Embed Size (px)
Citation preview

Eur. J. Biochem. 30, 53-59 (1972)
The Cytoplasmic DNAs of Cultured Human Cells Effects of Ethidium Bromide on Their Replication and Maintenance
Jiirgen KOCH Institut fur Biologische Chemie und Ernahrungswissenschaft, Universitat Hohenheim, Stuttgart-Hohenheim
(Received July 19, 1972)
Ethidium bromide a t a concentration of 1 pgxml-l in the medium completely inhibits the synthesis of mitochondrial DNA (e = 1.706 g x ml-l) in exponentially growing Chang liver cells (ATCC; CCL 13), as indicated by the fact that labeled thymidine is not incorporated into any of the three forms of mitochondrial DNA (closed circular, open circular and linear DNA). Existing mitochondrial DNA undergoes a structural change under these growth conditions. At first closed circular DNA with a higher super-helix density is generated. After prolonged exposure of the cells to the drug (48 h) no remaining closed circular DNA is detectable in the mitochondria. The bulk of the remaining mitochondrial DNA behaves during sedimentation like a linear DNA molecule with the length of the circumference of the circular mitochondrial DNA. Removal of the drug from the growth medium reverses this process of interconversion of the different forms of mito- chondrial DNA.
No such drastic effects of ethidium bromide on the light cytoplasmic DNA (e = 1.688 g x ml-l) have been observed.
Ethidium bromide acts as a powerful inducer of petite mutants in yeast producing a respiratory deficient phenotype [l]. The same intercalating drug has proven to be a selective and potent inhibitor of the incorporation in vivo of labeled thymidine into the supercoiled mitochondrial DNA in mammalian cells [Z-51. Application of the drug to trypano- somes causes alterations in the structure of their kinetoplast DNA [6]. A specific inhibition of mito- chondrial DNA synthesis by ethidium bromide in Physarum polycephalum also has been reported [7].
With respect to yeast cells, it has been claimed that the treatment with the drug results in the com- plete destruction of the parental mitochondrial DNA and in the synthesis of a mitochondrial DNA with abnormal properties [8]. In contrast, the effects of ethidium bromide on mammalian cells seem to be reversible, for example the reduction of cytochrome oxidase [9] activity and the inhibition of the normal synthesis of supercoiled mitochondrial DNA [3]. There are conflicting reports about the fate of the supercoiled mitochondrial DNA during the exposure of mammalian cells to ethidium bromide. While some researchers claim that the supercoiled mito- chondrial DNA is degraded during exposure to the drug [2,3], others report that this treatment leads to the formation of a supercoiled mitochondrial DNA
Enzyme. Deoxyribonuolease or deoxyribonuclease oligo- nucleotido-hydrolase (EC 3.1.4.5).
with a higher super-helix density [5] , due to nicking- closing cycles in the presence of the drug. The im- portance of a possibly reversible degradation of the mitochondrial DNA is obvious in view of the question of its continuity.
This report describes the effects in vivo of ethi- dium bromide on mitochondrial DNA (e = 1.706 g x ml-l) and light cytoplasmic DNA (e = 1.688 g xml-l) [41 in cultured human liver cells (Chang). The synthesis of the mitochondrial DNA (closed circular, open circular as well as linear forms) is effectively inhibited in the presence of the drug. Preexisting supercoiled mitochondrial DNA is con- verted in vivo into linear DNA during prolonged exposure to the drug. However continued growth in a medium free of ethidium bromide leads to a reversal of this process. No such drastic effects on the incorpo- ration of labeled thymidine into the light cytoplasmic DNA have been observed. The experiments described permit an evaluation of a previously stated hypothesis that the light cytoplasmic DNA might be a precursor of the closed circular mitochondrial DNA [15].
MATERIAL AND METHODS Cell Culture
A serially propagated cell line derived from hu- man liver (Chang, ATCC: CCL 13) was grown in Eagle’s basal medium supplemented with fetal
54 Effect of Ethidium Bromide on Cytoplasmic DNAs Eur. J. Biochem.
calf serum, 30 pg x ml-l tetracycline (Achromycin, Lederle Div. of American Cyanamid Co., U.S.A.) and 200 pg x ml-l gentamycin (Refobacin, E. Merck AG, Germany) in screw-cap bottles (monolayers, 37 "C). The cell line was checked regularly for mycoplasma contamination with negative results [4]. Cells were labeled with [Me-SH]thymidine (20.2 Ci x mmol-1) and [2-14C]thymidine (60 mCi x mmol-l) obtained from The Radiochemical Centre (Amersham, GB).
