Embed Size (px)
Citation preview

RESEARCH ARTICLE
The effect of gap junction-mediated transfer of miR-200bon osteogenesis and angiogenesis in a co-culture of MSCsand HUVECsXiaoting Fan, Yi Teng, Zhaoyang Ye, Yan Zhou* and Wen-Song Tan
ABSTRACTFor successful engineering of pre-vascularized bone tissue in vitro,understanding the interactions between vasculogenic cells and bone-forming cells is a prerequisite. Mounting evidence indicates thatmicroRNAs can serve as intercellular signals that allow cell–cellcommunication. Here, the role of the transfer of the microRNA miR-200b between vasculogenic and osteogenic cells was explored in aco-culture system. Rat bone-marrow derived mesenchymal stemcells (BMSCs) formed functional gap junctions composed ofconnexin 43 (Cx43, also known as GJA1) with human umbilicalvein endothelial cells (HUVECs), through which miR-200b couldtransfer from BMSCs to HUVECs to regulate osteogenesis andangiogenesis. As a negative regulator, the decrease in miR-200blevel in BMSCs derepressed the expression of VEGF-A, leading toincreased osteogenic differentiation. Once inside HUVECs, miR-200b reduced the angiogenic potential of HUVECs throughdownregulation of ZEB2, ETS1, KDR and GATA2. Additionally,TGF-β was found to trigger the transfer of miR-200b to HUVECs.Upon adding the TGF-β inhibitor SB431542 or TGF-β-neutralizingantibody, the formation of capillary-like structures in co-culture couldbe partially rescued. These findings may be fundamental to thedevelopment of a cell-based bone regeneration strategy.
KEY WORDS: Pre-vascularized bone, Mesenchymal stem cell,Endothelial cell, Gap junction, miR-200b
INTRODUCTIONCurrently, cell-based strategies to vascularize engineered bonetissue in vitro are arousing great interest for bone regeneration. It isanticipated that the vascular network in grafts could be improvedthrough co-culture of osteogenic cells and vasculogenic cells, andthat this network would amalgamate with the host vasculature andpromote survival and regeneration of bone tissue (Tsigkou et al.,2010). However, the problem of insufficient vascularization innewly formed bone tissue still exists. In vivo, bone development andremodeling depend on complex interactions between bone-formingcells and other cells present within the bone microenvironment,particularly endothelial cells (ECs), which might be one of thepivotal cell types involved in a complex interactive communicationnetwork in bone. For successful engineering of pre-vascularizedbone tissue in vitro, understanding the interactions between
vasculogenic cells (endothelial cells or endothelial progenitors)and osteogenic cells [osteoblasts or mesenchymal stem cells(MSCs)] is a prerequisite. However, the cellular and molecularbases of osteogenic–endothelial cell interactions, as wellas their impact on endothelial vascularization and osteogenicdifferentiation, are not fully understood.
The communication between vasculogenic cells and osteogeniccells may not only deploy diffusible soluble paracrine factors, butalso involve the junctional communication through the formation ofa multicellular network (Villars et al., 2000, 2002; Guillotin et al.,2004). Moreover, the secretion of soluble factors, such as VEGF,has been reported to rely on cell–cell contact via gap junctions(Grellier et al., 2009). Gap junctions are specialized intercellularchannels formed by membrane proteins called connexins, whichcan mediate intracellular exchange of regulatory ions, smallmolecules (maximum molecular mass of 1.5 kDa) (Moorer andStains, 2017), and siRNAs and microRNAs (miRNAs) (minordiameters of 1.0 nm) between adjacent cells (Valiunas et al., 2005;Inose et al., 2009). Connexin 43 (Cx43; also known as GJA1), apredominant gap junction protein in osteoblasts, has beendemonstrated to form gap junctions between ECs and osteogeniccells in co-culture in several studies. The functional inhibition ofgap junctions by means of treatment with 18α-glycyrrhetinic acid(18GA) or through inhibiting Cx43 synthesis with antisenseoligodeoxyribonucleotide can attenuate the effect of humanumbilical vein endothelial cells (HUVECs) on osteogenicdifferentiation in co-culture (Villars et al., 2002). In anotherstudy, gap junctions between apposed HUVECs and humanosteoprogenitor cells were shown to form from Cx43 after a short-term co-culture (48 h), and synthesis of Cx43 was required for thestimulated alkaline phosphatase (ALPase) activity in this co-culturesystem (Guillotin et al., 2008). Moreover, Herzog et al. revealed thatin co-culture of outgrowth endothelial cells (OECs) and osteoblastswith direct cell–cell contacts, both cellular organization of OECsinto vascular structures and osteogenic differentiation of osteoblastswere promoted via Cx43-based gap junctions (Herzog et al., 2014).
miRNAs are a group of non-coding endogenous RNAs that are∼21–25 nucleotides long, and usually function as negative regulatorsof gene expression through complementary binding to their targetmRNAs. Numerous miRNAs have been identified to play criticalregulatory roles in biological and pathological processes, such as celldifferentiation, angiogenesis and cancer. For instance, miRNAs arecritical to stem cell function, particularly in the maintenance ofstemness or the lineage-specific differentiation (Kim, 2005; Lu et al.,2005). In particular, miRNAs are able to regulate the function of ECsduring angiogenesis (Akhtar et al., 2015; Climent et al., 2015).Furthermore, several miRNAs have already been confirmed toregulate osteogenesis and angiogenesis simultaneously. Inhibition ofmiR-222 expression was shown to accelerate bone healing byReceived 29 January 2018; Accepted 4 June 2018
State Key Laboratory of Bioreactor Engineering, East China University of Scienceand Technology, Shanghai 200237, P. R. China.
*Author for correspondence ([email protected])
X.F., 0000-0002-0928-6207; Y.Z., 0000-0001-7865-3413
1
© 2018. Published by The Company of Biologists Ltd | Journal of Cell Science (2018) 131, jcs216135. doi:10.1242/jcs.216135
Journal
ofCe
llScience

stimulating both osteogenesis and angiogenesis (Yoshizuka et al.,2016). Bone regeneration has been to shown to be promoted throughmiR-26a mediating positive regulation of angiogenic-osteogeniccoupling (Li et al., 2013). Yang et al. reported that the miR-497–195cluster regulates the angiogenesis coupled with osteogenesisthrough maintaining endothelial Notch activity and HIF-1αstability (Yang et al., 2017).miR-200b, which belongs to the miR-200 family, is expressed in
a variety of cells and regulates key cellular functions, such as cellproliferation, motility, apoptosis, stem cell properties, angiogenesisand epithelial–mesenchymal transition (EMT) (Brabletz andBrabletz, 2010). As an anti-angiogenic miRNA, miR-200b targetsv-Ets erythroblastosis virus E26 oncogene homolog 1 (ETS1),thereby inhibiting the angiogenic response of human microvascularendothelial cells (HMECs). When transfected with miR-200b, theability of HMECs to migrate and to form tubes on Matrigel isinhibited (Chan et al., 2011). As an angiogenic inhibitor, bynegatively regulating VEGF signaling, via targeting VEGF and itsreceptor KDR, its therapeutic potential in cancer has also beenimplicated (Choi et al., 2011). It has been reported that the TGF-β/ZEB/miR-200b signaling network creates a double-negativefeedback loop that plays an essential role in the initiation of EMTand cancer metastasis (Xiong et al., 2012; Shen et al., 2015).Moreover, miR-141 and miR-200b could inhibit BMP-2-inducedpre-osteoblast differentiation through translational repression ofDlx5 (Vimalraj and Selvamurugan, 2013).Mounting evidence indicates that miRNAs can be considered as
intercellular signals that allow cells to communicate. Cells canrelease miRNAs to specifically modulate physiological processes inrecipient cells via exosomes (Montecalvo et al., 2012; Baglio et al.,2015) or gap junctions (Calderón and Retamal, 2016; Thuringeret al., 2016; Zong et al., 2016). In the present study, the objectivewas to gain an insight into the molecular mechanisms in cellcoupling and reciprocal interactions between ECs and MSCs. In adirect co-culture between bone marrow-derived MSCs (BMSCs)and HUVECs under osteogenic induction conditions, we found thatosteogenesis was significantly stimulated while angiogenesis wascompromised. It was therefore hypothesized that miR-200b mightfunction as a regulator in this process. Results demonstrated thatBMSCs communicated with HUVECs via miR-200b, which couldtransfer from BMSCs to HUVECs through gap junctions formed ofCx43. In particular, TGF-βwas found to trigger the transfer of miR-200b to HUVECs. As a negative regulator, the decrease of miR-200b in BMSCs induced upregulation of VEGF-A, which waspartly responsible for the stimulatory effects on osteogenicdifferentiation. Once inside HUVECs, miR-200b reduced theangiogenic potential of HUVECs by downregulating of ZEB2,ETS1, KDR and GATA2. Treatment with the TGF-β inhibitorSB431542 or TGF-β-neutralizing antibody in co-culture causedthe formation of capillary-like structures to be partially rescued.These findings could be fundamental to the engineering ofpre-vascularized bone tissue in vitro.
RESULTSCharacterization of rat BMSCsRat BMSCs at passage 3 were tagged with conjugated antibodiesagainst CD34, CD45 (also known as PTPRC), CD44, CD73(NT5E), CD105 (ENG), CD106 (VCAM1), CD29 (integrin β1,encoded by ITGB1) and CD90 (THY1). Flow cytometric analysisindicated that cells were strongly positive for the MSC markersCD29 and CD90 (≥99%), positive for CD44, weakly positive forCD106 and CD73, and negative for the hematopoietic stem cell
markers CD45, CD34 and CD105 (Fig. S1A). Results showed thatafter subculture, hematopoietic cells were eliminated from bonemarrow-derived cells, and BMSCs used in this study were ofhigh purity.
Rat BMSCs at passage 3 were induced for osteo-, adipo- andchondro-genesis for 21 days to assess their capacity for multi-lineage differentiation [with osteogenic induction medium (OIM),adipogenic induction medium (AIM) or chondrogenic medium; seeMaterials and Methods]. After induction, the differentiated cellswere stained with the indicated dyes (Fig. S1B). After osteogenicinduction, BMSCs were able to produce mineral nodules asevidenced by strong intensities of Alizarin Red S staining. Afteradipogenic induction, lipid vacuoles were seen within the cells uponOil Red O staining. For analyzing chondrogenic differentiation, weused Safranin O staining, which detects glycosaminoglycan (GAG),a characteristic extracellular matrix synthesized by chondrocytes.These cell differentiation assays demonstrated that BMSCs hadmulti-lineage differentiation potential.
