vegetative stages, N remobilization from stems and leaves is considered efficient duringmonocarpic senescence. However, the contribution of stems towards N management and the cellular mechanismsinvolved in foliar remobilization remain largely unknown. To reach this goal, the N fluxes at the whole plant levelfrom bolting to mature seeds and the processes involved in leaf N remobilization and proteolysis were investigatedin two contrasting genotypes (Aviso and Oase) cultivated under ample or restricted nitrate supply.
Results: During seed filling in both N conditions, Oase efficiently allocated the N from uptake to seeds while Avisofavoured a better N remobilization from stems and leaves towards seeds. Nitrate restriction decreased seed yieldand oil quality for both genotypes but Aviso had the best seed N filling. Under N limitation, Aviso had a better Nremobilization from leaves to stems before the onset of seed filling. Afterwards, the higher N remobilization fromstems and leaves of Aviso led to a higher final N amount in seeds. This high leaf N remobilization is associated witha better degradation/export of insoluble proteins, oligopeptides, nitrate and/or ammonia. By using an originalmethod based on the determination of Rubisco degradation in the presence of inhibitors of proteases, efficientproteolysis associated with cysteine proteases and proteasome activities was identified as the mechanism of Nremobilization.(Continued on next page)
* Correspondence: [email protected]é de Caen Basse-Normandie, F-14032 Caen, France2UCBN, UMR INRA–UCBN 950 Ecophysiologie Végétale, Agronomie &Nutritions N.C.S., F-14032 Caen, FranceFull list of author information is available at the end of the article
© 2015 Girondé et al.; licensee BioMed Central. This is an Open Access article distributed under the terms of the CreativeCommons Attribution License (http://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, andreproduction in any medium, provided the original work is properly credited. The Creative Commons Public DomainDedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article,unless otherwise stated.

Girondé et al. BMC Plant Biology (2015) 15:59 Page 2 of 21
(Continued from previous page)
Conclusion: The results confirm the importance of foliar N remobilization after bolting to satisfy seed filling andhighlight that an efficient proteolysis is mainly associated with (i) cysteine proteases and proteasome activities and(ii) a fine coordination between proteolysis and export mechanisms. In addition, the stem may act as transientstorage organs in the case of an asynchronism between leaf N remobilization and N demand for seed filling.
Keywords: Brassica napus, Leaf senescence, N remobilization efficiency, N use efficiency, Proteolysis, Proteasome
Background
Over the last four decades, oilseed rape (Brassica napusL.) has become the third most widely grown oleaginouscrop worldwide with a 2.4-fold increase in seed produc-tion between 1992 and 2012 [1]. This crop represents amajor renewable resource for human food (oil), animalfeed (meal) and numerous non-food uses (biofuel, lubri-cants, high added-value products derived from greenchemistry). To increase the level of seed production withcurrent genotypes, the use of N fertilizers has increasedby 430% over the last forty years [2]. Despite the highcapacity of mineral N absorption of oilseed rape [3], only50% of N from fertilizer is recovered in seeds at harvest[4]. This low N Use Efficiency (NUE) is related to theloss of N by leaf drop (up to 100 kg.N.ha−1.year−1, [5]),reflecting the weak N Remobilization Efficiency (NRE)of oilseed rape. Therefore, in a context of imposedlimitations on N-fertilizer inputs, improving NUE isbecoming a priority in order to maintain/increase seedyield and decrease (i) the risk of water pollution by ni-trate, (ii) the emission of greenhouse gases contributingto global warming, and (iii) the economic costs of oil-seed rape crops.NUE can be represented as two main components: the
N Uptake Efficiency (NUpE) and the N Utilization Effi-ciency (NUtE), itself subdivided into two other compo-nents, N Assimilation Efficiency (NAE) and NRemobilization Efficiency (NRE) [6]. Even if N uptake ofwinter oilseed rape is considered to be efficient at vege-tative stages [3], an N uptake that remains significantuntil flowering is associated with genotypes having highseed yield [7]. A mineral N input during seed filling in-creases seed yield of spring oilseed rape (cv. Aries, [8])and recent studies on winter oilseed rape [9-13] have re-ported that a N uptake during reproductive stages ap-pears to be a determinant trait for seed yield of winteroilseed rape, especially in restricted N supply. Neverthe-less, in field conditions, the mineral N availability highlyfluctuates during reproductive stages due to environ-mental factors, such as water deficit in the soil. Toobtain a high NUE, the N coming from uptake has to bewell managed by the plant. Consequently, an improve-ment of NUtE is also necessary to enhance NUE.Concerning the NAE, transgenic approaches have beentargeted to enzymes involved in N metabolism such as
nitrate reductase, nitrite reductase or alanine amino-transferase (AlaT) in various species such as Arabidopsisand tobacco, with more or less success (for review [14]).A promising result was obtained with oilseed rape plantsoverexpressing an AlaT, which need 40% less N fertilizerto reach similar yield as the wild type [14]. However, astudy of 40 spring oilseed rape genotypes has suggestedthat NRE is a major determinant of NUtE [15] and anefficient assimilation needs to be correlated to anenhanced N remobilization to improve the seed Nfilling, which is essential to improve/maintain seed yield,in a context of N input reduction.
The N remobilization associated with leaf senescenceis considered as crucial for oilseed rape yield [16]. Dur-ing the vegetative stages, N is remobilized from the olderleaves to the younger leaves along the axis of the plantsvia the sequential senescence, but senescent leaves canfall with a high level of residual N (up to 3.5% of drymatter) leading to a significant return of N to the soil[5,17]. By developing a modelling approach as a mean toidentify potential methods for improving the NUE ofoilseed rape, it was estimated that a 1% decrease in theresidual N in fallen leaves (from 3.5% to 2.5% of drymatter), resulting from an enhanced foliar N remobiliza-tion, may increase seed yield by 5-10% [18]. After bolt-ing, and especially during monocarpic senescence whenthe N is remobilized from vegetative organs to seeds, thelow soil N availability during spring and at the beginningof summer [19] makes N remobilization crucial for seedyield and seed filling. During this period of developmentthe main source organs are leaves, stems and pod walls[16,17,20,21]. During reproductive stages, the N amountin source leaves is highly remobilized (86% of N presentat the beginning of flowering in field conditions) [12],leading to a low residual N in dead leaves (less than 2%of dry matter; [17]) and resulting in the reduction of therisk of N pollution. These results highlight an efficient Nremobilization from source organs during monocarpicsenescence, but also reveal variations for the residual Namount in stems compared with leaves of four winteroilseed rape genotypes [12]. In addition, a genotypic andN-supply interaction for N content in stems was foundfor 12 genotypes of spring canola [22] and the genotypewith the best N remobilization was able to better remo-bilize N from stems and pod walls to seeds [23]. These

Girondé et al. BMC Plant Biology (2015) 15:59 Page 3 of 21
results suggest that stems can also be important organfor improving seed N-filling, but the role of stems in Nmanagement at the whole plant level for winter oilseedrape remains largely unknown.Although NRE seems to be a major lever for improv-
ing the NUE in oilseed rape, the cellular mechanismsassociated with N remobilization from senescing leaves(proteolysis and the N export) remain largely an enigma.During leaf senescence, the degradation of proteins (themain form of N storage in leaves) into amino acids orpeptides is performed by different classes of proteases.Studies on Arabidopsis reported the predominantinvolvement of cysteine and serine proteases (for review[24]), but also a role for the proteasome in the degrad-ation of carbonylated proteins, which are accumulatedduring senescence [25]. The Ribulose-1,5-biphosphatecarboxylase/oxygenase (Rubisco; EC 4.1.1.39) can repre-sent up to 65% of the soluble proteins in C3 plants [26]and 20–30% of total leaf N [27-29]. Consequently,Rubisco is the major source of N for remobilization andits budget is very relevant for the plant. In winter oilseedrape, cysteine, aspartic, and metallo-proteases as well asthe 26S proteasome are supposed to play an importantrole in foliar N remobilization during senescence in thevegetative stages [30,31] but there is no evidenceconcerning the proteolysis activities that are involved inleaf senescence and degradation of Rubisco after thebolting stage.Among the enzymes involved in the conversion of
amino acids into transportable forms, a recent studyhighlighted the importance of asparagine synthetase (AS;EC 6.3.5.4) in N remobilization during senescence inArabidopsis [32]. In addition, the glutamine synthetase(GS; EC 6.3.1.2) catalyses the assimilation of ammoniuminto glutamate to synthesize glutamine. Glutamine wasfound at high levels in phloem sap of oilseed rape (cv.DSV15 and Duplo; [33]) and consequently, GSs aresupposed to be largely involved in N remobilization pro-cesses during senescence in winter wheat [34] and maize[35], especially the cytosolic form (GS1) in the case ofoilseed rape [36,37]. In parallel, glutamate dehydrogen-ase (GDH; EC 1.4.1.2; [38]) may catalyse a glutamate de-amination [39,40] which provides ammonium for the GSactivity in senescing leaves. In addition, in case of highlevels of ammonium, GDH can incorporate ammoniumon α-ketoglutarate to produce glutamate [41]. A signifi-cant quantity of glutamate has been found in phloemsap of oilseed rape [33,42] and an increase of both GDHactivities in sliced leaves of oilseed rape was associatedwith a decrease in soluble proteins and ammonium [43],suggesting an important role for GDH in foliar N remo-bilization. The phloem loading of the resulting aminoacids is regarded as not limiting in oilseed rape at thevegetative stages [44]. The efficient export of amino
acids and the involvement of GS1 and GDH in N remo-bilization need to be confirmed in leaves as they senesceafter bolting because they are proposed to have an effi-cient N remobilization.The aim of this study was to identify the physiological
traits involved in the high NUE and N remobilization inrestricted N supply conditions, at whole plant (frombolting to mature seeds) and cellular levels (from boltingto flowering stages). To reach this goal, two genotypes(Aviso and Oase), known to have different strategies fol-lowing a nitrate limitation at the vegetative stage [45],were selected. Long term pulse-chase 15 N-labelling wasperformed to precisely define the endogenous and ex-ogenous N fluxes at the whole plant level between thebolting and mature seed stages, under restricted andample nitrate supply. A specific 15 N-labelling was alsoused in order to determine a relevant estimation of NUEand its different components (NUpE, NUtE and NRE).This study (i) highlights the role of leaves and stems inthe remobilization of N towards seeds and (ii) allows theidentification of the physiological traits associated with ahigh NUE in response to restricted N supply. Moreover,a study of foliar N remobilization in source leaves wasperformed through the analysis of foliar N compounds,the GS and GDH activities, and the development of anew method to study proteolytic activities, using en-dogenous Rubisco as a substrate, in order to determinethe involvement of different classes of proteases in the Nremobilization which occurs during this growth stage.
ResultsImpact of N limitation on growth, yield components, seedand oil compositionWhen plants were well supplied with nitrate (HN), thetotal DM (Figure 1A, B and Additional file 1) was higherfor Oase (34 g) compared with Aviso (28 g) during seedfilling (D70 to D99), mainly due to differences in thestem DM (12.1 g for Oase vs 8.4 g for Aviso). Under ourexperimental conditions, the occurrence of the differentstages of development was not significantly differentbetween both genotypes or between both N treatmentsfor a given genotype. In low nitrate (LN) conditions, nodifference in the total DM was observed between geno-types (Figure 1C, D and Additional file 1). As expected,compared with HN, DM was reduced in leaves fromD42, in stems at D70 and in pod walls at D99. Inaddition, DM declined significantly in siliques at D42and increased in stems at D70 for Aviso while DM of si-liques and stems declined at D70 for Oase. The rootDM at D99 was lower in Aviso under LN compared withHN. Under LN supply, a similar decrease in seed pro-duction was observed for both genotypes (−41.8% forAviso, −42.8% for Oase; Figure 1C and D).