Cell Practionation and Isolation of DNA
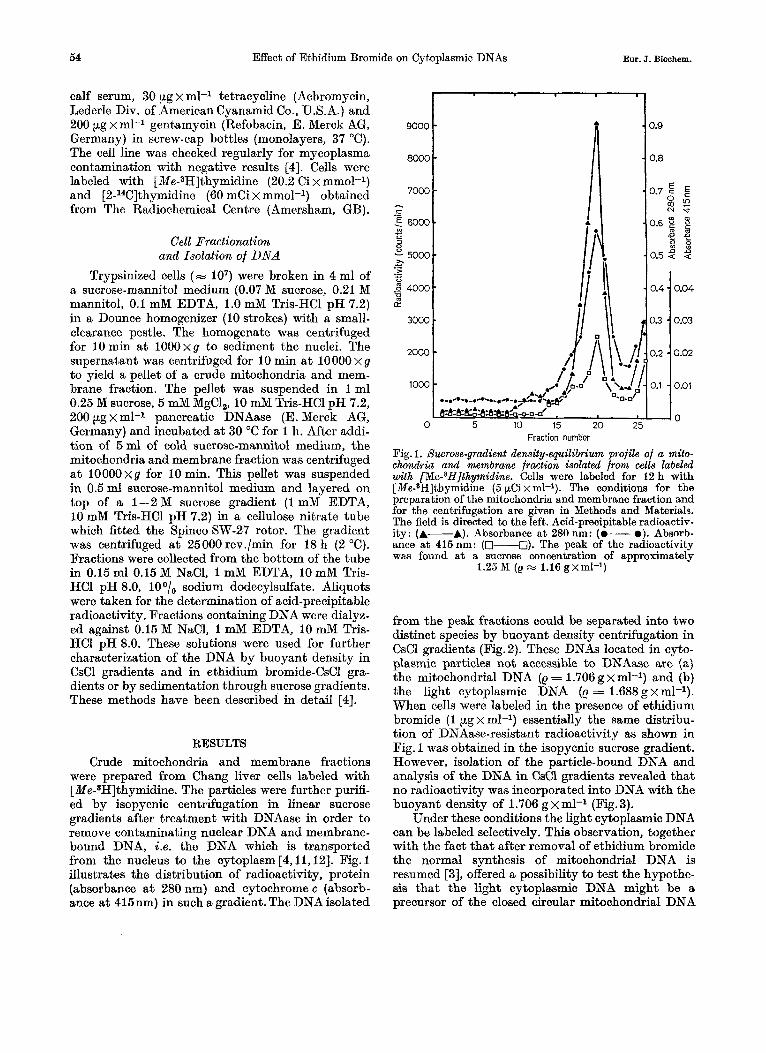
Trypsinized cells (- lo7) were broken in 4 ml of a sucrose-mannitol medium (0.07 M sucrose, 0.21 M mannitol, 0.1 mM EDTA, 1.0 mM Tris-HC1 pH 7.2) in a Dounce homogenizer (10 strokes) with a small- clearance pestle. The homogenate was centrifuged for 10 min a t 100Oxg to sediment the nuclei. The supernatant was centrifuged for 10 min a t 100OOxg to yield a pellet of a crude mitochondria and mem- brane fraction. The pellet was suspended in 1 ml 0.25 M sucrose, 5 mM MgCl,, 10 mM Tris-HC1 pH 7.2, 200 pg x ml-I pancreatic DNAase (E. Merck AG, Germany) and incubated a t 30 "C for 1 h. After addi- tion of 5ml of cold sucrose-mannitol medium, the mitochondria and membrane fraction was centrifuged a t 100OOxg for 10min. This pellet was suspended in 0.5ml sucrose-mannitol medium and layered on top of a 1-2 M sucrose gradient (1 mM EDTA, I0 mM Tris-HC1 pH 7.2) in a cellulose nitrate tube whioh fitted the SpincoSW-27 rotor. The gradient was centrifuged a t 25000 rev./min for 18 h (2 "C). Fractions were collected from the bottom of the tube in 0.15 ml 0.15 M NaC1, 1 mM EDTA, 10 mM Tris- HC1 pH 8.0, loo/, sodium dodecylsulfate. Aliquots were taken for the determination of acid-precipitable radioactivity. Fractions containing DNA were dialye- ed against 0.15 M NaC1, 1 mM EDTA, 10 mM Tris- HCI pH 8.0. These solutions were used for further characterization of the DNA by buoyant density in CsCl gradients and in ethidium bromide-CsCl gra- dients or by sedimentation through sucrose gradients. These methods have been described in detail [4].
RESULTS Crude mitochondria and membrane fractions
were prepared from Chang liver cells labeled with [Me-8H]thymidine. The particles were further purifi- ed by isopycnic centrifugation in linear sucrose gradients after treatment with DNAase in order to remove contaminating nuclear DNA and membrane- bound DNA, i .e. the DNA which is transported from the nucleus to the cytoplasm [4,11,12]. Fig. 1 illustrates the distribution of radioactivity, protein (absorbance at 280 nm) and cytochrome c (absorb- ance a t 415nm) in such zl gradient. The DNA isolated
10 15 20 25 0 5 Fraction number
Fig. 1. Sucrose-gradient density-equilibrium profile of a mito- chondria and membrane fraction isolated from cells labeled with [Me-8H]thymidine. Cells were labeled for 12 h with [Me-8H]thymidine (5 pCi x ml-I). The conditions for the preparation of the mitochondria and membrane fraction and for the centrifugation are given in Methods and Materials. The field is directed to the left. Acid-precipitable radioactiv- ity: (A-A). Absorbance a t 280 nm: (0- e). Absorb- ance a t 415nm: (-0). The peak of the radioactivity was found a t a sucrose concentration of approximately
1.25 M (e x 1.16 gxml-I)
from the peak fractions could be separated into two distinct species by buoyant density centrifugation in CsCl gradients (Fig.2). These DNAs located in cyto- plasmic particles not accessible to DNAase are (a) the mitochondrial DNA (p = 1.706 g x ml-l) and (b) the light cytoplasmic DNA (e = 1.688 g x ml-l). When cells were labeled in the presence of ethidium bromide (1 pg x ml-l) essentially the same distribu- tion of DNAase-resistant radioactivity as shown in Fig. 1 was obtained in the isopycnic sucrose gradient. However, isolation of the particle-bound DNA and analysis of the DNA in CsCl gradients revealed that no radioactivity was incorporated into DNA with the buoyant density of 1.706 g x ml-l (Fig. 3).
