Embed Size (px)
Citation preview

R E S E A R C H P A P E R
The effect of three anaesthetic protocols on the stress
response in cane toads (Rhinella marina)
Sandra E Hernandez, Conrad Sernia & Adrian J Bradley
School of Biomedical Sciences, The University of Queensland, Brisbane, Queensland, Australia
Correspondence: Sandra E Hernandez, School of Biomedical Sciences, The University of Queensland, Brisbane, 4072 Queensland, Australia.
E-mail: [email protected]
Abstract
Objective Three anaesthetics (MS222, clove oil and
a mixture of ketamine/diazepam) were administered
to cane toads to determine their effect on the
hypothalamic-pituitary-adrenal (HPA) axis. Time to
induction and recovery and any adverse events
were also evaluated.
Study design Prospective randomized experimental
trial.
Animals Thirty adult male cane toads (Rhinella
marina) with body mass ranging between 130 and
250 g were captured from the field.
Methods Three groups of 10 toads were anaesthe-
tized with ketamine (200 mg kg)1) and diazepam
(0.2 mg kg)1) by intramuscular injection, MS222
(3 g L)1) or clove-oil (0.3 mL L)1) both by immer-
sion. Blood samples were collected to determine
plasma corticosterone concentrations. Induction
and recovery time were recorded in each treatment.
After full recovery animals were euthanized and a
complete post-mortem examination was performed.
Results Significant differences were found in the
activation of the HPA axis and in the times of
induction and recovery between treatments
(p < 0.001). Animals anaesthetized with clove-oil
had the highest levels of corticosterone in plasma
(42.5 ± 21.6 ng mL)1). No differences were found
between ketamine/diazepam (15.0 ± 13.3 ng mL)1)
and MS222 (22.0 ± 13.6 ng mL)1) groups. The
mean ± SD induction (minutes) and recovery
(hours) times respectively were; ketamine/diazepam
66.5 ± 11 and 8 ± 3, clove oil 39 ± 12 and 7.6 ± 3,
and MS222 42.5 ± 11 and 1.5 ± 0.5. Clove oil
exposure had 30% mortality. Death followed a
period of respiratory distress with changes consistent
with non-cardiogenic oedema observed at post-
mortem examination.
Conclusions and Clinical relevance Based on shorter
induction and recovery times and minimal
activation of HPA, MS222 is the anaesthetic of
choice in cane toads. If it is not possible to use
immersion methods of anaesthesia, ketamine/
diazepam can be used but induction and recovery
times are prolonged. Clove oil had unacceptable
mortality in this study and should be used with
extreme caution.
Keywords anaesthesia, cane toads, clove-oil, keta-
mine, MS222, stress.
Introduction
Many laboratory or field interventions require the
use of anaesthetics to reduce distress and pain in
animals. An ideal anaesthetic will provide anaes-
thesia (loss of consciousness and reduced reflex
response), analgesia and amnesia for the course of
the intervention. In the case of field anaesthesia it is
vital that induction and recovery times are short or
that the anaesthetic can be easily antagonized.
1
Veterinary Anaesthesia and Analgesia, 2012 doi:10.1111/j.1467-2995.2012.00753.x

Since it is difficult to assess pain and distress by
direct observation in amphibians, mediators of pain
or stress that are released commonly in vertebrates
may be used as markers. The level of activation of the
hypothalamic-pituitary-adrenal (HPA) axis is con-
sidered an acceptable method of assessing the
response of an organism to a stressor (Sapolsky et al.
2000). As part of the early stress response catechol-
amines (epinephrine and norepinephrine), followed
soon after by glucocorticoids (cortisol and corticoste-
rone) usually increase in concentration in the blood
(McEwen & Wingfield 2003). It is common to use
glucocorticoids as markers of stress, because the
plasma concentration of this class of adrenocortical
hormones increases in response to handling, surgical
intervention, or anaesthesia (Bentson et al. 2003). In
amphibians, plasma corticosterone concentration is
a good indicator of physiological or environmen-
tally induced stress since it can be related to the
stage of development (Krug et al. 1983; Wright
et al. 2003), seasonal changes (Mukherji 1968;
Piezzi & Burgos 1968), level of activity and energy
expenditure (Emerson & Hess 2001; Romero et al.
2004), and to anthropogenic stressors (Hopkins
et al. 1999). However, no studies have been per-
formed to determine the effect of general anaesthe-
sia on the HPA axis in amphibians.
