Embed Size (px)
Citation preview

The Crystalline Style in Gastropods.
By
N. A. Mackintosh, A.R.C.S., B.Sc,
Assistant Naturalist to the ' Discovery' Expedition.
With Plates 20 and 21 and 3 Text-figures.
CONTENTS.PAGE
1. INTRODUCTION . . . . . . . . . 317
2. T H E CRYSTALLINE STYLE I N C R E P I D U L A F O R N I C A T A . 319
(1) Material 319(2) The Style-bearing Region 320
(a) Gross Anatomy . . . . . . . 320(6) Histology 323
(3) The Crystalline Style 327(a) Form 327(b) C o m p o s i t i o n 3 2 8( c ) F u n c t i o n 3 2 9
3 . G E N E R A L C O N S I D E R A T I O N S . . . . . . . 3 3 2
4 . S U M M A R Y 3 3 9
5 . L I T E R A T U R E 3 4 0
6 . E X P L A N A T I O N o r P L A T E S . . . . . . . 3 4 2
1. INTRODUCTION.
IT is well known that the presence of a crystalline style isthe normal condition among Lamellibranchs, but the occasionalappearance of this organ in Gastropoda is often overlooked.A summary of the Gastropod genera in which it is knownto occur is given by Kobson (27). It is not claimed that thislist is exhaustive, indeed it may be found that a style occursin many more forms than are so far recorded as possessing it,but we see from the list that it is present most commonly in theTaenioglossa, and that one or two cases are recorded amongst theDocoglossa and the Ehipidoglossa. Now although the crystallinestyle in Lamellibranchs has been the subject of numerous and

318 N. A. MACKINTOSH
exhaustive investigations, no really complete description ofthis organ has yet been given in the case of any Gastropod.The literature dealing with the subject has been confinedprincipally to the general shape of the style-bearing region,the relation of the style-sac to the intestine, &c, or has con-sisted merely of brief references to the occurrence of a style.For instance, there are papers by Moore (18), Woodward (30),and others, and that of Eobson mentioned above, in whichthe occurrence of a style in Gastropods is discussed, but whichdo not go into details, and again Bregenzer (3), in describing thegeneral anatomy of B i t h y n e l l a d u n k e r i , devotes somethirty lines to a description of the style and style-sac. Thelast is, in fact, as complete an account of the style in anyindividual species as I have been able to find.
The object of the present paper is to give a detailed accountof the nature and function of the Gastropod crystalline styleand the structures with which it is associated, in order to providea basis upon which a comparison can be made with the con-ditions found in Lamellibranchs, and to consider what is theexplanation of its occurrence in a few isolated genera ofGastropods. For this purpose the greater part of the workhas been devoted to a thorough investigation of the style inCrepidula, and the rest to a discussion of the style in Gastropodsfrom a general point of view. I have selected Crepidula asthe most convenient type, since the style is large in comparisonwith that of most other Gastropods in which it occurs, andbecause living specimens are easily obtained in large numbers.
To consider in detail every aspect of the subject whichhas been studied in regard to the style in Lamellibranchswould be rather beyond the scope of the present paper, but inorder that it may be as complete as possible the matter istreated from the point of view of the structure and histologyof the style-bearing region, and the form and general physiologyof the style itself. It should be mentioned here that thehistology of the style-sac is considered in rather more detailthan might appear to be strictly necessary for purposes ofcomparison with other forms, but investigation of the minute

CRYSTALLINE STYLE IN GASTROPODS 319
structure of the style-bearing organs has revealed such interest-ing features as to deserve some special attention.
The Slipper Limpet, C r e p i d u l a f o r n i c a t a , has beenthe subject of various investigations, of which the most impor-tant from our point of view are two papers by Orton (21, 22).Prom these it may be seen that Crepidula was introduced intoEngland from America about 1880, and that it has spreadrapidly over certain grounds round the south and east coasts,and is especially abundant on the oyster beds. Its food is thesame as that of the oyster, and its method of feeding is alsosimilar in many ways. The contents of the gut were examinedby Orton, who found diatom shells, vegetable debris, radio-larian, foraminiferan and peridinean tests, sponge spicules,and sand grains in both animals. A thorough account of themode of feeding in Crepidula is given by Orton.
The occurrence of a style in this Gastropod was also pointedout by Orton (23) some years later. The presence of thisorgan is interesting-in connexion with the similarity in thefood and feeding in this species to that of the oyster. This,however, is a matter which will be considered later.
The investigations which are now to be described werecarried out in the Zoological Department of the ImperialCollege of Science.
2. THE CRYSTALLINE STYLE IN C R E P I D U L A F O R N I C A T A .
(1) M a t e r i a l .Living specimens of Crepidula were obtained from Whit-
stable. It is quite easy to keep Crepidula alive in tanks fora considerable period, but the style disappears if the -wateris allowed to become foul. Owing to a very limited supplyof fresh sea-water and to the fact that the animals were notliving under natural conditions, it was not possible to makeany extensive experimental studies, but there was no greatdifficulty in testing the biochemical nature of the style andin-studying its relation to the food-stream in freshly deliveredspecimens.