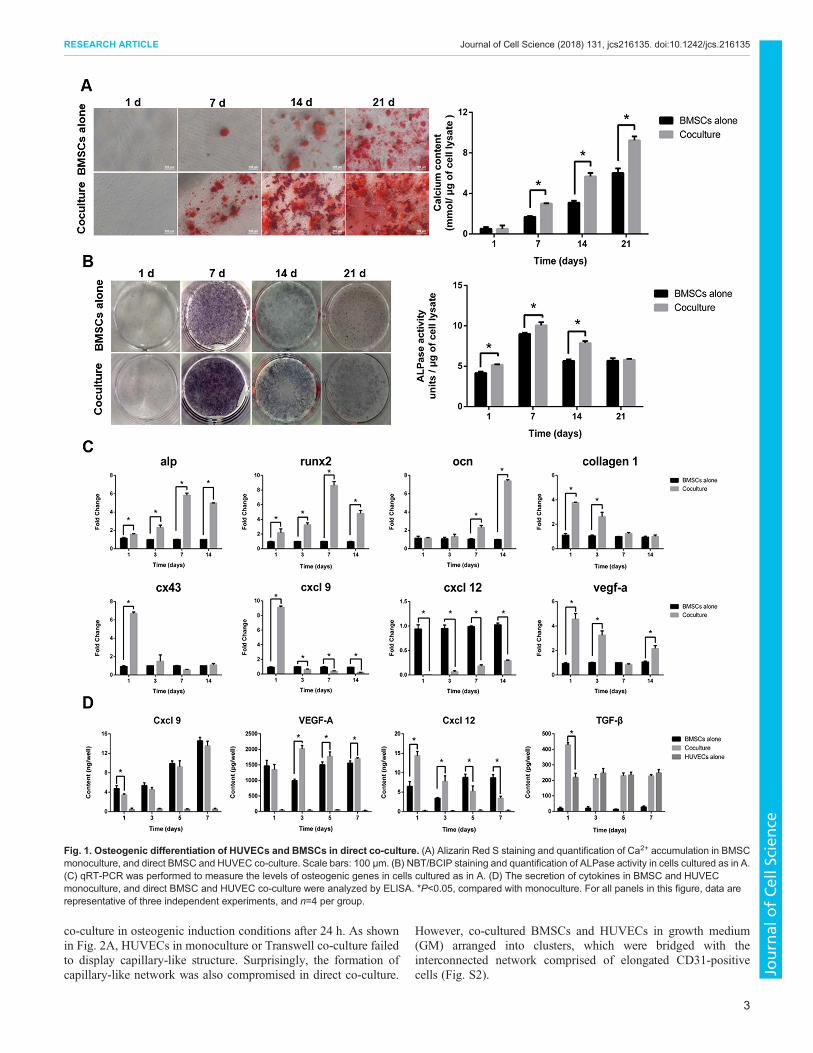
Osteogenic differentiation in co-culture of HUVECs andBMSCsAs shown in Fig. 1A, co-culture of HUVECs and BMSCs withdirect cell–cell contact in osteogenic induction conditions induced asignificant amount of Ca2+ accumulation within 14 days of co-culture as compared to what was seen with BMSC monoculture, asshown by the stronger intensities of Alizarin Red S staining andgreater number of mineralized nodules. The activity of ALPasewithin 14 days of osteogenic induction, as detected by Nitro Bluetetrazolium and 5-bromo-4-chloro-3-indolyl phosphate (NBT/BCIP) staining showed a noticeable difference between co-cultureand monoculture (Fig. 1B). More intensive NBT/BCIP staining andhigher ALPase activity was observed in co-culture.
qRT-RCR analysis confirmed that the osteoblastic phenotypemarkers genes, such as Alp, Runx2, Ocn (also known as Bglap) andcollagen 1 (also known as Col1a1), were greatly upregulated in directco-culture compared with the level in BMSC monoculture (Fig. 1C).Importantly, the level of Cx43, a predominant gap junctionalmolecule, was significantly increased by 6.82-fold at the early timepoint in co-culture (1 day). Chemokine (C-X-C motif) ligand 9(Cxcl9) was also enhanced by 9.23-fold on the first day of co-culture.The level of another critical chemokine for cell migration and celldifferentiation, Cxcl12, in co-culture was lower than that in BMSCmonoculturewithin 14 days. Direct cell contacts between BMSCs andHUVECs stimulated the expression of Vegf-a by 4.43-fold on day 1 inco-cultured BMSCs compared with monoculture.
The amount of cytokine secreted in culture medium was alsoquantified by using ELISA. As shown in Fig. 1D, more Cxcl9,VEGF-A and Cxcl12 was secreted by BMSCs, and more TGF-βwas produced by HUVECs.Within 7 days, the secretion of Cxcl9 inco-culture was almost equivalent to the amount in BMSCmonoculture. The content of VEGF-A was significantly enhancedin co-culture compared with that in monoculture. Co-culture led to ahigher level of Cxcl12 production on day 1 and 3, but a lower levelon day 5 and 7 compared with monoculture. TGF-β reached thepeak value at the early time point (1 day) in co-culture, which wassignificantly higher than that in HUVEC monoculture.
Angiogenesis in co-cultures of HUVECs and BMSCsTo observe the formation of capillary-like structures,immunofluorescence staining was performed to detect theexpression of CD31 in HUVECs in monolayers, co-culture inTranswell inserts (hereafter called Transwell co-culture) and direct
2
RESEARCH ARTICLE Journal of Cell Science (2018) 131, jcs216135. doi:10.1242/jcs.216135
Journal
ofCe
llScience
co-culture in osteogenic induction conditions after 24 h. As shownin Fig. 2A, HUVECs in monoculture or Transwell co-culture failedto display capillary-like structure. Surprisingly, the formation ofcapillary-like network was also compromised in direct co-culture.
However, co-cultured BMSCs and HUVECs in growth medium(GM) arranged into clusters, which were bridged with theinterconnected network comprised of elongated CD31-positivecells (Fig. S2).
Fig. 1. Osteogenic differentiation of HUVECs and BMSCs in direct co-culture. (A) Alizarin Red S staining and quantification of Ca2+ accumulation in BMSCmonoculture, and direct BMSC and HUVEC co-culture. Scale bars: 100 μm. (B) NBT/BCIP staining and quantification of ALPase activity in cells cultured as in A.(C) qRT-PCR was performed to measure the levels of osteogenic genes in cells cultured as in A. (D) The secretion of cytokines in BMSC and HUVECmonoculture, and direct BMSC and HUVEC co-culture were analyzed by ELISA. *P<0.05, compared with monoculture. For all panels in this figure, data arerepresentative of three independent experiments, and n=4 per group.
3
RESEARCH ARTICLE Journal of Cell Science (2018) 131, jcs216135. doi:10.1242/jcs.216135
Journal
ofCe
llScience
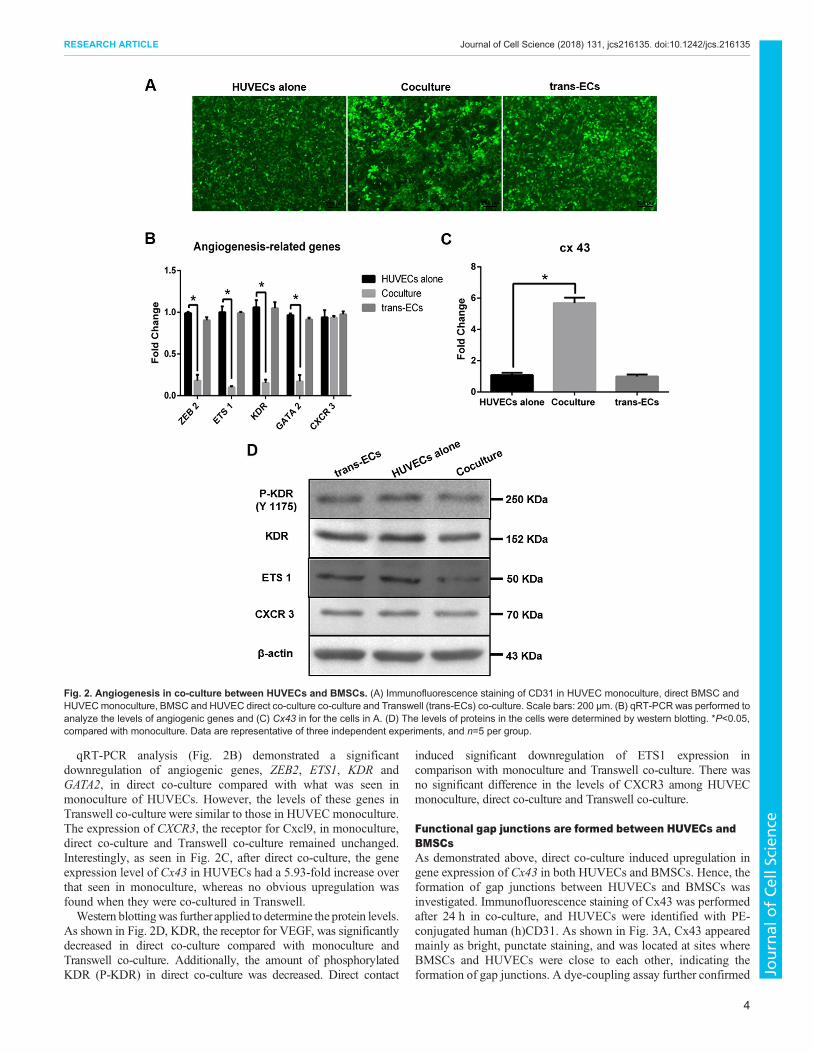
qRT-PCR analysis (Fig. 2B) demonstrated a significantdownregulation of angiogenic genes, ZEB2, ETS1, KDR andGATA2, in direct co-culture compared with what was seen inmonoculture of HUVECs. However, the levels of these genes inTranswell co-culture were similar to those in HUVEC monoculture.The expression of CXCR3, the receptor for Cxcl9, in monoculture,direct co-culture and Transwell co-culture remained unchanged.Interestingly, as seen in Fig. 2C, after direct co-culture, the geneexpression level of Cx43 in HUVECs had a 5.93-fold increase overthat seen in monoculture, whereas no obvious upregulation wasfound when they were co-cultured in Transwell.Western blottingwas further applied to determine the protein levels.
As shown in Fig. 2D, KDR, the receptor for VEGF, was significantlydecreased in direct co-culture compared with monoculture andTranswell co-culture. Additionally, the amount of phosphorylatedKDR (P-KDR) in direct co-culture was decreased. Direct contact
induced significant downregulation of ETS1 expression incomparison with monoculture and Transwell co-culture. There wasno significant difference in the levels of CXCR3 among HUVECmonoculture, direct co-culture and Transwell co-culture.
Functional gap junctions are formed between HUVECs andBMSCsAs demonstrated above, direct co-culture induced upregulation ingene expression of Cx43 in both HUVECs and BMSCs. Hence, theformation of gap junctions between HUVECs and BMSCs wasinvestigated. Immunofluorescence staining of Cx43 was performedafter 24 h in co-culture, and HUVECs were identified with PE-conjugated human (h)CD31. As shown in Fig. 3A, Cx43 appearedmainly as bright, punctate staining, and was located at sites whereBMSCs and HUVECs were close to each other, indicating theformation of gap junctions. A dye-coupling assay further confirmed
Fig. 2. Angiogenesis in co-culture between HUVECs and BMSCs. (A) Immunofluorescence staining of CD31 in HUVEC monoculture, direct BMSC andHUVECmonoculture, BMSC and HUVEC direct co-culture co-culture and Transwell (trans-ECs) co-culture. Scale bars: 200 μm. (B) qRT-PCR was performed toanalyze the levels of angiogenic genes and (C) Cx43 in for the cells in A. (D) The levels of proteins in the cells were determined by western blotting. *P<0.05,compared with monoculture. Data are representative of three independent experiments, and n=5 per group.
4
RESEARCH ARTICLE Journal of Cell Science (2018) 131, jcs216135. doi:10.1242/jcs.216135
Journal
ofCe
llScience
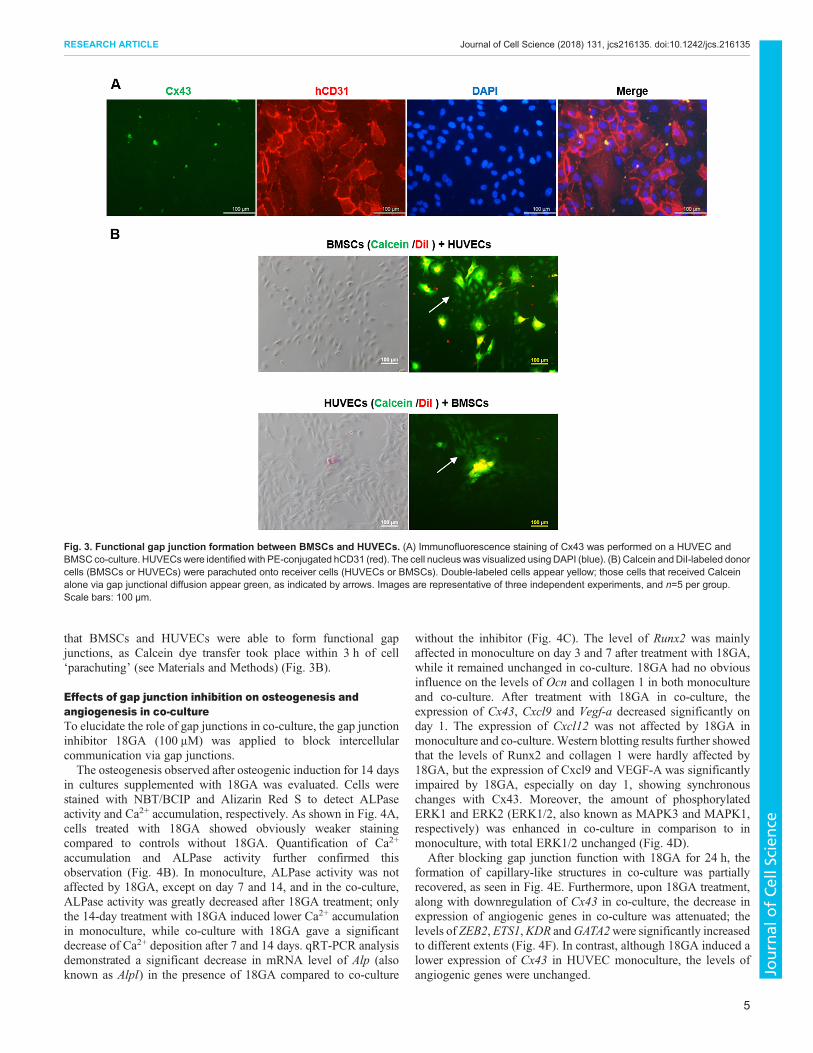
that BMSCs and HUVECs were able to form functional gapjunctions, as Calcein dye transfer took place within 3 h of cell‘parachuting’ (see Materials and Methods) (Fig. 3B).