Under these conditions the light cytoplasmic DNA can be labeled selectively. This observation, together with the fact that after removal of ethidium bromide the normal synthesis of mitochondrial DNA is resumed [3], offered a possibility to test the hypothe- sis that the light cytoplasmic DNA might be a precursor of the closed circular mitochondrial DNA
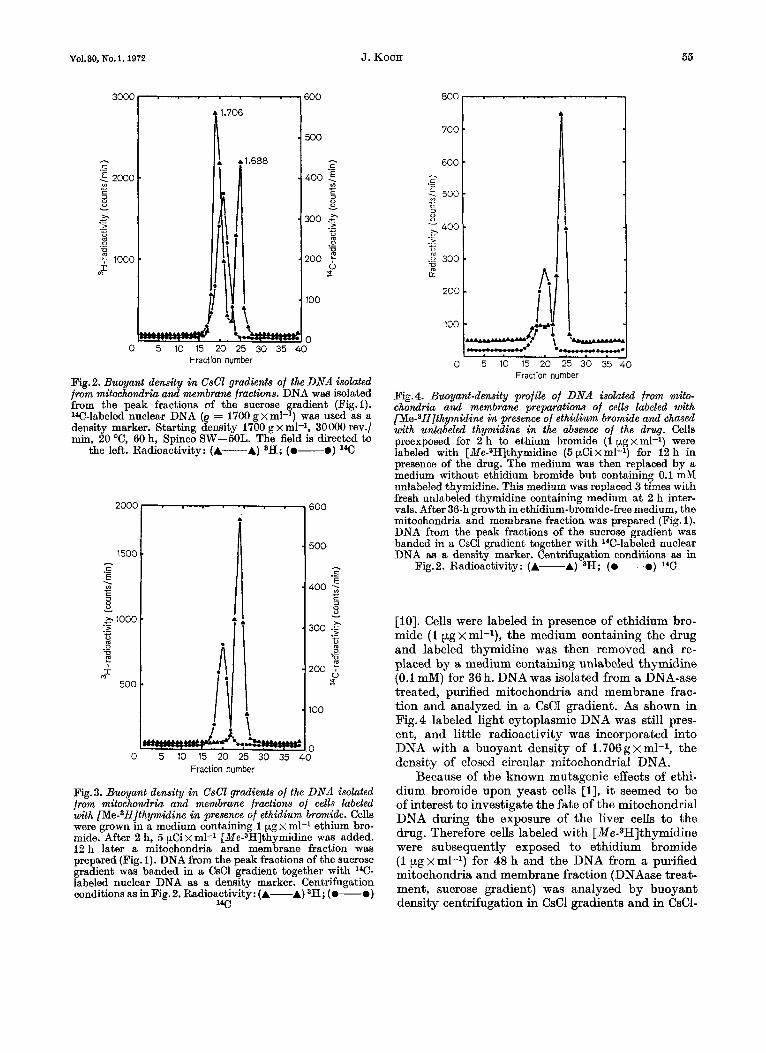
Vo1.30,No.l. 1972 J. KOCH 56
Fraction number
Fig.2. Buoyant density in CsCl gradients of the DNA isolated from mitochondria and membram fractions. DNA was isolated from the peak fractions of the sucrose gradient (Fig.1). W-labeled nuclear DNA (e = 1700 g x ml-l) was used as a density marker. Starting density 1700 g x rnl-l, 30000 rev./ min, 20 "C, 60 h, Spinco SW-5OL. The field is directed to
the left. Radioactivity: (A-A) aH; (0-0) lCC
2000 I . . . . . , ,
1500 - c .- E . v)
E 2 I
8 3 1000 - ._ >
u m .- I
._ P r
500
0 Fraction number
Fig. 3. Buoyant density in CsCl gradients of the DNA isolated from mitochondria and membrane fractions of cells labeled with [Me-3H]thymidine in presence of ethidium bromide. Cells were grown in a medium containing 1 pg x ml-1 ethium bro- mide. After 2 h, 5 pCi x ml-l [Me-SH]thymidine was added. 12 h later a mitochondria and membrane fraction was prepared (Fig. 1). DNA from the peak fractions of the sucrose gradient was banded in a CsCl gradient together with lac- labeled nuclear DNA as a density marker. Centrifugation conditions as inFig.2. Radioactivity: (A-A) 3H; (0-0)
"C
8001 . . . . . . .
