Embed Size (px)
Citation preview

Brain Research 958 (2002) 459–462www.elsevier.com/ locate/brainres
Short communication
T he difference of osteocalcin-immunoreactive neurons in the rat dorsalroot and trigeminal ganglia: co-expression with nociceptive transducers
and central projectiona,b , a,b*Hiroyuki Ichikawa , Tomosada Sugimoto
aDepartment of Oral Function and Anatomy, Okayama University Graduate School of Medicine and Dentistry, 2-5-1 Shikata-cho,Okayama 700-8525, Japan
bBiodental Research Center, Okayama University Graduate School of Medicine and Dentistry, 2-5-1 Shikata-cho, Okayama 700-8525, Japan
Accepted 23 September 2002
Abstract
The co-expression of osteocalcin (OC) with the capsaicin receptor (VR1) and vanilloid receptor 1-like receptor (VRL-1) was examinedin the dorsal root (DRG) and trigeminal ganglia (TG). Virtually all OC-immunoreactive (ir) DRG neurons were devoid of VR1- andVRL-1-immunoreactivity (ir). In the TG, 14.1% of OC-ir neurons were also immunoreactive for VR1. Only 1.7% of OC-ir TG neuronsco-expressed VRL-1-ir. The distribution of OC-ir was also examined in the spinal cord and trigeminal sensory nuclei. In the spinal cord,the superficial laminae of the dorsal horn were devoid of OC-ir. The neuropil was weakly stained in other regions of the spinal horns. Themedullary dorsal horn (MDH) contained numerous OC-ir varicose fibers in laminae I and II. These fibers were occasionally observedoriginating from the spinal trigeminal tract. The neuropil was weakly stained in deeper laminae of the MDH, and the rostral parts of thetrigeminal sensory nuclei. The present study suggests that OC-ir TG nociceptors send their unmyelinated axons to the superficial laminaeof the MDH. 2002 Elsevier Science B.V. All rights reserved.
Theme: Sensory systems
Topic: Somatic and visceral afferents
Keywords: Immunohistochemistry; Osteocalcin; Trigeminal ganglion; Trigeminal sensory nucleus; VR1; VRL-1
Osteocalcin (OC), an extracellular protein of bone, has a CGRP-ir primary sensory neurons supply their peripheralmolecular weight of 5700 and contains three calcium- receptive field with free nerve endings, and are thought tobinding amino acid residues [3,6,12]. This calcium-binding participate in nociception [11,14]. Therefore, OC-immuno-protein (CaBP) is synthesized by osteoblasts, accumulates reactivity (ir) may be also expressed by the trigeminalin bone, and regulates bone formation [3,6]. Recently we primary nociceptive neurons.have demonstrated the presence of OC in primary sensory The capsaicin (vanilloid) receptor VR1 is a nociceptiveneurons [9]. In the dorsal root (DRG) and trigeminal (TG) transducer which can be activated by vanilloid compounds,ganglia, OC-immunoreactivity (-ir) was predominantly protons and heat (.43 8C) [1]. In the DRG and TG,VR1 islocalized to large neurons. These neurons are considered to localized to small and medium-sized neurons [4,8]. Suchinclude muscular proprioceptors and low-threshold mech- neurons supply their peripheral receptive fields with freeanoreceptors. In the TG, however, a substantial proportion nerve endings [4]. In addition, VR1-ir DRG neuronsof OC-immunoreactive (-ir) neurons co-express calcitonin project to the superficial dorsal horn of the spinal cord [4].gene-related peptide (CGRP), a marker specific to small to On the other hand, vanilloid receptor 1-like receptormedium-sized neurons in the sensory ganglia [9–11,15]. (VRL-1), a newly cloned capsaicin-receptor homologue, is
localized to medium-sized and large neurons in the DRGand TG [2,9]. This transducer dose not respond to cap-*Corresponding author. Fax:181-86-235-6612.
