Embed Size (px)
Citation preview

290 Neuroscwnce Research, 10 ( 1991 ) 290- 295 ' 1991 Elsevier Scientific Publishers Ireland, Ltd. 0168-0102/')1/$(13 5(I
NEURES 00437
The effect of microstimulation of the oculomotor vermis on discharges of fastigial neurons and visually-directed
saccades in macaques
Kenji Ohtsuka* and Hiroharu Noda Visual Science Department. School of Optometry, Indiana Unwersity, Bloomington, IN (US .A)
(Received 13 September 1990; Revised version received 28 November 1990; Accepted 28 December 1990)
Key words: Vermal microstimulation; Saccadic burst neurons; Saccadic eye movements; Evoked saccades; Purkinje-cell inhibition
SUMMARY
The present study confirmed our previous reports that neurons in the fastigial ocuiomotor region (FOR) of the macaque show presaccadic bursts during contralateral saccades and that the burst duration is closely related to the duration of the accompanying saccade. Furthermore, when the burst duration was reduced by sub- threshold electrical stimulation applied to the oculomotor vermis prior to the onset of the burst, the impending visually-directed saccade became hypometric. The reduction in the burst duration was closely related to the degree of the hypometria. Since saccadic burst neurons in the FOR constitute the sole output channel for saccadic signals of the ocuiomotor vermis, the findings support the hypothesis that the cerebellum can regulate the amplitude of eye movements.
It has been shown that saccade- re la ted neurons in the deep cerebe l la r nuclei aggregate in a cauda l region of the fast igial nucleus, des igna ted as the fast igial o c u l o m o t o r region ( F O R ) 5. The F O R coincides ana tomica l ly wi th the a rea which receives Purk in je cell (P-cell) axons f rom lobules VIc and VII , name ly f rom the oc u lomo to r vermis s. N e u rons in the F O R show burs ts of f ir ing dur ing saccades in the d i rec t ion con t ra l a t e ra l to the recording side, s tar t ing as ear ly as 19 ms p r io r to the onset of saocades. Un l ike the long- lead burs t neurons in the b r a i n s t e m tha t show a g radua l bu i ld -up over 1 0 - 3 0 ms, the saccadic burs t neurons in the F O R show an a b r u p t onset , a regular d i scharge du r ing the burs t and a fair ly sharp offset which is fol lowed, in m a n y cases, by a depress ion. The du ra t ion of the bu r s t was highly cor re la ted with that of the a c c o m p a n y i n g saccade 6 These response features of the F O R neurons suggest tha t the ou tpu t of the ce rebe l lum is well def ined as to the du ra t ion of saccade and, therefore, it is su i tab le for regula t ing the
* Present address: Department of Ophthalmology. Sapporo Medical College, Sapporo, Japan. Correspondence: Dr. Hiroharu Noda, School of Optometry, Indiana University, 800 E Atwater, Bloomington, IN 47405, U.S.A.

291
amplitude of visually-directed saccades. Suppose this hypothesis is correct, alterations in the burst duration of FOR units would accompany some changes in the amplitude of saccades. In this study, we were able to alter the burst duration of FOR neurons by applying subthreshold stimulation to the oculomotor vermis and confirmed that the amplitude of the accompanying targeting saccade was changed. We present the evidence that a reduction in the burst duration of FOR neurons is accompanied by a hypometric saccade, supporting the hypothesis that cerebellar output carries the information for controlling the amplitude of saccades.
Experiments were conducted on 3 adolescent pig-tailed monkeys (Macaca nemestrina), weighing from 4 -5 kg. The procedures of training monkeys, surgical operations and experimental conditions for chronic single-unit recordings and microstimulation, histo- logical preparation of the brains, and the identification of recording and stimulating sites have already been described in detail 6. In brief, each monkey had been trained to make saccades to a visual target (red LED) which was flashed (for 25 ms) as soon as a fixation target was extinguished. Eye movements were recorded with magnetic search-coil after implanting the coil on one eye, using the method described by Judge et al. z. The stimulation site in the oculomotor vermis was determined in each experiment after recording saccade-related neuronal activity and finding the threshold minima for evoking saccades, using the same microelectrode. The variables of stimulation were negative 0.2-ms pulses applied at 600/s for 20-50 ms. Currents varied but were usually < 10 #A. The spike trains and the horizontal and vertical components of eye-position signals were digitized on a computer at 500/s. In measuring the burst duration of FOR neurons associated with saccades, the spikes preceding and following two consecutive interspike intervals, in which both instantaneous firing rates (reciprocal of interval) exceeded 150% of the spontaneous level, were considered as the first and the last spikes of the burst, respectively.