700
600
200
100
*--..- 0 5 10 15 20 25 30 35 3
Fraction number
Pig.4. Buoyant-density profile of DNA isolated from mito- chondria and membrane preparations of cells labeled with [Me-SH]thymidine in presence of ethidium bromide and chased with unlabeled thymidine in the absence of the drug. Cells preexposed for 2 h to ethium bromide (1 pgxml-l) were labeled with [Me-3H]thymidine (5 pCixml-l) for 12 h in presence of the drug. The medium was then replaced by a medium without ethidium bromide but containing 0.1 mM unlabeled thymidine. This medium was replaced 3 times with fresh unlabeled thymidine containing medium at 2 h inter- vals. After 36-h growth in ethidiurn-bromide-free medium, the mitochondria and membrane fraction was prepared (Fig. 1). DNA from the peak fractions of the sucrose gradient was banded in a CsCl gradient together with 14C-labeled nuclear DNA a5 a density marker. Centrifugation conditions as in
Fig.2. Radioactivity: (A-A) SH; (0-0) 1*C
[lo]. Cells were labeled in presence of ethidium bro- mide (1 pg x ml-l), the medium containing the drug and labeled thymidine was then removed and re- placed by a medium containing unlabeled thymidine (0.1 mM) for 36 h. DNA was isolated from a DNA-ase treated, purified mitochondria and membrane frac- tion and analyzed in a CsCl gradient. As shown in Fig.4 labeled light cytoplasmic DNA was still pres- ent, and little radioactivity was incorporated into DNA with a buoyant density of 1.706 g x ml-l, the density of closed circular mitochondria1 DNA.
Because of the known mutagenic effects of ethi- dium bromide upon yeast cells [I], it seemed to be of interest to investigate the fate of the mitochondria1 DNA during the exposure of the liver cells to the drug. Therefore cells labeled with [Me-3H]thymidine were subsequently exposed to ethidium bromide (1 pg x ml-l) for 48 h and the DNA from a purified mitochondria and membrane fraction (DNAase treat- ment, sucrose gradient) was analyzed by buoyant density centrifugation in CsCl gradients and in CsC1-
56 Effect of Ethidium Bromide on Cytoplasmic DNAs Eur. J. Biochem.
ethidium bromide gradients and by band sedimenta- tion. The medium of the cells serving as a control did not contain ethidium bromide during the 48-h chase.
3000
- .; 2000 . v)
c c
s - 1.
z 0
0
- ._ ._ c
._ P 1000 cz
0
I ' ' = " A l 6 A
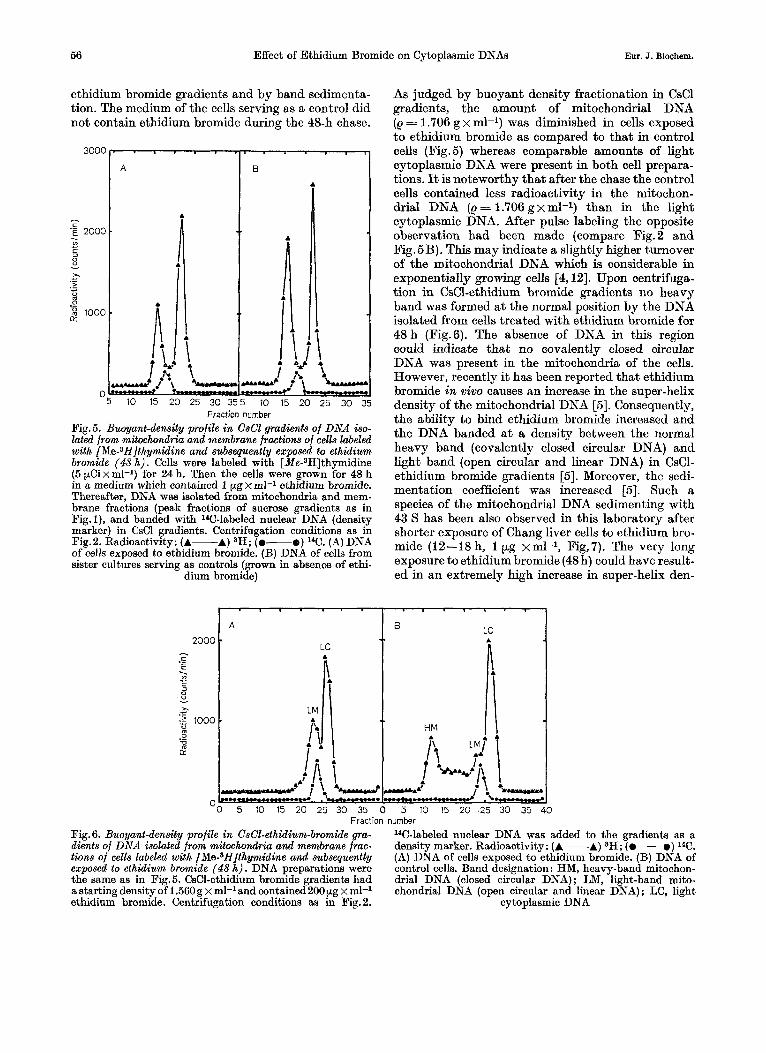
Fraction number Fig.5. Buoyant-density profile in CsCl gradients of DNA iso- lated from mitochondria and membrane fractions of cells labeled with [Me-SHIthymidine and subsequently exposed to ethidium bromide (48 h) . Cells were labeled with [Me-SH]thymidine (5 yCixml-l) for 24 h. Then the cells were grown for 48 h in a medium which contained 1 pgxml-l ethidium bromide. Thereafter, DNA was isolated from mitochondria and mem- brane fractions (peak fractions of sucrose gradients as in Fig. i), and banded with l*C-labeled nuclear DNA (density marker) in CsCl gradients. Centrifugation conditions as in Fig.2. Radioactivity: (A-A) 3H; (0-0) lac. (A) DNA of cells exposed to ethidium bromide. (B) DNA of cells from sister cultures serving as controls (grown in absence of ethi-