820 N. A. MACKINTOSH
(2) The S t y l e - b e a r i n g E e g i o n .
(a) Gross Anatomy.In describing the general structure of the style-bearing
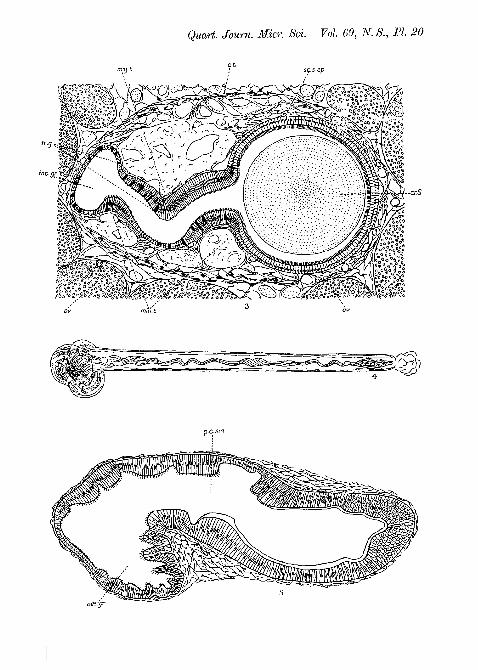
region it will perhaps be well first to explain the course takenby the alimentary canal (Text-fig. 1). The oesophagus is along narrow tube which runs in a fairly straight line from thehead to the stomach, into which it opens about half-wayalong the ventral wall. The stomach lies on the right-handside of the visceral hump, which is the form of a flat lobeenclosed in the upper chamber of the shell and joined to thefoot only at its anterior end. The intestine passes out fromthe left anterior corner of the stomach and runs diagonallyacross the visceral hump. It then loops back and runs intothe mantle where it opens into the rectum.
The style-sac in Crepidula is of the type which is incom-pletely separated from the intestine. The first straight sectionof the latter is divided into two channels by dorsal and ventraltyphlosoles. As viewed from the stomach, the left-handchannel is considerably smaller than the right. The formeris the intestine proper and the latter is the style-sac (fig. 3,PI. 20). The typhlosoles do not quite meet one another,but leave a slit through which the style-sac and intestine arein communication. It will be convenient to refer to thelatter as the intestinal groove, as the style-sac is morpho-logically part of the intestine itself.
The dorsal typhlosole is rather deeper than the ventral,and, using the terms employed by Nelson (20) in the case ofLamellibranchs, we may refer to it as the major typhlosole,and to the ventral as the minor typhlosole. The latter israther concave in the middle line, consisting of a kind of doubleridge, and can in fact almost be regarded as two typhlosoles.The major typhlosole is convex and fits into the concavegroove of the minor typhlosole so that the slit connectingthe style-sac and intestinal groove is V-shaped in cross-section.
The style-sac and typhlosoles gradually diminish in sizeas they become more remote from the stomach. They persist
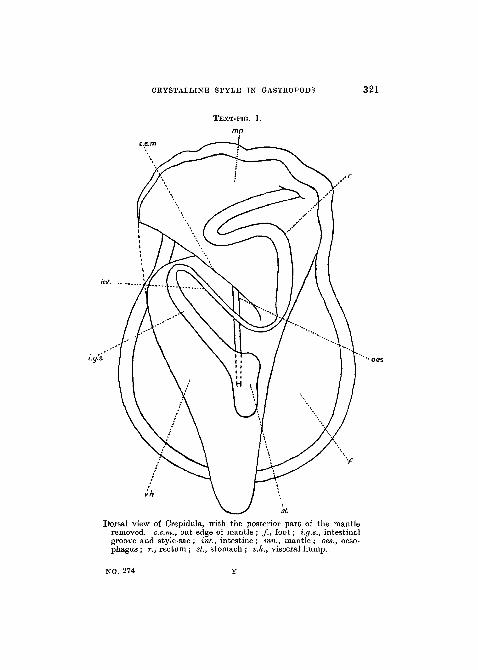
CRYSTALLINE STYLE IN GASTROPODS 321
TEXT-FIG. 1.
i.g.s.
y.h
st
Dorsal view of Crepidula, with the posterior part of the mantleremoved, c.e.m., out edge of mantle ; / . , foot; i.g.s., intestinalgroove and style-sac ; int., intestine ; mn., mantle ; oes., oeso-phagus; r., rectum; St., stomach ; v.h., visceral hump.
NO. 274

322 . N. A. MACKINTOSH
along the whole of the first straight section of the intestineand end without any indication of a divergent caecum, suchas is found in some Lamellibranchs possessing an incompletelyseparated style-sac. That is to say that the style-sac inCrepidula is in communication with the intestinal groovealong the whole of its length.
At the proximal end of the stj'le-sac the minor typhlosolestops short, forming a kind of lip, but the major typhlosoleis to some extent continued as a ridge across the roof of thestomach.
The stomach itself (fig. 1, PI. 20) is roughly pear-shaped, andthe anterior part, from which the intestine opens out, isconsiderably deeper than the rest, forming a cavity in whichthe head of the style works. The posterior part is shallower,and it is into this that the oesophagus opens on the lower left-hand side. Here there is a peculiar anatomical feature. Theoesophagus is continued as a deep groove which curves rightround the posterior half of the stomach, and serves, as willbe explained later, to convey the food-stream round to thehead of the style in the main cavity. This groove, which isshown in section in fig. 5, PI. 20, is to all intents and purposesa part of the oesophagus itself, for the epithelium of the latteris thrown up into sharp longitudinal folds which continueunchanged in the groove. Various parts of the epithelium ofthe stomach are also thrown up into parallel folds.
A large part of the anterior cavity of the stomach is coveredAvith a cutieular lining. Opposite the opening of the style-sacthis lining is greatly thickened and forms a large, blade-likegastric shield upon which the head of the style impinges(fig. 2, PI. 20). No definite shape can be assigned to the gastricshield as it merges imperceptibly into the cutieular lining.
The hepatopancreas lies round about the stomach, mostlyon the left-hand side. Its main ducts open into the anteriorcavity near the lip of the style-sac. It is here that all thegastric processes appear to take place, in fact the posteriorpart which is traversed by the oesophageal groove seems to bequite unnecessarily large.

CRYSTALLINE STYLE IN GASTROPODS 323
The actual working of the style will be considered laterwhen its function is discussed.
(b) Histology.In the investigation of the histological features of the style-
bearing region the following technique was used. For generalroutine work on the anatomy and histology, involving sectionscut right across the style-bearing region, the material was fixedin Bouin's solution and stained principally in Ehrlich's haematoxylin and orange G or eosin, or in iron haematoxylin. Othermethods which gave useful results were the Champy-Kulltechnique, Memming's orange combination, and Mallory'striple stain. Mucicarmine was used for the detection of mucusglands. 1'or the study of the intracellular fibre apparatusiron haematoxylin was the most useful method. Macerationin boracic acid gave unsatisfactory results.
The style-sac is lined with an epithelium consisting of largecolumnar cells, covered at their distal ends with a dense massof long cilia. The cilia are all of exactly equal length, and forma perfectly even surface upon which the style rests.
Where the epithelium of the style-sac joins that of thetyphlosoles there is a distinct line of demarcation, especiallyin respect of the cilia which abruptly become shorter andnarrower. The epithelium of at least the main ridge of themajor typhlosole consists of cells whose length is twenty orthirty times as great as the diameter. On the intestinal aspectof the major typhlosole the epithelium becomes graduallyshallower and the cilia more sparse, until it passes into theepithelium of the intestinal groove which consists principallyof small, almost cubical, feebly ciliated cells.
The minor typhlosole, as already explained, consists oftwo ridges. The cells on the ridges are relatively long witha fair development of cilia, while the cells of the concave partbetween them are short, resembling those of the intestine, withpoorly developed cilia.
Beneath the epithelium the substance of the typhlosoles iscomposed of a pale, almost structureless tissue, rather similar
Y2

