Embed Size (px)
Citation preview

i.
THE EFFECTS OF GLYCERYL TRINITRATE AND
OVARIECTOMY ON FEMORO-TIBIAL ARTICULAR
CARTILAGE, SYNOVIUM AND SUBCHONDRAL BONE IN
NORMAL AND OSTEOARTHRITIC EWES
Martin Andrew Cake BSc BVMS
This thesis is presented for the degree of Doctor of Philosophy of Murdoch University
2002

i.
I declare that this thesis is my own account of my research, and contains as its main content work which has not previously been submitted for degree at any
tertiary educational institution.
Martin Cake 8th February 2002

ii.
ABSTRACT Nitric oxide (NO) alters chondrocyte metabolism, and is thought to be a key catabolic mediator in
osteoarthritis. NO is also an important modifier of bone metabolism, and may partially mediate the
bone-sparing effects of oestrogen. Oestrogen has also been linked to the modulation of
osteoarthritis, though its role is not clear. The aim of this study was to examine the structural and
metabolic effects of ovariectomy and the NO donor glyceryl trinitrate (GTN) on (1) normal ovine
femoro-tibial joint tissues, and (2) the progression of joint lesions in the established ovine
meniscectomy model of osteoarthritis.
Preliminary investigations tested a novel computer-assisted histomorphometric method of assessing
osteochondral changes post-meniscectomy, in a trial of a putative disease-modifying osteoarthritis
compound. Quantitative assessment revealed a subtle protective effect not evident by qualitative
methods. These techniques were then used to test the experimental hypotheses in a combined trial
involving 48 aged ewes, variously subjected to ovariectomy, bilateral lateral meniscectomy, and/or
topical GTN therapy. At six months, joint tissues were analysed using histology, histomorphometry,
dynamic biomechanical testing, serum markers, bone densitometry, and tissue culture of synovial
fibroblasts and explants of cartilage and bone. Ovariectomy modified cartilage structure and
chondrocyte metabolism, and induced subchondral bone remodelling. Prior ovariectomy altered the
development of OA lesions post-meniscectomy, producing thicker but biomechanically inferior
cartilage and elevated metabolic activity in subchondral bone. GTN treatment of normal sheep
induced thinner, structurally-altered cartilage in normal sheep, and accentuated cartilage and
subchondral bone lesions post-meniscectomy. These results support an important homeostatic role
for oestrogen in joint tissues, and show that GTN, a commonly used angina therapy, can induce
structural alterations in joint tissues and potentially accelerate the progression of concurrent OA.
Results also advance understanding of the role of synovial and subchondral bone changes in the
pathogenesis of this OA model.

iii.
TABLE OF CONTENTS
Declaration i Abstract ii Table of Contents iii Index to Figures vi Index to Tables ix Publications and conference presentations resulting from this work x Acknowledgements xii List of Abbreviations xiii Preface xv
Chapter 1: INTRODUCTION 1 1.1 LITERATURE REVIEW
1.1.1 Osteoarthritis 1
1.1.1.1 Definition and classification of osteoarthritis 1 1.1.1.2 Aetiology and epidemiology of OA 2
1.1.2 Structure of normal and osteoarthritic joint tissues 3
1.1.2.1 Articular cartilage 3 1.1.2.1.1 Chondrocytes 4 1.1.2.1.2 Cartilage matrix 5 1.1.2.1.3 Cartilage changes in osteoarthritis 9
1.1.2.2 Synovial Membrane 12 1.1.2.2.1 Structure of normal synovium 12 1.1.2.2.2 Synovial changes in osteoarthritis 15 1.1.2.2 3 Synovium-cartilage interactions in osteoarthritis 16 1.1.2.2.4 Rheumatoid arthritis 18 1.1.2.2.5 Synovial fluid 18
1.1.2.3 Subchondral Bone 20 1.1.2.3.1 Normal structure of subchondral bone 20 1.1.2.3 2 Subchondral bone changes in osteoarthritis 21 1 1.2.3.3 Role of subchondral bone changes in the
pathogenesis of OA 23 1.1.2.3 4 Osteophytes 26 1.1.2.3.5 Markers of bone turnover in OA 27
1.1.3 Pathogenetic mediators of osteoarthritis 27
1.1.3.1 Enzymes and Enzyme Inhibitors 27 1.1.3.1.1 Matrix metalloproteases 28 1.1.3.1.2 Tissue inhibitor of matrix
metalloproteases (TIMP) 32 1 1.3.1.3 Serine and CysteineProteases 32
1.1.3.2 Cytokines 34 1.1.3.2.1 Interleukin-1 (IL-1) 35 1 1.3.2.2 Tumour Necrosis Factor- (TNF- ) 38 1.1.3.2.3 Interleukin-6 (IL-6) 39 1.1.3.2.4 Other cytokines 41 1 1 3.2 5 Growth Factors 42
1.1.3.3 Nitric Oxide 46 1.1.3.3.1 Nitric oxide and cartilage 48 1.1.3.3.2 Nitric oxide and synovium 50 1.1.3.3.3 Nitric oxide and bone 50
1 1.3.4 Eicosanoids 51

iv.
1.1.3.4.1 Effect of prostaglandin E2 on joint tissues 52 1.1.3.4.2 Interaction between nitric oxide and PGE2 54
1.1.4 Oestrogen in bone and joint homeostasis 55
1.1.4.1 Evidence linking oestrogens and arthritis 57 1.1.4.1.1 Clinical and epidemiological evidence 57 1.1.4.1.2 Experimental Evidence 59
1.1.4.2 Oestrogen and cartilage 60 1.1.4.3 Oestrogen and synovium 62 1.1.4.4 Oestrogen and bone 62
1.1.5 Animal models of osteoarthritis and osteoporosis 65
1.1.5.1 Animal models of OA 65 1.1.5.2 The ovine meniscectomy model of OA 67 1.1.5.3 The ovine ovariectomy model of postmenopausal osteoporosis 68
1.1.6 In vitro investigation of synovial tissue 70
1.1.6.1 Culturemethodsofsynovialtissueinvitro 70 1.1.6.2 Culturerequirementsofsynoviocytes 72 1.1.6.3 Differences between normal and arthritic synovium in culture 72
1.1.7 Pharmaceutical management of osteoarthritis 75
1.1.7.1 Non-steroidal anti-inflarnmatory drugs (NSAIDs) 75 1.1.7.2 Corticosteroids 79
1.1.7.3 Sulphated heparinoids 79 1.1.7.4 Hyaluronanpreparations 80
1.1.7.5 Oral supplements 81 1.1.7.6 Potential future therapies 82
1.2 STUDY BACKGROUND 84
1.3 AIMS AND HYPOTHESES 87
Chapter 2: DEVELOPMENT OF NOVEL METHODS OF
HISTOLOGIC ASSESSMENT OF CARTILAGE AND
SYNOVIUM, IN A TRIAL OF ORAL AVOCADO-SOYA
UNSAPONIFIABLES 89 2.1 INTRODUCTION 89
2.2 MATERIALS AND METHODS 91
2.2.1 Trial protocol 91
2.2.1.1 Source of animals 91 2.2.1.2 Meniscectomy 91 2.2.1.3 Trial protocol 93 2.2.1.4 Sacrifice and tissue processing 93
2.2.2 Gross pathology 94
2.2.3 Cartilage histopathological seoring 95
2.2.4 Histomorphometric image analysis 95
2.2.5 Synovial histopathological scoring 98
2.2.6 Statistical analysis 99
2.3 RESULTS 100
2.3.1 Body weight 100
2.3.2 Gross pathology 100
2.3.3 Cartilage histopathological scoring 100
2.3.4 Histomorphometric image analysis 101
2.3.5 Synovial histopathological scoring 102
2.4 DISCUSSION 111

v.
Chapter 3: EFFECTS OF GLYCERYL TRINITRATE, OVARIECTOMY,
AND MENISCECTOMY ON FEMORO-TIBIAL JOINT
TISSUES OF AGED EWES 116 3.1 INTRODUCTION 116
3.2 MATERIALS AND METHODS 118
3.2.1 Trial protocol 118
3.2.1.1 Source of animals 118 3.2.1.2 Surgery 119
3.2.1.2.1 Ovariectomy 119 3.2.1.2.2 Meniscectomy 119
3.2.1.3 Glyceryl trinitrate treatment 120 3.2.1.4 Animal maintenance and sampling 120 3.2.1.5 Sacrifice and tissue processing 121
3.2.2 Gross pathology 122
3.2.3 Dynamic biomechanical testing 122
3.2.4 Histological assessment 124 3.2.4.1 Cartilage histology 124 3.2.4.2 Collagen birefringence 125 3.2.4.3 Synovial histopathology 125 3.2.4.4 Undecalcified bone sections 126
3.2.5 Bone densitometry 128
3.2.6 Blood and urine assays 129
3.2.6.1 Plasma 17ß-oestradiol 129 3.2.6.2 Serum osteocalcin 130 3.2.6.3 Urine collagen crosslinks 130
3.2.7 Cell culture procedures and assays on conditioned media (CM) 131
3.2.7.1 Patellar cartilage explant culture 132 3.2.7.2 Subchondral bone explant culture 132 3.2.7.3 Synovial fibroblast culture 133 3.2.7.4 DNA assay 135 3.2.7.5 Nitric oxide 136 3.2.7.6 MMP activity 136 3.2.7.7 35SO4-labelled proteoglycan synthesis 137 3.2.7.8 PGE2 138 3.2.7.9 Urokinase-type plasminogen activator (uPA) activity 138 3.2.7.10 IL-6 bioassay 139 3.2.7.11 IL-1 activity (lymphocyte proliferation assay) 140
3.2.8 Oestrogen receptor assay 141
3.2.9 Statistical methods 142
3.3 RESULTS 143
3.3.1 Body weights 143
3.3.2 Gross pathology 143
3.3.3 Biomechanical testing 144
3.3.4 Histological assessments 145
3.3.4.1 Cartilage histology 145 3.3.4.1.1 Cartilage histopathological scores 145 3.3.4.1.2 Computerised histomorphometry 147
3.3.4.2 Collagen birefringence 149 3.3.4.3 Synovial histopathological scores 149 3.3.4.4 Undecalcified bone histomorphometry 150

vi.
3.3.5 Bone mineral density 150
3.3.6 Blood and urine markers 151
3.3.6.1 Plasma 17ß-oestradiol 151 3.3.6.2 Serum osteocalcin 151 3.3.6.3 Urine collagen crosslinks 152
3.3.7 Assays on tissue-culture conditioned media 152
3.3.7.1 Patellar cartilage explants 152 3.3.7.2 Subchondral bone explants 153 3.3.7.3 Synovial cultures 154
3.3.7.3.1 Passaged synovial fibroblast cultures 154 3.3.7.3.2 Primary synovial cultures 154
3.3.8 Oestrogen receptors 156
3.4 DISCUSSION 198
3.4.1 Effect of lateral meniscectomy on femoro-tibial
joint tissues of aged ewes 198
3.4.1.1 Cartilage changes after meniscectomy 198 3.4.1.2 Synovial changes after meniscectomy 199 3.4.1.3 Bone responses after meniscectomy 202
3.4.2 Effect of ovarieetomy on femoro-tibial joint tissues of aged ewes 206
3.4.2.1 Cartilage changes after ovariectomy 206 3.4.2.2 Synovial changes after ovariectomy 210 3.4.2.3 Bone changes after ovariectomy 211
3.4.3 Effect of prior ovariectomy on the development of
femoro-tibial joint lesions after menisectomy of aged ewes 213
3.4.3.1 Cartilage changes after ovariectomy and meniscectomy 213 3.4.3.2 Synovial changes after ovariectomy and meniscectomy 214
3.4.3.3 Bone changes after ovariectomy and meniscectomy 215 3.4.4 Effect of topical glyceryl-trinitrate on femoro-tibial
joint tissues in normal and ovariectomised sheep 216
3.4.5 Effect of topical glyceryl trinitrate on the development of
femoro-tibial joint lesions after ovine meniscectomy 221
Chapter 4: CONCLUSION AND SUMMARY 224
Appendix 1: SELECTED RESULTS FROM ANOTHER TRIAL OF
OVARIECTOMISED AND MENISCECTOMISED EWES 229
References 231

vii.
INDEX TO FIGURES 1.1 Schematic representation of articular cartilagezones 5 1.2 Structure of an aggrecan monomer and its attachment to hyaluronan 6 1.3 Schematic representation of the interaction of various matrix components 9 1.4 Schematic representation of the aetiopathogenesis of osteoarthritis 10 1.5 The enzymatic production of NO from arginine, as
catalysed by nitric oxide synthase 47 1.6 Dorsal view of the ovine tibial condyles with menisci intact 69 2.1 The cartilage histopathological scoring system used in this study 96 2.2 Schematic representation of the femoro-tibial joint in cross-section,
showing division for the purposes of histomorphometric analysis 97 2.3 The synovial histopathological scoring system used in this study 98 2.4 Scanned toluidine blue-stained sections of lateral tibial plateau 103 2.5 Gross appearance of non-operated control and meniscectomised joints 104 2.6 Histological appearance of toluidine blue-stained articular cartilage 104 2.7 Body weights 105 2.8 Cartilage thickness and proteoglycan staining - 3 months 107 2.9 Proteoglycan staining - 6 months 108 2.10 Cartilage and subchondral bone plate thickness (LFC/MFC) - 6 months 109 2.11 Cartilage and subchondral bone plate thickness (LTP/MTP) - 6 months 110 3.1 Structure of glyceryl trinitrate 117 3.2 The 2% glyceryl trinitrate ointment used in this trial 157 3.3 Sample DEXA scan of the tibial plateau 157 3.4 Application of glyceryl trinitrate ointment 158 3.5 Biomechanical testing of tibial plateau cartilage 159 3.6 Schematic showing the point locations where biomechanical
testing was performed 160 3.7 Polarised light-microscopic appearance of articular cartilage showing
collagen birefringence pattern 161 3.8 Scanned Masson's trichrome stained sections showing thickening of the
subchondral bone plate 162 3.9 Masson's trichrome stained section showing neovascularisation of cartilage 163 3.10 Ovine synovial fibroblasts 163 3.11 Solochrome cyanin-stained undecalcified bone section 164 3.12 Fluorescent labelling of undecalcified bone 164 3 13 MRI scans of normal and meniscectomised joints 165 3.14 Sheep bodyweights 166 3.15 Gross pathology scores 167 3.16 Correlation between gross appearance of synovium and
cartilage erosions and osteophyte formation 167 3.17 Dynamic shear modulus, phase lag, and thickness (non-meniscectomised) 168 3.18 Dynamic shear modulus, phase lag, and thickness (meniscectomised) 169 3.19 Calcified cartilage neovascularisation 171 3.20 Cartilage and subchondral plate thickness - LFC/MFC (non-meniscectomised) 172 3.21 Cartilage and subchondral plate thickness - LTP/MTP (non-meniscectomised) 173 3.22 Cartilage and subchondral plate thickness - LFC/MFC (meniscectomised) 174 3.23 Cartilage and subchondral plate thickness - LTP/MTP (meniscectomised) 175 3.24 Proteoglycan content (LFC/MFC) 176 3.25 Proteoglycan content (patella) 177 3.26 Osteophyte size 177

viii.
3.27 Collagen birefringence (LFC) 178 3.28 Collagen birefringence (LTP) 179 3.29 Collagen birefringence (MTP) 180 3.30 Cellularity of the synovial intimal layer 181 3.31 Surface parameters - LTP cancellous bone 182 3.32 Trabecular bone volume - LTP 182 3.33 Labelled surface - LTP cancellous bone 183 3.34 Bone mineral density -extra-articular sites 184 3.35 Subchondral bone mineral density 185 3 36 Cubic bone mineral density 186 3.37 Scattergram: relationship bet~veen subchondral thickness and BMD 186 3.38 Plasma 17ß-oestradiol 187,188 3.39 Serum osteocalcin 189 3.40 Urine collagen crosslinks 189 3.41 Cartilage explants - nitric oxide release 190 3.42 Cartilage explants - 35SO4-proteoglycan 190 3.43 Cartilage explants - MMP activity 191 3.44 Bone explants - nitric oxide 191 3.45 Bone explants - PGE2 192 3.46 Bone explants - urokinase activity 192 3.47 Synovial fibroblasts - nitric oxide 193 3.48 Synovial fibroblasts - PGE2 193 3.49 Synovial fibroblasts - MMP activity 194 3.50 Synovial fibroblasts - urokinase activity 194 3.51 Synovial fibroblasts - proliferation 195 3.52 Synovial fibroblasts - confluent density 195 3.53 Primary synovial cultures - IL-6 bioactivity 196 3.54 Primary synovial cultures - IL-1 bioactivity 196 3.55 Oestrogen receptors 197

ix.
INDEX TO TABLES 1.1 Classification of matrix metalloproteases (MMPs) found in cartilage 29 2.1 Gross pathology scores - 6 months 105 2.2 Mean aggregate histopathology scores - 6 months 106 2.3 Synovial membrane histopathology scores - 6 months 106 3.1 Treatment intervals (weeks) at time of sacrifice, by treatment group 121 3.2 Mean aggregate histopathology scores 170 3.3 Mean scores for calcified cartilage pathology 170 3 4 Chondrocyte density 170 3.5 Synovial membrane histopathology scores 181 3.6 Subchondral bone plate calcein score 183

x.
PUBLICATIONS AND CONFERENCE PRESENTATIONS
RESULTING FROM THIS WORK * * updated for this electronic version of thesis Dec 2005 REFEREED JOURNAL ARTICLES Cake MA, Read RA, Guillou B, Ghosh P (2000). Modification of articular cartilage and subchondral bone pathology in an ovine meniscectomy model of osteoarthritis by avocado and soya unsaponifiables (ASU). Osteoarthritis and Cartilage 8: 404-11 Appleyard RC, Burkhardt D, Ghosh P, Read R, Cake M, Swain MV, Murrell GAC (2003). Topographical analysis of the structural, biochemical and dynamic biomechanical properties of cartilage in an ovine model of osteoarthritis. Osteoarthritis and Cartilage 11: 65-77. Cake MA, Appleyard RC, Read RA, Ghosh P, Swain MV, Murrell GC (2003). Topical administration of the nitric oxide donor glyceryl trinitrate modifies the structural and biomechanical properties of ovine articular cartilage. Osteoarthritis and Cartilage 11: 872-8 Cake MA, Read RA, Appleyard RC, Hwa S-Y, Ghosh P (2004). The nitric oxide donor glyceryl trinitrate increases subchondral bone sclerosis and cartilage degeneration following ovine meniscectomy. Osteoarthritis and Cartilage 12: 974-981 Cake MA, Appleyard RC, Read RA, Smith MM, Murrell GC, Ghosh P (2005) Ovariectomy alters the structural and biomechanical properties of ovine femoro-tibial articular cartilage and increases cartilage iNOS. Osteoarthritis and Cartilage 13(12):1066-1075 PRESENTATIONS AT SCIENTIFIC MEETINGS
Cake MA, Read RA, Ghosh P (1999). Chondroprotective effects of Piasclidine 300 in an ovine meniscectomy model of OA. Australian Rheumatology Association (ARA), 42nd Annual Scientific
Conference, Perth WA, 23-26 May 1999
Cake MA, Read RA, Hwa S-Y, Ghosh P (2001). Glyceryl trinitrate increases subchondral bone thickness and density in an ovine meniscectomy model of osteoarthritis. Osteoarthritis and
Cartilage 9 (Suppl B): S54 [6th World Congress on Osteoarthritis, Osteoarthritis Research Society
International (OARSI) World Congress, Washington USA, 30 September - 3 October 2001]
Appleyard RC, Cake M, Ghosh P, Read R, Swain M, Murrell GAC (2001). Effects of a nitric oxide radical donor (glyceryl trinitrate) on joint articular cartilage of normal and ovariectomised ewes. Proceedings of the Matrix Biology Society of Australia and New Zealand (MBSANZ) Silver Jubilee
Scientific Conference, 4-7 October 2001 Cake MA, Appleyard RC, Read RA, Ghosh P, Swain MV, Murrell GAC (2002). Effects of topical glyceryl trinitrate on femoro-tibial articular cartilage and subchondral bone in normal and ovariectomised sheep. Trans. Orthop. Res. Soc. 2002 Abstract 0154 [Orthopaedic Research Society 48th Annual Meeting, Dallas USA, 10-13 February 2002]