Figure 1 Kinetics of dry matter (DM) of Aviso and Oase under ample or low N supply. The plants were supplied with ample (HN, 3.75 mM)or low (LN, 0.375 mM) concentrations of nitrate. Dry matter is expressed in g per plant for Aviso (A, C) and Oase (B, D) at 0 (early bolting), 42(pod formation), 70 (start of seed filling) and 99 (mature seeds) days after the beginning of bolting (D0). Seeds and pod walls were grouped andcalled siliques at D42 and they were separated from D70 onwards. Data are indicated as the mean value ± standard error (vertical bars). Differentletters (a, b, c) indicate that the total dry matter is significantly different between two dates. Hashes represent significant differences betweengenotypes in HN or LN conditions and the asterisks represent significant differences between N treatments (n = 4 plants; p < 0.05).
Table 1 Seed composition and nitrogen harvest index of Aviso and Oase under ample or low N supply
Aviso Oase
HN LN HN LN
Yield components C:N ratio 11.31 ± 0.19 14.29 ± 1,01* 11.19 ± 0.25 18.36 ± 0.70**#
Seed N amount (mg) 421.48 ± 40.08 194.36 ± 8.05** 432.84 ± 5.93 168.22 ± 7.77**#
N Harvesting Index (NHI) 63.69 ± 3.33 72.05 ± 2.14 65.01 ± 1.24 73.97 ± 1.68*
Seed composition Proteins (%) 32.20 ± 0.39 25.83 ± 1.20* 31.68 ± 1.87 19.48 ± 0.77*#
Fatty acids (%) 33.99 ± 1.31 38.32 ± 1.45* 30.10 ± 4.27 44.37 ± 1.75*#
C18:1 (oleic; %) 53.13 ± 0.46 55.86 ± 0.8 58.76 ± 3.13 64.61 ± 1.08*#
C18:2 (linolenic ω6; %) 21.91 ± 0.34 21.43 ± 0.33 15.34 ± 0.40# 17.29 ± 0.29*#
C18:3 (linoleic ω3; %) 11.53 ± 0.19 10.91 ± .022 8.05 ± 0.59# 7.56 ± 0.29#
C22:1 (erucic; %) 2.13 ± 0.13 8.26 ± 0.83* 3.57 ± 2.39 4.28 ± 2.19#
The plants were supplied with an ample (HN, 3.75 mM) or low (LN, 0.375 mM) nitrate concentration. The N amount in seed is estimated in milligrams. C: N ratio:ratio C: N in seeds; NHI: N Harvesting Index (mg N in seeds per mg N in shoot). The seed composition (percentage of proteins and fatty acids) and the oilcomposition (percentage in oleic, linolenic, linoleic and erucic acids) was determined by NIRS method. Asterisks represent significant differences between Ntreatments and hashes represent significant differences between the two genotypes (n = 4 plants; #,*= p < 0.05; **= p < 0.01).
Girondé et al. BMC Plant Biology (2015) 15:59 Page 4 of 21

Girondé et al. BMC Plant Biology (2015) 15:59 Page 5 of 21
In HN plants, the yield components (C:N ratio, seed Namount, N Harvest Index (NHI); Table 1) as well as theseed composition (proteins and oil in % of dry matter(DM); Table 1) were similar for both genotypes, exceptfor the percentage of omega-3 (C18:3) and omega-6(C18:2) precursors, which were higher for Aviso thanOase. However, the C18:2/C18:3 ratio was similar forboth genotypes (1.9). As expected, an N limitation led toa strong decrease in the N amount of seeds (−53% inAviso, −61% in Oase; Table 1). Compared with HN con-ditions, NHI significantly increased in Oase and tendedto increase in Aviso (p = 0.06) in response to LN treat-ment (Table 1). Seeds of Oase LN plants had a higher C:N ratio associated with a higher oil percentage, whichwas to the detriment of proteins (20% proteins in DM vs26% for Aviso). For oil composition, an increase of oleicacid (C18:1) proportion was observed under N limitationfor both genotypes (Table 1). While C18:3 and C18:2
Figure 2 Kinetics of the N amount in Aviso and Oase under ample orlow (LN, 0.375 mM) concentrations of nitrate. The N amount is expressed in(pod formation), 70 (start of seed filling) and 99 (mature seeds) days after tcalled siliques at D42 and they were separated from D70 onwards. Data areletters (a, b, c) indicate that the total N amount is significantly different betgenotypes in HN or LN conditions and the asterisks represent significant d
percentages did not differ under both N conditions forAviso, an increase in C18:2 (+2%) and in the C18:2/C18:3 ratio (2.3) occurred under LN for Oase. Theproportion of erucic acid also increased under LN condi-tions for Aviso (8% of oil).
Effects of nitrate limitation on N amount and NUEcomponentsIn HN plants, the total N amount did not differ betweenthe genotypes (Figure 2A, B and Additional file 2). How-ever, the N amount in leaves and flowers at the finalstage of development was significantly higher for Oasethan for Aviso. As expected, the total N amount was re-duced for both genotypes in LN conditions (Figure 2C,D and Additional file 2), resulting from a decline in theN amount in nearly all organs from D42. Exceptionswere flowers present at any time during the experiment,roots at D42 for Oase, stems at D70 and leaves at D99
low N supply. The plants were supplied with ample (HN, 3.75 mM) ormg per plant for Aviso (A, C) and Oase (B, D) at 0 (early bolting), 42
he beginning of bolting (D0). Seeds and pod walls were grouped andindicated as the mean value ± standard error (vertical bars). Different
ween two dates. Hashes represent significant differences betweenifferences between N treatments (n = 4 plants; p < 0.05).

Girondé et al. BMC Plant Biology (2015) 15:59 Page 6 of 21
for Aviso, which all showed a similar N amount in bothN conditions. In LN plants, the final N amount in seedsand pod walls of Oase was lower than in Aviso (D99).The N use efficiency (NUE), N utilization efficiency
(NUtE), and N remobilization efficiency (NRE) werenegatively correlated to N supply while the N uptakeefficiency (NUpE) was positively correlated to N supply(Table 2). The NUE was higher for Oase (2.29) than forAviso (1.98) in HN conditions while no differences wereobserved in the LN treatment (2.86 for Oase and 3 forAviso). The NUtE increased in response to LN supplybut no differences were observed between the genotypes,whatever the N supply. A strong genotype/treatmentinteraction effect was observed for NUpE between D70and D99. In HN conditions, the NUpE of Oase wasaround 81%, suggesting that the N distributed to seedsduring this period is mainly provided by the N that isnewly taken up by the roots. The NUpE of Oase is about2-fold higher than Aviso in both N conditions. The glo-bal NRE, i.e. the NRE calculated on the whole growingcycle (D0-D99; Table 2), reached about 55-60% for bothgenotypes, indicating that a large part of the N stored insource organs at the bolting stage (D0) is remobilizedduring the experiment. When NRE was calculated step-wise (i.e. between D0-D42, D42-D70 and D70-D99), aregular decrease occurred throughout the steps, what-ever the treatment and the genotype. In HN plants, NREwas 1.2-fold higher in Oase than Aviso between D42 andD70 while it was reduced by about 50% after D70 (32.7
Table 2 NUE, NUtE, NUpE and NRE of Aviso and Oase under a
Genotypes (G)
Aviso Oase
N treatment (N) HN LN HN
NUEa 1.98 ± 0.06 3.00 ± 0.12** 2.29 ± 0.11
NUtEb 13.25 ± 0.66 20.27 ± 2.00* 13.67 ± 0.5
NUpEc (%; D70-D99) 45.86 ± 8.10 32.00 ± 5.30 81.57 ± 4.2
NREd (%) Global
0-99 59.12 ± 1.99 74.10 ± 3.22** 52.83 ± 2.0
Stepwise
0-42 72.28 ± 3.48 86.62 ± 4.37* 77.59 ± 4.5
42-70 51.17 ± 1.46 77.80 ± 4.16** 59.90 ± 2.9
70-99 32.70 ± 2.75 39.96 ± 2.19 14.91 ± 1.1
The plants were supplied with an ample (HN, 3.75 mM) or low (LN, 0.375 mM) nitraformation, D70 to seed filling and D99 to mature seeds. The main source of variatiogenotype x N treatment interaction were tested. The resulting F values are presenteNUE, NUtE, NUpE and NRE, respectively.a:NUE (Nitrogen Use Efficiency) corresponds to the mg of N in seeds per mg of thetotal DM (including roots, pod walls and seeds).b:NUtE (Nitrogen Utilization Efficiency) corresponds to the g of seed DM produced pc:NUpE (Nitrogen Uptake Efficiency) is estimated as the percentage of N derived frobetween D70 and D99 (% of N distributed to seeds that derived from uptake).d:NRE (Nitrogen Remobilization Efficiency) is determined as the percentage of N storelative to the total N amount in all sources) between D0 (bolting stage) and final hAsterisks represent significant differences between N treatments and hashes repres0.05; **= p < 0.01; ***= p < 0.001).
for Aviso vs 14.9 for Oase). In response to LN supply,global NRE increases for both genotypes, but a genotypiceffect was highlighted between D70 and D99, where theNRE of Aviso was 1.4-fold higher than Oase (Table 2).However, no genotype/treatment interaction was ob-served for NRE.
N fluxes at the whole plant level in HN conditionsThe 15 N labelling method used in this study gave theopportunity to determine precisely the N fluxes at thewhole plant level (remobilization and uptake) betweenD0-D42, D42-D70 and D70-D99. Due to the fact thatgenotype and N treatment effects were observed forNRE solely between D70 and D99 in HN plants (Table 2),only the N fluxes between these two growing stages aregiven in Figure 3 (for other growing stages, N fluxes aregiven in Additional files 3 and 4). In HN conditions, Nremobilization was around 3-fold higher in Aviso(Figure 3A) than Oase (Figure 3B). For both genotypes,the stem was the main source organ: 47% (i.e. 57.32 mg N)and 59% (i.e. 24.70 mg N) of the total N remobilized inAviso and Oase, respectively. For Aviso, the other sourceorgans were the leaves, pod walls, flowers and roots whilefor Oase the source organs were leaves and roots. ForOase, the N amount remobilized from leaves was 4.4-foldlower than for Aviso. The N remobilized from sourceorgans was mainly distributed to the seeds with asignificantly greater amount in Aviso (119.18 mg N)than Oase (31.66 mg N). Contrary to Aviso, the flowers of
mple or low N supply
Source of variation
G N G x N
LN FG FN r FGxN# 2.86 ± 0.08** 1.048 89.81*** −0.90 7.13*
0 23.45 ± 0.55* 3.38 73.78*** −0.90 1.10
5# 70.32 ± 7.88# 8.72* 60.81*** 0.76 22.96**
3 76.08 ± 4.10** 0.70 55.28*** −0.89 2.59
2 83.26 ± 4.16 0.07 7.74* −0.60 1.45
2# 75.65 ± 11.29 0.37 15.40** −0.73 1.01
3# 27.70 ± 1.75**# 71.99*** 31.99*** −0.52 2.43
te concentration. D0 corresponds to the early bolting stage, D42 to podn is deduced from an ANOVA test where genotypes, N treatment, andd below. The r values correspond to the correlation between N supply and
N amount in the whole plant relative to the seed DM (g) produced per g of
er mg of N in shoots (including pod walls and seeds).m uptake in the total N (from remobilization and uptake) distributed to seeds
red in all source organs which is remobilized (the total remobilized N amountarvest (D99) or between D0 and D42, D42 and D70 and D70 and D99.ent significant differences between the two genotypes (n = 4 plants; #,*= p <