E-mail address: [email protected](H. Ichikawa). saicin, and is activated by high temperatures with a
0006-8993/02/$ – see front matter 2002 Elsevier Science B.V. All rights reserved.PI I : S0006-8993( 02 )03701-0

460 H. Ichikawa, T. Sugimoto / Brain Research 958 (2002) 459–462
threshold .52 8C [2]. VRL-1-ir DRG and TG neurons ImmunoResearch Labs) and lissamine rhodamine B chlo-project to the superficial laminae of the spinal and medul- ride-conjugated donkey anti-rabbit IgG (1:500, Jacksonlary dorsal horns, respectively [2,9]. The co-expression of ImmunoResearch Labs).VR1and VRL-1 with CGRP is also demonstrated in the For demonstration of OC in the spinal cord and trigemi-sensory ganglia [2,4,8,7]. nal sensory nuclei, the spinal cord and brainstem were
In this study, we examine the co-expression of OC with frozen-sectioned at 50mm and treated as free-floatingVR1 and VRL-1 in the DRG and TG to investigate sections. Sections were incubated with goat anti-OC serumwhether primary nociceptive neurons contain this CaBP. (1:2000) for 24 h at room temperature, followed byThe distribution of OC-ir was also investigated in the biotinylated horse anti-mouse IgG and avidin–biotin–spinal cord and trigeminal sensory nuclei (principal sen- horseradish peroxidase complex (Vector Laboratories).sory nucleus and spinal trigeminal nucleus) to search the Following nickel ammonium sulfate-intensifiedcentral projection of OC-ir primary sensory neurons. diaminobenzidine reaction, the sections were dry-mounted
Eight DRGs and two spinal cords of the fourth and fifth on gelatin-coated glass slides, dehydrated in a gradedlumbar segments, 4TGs and four brainstems were obtained series of alcohols, cleared in xylene and cover-slipped withfrom six male Sprague–Dawley rats (180–250 g). Rats Entellan (Merck).were anesthetized with ether to the level at which respira- The specificities of antisera used in this study have beention was markedly suppressed, and transvascularly per- described elsewhere [2,9,8].fused with 50 ml of saline followed by 500 ml of 4% The experiments were carried out under the control offormaldehyde in 0.1 M phosphate buffer (pH 7.4). The the Animal Research Control Committee in accordanceDRGs and TGs were dissected, immersed in a phosphate- with The Guidelines for Animal Experiments of Okayamabuffered saline containing 20% sucrose overnight, frozen University Medical School, Government Animal Protec-sectioned at 12mm, and thaw-mounted on gelatin-coated tion and Management Law (No. 105), and Japaneseglass slides. Government Notification on Feeding and Safekeeping of
For simultaneous visualization of OC with VR1 or Animals (No. 6). All efforts were made to minimize theVRL-1, a double-immunofluorescence method was used. number of animals used and their suffering.The sections were incubated for 24 h at room temperature As described previously [2,4,9,8,7], the DRG and TGwith a mixture of goat anti-OC serum (1:100, Biomedical contained abundant OC-, VR1- or VRL-1-ir neurons (Fig.Technologies) and either rabbit anti-VR1 serum (1:500, 1A–H). OC-ir neurons were medium-sized to large in theNeuromics) or rabbit anti-VRL-1 serum [2]. The sections DRG (Fig. 1A,C), whereas such neurons were of variouswere then treated with a mixture of fluorescein isothio- sizes in the TG (Figs 1E,G). OC-ir granules were distribut-cyanate-conjugated donkey anti-goat IgG (1:100, Jackson ed within the cytoplasm of large neurons in these ganglia.
Fig. 1. Double immunofluorescent microphotographs for OC-ir (A,C,E,G) with VR1-ir (B,F) or VRL-1-ir (D,H) in the DRG (A–D) and TG (E–H). Panels(A–C) and (D–H) show the same fields of view, respectively. OC-ir neurons are devoid of VR1- and VRL1-ir in the DRG. In the TG, however, manyOC-ir neurons co-express VR1-ir (arrows in E, F). Arrows point to a trigeminal neuron which exhibits both OC- (G) and VRL-1-ir (H). Scale bar550mm(A). All panels are at the same magnification.
H. Ichikawa, T. Sugimoto / Brain Research 958 (2002) 459–462 461
In small and medium-sized TG neurons, strong OC- flocculent OC-ir except for laminae I and II that lacked itimmunofluorescence was occasionally observed throughout (Fig. 2A). The ir of neuropil was weak in other regions ofthe cytoplasm (Fig. 1E,G). the dorsal and ventral horns.