xi.
Appleyard RC, Burkhardt D, Ghosh P, Read RA, Cake MA, Swain MV, Murrell GAC (2002). Topographical variation in biomechanical and biochemical properties of cartilage in an ovine osteoarthritis model: assessment with a novel arthroscopic indentation device. Trans. Orthop. Res.
Soc. 2002 Abstract 0667 [Orthopaedic Research Society 48th Annual Meeting, Dallas USA, 10-13
February 2002]
Cake MA, Read RA, Smith SM, Ghosh P (2002). Ovariectomy increases nitric oxide production in ovine patellar cartilage. Osteoarthritis and Cartilage 10 (Suppl. A):S9 [2002 OARSI World
Congress in Osteoarthritis, Osteoarthritis Research Society International (OARSI), Sydney 22-25th
September 2002]

xii.
ACKNOWLEDGEMENTS I would like to sincerely thank to the following people for their assistance with this study: Associate Professors Rick Read (Murdoch University) and Peter Ghosh (University of Sydney) for their constant guidance and inspiration. Mrs Diana Pethick, David Brockway, Peter Clune, Jim Poynton, and other staff of the Division of Veterinary and Biological Sciences, Murdoch University, for their tireless assistance with all animal procedures. All staff of Raymond Purves Bone and Joint Research Laboratories, Institute of Bone and Joint Research (Royal North Shore Hospital, Sydney) for their friendship, stimulating discussion, and helpful suggestions. In particular, thanks to Ms Susan Smith for her competent preparation of the histological sections, and Hwa Su-Yang for his help with DEXA measurements. Ms Joanna Makowey and Prof. Philip Sambrook of the Department of Rheumatology, Royal North Shore Hospital for the use of the Hologic QDR 4500W densitometer. Richard Appleyard and George Murrell of the Orthopaedic Research Institute, St George Hospital (University of NSW) for performing biomechanical testing and assisting with birefringence studies. Dr Els Meusen and Garry Barcham of the Centre for Animal Biotechnology, University of Melbourne for kindly providing the ovine recombinant IL-113. The Walter and Eliza Hall Institute, for kindly providing 7TD1 hybridoma cells. Graeme Worth and Bob Retallack of the Endocrinology laboratory, Sir Charles Gairdner Hospital, for performing the osteocalcin assays. Mark Walters and Roger Price (Sir Charles Gairdner Hospital) for their help with densitometry of lumbar vertebrae, and helpful discussions Graeme Martin and Margaret Blackberry of Animal Sciences, University of Western Australia for the use of their laboratories and assistance with plasma oestradiol assays. I would especially like to thank the Murdoch University Veterinary Trust for providing my research scholarship, and Boehringer Ingelheim Vetmedica for additional financial support. Finally, to Jen, Josie, Ellie and family, for all their love and support.

xiii.
ABBREVIATIONS
ACL Anterior cruciate ligament ACLT Anterior cruciate ligament transection ACTH Adrenocorticotropic hormone ADAM A disintegrin and metalloprotease ADAM-TS A disintegrin and metalloprotease with thrombospondin motifs ANOVA Analysis of variance APMA p-aminophenylmercuric acetate ASU Avocado-soya unsaponifiables bFGF Basic fibroblast growth factor BMD Bone mineral density BMI Body mass index CAF Chondrocyte activating factors CM Conditioned medium/media cNOS Constitutive (Ca2+-dependant) nitric oxide synthase COX Cyclooxygenase DEXA Dual-energy x-ray absorptiometry DMOADs Disease-modifying osteoarthritis drugs dpi Dots per inch DPyr Deoxypyridinoline EDRF Endothelial-derived relaxation factor EDTA Ethylenediaminetetraacetic acid EGF Epidermal growth factor eNOS Endothelial nitric oxide synthase ER Estrogen (oestrogen) receptor FGF Fibroblast growthfactor FSH Follicle-stimulating horrnone G* Corrected dynamic shear modulus GTN Glyceryl trinitrate HA Hyaluronic acid/ hyaluronan HEPES N-[2-hydroxyethyl]piperazine-N'-[2-ethanesulfonic acid] HPLC High-performance liquid chromatography HRT Hormone replacement therapy ICAM Intercellular adhesion molecule ICE Interleukin-1 converting enzyme IGF Insulin-like growth factor IGFBP Insulin-like growth factor binding protein IL Interleukin IL-1Ra Interleukin-1 receptor antagonist iNOS Inducible nitric oxide synthase kD KiloDaltons LFC Lateral femoral condyle LH Luteinising hormone LIF Leukocyte inhibitory factor L-NAME NG-nitro-L-arginine methylester (NOSI) L-NIL L-N6-iminoethyl-L-lysine (NOSI) L-NMA L-NG-monomethylarginine (NOSI) LTP Lateral tibial plateau MCM Monocyte-conditioned medium

xiv.
MFC Medial femoral condyle MHC Major histocompatability complex MMA Methyl methacrylate MMP Makix metalloprotease mRNA Messenger ribonucleic acid MT-MMP Membrane-type matrix metalloprotease MTP Medial tibial plateau MX Meniscectomised NO Nitric oxide NOC Non-operated control NOS Nitric oxide synthase NOSI Nitric oxide synthase inhibitor NSAIDs Non-steroidal anti-inflammatorydrugs OA Osteoarthritis OPG Osteoprotegerin OVX Ovariectomised PBM Peripheral blood mononuclear cells PDGF Platelet-derived growth factor PGE2 Prostaglandin E2 PLSD Protected least significant difference PBS Phosphate-buffered saline PS-GAG Polysulphated glycosaminoglycan PPS Pentosan polysulphate Pyr Pyridinoline RA Rheumatoid arthritis RANKL Receptor activator of NF- ß ligand RIA Radio-immunoassay ROI Region of interest rpm Revolutions perminute SCID Severe combined immunodeficiency SCP Subchondral bone plate SNAP s-nitrosyl-n-acetylpenicillamine (organic NO donor) TACE TNF- converting enzyme TG Trochlear groove TGF-ß Transforming growth factor-ß TIMP Tissue inhibitor of matrix metalloproteases tPA Tissue-type plasminogen activator UDPGD Uridine diphosphoglucose dehydrogenase UCC Uncalcified cartilage uPA Urokinase-type plasminogen activator uPAR Urokinase-type plasminogen activator receptor VCAM Vascular cell adhesion molecule VEGF Vascular endothelial growth factor

xv.
PREFACE
This thesis details the findings of sheep studies conducted between 1998 and 2002. Predominantly,
these studies investigate the effects of ovariectomy, and the exogenous nitric oxide donor glyceryl
trinitrate, on the structural and metabolic integrity of joint tissues in normal and osteoarthritic
(meniscectomised) sheep. A large number of techniques were used to fully investigate the effects of
these interventions, encompassing histology, biomechanical testing, bone densitometry, serum
markers, and extensive tissue culture. Most of these investigations were conducted by the author,
the most notable exception being the dynamic biomechanical testing of cartilage, which was carried
out in conjunction with Dr. Richard Appleyard of St. George Hospital, Sydney.
Due to the length and scope of these studies, efficient use of time and subjects required that multiple
hypotheses were tested within the design of a single large trial. For this reason, this thesis is
composed of only four chapters: (Chapter 1) an introductory literature review and statement of
hypotheses; (Chapter 2) a preliminary trial applying computerised image analysis methods to assess
osteoarthritic changes in this sheep model; (Chapter 3) an extensive trial testing the main
experimental hypotheses; and (Chapter 4) summary and conclusions. The testing of several
hypotheses simultaneously within a single large trial may make it difficult for the reader to follow
individual hypotheses through Chapter 3. It is important to note that the methods and results (3.2
and 3.3) relating to multiple hypotheses (stated in 1.3) are presented together (i.e. all eight
experimental groups), before discussing each hypothesis individually in 3.4.
Reference is made within the text (and Appendix 1) to findings from another trial involving
ovariectomised ewes, conducted simultaneously by our laboratories. Some of these findings have
been presented in preliminary abstract form and are referenced as published work (i.e. Smith et al.
(2000),769 Parker et al. (2000),611 Hwa et al. (2001)367). Some of the findings of the present study are
therefore described as confirmatory of these published works, but the reader should note these
studies were essentially conducted in parallel.

1.
CHAPTER 1
INTRODUCTION
1.1 LITERATURE REVIEW
1.1.1 OSTEOARTHRITIS
1.1.1.1 Definition and Classification of Osteoarthritis
Osteoarthritis (OA) is a disorder of moveable joints characterised by painful degeneration of
articular cartilage and osteochondral remodelling, including osteophyte formation at joint margins.17
Cartilage degeneration leads to progressive fibrillation, fissuring, ulceration, and finally full
thickness loss of cartilage, exposing the underlying subchondral bone.618 In the earliest stages of
OA, which are generally asymptomatic, cartilage lesions are small and focal and are accompanied
by a hypertrophic response in the surrounding cartilage.498 The term osteoarthritis implies a degree
of inflammation in the synovial membrane, which is often grossly evident but may be confined to
chronic low-grade inflammatory changes such as fibrosis. Similarly, the involvement of bony
tissues is implicit in the nomenclature and subchondral bone sclerosis is thought to be an early and
important change.618 There is increasing evidence that the synovium and subchondral bone are not
only the primary source of OA symptoms such as pain, but may also play an important role in the
disease pathogenesis via feedback interaction with cartilage, and have even been proposed as the
site of the initial lesion.618,629
A strict definition of OA is difficult, complicated by the surprisingly limited overlap between
various subsets such as symptomatic, radiographic, and self-reported OA.226 Classification by

2.
radiographic criteria is problematic, as a large proportion of patients with radiographically-defined
OA are asymptomatic. Despite this phenomenon, radiological criteria such as those proposed by
Kellgren and Lawrence (1957)414 and more recently Altman et al. (1995)21 remain the standard,
especially for epidemiological studies.
The heterogeneity of OA presents inherent difficulties in the classification of the disease. Subgroups
of OA have been identified by the joint involved (most commonly the hip, knee, hand, or spine in
humans), or by the aetiological distinction of primary (idiopathic) versus secondary OA, where
some predisposing cause is identified. A generalised form of primary OA has been described,
generally involving the hands (proximal and distal interphalangeal joints), knees, and hips of elderly
women.578 However, given the heterogeneous nature of the disease, OA is better conceptualised as
the final common pathway of a number of separate diseases, rather than being a single disease
entity.896
1.1.1.2 Aetiology and Epidemiology of OA
OA is the most common joint disorder in the world, with radiographic evidence of the disease
present in over 80% of people over the age of 75.224 The economic impact of OA is enormous; in
the USA, arthritic conditions represent the leading cause of disability in people over 15 years of
age, with a cost to the economy estimated at US$149 billion in 1992 alone.924 OA is also the most
common joint disorder in canine veterinary practice, with approximately 20% of dogs in the USA
over the age of 1 year affected.181,863 However, the prevalence of OA is imprecisely reported and
dependent on definition.
Risk factors for OA can be categorised into two broad mechanisms: those signalling a general
predisposition to the disease, and those causing abnormal loading to specific joint locations.724
Alternatively, they can be divided into those aetiological factors placing abnormal stress on normal

3.
cartilage, and those involving normal stress on abnormal cartilage. Consistently identified risk
factors for human OA include aging, obesity, high bone mass, major trauma (e.g. cruciate ligament
and meniscal injuries), repetitive use (often occupational), and developmental disorders such as
congenital dislocation of the hip.224
Interestingly, smoking is thought to be protective.722,724 Age is the most powerful predictor of OA
for all joints. It is unknown whether this is due to the cumulative effect of mechanical damage, or
intrinsic age-related changes in joint tissues.422 Obesity has been shown to have a strong association
with knee OA, but a less clear relationship with hip or hand involvement. In one Swedish study,
women with a high body mass index (BMI) at age 40 had a relative risk of knee replacement
surgery of 9.2.722 Gender also has a major influence on OA, especially in older patients. The
prevalence of OA is similar in men and women until about 50 years of age, after which females
show more prevalent, generalised, and severe OA.911 The coincidence of this observation with the
timing of the female menopause suggests that changes in sex hormones levels may potentially
influence the disease.
1.1.2 STRUCTURE OF NORMAL AND OSTEOARTHRITIC JOINT TISSUES
1.1.2.1 Articular Cartilage
The articular surfaces of synovial joints are covered with a smooth, resilient layer of hyaline
cartilage. This provides a durable weight-bearing surface with a very low coefficient of friction,
which is sufficiently 'spongy' to minimise peak stresses transmitted to the underlying bone.
Articular cartilage is a highly specialised tissue consisting of sparsely distributed chondrocytes
(only about 5% of its total volume), embedded in an abundant extracellular matrix. This matrix,
essentially a stiff fibre-reinforced gel, relies on two principal structural components for its
biomechanical properties: large hydrophilic proteoglycan polymers known as aggrecan, and a

4.
network of collagen fibrils, principally type II collagen. These two components provide cartilage
with its distinctive viscoelastic properties, the collagen chiefly providing resistance to deformation
and shear, while proteoglycans provide swelling pressure and pre-stress in the collagen network.745
Three cartilage layers, termed the superficial, intermediate, and deep zones, are arbitrarily
discriminated by histology and more recently by magnetic resonance imaging (Figure 1.1).158 These
layers are alternatively named according to the predominant orientation of collagen network, as the
tangential zone (fine fibrils densely packed parallel to articular surface), intermediate zone
(apparently random, oblique fibril orientation), and radial zone (large fibrils oriented perpendicular
to the cartilage surface). The cellular form of chondrocyte also varies by zone, from the elongated,
parallel-aligned cells of the superficial zone to the spherical form of deep zone chondrocytes.
Another classification of matrix compartments is defined by the alterations in the collagen network
in the proximity of chondrocytes: pericellular matrix (little collagen), territorial matrix (defined by a
fine collagen 'basket' around the chondrocyte), and interterritorial matrix (larger, more ordered,
parallel fibres). A chondrocyte, its surrounding capsule, and pericellular matrix form a 'chondron' –
regular spherical matrix units surrounding individual chondrocyte lacunae, which can be isolated by
specific digestion methods.647
1.1.2.1.1 Chondrocytes
Despite the avascular nature of cartilage, chondrocytes are very active metabolically. They
synthesise all macromolecules of the cartilage matrix, and regulate continuous turnover of matrix
components. The nutritional requirements of chondrocytes are met by passive diffusion, a process
assisted by intermittent joint compression. They are therefore largely dependant nutritionally on
synovial fluid;824 the extent of diffusion via the underlying bone is unknown. Phenotypically,
chondrocytes are not homogeneous but show regional variation in metabolic activity, particularly
between superficial and deep zone chondrocytes.329

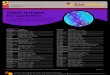
5.
Figure 1.1: Schematic representation of articular cartilage zones as defined by chondrocyte
morphology and predominant collagen orientation (reproduced, with permission, from Smith and Ghosh
(1998)771)
...
.
.. .. .
..
..
..
. .
.
..
.
..
. .
.. .
..
Superficial (10-20%)
Intermediate (40-60%)
Deep (30%)
Calcified cartilage
Subchondral bone
ZonesOrientation andmorphology of chondrocytes
Predominent orientation of
collagen fibrils
1.1.2.1.2 Cartilage Matrix
The extracellular matrix is composed primarily of water, which forms 70-80% of its wet weight,
variable with depth. This water is loosely bound to the hydrophilic proteoglycan-collagen
composite, and is able to extrude under compression. Such water movement is responsible for the
elasticity and resilience of cartilage, and may play a role in joint lubrication. Proteoglycans make up
approximately 22-38% of the dry weight of articular cartilage, the remainder being predominantly
collagen.502 The composition of articular cartilage is regionally variable; for example, the portion of
the tibial plateau not covered by the meniscus is thicker, less cellular, and biomechanically inferior
compared to that protected by the meniscus.41

6.
Figure 1.2: Structure of an aggrecan monomer and its attachment to hyaluronan. N, N-terminal
G1 domain; LP, link protein; HA, hyaluronic acid (after Lapcík et al. (1998)451)
Aggrecan
Most proteoglycans within articular cartilage are present as large aggregates in association with a
hyaluronic acid (HA) chain.315 Proteoglycans forming such aggregates are termed aggrecan. Many
individual aggrecan monomers are bound to a filamentous core formed by a single HA molecule;
this non-covalent bond involves a specific HA binding region (the G1 domain) and is stabilised by a
separate link glycoprotein (Figure 1.2). The proteoglycan monomers themselves have a similar
bottle brush-like structure, with glycosaminoglycan side-chains (principally keratan sulphate and
chondroitin sulphate) covalently attached to a central core protein.314
Due to their highly anionic nature, proteoglycans are very hydrophilic and expand greatly in volume
when fully hydrated. When combined into the massive aggrecan complex and enmeshed within the
Aggrecan core protein
Keratan sulphate side-chains
Chondroitin sulphate side-chains