Figure 3 N fluxes in Aviso (A) and Oase (B) in HN conditions between D70 and D99. The plants were supplied with an ampleconcentration of nitrate (HN, 3.75 mM of nitrate). D70 corresponds to the start of seed filling and D99 to the mature seed stage. Fluxes of N fromremobilization or uptake in the different organs are expressed as mg of N remobilized or taken up, respectively. A shaded box means that theorgan was not present during these growing stages. For fluxes of N remobilization, the N amount is indicated with a minus sign (−) when N isremobilized from a source organ, or it is indicated with a plus sign (+) when remobilized N is redistributed towards a sink organ. Data areindicated as the mean value ± standard error. Hashes represent significant differences between genotypes (n = 4 plants; p < 0.05).
Girondé et al. BMC Plant Biology (2015) 15:59 Page 7 of 21
Oase are sink organs (7.46 mg N) for remobilized N.There was no significant difference in the amount of Nuptake between genotypes (202.8 mg N for Aviso and217.6 mg N for Oase; Figure 3). However, the allocation ofN towards seeds is more important for Oase (135.6 mg N)than for Aviso (87.97 mg N). Despite this higher allocationof N towards seeds in Oase, the total N distributed toOase seeds was not the same as in Aviso (−40 mg N forOase compared with Aviso; Figure 3).
N fluxes at the whole plant level in response to nitratelimitationBetween D0 and D42, N remobilization increased forAviso in response to LN treatment (+20.6 mg N;Figure 4A) compared with HN (Additional file 3A), dueto a larger N remobilization from source leaves(+18 mg N). This remobilized N was mainly redistribu-ted towards siliques, allowing a similar amount of N tobe redistributed as HN plants (around 110 mg N). Incontrast to Aviso, Oase showed a similar total amount ofremobilized N in both N conditions (154.4 mg and150.6 mg N in LN and HN conditions, respectively;
Figure 4B and Additional file 3B). However, comparedwith HN, the redistribution of this remobilized N to si-liques was higher in Oase LN plants (+28 mg N) due toa lower loss via dead leaves and a lower redistribution tosink leaves (Figure 4B). The N remobilization in Oasewas lower than Aviso (−12 mg N) due to a lower contri-bution of roots and flowers. As expected, in LN plantsthe N uptake strongly decreased for Aviso (only37.82 mg N) and was not detected for Oase (Figure 4).In Aviso LN plants, the main sink organs for the N up-take were siliques, leaves and stems (Figure 4A). Thanksto the N uptake in Aviso, which supplemented the N re-mobilization, the total N amount distributed to siliques wassimilar in both genotypes (around 130 mg N; Figure 4).Between D42 and D70, no N uptake was detectable
and the total remobilized N amount decreased for bothgenotypes under N limitation (Figure 5) contrary to HNplants (Additional file 4). This was mainly related tolower contributions of leaves and pod walls, leading to alower N redistribution to seeds. The N remobilizationwas 1.2-fold higher for Aviso than Oase (Figure 5),mainly due to a 2-fold higher N remobilization from

Figure 4 N fluxes in Aviso (A) and Oase (B) in LN conditions between D0 and D42. The plants were supplied with a low concentration of nitrate (LN,0.375 mM of nitrate). D0 corresponds to early bolting and D42 to pod formation. Fluxes of N from remobilization or uptake in the different organs are expressedas mg of N remobilized or taken up, respectively. For fluxes of N remobilization, the N amount is indicated with a minus sign (−) when N is remobilized from asource organ, or it is indicated with a plus sign (+) when remobilized N is redistributed towards a sink organ. Data are indicated as the mean value± standarderror. Asterisks represent significant differences between treatments and hashes represent significant differences between genotypes (n=4 plants; p<0.05).
Figure 5 N fluxes in Aviso (A) and Oase (B) in LN conditions between D42 and D70. The plants were supplied with a low concentration ofnitrate (LN, 0.375 mM of nitrate). D42 corresponds to pod formation and D70 to the start of seed filling. Fluxes of N from remobilization or uptakein the different organs are expressed as mg of N remobilized or taken up, respectively. A shaded box means that the organ was not presentduring these growing stages. For fluxes of N remobilization, the N amount is indicated with a minus sign (−) when N is remobilized from asource organ, or it is indicated with a plus sign (+) when remobilized N is redistributed towards a sink organ. Data are indicated as the meanvalue ± standard error. Asterisks represent significant differences between treatments and hashes represent significant differences betweengenotypes (n = 4 plants; p < 0.05).
Girondé et al. BMC Plant Biology (2015) 15:59 Page 8 of 21

Girondé et al. BMC Plant Biology (2015) 15:59 Page 9 of 21
leaves. Nevertheless, the N redistribution to seeds wasnot significantly different between the genotypes(144.73 mg N for Aviso and 127.75 mg N for Oase).These results can be explained by the fact that flowersand stems are sink organs for Aviso contrary to Oase,and a higher N loss by dead leaves occurs for Aviso.Between D70 and D99, the N remobilization from all
source organs was low for both genotypes (Figure 6),leading to a lower total N remobilization in LN com-pared with HN plants (−59% for Aviso and −43% forOase; Figure 3). Consequently, a lower N amount wasredistributed to seeds (−73.13 mg N for Aviso and−14.22 mg N for Oase; Figure 6). Under LN conditions,Aviso had a higher amount of total remobilized N thanOase (+27 mg N) resulting in a higher redistribution ofremobilized N to seeds (+29 mg N). This was related toa higher remobilization from stems (+21.81 mg N) andsource leaves (+2.42 mg N) for Aviso compared withOase. The flowers were sink organs for remobilized N inOase LN plants (Figure 6B) in contrast to Aviso LNplants. It is noteworthy that contrary to the HN treat-ment, sink leaves were present for both genotypes(Figure 6). Unlike the previous period (D42-D70,Figure 5), a significant N uptake occurred in both
Figure 6 N fluxes in Aviso (A) and Oase (B) in LN conditions betweennitrate (LN, 0.375 mM of nitrate). D70 corresponds to the start of seed fillinin the different organs are expressed as mg N remobilized or taken up, reswith a minus sign (−) when N is remobilized from a source organ, or it is intowards a sink organ. Data are indicated as the mean value ± standard errotreatments and hashes represent significant differences between genotype
genotypes under LN conditions (Figure 6). The N uptakeand allocation of N taken up into seeds were respectively1.57- and 1.86-fold higher for Oase than for Aviso.Nevertheless, the total N amount distributed to theseeds of Oase LN plants (i.e. from N uptake and Nremobilization, 54.8 mg N) remained lower than thetotal N amount distributed to seeds of Aviso LN plants(66.2 mg N, Figure 6).
Impacts of nitrate limitation on chlorophyll levels, Ncompounds and amino acid metabolism during Nremobilization in a selected source leafIn order to study the remobilization at the foliar level inresponse to N limitation applied at the bolting stage(D0), selected mature leaves undergoing senescence dur-ing the experiment (called “source leaves”) were analysedin detail over 28 days. The leaf biomass (at D0), leaf area(from D0 to D28; Additional file 5) and chlorophyll con-tent (at D0; Figure 7A) in these selected source leaveswere not significantly different between Aviso and Oase,meaning that the leaf initial status was similar betweenboth genotypes. Consequently, it was possible to com-pare the processes involved in the N remobilization ofsource leaves of both genotypes. In HN conditions, the
D70 and D99. The plants were supplied with a low concentration ofg and D99 to mature seeds. Fluxes of N from remobilization or uptakepectively. For fluxes of N remobilization, the N amount is indicateddicated with a plus sign (+) when remobilized N is redistributedr (vertical bars). Asterisks represent significant differences betweens (n = 4 plants; p < 0.05).

Figure 7 Changes in chlorophyll level, total N, N-soluble proteins, N-amino acids and other N compounds in a source leaf. Plants ofAviso and Oase were supplied with ample (HN, 3.75 mM) or low (LN, 0.375 mM) nitrate concentrations. These data were obtained on a selected“source leaf”, determined as mature at D0 (early bolting) and becoming senescent during the experiment. The chlorophyll amount (A; SPADvalue) is expressed in an arbitrary unit. The amount of total N (B), soluble proteins (C), amino acids (D) and other N compounds (E) are expressedas mg of N per leaf for each fraction. The fraction of other N compounds that mainly corresponded to insoluble proteins, oligopeptides andammonia, was determined as follows: mg of total N amount - (mg of N-soluble proteins +mg of N-amino acids). All these data were quantifiedat 0, 7 (bolting stage), 14 (flower buds raised above the youngest leaves), 21 (first petals visible, but flower buds still closed) and 28 (flowering)days after the beginning of bolting (D0). Concerning the soluble proteins and the other N compounds, only one biological replicate remained atD28, and its value is indicated by a cross (x). Data are indicated as the mean value ± standard error (vertical bars). Letters a, b and c representdifferences in kinetics, asterisks indicate significant differences between treatments and hashes represent significant differences betweengenotypes (n = 4 plants; p < 0.05).
Girondé et al. BMC Plant Biology (2015) 15:59 Page 10 of 21
chlorophyll content decreased at D7 and remained con-stant for Aviso until D28, while it decreased all alongthe experiment for Oase. In response to LN treatment,a decrease in chlorophyll content was observed fromD21 for both genotypes (Figure 7A). In HN conditions,the N amount in the source leaf was 2-fold higher for
Aviso than for Oase and remained nearly constant dur-ing the 28 days for both genotypes (Figure 7B). Inresponse to LN conditions, the leaf N amount de-creased significantly from D21 in Aviso alone, resultingin a decline of 83% (−6.13 mg N) between D0 and D28(Figure 7B).