Anaesthesia in amphibians can be achieved by
injection, inhalation, topical application, or immer-
sion (dissolving the agent in water and placing
the animal in the solution) (Wright 2001b,c). Top-
ical and immersion administration are the most
commonly used, with MS222 (Tricaine methanesulf-
onate [3-aminobenzoic acid ethyl ester methanesul-
fate]) as the preferred choice (Wright 2001b,c) and
having the American Veterinary Medicine Associa-
tion (AVMA) and Australian approval (Haskell et al.
2004; Gentz 2007). Clove oil has been mentioned as a
possible alternative anaesthetic in amphibians (Gentz
2007). Clove oil and its principal chemical component
eugenol (4-allyl-2-methoxyphenol) have been used
regularly in human dentistry because of their local
anaesthetic and antiseptic properties (Sticht & Smith
1971). Clove oil has been used for chemical restraint in
amphibians, where it is reported to have anaesthetic
and analgesic activity (Guenette et al. 2007; Mitchell
et al. 2009; Goulet et al. 2010). Apart from its
chemical properties, clove oil is also attractive because
of its low cost and wide availability (Keene et al. 1998;
Walsh & Pease 2002; Iversen et al. 2003).
Ketamine, an N-methyl-D-aspartate (NMDA)
antagonist that produces dissociative anaesthesia,
has been reported as an option in amphibians with
diverse results (Wright 2001c). Used alone it gives
poor muscle relaxation and therefore is normally
given in combination with benzodiazepines. Diaze-
pam is most commonly reported in amphibians
(Wright 2001b) and gives central muscle relaxation
and additional tranquilization. Previous reports
show that while a number of topical and injectable
preparations have been used as anaesthetics in
amphibians, there has not been a systematic study
of their comparative effectiveness which includes
physiological parameters of stress. Knowledge of the
extent to which the chosen chemical restraint is
exacerbating the stress response, and whether it is
provoking secondary effects, is extremely important
in order to assess the physiological impact of general
anaesthesia on an animal.
In this study the effect of three anaesthetic proto-
cols on the stress response in cane toads (Rhinella
marina) was assessed. Plasma corticosterone was
measured to evaluate the effects of anaesthesia on the
HPA. Time to induction and recovery of anaesthesia
were also assessed, as were any adverse reactions
to the anaesthetic agents. The overall objective of
the study was to form conclusions regarding the
relative suitability of the agents for anaesthesia of
cane toads and potentially other amphibians.
Material and methods
Animal subjects
All procedures were approved by The University of
Queensland Animal Ethics Committee (SBMS/437/
09/URG/GOVTMEX/HSF/CFOC).
A total of 30 male cane toads Rhinella marina
weighing more than 100 g (range 130–250 g),
were captured from The University of Queensland
Lakes, St Lucia, SE Queensland, Australia. Toads
were housed individually with a wet bed, a depth of
2 cm of water and a dry refuge area containing a
section of poly-vinyl-chloride pipe (120 · 300 mm).
An ambient temperature of 22–24 �C was main-
tained with a light cycle of 12 hours of dark:
12 hours of light. All animals were fasted for
24 hours before exposure to the anaesthetics.
The experimental protocol consisted of three
groups of ten toads. Toads were assigned randomly
to a group as they were captured. The first group
received a dose of 200 mg kg)1 of ketamine
(Ketamil; Troy laboratories Pty Ltd., Australia)
combined with 0.2 mg kg)1 of diazepam (Pamlin;
Three anaesthetic protocols in cane toads SE Hernandez et al.
� 2012 The Authors. Veterinary Anaesthesia and Analgesia2 � 2012 Association of Veterinary Anaesthetists and the American College of Veterinary Anesthesiologists

Parnel Pty Ltd., Australia), administered intramus-
cularly (IM.) in the hind leg (semimembranosus)
(Wright 2001b,c). They were placed in a dry
container to avoid drowning during the induction
period. The containers utilized to maintain and
induce anaesthesia were opaque plastic containers
of 4 L in capacity with lids perforated to facilitate
ventilation. The second group was immersed in
3 g L)1 of MS222 (SIGMA-Aldrich, Ethyl 3-amino-
benzoate methanesulfonate 98%, MO, pH7) (Wright
2001b,c). The third group was immersed in
0.3 mL clove oil L)1 water (Wright 2001b). Clove
oil is not soluble in water at low temperature
(<15 �C), therefore it was prepared as a stock of
3 mL of clove oil (Biotech pharmaceuticals Pty Ltd.,
Australia) in 10 mL of ethanol (ETOH 90%). The
quantity of ethanol used to solubilise clove oil in this
study was 100 times lower than concentrations
reported in the literature (Keene et al. 1998; Ross &
Ross 2008) therefore no secondary effect attributable
to ethanol was expected. In Bufo sp. the inguinal area
has the highest capacity for transcutaneus absorp-
tion (Ogushi et al. 2010), so the volume of water
used to induce the animals was enough to submerge
a third of their height. Furthermore maintaining
the animals in a low volume of water facilitated the
introduction of a sponge or a piece of fabric to
the container enabling the head to be kept clear of
the water once the righting reflex was abolished.