7.
collagen network, proteoglycans create significant swelling pressure which gives cartilage its
turgidity, resilience and shock-absorbing capacity.848 By retaining nearly all available water,
proteoglycans also restrict water flow and sequester ions, and control fluid dynamics within the
cartilage matrix.
Non-aggregating Proteoglycans
A minor subpopulation of cartilage proteoglycans do not form aggregates with HA. The role of
these non-aggregating proteoglycans is unknown, though they are thought to be involved in
collagen fibril organisation, inhibiting mineralisation, regulating matrix turnover, and facilitating
interactions between chondrocytes and their surrounding matrix. Several small non-aggregating
proteoglycans are well characterised and are thought to have important roles. Decorin has a 36.5 kD
protein core and a single dermatan sulphate side-chain, and is thought to influence the morphology
of type I and II collagens.316 Fibromodulin, which has several keratan sulphate side-chains, localises
to the cartilage surface associated with the large glycoprotein fibronectin. Fibromodulin may
prevent the collagen interactions necessary for collagen auto-recognition and fibroblast adhesion.589
Biglycan has a small 38 kD core and a few dermatan sulphate side-chains. It is found mainly
pericellularly, though its function is unknown.316
Collagens
Articular cartilage contains the following types of collagen:
Fibrillar collagens
Type II collagen comprises 85-90% of the total collagen in articular cartilage. It is formed from a
triple helix of three identical alpha polypeptide chains. As the primary structural collagen in

8.
articular cartilage, it is responsible for the high tensile strength and ability to resist deformation
characteristic of this tissue. It is a product of the well-studied COL2A1 gene; mutations of this gene
have been associated with various connective tissue disorders such as achondroplasia.15
Type V and XI collagen are structurally similar, and are present in low amounts (<5% of total) in
association with type II fibrils. These collagens form small fibrils that closely associate with the
larger fibrils, and may function to modulate their diameter.80
Type I and III collagen are synthesised by chondrocytes in vitro but are minor components of
mature articular cartilage.
Non-fibrillar collagens
Type IV collagen is present in small amounts and may act to cross-link the larger fibrillar collagens.
Type VI collagen comprises 1-2% of total collagen, and is found mainly in the form of a fine
network which defines the pericellular capsule.647 It is relatively resistant to collagenase
degradation.62
Type IX collagen differs from type II in being the product of three separate genes, which code for
distinct alpha chains. It may act as a stabiliser between adjacent type II fibrils, limiting their
separation and aiding entrapment of proteoglycan aggregates (Figure 1.3).189
Type X collagen is normally synthesised only by hypertrophic chondrocytes of the calcified
cartilage zone, and is presumed to play a role in mineralisation of this layer.721,881

9.
Figure 1.3: Schematic representation of the interaction of various matrix components. Aggrecan
proteoglycan monomers elaborated by chondrocytes form large aggregates with cell surface-
associated hyaluronan. Minor collagens such as type IX, together with small non-aggregating
proteoglycans stabilise the network of larger type II collagen fibrils (reproduced, with permission, from
Smith and Ghosh (1998)771).
++++
+
+++
++
Type IX collagen
DecorinLink protein
Proteoglycan monomer
Proteoglycan aggregate
COMP
pCol II-c
Hyaluronan
Fibromodulin
Chondrocyte
Type II collagen fibrils
1.1.2.1.3 Cartilage Changes in Osteoarthritis
Conceptualisation of the pathogenesis of OA has altered over the past few decades, with recognition
that cartilage is a metabolically active tissue, and the changes observed are not simple wear
processes but are the results of imbalances in a dynamic equilibrium (Figure 1.4).629 Importantly, it
is now recognised that many of the processes of cartilage degradation can be mediated by the
chondrocyte alone. An imbalance in matrix homeostasis occurs, such that the equilibrium between
anabolic and catabolic events is shifted in favour of degradation. Anabolic activity increases
steadily up to a certain stage of degeneration, then dramatically decreases as repair mechanisms
become ineffective and cell death occurs.498

10.
Insert Figure 1.4 after 177

11.
Biochemically, OA progression is characterised by increased turnover and eventual net loss of
proteoglycan, early disruption of collagen architecture, and an increased water content. This
increase in hydration is one of the earliest detectable events in OA and implies failure of the elastic
restraint of aggrecan swelling provided by the collagen network. In early stages of the disease, the
anabolic repair response is predominant; however this is often defective, producing proteoglycans
of abnormal composition, distribution, and aggregating ability. The sulphation state of
glycosaminoglycans is increased, and the proportion of chondroitin 4-sulphate increases while that
of chondroitin-6-sulphate and keratan sulphate is decreased.499 Previously minor components such
as dermatan sulphate, decorin, biglycan, fibronectin are also proportionally increased.475 These
changes in chondrocyte metabolic activity are probably the result of continued cytokine stimulation
rather than a permanent phenotypic change, as they have experimentally been shown to reverse
after a few days in culture.864
Collagen changes occur early in the OA process, within a few days in experimental animal models.
Wider spacing of type II fibres is apparent, accompanied by the appearance of novel thinner fibres.
An increase in type II collagen synthesis is one of the early anabolic responses of OA chondrocytes,
which may also show abnormal production of type I, III and X collagens.9,881 It has been shown in
vitro that the key catabolic cytokine interleukin-1ß (IL-1ß) selectively increases chondrocyte
synthesis of type I and III collagens while depressing type II and IX collagen synthesis.721 Collagen
breakdown and turnover is also increased in OA cartilage. As the basal rate of collagen turnover is
lower than that of proteoglycans, the loss of the collagen network is a less reversible change than
aggrecan loss.636
Structurally, OA is characterised by the appearance of perpendicular clefts in the articular cartilage
surface, which become progressively deeper and coalesce to form regions of overt fibrillation. Loss
of proteoglycan staining progresses from the superficial layer into deeper zones with worsening
disease.498 Initially, hypercellularity is present with the formation of clusters of chondrocyte clones,

12.
later progressing to hypocellularity and the appearance of empty chondrocyte lacunae. Cell death
may be a central event in OA, a process possibly driven by chondrocyte apoptosis.322
Biomechanically, OA cartilage shows reduced static compressive stiffness, a change partly related
to increased matrix permeability, which is normally very low.455 Dynamic shear stiffness, measured
as the dynamic shear modulus (G*), is often severely reduced due to fibrillation, rupture of collagen
fibres and loss of proteoglycan-supplied collagen pre-stress.935 Phase angle ( ), a vector of viscous
and elastic properties ranging from 0° (perfectly elastic) to 90° (perfectly viscous), is increased
suggesting increased frictional energy dissipation due to loosening of collagen-proteoglycan
interactions.745,746
1.1.2.2 Synovial Membrane
1.1.2.2.1 Structure of Normal Synovium
The synovial cavity is lined by a thin layer of specialised cells, whose major function is to produce
and regulate the synovial fluid which lubricates and provides nutrition to articular cartilage. Normal
synovium consists of an intimal lining layer containing primarily those cells designated
‘synoviocytes’, and a subintimal layer of fibroblasts, macrophages, lymphocytes, mast cells, nerves,
and vascular elements in a loose connective tissue matrix.339,558 The lining layer is 1-3 cells deep in
normal synovium, with no basement membrane, though tight junctions between cells may be
present. 205,558,759,917 Decorin, biglycan, fibromodulin and fibronectin are co-distributed with collagen
in the loose subsynovial connective tissue.146,613 The synovial nerve supply was once thought to be
sparse, but a good supply of nociceptive unmyelinated C fibres is now recognised.501 Changes in
synovial membrane composition occur with age, with more numerous villi and increasing
irregularity of blood vessel and cell distribution.613

13.
Synoviocytes are unusual in their mixed characteristics of a mesenchymal origin, immunological
capability, and an epithelial role.753 Barland et al. (1962)58 was the first study to identify two distinct
synovial cell types, on the basis of their transmission electron microscopic characteristics. These
could be broadly characterised as phagocytic (type A) and secretory (type B) cells. This functional
definition has lead to the suggestion by several authors that synoviocytes represent a single cell type
with great morphologic flexibility.221,280,423 Other studies have concluded the two cell types are
totally distinct and no intermediate types exist.279
Type A synoviocytes are thought to function as phagocytic cells with antigen processing
capabilities. Phenotypically they resemble tissue macrophages, with a prominent heterochromatin-
rich nucleus, numerous peripheral cytoplasmic vacuoles, fine lamellar extensions of the cell surface,
a prominent Golgi apparatus, and scant endoplasmic reticulum.221,279,558 They express typical
macrophage markers such as CD11, CD14, CD68, and class II major histocompatibility complex
(MHC) antigens, as well as non-specific esterase activity.558,900 Type A synoviocytes comprise
approximately 30% of cells in the intima and subintima, with the proportion reportedly increasing
with age and progression of arthritis.138,288,613 However, a decrease in Type A synoviocytes has been
reported as an early pathologic change in canine hip dysplasia.289 In rheumatoid arthritis (RA), a
distinct subpopulation of Type A cells is recognised, termed dendritic cells. These CD13/CD33-
positive cells interact with T lymphocytes as antigen-presenting cells.48,558
Type B synoviocytes resemble tissue fibroblasts, with a bipolar or spindle-shaped form, extensive
rough endoplasmic reticulum with wide cisternae, less developed Golgi apparatus, pale
euchromatin-rich nuclei with a number of nucleoli, and small cytoplasmic secretory granules, but
fewer large vacuoles than Type A cells. 221,279,558 These morphological features point to their
primarily synthetic function, in the production of hyaluronan and matrix components such as
collagen. Several distinctive features distinguish Type B synoviocytes from other tissue fibroblasts:
(1) sustained release of interleukin-6 (IL-6) in culture;128 (2) expression of vascular cell adhesion

14.
molecule-1 (VCAM-1);231,553,899 and (3) high activity of uridine diphosphoglucose dehydrogenase
(UDPGD), an enzyme related to hyaluronan synthesis.231,377,900 However, membranes lined with
cells of high UDPGD activity have been shown to form spontaneously around prosthetic implants,
especially where shear stresses occur.205 Approximately 75-85% of the glycosaminoglycans
synthesised by synovial fibroblasts is hyaluronan.26,375 HA synthesis is continued by cells in tissue
culture, forming a loose pericellular matrix around each cell.242 Synovial fibroblasts also produce
small sulphated glycosaminoglycans, mainly in the form of dermatan sulphate.118,300
While VCAM-1 does appear to be a distinctive feature of synovial fibroblasts, expression is more
consistent in arthritic synovia.437,553,899 Intercellular adhesion molecule-1 (ICAM-1) is also
expressed following cytokine stimulation; up-regulated expression of these adhesion molecules in
RA has been linked to the binding of invading inflammatory cells.553 Cells of the immortalised
rheumatoid synovial fibroblast line RAMAK-1 constitutively express ICAM-1, but express VCAM-
1 only when stimulated by tumour necrosis factor- (TNF- ).409
The formerly debated origins of Type A and B cells have been clarified by the studies of Edwards et
al. (1994).205 Following bone marrow implantation from beige mice into irradiated histocompatible
normal mice, giant granules were identified only in cells with the characteristics of Type A cells.
This suggests that, in common with other tissue macrophages, Type A cells are derived from bone
marrow and subsequently migrate to the synovial intima to undergo maturation.376 Expression of
leucocyte common antigen by Type A cells also suggests their origin from haematopoietic stem
cells.48 Type B cells are derived either locally (by cell division), or from an unidentified stem cell
population in the subintima. However, the rate of cell division in synovial lining cells is low under
most conditions.
Synovial fibroblasts (Type B synoviocytes) were once considered to be relatively inactive secretory
and maintenance cells, but studies in RA have confirmed their potential for joint destruction when

15.
activated.558 Experiments in severe combined immunodeficiency (SCID) mice and refined in vitro
systems have shown that synovial fibroblasts are capable of cartilage destruction completely
independent of T-cell influence.558,740
1.1.2.2.2 Synovial Changes in Osteoarthritis
Whilst OA is not historically considered an inflammatory disease, some degree of synovitis is often
present and is likely to be important in the disease pathogenesis. It is becoming recognised that OA
synovium can demonstrate the same inflamed and activated phenotype as rheumatoid tissue, and
that the difference between the diseases is more quantitative than qualitative.214,472 In an important
study by Smith et al. (1997)768 examining synovial biopsies from patients with early OA, a
significant increase in thickness, vascularity, inflammatory cell infiltration, and cytokine production
was consistently detected even in early stages of the disease. Altered synovial vascularity (dilation
of existing vessels and angiogenesis) is a consistent change in OA. Despite this altered vascularity,
synovial fluid can be hypoxic and acidic in inflammatory conditions.797 Altered vessel permeability
and subsequent leakage of serum proteins promotes synovial oedema, increased synovial fluid
volume, and the accumulation of fibrin on the intimal surface.243,888 The same changes also admit
joint-derived HA into the circulation, and elevated serum HA levels have been shown to be
predictive of OA progression and synovitis.495,747
Infiltration of inflammatory cells (especially T and B lymphocytes) is variable and generally focal
in OA synovia, but can be considerable.470,568 A subset of OA patients seem to show greater severity
of inflammatory features, with prominent lymphoid aggregates.682 However, the importance of T-
cells in synovial inflammation is questionable, as T-cell-specific cytokines are absent from both OA
and rheumatoid synovia, and perivascular lymphoid aggregates are occasionally present in normal
human synovium.214,471,576 Mast cells are found in appreciable numbers in OA synovia, and may
play a role in synovial inflammation and joint effusion.94,337,640,652,680,681

16.
Mild to moderate synovitis is observed in around 50% of biopsies from OA dogs.863 Following
experimental anterior cruciate ligament transection (ACLT) in dogs, synovitis develops rapidly.567
Hyperplasia and inflammatory changes (subsynovial lymphocyte and plasma cell accumulation) are
prominent by 8 weeks post-operatively, and fibrosis develops within 13 weeks.473 However, careful
haemostasis during open surgery considerably reduces the incidence of synovitis when compared to
a blind ‘stab’ technique.567,569 In rabbit models of OA, synovial responses are rapid and severe, often
preceding observable cartilage changes.808 Synovial hyperplasia is a prominent event, observed
rapidly following the Hulth procedure (ACLT and hemimeniscectomy),484 joint immobilisation,430
10% NaCl injection,862 or repetitive impulse loading.880
The cause of synovitis in OA has not been identified, but breakdown products of cartilage are
assumed to play a role.210,629 Cartilage-derived proteoglycans provoke synovitis when injected intra-
articularly in rabbits, suggesting the effect of cartilage wear products may be from chemical factors
as much as the physical presence of particles.83,252 Breakdown products of type II collagen are
similarly antigenic.436 Embedded macroscopic cartilage shards are commonly observed in the
synovium in end-stage OA, but are absent in the initial phases and are not visibly associated with
inflammatory changes.571 Microcrystals of calcium pyrophosphate and hydroxyapatite are often
present in OA synovial fluids, though their significance is unknown.20,738,860
1.1.2.2.3 Synovium-Cartilage Interactions in Osteoarthritis
In a landmark series of experiments, Fell and Jubb (1977)222 demonstrated that normal synovium,
when minced, was capable of inducing degradation of co-cultured explants of normal cartilage.
Subsequent experiments showed that: (i) killed cartilage showed significantly less degradation,
suggesting that any direct enzymatic action was only partially responsible; and (ii) direct contact
was not required, highlighting for the first time the presence of soluble ‘chondrocyte activating
factors’ (CAF) capable of inducing chondrocytes to digest their own matrix.221 Meats et al.

17.
(1980)534 further showed that these synovial factors induced chondrocyte release of plasminogen
activator and prostaglandin E2 (PGE2). More recently, Scott et al. (1997)740 showed using a
rheumatoid synovial fibroblast / macrophage line co-culture system, that 90% of CAF activity could
be abolished by blocking antibodies to IL-1ß and TNF- , though IL-6 antibodies had a surprisingly
large effect. However, other studies have found a less dominant role for IL-1 and shown that other
as yet uncharacterised cytokines contribute to synoviocyte-chondrocyte cross-talk.57 The umbrella
of synovium-derived CAF probably also includes various growth factors, which are less well
characterised than catabolic cytokines, but may include transforming growth factor-ß (TGF-ß) and
basic fibroblast growth factor (bFGF).32,57,130
Current evidence suggests that in OA, pro-inflammatory cytokines may potentially be produced
first in the synovial membrane, then diffuse across the synovial fluid to affect chondrocytes.618
However, the true importance of such tissue cross-talk is unknown. In a study directly comparing
OA cartilage and synovium within the same joint, chondrocytes were seen to be the primary source
of the important mediators IL-1, TNF- and nitric oxide.538 Immunohistological studies of OA
synovia also demonstrate increased expression of matrix metalloproteases (MMPs) and their
inhibitors, predominantly in the intimal lining layer, in levels which correlate with the histological
severity of synovitis.232,593 It has been demonstrated that synovial fibroblasts are a major source of
these catabolic enzymes.539 However, the role of synovial-derived proteases in cartilage degradation
has not been clearly demonstrated, with current evidence suggesting they are less important than
endogenous cartilage enzymes.624
1.1.2.2.4 Rheumatoid Arthritis
Rheumatoid arthritis is a chronic polyarthritis characterised by severe synovial hyperplasia and
dense subsynovial lymphoplasmacytic infiltrate. The simultaneous involvement of multiple joints is
strongly suggestive of initiation by some circulating factor; one hypothesis suggests localisation of

18.
persistent bacterial or viral antigens to the joint as a causal pathway.310 Genetic factors also seem
important though it is unknown whether these relate to differences in systemic or local responses.737
Intimal hyperplasia is one of the earliest and most striking alterations in RA, and correlates directly
with the severity of joint erosion.339 Two main pathways seem to operate in RA: (i) infiltration of
the subsynovium by various T-cell types, which may (or may not) play a central role in initiating
and perpetuating inflammation through interactions with antigen-presenting cells;857 and (ii)
activation of synovial fibroblasts to form an extremely aggressive, transformed phenotype.324 RA
synovium often forms a vascular 'pannus' which adheres to and erodes affected bone and cartilage.
Lymphoid elements may make up over 5% of the total tissue mass in RA.765 It has been
demonstrated that patients with focal lymphoid aggregates produce more inflammatory cytokines
than patients with more diffuse mononuclear infiltration, suggesting this marks a greater degree of
immune activation.919 Certain characteristics of rheumatoid synovium, such as the invasive growth
of synovial xenografts implanted into SCID mice, are suggestive of neoplastic transformation.231
Certainly, the oncogene products egr-1, c-fos and c-jun are constitutively expressed in RA,201
though this is probably a reflection of chronic inflammation rather than any primary growth
abnormality.693
1.1.2.2.5 Synovial Fluid
Synovial joints contain a small volume of synovial fluid, which nourishes and lubricates the
articular cartilage. By forming a thin viscous film between congruent joint surfaces, synovial fluid
opposes distraction but not shearing movement.759 Though synovial fluid, and in particular its
hyaluronan content, is known to be important in cartilage-on-cartilage lubrication, the exact mode
of this lubrication is the subject of some debate, and may differ between high and low load states.
Thin film lubrication,485 boundary lubrication, “weeping” hydrostatic lubrication657, and “boosted”
lubrication (by squeeze-film entrapment and enrichment)882 have all been proposed.