Virtually all OC-ir DRG neurons were devoid of VR1-ir The trigeminal sensory nuclei contained various num-(2/432 or 0.5%, Fig. 1A,B). No OC-ir neurons co-ex- bers of OC-ir cells (Fig. 2B,D–F). In the medullary dorsalpressed VRL-1-ir in the DRG (0/334, Fig. 1C,D). In the horn (MDH), small OC-ir cells were sparsely distributedTG, 14.1% (60/426) of OC-ir neurons contained VR1-ir, (Fig. 2B). On the other hand, the rostral parts of thewhereas 8.2% (60/736) of VR1-ir neurons showed OC-ir. trigeminal sensory nuclei (subnuclei interpolaris and oralis,These neurons had strong OC-immunofluoresence through- and nucleus principalis) contained abundant OC-ir cellsout the cytoplasm (Fig. 1E,F). On the other hand, 1.7% (Fig. 2D–F). OC-ir cell bodies in the subnucleus inter-(5 /294) of OC-ir TG neurons exhibited VRL-1-ir and polaris, the dorsal part of the subnucleus oralis and the1.3% (5/399) of VRL-1-ir TG neurons contained OC-ir. nucleus principalis were small (Fig. 2D–F), while those inSuch neurons had granular OC-ir in the cytoplasm (Fig. the ventral part of subnucleus oralis were large (Fig. 2E).1G,H). The neuropil of the entire trigeminal sensory nuclei was
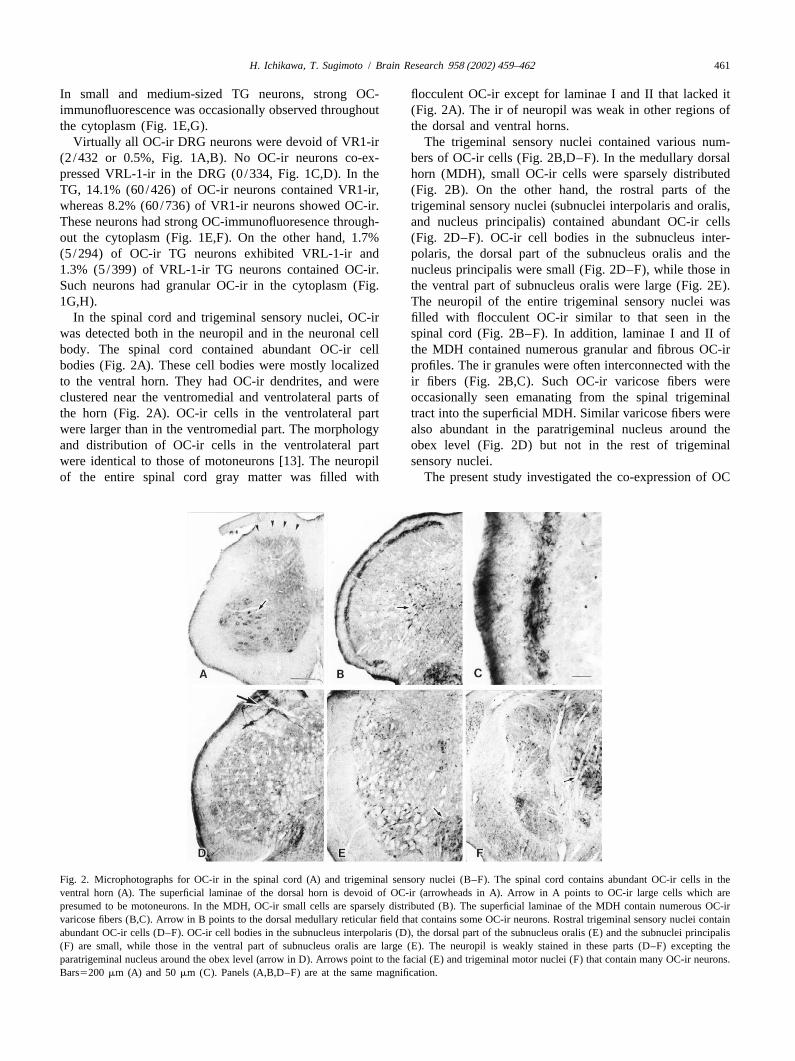
In the spinal cord and trigeminal sensory nuclei, OC-ir filled with flocculent OC-ir similar to that seen in thewas detected both in the neuropil and in the neuronal cell spinal cord (Fig. 2B–F). In addition, laminae I and II ofbody. The spinal cord contained abundant OC-ir cell the MDH contained numerous granular and fibrous OC-irbodies (Fig. 2A). These cell bodies were mostly localized profiles. The ir granules were often interconnected with theto the ventral horn. They had OC-ir dendrites, and were ir fibers (Fig. 2B,C). Such OC-ir varicose fibers wereclustered near the ventromedial and ventrolateral parts of occasionally seen emanating from the spinal trigeminalthe horn (Fig. 2A). OC-ir cells in the ventrolateral part tract into the superficial MDH. Similar varicose fibers werewere larger than in the ventromedial part. The morphology also abundant in the paratrigeminal nucleus around theand distribution of OC-ir cells in the ventrolateral part obex level (Fig. 2D) but not in the rest of trigeminalwere identical to those of motoneurons [13]. The neuropil sensory nuclei.of the entire spinal cord gray matter was filled with The present study investigated the co-expression of OC