324 N. A. MACKINTOSH
to that which occurs in Lamellibranchs, with few nuclei andinterspersed with lacunae. The style-bearing region is enclosedby a rather ill-defined layer of connective tissue, forminga, roughly circular sheath separating the whole from thesurrounding tissues.
Among the cells of the epithelium of the style-bearing regionare numbers of mucus glands. These occur in the intestinalgroove, on the inner slope of the right ridge of the minortyphlosole, and on the intestinal aspect of the major typhlosole.It is curious that the mucus glands are seen in many sectionsat a point near the junction of the style-sac and majortyphlosole, but definitely within the style-sac epithelium.It appears that there is a row stretching longitudinally downthe style-sac in this position. Mucus glands do not occurin any other part of the style-sac epithelium.
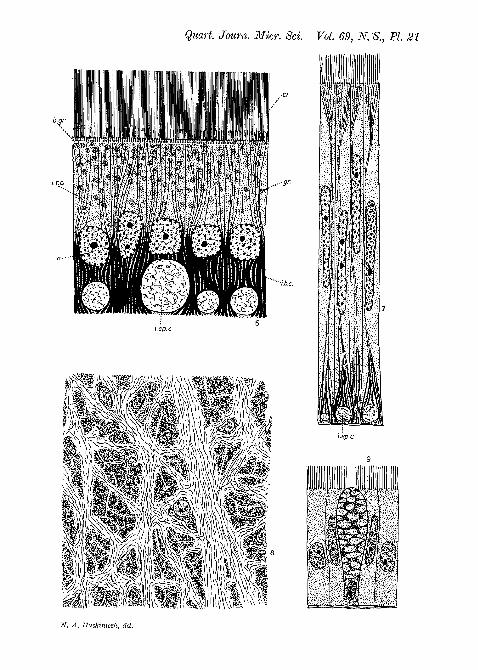
The mucus glands (fig. 9, PI. 21) do not call for any specialcomment. Each consists of a flask-like cell interpolated betweenthe epithelial cells. They are usually considerably distendedwith mucus, with the nucleus lying near the base. Theseglands show no essential difference from those described byGutheil (11) and others in the alimentary tract of Lamelli-branchs.
It has already been mentioned that the histology of thestyle-bearing region presents certain interesting features whichdeserve special attention. These are concerned principallywith the minute structure of the epithelial cells of the style-sac.The style-sac epithelium (fig. 6, PI. 21) consists of columnarcells whose size, shape, and arrangement are extremely uniform.The width of the cells is rather less than a quarter the length.The nuclei lie just within the lower half of the cells and maycontain one or two nucleoli. The cytoplasm above themcontains a variable number of large green granules which,though sometimes rather scanty, are often so densely crowdedas to form an opaque mass completely obscuring the upperhalf of each cell. The lower half of each cell is filled witha bundle of thick, rod-like bodies, which, in most cells, arepushed apart by a cylindrical canal which apparently pierces

CRYSTALLINE STYLE IN GASTROPODS 325
the base of the cells. These rod-like bodies and canals are ofspecial interest and are presently to be described more fully.It is necessary, however, first to describe the cilia at the endsof the cells. These cilia are very strikingly developed and forma dense, perfectly even carpet all over the style-sac. Theirlength is always precisely the same and equal to rather less thanhalf that of the cells. Their proximal ends appear to bethickened, for in stained sections there is a darker zone againstthe ends of the cells. Next to this zone there is a prominentdouble row of basal granules.
The cilia are continued into the base of the cell, forming aninternal fibrillar apparatus. It appears that by no means allthe external cilia are continued into the cell. The number ofactually visible internal fibres is at least relatively small.Gurwitsch found in cross-sections of certain ciliated epithelialcells that there were only 30 to 40 rootlet fibrils in each cellas against 200 to 300 basal granules. It is possible of coursethat many of these fibres tend to fuse together. In the ciliatedepithelial cells of many molluscs the rootlets of the ciliagradually converge and join in one fibre which may be tracedfar down into the cell. This is described by Englemann (8).In the case of the style-sac of Crepidula, however, though therootlets show some sign of convergence, they penetrate to thebase of the cells without fusing together. But their moststriking feature is that below the nucleus they becomeimmensely thickened, forming the heavy, rod-like bodiesmentioned above. These are probably composed of a hardbrittle substance, for they are inclined to crumble and causeconsiderable difficulties in section cutting. In successfulpreparations, however, which have been stained in iron haema-toxylin, they appear as a bundle of black rods. They alsostain strongly in Altmann's acid fuchsin and to some extentin safranin.
The most interesting feature, however, of the style-sacepithelium is a peculiar system of ' intra-epithelial' canalswhich are shown in figs. 6 and 8, PI. 21. In transverse sectionsone sees just within the basement membrane what at first

326 N. A. MACKINTOSH
appear to be large vacuoles of variable size, separating therod-like bases of the internal rootlets of the cilia. Frontalsections show that these are the elements of a complex systemof canals which ramify throughout the base of the epithelium,forming an anastomosing network (fig. 8, PI. 21). It is noteasy to determine whether the canals pass through or betweenthe cells, for the cell walls are obscured by the internal basesof the cilia; but I am of the opinion that they actually piercethe bases of the cells or at least divide the lower extremitiesinto two ' legs ', for they correspond roughly in number to thecells and lie directly below each nucleus. The larger canalsmostly run longitudinally down the style-sac, but they areconnected with one another by numerous smaller canals whichlie largely in oblique directions. They are filled with a palestringy substance, the identification of which has presentedsome difficulty. Its staining properties are almost identicalwith those of the crystalline style, but it cannot be the samematerial as the style substance is soluble in weak alkalis whilethis substance is not. Furthermore, the contents of the canals,when stained sufficiently strongly, is seen to consist of a wavybundle of fibres whose appearance is strongly suggestive ofthe white fibres of connective tissue. This suggestion is tosome extent supported by the staining reactions. They main-tain a whitish appearance after most stains, but show a certainaffinity for acid fuchsin and water-blue (' wasserblau '). Eventhese stains, however, have a rather feeble effect, and thecolouring is not so strong as is the case with the collagenousfibres of ordinary connective tissue. It seems possible, however,that we have here an apparatus whose function it is to lendextra strength to the epithelium. One sees that such a devicemight be of value, for the epithelium has to support thestrain of revolving and pushing forward the heavy style.
If the canals are actually of a collagenous nature, it mustbe supposed that its condition is not quite similar to that ofthe collagenous fibres of ordinary connective tissue, for itoften occurs in rather larger bundles than one would expectto find in the latter and its powers of staining are feebler.