Surgical procedure and evaluation of anaesthesia
Time of induction of anaesthesia was defined as the
time from the administration of the anaesthetic to
the time the animal lost righting reflex, palpebral
reflexes and involuntary muscular movement.
Righting reflex was tested placing the animal in the
dorsal position and observing whether the animal
tried to return to the ventral position. Palpebral
reflex was tested by gently tapping the palpebral
area with the finger and observing eyelid move-
ment. Involuntary muscular movement was tested
via the withdrawal reflex, by grasping each limb
distally and extending it. Patellar junction and tar-
sal junctions were tapped with a light object to test
myotactic reflex (Wright 2001c; Wright et al.
2001). Reflexes were assessed every 5 minutes
during induction. When the appropriate level of
anaesthesia was achieved, nociceptive withdrawal
was tested by applying a hard pinch on the inter-
digital skin in the hind limbs, inguinal area and
ventral area using straight blunt tipped tweezers.
A two centimetre incision was then made in the
skin below the xiphoid process in the abdominal
midline to expose the abdominal muscles and
ventral vein. A catheter 24 gauge · 19 mm was
inserted into the vein (Introcan Certo, Braun,
Germany). The catheter was fixed by suturing the
connecting hub to the pectoral muscles and sutur-
ing the skin around it, leaving the end of the
catheter with the cap exposed for blood sampling.
Catheters were placed to facilitate blood sampling
and to allow humane euthanasia at the end of the
study period. Once the placement of the catheter
was completed a blood sample (500 lL) was
collected into heparin treated syringes (Wright
2001a). The collected blood was placed into Eppen-
dorf tubes and centrifuged at 1,300 g for 5 minutes
to separate plasma from blood cells. Plasma was
stored at )20 �C for later corticosterone analysis.
Following surgery and blood collection flunixin
meglumine 1 mg kg)1 IM (Finadyne; Schering-
Plough Pty Ltd., Australia) an analgesic and
non-steroidal anti-inflammatory, was administered
IM in the front leg (deltoid muscle) (Wright 2001b).
Animals were then rinsed with fresh water and
returned to their cages to recover. Reflexes in
animals were monitored every 5 minutes for 1 hour
to determine recovery time. Reflex testing was then
conducted every 30 minutes for the next 2 hours
and finally every hour until recovery. A full
recovery was defined by the appearance of the
righting reflex and a return of ambulatory behav-
iour defined as the animal performing purposeful
and coordinated movement. After full recovery, all
animals were euthanized with 100 mg kg)1 of
pentobarbital (Lethabarb; Virbac Pty Ltd., Australia)
administered intravenously (IV) through the
catheter (Wright 2001b,c). After euthanasia all
animals were subject to a full post mortem exam-
ination. Special attention was paid to the examina-
tion of the respiratory, cardiovascular, hepatic and
renal systems.
Plasma corticosterone analysis
Plasma samples (20 lL) were assayed for cortico-
sterone by radioimmunoassay (Millis et al. 1999).
The tracer used was tritiated corticosterone (Corti-
costerone [1,2,6,7-3H] 2.59 TBq mmol)1; Perkin
Elmer Lab, MA, USA). Radioactivity was counted in
a liquid scintillation spectrometer (Beckman LS
6000 TA). For this study we used a polyclonal
antibody against corticosterone developed by
Three anaesthetic protocols in cane toads SE Hernandez et al.
� 2012 The Authors. Veterinary Anaesthesia and Analgesia� 2012 Association of Veterinary Anaesthetists and the American College of Veterinary Anesthesiologists 3
AbCam laboratories (Product Number: ab77798),
raised in sheep and based on corticosterone-
3-cmo-urease as immunogen. The cross-reactions
documented were: 0.67% to 11-dehydrocorticos-
terone, 1.5% to deoxycorticosterone, and <0.01% to
18-OH-DOC, cortisone, cortisol and aldosterone.
The plasma samples were measured in duplicate
and the corticosterone concentration was calculated
from a standard curve, as described by Dudley
(Dudley et al. 1985).