dium bromide)
As judged by buoyant density fractionation in CsCl gradients, the amount of mitochondrial DNA (e = I .706 g x ml-l) was diminished in cells exposed to ethidium bromide as compared to that in control cells (Fig. 5) whereas comparable amounts of light cytoplasmic DNA were present in both cell prepara- tions. It is noteworthy that after the chase the control cells contained less radioactivity in the mitochon- drial DNA (e = 1.706 gxml-l) than in the light cytoplasmic DNA. After pulse labeling the opposite observation had been made (compare Fig. 2 and Fig. 5B). This may indicate a slightly higher turnover of the mitochondrial DNA which is considerable in exponentially growing cells [4, i2]. Upon centrifuga- tion in CsC1-ethidium bromide gradients no heavy band was formed a t the normal position by the DNA isolated from cells treated with ethidium bromide for 48 h (Fig.6). The absence of DNA in this region could indicate that no covalently closed circular DNA was present in the mitochondria of the cells. However, recently it has been reported that ethidium bromide in vivo causes an increase in the super-helix density of the mitochondrial DNA [5]. Consequently, the ability to bind ethidium bromide increased and the DNA banded a t a density between the normal heavy band (covalently closed circular DNA) and light band (open circular and linear DNA) in CsC1- ethidium bromide gradients [5]. Moreover, the sedi- mentation coefficient was increased [5]. Such a species of the mitochondrial DNA sedimenting with 43 S has been also observed in this laboratory after shorter exposure of Chang liver cells to ethidium bro- mide (12- 18 h, 1 pg x ml-1, Fig, 7). The very long exposure to ethidium bromide (48 h) could have result- ed in an extremely high increase in super-helix den-
2000 - c ._ E . v)
c 0
m
CK
I
- % - .- .z 1000
P 0 ._
' 0 5 10 15 20 25 30 35 0 5 10 15 20 25 30 35 40 Fraction number
Fig. 6. Buoyant-density profile in CsCl-ethidium-bromide gra- dients of DNA isolated from mitochondria and membrane frac- tions of cells labeled with [Me-3H]thymidine and subsequently exposed to ethidium bromide (48 h ) . DNA preparations were the same as in Fig.5. CsC1-ethidium bromide gradients had a starting density of 1.560g x ml-land contained200pg x ml-l ethidium bromide. Centrifugation conditions as in Fig. 2.
l4C-labe1ed nuclear DNA was added to the gradients as a density marker. Radioactivity: (A-A) 3fI; (0-0) lac. (A) DNA of cells exposed to ethidium bromide. (B) DNA of control cells. Band designation: HM, heavy-band mitochon- drial DNA (closed circular DNA); LM, light-band mito- chondrial DNA (open circular and linear DNA) ; LC, light
cytoplasmic DNA
Vo1.30, No.1. 1972 J. KOCH 57
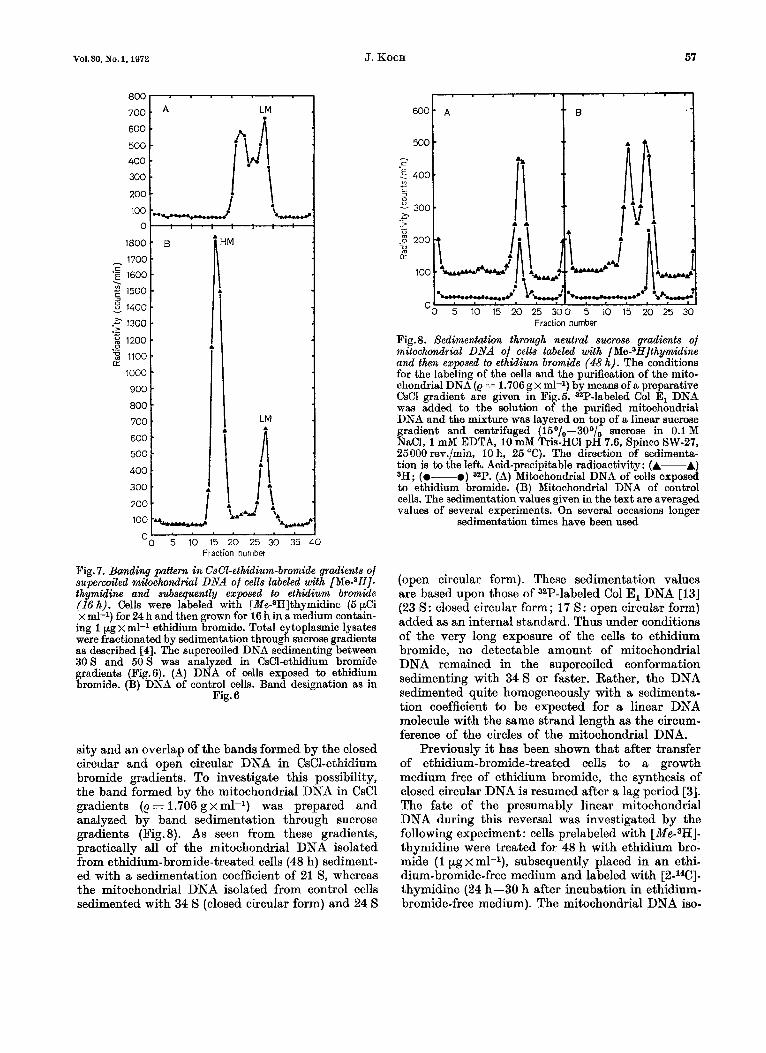
600 . 500 - 400 . 300 . 200 i . 100 'A%-
-0 5 10 15 20 25 300 5 10 15 20 25 30 Fraction number