Effects of gap junction inhibition on osteogenesis andangiogenesis in co-cultureTo elucidate the role of gap junctions in co-culture, the gap junctioninhibitor 18GA (100 μM) was applied to block intercellularcommunication via gap junctions.The osteogenesis observed after osteogenic induction for 14 days
in cultures supplemented with 18GA was evaluated. Cells werestained with NBT/BCIP and Alizarin Red S to detect ALPaseactivity and Ca2+ accumulation, respectively. As shown in Fig. 4A,cells treated with 18GA showed obviously weaker stainingcompared to controls without 18GA. Quantification of Ca2+
accumulation and ALPase activity further confirmed thisobservation (Fig. 4B). In monoculture, ALPase activity was notaffected by 18GA, except on day 7 and 14, and in the co-culture,ALPase activity was greatly decreased after 18GA treatment; onlythe 14-day treatment with 18GA induced lower Ca2+ accumulationin monoculture, while co-culture with 18GA gave a significantdecrease of Ca2+ deposition after 7 and 14 days. qRT-PCR analysisdemonstrated a significant decrease in mRNA level of Alp (alsoknown as Alpl) in the presence of 18GA compared to co-culture
without the inhibitor (Fig. 4C). The level of Runx2 was mainlyaffected in monoculture on day 3 and 7 after treatment with 18GA,while it remained unchanged in co-culture. 18GA had no obviousinfluence on the levels of Ocn and collagen 1 in both monocultureand co-culture. After treatment with 18GA in co-culture, theexpression of Cx43, Cxcl9 and Vegf-a decreased significantly onday 1. The expression of Cxcl12 was not affected by 18GA inmonoculture and co-culture.Western blotting results further showedthat the levels of Runx2 and collagen 1 were hardly affected by18GA, but the expression of Cxcl9 and VEGF-A was significantlyimpaired by 18GA, especially on day 1, showing synchronouschanges with Cx43. Moreover, the amount of phosphorylatedERK1 and ERK2 (ERK1/2, also known as MAPK3 and MAPK1,respectively) was enhanced in co-culture in comparison to inmonoculture, with total ERK1/2 unchanged (Fig. 4D).
After blocking gap junction function with 18GA for 24 h, theformation of capillary-like structures in co-culture was partiallyrecovered, as seen in Fig. 4E. Furthermore, upon 18GA treatment,along with downregulation of Cx43 in co-culture, the decrease inexpression of angiogenic genes in co-culture was attenuated; thelevels of ZEB2,ETS1,KDR andGATA2were significantly increasedto different extents (Fig. 4F). In contrast, although 18GA induced alower expression of Cx43 in HUVEC monoculture, the levels ofangiogenic genes were unchanged.
Fig. 3. Functional gap junction formation between BMSCs and HUVECs. (A) Immunofluorescence staining of Cx43 was performed on a HUVEC andBMSC co-culture. HUVECswere identified with PE-conjugated hCD31 (red). The cell nucleuswas visualized usingDAPI (blue). (B) Calcein andDiI-labeled donorcells (BMSCs or HUVECs) were parachuted onto receiver cells (HUVECs or BMSCs). Double-labeled cells appear yellow; those cells that received Calceinalone via gap junctional diffusion appear green, as indicated by arrows. Images are representative of three independent experiments, and n=5 per group.Scale bars: 100 μm.
5
RESEARCH ARTICLE Journal of Cell Science (2018) 131, jcs216135. doi:10.1242/jcs.216135
Journal
ofCe
llScience
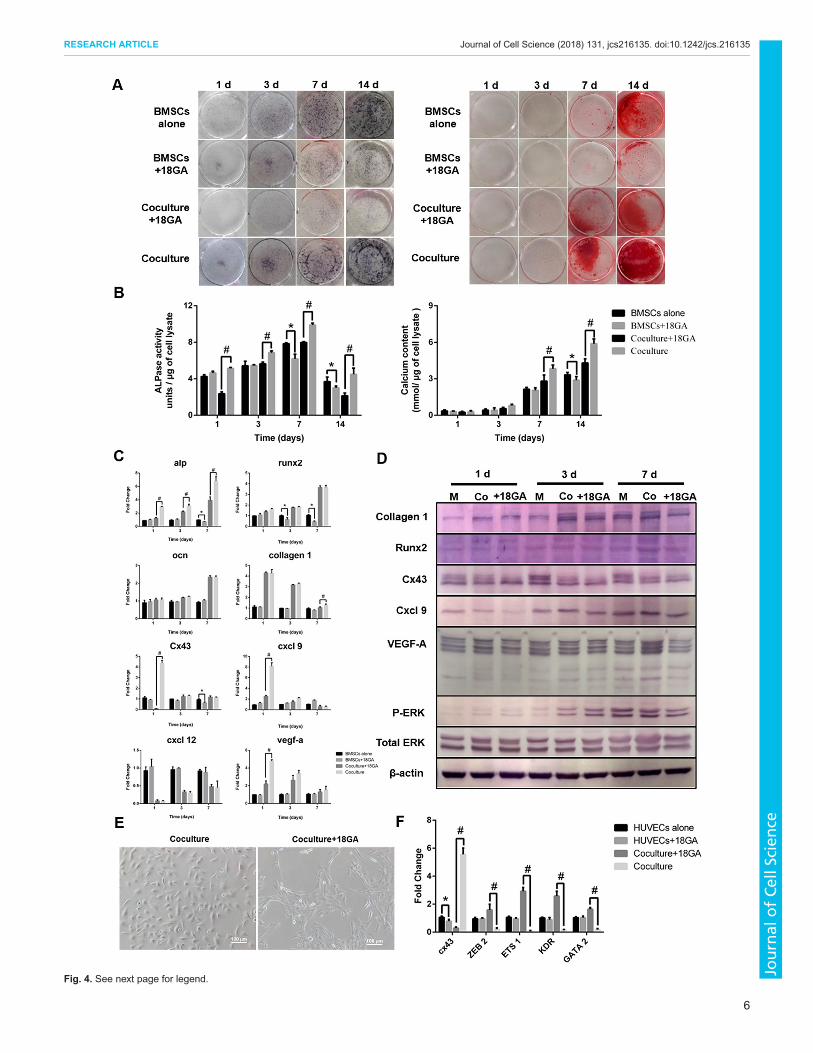
Fig. 4. See next page for legend.
6
RESEARCH ARTICLE Journal of Cell Science (2018) 131, jcs216135. doi:10.1242/jcs.216135
Journal
ofCe
llScience

Effects of gap junction activation on osteogenesis inco-cultureParathyroid hormone (PTH) was used to stimulate gap junctionformation by increasing the expression level of Cx43. A BMSCmonoculture and co-culturewere treated with PTH (10 nM), with noPTH treatment serving as control. During a 14-day induction, cellswere stained with NBT/BCIP and Alizarin Red S to detect ALPaseactivity and Ca2+ accumulation, respectively (Fig. S3A). Cellstreated with PTH showed obviously stronger Alizarin Red S stainingcompared to controls. As shown in Fig. S3B, a quantitativemeasurement indicated that ALPase activity was not affected inmonoculture by PTH, but was significantly increased in co-cultureon day 1. Treatment with PTH induced greater Ca2+ accumulation inmonoculture after 14 days and in co-culture after treatment forboth 7 and 14 days. qRT-PCR analysis demonstrated that asignificant increase in Alp mRNA level was observed in thepresence of PTH in co-culture compared to control on day 1, that thelevel of Runx2was altered in monoculture after treatment with PTH,but not in co-culture, and the levels of Ocn and collagen 1 werehardly affected by PTH in both monoculture and co-culture (Fig.S3C). After treatment with PTH in co-culture, the expression ofCx43 increased significantly on day 1, and Cxcl9 and Vegf-a werechanged accordingly, showing the close correlation between gapjunction formation and the levels of these mRNAs. By contrast,the level of Cxcl12 was hardly altered by PTH in monocultureand co-culture.
Effects of knocking down Cx43 on the expression of VEGFand Cxcl9 in co-cultureTo further investigate the role of gap junction in regulating Cxcl9and VEGF, siRNAs, named 3646 and 3710, respectively (seeMaterials and Methods), were used to specifically knockdownCx43 in BMSCs. As shown in Fig. S4A, after siRNA treatment for36 h, BMSCs showed a significant reduction in the Cx43 mRNA,level by 80% and 70% with siRNA 3646 and 3710, respectively,compared with control. When siRNA-transfected BMSCs weredirectly co-cultured with HUVECs in osteogenic inductionconditions for 24 h, the level of Cx43 was decreased by 50%,whereas the level of Cx43 in non-transfected BMSCs and HUVECsco-culture was increased by 2.8-fold. Semi-quantitative PCR furtherdemonstrated that siRNAs 3646 and 3710 could effectivelyknockdown Cx43 in BMSCs. However, the decrease of Cx43 inBMSCs did not directly affect the levels of Cxcl9 and Vegf-a. Afterco-culturing siRNA-transfected BMSCs with HUVECs, the levelsof Cxcl9 and Vegf-a were obviously lower than that in non-transfected co-cultures (Fig. S4B). The protein levels of Cx43,Cxcl9 and VEGF-A were also measured through western blotting(Fig. S4C). After siRNA transfection for 48 h, a significant
reduction of Cx43 was noticed in BMSCs, whereas the levels ofVEGF-A and Cxcl9 remained unchanged. For co-culture betweensiRNA-transfected BMSCs and HUVECs, the levels of Cxcl9 andVEGF-A were slightly lower than those in co-culture with non-transfected cells. These data indicate that Cxcl9 and VEGF are notdirectly regulated by Cx43 in BMSCs. Instead, moleculescommunicated through Cx43-based gap junctions betweenBMSCs and HUVECs might be responsible for regulating Cxcl9and VEGF.
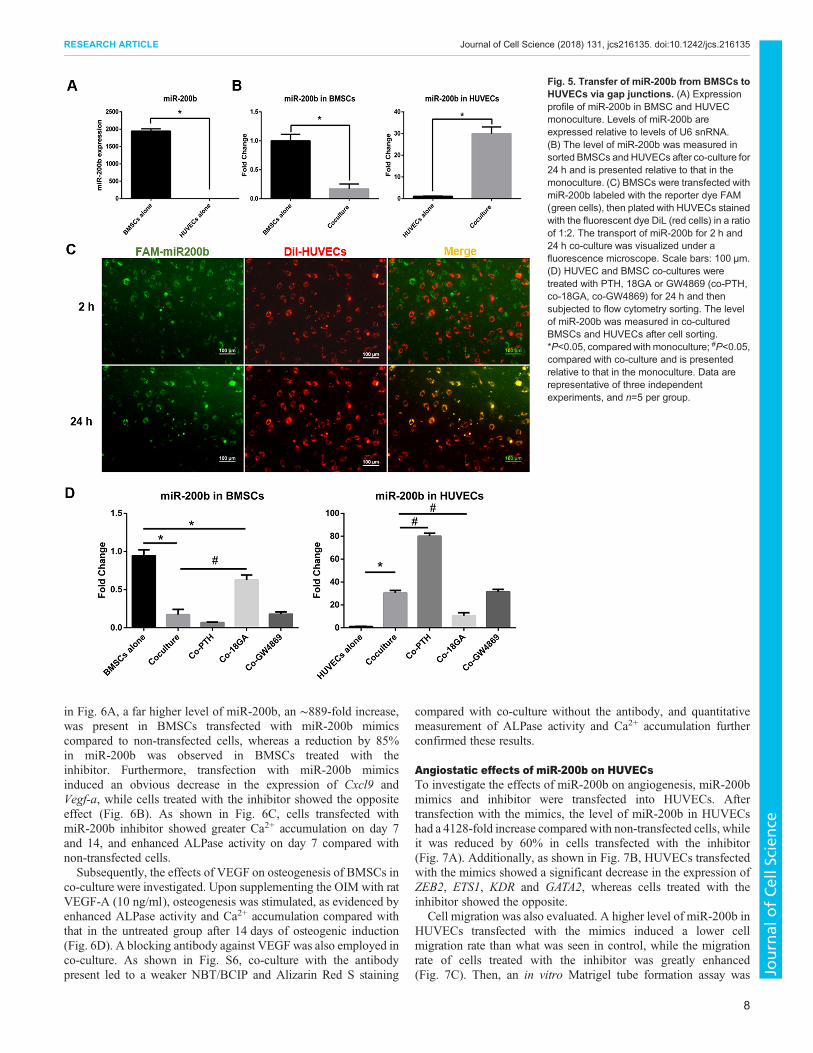
Transfer of miR-200b from BMSCs to HUVECs via gapjunctionsmiRNAs can regulate both osteogenesis and angiogenesis, andmiR-200b is an anti-angiogenic regulator (Pecot et al., 2013). Wetherefore hypothesized that miR-200b could move through gapjunctions between BMSCs and HUVECs to function as a regulatorin co-culture.