In a total of 98 saccadic burst units recorded from the FOR, there were 61 units (62. 2%) which were inhibited by microstimulation of the oculomotor vermis with current < 10 #A. The vermal stimulation evoked saccades with an ipsilateral horizontal compo- nent. The inhibition started with the onset of stimulation and outlasted the stimulation for a considerable period. The recovery from inhibition varied with the duration of stimulation. When a 50-ms pulse train was used, the period of complete inhibition following the cessation of stimulus ranged from 21 to 120 ms with an average 74.5 + 23.5 ms, measured in 21 FOR units.
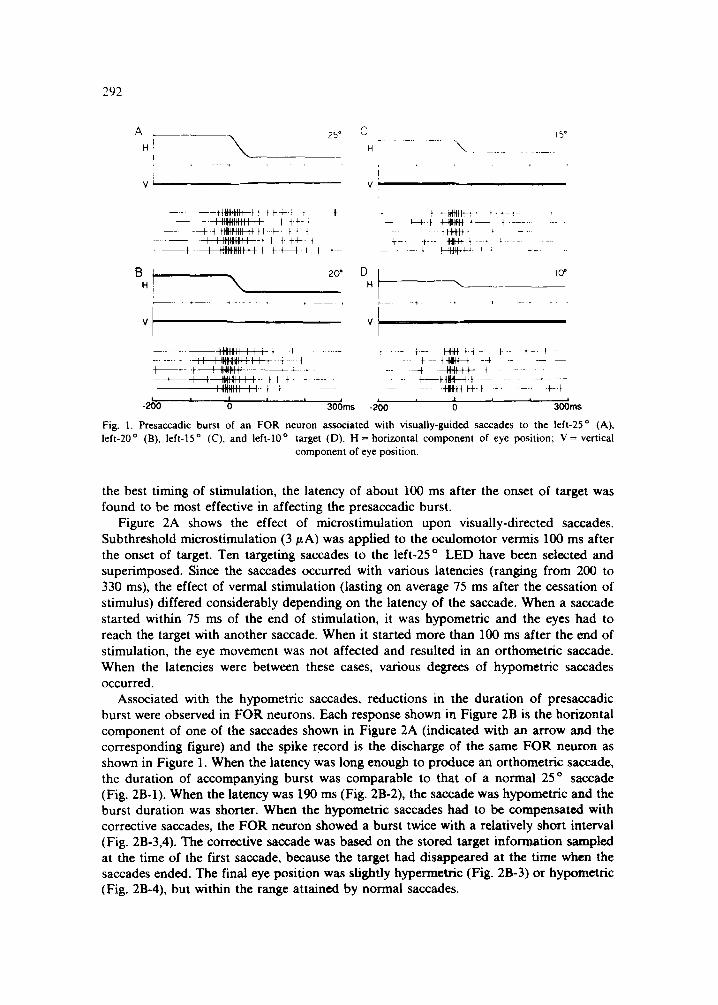
Figure 1 illustrates presaccadic bursts of an FOR unit during horizontal saccades from the central LED to one of the left-10 °, 15 °, 20 ° and 25 ° LEDs. The unit was recorded from the right FOR and the saccade direction for the optimal response was left-horizon- tal. The responses have been aligned with respect to the onset of saccades. The burst started approximately 20 ms prior to the saccade onset and the burst duration was highly correlated with the saccade duration, confirming the previous observation 6. As can be seen in the discharge of the neuron, a large saccade is associated with a longer burst but is not accompanied by a higher discharge rate. This observation is supported by the analysis of perisaccadic time histograms for this neuron (data not shown) as well as similar analyses for the other 97 saccadic burst units recorded from the FOR 6
In order to change the burst duration, the oculomotor vermis was stimulated during the latency period of visually-directed saccades. Since the purpose of the experiment was to alter the duration of the presaccadic burst and to test if the duration of the accompanying saccade is modified, the current was carefully kept subthreshold so that the stimulus itself did not cause any eye movements. After trials and errors, searching for
292
A ?-- H;
I
i
V ~
--\ 25* C
H . . . . . . , \ . I b *
I 1 . 1 1 1 , . ....... *{,,,,,,,,, } ~ t - - 4 - - ! :
. . . . . . . - - t - " ~ ' ~ ....... '~: I f - - - t - - ÷ ; ! • i i i i i t l i t l l i I t U ~ . l t . , i
- - ~ - i iiiiiiiiiii i i , .,,.0,,,,, ~ I-H-I t
BH.