Fig. 8. Sedimentation through neutral sucrose gradients of mitochondrial DNA of cells labeled with [Me-3HJthymidine and then exposed to ethidium bromide (48 h). The conditions for the labeling of the cells and the purification of the mito- chondrial DNA (e = 1.706 g X ml-l) by means of a preparative CsCl gradient are given in Fig.5. S2P-labeled Col E, DNA was added to the solution of the purified mitochondrial DNA and the mixture was lavered on to^ of a linear sucrose gradient and centrifuged ( i5°/0-300/; sucrose in 0.1 M NaCI, 1 mM EDTA, 10 mM Tris-HC1 DH 7.6, S~inco SW-27. 25000 rev./min, 10h, 25 "C). The dfrection kf sedimenta: tion is to the left. Acid-precipitable radioactivity: (A-A) 3H; (0-0) (A) Mitochondrial DNA of cells exposed to ethidium bromide. (B) Mitochondrial DNA of control cells. The sedimentation values given in the text are averaged values of several experiments. On several occasions longer
sedimentation times have been used
vO 5 10 15 20 25 30 35 40 Fraction number
Fig. 7. Banding pattern in CsCl-ethidium-bromide gradients of superwiled mitochondrial DNA of cells labeled with [Me-8H]- thymidine and subsequently exposed to ethidium bromide (16 h). Cells were labeled with [Me-8H]thymidine (6 $3 x ml-1) for 24 h and then grown for 16 h in a medium contain- ing 1 pg x ml-1 ethidium bromide. Total cytoplasmic lysates were fractionated by sedimentation through sucrose gradients as described [4]. The supercoiled DNA sedimenting between 30s and 5 0 s was analyzed in CsC1-ethidium bromide gradients (Fig.6). (A) DNA of cells exposed to ethidium bromide. (€3) DNA of control cells. Band designation as in
Fig. 6
sity and an overlap of the bands formed by the closed circular and open circular DNA in CsC1-ethidium bromide gradients. To investigate this possibility, the band formed by the mitochondrial DNA in CsCl gradients (e = 1.706 g x ml-l) was prepared and analyzed by band sedimentation through sucrose gradients (Fig.8). As seen from these gradients, practically all of the mitochondrial DNA isolated from ethidium-bromide-treated cells (48 h) sediment- ed with a sedimentation coefficient of 21 s, whereas the mitochondrial DNA isolated from control cells sedimented with 34 S (closed circular form) and 24 S
(open circular form). These sedimentation values are based upon those of 32P-labeled Col El DNA [13] (23 S: closed circular form; 17 S: open circular form) added as an internal standard. Thus under conditions of the very long exposure of the cells to ethidium bromide, no detectable amount of mitochondrial DNA remained in the supercoiled conformation sedimenting with 3 4 s or faster. Rather, the DNA sedimented quite homogeneously with a sedimenta- tion coefficient to be expected for a linear DNA molecule with the same strand length as the circum- ference of the circles of the mitochondrial DNA.
Previously it has been shown that after transfer of ethidium-bromide-treated cells t o a growth medium free of ethidium bromide, the synthesis of closed circular DNA is resumed after a lag period [3]. The fate of the presumably linear mitochondrial DNA during this reversal was investigated by the following experiment : cells prelabeled with [Me-*H]- thymidine were treated for 48 h with ethidium bro- mide (1 pg x ml-l), subsequently placed in an ethi- dium-bromide-free medium and labeled with [2-14C]- thymidine (24 h-30 h after incubation in ethidium- bromide-free medium). The mitochondrial DNA iso-