As shown in Fig. 5A, the level of miR-200b in BMSCmonoculture was significantly higher than that in HUVECmonoculture. After BMSCs and HUVECs were co-cultured for24 h, cells were sorted by flow cytometry after staining withphycoerythrin (PE)-conjugated anti-hCD31 antibody, with PE-positive cells being HUVECs, and PE-negative cells being BMSCs.The ratio between BMSCs and HUVECs after sorting was almostthe same as the original seeding ratio (2:1) (Fig. S5A). After co-culture, the level of miR-200b was significantly decreased by 88%in co-cultured BMSCs, whereas miR-200b had increased by 30-foldin co-cultured HUVECs (Fig. 5B).
To ascertain whether the transfer of miR-200b between cellsactually happened, BMSCs transfected with FAM-labeled miR-200b were cocultured with Dil-stained HUVECs. After co-culturefor 2 h, the green fluorescence in BMSCs was scattered outsideHUVECs, which indicated miR-200b was still inside BMSCs(Fig. 5C). However, after 24 h, most of the green spots began toenter HUVECs, overlapping with the red cell membrane (Fig. 5C).and appearing yellow in HUVECs.
To further determine whether the transfer of miR-200b wasmediated by gap junctions or secreted exosomes, the gap junctioninhibitor 18GA and activator PTH, and the exosome inhibitorGW4869 were applied to the direct co-culture for 24 h. Co-culturedcells were then subjected to flow cytometry sorting, and the level ofmiR-200b was measured by qRT-PCR. As shown in Fig. 5D, whenco-culture was carried out in the presence of 18GA, the decrease ofmiR-200b initially observed in co-cultured BMSCs was reversedto some extent, and the increase in co-cultured HUVECswas diminished. In contrast, PTH aggravated the decrease ofmiR-200b in co-cultured BMSCs and enhanced the increase inco-cultured HUVECs. However, the exosome inhibitor GW4869did not significantly affect the levels of miR-200b in BMSCsand HUVECs in co-culture. Transwell co-culture was alsoperformed for 24 h with or without GW4869. However, the levelof miR-200b was not affected by the presence of GW4869 (Fig.S5B). Moreover, with GW4869 treatment, the ALPase activity inBMSCs monoculture and co-culture was unaffected, and GW4869only slightly affected Ca2+ accumulation in the later stage (14 days)(Fig. S5C and D). These results indicate that the transfer of miR-200b from BMSCs to HUVECs takes place via gap junctions indirect co-culture.
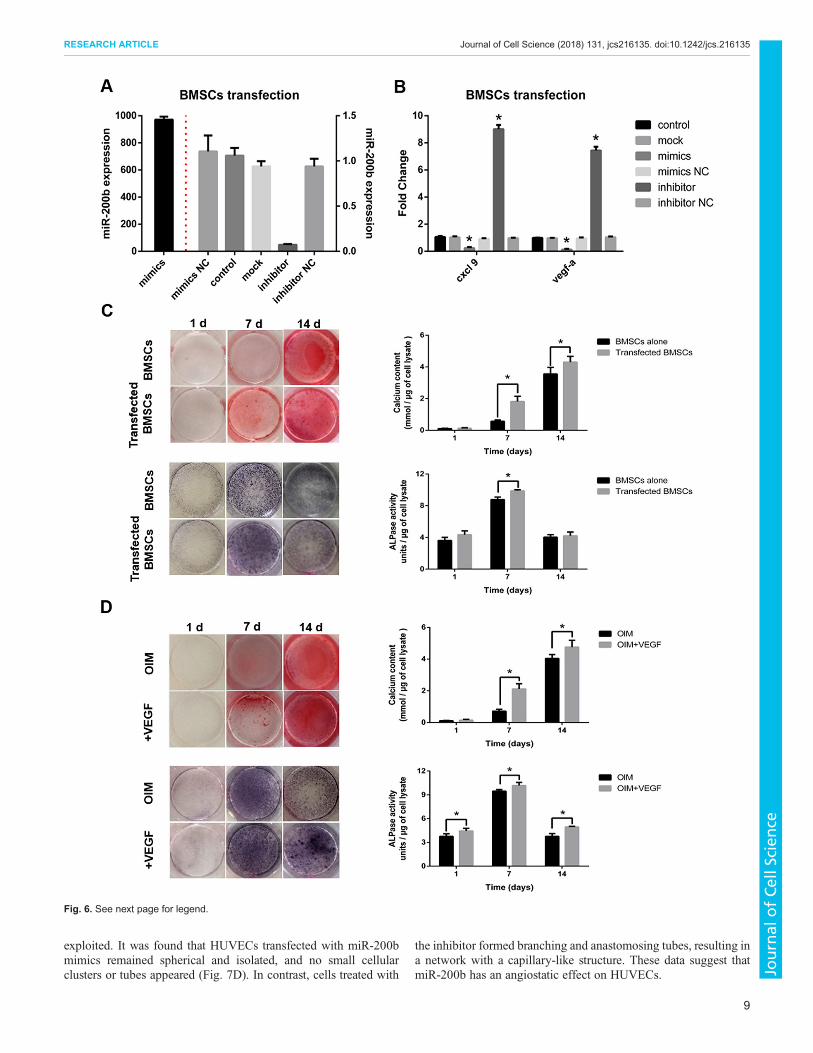
Regulation of osteogenesis of BMSCs by miR-200b via VEGFTo confirm the role of miR-200b on osteogenesis, BMSCswere treated with either miR-200b mimics or inhibitor. As shown
Fig. 4. Effects of treatment with gap junction inhibitor in co-culture.(A) After treatment with the gap junction blocker, 18GA, BMSCs inmonoculture, and HUVEC and BMSC co-cultures were stained with NBT/BCIPand Alizarin Red S. (B) Quantification of ALPase activity and Ca2+
accumulation. (C) qRT-PCRwas performed to analyze the levels of osteogenicgenes in BMSC monoculture, and HUVEC and BMSC co-culture after18GA treatment. (D) Western blotting was used to measure the expressionof proteins in monoculture and co-culture after 18GA treatment.(E) Representative images of co-cultures treated with or without 18GA for 24 h.Scale bars: 100 μm. (F) qRT-PCR was performed to measure the levels ofCx43 and angiogenic genes in monocultures and co-cultures after treatmentwith or without 18GA. *P<0.05, compared with monoculture; #P<0.05,compared with co-culture. For all panels in this figure, data are representativefor three independent experiments, and n=5 per group.
7
RESEARCH ARTICLE Journal of Cell Science (2018) 131, jcs216135. doi:10.1242/jcs.216135
Journal
ofCe
llScience
in Fig. 6A, a far higher level of miR-200b, an ∼889-fold increase,was present in BMSCs transfected with miR-200b mimicscompared to non-transfected cells, whereas a reduction by 85%in miR-200b was observed in BMSCs treated with theinhibitor. Furthermore, transfection with miR-200b mimicsinduced an obvious decrease in the expression of Cxcl9 andVegf-a, while cells treated with the inhibitor showed the oppositeeffect (Fig. 6B). As shown in Fig. 6C, cells transfected withmiR-200b inhibitor showed greater Ca2+ accumulation on day 7and 14, and enhanced ALPase activity on day 7 compared withnon-transfected cells.Subsequently, the effects of VEGF on osteogenesis of BMSCs in
co-culture were investigated. Upon supplementing the OIM with ratVEGF-A (10 ng/ml), osteogenesis was stimulated, as evidenced byenhanced ALPase activity and Ca2+ accumulation compared withthat in the untreated group after 14 days of osteogenic induction(Fig. 6D). A blocking antibody against VEGF was also employed inco-culture. As shown in Fig. S6, co-culture with the antibodypresent led to a weaker NBT/BCIP and Alizarin Red S staining
compared with co-culture without the antibody, and quantitativemeasurement of ALPase activity and Ca2+ accumulation furtherconfirmed these results.
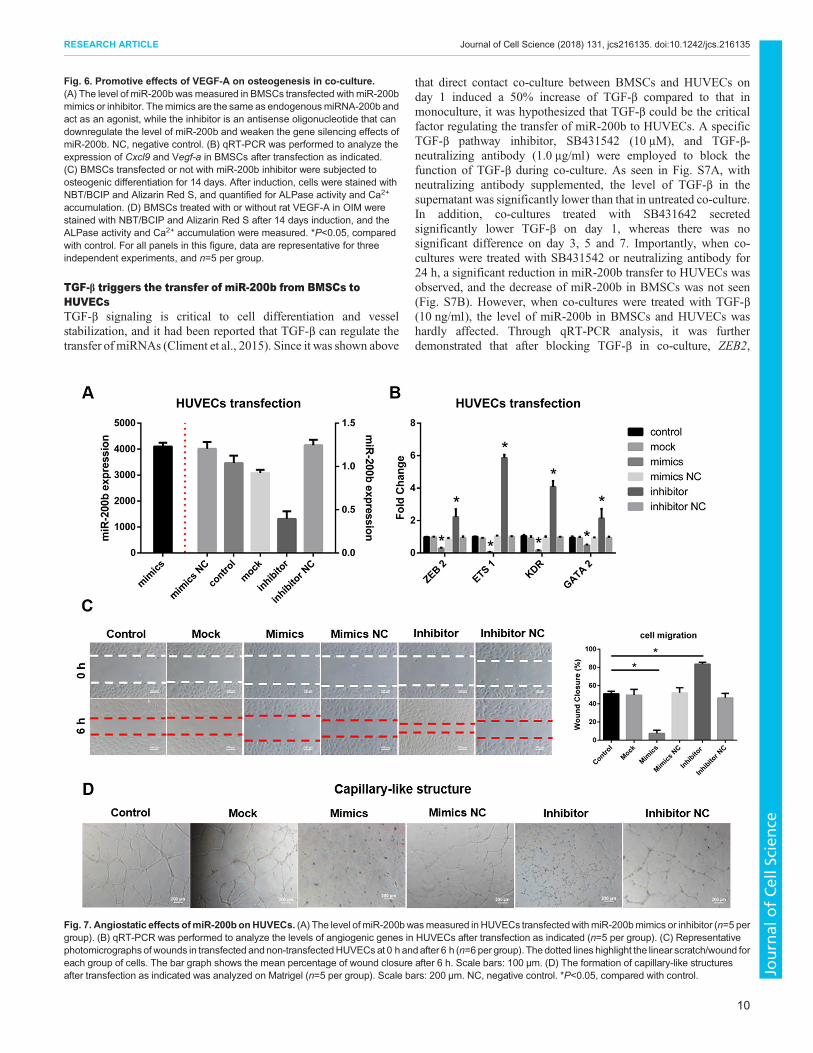
Angiostatic effects of miR-200b on HUVECsTo investigate the effects of miR-200b on angiogenesis, miR-200bmimics and inhibitor were transfected into HUVECs. Aftertransfection with the mimics, the level of miR-200b in HUVECshad a 4128-fold increase compared with non-transfected cells, whileit was reduced by 60% in cells transfected with the inhibitor(Fig. 7A). Additionally, as shown in Fig. 7B, HUVECs transfectedwith the mimics showed a significant decrease in the expression ofZEB2, ETS1, KDR and GATA2, whereas cells treated with theinhibitor showed the opposite.