19.
Synovial fluid is essentially an ultrafiltrate of plasma, supplemented with HA secreted by type B
synovial lining cells.889 Another synovial fluid protein not derived from plasma is lubricin, which is
thought to be important in boundary-layer lubrication of cartilage. The pressure within healthy
synovial fluid is normally sub-atmospheric, due to the swelling pressure exerted by the hyaluronan
matrix.759 In OA, the mechanisms allowing selective exclusion of serum components from synovial
fluid fail, resulting in: (i) increased protein content; (ii) increased fibrinogen content, hence a
tendency to clot; (iii) increased inflammatory cell content; and (iv) reduced viscosity due to dilution
of HA, despite an increase in HA synthesis by the inflamed synovium.567 The HA component of
synovial fluid is crucial to its rheologic properties, resistance to fluid leakage out of the joint
cavity,148 and maintenance of sub-atmospheric joint pressures.759 Reduction in both the quantity and
quality of synovial fluid HA has a major influence of the progressive deterioration of OA joints.
Locally-generated oxygen radicals may be another significant cause of HA degradation within
inflamed joints.730
Certain synovial fluid markers have been investigated as diagnostic indicators of OA disease
activity.601 The cytokine content of synovial fluid has been examined by numerous studies.465,678 IL-
6 and TNF- have shown promise as synovial fluid markers in anterior cruciate ligament (ACL)-
deficient dogs and humans.105,328 Reported synovial fluid levels of IL-6 vary widely – from 1 to
1000 pg/ml in OA patients, and up to 72,300 pg/ml in inflammatory arthropathies.678 Synovial IL-
1ß levels are an unreliable marker: while Westacott et al. (1990)898 found appreciable levels, other
studies have failed to detect IL-1 in OA synovial fluids.74,865 Cartilage-derived proteoglycan
fragments are also promising synovial fluid markers of joint disease, measured either as
glycosaminoglycan content by dye binding assays,113,167,372,668 or by immunological methods
recognising the HA binding region of aggrecan.105,113,910

20.
1.1.2.3 Subchondral Bone
1.1.2.3.1 Normal Structure of Subchondral Bone
The subchondral bone region is defined as the thin layer of cortical bone between the calcified
cartilage and the supportive epiphyseal cancellous bone. The calcified cartilage layer provides a
transition zone between the compliant cartilage and stiffer bone, freely traversed by type II collagen
fibres which are anchored to the subchondral bone. The calcified layer is distinct in containing type
X collagen, which is likely to be involved in the calcification process.721 During growth, the deepest
layers of calcified cartilage undergo endochondral ossification, while the superficial boundary, or
tidemark, advances into the uncalcified layers above. The calcified cartilage and its supporting
cortical bone form a functional unit often termed the subchondral bone plate. The irregular and
convoluted interface between these layers greatly strengthens the bond created by the thin cement-
like layer between them.388 The thickness of the subchondral plate is often proportional to the
thickness of the overlying cartilage; this phenomenon is not easily explained, as the stresses
responsible for modelling bone (chronic, low frequency) and cartilage (dynamic, high frequency)
are potentially different.545
The basic unit of bone, an osteon, represents a central Haversian canal and its surrounding
concentric lamellae of mineralised matrix.155 There are two major groups of cells within bone:
Osteoblasts are plump, strongly basophilic cells that line most bony surfaces. They arise from
osteoprogenitor stem cells within the bone marrow.71 Osteoblasts produce (and gradually become
buried in) the non-mineralised collagenous matrix known as osteoid. As osteoid mineralises the
enclosed cells lose much of their secretory activity and become phenotypically distinct as
osteocytes, which extend long cytoplasmic projections into minute bone channels known as
canaliculi.

21.
Osteoclasts are large multinucleate cells with abundant cytoplasm, mitochondria, and ribosomes.
Osteoclasts are distinct in their origin from the circulating monocyte-macrophage system.696 They
become activated on attachment to mineralised bone, forming a ruffled border in intimate contact
with the exposed bone surface. Enzymes active under acid conditions are secreted to dissolve bone,
producing resorption pits termed Howship's lacunae.
Bone is not static but is continuously remodelled according to Wolff's Law, which states that the
architecture of bone is dictated by the stresses acting upon it.912 Classical remodelling involves a
cycle of osteoclastic resorption, followed by deposition and eventual mineralisation of osteoid.
Osteoid primarily consists of type I collagen, deposited within a ground substance of proteoglycan
and non-collagenous proteins such as osteonectin and osteocalcin. Mineralisation involves the
formation of hydroxyapatite crystals (Ca10(PO4)6(OH)2) in a poorly understood process apparently
initiated by the osteocyte.
1.1.2.3.2 Subchondral Bone Changes in Osteoarthritis
Sclerosis and stiffening of the subchondral bone plate are well-recognised characteristics of OA,
though thinning of the underlying cancellous trabeculae has also been demonstrated. Rapid
appositional bone growth produces prominent layers of osteoid, while lagging mineralisation
creates a relatively hypomineralised matrix.39,293 Osteonecrosis is frequently observed, probably due
to ischaemia secondary to microfracture or thrombus formation. Resorption of necrotic regions
creates the large subchondral bone cysts commonly seen in some species. Calcified cartilage
remodelling occurs, resulting in increased vascularity and irregularity and duplication of the
tidemark.

22.
Two recent studies have shown metabolic differences in bone cells derived from the subchondral
region of human OA joints. Westacott et al. (1997)897 found that supernatants from OA bone cells
induced release of glycosaminoglycan from cartilage explants, whether they were viable or dead.
Hilal et al. (1998)350 found that both subchondral explants and primary cultures of osteoblast-like
cells show an altered phenotype in OA, with increased release of urokinase-type plasminogen
activator, insulin-like growth factor-1 (IGF-1), alkaline phosphatase, and osteocalcin. Such studies
show that active local release of cytokines and other mediators may be at least as important in
subchondral bone remodelling as mechanical mechanisms. They also raise the possibility that bone-
derived factors such as cytokines and catabolic enzymes might have a direct effect on overlying
cartilage if able to diffuse across the bone-cartilage interface.243 On the basis of these studies,
Martel-Pelletier et al. (1999)505 recently proposed that up-regulation of the IGF / IGF binding
protein (IGFBP) axis is central to the simultaneous events of attempted cartilage repair and
subchondral sclerosis. Whether systemic release of growth factors (IGF-1, TGF-ß) might also
mediate the differences in bone metabolism at distant skeletal sites that have been observed in OA
remains an intriguing theory.184
Altered subchondral bone turnover and metabolic activity appear to be early and central changes in
OA.350 Dieppe et al. (1993)194 demonstrated increased activity in OA subchondral bone by
scintigraphy and, importantly, found that this was predictive of joint space narrowing. Observed
elevations of type I collagen synthesis, MMP-2 and alkaline phosphatase activity, and integrin
expression in OA subchondral bone similarly demonstrates altered metabolic activity.54,107,500 A
localised increase in concentrations of growth factors such as TGF-ß, as has been observed in
similar tissue, may represent one stimulus for such changes.500 Other cytokines that may be
involved in OA bone remodelling (particularly bone loss) include IL-1, IL-6, IL-11, and TNF- .
Receptor activator of NF- ß ligand (RANKL), a potent stimulus of osteoclast formation and
activation,355 is detectable in rheumatoid synovium, but not healthy tissues.285 Fazzalari et al.
(2001)219 recently showed that while RANKL levels correlate strongly with bone turnover in normal

23.
femoral heads, the same association is not evident in OA bone, suggesting inflammatory cytokines
such as those mentioned above may assume a dominant role in the control of bone metabolism in
arthritic conditions.
1.1.2.3.3 Role of Subchondral Bone Changes in the Pathogenesis of OA
Radin et al. (1972)658 first proposed that sclerosis and stiffening of the subchondral bone plate may
be central to the pathogenesis of OA. They proposed that repetitive impulse loading results in
trabecular microfractures and subsequent remodelling and stiffening, which reduces compliance of
the subchondral bone structure and increases the stresses applied to cartilage. Advancement of the
tidemark effectively causes thinning of the uncalcified layer, which increases shear forces near the
base of the cartilage.363 Other authors have subsequently endorsed the theory that bone sclerosis
plays a major role in the pathogenesis of OA.412,505,506,618,656 However, whether this process truly
initiates or is an early secondary feature of cartilage loss remains the subject of debate. Several
observations complicate this hypothesis. Firstly, it is very difficult to separate cartilage and bone as
a mechanical unit; for example, in a study of aging, the stiffness of bone and cartilage was found to
be very closely correlated.195 Secondly, multiple studies have demonstrated that increasing
thickness of osteoarthritic subchondral bone is accompanied by hypomineralisation, due to
excessive osteoid production.39,293 This suggests a defect of the remodelling process, probably
related to the alterations in cellular metabolism already discussed.350,505 Thirdly, subchondral bone
stiffness may be related to a more generalised alteration in skeletal bone quality, as suggested by
Dequeker and coworkers.184,185

24.
Two large bodies of evidence suggest an integral link between bone density and OA, though the
exact nature of this association is far from clear:
Epidemiological Evidence
Multiple studies have concluded an inverse epidemiological relationship between osteoporosis and
OA.312,779 In several large cross-sectional populations, women with hip,153,580,798 hand,319,778
knee,319,778 or spine319 OA have been shown to have higher bone mineral density (BMD) than did
non-arthritic subjects. In another study, women with radiographically-defined knee OA were found
to have greater average BMD, less temporal change in BMD, and lower serum levels of osteocalcin
than unaffected women.779 However, the same may not hold for hand OA.779 In the iliac crest, a site
with no direct mechanical stimulus, bone density is increased in patients with hand OA, as are
levels of bone IGF-1 and TGF-ß 184 As it is difficult to conceive that this could be a secondary
change, such findings support the notion that an inherent difference in bone quality may predispose
to a form of primary OA in certain groups. Alternatively, if subchondral stiffening is an initiating
cause in OA, perhaps it occurs as part of a more generalised increase in bone biosynthesis.185
Certainly, patients affected by osteopetrosis are prone to subsequent development of OA.505
However, most of these studies have used relatively severe radiographic criteria of OA. Patients
with osteoporosis and osteoarthritis may also have lifestyle differences, such as greater exercise
levels in earlier life.403 The two populations also differ anthropometrically.184 For instance, the
higher body weight in the OA subset influences bone density not only mechanically, but also from
higher postmenopausal oestrogen levels associated with increased body fat and peripheral
conversion of androstenedione to estrone.185 A recent study involving female twin pairs, one with
hip OA and one without, found that hip OA was associated with a higher BMD of the ipsilateral
femoral neck, but not any of the other sites tested.31 This study, while confirming the presence of
local bone changes in association with OA, suggests that the more general relation between OA and
high BMD is likely to be due to shared genetic factors.

25.
Several studies have suggested a link between dynamic bone loss and OA. Karvonen et al. (1998)403
found that patients with relatively mild knee OA had a significant decrease of periarticular BMD,
despite no significant difference in spine or femoral shaft density. Local osteoporosis was found to
be more severe in superficial rather than deep subchondral regions. In a long-term prospective
study, Sowers et al. (1991)780 found that despite greater baseline bone density, women who
developed hand OA showed greater bone density loss than normal women. Such data argues against
a primary role for trauma-induced subchondral sclerosis in the pathogenesis of OA.403 Interestingly,
low serum levels of 1,25-dihydroxycholecalciferol, a well-known promoter of bone mineralisation,
are associated with increased incidence of hip OA449 and more rapid progression of knee OA,523
though whether this is related to its skeletal influence or direct effects on cartilage metabolism
remains to be established.
Experimental Evidence
Proof that subchondral bone changes play a primary initiating role in the pathogenesis of OA
depends on the observation of very early bone changes in natural and experimental models. Indeed,
in various animal models of spontaneous OA, such as in macaques and Dunkin-Hartley guinea pigs,
subchondral bone changes are often the earliest detectable changes, preceding detectable cartilage
fibrillation.54,89 In rabbits subjected to impulsive loading, subchondral thickening and remodelling
can be demonstrated within six weeks, prior to detectable changes in cartilage proteoglycan
content.659 However, surgical models (especially those involving unilateral surgery) are
compromised by their effect on limb loading. In meniscectomised rabbits, a decrease in bone
density is observed due to reduced limb usage post-surgery.540 In the canine ACLT model, the
subchondral region shows a pattern of early trabecular bone loss and later subchondral plate
thickening.179 Loss of trabecular bone mass can be demonstrated by micro-computerised
tomography from 3 months post-operatively, well before subchondral sclerosis is evident, and
persists to 54 months.179 Histomorphometric analysis shows reduced bone mass and increased bone
turnover as early as 6 weeks following ACLT.931 Many critics attribute this trabecular loss to

26.
decreased loading following surgery.90,590 However, even if the joint is denervated by
ganglionectomy (a procedure that also accelerates cartilage destruction), rapid loss of bone ensues,
with increased porosity of the subchondral plate and trabecular osteopenia.180 In this model,
therefore, thickening of subchondral bone does not appear to be necessary for the development of
cartilage changes, and occurs much later in the progression of disease.90,179,180 In a long-term (54
months) follow-up study of ACLT dogs, subchondral studies demonstrated a marked increase in
bone volume and active bone formation, as demonstrated by fluorochrome labelling. Further, these
changes were demonstrated only in the medial femoral condyle, not the lateral condyle.90 However
in a canine histomorphometric study of OA hip joints, greater subchondral trabecular area was
found to correlate with an increasing degree of cartilage pathology.800
1.1.2.3.4 Osteophytes
Osteophytes are bony outgrowths occurring at the margins of arthritic joints, which form by either
endochondral or intramembraneous ossification. The stimulus for osteophyte formation is unknown,
though cartilage damage, joint instability, inflammation, and stretching of the synovial membrane
have all been proposed as mechanisms.682 Experimental observations of osteophyte formation in the
absence of instability argue against this as a primary cause.388 The function of osteophytes is
thought to be mainly protective, countering instability by decreasing range of motion, and reducing
focal loading by expanding the effective weight-bearing surface.14 However, they are also a
significant source of pain due to periosteal stretching.388
1.1.2.3.5 Markers of Bone Turnover in OA
Several studies have shown alterations of serum and urine bone turnover markers in OA patients.
Pyridinoline (Pyr) and deoxypyridinoline (DPyr) are elevated in both OA and RA patients when
compared to reported levels in control populations.450,762,798 These molecules are important in cross-

27.
linkage of mature collagen and are released during its degradation. In rheumatoid patients, levels
are significantly elevated during active RA when compared with subjects with suppressed disease.11
Similar elevations are seen following intra-articular chymopapain injection in rabbits.846 However,
whether such markers reflect local changes or are associated with systemic changes in bone
turnover is unknown.54
Plasma osteocalcin levels provide a useful marker of osteoblast function. Due to its specific origin
in newly formed bone and short plasma half-life, osteocalcin levels reflect the current activity of
osteoblast-like cells,157 though variations in renal and extra-renal clearance rate also potentially
influence plasma levels.218 The lower serum osteocalcin levels seen in patients with incident hand or
knee OA therefore suggest a reduction of bone formation and turnover during active OA.779,798
Similarly, serum osteocalcin levels are reduced in patients with active RA when compared to
subjects with inactive or no disease.11 However, other studies have shown elevated serum
osteocalcin in patients with hand OA350 or with active psoriatic arthritis.240
1.1.3 PATHOGENETIC MEDIATORS OF OSTEOARTHRITIS
1.1.3.1 Enzymes and Enzyme Inhibitors
The importance of enzymatic degradation of cartilage, rather than simple mechanical disruption, has
been known for several decades.275,625 Proteolytic enzymes secreted by chondrocytes are
predominantly responsible for normal turnover of various extracellular matrix components.
Hyaluronan may be an exception here, as it is thought to be internalised and degraded intracellularly
after binding to CD44 cell surface markers.209,833 Overproduction of these enzymes in OA causes
structural degeneration through two main process: degradation and loss of proteoglycan, and

28.
cleavage and loosening of the fibrillar collagen network. Cartilage appears particularly susceptible
to protease attack, with even a single enzyme being able to cause major defects in its structural and
biomechanical properties.
1.1.3.1.1 Matrix Metalloproteases
While it must be recognised that matrix breakdown occurs through a complex cascade of proteolytic
events, the enzymes most consistently implicated are members of the matrix metalloprotease family
(Table 1.1). As well as the ability to degrade matrix components, MMPs share the following
common features: (i) they contain a zinc atom at the active site; (ii) they require Ca++ for full
activity, and are inhibited by calcium-chelating agents; (iii) they display peak activity at neutral pH;
and (iv) they are secreted as pro-enzymes which require proteolytic cleavage for full activity.612
Four MMP subclasses are elevated in OA: stromelysin (MMP-3), acting on proteoglycan and type
IX collagen; collagenases (MMP-1,-8,-13), acting on native fibrillar collagen; gelatinases (MMP-2,-
9), acting on denatured collagen; and membrane-type MT-MMPs, which may play a role in cell-
associated protease activity. In addition to its role in activating MMP-2, MT1-MMP may also act as
a collagenase in its own right.618 The recently characterised aggrecanases, members of the related
ADAMTS (A Disintegrin And Metalloprotease with ThromboSpondin motifs) family, specifically
degrade aggrecan.828
Many of the MMPs are not constitutively produced, but are induced in chondrocytes by IL-1ß and
other cytokines by complex mechanisms involving the AP-1 promoter element and the oncoproteins
c-jun and c-fos.291,420,444,467,620 Induction of collagenase has also been shown to involve Protein
Kinase C and NF- ß induction, but is antagonised by Protein Kinase A.115,192,872 Expression of
stromelysin and collagenase is generally closely linked, but enough examples of discoordinate
expression have been shown to suggest independent modes of regulation.87,244,488,509 Collagenase and
stromelysin levels are increased in OA cartilage in levels which correlate to histologic severity, both

29.
regionally and between patients.625 MMPs (especially stromelysin) are also markedly increased in
arthritic synovial fluids and represent a promising marker of diseased joints.144,432,477,879
Synovial tissue is also an important source of MMPs.932 For example, after partial meniscectomy of
rabbits, synovial stromelysin production precedes that of cartilage.536 It has been suggested that the
synovium is the main source of synovial fluid collagenase.323,366 Numerous in situ-hybridization
studies have localised expression of collagenase, stromelysin, and tissue inhibitor of matrix
metalloproteases (TIMP) mRNA to the synovial lining layer.232,284,524,932 Synoviocytes are also
known to produce collagenase-2311 and collagenase-3.895 However, synovial tissues produce
abundant quantities of the endogenous inhibitor TIMP,536 and it seems likely that synovium-derived
MMPs have an effect only on the most superficial layers of cartilage, if at all.
As MMPs are secreted as inactive pro-forms, activation by proteolytic cleavage is an important step
in their regulation.560 The complex cascade of MMP activation is only partially understood.612
Plasmin is thought to be an important activator of MMP-1, -3 and -9, but it inefficiently activates
MMP-2.893 Once activated, stromelysin itself activates MMP-1 and -9;618 its role in MMP-1
activation may be an obligatory step.825 Similarly, collagenase-1 and -3 activate the gelatinases, and
membrane-bound MT-MMPs are thought to be important activators of the MMP-2, -9, and -13.
Endogenous inhibitors such as TIMP are also important regulators of MMP activity. Serum protease
inhibitors such as 2-macroglobulin and 1-antitrypsin are not normally present within the joint, but
may enter synovial fluid in inflammatory states.871

30.
Table 1.1: Classification of the matrix metalloproteases found in articular cartilage.812,885
Name MMP
Number Mass
(kD) Native Substrates (*=major targets)
Collagenase-1 (interstitial collagenase) Collagenase-2 (neutrophil collagenase) Collagenase-3
1 8 13
55 75 65
Collagens I, II, III*, VII, VIII, X Collagens I*, II, III, VII, VIII, XI; proteoglycan core protein Collagens I, II*, III, IV, IX, X, XIV
Gelatinase A Gelatinase B
2 9
72 92
Gelatin*, collagen I, II, IV, V, VII, X,XI, XIV; PG core protein, elastin, laminin, fibronectin; pro-IL-1ß, pro-TNF-
As for gelatinase A Stromelysin-1 Matrilysin
3 7
57 28
Proteoglycan core protein & link protein*; non-helical regions of collagens III,IV,V,IX; laminin, fibronectin; pro-MMP-1 As for stromelysin-1
MT1-MMP 14 63 Collagen I, II, III; fibronectin, laminin, vibronectin; pro-MMP-2, pro-MMP-13
Aggrecanase-1 Aggrecanase-2
ADAM-TS4 ADAM-TS5
? ?
Aggrecan core protein Aggrecan core protein
Collagenase
Collagenase-1, -2, and -3 are all up-regulated in OA cartilage.625,671 The relative roles and
importance of these enzymes are unknown, though some differences in distribution have been
recognized, collagenase-1 being mainly expressed in the superficial layers of OA cartilage.244,505
Contrary to initial suggestions, recent studies indicate that collagenase-3 may play primarily a
remodelling rather than catabolic role.505 Supportive of this is the finding that TGF-ß preferentially
induces collagenase-3 over collagenase-1.820 However, collagenase-3 inhibitors effectively reduce
cartilage collagen damage in arthritis without co-inhibition of MMP-1.121 MMP-1 has not been
identified in rodents, yet these are well-known experimental subjects for arthritis models.121 Such
studies suggest that the true culprit(s) of collagenase degradation in OA is yet to be elucidated.