68 Effect of Ethidium Bromide on Cytoplasmic DNAs Eur. J. Biochem.
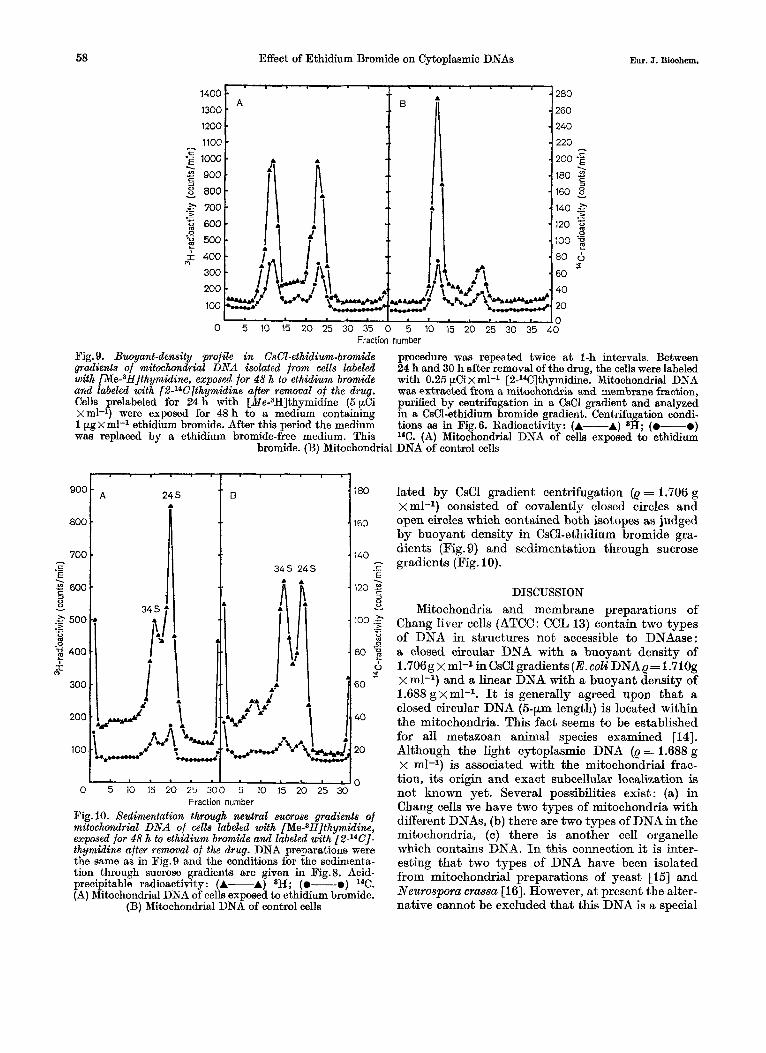
- 260
1200 - - 240
1300 -
0 5 10 15 20 25 30 35 0 5 10 15 20 25 30 35 40 Fraction number
Fig. 9. Buoyant-density profile in CsCl-ethidium-bromide procedure was repeated twice a t l-h intervals. Between gradients of mitochondrial DNA isolated from cells labeled 24 h and 30 h after removal of the drug, the cells were labeled with [Me-3H]thymidine, exposed for 48 h to ethidium bromide with 0.25 pCi x ml-l [2-14C]thymidine. Mitochondrial DNA and labeled with [2J4C]thymidine after removal of the drug. waa extracted from a mitochondria and membrane fraction, Cells prelabeled for 24 h with [Me-3H]thymidine (5 pCi purified by centrifugation in a CsCl gradient and analyzed xml-l) were exposed for 48 h to a medium containing in a CsC1-ethidium bromide gradient. Centrifugation condi- 1 pg x ml-1 ethidium bromide. After this period the medium tions as in Fig. 6. Radioactivity: (A-A) 8H; (0-0) was replaced by a ethidium bromide-free medium. This lac. (A) Mitochondrial DNA of cells exposed to ethidium
bromide. (B) Mitochondrial DNA of control cells
A 24 S
. I . I , ~ -
B
345 2 4 s
0 5 10 15 20 25 300 5 10 15 20 2 5 3 Fraction number
I80
60
- I40 c __ E .
120 2
I00 .$
30 p i,
3 8 u
.- - " 0 .-
0 50
40
20
5
Fig. 10. Xedimentation through neutral sucrose gradients of mitochondrial DNA of cells labeled with [Me-3H]thymidine, exposed for 48 h to ethidium bromide and labeled with [2-14C]- thymidine after removal of the drug. DNA preparations were the same as in Fig.9 and the conditions for the sedimenta- tion through sucrose gradients are given in Fig.8. Acid- precipitable radioactivity: (A-A) 3H; (.-.) 14C. (A) Mitochondrial DNA of cells exposed to ethidium bromide.
(B) Mitochondrial DNA of control cells
lated by CsCl gradient centrifugation (e = 1.706 g x ml-l) consisted of covalently closed circles and open circles which contained both isotopes as judged by buoyant density in CsC1-ethidium bromide gra- dients (Fig. 9) and sedimentation through sucrose gradients (Fig. lo).
DISCUSSION Mitochondria and membrane preparations of
Chang liver cells (ATCC: CCL 13) contain two types of DNA in structures not accessible to DNAase: a closed circular DNA with a buoyant density of 1.706g x ml-linCsClgradients(E.coliDNA~=l.710g x ml-l) and a linear DNA with a buoyant density of 1.688 g x ml-I. It is generally agreed UPOR that a closed circular DNA (5-pm length) is located within the mitochondria. This fact seems to be established for all metazoan animal species examined [la]. Although the light cytoplasmic DNA (e = 1.688 g x ml-l) is associated with the mitochondrial frac- tion, its origin and exact subcellular localization is not known yet. Several possibilities exist: (a) in Chang cells we have two types of mitochondria with Merent DNAs, (b) there are two types of DNA in the mitochondria, (c) there is another cell organelle which contains DNA. In this connection it is inter- esting that two types of DNA have been isolated from mitochondrial preparations of yeast [ 151 and Neurosporu crussa [16]. However, a t present the alter- native cannot be excluded that this DNA is a special

V01.30, No. 1, 1972 J. KOCH 59
feature of Chang cells. This would not be too sur- prising in view of the results obtained with the membrane-bound cytoplasmic DNA which is not common to all human cells studied [12].