Girondé et al. BMC Plant Biology (2015) 15:59 Page 11 of 21
Considering the different N fractions, it is noteworthythat whatever the genotype, a similar pattern of solubleprotein degradation (strong decrease at D7) wasobserved for both N conditions. However, the only avail-able biological replicate at D28 (n = 1) suggested a loweramount of soluble proteins at D28 in LN plantscompared with HN for both genotypes (Figure 7C). Eventhough the level of free amino acids was 1.7-fold higherin Aviso than in Oase, it decreased strongly in bothgenotypes at D28 in HN conditions (Figure 7D). In LNconditions, a lower amount of N-amino acids wasobserved for both genotypes at D21 compared with theHN treatment. Surprisingly, the fraction correspondingto the other N compounds increased in Aviso betweenD0 and D14 in both N conditions (+64% and +80% inHN and LN, respectively; Figure 7E). Thereafter, thefraction of other N compounds in the source leaf ofAviso remained almost stable in HN plants while it de-creased strongly in LN plants (−82% between D14 andD28). Concerning Oase, the other N compoundsremained low and constant during all the experiments,whatever the N supply.The activities of glutamate dehydrogenase (GDH) and
glutamine synthetase (GS), involved in the metabolismand remobilization of amino acids during senescence,were similar and constant in both N conditions forAviso, except for a 50% decrease in GS activity at D28 inboth N conditions and a putative 6-fold increase (n = 1)of GDH activity at D28 in LN conditions (Figure 8A andB). Concerning Oase, the GDH activity remained lowuntil D21 and increased by 1.5-fold at D28 in both Nconditions (Figure 8A). In HN plants, the GS activityremained constant until D21 and decreased by 43% atD28 (n = 1). In LN plants, the GS activity increased(1.5-fold) at D21 and putatively decreased at D28 (n = 1,Figure 8B). The immunoblots of cytosolic (GS1) andchloroplastic (GS2) glutamine synthetase (Figure 8C)revealed that the proportion of GS1 was not impactedby LN treatment in both genotypes, but it remainedhigher for Oase throughout the experiment (53% forOase and 39% for Aviso on average).
Proteolytic activities in the source leavesDifferent protease inhibitors (against cysteine, serine,aspartic or metallo- proteases and proteasome) wereused in order to identify the class of proteases involvedin the strong degradation of soluble proteins occurringin the source leaf at D7 (Figure 7C). Due to the fact thatRubisco represents a large proportion of the soluble pro-teins in leaves [26], the characterization of proteases wasdetermined via the analysis of the degradation of theRubisco large subunit (LSU, Figure 9A). In source leaf ofHN plants (Figure 9B), the LSU proteolysis was stronglyinhibited by iodoacetamide for Aviso and Oase (35.5 and
31.3% of inhibition, respectively) and by MG132 forOase (46.22% of inhibition), suggesting that the proteoly-sis is mainly carried out by cysteine proteases for Avisoand by the proteasome and cysteine proteases for Oase.The contribution of cysteine and aspartic proteases tothe LSU degradation in Aviso was the same in both Nconditions, while a slight increase of serine proteases(from 6.1 to 12.1% of inhibition; p = 0.12) and metallo-proteases (from 7.9 to 14.1% of inhibition; p = 0.17) anda significant increase of proteasome activity (60% ofinhibition) were observed (Figure 9B) in response to theLN treatment. The contribution of proteasome, cysteineand serine proteases for Oase remained similar in bothN conditions. Compared with HN plants, the contribu-tion of aspartic proteases decreased (4% of inhibition)while the participation of metalloproteases increased(27% of inhibition) in Oase LN plants (Figure 9B).
DiscussionEvidence of two different N management strategies thatlead to a similar seed yield under ample nitrate nutritionThe 15N-labelling method performed in this studyallowed the distinction between endogenous and ex-ogenous N fluxes and, therefore, the relevant determin-ation of the involvement of N uptake and remobilizationduring seed filling. Despite no significant differences inseed yield (Figure 1), the two genotypes of winter oilseedrape investigated in the present work showed contrast-ing N management during seed filling under amplenitrate supply. In fact, even if both genotypes used ex-ogenous and endogenous N for seed filling (Figure 3),Oase had the highest NUpE (82%) while Aviso had thehighest NRE (32%) during seed filling (Table 2). Ourdata suggest that allocation of N taken up into seeds ismore efficient in Oase than Aviso, while the processes ofN remobilization from source organs towards seeds dur-ing the final reproductive phases is higher in Aviso. Infact, during seed filling, the total N remobilization was3-fold higher for Aviso than for Oase, which was a con-sequence of a higher remobilization from leaves, stemsand pod walls (Figure 3), leading to a higher amount ofN redistributed towards seeds (Figure 2C and D). Theseresults confirm that leaves, stems and pod walls are themain source organs for N remobilization after flowering[8,16,17,20,46].
Under nitrate limitation, N management in Aviso is moreefficient than Oase for seed fillingDespite an increase of NHI, a decrease of seed yield wasobserved for Oase and Aviso in response to nitrate limi-tation (LN treatment) applied at the early bolting stage(Figure 1, Table 1). The seed oil percentage increased inresponse to LN conditions for both genotypes (Table 1)but the nutritional quality of oil of both genotypes was

Girondé et al. BMC Plant Biology (2015) 15:59 Page 12 of 21
affected: increase of (i) C18:2/C18:3 ratio for Oaseand (ii) proportion of erucic acid for Aviso (Table 1).These results confirm the strong relationshipbetween N supply and oil quality [47] and suggesta genotypic variability about the oil quality in responseto N limitation.
Figure 8 Changes in glutamate dehydrogenase activity, glutamine syPlants were supplied with ample (HN, 3.75 mM) or low (LN, 0.375 mM) nitrleaf’, determined as mature at D0 (early bolting) and becoming senescentA) was quantified as the synthesis of glutamate and is expressed as nmol o(GS; B) was determined by the nmol of glutamine produced.h−1.μg−1 proteblotting with specific antibodies and the percentage of GS1 among the tobolting), 14 (flower buds raised above the youngest leaves), 21 (first petalsbeginning of bolting (D0). Only one biological replicate remained at D28, aindicated as the mean value ± SE (vertical bars). Letters a, b and c representreatment and hashes represent significant differences between genotypes
As expected, in response to LN treatment, the N up-take was lower throughout the experiment (Figures 4, 5and 6) and the total N amount in seeds (mg N in totalseeds) decreased for both genotypes (Table 1), leading toa lower protein content in parallel to the increase in oilcontent [48-50]. Nevertheless, a significant N uptake
nthetase activity and their amounts (GS1, GS2) in a source leaf.ate concentrations. These data were obtained from a selected ‘sourceduring the experiment. The activity of glutamate dehydrogenase (GDH;f NADH used.h−1.μg−1 proteins. The activity of glutamine synthetaseins. The GS1 and GS2 amounts (C) were quantified after westerntal GS amount was estimated. Data were observed at 0, 7 (earlyvisible, but flower buds still closed) and 28 (flowering) days after thend its value is indicated by a cross (x). In panels A and B, data aret differences in kinetics, asterisks mean significant differences between(n = 4 plants; p < 0.05).

Girondé et al. BMC Plant Biology (2015) 15:59 Page 13 of 21
occurred during seed filling despite the low N supply,especially in Oase (Figure 6), supporting the fact thatthere is a significant N uptake capacity after flowering inoilseed rape [16,17]. Interestingly, growth of new leaveswas observed during seed filling in response to LNconditions (Figure 6). Even if these new organs representa sink for N, new leaves may help maintaining photosyn-thetic capacities during seed filling and raise the poten-tial seed yield by increasing the allocation of C to seeds[7,51]. In addition, photosynthetically active leaves couldparticipate in the improvement of N uptake [8], as previ-ously suggested for winter oilseed rape [52]. Indeed, theN uptake is associated with the availability of photoassi-milates in the roots [53]. Consequently, the higher Nuptake of Oase during seed filling (Figure 6) may be
Figure 9 Rubisco large subunit degradation in a source leaf with or wactivities by protease inhibitors (B). The Rubisco large subunit (LSU) in tis visualized on stain free SDS-PAGE and quantified for the four biological repeat 37°C (t1h) without inhibitors (PE, control conditions) or with specific proteas(PE + SPI; serine protease inhibitor), methanol (PE +Me), methanol and 1–10 ppepstatin A (PE +Me+ API; aspartic protease inhibitor), DMSO (PE + DMSO) orrepresentative biological repetition is shown in panel A and the percentage opresents the inhibition of the protease activities by the proteases inhibitors (epanel B, data are indicated as the mean value ± SE. An asterisks means that th# means a significant differences between genotypes (n = 4 plants; * or #= p <
related to the significant higher C content observedunder N limitation in Oase roots at D70 and D99 (+4%and +5% respectively compared with Aviso; data notshown).As in HN conditions, N from uptake is mainly used
for seed filling in Oase, but due to the low N supply, alower N amount is distributed to seeds compared withAviso (Figure 6). In addition, even though Oase had thehighest N uptake during seed filling, Aviso had the high-est total N uptake from the bolting to mature seedsstages (75 mg N) in comparison with Oase (58 mg N)(Figures 4, 5 and 6). This result agrees with the previousstudies reporting that the genotypic variability of Nuptake is mainly observed under restricted N supply[11,15]. However, because both genotypes have the same
ithout protease inhibitors (A) and the inhibition of the proteasehe soluble protein extract (PE) of the source leaf (7 days after bolting)titions by Image Lab software (Bio-Rad) at (t0) and after 1 h of incubatione inhibitors: iodoacetamide (PE + CPI; cystein protease inhibitor), aprotininhenanthroline (PE +Me+MI; metalloprotease inhibitor), methanol andDMSO and MG132 (PE + DMSO + PI; proteasome inhibitor). The mostf degradation (mean value ± SE, n = 4 plants) are indicated below. Panel Bxpressed as % of LSU degradation observed in control conditions (PE)). Ine LSU degradation is significantly different between N treatment and0.05).

Girondé et al. BMC Plant Biology (2015) 15:59 Page 14 of 21
seed yield under LN conditions (Figure 1), the positivecorrelation between N uptake and seed yield demon-strated in previous works on oilseed rape [9,11,12,15]was not confirmed in our experimental conditions. Thehigher N amount in seeds of Aviso could be due to thefact that, as in HN treatment, endogenous N is mainlyused for seed filling. Indeed, Aviso had a higher NREduring seed filling (Table 2), leading to a higher redistri-bution of the N remobilized from leaves and stemstoward seeds in comparison with Oase (Figure 6). Inorder to better understand the importance of N remobi-lization from leaves and stems in Aviso with its superiorNRE, these organs were studied in detail.
Impact of leaf N remobilization and its associated cellularprocesses in coping with N limitationAs previously observed for oilseed rape cv. Capitol[17,20], leaves are the main source organs between bolt-ing and early pod formation, whatever the genotype orthe N supply (Figure 4 and Additional file 1). In fact, themaintenance of the amount of N distributed to siliquesin Aviso under LN conditions is linked to an increase inN remobilization from source leaves (Figure 4A) associ-ated with an increase in the number of dead leaves(Additional file 6). The N remobilization in leaves ofOase (Figures 4B and 7B) and the number of dead leaves(Additional file 6) were similar in both N conditions,suggesting a similar progression of senescence for thisgenotype whatever the N supply, which has never beenhighlighted previously.The mechanisms involved in N remobilization were
then investigated in a selected source leaf, becomingsenescent between the bolting and the flowering stages.Proteins are the main form of N storage in source leaves,especially the soluble protein Rubisco [54], and for bothgenotypes a massive degradation of the soluble proteinsoccurs 7 days after bolting under both N conditions(Figure 7C). The aspartic and serine proteases seem tocontribute at a low level to the proteolysis in bothgenotypes (Figure 9B, Additional file 7), contrary toprevious results reported for Arabidopsis [24]. Our studyreveals a higher involvement of metalloproteases duringN remobilization in the source leaves of Oase inresponse to LN supply (Figure 9B). These results areconsistent with previous proteomics studies whichshowed the induction of metalloproteases (FtsH) duringleaf senescence at the vegetative stages of winter oilseedrape (cv. Capitol) in response to limitation/privation innitrate [31]. Because FtsHs are involved in the proteoly-sis of D1 or Lhcb3 proteins localized in the thylakoidmembrane [55], our results suggests that a nitrate limita-tion may lead to an improvement in the degradation ofthylakoid-bound proteins.
Except for Aviso in HN conditions, the classes of pro-teases having the greatest involvement in the massivein vitro proteolysis observed 7 days after bolting arecysteine proteases and the proteasome system(Figure 9B). The importance of cysteine proteases in leafproteolysis is in agreement with previous studies onwinter oilseed rape demonstrating that the cysteine pro-tease SAG12 is highly induced at the transcriptomic [56]and proteomic [31] levels during leaf senescence. The ef-ficient N remobilization observed for Oase in both Nconditions and for Aviso in LN treatment seems to becorrelated to a high contribution of the proteasome inthe proteolysis mechanisms (Figure 9B), which confirmthe previous studies on oilseed rape [31] and Arabidopsis[24]. In addition, in our in vitro conditions, the prote-asome seems to be able to degrade the LSU of Rubisco.However, in cell, the proteasome complex is involved indegradation of proteins previously tagged with ubiquitinand is mainly localized in the nucleus and cytosol(around pH 7.5, [57]). Some studies in Arabidopsisshown that Rubisco is degraded in acidic organelles suchas lytic vesicles or vacuoles (around pH 5, [58-60]) and/or directly in the stroma (characterized by a pH of 7.5and a low amount of ubiquitin [61]). Taken together,these studies suggest that Rubisco is probably notdegraded in planta by the proteasome system. However,a chloroplastic E3 enzyme, involved in the selectivity ofthe ubiquitin-proteasome system during plastid forma-tion was recently found in the outer membrane of chlo-roplasts [62]. In addition, the proteolysis by theproteasome can occur without the involvement of ubi-quitin in animal cells [63], suggesting that could alsohappen in plant cells. Consequently, the involvement ofthe proteasome pathway in the degradation of chloro-plastic proteins seems to be possible and should beinvestigated further.The efficient degradation of soluble proteins observed
in the senescing leaf of Aviso under both N conditions(Figure 7C) was not correlated with a concomitant de-crease in the total N amount (Figure 7B). In our study,there was no transient accumulation of N in the aminoacid fraction (Figure 7D), confirming that the export ofamino acids was not limiting, as indicated in earlierresults obtained for winter oilseed rape cv. Lirajet [44].Consequently, despite the decline in N-soluble proteins,the stability of total N in senescing leaf was relatedto a transient accumulation of other N compounds(Figure 7E), including transmembrane proteins, peptides,nitrate and/or ammonia. In addition, the degradationand/or export of these other N compounds seem to beresponsible of the higher N remobilization of the sourceleaf observed at D21 under LN conditions (Figure 7E).In Aviso LN plants, the high decrease of other N com-pounds at D21 is followed by an increase of the