÷ .. . .
2 0 "
i
V
t I I * I I H I . . . . . . . . , , . m . ', i i - ; 4 . . . . . . . . . . . . . . . . . . . . , i i l t b N u l ,
I t - - q ': . . . . . . . . . I I 1 '~ n i i i . . . . . . . . . ~ . . . . . . .
- ' I I " " " " ' , ' , ' , - ! t "~ . . . . . . . . . . . i n l l F I I I I t t l U . I I I I , . . . , , , , ',-- ~ ; . . . . . . . .
O0 l I I l l - 2 0 3 0 0 r n s
~--4- ~ = ~',$I . . . . .... ~¢~H-t ;
-t--- tt~4, ~ ..... , ,,#,!,~ ~_~. t
+ - - .
. . . . . . . [
D t ~o* H \
. . . . . . . + . . ~ . . . . . . . . .
I
. . . . . . . i - - HHPr ;-1 --- f . . . . . ~ ..... t -- --- ~ - - ~ 4 ~ 4 - - 1 ---+ . . . . . . . .
. . . . . -t" il~l~ ',+~ t . . . . . . . . . . . . t - - - - F t t ~ - - q ' - t . . . . . . . . . ~ " . . . . . .
. . . . . . . . . ~ H ~ ~- . . . . . . . . q---}-
- 2 0 3 0 0 m s
F i g . 1. P r e s a c c a d i c b u r s t o f a n F O R n e u r o n a s s o c i a t e d w i t h v i s u a l l y - g u i d e d s a c c a d e s t o t h e l e f t - 2 5 ° ( A ) ,
l e f t - 2 0 ° (B) , l e f t - 1 5 ° (C) , a n d l e f t - 1 0 ° t a r g e t ( D ) . H = h o r i z o n t a l c o m p o n e n t o f e y e p o s i t i o n ; V = v e r t i c a l
c o m p o n e n t o f e y e p o s i t i o n .
the best timing of stimulation, the latency of about 100 ms after the onset of target was found to be most effective in affecting the presaccadic burst.
Figure 2A shows the effect of microstimulation upon visually-directed saccades. Subthreshold microstimulation (3 ~tA) was applied to the oculomotor vermis 100 ms after the onset of target. Ten targeting saccades to the left-25 o LED have been selected and superimposed. Since the saccades occurred with various latencies (ranging from 200 to 330 ms), the effect of vermal stimulation (lasting on average 75 ms after the cessation of stimulus) differed considerably depending on the latency of the saccade. When a saccade started within 75 ms of the end of stimulation, it was hypometric and the eyes had to reach the target with another saccade. When it started more than 100 ms after the end of stimulation, the eye movement was not affected and resulted in an orthometric saccade. When the latencies were between these cases, various degrees of hypometric saccades occurred.
Associated with the hypometric saccades, reductions in the duration of presaccadic burst were observed in FOR neurons. Each response shown in Figure 2B is the horizontal component of one of the saccades shown in Figure 2A (indicated with an arrow and the corresponding figure) and the spike record is the discharge of the same FOR neuron as shown in Figure 1. When the latency was long enough to produce an orthometric saecade, the duration of accompanying burst was comparable to that of a normal 25 o saccade (Fig. 2B-l). When the latency was 190 ms (Fig. 2B-2), the saccade was hypometric and the burst duration was shorter. When the hypometric saccades had to be compensated with corrective saccades, the FOR neuron showed a burst twice with a relatively short interval (Fig. 2B-3,4). The corrective saccade was based on the stored target information sampled at the time of the first saccade, because the target had disappeared at the time when the saccades ended. The final eye position was slightly hypermetric (Fig. 2B-3) or hypometric (Fig. 2B-4), but within the range attained by normal saccades.

293
A
H
V
25*
200 m sec
[3
H H
, ~ s . . . . ~ . . . . . .
, - . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
I
Fig. 2. (A) Effect of subthreshold microstimulation (3 ~A) of the oculomotor visually-guided saccades executed from the central LED to the left-25 o LED. Note that the latency of saccade varied from 200 to 330 ms and that the effect of oculomotor vermis stimulation depended on the latency of saccade. The numbers indicating saccades correspond to the responses shown in B with the same numbers. (B) Changes in the duration of presaccadic burst of the same FOR unit as shown in Fig. 1, associated with the alterations in saccade duration caused by oculomotor vermis stimulation. H = horizontal component of eye position; V = velocity of the horizontal component. S = stimulation of the oculomotor vermis. T = target presentation. V = vertical compo-
nent of eye position. The bracket above spike records indicates the duration of presaccadic burst.
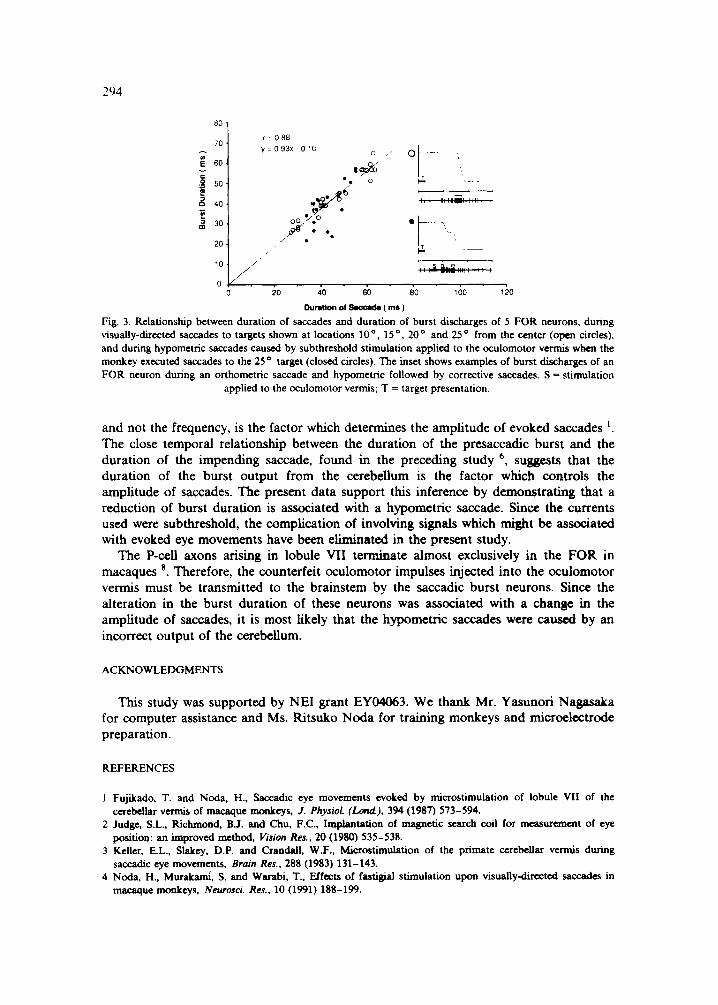
F igure 3 i l lustrates the re la t ionship be tween the burs t du ra t i on of F O R neurons and the saccade du ra t i on by compi l ing the da t a f rom 5 exper iments . Open circles represent the cont ro l da t a o b t a i n e d dur ing o r thomet r i c saccades to targets p resen ted at loca t ions 10 °, 15 °, 20 °, and 25 ° f rom the center, wi thout the cond i t ion ing s t imula t ion to the ocu lomoto r vermis. Closed circles represent the exper imenta l da t a in which saccades to 25 o target became hypomet r i c when the vermal s t imula t ion was app l i ed p r io r to the onset of the burst . When a saccade was small and the target had to be a t t a ined by a correc t ive saccade, the burs t du ra t ion for the correc t ive saccade was also p lo t t ed as ano the r point . W h e n the burs t du ra t ion was reduced by vermal s t imula t ion , the du ra t i on of the a c c o m p a n y i n g saccade was also shorter , ind ica t ing that the saccade was hypomet r ic .