Cell migration was also evaluated. A higher level of miR-200b inHUVECs transfected with the mimics induced a lower cellmigration rate than what was seen in control, while the migrationrate of cells treated with the inhibitor was greatly enhanced(Fig. 7C). Then, an in vitro Matrigel tube formation assay was
Fig. 5. Transfer of miR-200b from BMSCs toHUVECs via gap junctions. (A) Expressionprofile of miR-200b in BMSC and HUVECmonoculture. Levels of miR-200b areexpressed relative to levels of U6 snRNA.(B) The level of miR-200b was measured insorted BMSCs and HUVECs after co-culture for24 h and is presented relative to that in themonoculture. (C) BMSCs were transfected withmiR-200b labeled with the reporter dye FAM(green cells), then plated with HUVECs stainedwith the fluorescent dye DiL (red cells) in a ratioof 1:2. The transport of miR-200b for 2 h and24 h co-culture was visualized under afluorescence microscope. Scale bars: 100 μm.(D) HUVEC and BMSC co-cultures weretreated with PTH, 18GA or GW4869 (co-PTH,co-18GA, co-GW4869) for 24 h and thensubjected to flow cytometry sorting. The levelof miR-200b was measured in co-culturedBMSCs and HUVECs after cell sorting.*P<0.05, comparedwith monoculture; #P<0.05,compared with co-culture and is presentedrelative to that in the monoculture. Data arerepresentative of three independentexperiments, and n=5 per group.
8
RESEARCH ARTICLE Journal of Cell Science (2018) 131, jcs216135. doi:10.1242/jcs.216135
Journal
ofCe
llScience
exploited. It was found that HUVECs transfected with miR-200bmimics remained spherical and isolated, and no small cellularclusters or tubes appeared (Fig. 7D). In contrast, cells treated with
the inhibitor formed branching and anastomosing tubes, resulting ina network with a capillary-like structure. These data suggest thatmiR-200b has an angiostatic effect on HUVECs.
Fig. 6. See next page for legend.
9
RESEARCH ARTICLE Journal of Cell Science (2018) 131, jcs216135. doi:10.1242/jcs.216135
Journal
ofCe
llScience
TGF-β triggers the transfer of miR-200b from BMSCs toHUVECsTGF-β signaling is critical to cell differentiation and vesselstabilization, and it had been reported that TGF-β can regulate thetransfer of miRNAs (Climent et al., 2015). Since it was shown above
that direct contact co-culture between BMSCs and HUVECs onday 1 induced a 50% increase of TGF-β compared to that inmonoculture, it was hypothesized that TGF-β could be the criticalfactor regulating the transfer of miR-200b to HUVECs. A specificTGF-β pathway inhibitor, SB431542 (10 μM), and TGF-β-neutralizing antibody (1.0 μg/ml) were employed to block thefunction of TGF-β during co-culture. As seen in Fig. S7A, withneutralizing antibody supplemented, the level of TGF-β in thesupernatant was significantly lower than that in untreated co-culture.In addition, co-cultures treated with SB431642 secretedsignificantly lower TGF-β on day 1, whereas there was nosignificant difference on day 3, 5 and 7. Importantly, when co-cultures were treated with SB431542 or neutralizing antibody for24 h, a significant reduction in miR-200b transfer to HUVECs wasobserved, and the decrease of miR-200b in BMSCs was not seen(Fig. S7B). However, when co-cultures were treated with TGF-β(10 ng/ml), the level of miR-200b in BMSCs and HUVECs washardly affected. Through qRT-PCR analysis, it was furtherdemonstrated that after blocking TGF-β in co-culture, ZEB2,
Fig. 6. Promotive effects of VEGF-A on osteogenesis in co-culture.(A) The level of miR-200b wasmeasured in BMSCs transfected with miR-200bmimics or inhibitor. Themimics are the same as endogenousmiRNA-200b andact as an agonist, while the inhibitor is an antisense oligonucleotide that candownregulate the level of miR-200b and weaken the gene silencing effects ofmiR-200b. NC, negative control. (B) qRT-PCR was performed to analyze theexpression of Cxcl9 and Vegf-a in BMSCs after transfection as indicated.(C) BMSCs transfected or not with miR-200b inhibitor were subjected toosteogenic differentiation for 14 days. After induction, cells were stained withNBT/BCIP and Alizarin Red S, and quantified for ALPase activity and Ca2+
accumulation. (D) BMSCs treated with or without rat VEGF-A in OIM werestained with NBT/BCIP and Alizarin Red S after 14 days induction, and theALPase activity and Ca2+ accumulation were measured. *P<0.05, comparedwith control. For all panels in this figure, data are representative for threeindependent experiments, and n=5 per group.
Fig. 7. Angiostatic effects ofmiR-200bonHUVECs. (A) The level of miR-200bwasmeasured in HUVECs transfectedwithmiR-200bmimics or inhibitor (n=5 pergroup). (B) qRT-PCR was performed to analyze the levels of angiogenic genes in HUVECs after transfection as indicated (n=5 per group). (C) Representativephotomicrographsof wounds in transfected andnon-transfectedHUVECsat 0 h andafter 6 h (n=6per group). Thedotted lines highlight the linear scratch/wound foreach group of cells. The bar graph shows the mean percentage of wound closure after 6 h. Scale bars: 100 μm. (D) The formation of capillary-like structuresafter transfection as indicated was analyzed on Matrigel (n=5 per group). Scale bars: 200 μm. NC, negative control. *P<0.05, compared with control.
10
RESEARCH ARTICLE Journal of Cell Science (2018) 131, jcs216135. doi:10.1242/jcs.216135
Journal
ofCe
llScience

ETS1, KDR andGATA2were all significantly upregulated, althoughtheir expression levels were still lower than those in HUVECmonoculture (Fig. S7C).A vessel formation assay on Matrigel was further undertaken to
evaluate the angiogenesis in co-culture after supplementing TGF-β,SB431542 or neutralizing antibody. As shown in Fig. S7D,HUVECs alone on Matrigel could form intact capillary-likestructures, whereas cells in co-culture without any treatmentremained spherical and dispersed. Co-cultures treated with TGF-βexhibited similarly dispersed, heterogeneous cell clusters onMatrigel. In contrast, angiogenesis was partly recovered in co-cultures when they were treated with SB431542 or neutralizingantibody, as evidenced by formation of small cellular nests andshort tubes.
DISCUSSIONBone regeneration occurs in close spatial and temporal associationwith angiogenesis in vivo. Therefore, to engineer a vascularized bonetissue in vitro, it is necessary to take both osteogenesis andangiogenesis into consideration. By using co-culture betweenBMSCs and HUVECs, miR-200b was found to be an importantregulator for both osteogenesis and angiogenesis, in a manner thatdepended on the formation of functional gap junctions between thetwo cell types. A proposed mechanism of action is illustrated inFig. 8. BMSCs and HUVECs in direct co-culture could formfunctional gap junctions via Cx43, through which miR-200b couldtransfer from BMSCs to HUVECs. TGF-β secreted by HUVECswas the critical factor triggering the transfer of miR-200b. Asa negative regulator, the reduction of miR-200b in BMSCsinduced upregulation of VEGF, thus promoting the osteogenicdifferentiation. Once inside HUVECs, miR-200b could diminish theangiogenesis of HUVECs by reducing the expression of ETS1,GATA2, ZEB2 and KDR, thus inhibiting cell migration and theformation of a vascular-like network. Although VEGF couldpromote angiogenesis, the downregulation of its receptor KDR inHUVECs limited its function.Previous studies have concentrated on assessing the angiogenesis
in co-culture between ECs and MSCs in proliferative medium. Toaddress the impact of ECs on osteogenesis of MSCs, cells were also
co-cultured in either osteogenic or osteogenic factor-free medium.However, few studies have addressed osteogenesis and angiogenesissimultaneously in co-culture in osteogenic induction conditions. Inthe present work, rat BMSCs and HUVECswere directly co-culturedin osteogenicmedium containing endothelial cell growth supplement(ECGS) to keep endothelial cells alive. It was found that theosteogenic differentiation of BMSCs was significantly stimulated inco-culture compared to what was seen in monoculture within14 days. However, the formation of capillary-like structures in co-culture was compromised. Previous studies have shown that whenMSCs and ECs are co-cultured in proliferative medium, MSCs cansupport EC forming the capillary-like structures (Li et al., 2011,2015; Carrion et al., 2013). Consistent with these findings, inco-culture between BMSCs and HUVECs in growth medium,vascularized structures were seen in the present study. It is possiblethat the osteogenic state of BMSCs may not support the formation ofvascularized structure by ECs. In fact, the capillary-like structuresfailed to form in co-culture between the osteogenic-induced BMSCsand HUVECs (data not shown). Scherzed et al. have also reportedthat differentiated human MSCs showed restricted migration ability,which meant they were no longer able to support tube formation byECs (Scherzed et al., 2016). On the other hand, the impact of ECs onosteogenesis of MSCs is controversial. Most studies have found thatECs had positive effects on osteogenesis of MSCs, with enhancedosteogenic gene expression, Ca2+ accumulation and ALPase activity(Gershovich et al., 2013; Kim et al., 2013; Gurel Pekozer et al., 2016;Wu et al., 2016). Other studies have indicated that ECsmay negatively affect osteogenesis of MSCs. Duttenhoefer et al.found that endothelial progenitor cells impaired the osteogenicdifferentiation of BMSCs (Duttenhoefer et al., 2015). Co-culturebetween adipose tissue-derived MSCs and HUVECs led to less-efficient osteogenesis and repair of rat cranial defects than in theMSCmonoculture group (Ma et al., 2014). Meury et al. indicated that ECsinhibited osteogenesis of BMSCs through interfering with Osterix(also known as SP7) expression (Meury et al., 2006).
Communication, regulation of cellular processes and developmentin cell culture are impossible without connexins. They are importantand obligatory mediators that are involved throughout angiogenesisand osteogenesis. Cx43 is the most abundant connexin in the
Fig. 8. Proposed mechanism of action for miR-200b transferred through gap junctions inregulating angiogenesis and osteogenesis in co-culture.When BMSCs and HUVECs were co-culturedin direct contact, functional gap junctions composed ofCx43 are formed. TGF-β secreted by HUVECs triggersthe transfer of miR-200b from BMSCs to HUVECsvia gap junctions to regulate osteogenesis andangiogenesis. After transfer, the lower amounts ofmiR-200b in BMSCs leads to an increased expressionof VEGF-A. The enhanced VEGF-A has positiveimpacts on osteogenesis of BMSCs. Once insideHUVECs, miR-200b could reduce the levels ofangiogenic molecules, including ETS1, GATA2, ZEB2and KDR, thus inhibiting cell migration and preventingHUVECs from forming vascular-like structures.