It has been hypothesized that the light cyto- plasmic DNA might be a precursor of the closed circular DNA [lo]. The present results do not support this idea. Ethidium bromide completely inhibits the incorporation of labeled thymidine into the mito- chondrial DNA (e = 1.706 g x ml-l) but permits the labeling of the light cytoplasmic DNA. During a chase period in the absence of ethidium bromide the synthesis of mitochondrial DNA is resumed [3]. If the above stated hypothesis were correct, then one would expect that during this chase period, the radioactivity would move from the precursor into the closed circular mitochondrial DNA. Although some radioactivity appeared in the mitochondrial DNA in these experiments, the amount has not been sufficient to justify the assumption that the light cytoplasmic DNA as an entity is the precursor of the mitochondrial DNA. The origin of the small amount of the radioactivity which is found in the mitochondrial DNA after such a chase remains enig- matic. A second argument against a precursor- product relationship of these DNAs is the fact that the turnover of the presumed product is found to be somewhat higher than that of the precursor.
Ethidium bromide not only inhibits the synthesis of the mitochondrial DNA but also brings about a structural change in the existing mitochondrial DNA. The first change is that the supercoiling in- creases. This phenomenon has been attributed to nicking and reclosing of the unwound (by the inter- calating dye) DNA in vivo. After isolation and remo- val of the dye the DNA obtains a higher super-helix density [5]. Upon prolonged exposure of the cells to ethidium bromide, the mitochondrial DNA mole- cules evidently cannot be reclosed and eventually a conversion into linear DNA occurs. The relation of the sedimentation values of 21 S for the linear form, 24 S for the open circular form and 34 S for the closed circular form are in good agreement with the relative sedimentation coefficients of the three forms found in the I-system [17,18]. A further analogy with the
phage A-system is suggested by the fact that the linear DNA can be converted in vivo into closed circular DNA after removal of ethidium bromide. It will be of interest to see whether this linear mito- chondrial DNA possesses cohesive ends which can interact to form hydrogen-bonded circles.
I thank Mrs Helga v. Pfeil for excellent technical assi- stance, Dr W. Goebel for a kind gift of 32P-labeled Col E, DNA and the E. Merck AG (Darmstadt) for the generous supply of gentamycin. This work was supported by a grant from the Deutsche Forschungsgemeinschaft, Bonn-Bad Godesberg.
REFERENCES 1. Slonimski, P. P., Perrodin, G. & Croft, J. H. (1968) Bio-
2. Ness, M . M. K. (1970) Proc. Nut. Acad. Sci. U . S. A . 67, chem. Biophys. Res. Commun. 30, 232-239.
1926-1933. 3. Koch, J. & v. Pfeil, H. (1971) FEBSLett. 18,172-174. 4. Koch, J. (1972) Eur. J . Biochem. 26, 259-266. 5. Smith,C.A., Jordan, J.M. & Vinograd, J. (1971) J .
6. Riou, G. & Delain, E. (1969) Proc. Nut. A d . Sci.
7. Horwitz, H. B. & Holt, C. E. (1971) J . CeEZ Biol. 49,
8. Perlman, P. S. & Mahler, H. R. (1971) Nature New Biol.
9. Naum, Y . & Pious, D. A. (1971) Exp. Cell Res. 65,
MoE. Bi01. 59, 255-272.
U . S. A . 64, 618-625.
546-553.
231, 12-16.
335-339. 10. Koch, J. (1969) Eur. J . Biochem. 11, 296-304. 11. Koch, J. & v. Pfeil, H. (1971) FEBS Lett. 17, 312-314. 12. Koch, J. & v. Pfeil, H. (1972) FEBS Lett. 24, 53-56. 13. Bazaral, M., & Helsinki, D. R. (1969) J . Mol. Biol. 36,
14. Ashwell, M. & Work, T. S. (1970) Annu. Rev. Biochem.
15. Clark-Walker, G. D. (1972) Proc. Nut. Acad. Sci. U . S. A.
16. Reich, E. & Luck, D. J. L. (1966) Proc. Nut. Acad. Sci.
17. Hershey, A. D., Burgi, E. & Ingraham, I. (1963) Proc.
18. Bode, V. C. & Kaiser, A. D. (1965) J . Mol. Biol. 14,
185 - 194.
39, 251-290.
69, 388-392.
U . S. A . 55, 1600-1608.
Nut. A d . Sci. U. 5. A . 49, 748-755.
399-417.
J. Kook Institut fur Bioloizische Chemie und Ernahrungsw&senschaft Universitiit Hohenheim D-7000 Stuttgart-Hohenheim, GarbenstraBe 30 Federal Republic of Germany




![6 -Fluoro[4.3.0]bicyclo nucleic acid: synthesis ... · acids (LNAs) [13-15], hexitol nucleic acids (HNAs) [16,17], cyclohexenyl nucleic acids (CeNAs) [18,19], tricyclo-DNAs (tc-DNAs,](https://img.pdfslide.net/doc/110x75/600a55cffb08ea5e44429f99/6-fluoro430bicyclo-nucleic-acid-synthesis-acids-lnas-13-15-hexitol.jpg)