CliYSTALLINE STYLE IN GASTROPODS 327
I have been unable to find records of any structure whichcould be compared with this except in one case. It is verycurious that in the pharyngeal epithelium of LumbricusPolowzow (24) and Mayer (16) have described an anastomosingnetwork which is very similar to that which occurs in thestyle-sac of Crepidula. This is interesting in view of the factthat it may be supposed that the pharyngeal epithelium of theearthworm is also subjected to a certain amount of strain.Polowzow held that the network consisted of nerve-fibres,but Mayer considered it to be an elastic supporting tissue.
The architecture of the cells of the typhlosoles (fig. 7, PL 21)is in general similar to those of the style-sac, but the structureswhich have been considered above are less pronounced andthe cytoplasm does not contain the green granules which arecharacteristic of the style-sac epithelium. The canals at thebases of the cells are more scanty and mostly smaller, thoughvery large ones occur here and there. They become still lessnoticeable in the direction of the intestinal groove.
(3) The C r y s t a l l i n e S t y l e .
(a) Form.
The crystalline style (fig. 4, PI. 20) in its normal form infully grown individuals is a straight, flexible, transparent rodabout 20 mm. in length and of an average diameter of slightlymore than 1 mm. As will be seen later, it resembles in everyway the Lamellibranch style. The colour varies from a paleyellow to a rather faint reddish brown. It tapers slightlyfrom before backwards in accordance with the shape of thestyle-sac which it completely fills, unless it is in the process ofdissolution. When it is viewed in optical section a stringycore can be seen which is twisted in a rather irregular manner,and which expands into a kind of knot in places. The arrange-ment of this core is very variable, but it is often most prominentin the hinder part of the style, becoming more diffuse near thehead. At other times it can be clearly distinguished throughoutthe length of the style.

328 N. A. MACKINTOSH
The head of the style projects into the stomach, and, mixedwith the contents of the latter, fills the larger anterior cavityto which reference has already been made.
The inner structure of the style can often be studied largelyfrom transverse sections of the whole style-bearing region,but much better results are obtained if the style is removedand sectioned separately after fixation in Bouin or Carnoy.In transverse section it is seen to consist of co-axial layerssurrounding a granular core (fig. 3, PI. 20). The diameter ofthe core relative to that of the style is variable and dependson the condition of the style and the point at which the sectionis cut. The head of the style consists of a tangled mass of stylesubstance, food material, and mucus.
The style usually stains feebly or not at all. Saurefuchsingenerally gives the best results. The fact that mucicarmine istaken up to some extent suggests that mucin is a constituentof the style, if mucicarmine is really a specific stain.
(b) Composition.Some tests were made in order to gain some idea of the
biochemical nature of the style substance. Extensive investiga-tions of the composition of the style in Lamellibranchs havebeen made by A'arious workers, notably by Barrois (1), whoshowed that the style exists as a globulin Avith about 87 percent, of water and 1 per cent, of salts. These results Averelargely confirmed by Mitra (17).
Owing to limitation of time and material I haA'e been unableto make any quantitative investigations of the compositionof the style in Crepidula, but the following qualitative testsAvere made.
The style dissolves in Avater, giving a neutral solution whichis coagulated by boiling or by the addition of alcohol. A smallquantity of the style solution Avas acidified with dilute aceticacid. There was a slight precipitate Avhich Avas insoluble instrong acetic acid. This indicates the presence of mucin.A solution of styles was treated Avith an equal quantity ofsaturated ammonium sulphate solution and filtered. The

CRYSTALLINE STYLE IN GASTROPODS 329
precipitate was scraped from the paper, dissolved in a littlecold water, and boiled. A coagulum was obtained indicatingthe presence of globulin. The nitrate gave no heat coagulum.
From these tests it may be inferred that the style substanceexists principally in the form of a globulin, and that somemucin is also present. There is in fact no reason to doubtthat it is essentially similar in composition to that of Lamelli-branchs, which it so closely resembles in form, colour, andconsistency.
Globulins are generally insoluble in distilled water, but thesolubility of the style is probably due to the presence of salts,as was suggested by Mitra in the case of Anodonta.
The presence of an amylolytic enzyme in the Lamellibranchstyle was first demonstrated by Coupin (5) in the case ofCardium, and by Mitra, Yonge (31) and others in other species.I have tested for this enzyme in the case of Crepidula byincubating a solution of styles with an equal quantity of1 per cent, starch paste for two hours and subsequently testingwith Fehlings solution, and have found it to be present.
(c) Function.In considering the function of the style in Crepidula it will
be convenient to include under this heading its actual methodof working, together Avith the part it plays in the digestiveprocesses, and it will be best to begin with the animal's modeof feeding. This has been investigated by Orton (22), whosedescription may be summarized as follows. Crepidula feedsin much the same way as the oyster. Ingoing and outgoingcurrents pass through the mantle-cavity. Food particles arecollected on the gills and deposited in a ciliated groove, wherethey becomeembedded in mucus and worked into a cylindricalmass. This is passed up to the mouth at intervals where it isseized and drawn in by the radula. An alternative routewhich is usually taken by the larger particles leads to a pouchin front of the mouth, where the food is mixed with mucusand formed into a pellet.
The food material passes up the oesophagus as a thin cord

330 N. A. MACKINTOSH
of mucus in which the particles are embedded, and in thisform it arrives at the stomach and moves round the oesophagealgroove and thence forward to the anterior cavity of the stomachwhich is occupied by the head of the style with which it becomesentangled (Text-fig. 2).
It may be assumed from the existence of a spiral coresurrounded by co-axial layers, and by analogy with Lamelli-branchs, that, as the style is formed it is simultaneously
TEXT-FIG. 2.
Dorsal view of the stomach of Crepidula, to show the relations of thestyle to the stream of food particles embedded in mucus.Compare with fig. 1, PI. 20. cr.a., crystalline style; int.g.,intestinal groove; maj.t., major typhlosole; Min.t., minortyphlosole ; m.s., stream of mucus containing food particles ;o.oes., opening of the oesophagus.
rotated and pushed forward by the powerful cilia of the style-sac. In the stomach the string of food and mucus meets therevolving head of the style where the latter comes into contactwith the blade-like gastric shield (fig. 2, PI. 20), so that athorough admixture of all the stomach contents takes placebefore the food passes on to the intestine.
It is generally supposed that the gastric shield was developedopposite the style-sac as a necessary protection for the gastricepithelium. This is very probable, but in the case at least of