11
RESEARCH ARTICLE Journal of Cell Science (2018) 131, jcs216135. doi:10.1242/jcs.216135
Journal
ofCe
llScience

osteoblast lineage and plays critical roles in bone formation (Watkinset al., 2011). In addition, downregulation of endothelial Cx43 causesendothelial dysfunction and impaired angiogenesis (Wang et al.,2013). In vivo, osteogenic cells residing in a perivascular niche lie inclose proximity to ECs, and therefore gap junctional communicationbetween heterotypic cells is inevitable (Crisan et al., 2008). Severalstudies have revealed that by directly co-culturing osteogenic cellsand ECs, gap junctions were formed via Cx43, and only with directcontact, could osteogenesis be enhanced (Villars et al., 2000, 2002;Guillotin et al., 2008; Herzog et al., 2014). In addition, variouspathways acting downstream of Cx43 in bone, including thoseinvolving ERK, PKCδ, β-catenin and protein kinase A, have beenreported (Stains and Civitelli, 2005; Hebert and Stains, 2013; Guptaet al., 2016). Cx43-dependent ERK and PKCδ pathways furtherregulate the transcriptional activity of Runx2, with increased Cx43level promoting Runx2 activity, which drives the expression ofseveral osteoblast genes and vice versa (Niger et al., 2013). In thepresent study, when BMSCs and HUVECs were in direct co-culture,osteogenic differentiation of BMSCs was significantly enhanced.In addition, inhibition of Cx43 in BMSC monoculture couldsignificantly decrease the expression of Runx2. However, blockingthe gap junctional intercellular communication between the two celltypes could not induce the downregulation of Runx2. In addition,18GA, an inhibitor of gap junction function, had no obvious negativeimpact on the expression of ERK. In one report, inhibition of gapjunctions was shown to block the expression of Ocn and Bsp inosteoblasts, and its effect on the expression of collagen type I was low(Schiller et al., 2001).However, the effects of 18GAon the expressionofOcn and collagen I inBMSCmonoculture and co-culturewere veryminimal in the present study. Importantly, it was found that theexpression level of Cxcl9 and Vegf-a was regulated by gap junctionsformed between BMSCs and HUVECs. Moreover, by exploitingsiRNAs, a correlation between Cxcl9 and Vegf-a levels and gapjunction function was established. The elevated level of VEGF-Acould partly be responsible for the enhancement of osteogenesis inBMSCs in co-culture. The secreted Cxcl9 had almost no influence onALPase activity, and slightly inhibited Ca2+ accumulation in the latephase of osteogenesis (data not shown).Cxcl9, as an angiostatic factorsecreted by osteoblasts, could interact with VEGF and preventits binding to ECs and osteoblasts, thus abrogating angiogenesisand osteogenesis both in vivo and in vitro (Huang et al., 2016).In co-culture as described in the present study, Cxcl9 slightlyinhibited cell proliferation and migration and prevented HUVECsfrom forming capillary-like structures. However, Cxcl9 was notresponsible for downregulation of angiogenic genes, includingETS1,GATA2, ZEB2 and KDR (data not shown).The miR-200 family comprises five members, miR-200a, -200b,
-200c,miR-429 andmiR-141,which regulate the EMT, and all appearto regulate tumor angiogenesis by targeting and repressing theexpression of several key mRNAs. Several reports have linked lowmiR-200 levelswith stimulation of angiogenesis throughupregulationof ETS1, TUBB3, VEGF, KDR, SUZ12, ERRFI-1 and ZEB1/2;however, the clinical relevance ofmiR-200 expression appears tovaryfor different cancers (Pecot et al., 2013). ETS1, originally detected inlymphoid cells of adult tissues, has been identified as being the firsttranscription factor expressed during the angiogenesis of ECs in theembryo (Lelievre et al., 2001). It has been reported that miR-200btargets ETS1 and induces inhibition of angiogenic response in ECs(Chan et al., 2011). GATA2 is critical for proliferation anddifferentiation of early hematopoietic progenitors (Tsai et al., 1994).GATA2 regulates the expression of vascular endothelial growthfactor receptor-2 (VEGFR-2) during both vascular development
and angiogenesis, the process by which ECs form new bloodvessels from an existing vascular network (Mammoto et al., 2009).Transcription factors of the ZEB protein family (ZEB1 and ZEB2)and miR-200 family members can form a double-negative feedbackloop, which controls EMT and mesenchymal–epithelial transition(MET) programs in both development and tumorigenesis (Hillet al., 2013). It has also been demonstrated that KDR, as animportant VEGFR, is negatively regulated by miR-200b (Choiet al., 2011). In the present study, after co-culture, the level ofmiR-200b in HUVECs was significantly enhanced, whereas adecreased level was noticed in BMSCs. The critical role of miR-200b in co-culture was further confirmed upon the transfection ofBMSCs with its mimics and inhibitor. As an angiostatic factor, ahigher level of miR-200b in HUVECs reduced the expression ofETS1, GATA2, ZEB2 and KDR, thus inhibiting cell migration andthe formation of a vascular-like network. Moreover, inhibition ofmiR-200b in BMSCs derepressed the expression of VEGF andthus promoted osteogenesis.
Mounting evidence implies that miRNAs could be considered as alanguage that allows cells to communicate with each other, stressingtheir important roles in the intercellular signal transduction. Forexample, smooth muscle cells (SMCs) can deliver miR-143 and/or-145 (miR-143/145) to ECs via intercellular tubes. Once in ECs,miR-143/145 repressed hexokinase II and integrin β8 and therebyreduces the angiogenic potential of the recipient cell (Climent et al.,2015). Zhou et al. found that endothelial miR-126 acts as a keyintercellular mediator for an increase in SMC turnover by repressingthe expression of FOXO3, BCL2 and IRS1 (Zhou et al., 2013). Sunet al. demonstrated that osteoclasts secreted miRNA-enrichedexosomes, which mediated a transfer of miR-214 into osteoblaststo inhibit their function (Sun et al., 2016). MSC-derived exosomesmight serve as an important mediator of cell-to-cell communicationwithin the tumor microenvironment that would suppressangiogenesis by transferring anti-angiogenic molecules, such asmiR-16 (Lee et al., 2013). Additionally, miR-145 has been shownto transfer from microvascular ECs to colon cancer cells, thusinhibiting angiogenesis (Thuringer et al., 2016). However, the rolesof miRNAs in the crosstalk between MSCs and ECs remain largelyelusive. Liang et al. found human adipose-derived MSCs secretedexosomes enriched with miR-125a, which could regulateangiogenesis of ECs in vitro and in vivo (Liang et al., 2016). Here,we show that miR-200b is transferred from cell-to-cell via gapjunctions, as well as demonstrating its role in intercellularcommunication between MSCs and ECs. Therefore, it would bepossible to modulate the expression as well as transfer of miR-200bfor better formation of vascularized bone tissues. Taken together, ourfindings may be fundamental to the development of cell-based boneregeneration strategies by providing new insight into how an efficientstrategy for engineering pre-vascularized bone tissue in vitro couldbe developed.
MATERIALS AND METHODSCell isolation and cell cultureAll procedures on rats were performed in full compliancewith the guidelinesof the ethics committee at East China University of Science and Technology.We used 4-week-old Sprague Dawley rats (SD rats), purchased fromShanghai SLAC Laboratory Animal Co., Ltd, that were SPF grade, male andweighed ∼80–120 g. Bone marrow mesenchymal stem cells (BMSCs) wereisolated using a bonemarrow adherence method as previously described (Jinet al., 2018). Cells were cultured in minimum essential medium α medium(α-MEM) (Gibco) supplemented with 10% fetal bovine serum (FBS)(Hyclone) at 37°C in a 5% CO2 humidified atmosphere (proliferativemedium). Cells at passage 3 to 6 were used in our experiment. Cell surface
12
RESEARCH ARTICLE Journal of Cell Science (2018) 131, jcs216135. doi:10.1242/jcs.216135
Journal
ofCe
llScience

antigens were analyzed on a flow cytometer (BD FACS Calibur). Theantibodies used are shown in Table S1.
Human umbilical vein endothelial cells (HUVECs) were purchased fromScienCell and cultured in endothelial cell growth medium (ECM)(ScienCell), containing 5% FBS, 100× penicillin-streptomycin solutionand supplemented with 100× endothelial cell growth supplement (ECGS).Cells at passage 3 to 8 were used in our experiment.
Cell differentiation assayBMSCs at passage 3 were seeded in 24-well plates at a density of 1×104 cells/well and grown in α-MEM.After cell adhesion, the proliferative mediumwasdiscarded, and a specializedmedium, according to the desired differentiation,was directly added to the cell culture. The assays were performed in triplicatefor each group, with three independent experiments.
Osteogenic differentiationThe cultured cells were treated with osteogenic induction medium (OIM),comprising Dulbecco’s modified Eagle’s medium (DMEM) (Gibco)supplemented with 10% FBS, 10−7 M dexamethasone (Sigma), 10 mMβ-glycerol phosphate (Sigma) and 50 μg/ml L-ascorbic acid (Sigma). Themedium was replaced twice a week. According to different experiments,factors such as rat VEGF-A (10 ng/ml) (Peprotech) were added to OIM asneeded. After induction, mineralized nodules were detected by Alizarin RedS staining. ALP enzymatic activity was analyzed by using the NBT/BCIPalkaline phosphatase color development kit (Beyotime). The quantificationof Ca2+ accumulation and ALPase activity was performed as previouslydescribed (Fan et al., 2017).
Adipogenic differentiationThe cells were cultured in adipogenic induction medium consisting ofDMEM, 10% FBS, 1 μM dexamethasone, 0.1 mM indomethacin (Sigma),0.1 mM 3-isobutyl-1-methyl-xanthine (IBMX) (Sigma) and 10 mg/l insulin(Sigma). The medium was changed twice a week. After induction, cellswere stained with 0.2% Oil Red O solution (Sigma) to detect lipid dropletswithin the cells.
Chondrogenic differentiation1×106 cells were suspended in 20 μl α-MEM to form a high-density cellpellet. The cell pellet was induced using chondrogenic medium, comprisingDMEM, 1% ITS+ (BD Biosciences), 50 μg/ml L-ascorbic acid, 0.1 μMdexamethasone, 40 μg/ml L-proline (Sigma), 1 mM sodium pyruvate(Sigma) and supplemented with 10 ng/ml TGF-β1 (Peprotech). Themedium was changed every 3 days. After induction, cells were stained withSafranin O (Sigma).
Co-culture settingsTo carry out direct co-culture, both cell types were expanded separately inthe media described above and then co-cultured in 24-well plates orcell culture flasks. For co-culture, ECGS was used to maintain thesurvival of ECs. BMSCs and HUVECs were mixed and seeded at a ratio of1:2 in growth medium (1:1 α-MEM:ECM supplemented with 100× ECGS),at the density of 5000 cells/cm2 and 10,000 cells/cm2, respectively. Cellswere seeded in the growth medium. After cell adhesion, the growth mediumwas replaced with OIM, and according to the different experiments, 100 μM18GA (Sigma), 10 nM PTH (Sigma), 10 μM GW4869 (Sigma), 10 μMSB431542 (Sigma), 10 ng/ml TGF-β (Peprotech), 1.0 μg/ml neutralizingantibody of TGF-β (R&D) or 0.5 μg/ml VEGF blocking antibody (R&D)was added into the OIM.
Indirect co-culture was established using Transwell inserts with 0.4-μmfilters (Corning). Depending on different experiments, either BMSCs orHUVECs were plated in the Transwell inserts.
RNA isolation and qRT-PCRTotal RNA was extracted using Trizol reagent (Invitrogen) and cDNA wasobtained using MLV reverse transcriptase (Promega) following themanufacturer’s instructions. Quantitative real-time PCR (qRT-PCR) wasperformed on a RT-PCR system (Bio-Rad CFX96) using a SYBR mix(Roche) according to the manufacturer’s instructions. PCR conditions were
95°C for 10 min, followed by 40 cycles of amplification at 95°C for 10 s and57°C–63°C (depending on the primers used) for 20 s.GAPDHwas used as ahouse keeping gene.
For semi-quantitative PCR analysis, PCR conditions were 94°C for3 min, followed by 30 cycles of amplification at 94°C for 30 s, and 60°C for30 s. PCR products in a 25 μl aliquot were size separated by electrophoresisin 2% agarose gels.
For miRNA isolation, total RNAwas extracted with a miRNeasy Mini Kit(Qiagen) according to the manufacturer’s instructions. Reverse transcriptionof 1 μg RNAwas carried out with a miScript II RT Kit (Qiagen). AMiScriptSYBR Green PCR kit was used to quantitatively analyze the level of maturemiRNAs.U6 snRNAwas used as an internal control. The relative quantitativeexpression of miR-200b was normalized to the level of U6 snRNA. Thespecific primers for miR-200b and U6 were purchased from Qiagen.
Gene expression was analyzed according to the ΔΔCT method. Thespecifics of primers are listed in Table S2.
Western blottingCells were harvested and lysed in protein extraction reagent RIPA lysisbuffer (Beyotime) containing 10 mM phenylmethylsulphonyl fluoride(PMSF) (Beyotime) on ice for 30 min. After centrifugation for 1 h at12,000 g, the supernatantwas retained. Protein concentrationswere determinedby using the BCA assay (Beyotime) following the manufacturer’s instruction.After being heated for 5 min at 95°C in the sample buffer (Beyotime), equalaliquots of cell lysates were run on a 10% SDS-polyacrylamide gel. Theseparated proteins were electrically transferred onto PVDF membrane(Millipore). The membrane was blocked with 5% (w/v) non-fat dry milk inTBST buffer (20 mMTris-HCl pH 7.4, 150 mMNaCl, 0.05%Tween-20) for1 h at room temperature, and then incubated overnight at 4°Cwith the primaryantibodies (Table S3). The bound primary antibodies were detected withsecondary antibodies conjugated to alkaline phosphatase (ALPase) orhorseradish peroxidase (HRP), and visualized through NBT/BCIP staining(Beyotime) or enhanced chemiluminescence (Millipore). β-actin was used asan internal control, and its expression was used to standardize input protein toanalyze relative protein expression.