Statistical analysis
The D’Agostino-Pearson omnibus test demonstrated
that the variables had a normal distribution
(p > 0.05). ANOVA analysis was used to determine
differences between treatments, followed by a Tukey
test to compare group means. The null hypothesis
was rejected at p < 0.05. The data were expressed
as mean ± SD. All statistical analyses were con-
ducted using GraphPad Prism (GraphPad Software,
CA, USA, 2009).
Results
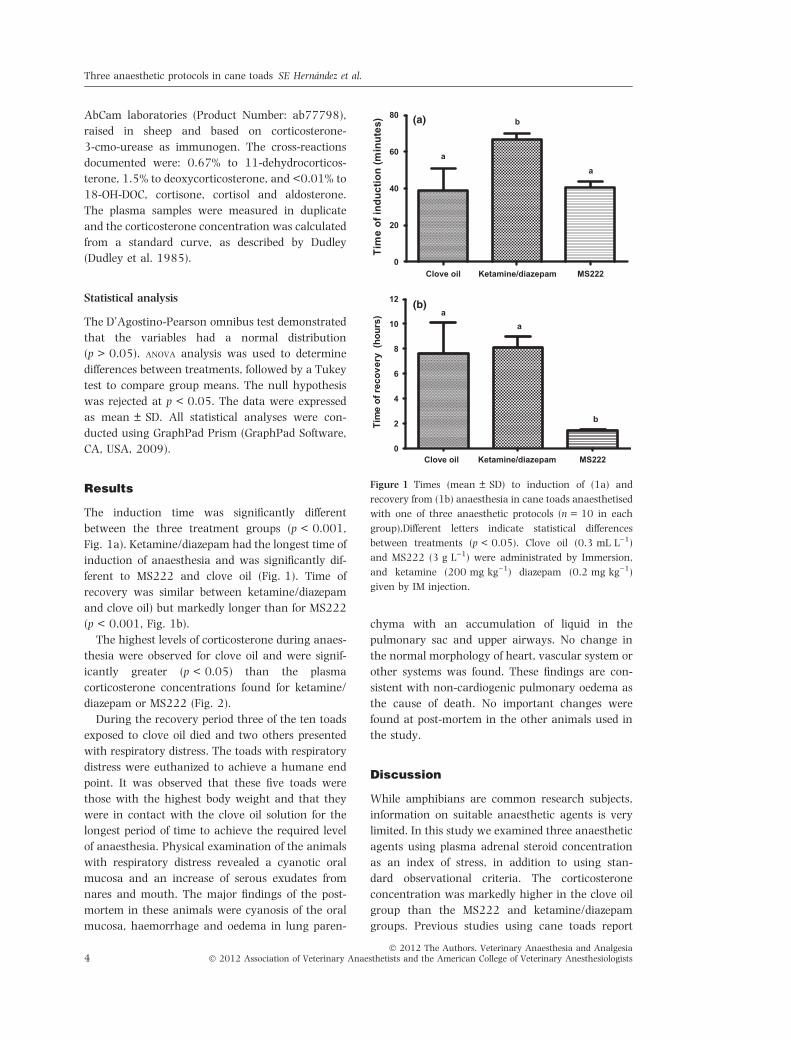
The induction time was significantly different
between the three treatment groups (p < 0.001,
Fig. 1a). Ketamine/diazepam had the longest time of
induction of anaesthesia and was significantly dif-
ferent to MS222 and clove oil (Fig. 1). Time of
recovery was similar between ketamine/diazepam
and clove oil) but markedly longer than for MS222
(p < 0.001, Fig. 1b).
The highest levels of corticosterone during anaes-
thesia were observed for clove oil and were signif-
icantly greater (p < 0.05) than the plasma
corticosterone concentrations found for ketamine/
diazepam or MS222 (Fig. 2).
During the recovery period three of the ten toads
exposed to clove oil died and two others presented
with respiratory distress. The toads with respiratory
distress were euthanized to achieve a humane end
point. It was observed that these five toads were
those with the highest body weight and that they
were in contact with the clove oil solution for the
longest period of time to achieve the required level
of anaesthesia. Physical examination of the animals
with respiratory distress revealed a cyanotic oral
mucosa and an increase of serous exudates from
nares and mouth. The major findings of the post-
mortem in these animals were cyanosis of the oral
mucosa, haemorrhage and oedema in lung paren-
chyma with an accumulation of liquid in the
pulmonary sac and upper airways. No change in
the normal morphology of heart, vascular system or
other systems was found. These findings are con-
sistent with non-cardiogenic pulmonary oedema as
the cause of death. No important changes were
found at post-mortem in the other animals used in
the study.