31.
Gelatinase
The gelatinases are so named for their ability to cleave fibrillar collagens only after their triple
helical structure is already disrupted. However, they may be involved in cleavage of minor collagen
types, and collagen-associated molecules such as fibronectin. MMP-2 (Gelatinase A) and MMP-9
(Gelatinase B) are unusual amongst MMPs in containing a fibronectin-like domain and a type V
collagen-like domain.812 They are regulated independently, with the 72kD MMP-2 being
constitutively produced in cartilage, while the 92kD MMP-9 is selectively expressed in OA, in a
distribution corresponding to fibrillated areas.551 Similarly, MMP-2 is constitutively expressed by
synovial fibroblasts, while MMP-9 is induced by pro-catabolic cytokines.415 MMP-9 is therefore a
useful marker, if not mediator, of cartilage damage and synoviocyte activation.
Stromelysin
Complete enzymatic digestion of proteoglycan is a complex process, probably involving multiple
MMPs. Stromelysin (MMP-3) is thought to be a key enzyme in destabilising aggrecan, through
proteolysis of both the core protein and associated link protein. It can also digest collagens, but
only secondary to initial collagenase attack.87 Stromelysin is found in normal cartilage, but is
greatly elevated in OA to levels correlating with disease severity.323,509,594 It is also expressed in
inflamed synovia, though cartilage expression appears to be more sustained.366
Aggrecanase
Loss of aggrecan from cartilage involves cleavage at the interglobular domain of the core protein. It
was recently discovered that cleavage occurs at two major sites, Asn341-Phe342 and Glu373-Ala374,
yielding the C-terminal neoepitopes VDIPEN and NITEGE respectively. While the VDIPEN
epitope was associated with previously known MMPs, it was proposed that an uncharacterised
enzyme ‘aggrecanase’ might be responsible for generating the NITEGE neoepitope. Aggrecanase-1
and -2 have since been purified and cloned, and their specificity for aggrecan cleavage
confirmed.828 A recent murine study has demonstrated differential distribution of the two

32.
neoepitopes in normal cartilage, with VDIPEN detected mainly around deep zone chondrocytes,
while NITEGE was present mainly in the upper zones. However, both were expressed adjacent to
the spontaneous OA lesions characteristic of STR/ort mice.122 Fragments from aggrecanase
cleavage are the major product of aggrecan catabolism detected in arthritic synovial fluid and
cartilage explant conditioned media, suggesting these enzymes may be a major mediator of
cartilage proteoglycan catabolism.43,478,723
1.1.3.1.2 Tissue Inhibitor of Matrix Metalloproteases (TIMP)
Tissue inhibitors of matrix metalloproteases (TIMP) are proteins which specifically bind to and
irreversibly inactivate MMPs.885 Four forms are currently recognised; of these, TIMP-1, -2 and -3
are found in articular cartilage.509 TIMP-1 may be only form in synovium and synovial fluid,336,505
while TIMP-2 seems to be the primary form elevated in OA cartilage.509 A special relationship has
been shown between TIMP-2 and MMP-2, with the inhibitor binding with high affinity to a region
separate from the enzyme’s active site, facilitating the formation of an MMP-2/TIMP-2/MT-MMP-
1 complex which may be important in MMP-2 activation.270,612 TIMP-1 has a similar association
with MMP-9.612 TIMPs are elevated within the OA joint, but are relatively deficient in comparison
to the proportionally greater increase in MMP expression.479 This is surprising given they are
induced by IL-1ß and similar cytokines via the same Protein Kinase C-dependant mechanisms as
the MMPs.192,193 Inhibition of TIMP expression by co-induced PGE2 may be responsible.193 TGF-ß,
as a particularly powerful inducer of TIMP, opposes this imbalance.130
1.1.3.1.3 Serine and Cysteine Proteases
The serine protease system of plasmin and its endogenous activator, plasminogen activator (PA),
contributes to cartilage degradation (in vitro at least) by activation of latent MMPs.98,150,533 In
addition, both PA and plasmin are thought to directly degrade various matrix components.533,548,744

33.
Plasmin has been shown to activate latent forms of growth factors, to activate IGF by releasing it
from IGFBPs, and to induce leukocyte recruitment independently of its proteolytic effects.102,506,809
PA is also likely to be involved in subchondral bone degradation.695 The involvement of PA in joint
degradation in vivo has not been proven, though Martel-Pelletier et al. (1991)507 found a significant
correlation between plasmin activity and active collagenase in human OA cartilage. In the only
such trial reported, Kikuchi et al. (1987)416 reported clinical improvement of RA patients following
intra-articular injection of a PA inhibitor.
Levels of PA and plasmin are both elevated in OA cartilage.507 PA is induced by IL-1 and TNF-
in chondrocytes98,106,212 and synoviocytes.308,459,529 PGE2 may partially mediate the effects of these
cytokines, as non-steroidal anti-inflammatory drugs (NSAIDs) partially reverse PA
induction.306,459,549 Leizer and Hamilton (1989)460 described a human monocyte-derived factor
capable of inducing PA activity in synovial fibroblast cultures, which was immunologically and
functionally distinct from known cytokines. This may have been TGF-ß, which can similarly
induce PA without concurrent induction of PGE2.307 Plasminogen is not produced by joint tissues.98
Two distinct forms of PA are generally described: urokinase-type (uPA) and tissue-type PA (tPA).
Early experiments (using a radio-labelled fibrin assay) identified tPA as the more commonly
induced form in IL-1-stimulated chondrocytes.100,106,150 More recent studies have shown uPA to be
the predominant form in OA cartilage and synovial fluid.416,507 In OA synovia, uPA also appears to
be the main form induced, while tPA is more commonly expressed by normal synoviocytes and
may actually be down-regulated in OA.102,535 This shift from tPA to uPA may reflect a change in
the role of synovial PA from fibrinolysis to matrix degradation.694 uPA can be activated at the cell
surface by binding to uPA-receptors (uPAR), producing local pericellular proteolysis. uPAR are
present on both synoviocytes and chondrocytes and are up-regulated in OA.102,744 tPA is activated

34.
by binding to fibrin.102 In the inhibitor-rich environment of the normal joint, cell-associated uPA
probably represents the most active pool.694 With increasing cytokine stimulation, secreted PA
comprises a greater proportion of total PA activity.212,308
Serine protease inhibitors are plentiful in cartilage.261 Two forms of PA inhibitor (PAI) are present
in joint tissues: PAI-1 which primarily inhibits tPA, and PAI-2 which more effectively inhibits
uPA.102,543 Synovial tissue produces an excess of PAI over PA, hence synovial fluid does not show
fibrinolytic activity without extraction procedures.416,626 PAI-1 levels are decreased in OA
cartilage,507 which possibly contributes to the increase in non-cell-associated PA described above.
In addition, poorly characterised low molecular weight (12 kD) serine protease inhibitors are
known to be endemic in cartilage.30
Interest is reviving in certain cysteine proteases such as cathepsin B, which is thought to be capable
of activating plasminogen activator and stromelysin-1. Cathepsin B and Cathepsin D are both
significantly up-regulated in OA synovia.249,537
1.1.3.2 Cytokines
The importance of cytokines in the pathogenesis of OA has become increasingly clear over the last
few decades of research. It is now thought that differences in the cytokine profiles of OA and RA
are more quantitative than qualitative; the common practice of using OA-derived synovial fluids as
a ‘non-inflammatory’ control is therefore dubious.876,896 Current evidence strongly suggests that IL-
1ß, and to some extent TNF- , initiate many of the catabolic events in OA joint tissues, such as the
production and activation of proteases, depressed or deranged matrix biosynthesis, and release of
inflammatory mediators such as nitric oxide and prostaglandins. 505 It is unclear whether IL-1 and
TNF- act in concert, or whether a functional hierarchy exists between these and other
cytokines.618 Some authors suggest that IL-1 is primarily responsible for cartilage destruction,

35.
while TNF- primarily mediates joint inflammation.505 Van den Berg (1998)857 has demonstrated
that the effects of cytokines on joint inflammation and cartilage destruction can occur uncoupled.
For example, cytokines such as IL-4, IL-6, IL-10 and TGF-ß enhance inflammation but potentially
reduce cartilage damage. As small soluble mediators, cytokines are central to the tissue 'cross-talk'
that occurs between cartilage, synovium, and bone within the OA joint. However, this highlights a
gap in current knowledge, in that the cytokine-driven model of OA fails to fully explain the focal
cartilage damage so characteristic of the disease.896
Cytokines are generally absent or present in only low levels in normal tissue, but are dramatically
up-regulated in response to stimuli such as cellular injury. The subsequent action of cytokines is
tissue-dependent, and is potentially controlled by many different mechanisms, such as the type and
quantity of specific cell-surface receptors. The presence of specific receptor antagonists, soluble
'decoy' receptors, or antagonistic cytokines may act to block these responses.394 Some cytokines
also require enzymatic cleavage for full activity. Post-receptor signalling is thought to represent
another important level of modulation, but remains poorly understood.
1.1.3.2.1 Interleukin-1 (IL-1)
Early experiments with conditioned media from cultured synovium222,534 and peripheral blood
mononuclear cells172,275 demonstrated the existence of a soluble factor which induced the release of
inflammatory and catabolic factors from cartilage and synovium. This 17kD factor was isolated
from porcine synovium and named ‘catabolin’.196 It was subsequently demonstrated in the early
1980’s that this factor was probably interleukin-1.175,529,712,831,913,914 Subsequent in vivo studies using
recombinant IL-1ß have shown it is capable of inducing, even from a single intra-articular
injection, many of the changes characteristic of OA such as loss of cartilage proteoglycan, reduced
matrix synthesis, and cellular infiltration of synovial fluid.42,528 Gene transfer-induced over-
expression of IL-1 in rabbits has been shown to produce a severe, RA-like disease.254 Conversely,

36.
blocking IL-1ß activity is extremely effective at preventing cartilage destruction.855 IL-1 is
currently recognised as the primary cytokine driving degenerative changes within the OA joint.
Other cytokine systems may predominate in more inflammatory arthritides, as illustrated by a
recent study showing significant differences in the relative importance of IL-1 in adjuvant arthritis
and collagen-induced arthritis in rats.70
Two forms of IL-1 are recognised, IL-1 and IL-1ß, though they share little homology (~27%) and
are products of separate genes.459 IL-1 is thought to be mainly cell-associated, while IL-1ß is
predominantly soluble and is the principal form produced within OA joints.276,428,576,627 The effects
of both are similar, though IL-1ß exhibits greater activity on an equimolar basis.922 IL-1
simultaneously suppresses cartilage matrix biosynthesis and stimulates matrix degradation, though
chondrocytes are much more sensitive to the former effect than the latter, especially in human
cartilage.259,822 The mechanism of this suppression is unknown, but possibly relates to reduced
expression of galactose-ß-1,3-glycuronosyltransferase I, a key enzyme in glycosaminoglycan
synthesis.278 IL-1 also suppresses synthesis of other matrix components, notably type II collagen.684
Conversely, IL-1 promotes chondrocyte production of the normally fibroblast-associated type I and
III collagen, but only when concurrent induction of PGE2 is suppressed.273 IL-1 is also a major co-
factor in inducing the IGF-1 hyporesponsiveness commonly present in OA, though this may be via
indirect means.867 At higher concentrations, MMP and TIMP are simultaneously but
disproportionately induced, resulting in an increased MMP/TIMP ratio.508,684 A similar change
increases the uPA / PAI-1 ratio.507 IL-1 is also responsible for many of the inflammatory changes in
OA synovium, inducing synthesis of collagenase, stromelysin, IL-6, PGE2, and
hyaluronan.114,301,435,627,684 IL-1 induces proliferation of synoviocytes, but simultaneously sensitises
them (possibly via NO) to Fas-mediated apoptosis, hence its mitogenic action is variable.427,773
Cellular release of IL-1 is induced by such varied stimuli as mechanical alteration, soluble matrix
breakdown products, collagen (in vitro), TNF- , and IL-1 itself (i.e. it is autoinductive).130 IL-1

37.
release is inhibited by PGE2. and certain leukotrienes.92 Both chondrocytes and synoviocytes are
capable of producing significant amounts of IL-1, creating debate as to which tissue is the more
important source. IL-1 within synovial fluid is thought to derive from synovium, produced as part
of a theoretical cartilage-synovium feedback loop in response to phagocytosed matrix breakdown
products. Supportive of this view is the predominant immunolocalisation of IL-1ß within the
superficial layer of OA cartilage.511,620,830 IL-1 is diffusely expressed throughout the lining layer of
OA synovia,182,214,511 though it can also be detected in normal tissue.182 However, several studies
have shown very low or undetectable levels of IL-1ß in OA synovial fluid, suggest that synovium-
derived IL-1 is unlikely to drive cartilage damage.74,105,865,896,898 Autocrine production by
chondrocytes is now recognised as significant and represents a more likely source of cartilage IL-
1.49,538
IL-1ß requires proteolytic maturation for full bioactivity, a function performed by the unique
cysteine protease interleukin-1ß converting enzyme (ICE).428 The cellular effects of IL-1 are
receptor-mediated. Cell membrane receptors of 80kD (Type I) and 68kD (type II) are recognised,
the smaller form possibly acting as a ‘decoy’ rather than a true signalling receptor.392 Only low
levels of receptor occupancy are necessary for maximal catabolic responses.627 Human
chondrocytes primarily express the Type I receptor, with cells derived from the superficial zone
expressing approximately twice the number of binding sites and are hence being more susceptible
to the effects of IL-1 than those from deeper layers.325 Receptor expression is also increased in OA
chondrocytes relative to normal cells.508,627 Human synoviocytes have been reported to express both
receptor types, with up-regulation of Type I IL-1 receptors in OA.709
IL-1 receptor antagonist (IL-1Ra) was first characterised in 1990, at that time the first description
of a naturally-occurring cytokine receptor antagonist.206 Since then other (mainly intracellular)
forms have been described.56,493 A large excess of IL-1Ra over IL-1ß is required to reduce receptor
occupancy sufficiently to block activity. IL-1Ra is induced in synovium by IL-1ß itself, a process

38.
which is enhanced by other cytokines such as interferon- and IL-4.133,510,741 However, IL-1Ra
production is deficient relative to IL-1 release, increasing the agonist:antagonist ratio.105,133,182,392,768
The post-receptor signalling mechanisms of IL-1 are poorly understood. Its effects on matrix
synthesis, matrix degradation, and PGE2 release all seem to be mediated through independent
mechanisms, as reflected by differences in dose and time dependence. The induction of the MMP
and plasmin systems involves Protein Kinase C-dependant mechanisms.193 IL-1-induced
suppression of proteoglycan synthesis is partially reduced by Protein Kinase C activation, though
PGE2 release is unaffected.44 Numerous tyrosine kinases and oncogene products (e.g. c-fos, c-jun)
are also involved in IL-1 signal transduction.
1.1.3.2.2 Tumour Necrosis Factor- (TNF- )
TNF- is generally described as having similar effects to IL-1 but with lesser potency, inducing the
release of MMPs, IL-6, PGE2, and superoxide by target cells.8,12,814,922 IL-1ß and TNF stimulate
production of each other,92,753 and it has been suggested that many of the IL-1-like actions of
TNF in vitro may actually be due to induction of endogenous IL-1. However, others have found
this effect to be limited.92,440 It has also been suggested that there is some synergy of their cartilage
degradative effects when both IL-1 and TNF- are present.340,542,550,713 While IL-1 and TNF- are
generally induced by the same stimuli, this may not always be the case in OA cartilage, as positive
correlations between the two cytokines are surprisingly lacking.896
TNF- is generally present in only low levels in OA joints,627 though it is thought to be the
dominant cytokine produced within rheumatoid joints.220 It has been shown to be the major
mediator of joint swelling and inflammation, but not cartilage destruction, in inflammatory rodent
models of arthritis.440,713,857 Intra-articular injection of TNF- in rabbits induces profound
(predominantly monocytic) cellular infiltration of the synovium, but causes minimal proteoglycan