CRYSTALLINE STYLE IN GASTROPODS 331
Crepidula, the shield has a further function in causing a muchmore thorough mixture of the stomach contents than wouldtake place if it were absent or of a less prominent shape. Itis of a sharp, blade-like form, and as shown in fig. 2, PL 20,Avhere it appears in section, it projects well into the revolvingalimentary mass so as to resist the rotation in such a way asto cause a most thorough entanglement of the various sub-stances present.
Prom the evidence presented by Nelson and Edmondson (7)it appears that in Lamellibranchs the style substance issecreted by the epithelial cells of the typhlosoles. Nelsondescribes and figures in the cytoplasm of these cells dropletsof a refractive fluid which are also seen outside the epithelium,and which appear to become incorporated with the style.I have also seen drops of a pale fluid substance in the typhlosoleepithelium of Crepidula, but have not been able definitely totrace them to the style.
For reasons already given I have not been able to carryout systematic experiments on the permanence of the style.If Crepidula is removed from the water the style disappearsafter two or three days, but it is reformed after a few hourswhen the animal is put back into the water.
There is no reason for supposing that the style is in anyessential different from that of Lamellibranchs as regardsfunction. It has been shown that it is similar in form andcomposition, and that it also contains an amylolytic enzyme.It appears that, as in Lamellibranchs, its principal functionsare to mix together the contents of the stomach, to keep thewhole mass in rotation, and to supply a starch-splittingferment.
It was suggested by Orton (23) that it would be interestingto ascertain whether the style had any power of dissolvingmucus in view of the fact that the food is conveyed to thestomach in a heavy thread of this substance. My experiments,however, in this direction have met with negative results.

3 3 2 N. A. MACKINTOSH
3. GENERAL CONSIDERATIONS.
The most important fact revealed by the foregoing accountof the crystalline style and its accompanying structures inCrepidula is the extraordinary resemblance to the conditionsfound in Lamellibranchs. The style-sac, being of the typewhich is incompletely separated from the intestine, may becompared with that of Anodonta, Lamsilis, and others.
It may now be stated that the style and style-bearing regionresemble those of the corresponding type in Lamellibranchsin the following features :
(1) The style-sac and intestine are separated from oneanother by typhlosoles, one of which is distinctly larger thanthe other.
(2) The intestinal part is in the form of a relatively smallgroove.
(3) The style and typhlosoles gradually diminish distally.(4) There is a gastric shield of the ordinary type.(5) The style-sac epithelium is similar to that of Lamelli-
branchs in regard to the shape of the cells and the denseexternal cilia.
(6) There is a distinct demarcation between the epitheliumof the style-sac and that of the typhlosoles.
(7) The supporting substance of the typhlosoles consists ofa pale vascular tissue with few nuclei.
(8) The whole style-bearing region is bound round by arather indistinct sheath of connective tissue.
(9) Mucus glands are numerous in the intestine and on partsof the typhlosoles.
(10) The upper parts of the cells lining the style-sac are filledwith dark granules.
(11) The style itself is similar in shape, colour, and con-sistency to that of Lamellibranchs.
(12) It is formed of co-axial layers surrounding a spiral core.(13) The style substance consists principally of a globulin
and contains an amylolytic enzyme.(14) The whole function and method of working must be
presumed to be similar to those of the Lamellibranch style.

CRYSTALLINE STYLE IN GASTROPODS 333
It should be explained that for this comparison Anodontais taken as the type Larnellibranch, at least so far as themorphology of the style-bearing region is concerned. But theabove points will be found to apply in general to most Lanielli-branchs possessing the incompletely separated style-sac. Myinformation regarding them is derived from my own sections ofAnodonta, and from various authors to whom reference hasbeen made, in particular from Nelson.
There are practically no differences between the Lamelli-branch style and that of Crepidula. It should be mentionedthat in Anodonta and some other bivalves the epithelium of thestyle-sac is transversely ridged so that the style rests on aseries of ' bearings ' instead of on a smooth surface as inCrepidula.
At all events the structure of the style and style-bearingregion in Crepidula is well within the limits of the variationsfound in this organ in the Larnellibranchs which have anincompletely- separated style-sac. When an organ such asthe crystalline style appears in two separate classes of thesame phylum one would expect to find certain resemblances,but such a close similarity as actually exists is astonishing.The significance of this must, however, be discussed later,for it is now necessary to consider the nature of the style insome other Gastropods.
Working on the relations of the style-sac and intestine inLamellibranchs, Matthias (15) distinguished three main types :(1) That in which the style lies in the intestine, or in which theintestine and style-sac are in wide and open communicationwith one another (Area, Ostrea, Leda, &c). (2) That in whichthe style-sac and intestine are definitely separated by typhlo-soles but communicate by a slit (Modiolaria, Jouanetia, &c).(3) That in which the style-sac is an independent sac completelyseparated from the intestine (Martesia, Teredo, Dreissensia,&c). Eobson (27) has pointed out the interesting fact that thesame three types are to be distinguished among Gastropods.He also gives a list of cases in which a style is known to occuramong Gastropoda. It will be convenient to draw up such a