Discussion
While amphibians are common research subjects,
information on suitable anaesthetic agents is very
limited. In this study we examined three anaesthetic
agents using plasma adrenal steroid concentration
as an index of stress, in addition to using stan-
dard observational criteria. The corticosterone
concentration was markedly higher in the clove oil
group than the MS222 and ketamine/diazepam
groups. Previous studies using cane toads report
Clove oil Ketamine/diazepam MS2220
20
40
60
80
a
b
a
Tim
e of
indu
ctio
n (m
inut
es)
Clove oil Ketamine/diazepam MS2220
2
4
6
8
10
12a
a
bTim
e of
reco
very
(ho
urs)
(a)
(b)
Figure 1 Times (mean ± SD) to induction of (1a) and
recovery from (1b) anaesthesia in cane toads anaesthetised
with one of three anaesthetic protocols (n = 10 in each
group).Different letters indicate statistical differences
between treatments (p < 0.05). Clove oil (0.3 mL L)1)
and MS222 (3 g L)1) were administrated by Immersion,
and ketamine (200 mg kg)1) diazepam (0.2 mg kg)1)
given by IM injection.
Three anaesthetic protocols in cane toads SE Hernandez et al.
� 2012 The Authors. Veterinary Anaesthesia and Analgesia4 � 2012 Association of Veterinary Anaesthetists and the American College of Veterinary Anesthesiologists

concentrations of corticosterone in plasma ranging
between 5–20 ng mL)1 (Orchinik et al. 1988).
These concentration are similar to those found in
the group anaesthetized with ketamine/diazepam
and MS222 but are approximately half of those
found when clove oil was used to induce anaes-
thesia. An increase in glucocorticoid concentration
in response to anaesthesia is an observation com-
mon in fish (Sink et al. 2007), dog (Fox et al. 1998)
and horses (Taylor 1991) and this study confirms
that it also occurs in the cane toad and presumably
in amphibians in general. Our observations with
clove oil are mirrored by a study in rainbow trout by
Sink et al. (2007) that reported clove oil to be less
effective than MS222 in reducing stress-induced
cortisol elevation. In addition, there was evidence of
a greater mortality in field-handled fish following
clove oil anaesthesia. These authors suggested that
the stress and higher mortality were due to the
irritant nature of clove oil odour, and possibly to
poor anaesthetic properties which rendered the fish
immobile without loss of consciousness (Sink et al.
2007). Data from rat nerve-muscle preparations
show that eugenol, the main component of clove oil,
is effective in blocking neuromuscular transmission
at low concentrations, thereby supporting the view
that clove oil may be causing neuromuscular
paralysis without anaesthesia (Ingvast-Larsson
et al. 2003). Despite these reservations eugenol can
cross the blood-brain barrier and reduce brain
activity when it is administered intraperitoneally in
rats (Sell & Carlini 1976). Furthermore IV admin-
istration of eugenol produces a reversible, dose-
dependent anaesthesia in male rats (Guenette et al.
2006). Therefore the route of administration is a
significant factor to consider since topical adminis-
tration would lead to paralysis while requiring a
much longer time for sedation and anaesthesia.
Studies on fish have shown that, even though the
primary route of adsorption of anaesthetic dissolved
in water is via the gills, the skin also plays an
important role. Studies in fish reported by Ferreira
et al. (1984) and Gilderhus (1989) showed that the
highest concentrations of anaesthetic are needed as
the size of the animal and skin thickness increases.
A report in amphibia (Xenopus laevis) exposed to
clove oil showed that body size influences induction
and recovery times (Goulet et al. 2010, 2011).
Young and smaller animals require shorter induc-
tion times and have a shorter surgical anaesthesia
period, compared with bigger animals (Goulet et al.
2010, 2011). Consequently age, body mass, body
surface area in contact with the water, and thick-
ness of the skin can affect the uptake of anaesthetics
diluted in water. In this study the effect of life cycle
was minimized by using toads with body weights of
more than 100 g, which is considered adult stage
(Zug et al. 2001).
The induction and recovery times found in the
present study were longer than those previously
reported for induction with MS222 (15–30 minutes)
(Wright 2001b,c; Ross & Ross 2008; Torreilles et al.
2009) and clove oil (15–30 minutes) (Lafortune
et al. 2000; Guenette et al. 2007; Goulet et al.
2010). Previous studies using MS222 and clove oil
were performed in aquatic amphibians completely
submerged in the water containing the anaesthetic.