Immunofluorescence stainingCells in 24-well plates were fixed in 4% (w/v) paraformaldehyde for 10 minat room temperature. After washing in PBS, cells were permeabilized with0.25% Triton X-100 for 10 min and blocked with 1% (w/v) bovine serumalbumin (BSA) for 30 min. Cells were incubated for 60 min at roomtemperature with primary antibodies (Table S3), each diluted at 1:100. Assecondary antibodies, Alexa Fluor 488 goat anti-rabbit-IgG (Santa CruzBiotechnology) at a dilution of 1:200 were used. The cell nucleus wasvisualized via DAPI (Invitrogen) staining. Fluorescence staining wasvisualized in a fluorescence microscope (Nikon Eclipse Ti).
Gap junction functional assayThe function of the gap junctions was assessed using a ‘parachute’ dye-coupling assay. Donor cells (BMSCs or HUVECs) were double-labeledwith 1 μM Calcein AM (Sigma), which is intracellularly convertedinto the gap junction-permeable dye Calcein, and 5 μM Dil (Beyotime),which is a membrane dye that cannot spread to coupled cells, for 30 min.Unincorporated dye was removed by three consecutive washes with culturemedium. Donor cells were then trypsinized and seeded onto the receivercells (HUVECs or BMSCs) at a 1:200 ratio, and the two cell types wereallowed to attach to the monolayer and form gap junctions for 3 h at 37°C.Fluorescence microscope was used for imaging.
Cell transfectionCells were transfected by means of Lipofectamine RNAiMAX according tothe manufacturer’s protocol (Invitrogen).
siRNAs targeting the rat Cx43 gene (3646, 5′-GCUGGUUACUGGU-GACAGA-3′; 3710, 5′-GAACUACAGCGCAGAGCAA-3′) (RiboBio)were used to specifically downregulate the level of Cx43 in BMSCs, andnonspecific sequence siRNAs were used as a negative controls (NC)(RiboBio). Cells without transfection served as control, and cells treatedwith transfection reagent were considered mock. After 24 to 48 h of siRNA
13
RESEARCH ARTICLE Journal of Cell Science (2018) 131, jcs216135. doi:10.1242/jcs.216135
Journal
ofCe
llScience

transfection, the knockdown efficiency was confirmed by qRT-PCR orwestern blot analysis.
miR-200b mimics (FAM labeled or not) or inhibitor (RiboBio) weretransfected into BMSCs and HUVECs to evaluate its function, and negativecontrol miRNAs (mimics NC and inhibitor NC) (RiboBio) were also used.MiR-200b mimics are synthetic double stranded RNAs, same as endogenousmiRNA-200b. TransfectionwithmiR-200bmimics can lead to a high level ofmiRNA-200b, and their regulatory role would be boosted, meaning that therewould be gain-of-function effects. miR-200b inhibitor is a synthetic single-stranded RNA, and functions as antisense oligonucleotides that can preventmiR-200b from binding to its target mRNAs, thereby weakening the genesilencing effects. Their negative controlmiRNAs are the oligonucleotides thatdo not react with any mRNAs in cells. After 24 h of transfection, the level ofmiR-200b was measured by qRT-PCR or visualized with a fluorescencemicroscope. Sequences were as follows: miR-200b mimics, sense, 5′-UA-AUACUGCCUGGUAAUGAUGA-3′ and antisense, 5′-UCAUCAUUAC-CAGGCAGUAUUA-3′; miR-200b mimics NC, sense, 5‘-UUUGUACUA-CACAAAAGUACUG-3′ and antisense, 5‘-CAGUACUUUUGUGUAGU-ACAAA-3′; miR-200b inhibitor, sense, 5′-UCAUCAUUACCAGGCAGU-AUUA-3′; miR-200b inhibitor NC, sense, 5‘-CAGUACUUUUGUGUA-GUACAAA-3′.
Enzyme-linked immunosorbent assayELISA was used to quantify the content of rat vascular endothelial growthfactor (VEGF) (E-EL-R2603c, Elabscience), rat chemokine (C-X-C motif)ligand 9 (Cxcl9) (E-EL-R0854c, Elabscience), rat chemokine (C-X-C motif)ligand 12 (Cxcl12) (E-EL-R0922c, Elabscience) and human TGF-β1 (E-EL-H0110c, Elabscience) expressed by cells. Culture media was collected andanalyzed at the specific time points. Assays were carried out according to themanufacturer’s protocol and analyzed on a microplate reader at a wavelengthof 450 nm (Biotek ELX800). The assays were performed in triplicates foreach group, with three independent experiments.
Flow cytometry cell sortingCo-cultures were incubated with 18GA (100 μM), PTH (10 nM), GW4869(10 μM), SB431542 (10 μM), TGF-β1 (10 ng/ml) or TGF-β-neutralizingantibody (1.0 μg/ml) in 75 cm2 culture flasks. After 24 h, cells weretrypsinized and incubated with PE-conjugated anti-hCD31 antibody (R&D;Table S1). Co-cultured BMDCs and HUVECs can be separated by flowcytometry based on the fluorescence dye.
In vitro migration assayHUVECs were seeded at the density of 5000 cells/cm2 in 24-well plates.Confluent cells (or transfected HUVECs) were serum-deprived for 16 h, anda linear wound was created in monolayer by scratching with a sterile pipettetip (200 μl yellow tip). Cells werewashed with PBS to remove floating cells.After additional 6 h, cell migration into the wound was assessed by using adigital inverted microscope. The percentage of wound closure for a giventime was calculated from at least five images.
In vitro tube formation assayHUVEC monoculture (or transfected HUVECs) or co-cultures were seededon growth factor-depleted Matrigel (BD). The Matrigel was assessed forcapillary-like structures 12 h later. Microscopic fields containing the tubestructure formed on the gel were photographed at low magnification (×40).Five fields per test condition were examined.
Data analysisAll the experiments were repeated at least three times using three rats toprovide BMSCs and from pooled HUVECs. All the data are presented asmeans±s.d. Statistical significance was evaluated using one-way ANOVAwith the Student–Newman–Keuls post-hoc test when data could be assessedas normally distributed. Non-normally distributed data were statisticallyanalyzed using the nonparametric Kruskal–Wallis test. A value of P<0.05was considered statistically significant.
Competing interestsThe authors declare no competing or financial interests.
Author contributionsMethodology: X.F.; Validation: Y.T.; Investigation: X.F., Y.T.; Data curation: X.F.;Writing - original draft: X.F.; Writing - review & editing: X.F., Y.T., Z.Y., Y.Z.;Visualization: Y.Z.; Supervision: Z.Y., Y.Z., W.-S.T.; Project administration: Z.Y.,Y.Z., W.-S.T.; Funding acquisition: Z.Y., Y.Z.
FundingThis research was supported by the National Natural Science Foundation of China(grant no. 31170951 and 81671841), the Natural Science Foundation of Shanghai(grant no. 16ZR1408700) and the Key Project of Science and Technology ofShanghai (grant no. 16JC1400203).
Supplementary informationSupplementary information available online athttp://jcs.biologists.org/lookup/doi/10.1242/jcs.216135.supplemental
ReferencesAkhtar, S., Hartmann, P., Karshovska, E., Rinderknecht, F. A., Subramanian, P.,
Gremse, F., Grommes, J., Jacobs, M., Kiessling, F., Weber, C. et al. (2015).Endothelial hypoxia-inducible factor-1alpha promotes atherosclerosis andmonocyterecruitment by upregulating microRNA-19a. Hypertension 66, 1220-1226.
Baglio, S. R., Rooijers, K., Koppers-Lalic, D., Verweij, F. J., Perez Lanzon, M.,Zini, N., Naaijkens, B., Perut, F., Niessen, H. W. M., Baldini, N. et al. (2015).Human bone marrow- and adipose-mesenchymal stem cells secrete exosomesenriched in distinctive miRNA and tRNA species. Stem Cell Res. Ther. 6, 127.
Brabletz, S. and Brabletz, T. (2010). The ZEB/miR-200 feedback loop–a motor ofcellular plasticity in development and cancer? EMBO Rep. 11, 670-677.
Calderon, J. F. and Retamal, M. A. (2016). Regulation of connexins expressionlevels by microRNAs, an update. Front. Physiol. 7, 558.
Carrion, B., Kong, Y. P., Kaigler, D. and Putnam, A. J. (2013). Bone marrow-derived mesenchymal stem cells enhance angiogenesis via their alpha6beta1integrin receptor. Exp. Cell Res. 319, 2964-2976.
Chan, Y. C., Khanna, S., Roy, S. and Sen, C. K. (2011). miR-200b targets Ets-1and is down-regulated by hypoxia to induce angiogenic response of endothelialcells. J. Biol. Chem. 286, 2047-2056.
Choi, Y.-C., Yoon, S., Jeong, Y., Yoon, J. and Baek, K. (2011). Regulation ofvascular endothelial growth factor signaling by miR-200b. Mol. Cells 32, 77-82.
Climent, M., Quintavalle, M., Miragoli, M., Chen, J., Condorelli, G. and Elia, L.(2015). TGFbeta triggers miR-143/145 transfer from smooth muscle cells toendothelial cells, therebymodulating vessel stabilization.Circ. Res. 116, 1753-1764.
Crisan, M., Yap, S., Casteilla, L., Chen, C.-W., Corselli, M., Park, T. S., Andriolo,G., Sun, B., Zheng, B., Zhang, L. et al. (2008). A perivascular origin formesenchymal stem cells in multiple human organs. Cell Stem Cell 3, 301-313.
Duttenhoefer, F., Lara de Freitas, R., Loibl, M., Bittermann, G., Geoff Richards,R., Alini, M. and Verrier, S. (2015). Endothelial progenitor cell fraction containedin bone marrow-derived mesenchymal stem cell populations impairs osteogenicdifferentiation. Biomed. Res. Int. 2015, 659542.
Fan, X., Li, L., Ye, Z., Zhou, Y. and Tan,W. S. (2017). Regulation of osteogenesis ofhuman amniotic mesenchymal stem cells by sodium butyrate. Cell Biol. Int. 42,457-469.
Gershovich, J. G., Dahlin, R. L., Kasper, F. K. andMikos, A. G. (2013). Enhancedosteogenesis in cocultures with human mesenchymal stem cells and endothelialcells on polymeric microfiber scaffolds. Tissue Eng. Part A 19, 2565-2576.
Grellier, M., Ferreira-Tojais, N., Bourget, C., Bareille, R., Guillemot, F. andAmedee, J. (2009). Role of vascular endothelial growth factor in thecommunication between human osteoprogenitors and endothelial cells. J. Cell.Biochem. 106, 390-398.
Guillotin, B., Bourget, C., Remy-Zolgadri, M., Bareille, R., Fernandez, P.,Conrad, V. andAmedee-Vilamitjana, J. (2004). Human primary endothelial cellsstimulate human osteoprogenitor cell differentiation. Cell. Physiol. Biochem. 14,325-332.
Guillotin, B., Bareille, R., Bourget, C., Bordenave, L. and Amedee, J. (2008).Interaction between human umbilical vein endothelial cells and humanosteoprogenitors triggers pleiotropic effect that may support osteoblasticfunction. Bone 42, 1080-1091.
Gupta, A., Anderson, H., Buo, A. M., Moorer, M. C., Ren, M. and Stains, J. P.(2016). Communication of cAMP by connexin43 gap junctions regulatesosteoblast signaling and gene expression. Cell. Signal. 28, 1048-1057.
Gurel Pekozer, G., Torun Kose, G. and Hasirci, V. (2016). Influence of co-cultureon osteogenesis and angiogenesis of bone marrow mesenchymal stem cells andaortic endothelial cells. Microvasc. Res. 108, 1-9.
Hebert, C. and Stains, J. P. (2013). An intact connexin43 is required to enhancesignaling and gene expression in osteoblast-like cells. J. Cell. Biochem. 114,2542-2550.
Herzog, D. P. E., Dohle, E., Bischoff, I. and Kirkpatrick, C. J. (2014). Cellcommunication in a coculture system consisting of outgrowth endothelial cells andprimary osteoblasts. Biomed. Res. Int. 2014, 320123.