334 N. A. MACKINTOSH
list here and to subdivide it according to the three types distin-guished by Matthias in Lamellibranchs. The names in bracketsrefer to the authors who record the presence of a style in eachgenus.
Stage 1. Style free in the intestine or in wide and opencommunication therewith.
Patella (Gibson, 10), Gyclostoma (Garnault, 9), Fissurella(Haller, 12).
Stage 2. Style-sac in restricted communication with theintestine.
Urepidula (Orton, 23), Paludestrina and Hypsolia (Eobson,26, 28).
Stage 3. Style-sac completely separated from the intestine.Adeorbis (Woodward, 30), Aporrhais (Digby, 6), Bythoceras,
Chytra, Limnotrochus, Nassopsis, Paramelania (I find thata style is also present in Melania), Spekia, Tanganicia, andTyphobia (Moore, 18), Lithoglyphus (von Ihering, 14), Ptero-cera (Huxley, 13), Turritella (Bandies, 25).
Bithynia (Moquin-Tandon, 19), Bithynella (Bregenzer, 3),Ampullaria (Bouvier, 2), Eissoa (Simroth, 29), Calyptraeaand Capulus (Orton, 23), are also known to possess a crystallinestyle, but the relations between the style and intestine havenot been very clearly described. In Rissoa and Ampullaria noactual style is recorded, but its presence may be inferred fromthe characteristic sac shown in illustrations of the stomach.
The presence of a style in T r o c h u s t u r r i t us has beenrecorded by Collier (4), but, as is pointed out by Robson, thisis not mentioned by Haller or Randies, who have examined thisspecies. I have also examined a specimen of Trochus but wasunable to find any sign of a style.
The species possessing a style which I have actually examinedin addition to Crepidula are Paludestrina, Patella, and Melania.The style which occurs in Patella is very peculiar. If the animalbe removed from the shell and a loop of the intestine runninground the surface of the visceral hump be opened, an opaquewhite rod will be found therein. When sectioned this rod isfound to consist of co-axial layers of a granular substance similar
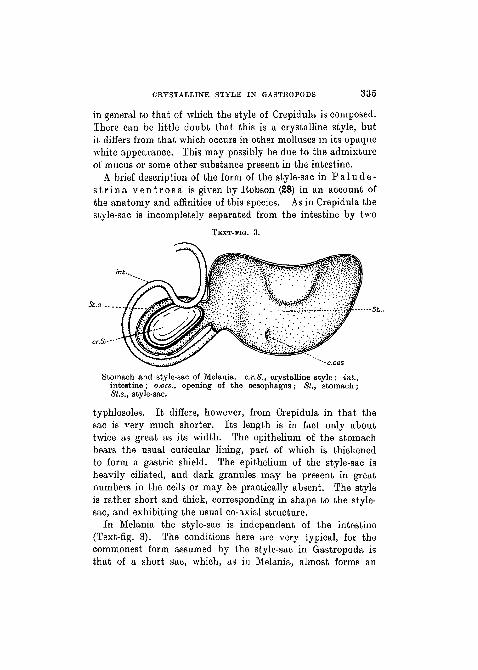
CRYSTALLINE STYLE IN GASTROPODS 335
in general to that of which the style of Crepidula is composed.There can be little doubt that this is a crystalline style, butit differs from that which occurs in other molluscs in its opaqueAvhite appearance. This may possibly be due to the admixtureof mucus or some other substance present in the intestine.
A brief description of the form of the style-sac in P a 1 u d e -s t r i n a v e n t r o s a is given by Eobson (28) in an account ofthe anatomy and affinities of this species. As in Crepidula thestyle-sac is incompletely separated from the intestine by two
TEXT-FIG. 3.
St.s
Stomach and style-sac of Melania. c.r.S., crystalline style; hit.,intestine; o.oe-s., opening of the oesophagus; St., stomach;St.s., style-sac.
typhlosoles. It differs, however, from Crepidula in that thesac is very much shorter. Its length is in fact only abouttwice as great as its width. The epithelium of the stomachbears the usual cuticular lining, part of which is thickenedto form a gastric shield. The epithelium of the style-sac isheavily ciliated, and dark granules may be present in greatnumbers in the cells or may be practically absent. The styleis rather short and thick, corresponding in shape to the style-sac, and exhibiting the usual co-axial structure.
In Melania the style-sac is independent of the intestine(Text-fig. 3). The conditions here are very typical, for thecommonest form assumed by the style-sac in Gastropoda isthat of a short sac, which, as in Melania, almost forms an

336 N. A. MACKINTOSH
extra, anterior chamber to the stomach and lies frequentlyin the ' crook ' of the intestine. Other examples of this aresupplied by Adeorbis, Aporrhais, Bythoceras, Chytra, Limno-trochus, Lithoglyphus, Nassopsis, Spekia, Tanganicia, andTyphobia.
The conditions in Pterocera are interesting as here the styleand style-sac, while bearing the same relations to the stomachand intestine as in the forms described above, are immenselyelongated. The style was first recorded by Huxley, but hasbeen more fully described by Woodward (30).
We are now in a position to make a general comparison ofthe crystalline style in Gastropoda with that of Lamellibranchia.
It has already been seen that the Gastropod style as it occurs inCrepidula bears an astonishing resemblance to that of Lamelli-branchs in respect of the form and function of the style and theanatomy of the style-sac, and it may confidently be stated thatthe style and its associated structures in this genus differ lessfrom such Lamellibranchs as Anodonta than many Lamelli-branchs differ from one another in this respect. The condi-tions which obtain in other Gastropods have also been reviewed,and it has been seen that they may be classified in the samethree categories as have been applied to Lamellibranchs inrespect of the relations of the style-sac and intestine. Thereis thus not only an individual but also a general resemblancebetween the two groups. It appears in fact that the onlydifferences are that whereas it is practically universal in Lamelli-branchs it is comparatively rare in Gastropods, and that ingeneral it is a more unobtrusive organ in the latter group.It has been pointed out that the commonest condition of thestyle in Gastropods is that in which it occurs as a short, bluntrod contained in a bulb-like sac which is generally not largerthan the stomach itself. In Lamellibranchs, on the other hand,the style, whether enclosed in a separate sac or not, is normallyof considerable length in comparison with the size of thestomach.
It is now necessary to consider what is the explanation ofthe occurrence of a crystalline style in Gastropods, of its

CRYSTALLINE STYLE IN GASTROPODS 337
appearance in only a few genera, and of its resemblance to theLamellibranch style.
Either the style is entirely homologous in the two groupsor it is a very striking example of parallel evolution. Nowif the resemblance of the style in the two groups rested onlyupon such features as are to be deemed inseparably correlatedwith its function, it might be that parallel evolution hadtaken place. But the extraordinary and detailed resemblanceof the style and style-bearing region in Crepidula to that ofcertain Lamellibranchs, in which the resemblance includessuch points as the shape and structure of the typhlosoles, thehistological details of the epithelium and the inner structureof the style itself, must lead to the conclusion that the organis strictly homologous in the two groups.
This being the case, two more alternatives must be con-sidered. Either the style was possessed by the commonancestor of both groups, and was retained in the Lamelli-branchs, but lost by all the Gastropods except for a few ;or we may accept the principle of orthogenesis and say thatthe tendency to produce a style was present in the commonancestor, that the tendency remained latent in the Gastropodsas a whole, but manifested itself in some cases, and that inLamellibranchs the presence of a style became general. It isdifficult to choose between these alternatives without enteringinto a discussion of the whole mechanism of evolution. Itshould, however, be pointed out that if the first of these alterna-tives be accepted it must be supposed that in such forms asCrepidula, where the style is more highly specialized than in,say, Patella, the similarity of the style to that of Lamelli-branchs is correlated with the similarity of the food, otherwisewe should expect to find a progressive divergence in the twogroups from the undifferentiated to the independent style-sac.
Whichever explanation is accepted, we have still to find theanswer to the question why a style is present in those particularGastropods which possess it. One cannot fail to be struck bythe fact that in Crepidula, whose style bears so close a resem-blance to that of Lamellibranchs, the food and feeding is also
NO. 274 Z