In this study we used a species that spends most of its
life out of the water and its skin is thicker compared
with aquatic species. Furthermore the animals in this
study were not totally submerged in the water
containing the anaesthetic to avoid animals drown-
ing during the induction period. Therefore the total
skin area in contact with the anaesthetic was reduced
and in consequence this increased the time of
induction. The recovery time for MS222 was similar
to previous reports (40 minutes) (Wright 2001c;
Mitchell et al. 2009), but for clove oil (2–3 hours)
(Lafortune et al. 2000; Guenette et al. 2007; Goulet
et al. 2010) it was longer. One probable explanation
is that as the animals have to stay for longer in
contact with the anaesthetic, the recovery times in
the animals increase too. Therefore time of exposure
to clove oil seems to be another factor that needs
careful consideration. Moreover, the mortality rate in
the animals exposed to clove oil was 30%, increasing
to 50% after our decision to euthanase toads in
Clove oil Ketamine/diazepam MS2220
20
40
60
80
a
bb
Cor
ticos
tero
ne (n
g m
L–1 p
lasm
a)
Figure 2 Plasma levels of corticosterone (means ± SD) in
cane toads during anaesthesia induced with one of three
anaesthetic protocols.Different letters indicate statistical
differences between treatments (p < 0.05).
Three anaesthetic protocols in cane toads SE Hernandez et al.
� 2012 The Authors. Veterinary Anaesthesia and Analgesia� 2012 Association of Veterinary Anaesthetists and the American College of Veterinary Anesthesiologists 5

respiratory distress, which may will have failed to
recover if they had been left to do so.
Our observations on adverse reactions to clove oil
are supported by findings in frogs (Xenopus laevis)
exposed to eugenol (Goulet et al. 2011). Histopathol-
ogy on the respiratory system showed that clove oil
provoked hyperplasia of the epithelium around alve-
oli in the lung sacs and hyaline membranes in the
lung parenchyma. There is a report of interstitial
pulmonary haemorrhage and oedema, after tracheal
instillation of clove oil in rats and hamsters (LaVoie
et al. 1986). In humans the use of clove cigarettes
has been linked to pulmonary oedema (LaVoie et al.
1986; JAMA 1988), and there is a case report of non-
cardiogenic pulmonary oedema following IV injec-
tion of clove oil (Kirsch et al. 1990). The mechanism
by which clove oil induces inflammatory responses is
not clear. In vitro studies suggest the redirection of
arachidonic acid into the inflammatory pathways
involving prostaglandins, thromboxane and leuko-
trienes may be involved (Rasheed et al. 1984).
It is worth noting that among all the groups
ketamine/diazepam had the longest period of induc-
tion of anaesthesia. This finding is similar to those
reported for other amphibian studies (30–60 min-
utes) (Wright 2001b,c), even though our dose was
higher. In the case of recovery, ketamine/diazepam
and clove oil have the longest periods of recovery, a
finding consistent with previous reports (18 hours)
(Wright 2001c; Mitchell et al. 2009).
In conclusion MS222 anaesthesia of cane toads is
superior to ketamine/diazepam or clove oil anaes-
thesia due to shorter induction and recovery times,
and minimal activation of the HPA axis. If it is not
practical to perform immersion anaesthesia, a
combination of ketamine and diazepam can be used
IM. Induction and recovery times are prolonged
when using this method, making it unsuitable for
field use. However it appears to be no more stressful
than MS222 anaesthesia. Clove oil administration
in cane toads caused adverse changes in the
respiratory system leading to an unacceptable high
mortality rate. Therefore clove oil should be used
with extreme caution until further investigations
into the mode of action, general toxicity, and routes
of administration have been performed.
Acknowledgements
The National Council of Science and Technology
(CONACYT), Mexico for providing scholarship
support to SEH.
References
Bentson K, Capitanio J, Mendoza S (2003) Cortisol
responses to immobilization with Telazol or ketamine in
baboons (Papio cynocephalus/anubis) and rhesus maca-
ques (Macaca mulatta). J Med Primatol 32, 148–160.
Dudley RA, Edwards P, Ekins RP et al. (1985) Guidelines
for immunoassay data processing. Clin Chem 31, 1264–
1271.
Emerson SB, Hess DL (2001) Glucocorticoids, androgens,
testis mass, and the energetics of vocalization in breed-
ing male frogs. Horm Behav 39, 59–69.