Girondé et al. BMC Plant Biology (2015) 15:59 Page 15 of 21
amination activity related to GDH (Figure 8A), whichcatalyses the incorporation of ammonium on α-ketogluratate to produce glutamate in case of accumula-tion of ammonium [64] released by the catabolism ofproteins and peptides [65]. The better degradation oftransmembrane proteins and peptides in LN conditionscould lead to an ammonium accumulation and, there-fore, may explain the increase of amination by the GDHin order to detoxify the cell and produce glutamate,which is found in phloem sap of oilseed rape [33]. If thishypothesis is true, the lower level of other N compoundsobserved in source leaf of Oase in both N conditions(Figure 7B) could be due to an efficient degradation oftransmembrane proteins and/or peptides into aminoacids. The amino acids are not accumulated in thesource leaf (Figure 7D), suggesting that they are wellexported towards the phloem vessels. This could be re-lated to the fact that GS1, the cytosolic isoform of GSwhich is known to be involved in N remobilization pro-cesses [36], is present in higher proportions in thesource leaf of Oase than in Aviso one (Figure 8C). Thesedata suppose that an efficient degradation of transmem-brane proteins and/or peptides and a rapid conversionof amino acids into their transportable forms (such asglutamate and glutamine) by enzymes like GDH or GS1in order to enhance their exportation from the meso-phyll cells towards the phloem is an important physio-logical trait for the optimization of NRE. These resultshave to be confirmed and other enzymes involved in theconversion of amino acids, such as asparagine synthetasein Arabidopsis [32], have to be investigated in oilseedrape leaves.
Stems play a pivotal role in N management as a transientN storage organIn LN supply, the highest N amount in seeds of Avisocompared with Oase is linked to a large N remobiliza-tion (3-fold higher) from the stem to seeds during seedfilling (Figure 6). As previously observed [9], the highestN remobilization is not related to a lower residual N inthe stem at D99 (Additional files 3 and 4), but to a sig-nificant higher distribution of N towards the stem beforeseed filling (Figures 4 and 5). Consequently, the increaseof N storage capacity in stems before the onset of seedfilling may significantly improve the N remobilizationfrom stems towards seeds. The same conclusion wasobserved about leaves during seed filling. However, whileleaves were crucial source organs before early silique for-mation, their participation remained low during seed fill-ing (only 7 and 5% of total N remobilization for Avisoand Oase, respectively; Figure 6). A high N remobiliza-tion from leaves towards the stem was previously associ-ated with a high NUE in spring oilseed rape [23]. Takentogether, results suggest that the stem is a crucial
buffering organ for transient N storage in order to com-pensate the delay between the large N remobilizationfrom leaves and the sink N demand for seed filling.
ConclusionsAs previously observed under restricted N supply, thehigher N uptake from bolting to flowering is associatedwith the most efficient genotype for seed N filling (Avisoin our study). Nevertheless, under field conditions, themineral N availability strongly fluctuates and is usuallylow during seed filling, meaning that a higher N uptakecould not be sufficient to significantly improve the NUE.The investigation of the contrasting N management inAviso and Oase genotypes confirms that remobilizationduring monocarpic senescence is efficient for leaves andstems. Consequently, an efficient remobilization andutilization of N taken up and N stored before the onsetof seed filling is probably one of the main physiologicaltrait for attaining a high NUE in an agronomic contextof mineral N restriction. Up to the early phase of seedfilling, N is mainly stored in leaves, and therefore an effi-cient foliar N remobilization is important to limit N lossfrom dead leaves and improving seed N filling togetherwith a limitation of pollution risk. Our findings suggestthat an efficient foliar NRE is mainly associated to (i) ahigh proteolysis by the cysteine proteases and prote-asome system and (ii) a strong coordination betweenproteolysis, the export/degradation of oligopeptides andthe export of amino-acids (involving the enzymes GSand GDH). Future developments are needed to confirmthe relevance of these classes of proteases before theirpotential use in a “candidate protein-strategy” to identifyQTLs related to high NRE. Our study also highlightsthat the improvement of NUE and seed N filling underlow mineral N availability is highly related to an efficientstorage of the N remobilized from leaves into the stem.Thus, it is likely that the stem may act as a transientstorage organ in case of asynchronism between themassive remobilization of N coming from source leavesand its further utilization by seeds. Such physiologicaltraits (efficient leaf N remobilization, transient N storagein stems) may be particularly relevant to breedingprograms aimed at identifying Brassica napus genotypeshaving high NUE in response to the fluctuation of ni-trate availability and low N fertilizer inputs.
MethodsGreenhouse experimentationTwo genotypes of winter oilseed rape (Brassica napus L.cv. Aviso and Oase) were cultivated in greenhouse con-ditions (Additional file 5). Plantlets were supplied for thefirst 2 months with a 25% Hoagland solution (1.25 mMCa(NO3)2 4H2O; 1.25 mM KNO3; 0.5 mM MgSO4;
0.25 mM KH2PO4; 0.2 mM EDTA-2NaFe-3H2O; 14 μM

Girondé et al. BMC Plant Biology (2015) 15:59 Page 16 of 21
H3BO3; 5 μM MnSO4; 3 μM ZnSO4; 0.7 μM (NH4)6Mo7O24; 0.7 μM CuSO4; 0.1 μM CoCl2). Thereafter,plants were vernalized for 8 weeks under a long photo-period and low temperature (day: 10 h, 10°C; night:14 h, 4°C; 90 mL of 25% Hoagland solution per plantrenewed twice a week). During pre-culture andvernalization, light was supplied by High PressureSodium Lamps (Philips, MASTER GreenPower T400W)with a PAR (Photosynthetically Active Radiation) of400 μmol photon.s−1.m−2 at the top of the canopy. Aftervernalization, plants were transferred to a greenhouse inpots containing perlite and vermiculite (2:1, v/v; oneplant per pot; 40 plants per square metre), and exposedto a thermoperiod of 20°C (day) and 15°C (night). To es-timate the N fluxes at the whole plant level, long termpulse chase 15N-labelling (at 2% atom excess) wasapplied from the first days of germination until the earlybolting stage (Day 0 of the experiment (D0) correspond-ing to GS31, a growth stage determined from the scaleof BBCH (Biologische Bundesanstalt, Bundessortenamtund CHemische Industrie)). At D0, 15N-NO3 labellingwas stopped and two N treatments with unlabelled-NO3
were applied until the final harvest stage (D99; corre-sponding to GS99 i.e. the final stage of developmentwith mature seeds and the whole plant in senescence):high nitrate (HN, 3.75 mM) and low nitrate (LN,0.375 mM). Each day the nutrient solution was suppliedautomatically in an increasing volume as a function ofthe growth stages: 90, 120, 150, and 180 mL per plant atthe start of the bolting, visible bud, flowering, and seedmaturation stages, respectively.Plants were harvested at different growing stages: D0
(GS31), D7 (GS32: bolting stage), D14 (GS53: flowerbuds raised above the youngest leaves), D21 (GS59: firstpetals are visible), D28 (GS60: flowering), D42 (GS70:end of flowering), D70 (GS73: pod filling) and D99(GS99: mature seeds, whole plant in senescence). Plantswere separated into roots (including taproot and second-ary roots), leaves, stem (including the ramifications),siliques, seeds and pod walls (when siliques were dehis-cent). The dry matter of each organ was determinedafter freeze-drying. The thousand seed weight and thetotal seed number were deduced from the weight of onehundred seeds after 48 h at 30°C.In order to analyse foliar N remobilization, leaves were
classified into three groups (dead leaves, old +matureleaves and young leaves) on the basis of their nodal pos-ition, their leaf area was determined by a LI-COR 300area meter (LI-COR, Lincoln, NE, USA) and theirchlorophyll content measured by SPAD (Soil PlantAnalysis Development; Minolta, SPAD-502 model).Throughout the experiment, dead leaves were countedand harvested. Moreover, at D0, a mature leaf that hadbecome senescent during the experiment (i.e. a “source”
leaf for N) was tagged and studied in order tocharacterize the processes associated with leaf N remo-bilization from bolting (D0) to flowering (D28) (seeAdditional file 5 for more details about the selection ofthese source leaves). One half of this mature leaf wasfreeze-dried, weighed for dry matter and ground to afine powder for further biochemical analyses. The otherhalf of the mature leaf was immediately frozen in liquidnitrogen and stored at −80°C until further protein andmolecular analyses.
Determination of oil content, fatty acid composition andprotein contents in seedsAs previously described by Dubousset et al. [66], seedsamples (about 3 g) were scanned on a monochromatorNear Infra-Red System (NIRSystem model 6500, FOSSNIRSystem Inc, Silver Spring, MD, USA) equipped withthe transport module, in the reflectance mode. Theresults were predicted from an external calibrationestablished for oil content and composition of fatty acids(CRAW, Gembloux, Belgium). Three determinationswere performed for each biological replicate.
Total N and 15N quantification, whole plant N fluxes anddetermination of NUE componentsThe total N and 15N amounts in each organ were ana-lysed by an elemental analyser (EA3000, EuroVector,Milan, Italy) linked to a continuous flow isotope massspectrometer (IRMS, IsoPrime GV instruments,Manchester, UK). Due to the long term pulse chase label-ling described below, the 15N amount is representative ofthe total N amount in every organ; the variation of the15N amount allows estimation of the endogenous N fluxesrelated to N remobilization and the exogenous N fluxcoming from uptake. The determination of source or sinkstatus of leaves or other organs was done on the basis oftheir 15N amount, where a sink organ is characterized by again of 15N and a source organ by a decrease in the 15Namount. The calculation of the whole plant N fluxes aredescribed in detail in Salon et al. [67].The N use efficiency (NUE) was calculated as:
NUE ¼ QNseed= QNwhole plant� �
= DMseed= DMwhole plant� �
where QNseed corresponds to the N amount (mg) inseeds, QNwhole plant to the N in the whole plant (includ-ing roots and seeds), DMseed to the seed dry matter (g)and DMwhole plant refers to the whole plant dry matter(g) observed at the final stage (including roots and seeds;GS99).The N utilization efficiency (NUtE) corresponds to the
seed dry matter (DMseed) production per g of N inshoots (QNshoot) and was estimated by the followingcalculation:

Girondé et al. BMC Plant Biology (2015) 15:59 Page 17 of 21
NUtE ¼ DMseed=QNshoot
Based on the calculation of endogenous and exogen-ous N fluxes, the N remobilization efficiency (NRE; totalremobilized N amount relative to the total N stored insource organs) and N uptake efficiency (NUpE; Namount allocated to seeds from uptake relative to totalN distributed to seeds) were defined as follows (as apercentage):
NRE ¼ ΣQNremobilization�100 = ΣQNsourcet þ ΣQNuptake
� �
NUpE ¼ QNSuptake� 100 = QNSdistributed
where ΣQNremobilization corresponds to the total Namount remobilized between t and t + Δt, ΣQNsource t isthe N amount in all source organs at t and ΣQNuptake
corresponds to the amount of N taken up that is allo-cated to all source organs between t and t +Δt. TheQNSuptake corresponds to the N amount allocated toseeds from the uptake and the QNSdistributed to the totalN amount distributed to seeds (from uptake andremobilization).
Analysis of N-soluble proteins, N-free amino acids andother N compounds in leavesSoluble proteins were extracted from 200 mg of frozenfresh matter previously ground in a mortar with liquidnitrogen and 500 μL of citrate-phosphate buffer (20 mMcitrate, 160 mM phosphate, pH 6.8 containing 50 mg ofpolyvinylpolypyrrolidone (PVPP)). After centrifugation(1 h, 12 000 g, 4°C), the resulting supernatant, containingthe soluble proteins (protein extract, PE), was trans-ferred to a microtube and completed to 500 μL. Theconcentration of the soluble proteins was determined inan equivalent of bovine serum albumin (BSA) usingprotein-dye staining [68]. To obtain the protein amountin equivalent nitrogen, an average N content of 16% wasapplied. For amino acid extraction, 1 mL of sodiumphosphate 100 mM (pH 7.5) was added to 25 mg of leafpowder obtained after grinding of freeze-dried leaf sam-ples. The mixture was incubated twice (30 min at 80°C)with 1 mL of 80% ethanol and 1 mL of 50% ethanol, re-spectively. After each incubation, the resulting mixtureswere centrifuged (10 min, 12 000 g, 4°C) and the result-ing supernatants containing amino acids were pooled,evaporated and re-suspended in 500 μL water. To quan-tify the amino acid concentration, 10 μL of amino acidextract were added to 90 μL of water and 1 mL of nin-hydrin reagent (112 mM ninhydrin and 3.85 mM tinchloride in 100 mM citrate buffer pH 5, 50% (v/v)DMSO). After incubation for 20 min in boiling waterand 10 min on ice, 5 mL of 50% ethanol were added andthe absorbance was read at 570 nm. The amino acidconcentration was calculated using L-leucine as
standards. The N-amino acid amounts were deduced byusing an average value for molecular mass (110 g.mol−1)and the N ratio (7.86) obtained from previous studies inleaf samples of oilseed rape was used to determine theN-amino acid amounts. In order to determine theamount of other N compounds (including insoluble pro-teins, oligopeptides, nitrate and ammonia), the N-solubleprotein and N-amino acid amounts have been subtractedfrom the total N amount.
Analysis of glutamate dehydrogenase (GDH) andglutamine synthetase (GS) in the source leafFor GS and GDH analysis, proteins were extracted from150 mg of fresh matter of leaves (previously ground in amortar with liquid nitrogen) with 1 mL of extractionbuffer (10 mM Na-EDTA, 10 mM MgCl2, 250 mMTris–HCl pH 7.6, 13.3 mM β-mercaptoethanol and2 mM leupeptine) containing 50 mg of polyvinylpyrroli-done (PVP). The GDH activity in the sense of glutamatesynthesis (expressed in nmol of NAD(H) used.h−1.μgprotein−1) was performed as previously described byMasclaux et al. [69]. The same protein extract was usedto determine the GS activity (nmol glutamine.h−1.μgprotein−1) by the method of O’Neal and Joy [70]. Inorder to monitor the kinetic evolution of GS1 and GS2,a western blot was performed. For this, the four bio-logical replicates of protein extracts were pooled and10 μg were denatured with Laemmli 2X buffer [71] con-taining β-mercaptoethanol (5% (v/v)). The proteins wereseparated on an SDS-PAGE gel (5.5% polyacrylamide(w/v) for the stacking gel and 10% polyacrylamide (w/v)for the resolving gel) and transferred to a polyvinylidenedifluoride (PVDF) membrane. The protein detection ofboth GS (GS1 at 39 kDa and GS2 at 44 kDa) was madeusing rabbit polyclonal antibodies (1/100e) derived byEurogenetec (Seraing, Belgium) against the peptideAYGENERRLTG. The antibodies were detected by sec-ondary antibodies from goat (1/12 000e) coupled with al-kaline phosphatase. The relative amount of GS1(cytosolic isoform) and GS2 (chloroplastic isoform) wasquantified by Image Lab software (Bio-Rad).
Analysis of protease activities involved in Rubisco largesubunit (LSU) degradation in the source leafIn order to identify the proteases involved in the degrad-ation of the Rubisco LSU, a new method using stain freegels (Bio-Rad®) was performed. These gels allow the de-tection of proteins immediately after the end of theSDS-PAGE and in less than 2 min, and the method isbased on a UV-induced trihalo-compound modificationof tryptophan residues contained in proteins (for details,see Kazmin et al. [72]). In order to identify the proteaseclasses involved in the LSU degradation, the soluble pro-tein extract (PE, 6 μg) obtained from source leaves at D7

Girondé et al. BMC Plant Biology (2015) 15:59 Page 18 of 21
was either incubated for 1 h at 37°C (t1h) or not (t0) insodium acetate buffer 250 mM (pH 5, 0.16% β-mercaptoethanol (v/v)) with or without specificprotease-class inhibitors. Iodoacetamide (14.5 mM) wasused as an inhibitor of cysteine proteases (PE + CPI) andaprotinin (34 μM) was the serine proteases inhibitor(PE + SPI). For analysis of metalloprotease and aspartateprotease activities, 5.5 mM of 1–10 phenanthroline(PE +Me +MI) and 10.2 μM of pepstatin A (PE +Me +API) were used, respectively. Finally, 40 μM of MG132(benzyloxycarbonyl-leu-leu-leucinol) were used to in-hibit proteasome (PE + DMSO + PI). Due to the solubil-ity of 1–10 phenanthroline and pepstatin inhibitors in1% methanol, the total protease activity was also quanti-fied with 0.5 % methanol (v/v; PE +Me). In the sameway, due to the solubility of MG132 in DMSO, the totalprotease activity was carried out with 0.5 % DMSO (PE+ DMSO). At t0 or after incubation for 1 h at 37°C (t1h),Laemmli 2X buffer [71] containing β-mercaptoethanol(5% (v/v)) was added to the protein samples (one volumeof Laemmli 2X buffer per volume of proteins) beforeprotein denaturation for 7 min in boiling water. Theresulting samples were loaded on SDS-PAGE (Mini-PROTEAN TGX Stain Free Gels 4-15%, Bio-Rad®) andthe LSU amount at t0 (LSUt0) and after 1 h of incubation(LSUt1h) were visualized and quantified using Image Labsoftware (Bio-Rad®). The percentage of LSU degradation(% Deg) with or without protease inhibitors was firstlydetermined and used to deduce the inhibition of LSUproteolysis (% Inh) by the different protease inhibitors asfollows:
% Deg ¼ LSUt0− LSUt1hð Þ �100 = LSUt0
%Inh ¼ LSUPEð Þ – LSUPEþInhð Þ �100 = LSUPEð Þwhere LSUPE is the amount of LSU degraded without
inhibitor and LSUPE+Inh, is the amount of LSU degradedin the presence of protease inhibitors.
Statistical analysisThe normality of the data was studied with the Ryan-Joiner test at 95%. Analysis of variance (ANOVA) andthe Tukey test were used to compare the means. Whenthe normality law of data was not respected, thenon-parametric test of Kruskal-Wallis was carried out.Statistical significance was postulated at P < 0.05. Fourbiological repetitions corresponding to four differentplants (n = 4) were used and the data presented here areexpressed ± standard error (SE).
Additional files
Additional file 1: Dry matter of every organs of Aviso and Oase inLN and HN conditions. The data are expressed in grams. The plants
were cultivated in ample (HN, 3.75 mM) or low (LN, 0.375 mM) nitrateconcentrations and the data obtained at D0 (early bolting stage), D42(pod formation), D70 (start of seed filling) and D99 (mature seed stage)are presented in this table. Grey box means that the organ was absent atthis growing stage. Letters a, b, c represent the significant differences oforgans between two harvest dates, the asterisks indicate significantdifferences between N treatments and hashes indicate differencesbetween genotypes (n = 4 plants; p < 0.05).
Additional file 2: N amount of every organs of Aviso and Oase inLN and HN conditions. The data are expressed in milligrams. The plantswere cultivated in ample (HN, 3.75 mM) or low (LN, 0.375 mM) nitrateconcentrations and the data obtained at D0 (early bolting stage), D42(pod formation), D70 (start of seed filling) and D99 (mature seed stage)are presented in this table. Grey box means that the organ was absent atthis growing stage. Letters a, b, c represent the significant differences oforgans between two harvest dates, the asterisks mean significantdifferences between N treatments and hashes mean differences betweengenotypes (n = 4 plants; p < 0.05).
Additional file 3: N fluxes in Aviso (A) and Oase (B) in HN conditionbetween D0 and D42. The plants were supplied with highconcentration of nitrate (HN, 0.375 mM of nitrate) and D0 corresponds toearly bolting and D42 to pod formation. Fluxes of N from remobilizationor uptake in the different organs are expressed as mg of N remobilizedor taken up, respectively. For fluxes of N remobilisation, N amount isindicated with a minus sign (-) when N is remobilized from a sourceorgan while it is indicated with a plus sign (+) when remobilized N isredistributed towards a sink organ. Asterisks represent significantdifferences between genotypes (n = 4 plants; *= p < 0.05).
Additional file 4: N fluxes in Aviso (A) and Oase (B) in HN conditionbetween D42 and D70. The plants were supplied with high concentrationof nitrate (HN, 0.375 mM of nitrate) and D42 corresponds to pod formationand D70 to the start of seed filling. Fluxes of N from remobilization or uptakein the different organs are expressed as mg of N remobilized or taken up,respectively. A shaded box means that the organ is not present during thesegrowing stages. For fluxes of N remobilisation, N amount is indicated with aminus sign (-) when N is remobilized from a source organ while it is indicatedwith a plus sign (+) when remobilized N is redistributed towards a sink organ.Asterisks represent significant differences between genotypes (n= 4 plants;*= p< 0.05).
Additional file 5: Description of the experimental design. Twogenotypes of winter oilseed rape (Brassica napus L. cv. Aviso and Oase)were cultivated in greenhouse conditions. After sterilization, seeds weresowed on watered vermiculite and when the first leaf appeared, plantletswere supplied during 2 months with a 25% Hoagland solution (for detailssee Material and Methods). Thereafter, plants were vernalized during8 weeks and were transferred under greenhouse in pot containing perliteand vermiculite with a thermoperiod of 20°C (day) and 15°C (night). Toestimate the N fluxes at whole plant level, a long term pulse chaselabelling (at 2% atom excess) was applied from 43 days after sowing untilearly bolting growing stage (Day 0 of the experiment (D0) correspondingto GS31), including the vernalization period (8 weeks). At D0, 15N-NO3
labelling was stopped and two N treatments with unlabelled-NO3 wereapplied until the final harvest (D99; mature seeds i.e. GS99 stage): highnitrate (HN, 3.75 mM) and low nitrate (LN, 0.375 mM). Plants wereharvested at different growing stage. Before the start of N limitation (D0),one mature leaf undergoing senescence (called “source leaf“, see pictureshowing the source leaf selected in Aviso plants) was studied in detailsfor analysis of the physiological and molecular processes involved in leafN remobilization. The “source leaf” was selected on the following criteria:leaf that attained its maximal expansion (leaf area), with a high chlorophyllcontent and having the same position in the canopy in both genotypes. Theleaf biomass at D0 (3.03 ± 0.06 g FW for Aviso vs 2.77 ± 0.13 g FW for Oase),leaf area from D0 to D28 (103.57 ± 4.71 cm2 for Aviso vs 99.45 ± 3.89 cm2 forOase) and chlorophyll content at D0 (SPAD value of 51.25 ± 0.71 for Aviso vs52.53 ± 1.28 for Oase) in source leaves were not significantly different betweenAviso and Oase.
Additional file 6: Number and residual N in dead leaves of Avisoand Oase. The plants were cultivated in ample (HN, 3.75 mM) or low(LN, 0.375 mM) nitrate concentrations and the data obtained at D0 (early