39.
loss.340 Bryson et al. (1998)98 found that while a uPA inhibitor was potent in preventing
proteoglycan release from TNF- -stimulated cartilage, it was less effective against IL-1ß-induced
degradation, suggesting a difference in the pathways by which these cytokines induce proteolysis.
TNF- is synthesised in a latent form that requires proteolytic cleavage for activation. TNF- -
converting enzyme (TACE), otherwise known as ADAM-17, is one of several MMPs capable of
cleaving the membrane-bound latent TNF- into a soluble trimer of 17kD subunits.320 Two
receptors, TNF-R55 and TNF-R75, are described according to their molecular size of 55kD and
75kD respectively. The 55kD form has been shown to mediate cellular responses to TNF- , and is
up-regulated in OA chondrocytes and synovial fibroblasts.13,886 Corresponding soluble forms are
described, TNF-sR55 and TNF-sR75, with a relative increase in synovial fibroblast TNF-sR75 release
seen following IL-1ß or TNF- stimulation. However it is currently unknown whether these
binding proteins function to transport or to sequester active TNF- ligand, thereby promoting or
antagonising its actions.512
1.1.3.2.3 Interleukin-6 (IL-6)
This multi-functional cytokine is best known for its role in inducing hepatic release of acute phase
proteins, and stimulation of B-lymphocyte proliferation. Given these effects, circulating synovium-
derived IL-6 may mediate some of the systemic effects recognised in arthritis, such as elevated
levels of gamma globulins and acute phase proteins such as 1-antitrypsin.296 In cartilage, IL-6 has
been described as a natural chondroprotective agent that attenuates the actions of more catabolic
mediators such as IL-1ß, and promotes chondrocyte proliferation and biosynthetic activity.259
However it is also potentially a catabolic mediator in that it promotes synovial inflammation and
local bone resorption.296

40.
Serum levels of IL-6 are elevated in OA patients.401 Similarly, patients with active RA have
significantly higher serum levels of IL-6 than those with suppressed disease.11 Synovial fluid IL-6
is elevated following experimental canine ACLT, and correlates strongly with increased
proteoglycan synthesis.865 The synovial membrane is a rich source of IL-6,64,296,547,698 especially in
RA.229,296,698 Immunolocalisation studies have found IL-6 is mainly associated with macrophage-
like (Type A) cells in the synovial lining,229 but also correlates histologically with plasma cell
infiltration.229,547 Articular chondrocytes are also an important source of IL-6.295 IL-1ß appears to be
the most potent stimulus for its production in vitro,132,295,627 though TNF- , TGF-ß, interferon- , IL-
4, IL-10, IL-13, insulin, and bFGF all increase IL-6 release.259,295,296,583,698 Paracrine stimuli
affecting IL-6 production appear complex - the addition of monocytes, but not monocyte
conditioned medium (MCM), increases synovial fibroblast IL-6 production in the absence of
detectable levels of IL-1ß or TNF- .128 Studies of the IL-6 gene promoter have revealed control by
multiple transcription factors including NF- ß, AP-1, and Protein Kinase A and C.64,933
IL-6 has multiple, somewhat pleiomorphic effects within the OA joint. It promotes the infiltration
of inflammatory cells into synovium, and may also regulate synovial fibroblast proliferation.296 In
synovial fluid, levels of IL-6 correlate with inflammatory indices such as leukocyte count and total
protein content.74 However, IL-6 has no direct effect on PGE2, glycosaminoglycan, or collagen
synthesis by synoviocytes.375,480 The primary receptor IL-6R is reportedly absent from synovial
fibroblasts; however, soluble receptor/cytokine complexes in synovial fluid have the potential to
interact with the gp130 subunit, inducing cell proliferation.64,375 In cartilage, IL-6 stimulates
chondrocyte proliferation,295,865 and may be a major stimulus for the formation of chondrocyte
‘clones’ in OA cartilage. Its effect on proteoglycan synthesis is unclear, having been shown to both
inhibit and increase synthesis under different conditions.344,583 Nietfeld et al. (1990)583 found that
while IL-6 alone was a weak inhibitor of proteoglycan synthesis, blocking antibodies to IL-6 were
able to significantly reduce the effect of IL-1 on matrix synthesis. Its role in proteolytic damage is
similarly unclear. IL-6 alone increases TIMP production without increasing MMP production,

41.
suggesting its net influence is probably to limit proteolytic damage.375,480,618,865 However, IL-6
augments IL-1ß-stimulated MMP production in a dose-dependant manner.234,375 IL-6 also induces
TGF-ß, IL-1Ra, TNF-sR55, uPA, and PAI-1 release, while reducing IL-8 production.344,375,857
Together, this suggests IL-6 may play an important regulatory role in amplifying the intermediate
events which follow IL-1ß stimulation.583
1.1.3.2.4 Other cytokines
The following cytokines have also been identified as having a potential role in the pathogenesis of
OA:
Interleukin-4 (IL-4) exhibits predominantly anti-inflammatory effects on synovial fibroblasts,
reducing IL-1ß, leukocyte inhibitory factor (LIF), IL-8, PGE2, collagenase and stromelysin release,
while increasing expression of IL-1Ra.85,133,178,741,823,857 IL-4 has been shown to reduce TNF-R55
levels in rheumatoid synovial fibroblasts by promoting receptor shedding, and this may represent
one mode by which this monokine antagonises the activities of pro-inflammatory cytokines.823
However synovial fibroblasts were found not to express IL-4 receptors in one study.183
Interleukin-8 (IL-8) is known primarily for its effects on neutrophil chemotaxis. It is released
constitutively by OA synovial fibroblasts, but is greatly up-regulated after IL-1ß stimulation.741
Chondrocytes may also be an important reservoir of IL-8.344
Interleukin-10 (IL-10) plays a predominantly anti-inflammatory and immunoregulatory role in
leukocyte cross-talk, and its over-expression by activated T-cells may contribute to the endogenous
immunosuppression present in the rheumatoid joint.147,408 IL-10 inhibits iNOS induction and
increases proteoglycan synthesis in cartilage. While it increases collagenase and stromelysin
production by skin fibroblasts,677 IL-10 has been shown to reduce cyclooxygenase-2 (COX-2) and

42.
phospholipase-A2 expression and increase IL-1Ra release by isolated synovial fibroblasts,12,742 and
to reduce synovial activation in the SCID mouse model.557
Interleukin-11 (IL-11) is elevated in synovial fluid of OA patients.832 It may play a protective role,
by inducing TIMP release.596
Interleukin-13 (IL-13) may be an important anti-inflammatory cytokine, at least in synovial tissue.
It reduces IL-1ß, TNF- , stromelysin, COX-2 and PGE2 production by OA synovial fibroblasts,
and increases IL-1Ra release.12,394
Interleukin-17 (IL-17) is a recently discovered cytokine induced by IL-1ß and TNF- in some cell
types.505 While it has been shown to increase iNOS expression in articular cartilage, its net role is
OA in not known.51,618 IL-17 may mediate the osteoclastic bone resorption associated with some
forms of arthritis.749
Leukocyte inhibitory factor (LIF), though a member of the IL-6 superfamily, has a clearer catabolic
role in cartilage, by stimulating proteoglycan resorption, MMP synthesis, and NO production. LIF
is up-regulated in both OA synovia and chondrocytes, and is induced by IL-1 and TNF- .309,482 LIF
may potentially form a positive feedback loop with IL-1ß, which would naturally amplify the
'activated' state of OA chondrocytes.344
1.1.3.2.5 Growth Factors
The degradative effect of catabolic cytokines is inhibited or antagonised by a range of anabolic
mediators (generally termed growth factors) which act to promote connective tissue formation, and
to maintain the proliferative and/or biosynthetic activity of connective tissue cells. In cartilage, the
balance between anabolic and catabolic cytokines regulates the homeostatic turnover of cartilage

43.
matrix components. In particular, TGF-ß and IGF-1 have been identified as potent promoters of
cartilage anabolism, stimulating chondrocyte proliferation, matrix biosynthesis, and antagonising
the deleterious effects of cytokines such as IL-1ß. It has been speculated that reduced production
of, or responsiveness to, such growth factors may contribute to the increased incidence of OA in
aged patients.896
Transforming growth factor-ß (TGF-ß)
TGF-ß is abundantly expressed in normal and inflamed synovial tissue,93,136,446,558,563,817,878 and is
spontaneously produced by synoviocytes in culture.446 IL-6 may be an important stimulus of its
increased expression in arthritic synovia.870 Synovial fluid levels of TGF-ß are elevated in arthritic
conditions.93,465,826 Cartilage also contains large quantities of TGF-ß, especially in OA.864 TGF-ß
has been described as pleiotropic in its actions, causing seemingly different effects depending on
experimental conditions and target cell type.670,754,857 TGF-ß is known to stimulate chondrocyte
proliferation, collagen and proteoglycan synthesis,650,653,672,673 and TIMP expression,804 and to
down-regulate IL-1 receptor expression.675 Its effect in inducing proteoglycan synthesis is potent;
however TGF-ß does not appear to have any additional effect in combination with IGF-1.852 In
cartilage, TGF-ß is protective of IL-1-induced degradation, partially reversing the inhibition of
proteoglycan synthesis29,124,268,852,857 and reducing proteolytic activity.124 It causes similar responses
in synoviocytes,446,916 also reducing collagenase expression,516 an effect which may be via down-
regulation of the c-jun family of oncogenes as has been proposed in dermal fibroblasts.516 In
gingival fibroblasts, TGF-ß independently reduces collagenase production and increases TIMP and
PAI-1 levels, but increases production of MMP-2.603 Synoviocyte production of hyaluronan, and its
molecular weight, is decreased by TGF-ß in vitro.410 It has also been shown to be a strong inhibitor
of nitric oxide production in cultured rabbit synoviocytes and other cell types.795,822 The net effect
of these diverse activities in the osteoarthritic joint is difficult to determine.

44.
As TGF-ß is secreted in latent form and most cells express TGF-ß receptors constitutively, its
activation by proteases such as plasmin and cathepsin may be an important regulatory step.446,506
Various isoforms of TGF-ß have different distributions within the synovial membrane: TGF-ß1 and
TGF-ß2 seem associated with synovial lining cells, fibroblasts, and endothelial cells,136,563 while
TGF-ß3 is localised to macrophages of the sublining layer,563 and appears to be selectively induced
by IL-1ß.870
Insulin-like growth factor (IGF)
The insulin-like growth factors are small peptides highly homologous with pro-insulin.506 In
particular, IGF-1 seems to be one of the most important cytokines acting to maintain normal
proteoglycan synthesis,506,576,727 and has been shown to be the primary cartilage anabolic factor
present in serum and synovial fluid.245 Though it only partially antagonises IL-1ß-induced
suppression of proteoglycan synthesis, it accelerates cartilage recovery when fluctuating IL-1ß
levels fall below the threshold required for suppression.576 IGF-2 has similar properties but is
generally associated with embryonic stages of growth,859 and may be involved in mechanical
strain-related proliferation of certain cell types.129 Chondrocyte production of IGF-1 is increased in
OA, with elevated levels in arthritic synovial fluid.202,735 IGF-1 has been shown to auto-induce
further IGF-1transcription.586 While IGF levels are up-regulated in OA cartilage, cellular
responsiveness is concurrently reduced, a phenomenon often attributed to the presence of IGF
binding proteins.506,821 IGFBP-2, -3, and -4 are all increased in osteoarthritic cartilage, their
expression induced by IGF-1 itself.131,821 However, Chevalier and Tyler (1996)131 found that
recombinant IGFBP-3 improved anabolic responses to IGF in human cartilage explants.
Epidemiological studies have found some evidence of an association between serum IGF levels and
osteoarthritis. While two studies have found no association between high IGF levels and knee
OA,237,354 others have identified positive correlations with osteophyte growth736 and more severe,

45.
bilateral disease.476 The latter study also found an association with OA of the distal interphalangeal
joint, a common manifestation of so-called 'generalised OA'.476 Any association between IGF and
OA may relate to its role in promoting greater bone density, as circulating IGF-1 has been shown to
correlate positively with skeletal bone mineral density.868 Osteoporosis has been linked with low
serum levels of IGF-1 and IGFBP-3,506 though no association was found in the large Chingford
study group.476 IGF may be a particularly important mediator of the subchondral bone sclerosis that
often accompanies OA.184,506,896
Exposure of vascular epithelium to IGF-1 causes rapid induction of nitric oxide release, apparently
by induction of iNOS.332,559,732,839 However IGF-1 apparently inhibits IL-1ß-induced iNOS
induction within the circulation.732,733 The opposite has been shown in renal mesangial cells.294 The
influence of IGF on iNOS induction in joint tissues has not been studied.
Other growth factors
Epidermal growth factor (EGF) has been detected in high levels in rheumatoid synovia, where it
may contribute to thickening and neovascularisation of the lining layer.214 In cultured fibroblasts, it
increases hyaluronan synthesis461 but antagonises the effect of TGF-ß on collagen synthesis.687 In
cartilage, EGF may decrease proteoglycan synthesis while stimulating chondrocyte proliferation.650
Platelet-derived growth factor (PDGF) is a potent stimulus for chondrocyte and synovial
proliferation.441,651 It prevents induction of iNOS by IL-1ß, but not TNF- .795 Direct oncogene-
triggered 'transformation' of synovial cells has been demonstrated following PDGF exposure.445
Basic Fibroblast growth factor (bFGF) is an autocrine growth factor produced by both
synoviocytes and chondrocytes.558 It has both anabolic and catabolic functions in matrix
homeostasis.670

46.
Vascular endothelial growth factor (VEGF) is secreted in significant amounts by rheumatoid and
OA synovial fibroblasts, especially after induction by IL-1ß or hypoxia.73,377 It is likely to be an
important stimulus for synovial angiogenesis, which is important in sustaining chronic proliferative
inflammatory states such as RA.
1.1.3.3 Nitric Oxide
Nitric oxide (NO) is a highly reactive, gaseous free radical which is emerging as an important
mediator in many physiological and pathophysiological processes, such as vascular tone, neuronal
transmission, platelet function, immune function, and angiogenesis.211,215,381,552,936 NO was initially
identified as the long sought endothelial-derived relaxation factor (EDRF) and is probably the
major influence on basal vascular tone, producing vasodilation. Its small size and unpaired electron
make NO a highly diffusible and reactive molecule, which passes readily through cell membranes
to act as a local signalling or effector mechanism.381 It has a very short half life, in the order of
seconds. However, associated products such as peroxynitrite and S-nitrosothiols have longer half-
lives and may mediate some of its effects.292,329 Experimentally, the stable intermediates nitrite and
nitrate are usually measured as an index of NO production itself.
NO is generated enzymatically by nitric oxide synthase (NOS) by oxidation of L-arginine, yielding
L-citrulline as a coproduct (Figure 1.5). To date three NOS forms have been recognized: a neuronal
(nNOS) and endothelial form (eNOS), both of which are constitutively expressed and regulated by
intracellular Ca2+ concentration, and an inducible form (iNOS) found in many cell types, which is
instead regulated at the transcriptional level.211 iNOS is not normally present in cells, but is induced
in inflammatory states in response to cytokines such as IL-1ß, TNF- , and gamma interferon, or
bacterial products such as endotoxin.381 NO production by iNOS is several orders of magnitude
greater than that of constitutive forms.215

47.
Figure 1.5: The enzymatic production of NO from arginine, as catalysed by nitric oxide synthase.
NADP, nicotinamide adenine dinucleotide phospate.
There is strong evidence that NO plays a role in arthritic conditions.381 Synovial fluid levels of
nitrite/nitrate are elevated in arthritic joints,216,378,680,815 as is the level in serum and urine.216,281,786,799
The main source of NO within the OA joint appears to be chondrocytes themselves.7,538,676 Standler
et al. (1991)791 first showed that like many other cell types, articular chondrocytes produce NO in
response to cytokines. Many subsequent studies have confirmed the expression of iNOS in OA
cartilage and chondrocytes in a number of species.25,49,538,609,637,676 OA cartilage demonstrates
greater capacity for NO production than normal cartilage, with IL-1ß apparently the main stimulus,
though TNF- and the newly-discovered IL-17 may also be important.51,538,637,785 Superficial
chondrocytes produce significantly more NO than chondrocytes derived from deeper
cartilage.139,326,329
Studies in rheumatoid patients demonstrate that synovial tissue also has the capacity for high output
of NO given the correct stimulus.532 Immunohistochemical studies have confirmed synovial iNOS
expression but differ over the cell types involved.282,538,714 McInnes et al. (1996)532 clarified this
issue by demonstrating, using an antiserum specifically raised against human iNOS, that synovial
fibroblasts (Type B synoviocytes) were the major expressors of iNOS in human synovia. They
showed substantial spontaneous and enterotoxin-stimulated NO production in primary synovial
L-Arginine L-citrulline + NO•
O2 , NADPH H2O, NADP+
NITRIC OXIDE SYNTHASE

48.
cultures sourced from both RA and OA patients. Other studies have found NO production by
synovial cultures to be inconsistent.676,714,795 This may be due to species differences, or may
indicate a requirement for a more complex cytokine stimulus than is usually provided
experimentally.894
Cells within fibrous structures such as menisci may also represent an important source of intra-
articular NO production.109,426,457,813 A recent rabbit study observed that nitric oxide release by
meniscal explants was much greater than that of synovial tissue.813 Cao et al. (1998)109 suggested
that within menisci, chondrocytic cells were responsible for most cytokine-stimulated NO
production. However, the observation of substantial NO release by canine cruciate ligaments
confirms that fibroblasts may also contribute to intra-articular levels.785
1.1.3.3.1 Nitric Oxide and Cartilage
The effect of NO induction within the arthritic joint is unclear. There is mounting evidence that it
plays a catabolic role, but also some evidence of chondroprotection, suggesting that its effect may
differ in matrix homeostasis and inflammatory states.381 Taskiran et al. (1994)822 first suggested
that NO mediates much of the effect of IL-1ß in suppressing proteoglycan. Recently, transfection
of rabbit chondrocytes with the iNOS gene confirmed the ability of NO to inhibit matrix production
independently of IL-1.801 However, the magnitude of this effect is not entirely consistent and varies
between species,349 culture systems,393 and chondrocytes derived from different zones.139,326,327,346,794
The mechanism of inhibition is unknown, but may be via interference with glycosaminoglycan
sulphation,349 cytoskeletal integrin-signalling complexes,141 or mitochondrial respiration.386,827
Alternatively, inhibition of matrix synthesis may be primarily by prevention of anabolic responses
mediated by endogenous cartilage growth factors. Arthritic cartilage is known to be
hyporesponsive to IGF-1 and recent studies have identified NO as a key element of this
phenomenon, possibly by inhibition of receptor autophosphorylation.803,858 NO also inhibits

49.
chondrocyte TGF-ß production, to similar effect.801,802 In addition to suppression of proteoglycan
synthesis, Cao et al. (1997)110 showed that NO reduces type II collagen production, possibly by
inhibition of prolyl-hydroxylase. NO has also been reported to reduce IL-1ß receptor antagonist
(IL-1Ra) expression and enhance cartilage MMP activity.637 Murrell et al. (1995)561 reported
inhibition of collagenolytic and caseinolytic activity by the NOS inhibitor NG-nitro-L-arginine
methylester (L-NAME), and induction of activity by the NO-donor S-nitrosylacetylpenicillamine
(SNAP), in cultured bovine chondrocytes and cartilage explants as well as tissue from a single
human patient. However, Bird et al. (2000)76 recently reported that NO inhibited proteoglycan
degradation in equine cartilage explants, specifically demonstrating reduced production of
aggrecanase-generated proteoglycan fragments of the NITEGE neoepitope.
Other cartilage studies have similarly demonstrated that NO may confer some protection against
the degradative effects of IL-1ß. Stefanovic-Racic et al. (1996)794 found that in bovine cartilage,
NO had no effect on PG synthesis, and that the competitive NOS inhibitor L-NG-
monomethylarginine (L-NMA) enhanced early IL1-induced catabolism. This protective effect has
also been demonstrated in human and rabbit articular cartilage,327,793 and lapine menisci.109
Endogenous NO also downregulates chondrocyte production of IL-6 and IL-8, cytokines which
themselves play both pro- and anti-catabolic roles.346 These protective effects of NO may be due to
disruption of anchorage-dependent integrin signalling, thus acting as a non-specific ‘brake’ in
preventing both anabolic and catabolic responses.140
NO has also been linked to chondrocyte apoptosis.78,321,481 However, in vivo evidence for this relies
mainly on regional correlations between apoptosis and NO or peroxynitrate formation,109,426 and as
such remains circumstantial. In vitro evidence of NO-mediated apoptosis has not been entirely
consistent, possibly due to co-dependence on other oxygen radicals.79 Interestingly, Notoya et al.
(2000)588 recently suggested that PGE2 was necessary to sensitise human OA chondrocytes to NO-
induced apoptosis.