Fig. 2. Microphotographs for OC-ir in the spinal cord (A) and trigeminal sensory nuclei (B–F). The spinal cord contains abundant OC-ir cells in theventral horn (A). The superficial laminae of the dorsal horn is devoid of OC-ir (arrowheads in A). Arrow in A points to OC-ir large cells which arepresumed to be motoneurons. In the MDH, OC-ir small cells are sparsely distributed (B). The superficial laminae of the MDH contain numerous OC-irvaricose fibers (B,C). Arrow in B points to the dorsal medullary reticular field that contains some OC-ir neurons. Rostral trigeminal sensory nuclei containabundant OC-ir cells (D–F). OC-ir cell bodies in the subnucleus interpolaris (D), the dorsal part of the subnucleus oralis (E) and the subnuclei principalis(F) are small, while those in the ventral part of subnucleus oralis are large (E). The neuropil is weakly stained in these parts (D–F) excepting theparatrigeminal nucleus around the obex level (arrow in D). Arrows point to the facial (E) and trigeminal motor nuclei (F) that contain many OC-ir neurons.Bars5200 mm (A) and 50mm (C). Panels (A,B,D–F) are at the same magnification.

462 H. Ichikawa, T. Sugimoto / Brain Research 958 (2002) 459–462
with VR1 or VRL-1 in the sensory ganglia. Virtually all R eferencesOC-ir neurons were devoid of these transducers in the
[1] M.J. Caterina, M.A. Schumacher, M. Tominaga, T.A. Rosen, J.D.DRG. The findings may suggest that OC-ir neurons do notLevine, D. Julius, The capsaicin receptor: a heat-activated ioninclude nociceptors in the DRG. This idea can be sup-channel in the pain pathway, Nature 389 (1997) 816–824.ported by the present observation that the laminae I and II
[2] M.J. Caterina, T.A. Rosen, M. Tominaga, A.J. Brake, D. Julius, Aof the spinal dorsal horn were devoid of OC-ir, because the capsaicin-receptor homologue with a high threshold for noxiouslaminae are thought to be the projection site of nociceptive heat, Nature 398 (1999) 436–441.
[3] C. Desbois, K. Gerard, Osteocalcin cluster: implications for func-DRG neurons [2,4,5,17].tional studies, J. Cell Biochem. 57 (1995) 379–383.In the TG, 14% of OC-ir TG neurons were also
[4] A. Guo, L. Vulchanova, J. Wang, X. Li, R. Elde, Immunocytochemi-immunoreactive for VR1. On the other hand, only a small cal localization of the vanilloid receptor 1 (VR1): relationship tonumber of OC-ir neurons showed VRL-1-ir. In sensory neuropeptides, the P2X3 purinoceptor and IB4 binding sites, Eur. J.ganglia, VR1-ir neurons have small to medium-sized cell Neurosci. 11 (1999) 946–958.