Girondé et al. BMC Plant Biology (2015) 15:59 Page 19 of 21
bolting stage), D42 (pod formation), D70 (start of seed filling) and D99(mature seed stage) are presented in this table. Asterisks representdifferences between N treatments and hashes differences betweengenotypes (n = 4 plants; *= p < 0.05).
Additional file 7: Rubisco large subunit degradation in a selectedsource leaf with or without protease inhibitors. The Rubisco largesubunit (LSU) in the soluble protein extract (PE) of the source leaf (7 daysafter bolting) is visualized on stain free SDS-PAGE and quantified for thefour biological repetitions (A, B, C and D) by Image Lab software (Bio-Rad)at (t0) and after 1 h of incubation at 37°C (t1h) without inhibitors (PE) orwith iodoacetamide (PE + CPI; cystein protease inhibitor), aprotinin(PE + SPI; serine protease inhibitor), methanol (PE + Me), methanol and1-10 phenanthroline (PE + Me + MI; metalloprotease inhibitor), methanoland pepstatin A (PE + Me + API; aspartic protease inhibitor), DMSO(PE + DMSO) or DMSO and MG132 (PE + DMSO+ PI; proteasome inhibitor).The percentage of degradation is indicated below the gel (in red). The meanof the percentage of degradation of the 4 biological repetitions ± standarderror is indicated in the table on the right of the gel.
AbbreviationsN: Nitrogen; HN: High nitrogen; LN: Low nitrogen; DM: Dry matter;TSW: Thousand seed weight; LSU: Rubisco large subunit; GS: Glutaminesynthetase; GDH: Glutamate dehydrogenase; NUE: Nitrogen use efficiency;NUtE: Nitrogen utilization efficiency; NAE: Nitrogen assimilation efficiency;NRE: Nitrogen remobilization efficiency; NUpE: Nitrogen uptake efficiency;FtsH: Filamentation temperature-sensitive H.
Competing interestsThe authors have declared that they have no competing interests.
Authors’ contributionsAG, PE, JT, AB, FLC, LL, MO, MFN, NN, DC and JCA have made substantialcontributions to conception and design of experiment. AG, JT and JCAcarried out the N flux calculations. AG, CMD and FS performed the GS andGDH analysis. AG made other measurements and analyses, includingstatistical analyses, interpretation of data and drafting the article. All authorshave been involved in revising the article critically for important intellectualcontent. All authors read and approved the final manuscript.
AcknowledgementsThis work was funded by the French National Research Agency (ANR-07-GPLA-016 GENERGY: Improvement of the oil yield of the rapeseed crop in the contextof bio fuel production; ANR-11-BTBR-004 RAPSODYN - Investments for theFuture: Optimisation of the RAPeSeed Oil content anD Yield under low Nitrogeninput) and by a Ph.D. grant to Mrs Alexandra Girondé from the French Ministryof Research. The authors would like to thank Ms Marie-Paule Bataillé, Ms Anne-Françoise Ameline, Mr Raphaël Ségura and Mr Cédric Scher for their technicalhelp in plant harvest and N analyses. The authors also thank Sophie Rolland forher valuable help in NIRS analysis.
Author details1Université de Caen Basse-Normandie, F-14032 Caen, France. 2UCBN, UMRINRA–UCBN 950 Ecophysiologie Végétale, Agronomie & Nutritions N.C.S.,F-14032 Caen, France. 3INRA, UMR INRA–UCBN 950 Ecophysiologie Végétale,Agronomie & Nutritions N.C.S., F-14032 Caen, France. 4INRA, UMR 1349Institut de Génétique, Environnement et Protection des Plantes, INRA,Agrocampus Ouest, Université de Rennes 1, F-35653 Le Rheu, France. 5UMR1345 Institut de Recherche en Horticulture et Semences, SFR 4207 QUASAV,PRES L’UNAM, Université d’Angers, F-49045 Angers, France. 6UMR 1345Institut de Recherche en Horticulture et Semences, AgroCampus-Ouest,F-49045 Angers, France. 7Département Adaptation des Plantes àl’Environnement, UMR 1318, INRA, Institut Jean-Pierre Bourgin, RD10, 78026Versailles, Cedex, France.
Received: 15 December 2014 Accepted: 23 January 2015
References1. Carré P, Pouzet A. Rapeseed market, worldwide and in Europe. Oilseeds Fats
Crops Lipids. 2014;21(1):1–12.
2. Mosier AR. Environmental challenges associated with needed increases inglobal nitrogen fixation. Nutr Cycl Agroecosyst. 2002;63:101–16.
3. Laîné P, Ourry A, Macduff J, Boucaud J, Salette J. Kinetic parameters ofnitrate uptake by different catch crop species: effects of low temperaturesor previous nitrate starvation. Physiol Plant. 1993;88(1):85–92.
4. Schjoerring JK, Bock JGH, Gammelvind L, Jensen CR, Mogensen VO.Nitrogen incorporation and remobilization in different shoot componentsof field-grown winter oilseed rape (Brassica napus L.) as affected by rate ofnitrogen application and irrigation. Plant and Soil. 1995;177:255–64.
5. Dejoux J-F, Recous S, Meynard J-M, Trinsoutrot I, Leterme P. The fate ofnitrogen from winter-frozen rapeseed leaves: mineralization, fluxes to theenvironment and uptake by rapeseed crop in spring. Plant and Soil.2000;218:257–72.
6. Good AG, Shrawat AK, Muench DG. Can less yield more? Is reducingnutrient input into the environment compatible with maintaining cropproduction? Trends Plant Sci. 2004;9(12):597–605.
7. Schulte auf’m Erley G, Wijaya K-A, Ulas A, Becker H, Wiesler F, Horst WJ. Leafsenescence and N uptake parameters as selection traits for nitrogen efficiencyof oilseed rape cultivars. Physiol Plant. 2007;130(4):519–31.
8. Dreccer MF, Schapendonk AHCM, Slafer GA, Rabbinge R. Comparativeresponse of wheat and oilseed rape to nitrogen supply: absorption andutilisation efficiency of radiation and nitrogen during the reproductivestages determining yield. Plant and Soil.2000;220:189–205.
9. Wiesler F, Behrens T, Horst WL. Nitrogen efficiency of contrasting rapeideotypes. In: Horst WJ, editor. Plant nutrition - Food security and sustainabilityof agro-ecosystems. Netherlands: Kluwer Academic Publishers; 2001. p. 60–1.
10. Berry PM, Spink J, Foulkes MJ, White PJ. The physiological basis of genotypicdifferences in nitrogen use efficiency in oilseed rape (Brassica napus L.).Field Crop Res. 2010;119(2–3):365–73.
11. Schulte auf’m Erley G, Behrens T, Ulas A, Wiesler F, Horst WJ. Agronomictraits contributing to nitrogen efficiency of winter oilseed rape cultivars.Field Crop Res. 2011;124:114–23.
12. Ulas A, Behrens T, Wiesler F, Horst WJ, Schulte auf’m Erley G. Doesgenotypic variation in nitrogen remobilisation efficiency contribute tonitrogen efficiency of winter oilseed-rape cultivars (Brassica napus L.). PlantSoil. 2013;371:463–71.
13. Kessel B, Schierholt A, Becker HC. Nitrogen use efficiency in a geneticallydiverse set of winter oilseed rape (Brassica apus L.). Crop Sci.2012;52:2546–54.
14. Good AG, Beatty PH. Biotechnological approaches to improving nitrogenuse efficiency in plants: alanine aminotransferase as a case study. In:Hawkesford MJ, Barraclough P, editors. The molecular and physiologicalbasis of nutrient use efficiency in crops. Oxford, UK: John Wiley & Sons, Inc;2011. p. 5–19.
15. Yau SK, Thurling N. Variation in nitrogen response among spring rape(Brassica napus) cultivars and its relationship to nitrogen uptake andutilization. Field Crop Res. 1987;16:139–55.
16. Noquet C, Avice J-C, Rossato L, Beauclair P, Henry M-P, Ourry A. Effects ofaltered source-sink relationships on N allocation and vegetative storage proteinaccumulation in Brassica napus L. Plant Sci. 2004;166:1007–18.
17. Malagoli P, Laine P, Rossato L, Ourry A. Dynamics of nitrogen uptake andmobilization in field-grown winter oilseed rape (Brassica napus) from stemextension to harvest: I. Global N flows between vegetative and reproductivetissues in relation to leaf fall and their residual N. Ann Bot. 2005;95(5):853–61.
18. Malagoli P, Laine P, Rossato L, Ourry A. Dynamics of nitrogen uptake andmobilization in field-grown winter oilseed rape (Brassica napus) from stemextension to harvest. II. An 15 N-labelling-based simulation model of Npartitioning between vegetative and reproductive tissues. Ann Bot.2005;95(7):1187–98.
19. Merrien A, Palleau JP, Maisonneuve C. Mineral element requirements ofrape cultivated in France. In: Physiologie et élaboration du rendement ducolza. Paris: Cetiom; 1988. p. 34–46.
20. Rossato L, Laîné P, Ourry A. Nitrogen storage and remobilization in Brassicanapus L. during the growth cycle: nitrogen fluxes within the plant andchanges in soluble protein patterns. J Exp Bot. 2001;52(361):1655–63.
21. Gombert J, Le Dily F, Lothier J, Etienne P, Rossato L, Allirand J-M, et al. Effectof nitrogen fertilization on nitrogen dynamics in oilseed rape using 15 N-labelingfield experiment. J Plant Nutr Soil Sci. 2010;173(6):875–84.
22. Balint T, Rengel Z. Nitrogen efficiency of canola genotypes varies betweenvegetative stage and grain maturity. Euphytica. 2008;164:421–32.