50.
1.1.3.3.2 Nitric Oxide and Synovium
The effect of exogenous NO on synoviocytes has not been adequately examined. NO certainly has
the potential to be an important mediator of inflammation, by modulating events as diverse as
blood flow, endothelial permeability, leucocyte migration, and immune cell activation. Via its
actions on blood vessels, NO may be involved synovial hyperaemia and joint swelling.282 However,
its direct metabolic effects on synoviocytes are unclear. In rabbit synovial fibroblasts, L–NMA had
no effect on PGE2, MMP or lactate production,7 though NO was shown to enhance MMP-2
production in rheumatoid human synoviocytes.353 NO has also been shown to induce TNF-
production in cultured synoviocyte and macrophage lines.352,532 Were this to occur in vivo, NO may
represent an important activator of synovial inflammation.
1.1.3.3.3 Nitric Oxide and Bone
NO has recently been identified as an important regulator of bone, another in the long list of
mediators involved in cell-to-cell signalling between osteoblasts and osteoclasts. Human and rat
osteoblasts have been shown to produce NO in response to cytokine stimulation.362,483,665 In mouse
calvarial bone cultures, such induction of NO is associated with reduced bone resorption.664
Induction of high levels of NO may represent a protective mechanism against cytokine-stimulated
bone resorption in pathological conditions.483 Direct studies in cultured osteoclasts have confirmed
that NO is a potent inhibitor of osteoclast function,88,405,486 and of osteoclast differentiation from
precursor cell types.88 NO (in culture at least) may be partly responsible for cytokine-associated
inhibition of osteoblast proliferation and activity,171,362,665 and at high concentrations, osteoblast cell
death.170 However, as in osteoclasts, constitutive production of small amounts of NO appears to be
needed for osteoblast growth.211

51.
While the in vitro effects of NO in bone are somewhat variable,483,664 its dominant role in vivo
appears to be tonic inhibitory restraint of bone resorption. This is supported by the observation that
in rats, the iNOS inhibitor aminoguanidine potentiates loss of bone mineral density.405 while
Wimalawansa and co-workers have demonstrated that topical use of a NO donor (glyceryl
trinitrate) alleviates ovariectomy-induced bone loss in rats.903,905,906 A subsequent study showed that
administration of exogenous NO also counteracts prednisolone-induced bone loss in rats.904 NO
may also be involved in the responses of bone to mechanical stimuli134,644 and fracture healing.199
Evidence from iNOS deficient mice suggests that NO may also contribute to osteophyte formation,
possibly via its influence on TGF-ß production.858 NO is also thought to promote angiogenesis in
some tissues, by feedback interaction with fibroblast growth factor-2 (FGF-2).937
1.1.3.4 Eicosanoids
Prostaglandins are important mediators of classical inflammatory changes such as pain,
vasodilation, increased vascular permeability, and immune responses.542 Prostaglandins are
generated from cell membrane phospholipids in an enzyme cascade involving phospholipase A2
(PLA2)-generated arachidonic acid, which is converted by cyclooxygenase (COX) to various
prostaglandins. As with NOS, both constitutively expressed and inducible forms of COX are now
recognised. COX-1 is expressed under basal conditions in many cells, and is generally prescribed
‘housekeeping’ functions, of particular importance in the gastrointestinal tract, platelets, renal
medulla, and vascular endothelium. In contrast, a second form of cyclooxygenase, COX-2, is not
expressed under normal conditions in most tissues, but is strongly induced by a variety of
inflammatory stimuli.575
Production of eicosanoids by synovial tissue has been recognized and studied for decades.530,534
PGE2 is the predominant eicosanoid produce by synoviocytes, though others including PGI2
(prostacyclin) are produced in appreciable amounts.172,305,689,715 Early researchers noted that PGE2

52.
production was increased in synovial fibroblasts derived from rheumatoid joints, with IL-1ß
subsequently identified as the major stimulus for its production.56,95,114,175,480 Induction of COX-2 is
currently thought to be the prime determinant of synovial PGE2 release,371,618,688 though other
mechanisms have been proposed, including increased expression of a secretory subtype of PLA2,379
and significant levels of COX-1 are also present.159
Though Meats et al.534 first reported in 1980 that chondrocytes in culture could be stimulated to
produce large amounts of PGE2, the potential importance of cartilage prostaglandin production has
only recently been recognized. Subsequent studies have recognised that while some COX-1 activity
is present in normal chondrocytes, COX-2 is indeed induced in cytokine-stimulated cartilage.77
Much interest was sparked by the report of Amin et al. (1997),22 who found that COX-2 is
‘superinduced’ in OA cartilage – to the extent that the spontaneous release of PGE2 by OA
cartilage may be an order of magnitude greater than that of even IL-1ß-stimulated normal cartilage.
This raises the prospect that cartilage may be the major source of PGE2 within the OA joint, though
a recent study in horses suggested synovial tissue was instead the major producer.874 The
proportionately greater cell mass in synovium, particularly in inflammatory states, increases its
contribution to synovial fluid PGE2 levels.
1.1.3.4.1 Effect of Prostaglandin E2 (PGE2) on Joint Tissues
In arthritic tissues, PGE2 has been traditionally thought of as a pro-inflammatory mediator.
Certainly, it is a key driver of inflammatory changes in the synovial microvasculature, such as
hyperaemia and increased permeability to proteins.759 In culture, PGE2 induces some of the major
phenotypic changes of synoviocytes derived from arthritic joints. A PGE2-mediated increase in
intracellular cAMP seems to be responsible for the characteristic stellate or ‘dendritic’ morphology
of cytokine-stimulated synovial fibroblasts in vitro.55 Also, PGE2 is responsible for the reduced
growth rate of rheumatoid synovial fibroblasts in culture, as IL-1ß or platelet-derived growth factor

53.
(PDGF) are mitogenic only when cyclooxygenase is inhibited.95,441 However, as PGE2-induced
Protein Kinase A generally opposes the (mainly catabolic) Protein Kinase C dependant
pathways,193 the net effect of PGE2 is generally to oppose the actions of pro-inflammatory
cytokines. In synovial tissue, PGE2 increases hyaluronic acid synthesis,168 reduces TNF-
expression,188 and attenuates expression of MMP-1 and MMP-3.191,192 However, PGE2
simultaneously down-regulates the endogenous MMP antagonist tissue inhibitor of
metalloproteinases-1 (TIMP-1),193 increases synovial plasminogen activator activity,549 and
increases IL-6 and IL-8 production.346
In the avascular, aneural substance of cartilage, where prostaglandins cannot be involved in overt
inflammation, it appears that they may yet play an important role in the regulation of chondrocyte
metabolism. Surprisingly, the effects of PGE2 in cartilage are little explored in the literature. While
evidence is not overwhelming, it is widely thought that PGE2 antagonises the cartilage effects of
IL-1ß in a negative feedback loop, as it does in synovium.259 There is some evidence that PGE2
reverses IL-1ß-induced depression of proteoglycan synthesis, increases type II collagen synthesis
(by a direct effect on COL2A1 gene expression), and decreases type I collagen
synthesis.197,259,272,273 Di Battista and coworkers have proposed a possible mechanism for this effect,
involving an autocrine feedback loop with IGF-1 and the IGF binding proteins IGFBP-3 and -
4.186,187,190 They hypothesised that the actions of locally synthesised eicosanoids in activating the
IGF-1 / IGFBP system may be important during cartilage repair processes.187 However, recent
interest has focussed on the interaction between PGE2 and MMPs, particularly evidence that it may
increase the release and activation of MMPs in certain tissues.3,24,530 PGE2 has also recently shown
to induce apoptosis in cultured chondrocytes, via a cAMP dependant pathway.546,588
In bone, prostaglandins are recognised as a potent inducer of bone resorption. PGE2 increases the
size and number of osteoclasts, and COX-2 seems to be necessary in vivo for osteoclastogenesis.
PGE2 may also repress osteoblast parathyroid hormone (PTH)-dependant responses.351 Clinically, it

54.
has been implicated in bone resorptive processes such as human RA96 and equine subchondral cyst
formation.873 However, other models equally implicate prostaglandins in bone formation.407,587 The
response of bone to prostaglandins is complex and likely differs in physiological and
pathophysiological states. The effect of NSAIDs on bone has been insufficiently examined, though
interestingly, long-term use of aspirin has been associated with greater bone mass in
postmenopausal women.63
1.1.3.4.2 Interaction Between Nitric Oxide and PGE2
Numerous studies have shown that NO appears to regulate cellular PGE2 production, though the
results obtained vary in different cell types, pathophysiological states, and experimental
conditions.22,789 In cartilage, the majority of studies suggest that NO augments PGE2
release.78,135,494,574,588,717 This is consistent with evidence that NO, via peroxynitrite, directly
activates the cyclooxygenase enzyme.448,716,717 However, other equally valid studies have shown
either no effect,383 or that NO instead shows an inverse relationship with PGE2 in cartilage.22,346,791
It has been suggested that NO may act at the level of COX expression.381 Amin and coworkers
have presented evidence that NO may differentially enhance COX-1-derived prostaglandins, while
inhibiting those produced by COX-2.24 However, Nedelec et al. (2001)574 found that NOS
inhibition specifically decreased induced but not basal COX activity. The subject remains a matter
of considerable controversy despite numerous investigations.
Conversely, the effect of PGE2 on NO production by joint tissues has received little attention. Onga
et al. (2000)599 recently examined the effect of PGE2 and forskolin, a direct adenylate cyclase
activator, on bovine articular chondrocytes. While they did not affect basal NO production, these
agents synergistically augmented IL-1ß-stimulated NO release and iNOS expression. This is
consistent with the observation that in many cell types, NSAIDs suppress iNOS
induction.5,112,360,591,718 In cartilage however, this effect seems to vary between different NSAIDs,

55.
suggesting that it is in fact unrelated to COX inhibition. For example, aspirin, diclofenac and
tenidap inhibit NO release,50,84 while naproxen, indomethacin and glucocorticoids have little
effect.3,628
The interaction of the NO and prostaglandin pathways in bone is also potentially important, given
the potent effects of each on bone resorption, but remains poorly understood. In an osteoblastic cell
line, NO appeared to mediate cytokine-induced induction of PGE2, which in turn had an inhibitory
effect on NO.402 However, SNAP had no effect on PGE2 concentration in a bone organ culture
system.664
1.1.4 OESTROGEN IN BONE AND JOINT HOMEOSTASIS
A growing body of epidemiological, clinical, and experimental evidence suggests that sex steroids,
particularly oestrogen, may modify the metabolic activity of joint tissues, and as such may be an
important player in the pathogenesis of OA. The effects of oestrogen in promoting bone density are
well known, and may represent one possible route by which it may influence the arthritic process.
Its role in the immune system is also well studied, and it is possible it may modulate the immune
processes of arthritis rather than cartilage events.162 However, strong evidence also suggests that
oestrogens have a direct influence on cartilage metabolism.
Being steroid hormones, oestrogens are highly lipid soluble and pass freely across cell membranes
to enter cells by passive diffusion. Binding to specific receptors occurs intracellularly; the exact
location of these receptors has been the subject of debate, but is probably the nuclear
compartment.143,419,891 The oestrogen-receptor complex then binds directly to the promoter region of
target genes.602 Since the cellular actions of oestrogen vary in response time, there appears to be a
cascade of gene responses. Recent evidence suggests that extremely rapid, non-genomic responses
also occur.433,602 Initial increases in nuclear proto-oncogenes c-myc, c-jun and c-fos suggest the AP-

56.
1 pathway may be one important route by which oestrogen affects cellular proliferation and
differentiation.165 This promoter pathway is also involved in post-receptor mechanisms of IL-1
activity. Oestrogen was found to be synergistic with lipopolysaccharides in enhancing IL-1ß
promoter activity, via oestrogen receptor-mediated processes, in a macrophage cell line.704
However, other reports have shown the opposite both in vitro and in vivo.704
Two functional oestrogen receptors are currently known, ER and ERß, which are the product of
separate genes.439,602 It has been shown that the two receptors signal in opposite ways via the AP-1
promoter element, suggesting they may have distinct and often opposite effects on cellular
activity.606 Therefore the ratio of ER to ERß is likely to be an important determinant of the cellular
effects of oestrogen, in addition to total receptor number.602,757 In discussing agonists/antagonists of
these receptors, the term ‘selective estrogen receptor modulator’ or SERM has come into use, to
reflect the highly tissue-specific action of such compounds 298
If oestrogens do play a role in joint tissue metabolism, local production or conversion from weaker
oestrogens might be important. Bellino (1992)68 examined rabbit cartilage and showed appreciable
17ß-hydroxysteroid dehydrogenase activity (which converts weak oestrone to potent oestradiol), but
not aromatase activity (required for de novo synthesis of oestradiol). This provides a mechanism for
local conversion of peripheral oestrone in postmenopausal women. Recent studies suggest
osteoblasts may possess both enzymes.711
Of particular relevance to discussion of the effects of oestrogen in joint tissues is its now well-
documented relationship with the IGF-1 system. Importantly, Fernihough et al. (1999)227 showed
that oestrogen supplementation of ovariectomised monkeys increased synovial fluid levels of IGF-1
and IGF-2, and IGFBP-1 and -3. This effect was dose-responsive but was reduced by treatment in
combination with progestogen.227 Further, elevated chondrocyte expression of IGFBP-2 was
demonstrated, in association with an increase in proteoglycan synthesis.685 However, these findings

57.
are somewhat contrary to previous observations that: (i) serum IGF-1 is known to increase five
weeks after ovariectomy in rats;811 (ii) oestrogen represses IGF-1 expression in HeLa cells and rat
vascular smooth muscle cells by an ER -dependant mechanism,728 while simultaneously
potentiating the IGF-1 receptor signalling pathway;398 and (iii) oral oestrogen replacement therapy
in women has been shown to reduce total serum IGF-1, IGFBP-3, and the acid-labile subunit of the
circulating ternary IGF-1 complex, though increases in IGFBP-1 and free IGF-1 have been
recorded.111,248,399,604,868 Interestingly though, oestradiol by either transdermal111,399,604 or intranasal248
routes has no apparent effect on IGF-1 levels in women. Serological associations between oestrogen
and IGF-1 have also been demonstrated in males.142,458 In the osteoblastoid ROS and SaOS-2 cell
lines, oestrogen-related proliferation has been associated with IGF-1,129,573 while mechanical strain-
related proliferation has been shown to act separately via IGF-2 receptors.129
1.1.4.1 Evidence Linking Oestrogens and Arthritis
1.1.4.1.1 Clinical and Epidemiological Evidence
The epidemiological patterns of OA strongly suggest a link with oestrogen.226 The incidence and
prevalence of OA increases steeply in women after the age of 50, corresponding to the time of
menopause.225,578 OA involving multiple joints is more frequent and more severe in women.556 A
subset of OA has long been recognised in middle-aged women, occurring around the time of
menopause, characterised by chronic recurrent “inflammatory OA” of multiple joints, especially the
interphalangeal joints (causing classical Heberden’s nodes), carpometacarpal joints, and cervical
spine.20,154,782 This ‘generalised OA’ has a strong genetic influence, possibly through oestrogen
receptor gene polymorphisms.850 It is unclear whether short-term hormonal fluctuations and
imbalances at the time of menopause, or the permanent decline in hormone levels which follows, is
responsible.578 In reviewing the subject, Spector and Campion (1989)782 postulated that an absolute
oestrogen excess, and/or a relative excess of oestrogen relative to progesterone during the

58.
perimenopausal period, may cause this phenomenon. Oestrogen excess may also explain other
epidemiological findings, such as the strong association between hysterectomy and OA (the
commonest indications for hysterectomy being those associated with hyperoestrogenism),781 the
well-documented inverse relationship of osteoporosis and OA,153,312,319,580,778,779 and the correlation
of obesity and OA in non-weight-bearing joints (obesity in women is associated with
hyperoestrogenism due to peripheral conversion of androstenedione to oestrone in adipose
tissue).782 However, results from studies directly measuring endogenous oestrogen levels have been
inconclusive.226,578,778 One study found a weak association between high oestrogen levels and
advanced knee (but not hand) OA, and a negative association with testosterone levels.778 Another
study found that women with generalised OA had lower levels of sex hormone binding globulin,
possibly allowing higher levels of free circulating oestrogen.784 Synovial oestradiol levels are
elevated in OA patients when compared to normal postmenopausal women, in levels correlated
highly with OA severity.838
On the other hand, there is fair (but not total) agreement amongst epidemiological studies (reviewed
by Felson and Nevitt (1999)226) that hormone (i.e. oestrogen) replacement therapy (HRT) in women
is associated with reduced incidence and prevalence of OA.577,598,719,722,778,783,934 A meta-analysis of
some of these studies found a pooled odds ratio of 0.76.578 In the Framingham study cohort of 551
elderly women, current use of oestrogen replacement therapy was found to have a moderate
protective effect against progression of radiographic knee OA.934 The same protective trend was
noted in the middle-aged Chingford study cohort.318,783 However, several studies have shown either
no effect or an increased relative OA risk with HRT.490,579,722,778 Adjustment for all confounding
factors is difficult because women choosing HRT differ in many ways from those who do not,
including frequency of utilisation of health services.225,578,598 In addition, women with osteoporosis
are more likely to be prescribed oestrogen, and osteoporosis is protective of OA.225 Such conflicting
lines of evidence are difficult to reconcile. It may be that the effect of oestrogen exposure depends
on menopausal status or stage of OA.225

59.
Other forms of arthritis are also apparently modified by hormonal influences. RA is more frequent
in females (gender ratio of approximately 2:1), and carries a worse prognosis in women.452,724 While
epidemiological studies remain inconclusive, there seems a consensus that oral contraceptives are
partially protective of severe RA.724,911 The anti-oestrogenic compound tamoxifen (commonly used
to treat breast cancer) has been circumstantially associated with symptomatic improvement in
rheumatoid patients, though it has also been suspected of causing inflammatory arthritis in its own
right. One small cohort study (of 49 patients) examining this matter found no effect.233
1.1.4.1.2 Experimental Evidence
To date, animal experiments investigating the effects of oestrogen on joint integrity are of
questionable relevance due to the high doses of oestrogen, mode of administration, species and age
of the animals used.835 Silberberg and Silberberg (1963)758 showed that oestrogen injections reduced
the severity of spontaneous OA lesions in male inbred mice. Later studies by Rosner and coworkers
demonstrated that subcutaneously administered oestrogen worsened osteoarthritic changes in both
ovariectomised and entire hemi-meniscectomised rabbits.699,701 Tamoxifen reduced erosive changes
in the same model.699 In a rabbit model of immune synovitis, tamoxifen also reduced gross and
histologic pathology.700
Tsai and coworkers conducted another series of experiments in normal rabbits, demonstrating OA-
like changes after intra-articular injection of oestradiol benzoate, an effect which could be blocked
by tamoxifen.834,837 The effect was apparently cytotoxic, with cell necrosis, chondrocyte cloning,
and pitting of cartilage.837 However, the doses of oestradiol used in these studies (equivalent of 0.3
mg/kg/day) are clearly supraphysiological.578 Chander and Desa (1991)123 also demonstrated that
large doses of oestrogen accelerated the loss of glycosaminoglycan from rat femoral heads
implanted subcutaneously into female mice, an effect which again could be blocked by tamoxifen.