Ferreira JT, Schoonbee HJ, Smit GL (1984) The uptake of
the anaesthetic benzocaine hydrochloride by the gills
and the skin of three freshwater fish species. J Fish Biol
25, 35–41.
Fox SM, Mellor DJ, Lawoko CRO et al. (1998) Changes in
plasma cortisol concentrations in bitches in response to
different combinations of halothane and butorphanol,
with or without ovariohysterectomy. Res Vet Sci 65,
125–133.
Gentz EJ (2007) Medicine and surgery of amphibians.
ILAR 48, 255–259.
Gilderhus PA (1989) Efficacy of benzocaine as an anes-
thetic for salmonid fishes. North Am J Fish Manag 9,
150–153.
Goulet F, Helie P, Vachon P (2010) Eugenol anesthesia in
African clawed frogs (Xenopus laevis) of different body
weights. J Am Assoc Lab Anim Sci 49, 460–463.
Goulet F, Vachon P, Helie P (2011) Evaluation of the
toxicity of eugenol at anesthetic doses in African clawed
frogs (Xenopus laevis). Toxicol Pathol 39, 471–477.
Guenette SA, Beaudry F, Marier JF et al. (2006) Pharma-
cokinetics and anesthetic activity of eugenol in male
Sprague-Dawley rats. J Vet Pharmacol Ther 29, 265–
270.
Guenette SA, Helie P, Beaudry F et al. (2007) Eugenol for
anesthesia of African clawed frogs (Xenopus laevis). Vet
Anaesth Analg 34, 164–170.
Haskell SRR, Payne MA, Webb AI et al. (2004) Current
approved drugs for aquatic species. J Am Vet Med Assoc
224, 50–51.
Hopkins WA, Mendonca MT, Congdon JD (1999)
Responsiveness of the hypothalamo-pituitary-interrenal
axis in an amphibian (Bufo terrestris) exposed to coal
combustion wastes. Comp Biochem Physiol C Pharma-
col Toxicol Endocrinol 122, 191–196.
Ingvast-Larsson JC, Axen VC, Kiessling AK (2003) Effects
of isoeugenol on in vitro neuromuscular blockade of rat
phrenic nerve-diaphragm preparations. Am J Vet Res
64, 690–693.
Iversen M, Finstad B, McKinley RS et al. (2003) The
efficacy of metomidate, clove oil, Aqui-S(TM) and
Benzoak� as anaesthetics in Atlantic salmon (Salmo
salar L.) smolts, and their potential stress-reducing
capacity. Aquaculture 221, 549–566.
Three anaesthetic protocols in cane toads SE Hernandez et al.
� 2012 The Authors. Veterinary Anaesthesia and Analgesia6 � 2012 Association of Veterinary Anaesthetists and the American College of Veterinary Anesthesiologists

JAMA (1988) Evaluation of the health hazard of clove
cigarettes. Council on scientific affairs. JAMA 260,
3641–3644.
Keene JL, Noakes DLG, Moccia RD et al. (1998) The
efficacy of clove oil as an anaesthetic for rainbow trout,
Oncorhynchus mykiss (Walbaum). Aquac Res 29, 89–
101.
Kirsch CM, Yenokida GG, Jensen WA et al. (1990) Non-
cardiogenic pulmonary oedema due to the intravenous
administration of clove oil. Thorax 45, 235–236.
Krug EC, Honn KV, Battista J et al. (1983) Corticosteroids
in serum of Rana catesbeiana during development and
metamorphosis. Gen Comp Endocrinol 52, 232–241.
Lafortune M, Mitchell MA, Smith JA (2000) Evaluation of
medetomidine, clove oil and propofol for anesthesia of
leopard frogs, Rana pipiens. J Herpe Med Surg 11, 13–18.
LaVoie EJ, Adams JD, Reinhardt J et al. (1986) Toxicity
studies on clove cigarette smoke and constituents of
clove: determination of the LD50 of eugenol by intra-
tracheal instillation in rats and hamsters. Arch Toxicol
59, 78–81.
McEwen BS, Wingfield JC (2003) The concept of allostasis
in biology and biomedicine. Horm Behav 43, 2–15.
Millis AL, Taggart DA, Bradley AJ et al. (1999) Repro-
ductive biology of the brush-tailed phascogale, Phasco-
gale tapoatafa (Marsupialia: Dasyuridae). J Zool 248,
325–335.
Mitchell MA, Riggs SM, Singleton CB et al. (2009)
Evaluating the clinical and cardiopulmonary effects of
clove oil and propofol in tiger salamanders (Ambystoma
tigrinum). J Exot Pet Med 18, 50–56.