338 N. A. MACKINTOSH
strikingly similar to that of such forms as the oyster. It willtherefore be worth while to inquire whether any correlationcan be traced between the habits or environment and theoccurrence of a style in Gastropods. As the function of thestyle may be broadly defined as to assist in some special wayin the digestion of the food, we must look for this correla-tion in the nature of the food itself, in the method of feeding,or in anything which may have an indirect influence on thecondition of the food when it reaches the stomach, such asa fresh- or salt-water habitat.
Unfortunately the answer to this question cannot be finallysettled here, but the problem can be narrowed down to someextent.
So far as external environment is concerned, there appearsto be no correlation. The style is represented in salt-water,fresh-water, and terrestrial forms. It also occurs in activeand sluggish forms. From the example of Crepidula it mightbe suggested that the actual method of feeding, in which thefood is caught in a string of mucus, might be concerned insome way, but in Paludestrina, to take one example, the foodis taken in simply by the eversion and withdrawal of theradula. It therefore seems probable that if the actual presenceof a style is correlated in any way with the mode of life of theorganism it would be concerned with the nature of the fooditself. Unfortunately this suggestion cannot be definitelyverified as precise information on the food of Gastropods isvery scanty. We have, however, the outstanding fact thatthe food of Crepidula is identical with that of the oyster, andthe style is practically identical with that of certain Lamelli-branchs. If there is no correlation here it may at least besaid that the coincidence is a very striking one. In thisconnexion it should be pointed out that Patella, like Crepidulaand the oyster, very probably feeds largely on diatoms. Nowdiatoms are a very general article of food among Lamellibranchs,and it is possible that the appearance of a style is concernedin some way with the digestion of organisms of this type.

CRYSTALLINE STYLE IN GASTROPODS 3 3 9
4. SUMMARY.
1. Whereas in Laniellibranchia a crystalline style is verycommon, in Gastropoda it occurs in a few genera only.
2. Crepidula possesses a crystalline style contained in a sacwhich is partially differentiated from the intestine, beingseparated therefrom by dorsal and ventral typhlosoles.
The stomach bears a gastric shield and a peculiar groovewhich is the continuation of the oesophagus. The head of thestyle lies in the anterior part of the stomach and impinges onthe gastric shield.
8. The epithelium of the style-sac consists of densely ciliatedcells of moderate length in which the internal rootlets of thecilia extend through the length of the cell and become greatlythickened at the base. The epithelium is specially charac-terized by the presence of an ' intra-epithelial' network ofcanals containing numerous fibres whose function is probablya mechanical one. The same structures are present, thoughless prominent in the epithelium of the typhlosoles, the cells ofwhich are longer and narrower than those of the style-sacepithelium. "
4. The crystalline style is a straight transparent. rod ofgelatinous consistency. It is built up of co-axial layers sur-rounding a spiral core. It is composed principally of globulinand contains an amylolytic enzyme.
Pood is brought to the stomach embedded in a stream ofmucus which becomes entangled with the head of the style.The function of the style appears to be to mix together thecontents of the stomach and to supply a starch-splittingenzyme.
5. The crystalline style and style-bearing region of Crepidularesemble in great detail those of certain Larnellibranchs inrespect of the shape, proportions, and structure of the style-sac, typhlosoles, and intestine, of the presence of a gastricshield and of the form, composition, and function of the style.
6. The crystalline style in Gastropods may be classifiedunder the same three types as are distinguished in Lamelli-

840 N. A. MACKINTOSH
branchs in respect of the relations between the style-sac andintestine.
A list of Gastropods is given in which a style is known tooccur, and a brief account is given of the various forms assumedby the style and style-sac in this class.
7. The crystalline style of Crepidula resembles that ofLamellibranchs in such detail that this organ must be regardedas strictly homologous in the two groups. It is to be supposedeither that it has been lost in all but a few Gastropods, or thatits appearance in this group is to be explained on the principleof orthogenesis.
8. In view of the fact that the food of Crepidula is identicalwith that of the oyster, the retention or appearance of a stylein certain Gastropods might be correlated with the feedinghabits. Investigations in this direction, however, have notmet with very definite results.
5. L I T E R A T U R E .
1. Barrois, T. (1889—90).—" Le stylet cristallin des Lamellibranches ",' Revue biol. du Nord de la France', 3, 4, and 5.
2. Bouvier, E. (1880).—" fitude sur l'organiaation des Ampullaires ",' M£m. Soc. Philomathique Paris ' (centenary volume).
3. Bregenzer, A. (1916).—" Anatomie und Histologie von Bithnelladunkeri ", ' Zool. Jahrb. (Anat.)', 39, Heft 2.
4. Collier, C. (1829).—" Observations on univalves", ' New Philo-sophical Journal', 7.
5. Coupin, H. (1900).—" Sur les fonctions de la tige cristalline desAcephales ", ' C. R. Acad. Sci. Paris ', 130.
6. Digby, L. (1900).—" On the structure and affinities of the Tan-ganyika Gastropods Chytra and Limnotrochus ", ' Journ. Linn.Soc. (Zool.)', 28.
7. Edmondson, C. and H. (1920).—" The reformation of the crystallinestyle of Mya arenaria after extraction ", ' Journ. Exp. Zool.', 30.
8. Engelmann, T. W. (1880).—" Zur Anatomie und Physiologic derFlimmerzellen ", ' Pfliigers Arch.', 23.
9. Garnault, P. (1887).—" Recherches anatomiques et histologiques surle Cylostoma elegans ", ' Actes Soc. Linn. Bordeaux ' , 4 1 .
10. Gibson, R. J. H. (1885).—" Anatomy and physiology of Patellavulgata ", 'Trans. Roy. Soc. Edinburgh', 31.