Girondé et al. BMC Plant Biology (2015) 15:59 Page 20 of 21
23. Balint T, Rengel Z. Nitrogen and sulfur uptake and remobilisation in canolagenotypes with varied N- and S-use efficiency differ at vegetative andmaturity stages. Crop Pasture Sci. 2011;62:299–312.
24. Roberts IN, Caputo C, Criado MV, Funk C. Senescence-associated proteasesin plants. Physiol Plant. 2012;145:130–9.
25. Jain V, Kaiser W, Huber SC. Cytokinin inhibits the proteasome-mediateddegradation of carbonylated proteins in Arabidopsis leaves. Plant Cell Physiol.2008;49(5):843–52.
26. Ellis RJ. The most abundant protein in the world. Trends Biochem Sci.1979;4(11):241–4.
27. Evans JR. Photosynthesis and nitrogen relationships in leaves of C3 plants.Oecologia. 1989;78:9–19.
28. Makino A, Mae T, Ohira K. Relation between nitrogen and ribulose-1,5-bisphosphate carboxylase in rice leaves from emergence through senescence.Plant Cell Physiol. 1984;25(3):429–37.
29. Kumar PA, Parry MAJ, Mitchell RAC, Ahmad A, Abrol YP. Photosynthesis andnitrogen-use efficiency. In: Foyer CH, Noctor G, editors. Photosynthetic nitrogenassimilation and associated carbon and respiratory metabolism, vol. 12.Netherlands: Springer; 2002. p. 23–34.
30. Avice J-C, Etienne P. Leaf senescence and nitrogen remobilization efficiencyin oilseed rape (Brassica napus L.). J Exp Bot. 2014;65(14):3813–24.
31. Desclos M, Etienne P, Coquet L, Jouenne T, Bonnefoy J, Segura R, et al. Acombined 15 N tracing/proteomics study in Brassica napus reveals thechronology of proteomics events associated with N remobilisation duringleaf senescence induced by nitrate limitation or starvation. Proteomics.2009;9:3580–608.
32. Gaufichon L, Masclaux-Daubresse C, Tcherkez G, Reisdorf-Cren M, SakakibaraY, Hase T, et al. Arabidopsis thaliana ASN2 encoding asparagine synthetase isinvolved in the control of nitrogen assimilation and export during vegetativegrowth. Plant Cell Environ. 2013;36(2):328–42.
33. Lohaus G, Moellers C. Phloem transport of amino acids in two Brassicanapus L. genotypes and one B.carinata genotype in relation to their seedprotein content. Planta. 2000;211:833–40.
34. Kichey T, Hirel B, Heumez E, Dubois F, Le Gouis J. In winter wheat (Triticumaestivum L.), post-anthesis nitrogen uptake and remobilisation to the graincorrelates with agronomic traits and nitrogen physiological markers. FieldCrop Res. 2007;102:22–32.
35. Martin A, Lee J, Kichey T, Gerentes D, Zivy M, Tatout C, et al. Two cytosolicglutamine synthetase isoforms of maize are specifically involved in thecontrol of grain production. Plant Cell. 2006;18:3252–74.
36. Ochs G, Schock G, Trischler M, Kosemund K, Wild A. Complexity andexpression of the glutamine synthetase multigene family in theamphidiploid crop Brassica napus. Plant Mol Biol. 1999;39:395–405.
37. Orsel M, Moison M, Clouet V, Thomas J, Leprince F, Canoy A-S, et al. Sixteencytosolic glutamine synthetase genes identified in the Brassica napus L.genome are differentially regulated depending on nitrogen regimes andleaf senescence. J Exp Bot. 2014;65(14):3927–47.
38. Turano FJ, Thakkar SS, Fang T, Weisemann JM. Characterization andexpression of NAD(H)-dependent glutamate deshydrogenase genes inArabidopsis. Plant Physiol. 1997;113:1329–41.
39. Masclaux-Daubresse C, Reisdorf-Cren M, Pageau K, Lelandais M, GrandjeanO, Kronenberger J, et al. Glutamine synthetase-glutamate synthase pathwayand glutamate dehydrogenase play distinct roles in the sink-source nitrogencycle in tobacco. Plant Physiol. 2006;140:444–56.
40. Purnell MP, Botella JR. Tobacco isoenzyme 1 of NAD(H)-dependentglutamate dehydrogenase catabolizes glutamate in vivo. Plant Physiol.2007;143:530–9.
41. Skopelitis DS, Paranychianakis NV, Paschalidis KA, Pliakonis ED, Delis ID,Yakoumakis DI, et al. Abiotic stress generates ROS that signal expression ofanionic glutamate dehydrogenases to form glutamate for proline synthesisin tobacco and grapevine. Plant Cell. 2006;18:2767–81.
42. Lohaus G, Schwerdtfeger M. Comparison of sugars, iridoid glycosides andamino acids in nectar and phloem sap of Maurandya barclayana,Lophospermum erubescens, and Brassica napus. PLoS One 2014, 9(1):DOI:10.1371/journal.pone.0087689.
43. Watanabe M, Nakayama H, Watanabe Y, Shimada N. Mechanical slicing-induced alteration of GDH isoenzyme patterns in Brassica napus leafprotoplasts. J Plant Physiol. 1994;143:87–91.
44. Tilsner J, Kassner N, Struck C, Lohaus G. Amino acid contents and transportin oilseed rape (Brassica napus L.) under different nitrogen conditions.Planta. 2005;221(3):328–38.
45. Girondé A, Poret M, Etienne P, Trouverie J, Bouchereau A, Le Cahérec F,et al. A profiling approach of the natural variability of foliar N remobilizationat the rosette stage gives clues to understand the limiting processesinvolved in the low N use efficiency of winter oilseed rape. J Exp Bot. inpress:JEXBOT/2014/137661.
46. Hocking PJ, Randall PJ, DeMarco D. The response of dryland canola tonitrogen fetilizer: partitioning and mobilization of dry matter and nitrogen,and nitrogen effects on yield components. Field Crop Res. 1997;54:201–20.
47. Ahmad A, Abdin MZ. Interactive effect of sulphur and nitrogen on the oiland protein contents and on the fatty acid profiles of oil in the seeds ofrapeseed (Brassica campestris L.) and mustard (Brassica juncea L. Czern. andCoss). Journal of Agronomy and Crop Science. 2000;185:49–54.
48. Zhao J, Becker HC, Zhang D, Zhang Y, Ecke W. Conditional QTL mapping ofoil content in rapeseed with respect to protein content and traits related toplant development and grain yield. Theor Appl Genet. 2006;113:33–8.
49. Asare E, Scarisbrick DH. Rate of nitrogen and sulphur fertilizers on yield,yield components and seed quality of oilseed rape (Brassica napus L.). FieldCrop Res. 1995;44:41–6.
50. Taylor AJ, Smith CJ, Wilson IB. Effect of irrigation and nitrogen fertilizer onyield, oil content, nitrogen accumulation and water use of canola (Brassicanapus L.). Fertilizer Res. 1991;29:249–60.
51. Long SP, Zhu X-G, Naidu SL, Ort DR. Can improvement in photosynthesis in-crease crop yields? Plant. Cell Environ. 2006;29:315–30.
52. Wiesler F, Behrens T, Horst WJ. The role of nitrogen-efficiency cultivars insustainable agriculture. Sci World. 2001;1(S2):61–9.
53. Wardlaw IF. The control and pattern of movement of carbohydrates inplants. Bot Rev. 1968;34(1):79–105.
54. Makino A, Sakashita H, Hidema J, Mae T, Ojima K, Osmond B. Distinctiveresponses of ribulose-1,5-bisphosphate carboxylase and carbonic anhydrasein wheat leaves to nitrogen nutrition and their possible relationships toC02-transfer resistance. Plant Physiol. 1992;100:1737–43.
55. Wagner R, Aigner H, Funk C. FtsH proteases located in the plant chloroplast.Physiol Plant. 2012;145:203–14.
56. Buchanan-Wollaston V, Ainsworth C. Leaf senescence in Brassica napus:cloning of senescence related genes by subtractive hybridisation. Plant MolBiol. 1997;33:821–34.
57. Sullivan JA, Shirasu K, Deng XW. The diverse roles of ubiquitine and the 26Sproteasome in the life of plants. Nat Rev Genet. 2003;4:948–58.
58. Hörtensteiner S, Feller U. Nitrogen metabolism and remobilization duringsenescence. J Exp Bot. 2002;53(370):927–37.
59. Martínez DE, Costa ML, Guiamet JJ. Senescence-associated degradation of chloroplastproteins inside and outside the organelle. Plant Biol. 2008;10 Suppl 1:15–22.
60. Feller U, Anders I, Mae T. Rubiscolytics: fate of Rubisco after its enzymaticfunction in a cell is terminated. J Exp Bot. 2008;59(7):1615–24.
61. Beers EP, Moreno TN, Callis J. Subcellular localization of ubiquitin andubiquitinated proteins in Arabidopsis thaliana. J Biol Chem. 1992;267(22):15432–9.
62. Ling Q, Huang W, Baldwin A, Jarvis P. Chloroplast biogenesis is regulated bydirect action of the ubiquitin-proteasome system. Science. 2012;338:655–9.
63. Vierstra RD. Proteolysis in plants: mechanisms and functions. Plant Mol Biol.1996;32:275–302.
64. Masclaux-Daubresse C, Daniel-Vedele F, Dechorgnat J, Chardon F, GaufichonL, Suzuki A. Nitrogen uptake, assimilation and remobilization in plants:challenges for sustainable and productive agriculture. Ann Bot. 2010;105(7):1141–57.
65. Damodaran M, Ananta-Narayanan P. Enzymic proteolysis. Liberation ofammonia from proteins. Biochem J. 1938;32(10):1877–89.
66. Dubousset L, Etienne P, Avice J-C. Is the remobilization of S and N reservesfor seed filling of winter oilseed rape modulated by sulphate restrictionsoccuring at different growth stages? J Exp Bot. 2010;61(15):4313–24.
67. Salon C, Bataillé M-P, Gallardo K, Jeudy C, Santoni A-L, Trouverie J, et al. 34Sand 15 N labelling to model S and N flux in plants and determine thedifferent components of N and S use efficiency. Methods Mol Biol.2014;1090:335–46.
68. Bradford MM. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding. Anal Biochem. 1976;72(1–2):248–54.
69. Masclaux C, Valadier M-H, Brugière N, Morot-Gaudry J-F, Hirel B.Characterization of the sink/source transition in tobacco (Nicotiana tabacumL.) shoots in relation to nitrogen management and leaf senescence. Planta.2000;21:510–8.

Girondé et al. BMC Plant Biology (2015) 15:59 Page 21 of 21
70. O’Neal D, Joy KW. Glutamine synthetase of Pea leaves. I. Purification,stabilization, and pH optima. Arch Biochem Biophys. 1973;159:113–22.
71. Laemmli UK. Cleavage of structural proteins during the heat bacteriophageT4. Nature. 1970;227:680–5.
72. Kazmin D, Edwards RA, Turner RJ, Larson E, Starkey J. Visualization ofproteins in acrylamide gels using ultraviolet illumination. Anal Biochem.2002;301:91–6.
Submit your next manuscript to BioMed Centraland take full advantage of:
• Convenient online submission
• Thorough peer review
• No space constraints or color figure charges
• Immediate publication on acceptance
• Inclusion in PubMed, CAS, Scopus and Google Scholar
• Research which is freely available for redistribution
Submit your manuscript at www.biomedcentral.com/submit





