14
RESEARCH ARTICLE Journal of Cell Science (2018) 131, jcs216135. doi:10.1242/jcs.216135
Journal
ofCe
llScience

Hill, L., Browne, G. and Tulchinsky, E. (2013). ZEB/miR-200 feedback loop: at thecrossroads of signal transduction in cancer. Int. J. Cancer 132, 745-754.
Huang, B., Wang, W., Li, Q., Wang, Z., Yan, B., Zhang, Z., Wang, L., Huang, M.,Jia, C., Lu, J. et al. (2016). Osteoblasts secrete Cxcl9 to regulate angiogenesis inbone. Nat. Commun. 7, 13885.
Inose, H., Ochi, H., Kimura, A., Fujita, K., Xu, R., Sato, S., Iwasaki, M.,Sunamura, S., Takeuchi, Y., Fukumoto, S. et al. (2009). A microRNA regulatorymechanism of osteoblast differentiation. Proc. Natl. Acad. Sci. USA 106,20794-20799.
Jin, C., Tian, H., Li, J., Jia, S., Li, S., Xu, G. T., Xu, L. and Lu, L. (2018). Stem celleducation for medical students at Tongji University, Primary cell culture anddirectional differentiation of rat bone marrow mesenchymal stem cells. Biochem.Mol. Biol. Educ. 46, 151-154.
Kim, V. N. (2005). MicroRNAbiogenesis: coordinated cropping and dicing.Nat. Rev.Mol. Cell Biol. 6, 376-385.
Kim, J., Kim, H. N., Lim, K.-T., Kim, Y., Pandey, S., Garg, P., Choung, Y.-H.,Choung, P. H., Suh, K.-Y. and Chung, J. H. (2013). Synergistic effects ofnanotopography and co-culture with endothelial cells on osteogenesis ofmesenchymal stem cells. Biomaterials 34, 7257-7268.
Lee, J.-K., Park, S.-R., Jung, B.-K., Jeon, Y.-K., Lee, Y.-S., Kim, M.-K., Kim, Y.-G.,Jang, J.-Y. and Kim, C.-W. (2013). Exosomes derived from mesenchymal stemcells suppress angiogenesis by down-regulating VEGF expression in breastcancer cells. PLoS ONE 8, e84256.
Lelievre, E., Lionneton, F., Soncin, F. and Vandenbunder, B. (2001). The Etsfamily contains transcriptional activators and repressors involved in angiogenesis.Int. J. Biochem. Cell Biol. 33, 391-407.
Li, H., Daculsi, R., Grellier, M., Bareille, R., Bourget, C., Remy, M. and Amedee,J. (2011). The role of vascular actors in two dimensional dialogue of human bonemarrow stromal cell and endothelial cell for inducing self-assembled network.PLoS ONE 6, e16767.
Li, Y., Fan, L., Liu, S., Liu, W., Zhang, H., Zhou, T., Wu, D., Yang, P., Shen, L.,Chen, J. et al. (2013). The promotion of bone regeneration through positiveregulation of angiogenic-osteogenic coupling using microRNA-26a. Biomaterials34, 5048-5058.
Li, J., Ma, Y., Teng, R., Guan, Q., Lang, J., Fang, J., Long, H., Tian, G. andWu, Q.(2015). Transcriptional profiling reveals crosstalk between mesenchymal stemcells and endothelial cells promoting prevascularization by reciprocalmechanisms. Stem Cells Dev. 24, 610-623.
Liang, X., Zhang, L., Wang, S., Han, Q. and Zhao, R. C. (2016). Exosomessecreted by mesenchymal stem cells promote endothelial cell angiogenesis bytransferring miR-125a. J. Cell Sci. 129, 2182-2189.
Lu, J., Getz, G., Miska, E. A., Alvarez-Saavedra, E., Lamb, J., Peck, D., Sweet-Cordero, A., Ebert, B. L., Mak, R. H., Ferrando, A. A. et al. (2005). MicroRNAexpression profiles classify human cancers. Nature 435, 834-838.
Ma, J., Both, S. K., Ji, W., Yang, F., Prins, H.-J., Helder, M. N., Pan, J., Cui, F.-Z.,Jansen, J. A. and van den Beucken, J. J. P. (2014). Adipose tissue-derivedmesenchymal stem cells asmonocultures or cocultures with human umbilical veinendothelial cells: performance in vitro and in rat cranial defects. J. Biomed. Mater.Res. A 102, 1026-1036.
Mammoto, A., Connor, K. M., Mammoto, T., Yung, C. W., Huh, D., Aderman,C. M., Mostoslavsky, G., Smith, L. E. H. and Ingber, D. E. (2009). Amechanosensitive transcriptional mechanism that controls angiogenesis. Nature457, 1103-1108.
Meury, T., Verrier, S. and Alini, M. (2006). Human endothelial cells inhibit BMSCdifferentiation into mature osteoblasts in vitro by interfering with osterixexpression. J. Cell. Biochem. 98, 992-1006.
Montecalvo, A., Larregina, A. T., Shufesky, W. J., Stolz, D. B., Sullivan, M. L. G.,Karlsson, J. M., Baty, C. J., Gibson, G. A., Erdos, G., Wang, Z. et al. (2012).Mechanism of transfer of functional microRNAs betweenmouse dendritic cells viaexosomes. Blood 119, 756-766.
Moorer, M. C. and Stains, J. P. (2017). Connexin43 and the intercellular signalingnetwork regulating skeletal remodeling. Curr. Osteoporos Rep. 15, 24-31.
Niger, C., Luciotti, M. A., Buo, A. M., Hebert, C., Ma, V. and Stains, J. P. (2013).The regulation of runt-related transcription factor 2 by fibroblast growth factor-2and connexin43 requires the inositol polyphosphate/protein kinase Cdeltacascade. J. Bone Miner. Res. 28, 1468-1477.
Pecot, C. V., Rupaimoole, R., Yang, D., Akbani, R., Ivan, C., Lu, C., Wu, S., Han,H.-D., Shah, M. Y., Rodriguez-Aguayo, C. et al. (2013). Tumour angiogenesisregulation by the miR-200 family. Nat. Commun. 4, 2427.
Scherzed, A., Hackenberg, S., Froelich, K., Rak, K., Schendzielorz, P., Gehrke,T., Hagen, R. and Kleinsasser, N. (2016). The differentiation of hMSCscounteracts their migration capability and pro-angiogenic effects in vitro. Oncol.Rep. 35, 219-226.
Schiller, P. C., D’Ippolito, G., Balkan, W., Roos, B. A. and Howard, G. A. (2001).Gap-junctional communication is required for the maturation process ofosteoblastic cells in culture. Bone 28, 362-369.
Shen, A., Lin, W., Chen, Y., Liu, L., Chen, H., Zhuang, Q., Lin, J., Sferra, T. J. andPeng, J. (2015). Pien Tze Huang inhibits metastasis of human colorectalcarcinoma cells via modulation of TGF-beta1/ZEB/miR-200 signaling network.Int. J. Oncol. 46, 685-690.
Stains, J. P. and Civitelli, R. (2005). Gap junctions regulate extracellular signal-regulated kinase signaling to affect gene transcription. Mol. Biol. Cell 16, 64-72.
Sun, W., Zhao, C., Li, Y., Wang, L., Nie, G., Peng, J., Wang, A., Zhang, P., Tian,W., Li, Q. et al. (2016). Osteoclast-derived microRNA-containing exosomesselectively inhibit osteoblast activity. Cell Discov. 2, 16015.
Thuringer, D., Jego, G., Berthenet, K., Hammann, A., Solary, E. and Garrido, C.(2016). Gap junction-mediated transfer of miR-145-5p from microvascularendothelial cells to colon cancer cells inhibits angiogenesis. Oncotarget 7,28160-28168.
Tsai, F.-Y., Keller, G., Kuo, F. C., Weiss, M., Chen, J., Rosenblatt, M., Alt, F. W.and Orkin, S. H. (1994). An early haematopoietic defect in mice lacking thetranscription factor GATA-2. Nature 371, 221-226.
Tsigkou, O., Pomerantseva, I., Spencer, J. A., Redondo, P. A., Hart, A. R.,O’Doherty, E., Lin, Y., Friedrich, C. C., Daheron, L., Lin, C. P. et al. (2010).Engineered vascularized bone grafts. Proc. Natl. Acad. Sci. USA 107, 3311-3316.
Valiunas, V., Polosina, Y. Y., Miller, H., Potapova, I. A., Valiuniene, L., Doronin,S., Mathias, R. T., Robinson, R. B., Rosen, M. R., Cohen, I. S. et al. (2005).Connexin-specific cell-to-cell transfer of short interfering RNA by gap junctions.J. Physiol. 568, 459-468.
Villars, F., Bordenave, L., Bareille, R. and Amedee, J. (2000). Effect of humanendothelial cells on human bone marrow stromal cell phenotype: role of VEGF?.J. Cell. Biochem. 79, 672-685.
Villars, F., Guillotin, B., Amedee, T., Dutoya, S., Bordenave, L., Bareille, R. andAmedee, J. (2002). Effect of HUVEC on human osteoprogenitor celldifferentiation needs heterotypic gap junction communication. Am. J. Physiol.Cell Physiol. 282, C775-C785.
Vimalraj, S. and Selvamurugan, N. (2013). MicroRNAs: synthesis, gene regulationand osteoblast differentiation. Curr. Issues Mol. Biol. 15, 7-18.
Wang, H.-H., Su, C.-H., Wu, Y.-J., Li, J.-Y., Tseng, Y.-M., Lin, Y.-C., Hsieh, C.-L.,Tsai, C.-H. and Yeh, H.-I. (2013). Reduction of connexin43 in human endothelialprogenitor cells impairs the angiogenic potential. Angiogenesis 16, 553-560.
Watkins, M., Grimston, S. K., Norris, J. Y., Guillotin, B., Shaw, A., Beniash, E.and Civitelli, R. (2011). Osteoblast connexin43 modulates skeletal architectureby regulating both arms of bone remodeling. Mol. Biol. Cell 22, 1240-1251.
Wu, L., Zhao, X., He, B., Jiang, J., Xie, X. J. and Liu, L. (2016). The possible rolesof biological bone constructed with peripheral blood derived EPCs and BMSCs inosteogenesis and angiogenesis. Biomed. Res. Int. 2016, 8168943.
Xiong, M., Jiang, L., Zhou, Y., Qiu, W., Fang, L., Tan, R., Wen, P. and Yang, J.(2012). The miR-200 family regulates TGF-beta1-induced renal tubular epithelialto mesenchymal transition through Smad pathway by targeting ZEB1 and ZEB2expression. Am. J. Physiol. Renal. Physiol. 302, F369-F379.
Yang, M., Li, C.-J., Sun, X., Guo, Q., Xiao, Y., Su, T., Tu, M.-L., Peng, H., Lu, Q.,Liu, Q. et al. (2017). MiR-497∼195 cluster regulates angiogenesis duringcoupling with osteogenesis by maintaining endothelial Notch and HIF-1alphaactivity. Nat. Commun. 8, 16003.
Yoshizuka, M., Nakasa, T., Kawanishi, Y., Hachisuka, S., Furuta, T., Miyaki, S.,Adachi, N. and Ochi, M. (2016). Inhibition of microRNA-222 expressionaccelerates bone healing with enhancement of osteogenesis, chondrogenesis,and angiogenesis in a rat refractory fracture model. J. Orthop. Sci. 21, 852-858.
Zhou, J., Li, Y.-S., Nguyen, P., Wang, K.-C., Weiss, A., Kuo, Y.-C., Chiu, J.-J.,Shyy, J. Y. and Chien, S. (2013). Regulation of vascular smooth muscle cellturnover by endothelial cell-secreted microRNA-126: role of shear stress. Circ.Res. 113, 40-51.
Zong, L., Zhu, Y., Liang, R. and Zhao, H.-B. (2016). Gap junction mediated miRNAintercellular transfer and gene regulation: a novel mechanism for intercellulargenetic communication. Sci. Rep. 6, 19884.
15
RESEARCH ARTICLE Journal of Cell Science (2018) 131, jcs216135. doi:10.1242/jcs.216135
Journal
ofCe
llScience










![Leukocytes express activation lipopolysaccharide appear ... · free)/1% goat serum] for 30 min at roomtemperature. Then theywereincubatedwithanti-Cx43 antibody(1:100inblocking solution)](https://img.pdfslide.net/doc/110x75/603f27b1951d67438b291286/leukocytes-express-activation-lipopolysaccharide-appear-free1-goat-serum.jpg)