The present exper imen t has shown that sub th resho ld mic ros t imu la t ion of vermai lobule VII ( the ocu lomo to r vermis) causes a con t ra la te ra l ly d i rec ted saccade to be hypomet r ic . This result is cons is ten t with the obse rva t ion of Kel le r et al. 3 that , when the pos te r ior vermis was s t imula ted dur ing saccades, con t ra la te ra l ly d i rec ted saccades were s lowed from their no rma l t ra jec tory and were made hypomet r ic . Our s tudy 4 also showed that, when the eyes were moved before or dur ing a ta rget ing saccade by s t imula t ion of the cerebel lum, the resul t ing eye movemen t was a lways dysmet r ic : ips i la te ra l ly d i rec ted saccades were m a d e hyperme t r i c and con t ra la te ra l saccades were hypomet r ic . Micro- s t imula t ion of the cerebe l lum has shown that the du ra t i on of the repet i t ive s t imula t ion ,
294
80,
70.
r: 60, v
-~ 50.
/~ 40.
m ~ 30,
r = 0 8 8
y = O 9 3 x - O ~ C c 0 ...... . . .,
2 0 . , . . / " . _ _ _
1 0 , /
o /" o ~o ~o ~ ~o ~6o ~o
l~ratlo, of ~ ( ms )
Fig. 3. Relationship between duration of saccades and duration of burst discharges of 5 FOR neurons, during visually-directed saccades to targets shown at locations 10 o 15 o 20 o a n d 25 o from the center (open circles), and during hypometric saccades caused by subthreshold stimulation applied to the oculomotor vermis when the monkey executed saccades to the 25 o target (closed circles). The inset shows examples of burst discharges of an FOR neuron during an orthometric saccade and hypometric followed by corrective saccades. S = stimulation
applied to the oculomotor vermis; T = target presentation.
and not the frequency, is the factor which determines the amplitude of evoked saccades L The close temporal relationship between the duration of the presaccadic burst and the duration of the impending saccade, found in the preceding study 6, suggests that the duration of the burst output from the cerebellum is the factor which controls the amplitude of saccades. The present data support this inference by demonstrating that a reduction of burst duration is associated with a hypometric saccade. Since the currents used were subthreshold, the complication of involving signals which might be associated with evoked eye movements have been eliminated in the present study.
The P-cell axons arising in lobule VII terminate almost exclusively in the FOR in macaques 6. Therefore, the counterfeit oculomotor impulses injected into the oculomotor vermis must be transmitted to the brainstem by the saccadic burst neurons. Since the alteration in the burst duration of these neurons was associated with a chan[le in the amplitude of saccades, it is most likely that the hypometric saccades were caused by an incorrect output of the cerebellum.
ACKNOWLEDGMENTS
This study was supported by NEI grant EY04063. We thank Mr. Yasunori Nagasaka for computer assistance and Ms. Ritsuko Noda for training monkeys and microelectrode preparation.
REFERENCES
1 Fujikado. T. and Noda, H., Saccadic eye movements evoked by microstimulation of Iobule VII of the cerebellar vermis of macaque monkeys, J. PhysioL (Lond), 394 (1987) 573-594.
2 Judge, S.L., Richmond, B.J. and Chu, F.C., Implantation of magnetic search coil for measurement of eye position: an improved method, Vision Res., 20 (1980) 535-538.
3 Keller, E.L., Slakey, D.P. and Crandall, W.F., Mierostimulation of the primate cerebellar vermis during saccadic eye movements, Brain Res., 288 (1983) 131-143.
4 Noda, H., Murakami, S. and Warabi, T., Effects of fastigial stimulation upon visually-directed saccades in macaque monkeys, NeuroscL Res., 10 (1991) 188-199.

295
5 Noda, H. and Fujikado, T., Topography of the oculomotor area of the cerebellar vermis in macaques as determined by microstimulation, J. Neurophysiol., 8 (1987) 359-378.
60htsuka, K. and Noda, H., Saccadic burst neurons in the oculomotor region of the fastigial nucleus of macaque monkeys, J. Neurophyswl., (1991) in press.
70htsuka, K. and Noda, H., Direction-selective saccadic-burst neurons in the fastigial oculomotor region of the macaque, Exp. Brain Res., 81 (1990) 659-662.
8 Yamada, J. and Noda, H., Afferent and efferent connections of the oculomotor cerebellar vermis in the macaque monkey, J. Comp. Neurol., 265 (1987) 224-241.





