[5] D.L. Hammond, M.A. Ruda, Developmental alterations in nocicep-bodies, whereas VRL-1-ir neurons are medium-sized totive threshold, immunoreactive calcitonin gene-related peptide andlarge [2,4,8,7]. Previous studies have indicated that VR1-irsubstance P, and fluoride-resistant acid phosphatase in neonatally
and VRL-1-ir TG neurons have unmyelinated and myeli- capsaicin-treated rats, J. Comp. Neurol. 312 (1991) 436–540.nated axons, respectively [2,4,7]. Therefore, it is likely that [6] P.V. Hauschka, F.H. Wians Jr., Osteocalcin-hydroxyapatite inter-small to medium-sized OC-ir TG neurons include nocicep- action in the extracellular organic matrix of bone, Anat. Rec. 224
(1989) 180–188.tors with unmyelinated axons.[7] H. Ichikawa, T. Sugimoto, VRL-1-immunoreactive primary sensoryIn the present study, numerous OC-ir varicose fibers
neurons in the rat trigeminal nervous system, Neuroscience 101were observed in laminae I and II of the MDH and the (2000) 719–725.paratrigeminal nucleus, the major termination site of TG [8] H. Ichikawa, T. Sugimoto, VR1-immunoreactive primary sensorynociceptors [16]. Because these fibers were occasionally neurons in the rat trigeminal ganglion, Brain Res. 890 (2001)
184–188.seen originating from the spinal trigeminal tract, they are[9] H. Ichikawa, T. Itota, Y. Torii, K. Inoue, T. Sugimoto, Osteocalcin-considered to be at least partly derived from the TG. OC-ir
immunoreactive primary sensory neurons in the rat spinal andnociceptive neurons may project to the superficial laminae trigeminal nervous systems, Brain Res. 838 (1999) 205–209.of the MDH. This suggestion is supported by the previous ¨[10] G. Ju, T. Hokfelt, E. Brodin, J. Fahrenkrug, J.A. Fischer, P. Frey,findings that small to medium-sized TG neurons co-ex- R.P. Elde, J.C. Brown, Primary sensory neurons of the rat showing
calcitonin gene-related peptide immunoreactivity and their relationpress OC- and CGRP-ir and that CGRP-ir TG neuronsto substance P-, somatostatin-, galanin-, vasoactive intestinal poly-project to laminae I and II of the MDH and the parat-peptide- and cholecystokinin-immunoreactive ganglion cells, Cell
rigeminal nucleus [9,16]. It may be deduced that OC-ir TG Tiss. Res. 247 (1987) 417–431.nociceptors have small to medium-sized cell bodies and [11] L. Kruger, J.D. Silverman, P.W. Mantyh, C. Sternini, N.C. Brecha,send their unmyelinated axons to the superficial laminae of Peripheral patterns of calcitonin-gene-related peptide general
somatic sensory innervation: cutaneous and deep terminations, J.the MDH.Comp. Neurol. 280 (1989) 291–302.In conclusion, we have described OC-ir primary
[12] J.B. Lian, C.M. Gundberg, Osteocalcin. Biochemical considerationsnociceptive neurons in the TG. A substantial proportion and clinical applications, Clin. Orthop. 226 (1988) 267–291.(14%) of OC-ir TG neurons were immunoreactive for [13] F. Piehl, U. Arvidsson, H. Johnson, S. Cullheim, A. Dagerlind, B.VR1. Only a small number of neurons co-expressed OC- Ulfhake, Y. Cao, R. Elde, R.F. Pettersson, L. Terenius, GAP-43,
aFGF, CCK and alpha- and beta-CGRP in rat spinal motoneuronsand VRL-ir in the TG. Laminae I and II of the MDHsubjected to axotomy and/or dorsal root severance, Eur. J. Neurosci.contained numerous OC-ir varicose fibers. OC-ir TG5 (1993) 1321–1333.
nociceptors may have small to medium-sized cell bodies, [14] J.D. Silverman, L. Kruger, Calcitonin-gene-related-peptide-immuno-and send their unmyelinated axons to the superficial reactive innervation of the rat head with emphasis on specializedlaminae of the MDH. sensory structures, J. Com. Neurol. 280 (1989) 303–330.
[15] G. Skofitsch, D.M. Jacobowitz, Calcitonin gene-related peptidecoexists with substance P in capsaicin sensitive neurons and sensoryganglia of the rat, Peptides 6 (1985) 747–754.
A cknowledgements [16] T. Sugimoto, Y. Fujiyoshi, C. Xiao, Y.F. He, H. Ichikawa, Centralprojection of calcitonin gene-related peptide (CGRP)- and substance
We greatly appreciate Dr. David Julius at the Depart- P (SP)-immunoreactive trigeminal primary neurons in the rat, J.Comp. Neurol. 378 (1997) 425–442.ment of Cellular and Molecular Pharmacology, University
[17] R.J. Traub, B. Allen, E. Humphrey, M.A. Ruda, Analysis ofof California for his kind gift of the anti-VRL-1-serum.calcitonin gene-related peptide-like immunoreactivity in the cat
This work was supported by a grant from the Japanese dorsal spinal cord and dorsal root ganglia provide evidence for aMinistry of Education, Culture, Sports, Science and Tech- multisegmental projection of nociceptive C-fiber primary afferents,nology to H.I. (No. 14571733). J. Comp. Neurol. 302 (1990) 562–574.