Mukherji M (1968) Adrenal cortex in stress and hiberna-
tion–a histochemical study in toad (Bufo melanostictus).
Acta Histochem 29, 297–303.
Ogushi Y, Tsuzuki A, Sato M et al. (2010) The water-
absorption region of ventral skin of several semiterres-
trial and aquatic anuran amphibians identified by
aquaporins. Am J Physiol Regul Integr Comp Physiol
299, R1150–R1162.
Orchinik M, Licht P, Crews D (1988) Plasma steroid
concentrations change in response to sexual behavior in
Bufo marinus. Horm Behav 22, 338–350.
Piezzi RS, Burgos MH (1968) The toad adrenal gland. I.
Cortical cells during summer and winter. Gen Comp
Endocrinol 10, 344–354.
Rasheed A, Laekeman G, Totte J et al. (1984) Eugenol
and prostaglandin biosynthesis. N Engl J Med 310, 50–
51.
Romero SM, Pereira AF, Garofalo MA et al. (2004) Effects
of exercise on plasma catecholamine levels in the toad,
Bufo paracnemis: role of the adrenals and neural control.
J Exp Zool A Comp Exp Biol 301, 911–918.
Ross LG, Ross B (2008) Anaesthesia of amphibians and
reptiles. In: Anaesthetic and Sedative Techniques for
Aquatic Animals, Ross LG, Ross B (eds). Institute
of Aquaculture, University of Stirling, Stirling, UK.
pp. 179–190.
Sapolsky RM, Romero LM, Munck AU (2000) How do
glucocorticoids influence stress responses? Integrating
permissive, suppressive, stimulatory, and preparative
actions. Endocr Rev 21, 55–89.
Sell AB, Carlini EA (1976) Anesthetic action of methyl-
eugenol and other eugenol derivatives. Pharmacology
14, 367–377.
Sink TD, Strange RJ, Sawyers RE (2007) Clove oil used at
lower concentrations is less effective than MS-222 at
reducing cortisol stress responses in anesthetized rain-
bow trout. North Am J Fish Manag 27, 156–161.
Sticht FD, Smith RM (1971) Eugenol: some pharmacologic
observations. J Dent Res 50, 1531–1535.
Taylor PM (1991) Current developments in understanding
stress responses to anaesthesia in horses. Veterinary
Anaesth Analg 18, 317–318.
Torreilles SL, McClure DE, Green SL (2009) Evaluation and
refinement of euthanasia methods for Xenopus laevis.
J Am Assoc Lab Anim Sci 48, 512–516.
Walsh CT, Pease BC (2002) The use of clove oil as an
anaesthetic for the longfinned eel, Anguilla reinhardtii
(Steindachner). Aquac Res 33, 627–635.
Wright KM (2001a) Amphibian hematology. In:
Amphibian Medicine and Captive Husbandry, Wright
KM, Whitaker BR (eds). Krieger Publishing company,
Malabar, FL, USA. pp. 129–146.
Wright KM (2001b) Amphibians. In: Exotic Animal
Formulary, Carpenter JW (ed.). Elsevier Saunders,
St. Louis, MO, USA. pp. 33–54.
Wright KM (2001c) Restraint techniques and euthanasia.
In: Amphibian Medicine and Captive Husbandry,
Wright KM, Whitaker BR (eds). Kreiger Publishing
Company, Malabar, FL, USA. pp. 111–122.
Wright KM, Brent R, Whitaker MS (2001) Clinical
techniques. In: Amphibian Medicine and Captive
Husbandry, Wright KM, Whitaker BR (eds). Krieger
Publishing Company, Malabar, FL, USA. pp. 89–110.
Wright ML, Guertin CJ, Duffy JL et al. (2003) Develop-
mental and diel profiles of plasma corticosteroids in
the bullfrog, Rana catesbeiana. Comp Biochem Physiol
A Mol Integr Physiol 135, 585–595.
Zug GR, Vitt JV, Caldwell JP (2001) Frogs. In: Herpetology
an Introductory Biology of Amphibians and Reptiles,
Zug GR, Vitt JV, Caldwell JP (eds). Academic Press, San
Diego, CA. pp. 389–390.
Received 26 July 2011; accepted 27 January 2012.
Three anaesthetic protocols in cane toads SE Hernandez et al.
� 2012 The Authors. Veterinary Anaesthesia and Analgesia� 2012 Association of Veterinary Anaesthetists and the American College of Veterinary Anesthesiologists 7





