CRYSTALLINE STYLE IN GASTROPODS 341
11. Gutheil, F. (1911).—" t)ber den Darmkanal und die Mitteldarmdriisevon Anodonta cellensis ", ' Zeitschr. wiss. Zool.', 99.
12. Haller, B. (1888).—" Die Morphologie der Prosobranchier ", ' Morph.Jahrb.', 14.
13. Huxley, T. H. (1853).—" On the morphology of the CephalousMollusca ", ' Phil. Trans. Roy. Soc.', 143, pt. 1.
14. von Ihering, H. (1885).—" Zur Kenntniss der Gattung Lithoglyphus ",' Mai. Blatt.', 7.
15. Matthias, M. (1914).—" Untersuehungen iiber den Darmkanal unddas Herz einiger Arcaceen ", ' Jena. Zeitschr. f. Naturwiss.', 52.
16. Mayer, L. (1913).—"Die intracellularen Fibrillen in den Epithel-zellen von Oligochaten und das Skelett der Muskelzellen ", ' Arch,f. Zellforschung ', 11.
17. Mitra, S. B. (1901).—"The crystalline style of Lamellibranehs",' Quart. Journ. Micr. Sci.', 44.
18. Moore, J. E. S. (1888-9).—" The Molluscs of the great African lakes ",ibid., 41-2.
19. Moquin-Tandon, A. (1885).—' Hist. Moll. France '.20. Nelson, T. C. (1918).—" On the origin, nature, and function of the
crystalline style of Lamellibranehs ", ' Journ. Morph.', 31.21. Orton, J. H. (1912).—" An account of the natural history of the Slipper
Limpet (Crepidula fornicata) ", ' Journ. Mar. Bio. Ass.', 9, no. 3.22. (1912).—" The mode of feeding in Crepidula, &c", ibid.23. (1922).—Letter to ' Nature ', 110, p. 149.24. Polowzow, W. (1904).—" t)ber contractile Fasern in einer Flimmer-
zellenart und ihre functionelle Bedeutung ", ' Arch. f. micr. Anat.',63. |
25. Randies, W. B. (1902).—" On the presence of a crystalline style inTurritella communis ", ' Anat. Anz.', 21.
26. Robson, G. C. (1921).—" On the anatomy and affinities of Hypsobianosophora ", ' Ann. and Mag. Nat. Hist.', 7.
27. (1922).—" On the connexion between the style-sac and intestinein Gastropoda and Lamellibranchia ", ' Proc. Mai. Soc.', 15, pt. 1.
28. (1922).—" On the anatomy and affinities of Paludestrina ven-trosa ",•' Quart. Journ. Micr. Sci.', 66.
29. Simroth, H.—' Bronn's Classen und Ordnungen des Thier-Reichs.Gastropoda Prosobranchia.'
30. Woodward, M. F. (1894).—" On the anatomy of Pterocera, with somenotes on the crystalline style ", ' Proc. Mai. Soc.', 1.
31. Yonge, C. M. (1923).—" Studies on the comparative physiology ofdigestion. I. The mechanism of feeding and assimilation in theLamellibranch Mya.", ' Brit. Journ. Bxp. Zoo!.', 1, no. 1.

342 N. A. MACKINTOSH
6. EXPLANATION OP PLATES 20 AND 21.
(All figures refer to Crepidula.)
ABBREVIATIONS.
a.c.sm., anterior chamber of stomach; b.gr., basal granules; ci, cilia;cr.s., crystalline styles; c.t., connective tissue; d.hep., ducts of hepato-pancreas ; gr., granules ; g.s., gastric shield ; hd.s., head of style ; hep.,hepatopancreas; i.b.c, internal bases of cilia; i.ep.c, ' intra-epithelial'canals; i.gr., intestinal groove ; i.r.c, internal rootlets of cilia; maj.t.,major typhlosole ; min.t., minor typhlosole ; vi.g., mucus glands ; n.,nucleus ; oes., oesophagus ; oes.gr., oesophageal groove ; o.oes., openingof the oesophagus ; ov., ovary; p.c.sm., posterior chamber of the stomach ;st.s., style-sac ; st.s.ep., style-sac epithelium.
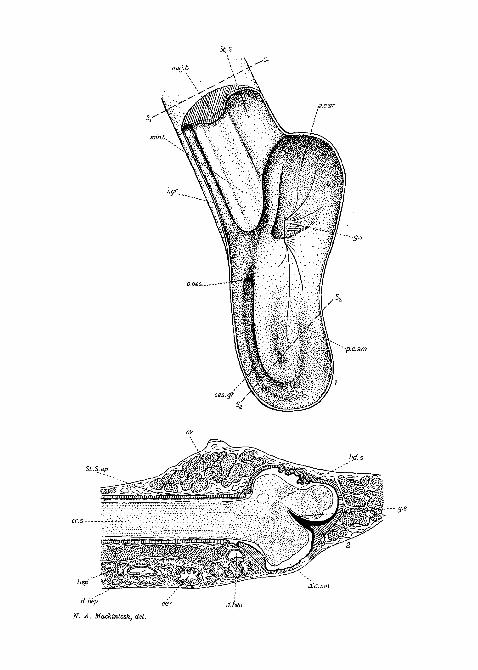
PLATE 20.
Fig. 1.—Dorsal view of the stomach. The dorsal wall has been removedfrom the stomach and the first part of the style-bearing region. Theminor typhlosole is thus seen in surface view and the major typhlosole insection.
Fig. 2.—Longitudinal section through the style-sac and style.Fig. 3.—Transverse section through the style-bearing region («j—sL in
fig. 1).Fig. 4.—The crystalline style.Fig. 5.—Vertical section through posterior part of stomach, showing
the oesophageal groove in transverse section (s2—s2 in fig. 1).
PLATE 21.
Fig. 6.—Transverse section of the style-sac epithelium.Fig. 7.—Transverse section of the epithelium of the major typhlosole.Fig. 8.—Frontal section of the style-sac epithelium, showing the ' intra-
epithelial * network. The larger canals are roughly parallel to the axisof the style-sac.
Fig. 9.—Mucus gland in the typhlosole epithelium.
-9-s
cr.s ; .--<:->
N. A. Mackintosh, del.
Quart. Journ. Micr. Sci. Vol. 69, N. S., PI. 30
>~crS
' < &
p.C.sm
oesfgr
Quart. Journ. Micr. Set. Vol. 69, N.S., PL 21
N. A. Mackintosh, del.