60.
1.1.4.2 Oestrogen and Cartilage
It has been shown in many species that exogenous oestradiol depresses proteoglycan synthesis by
chondrocytes,582,699,701,782,835 though in bovine cartilage, this effect has only been demonstrated at
supraphysiological concentrations.487 However, it is not clear whether this suppression of synthesis
leads to net loss of proteoglycan.487,782 In vitro, 17ß-oestradiol inhibits chondrocyte proliferation,
but increases production of collagen and other proteins.487,739 This change may effectively signal
chondrocyte differentiation, as has been shown in growth plate cartilage.572 It is not clear whether
these effects are direct (receptor-mediated) or are dependent on some second messenger. Oestradiol
enhances IL-6 production by IL-1-stimulated chondrocytes.295 17ß-oestradiol also reduces basal
COX-2 expression in bovine chondrocytes, but has no effect on cytokine-induced levels.720
If oestradiol is an important regulator of chondrocyte metabolism, this implies inherent gender
differences in cartilage. Male children have greater weight-adjusted cartilage volume than females,
even from a young age.390 In comparing femoral head cartilage from male and female rats, Larbre et
al. (1994)452 found that female cartilage contained less proteoglycan and collagen, showed greater
spontaneous proteoglycan loss and lower proteoglycan synthesis, and was more sensitive to IL-1-
induced inhibition of matrix synthesis. However, as female cartilage was less sensitive to IL-1-
induced loss of proteoglycan, the net effect on glycosaminoglycan content was similar in both
sexes. Some gender differences in metabolic responses have been shown in human cartilage, such
as in the effects of 24,25-dihydroxy-vitamin D3.739 Hormonal influences must also account for the
reduction in type II collagen, biglycan, TNF- , iNOS, and COX-2 expression seen in articular
cartilage of pregnant rabbits relative to non-pregnant controls.335
Turner et al. (1997)841 made the important observation that simple ovariectomy of normal, aged
sheep has an adverse effect on the biomechanical integrity of cartilage, as evidenced by a reduction
in aggregate modulus (compressive stiffness) and shear modulus. Räsänen and Messner (1999)667

61.
showed an increase in static stiffness and cartilage thickness in young rabbits after ovariectomy.
Similarly, ovariectomy of young rats rapidly increases cartilage thickness in the temporomandibular
joint.597,918,923 These findings should be interpreted with some caution, as results based on pubertal
growing animals may not be relevant to adult animals. Ovariectomy of adult rats increases cartilage
collagen and protein content4 and creatine kinase activity.777
The presence of high-affinity oestrogen receptors has been demonstrated in rabbit, sheep, rat, mice,
dog, baboon, and human articular cartilage.69,642,685,702,836,842,851,928 However, Mackintosh and Mason
(1988)487 were unable to demonstrate oestrogen receptors in bovine chondrocytes. Tsai et al.
(1992)838 examined oestrogen binding sites in human OA cartilage collected at knee joint
replacement, and showed that levels were greater in femoral and tibial cartilage of the medial
compartment than in equivalent tissues from the lateral joint compartment. In this small group of
patients, oestrogen receptors were found to be significantly correlated with OA severity, but were
absent from patients with traumatic OA secondary to articular fractures.838 Both ER and ERß have
been detected in human and monkey cartilage,685,851 though a recent report suggested that ERß may
be the predominant receptor present in human cartilage.395
One problem raised by the apparent importance of oestrogen in cartilage homeostasis is explaining
how this may apply to male animals. In fact, oestrogen-dependant mechanisms may also exist in
male tissues. Recent studies have shown that bone homeostasis in men appears to be oestrogen
dependant.669 Metabolites of dihydrotestosterone such as 5 -androstane-3ß,17ß-diol have a good
affinity for oestrogen receptors and may replace oestradiol in this function.298
1.1.4.3 Oestrogen and Synovium
Several authors suggest that synovial tissue may also be a significant target of oestrogen, not least
its influence on disease activity in rheumatoid arthritis.164,849 Given the known modulating effects of

62.
androgens and oestrogens on cytokine synthesis by monocyte/macrophage populations, synovial
macrophages may be a major target of sex hormones within the joint.161,164 Ushiyama et al.
(1995)849 demonstrated oestrogen binding and expression of oestrogen receptor-related protein
(p29) in human synovium. Cutolo et al. (1996)163 used kinetic binding studies to confirm the
presence of specific oestrogen receptors in cells characterised as synovial macrophages. Both Type
I (high affinity, low capacity) and Type II (lower affinity, high capacity) receptors were observed.
Ghosh and Seshandri (1986)266 reported ‘large amounts’ of oestrogen and progesterone receptors in
a rheumatoid synovial cell line.
The effect of oestrogen stimulation in synovial tissue remains poorly examined. The addition of
oestrogen caused a mild increase in IL-1ß-stimulated IL-6 production by human rheumatoid
synovial fibroblasts.411 Oestrogen has been shown to increase IL-1 production in peripheral
macrophages, though this effect may be biphasic at higher concentrations.161,361 This seems feasible
given the close association of oestrogen and IL-1 via the AP-1 promoter.
1.1.4.4 Oestrogen and Bone
The exact mechanisms by which oestrogens exert their well-known effects on bone metabolism are
unclear. Multiple pathways and mediators appear to be involved, as reviewed by Oursler (1998).602
Firstly, oestrogen modifies the proliferation and cellular responses of osteoblasts. Oestrogen
receptors are known to be present in osteoblast-like cells.46,208,711 In osteoblastic precursor cells,
oestrogen has been reported to both stimulate and inhibit proliferation in vitro, suggesting the effect
may be dependant on maturation state.602 Similarly, osteoblast matrix production is promoted by
oestrogen, but only under certain experimental conditions. Oestrogen also modulates the release of
growth factors such as IGF-1 and TGF-ß, and osteoblast responsiveness to other hormones such as
vitamin D3, progesterone, growth hormone, and parathyroid hormone.207 It has been shown in vitro
that oestrogen promotes the proliferative response by osteoblasts to mechanical strain.169 This may

63.
explain why the homeostatic strain-related adaptive responses of bone fail to compensate for
postmenopausal bone loss.
Secondly, oestrogen influences osteoclast differentiation and activation, probably via complex
cytokine networks including IL-1, TNF- and IL-6.356,602 Evidence for such interactions is multiple
but somewhat confused. IL-6 is known to be important in osteoclastogenesis,385 and may represent
an important pro-resorptive factor released by osteoblasts and other cells. Oestrogen has been
shown to inhibit IL-6 production in osteoblast-like cell lines.267,404,429 In one murine study, a
blocking antibody to IL-6 was able to prevent any post-ovariectomy increase in
osteoclastogenesis.385 Similarly, ovariectomy has no effect on bone mass in IL-6 knockout mice.645
However, other studies have found that blocking IL-6 antibodies fail to prevent bone loss in
ovariectomised rats and mice.418 The response may be site-specific, or other cytokines, such as IL-1
and TNF- may be involved.602 Both are potent inducers of bone resorption and their production by
certain cell types appears to be suppressed by oestrogen. Peripheral blood monocytes (PBM) from
patients with osteoporosis produce greater amounts of IL-1 and TNF- than those from age-
matched patients.691 This effect appears to be oestrogen-dependant, as the same effect has been
shown in ovariectomised women,605 and PBM cytokine production is normalised (and IL-1Ra
release increased) by oestrogen replacement therapy.691 This is somewhat surprising, as in other
experimental systems using monocyte cultures, oestrogen has been found to potentiate the basal
production of IL-1ß.161 Both IL-1 and IL-6 increase RANKL expression by osteoblasts,287 and
blocking antibodies to IL-1 and TNF- are able to prevent bone loss in ovariectomised mice.418
However, the relative importance of such cytokines in oestrogen-deficient bone loss is debatable.
Such second messengers may not be necessary, as oestrogen has been shown to suppress
differentiation of osteoclasts from blood precursors through a direct receptor-mediated
mechanism.748 Direct associations between oestrogens and the RANKL/osteoprotegerin (OPG) axis
have recently been reported, and may prove to be important. For example, oestrogen may decrease
responsiveness of osteoblastic precursors to RANKL.748,788

64.
Thirdly, oestrogen may modulate the activity of mature osteoclasts directly.602 It has been shown
that osteoclasts express oestrogen receptors,600 and that oestradiol produces a dose-dependant
reduction in resorptive activity when added to cultures of osteoclasts. In addition to down-
regulation of resorptive enzymes, rapid non-genomic effects may be involved, via specific trans-
membrane oestrogen receptors.602 Oestrogen also appears to induce osteoclast apoptosis.400
Another possible mechanism by which oestrogen regulates bone metabolism is by influencing nitric
oxide production. The known protective effects of oestrogen in the cardiovascular system are
suspected to be mediated by NO. For example, the aorta of female rabbits has higher basal NO
production than that of male animals.330 Oestrogen has been shown to increase production of NO in
a range of guinea pig tissues, including uterine artery, heart, skeletal muscle, and brain.884,890
Oestrogen is also known to cause vasodilation, an effect can be partially blocked by the NOS
inhibitor L-NMA.666 A link is also suggested by the observation that circulating nitrate levels in
women increase with follicular development,676 and after administration of exogenous
oestradiol,370,666 though the exact source of this nitrate is unknown.
More recently is has been proposed that NO may mediate the bone-sparing effects of oestrogen.
Wimalawansa et al. (1996)906 showed that while both glyceryl trinitrate and exogenous oestrogen
alleviated ovariectomy-induced bone loss in rats, oestrogen was totally ineffective in the presence
L-NAME, suggesting that much of the effects of oestrogen on bone resorption may be mediated by
NO. Other researchers have shown in vivo that oestradiol increases osteoblast NOS activity and
expression approximately two- to three-fold, probably via induction of the eNOS isoform.36 As NO
has been shown to reduce osteoclast formation and activity, this link seems entirely
plausible.483,486,664
Whilst oestrogen appears to increase constitutive NO synthesis, there is some evidence that it may
reduce NO production via iNOS. Ovariectomised rats demonstrate a greater increase in plasma

65.
nitrate levels following endotoxin treatment than entire animals, and this increase can be attenuated
by pre-treatment with oestradiol.406 Oestrogens may therefore be beneficial in pathological
conditions associated with iNOS induction.
1.1.5 ANIMAL MODELS OF OSTEOARTHRITIS AND OSTEOPOROSIS
1.1.5.1 Animal Models of OA
While the degeneration of articular cartilage is an early event in OA and is of central importance to
its pathogenesis, this process is generally asymptomatic. Radiographic evidence of the disease may
not be evident until cartilage disruption is well advanced.258 Similarly, symptoms of pain and
stiffness may be present only in advanced disease, and correlate poorly to radiographic or
pathologic indices. As joint tissues are only feasibly obtained from clinical end-stage disease, study
of early events is difficult and properly matched control tissues are difficult to obtain. The use of
animal models therefore provides the only practical method for sequential evaluation of the factors
initiating (and perpetuating) the complex morphological, biochemical, metabolic, and molecular
alterations seen in OA.
Brandt (1999)89 recognises three broad categories of animal OA models: (i) spontaneous or genetic
OA (e.g. Dunkin Hartley guinea pigs, STR/ort mice, canine hip dysplasia); (ii) chemically-induced
OA, generally by intra-articular injection of an irritant substance (iodoacetate, papain, hypertonic
saline); and (iii) physically-induced OA, including anterior cruciate ligament transection (ACLT),
immobilisation, myectomy, and meniscectomy procedures. Each method has its own advantages
and pitfalls. Because the aetiology of OA is unknown, particularly with respect to the importance of
overt inflammatory processes, models involving inflammatory responses are of questionable use.
Spontaneous models are caused by specific genetic defects and hence relate to specific subsets of
OA. Similarly, surgical models may be more relevant to post-traumatic OA than primary

66.
(idiopathic) OA, and it is unknown whether ideal treatment modalities may differ between these
subgroups.89 One criticism which has been applied to all animal models is the short time scale
required to establish advanced disease, compared to the decades-long progression of human OA.260
Models based on surgical disruption of the stifle joint have most frequently employed rabbits or
dogs. Rabbit models have involved partial or total meniscectomy, ACLT, or combined procedures
such as the Hulth procedure.152,364,430,927 The utility of rabbit models may be limited due the different
range of motion and stresses on the knee joint compared to that of bipeds, and the particularly labile
nature of lapine cartilage.260 Even injury as slight as intra-articular injection of sterile physiological
saline is able to induce proteoglycan defects in rabbit articular cartilage.246
The canine ACLT or Pond-Nuki model is widely utilised and well described.646 However, it has
been demonstrated that the blind 'stab' technique usually employed creates intra-articular
haemorrhage and subsequent inflammation which significantly contributes to the pathologic
process.569 Hypertrophic repair of cartilage is predominant in this model, and long experimental
periods are required for progression to full-thickness cartilage lesions, though this can be
accelerated by neurectomy.89,90,412 It has been proposed that many of the joint changes characteristic
of this model may be attributable to the decrease in loading following unilateral surgery, rather than
any intrinsic OA-like disease process.590
1.1.5.2 The Ovine Meniscectomy Model of OA
The knee joint menisci are fibrocartilaginous semi-lunar pads interposed between the femoral and
tibial condyles. They are composed predominantly of a dense type I collagen framework and
hydrated aggrecan, with the majority of collagen fibres oriented circumferentially. The menisci
function to improve congruity between the convex femoral condyles and the comparably flat tibial
articular surface, hence increasing the effective contact area of the articulation and reducing focal

67.
peak forces.67 It has been estimated that 50-70% of the total transarticular load may be transmitted
via the menisci.541 The menisci also have roles in shock absorption, stabilisation, joint lubrication,
and possibly in providing sensory feedback during joint compression.
It has long been recognised that total meniscectomy results in eventual cartilage degeneration and
joint remodelling in human patients.541 Experimental meniscectomy of sheep reliably creates a
useful model of OA, which primarily develops secondary to altered joint congruity and load
distribution in the meniscus-deficient femoro-tibial compartment.258,260,264,474 Medial meniscectomy
in sheep has been shown to reduce the mean area of femoro-tibial contact from 78.6 mm2 to 24.8
mm2.67 Biomechanical alterations also cause wide-ranging changes to more distant sites such as the
patella and even the femoral head.33 Synovitis, synovial fluid alterations, and subchondral bone
remodelling accompany cartilage changes and may play a role in disease progression.38,474 The
advantages of the sheep model are multiple: husbandry practices in Australia allow purchase of
genotypically and phenotypically homogenous groups of animals from a single source, and the use
of food animals is ethically and financially preferable to companion animal or primate models. The
large joint permits serial collection of synovial fluid, and allows both topographic division and the
harvest of large volumes of cartilage for multiple analyses.258 The anatomy of the ovine stifle is
comparatively similar to the human knee, and is relatively extended in normal stance, compared to
the more flexed posture of rodents and rabbits.520 The lesions induced are reproducible, slowly
progressive, and correspond faithfully to the pathological changes found in naturally occurring
human OA. Focal cartilage fibrillation or erosion develops within a few months, accompanied by
matrix proliferation and marginal osteophyte formation.474 Increased loss of proteoglycans from
cartilage, reduced incorporation of 35S-labelled proteoglycans, increased chondroitin-4-sulphate,
and changes in proteoglycan epitopes have all been documented.258,474
The differences between ovine medial and lateral meniscectomy models have been examined in
detail.264 The ovine lateral meniscus covers a greater proportion of the tibial condyle than the

68.
medial, as has been demonstrated in the human knee (Figure 1.6).41,143,541 The medial compartment
is reported to bear a greater proportion of total load than the lateral compartment in the human knee;
the greater bone density of the medial compartment suggests this is probably also true in sheep.39,41
Contact stresses are therefore lower in the intact lateral compartment than the medial under
conditions of normal weight-bearing. The higher stresses therefore imposed on articular cartilage
following lateral meniscectomy may explain the more rapid cartilage deterioration when compared
to that following medial surgery.264
Figure 1.6: Dorsal view of the ovine tibial condyles with menisci intact. Note the difference in the
caudal attachments of the medial (menisco-tibial ligament) and lateral (menisco-femoral ligament)
menisci, and the close association between the lateral meniscus and the popliteus tendon. MED,
medial; LAT, lateral.
CRANIAL
LAT MED
CAUDAL

69.
1.1.5.3 The Ovine Ovariectomy Model of Postmenopausal Osteoporosis
Osteoporosis is a systemic skeletal disease defined by loss of bone volume and bone mineral
density, and micro-architectural deterioration of bone, leading to an increase in bone fragility and
incident fracture. Women are prone to a postmenopausal form of osteoporosis, characterised by
rapid bone loss in the 5-10 years following menopause, as distinct from the slower age-dependant
loss of bone density occurring in both sexes.615 BMD is the variable most often measured, and is
highly correlated with bone strength.615 However, recognition of the importance of other factors
such as trabecular connectivity has lead to the view that densitometry may be inappropriate as a
single predictor of fracture risk in osteoporosis.616 The most important osteoporotic changes occur
in trabecular bone, though cortical bone loss is also important in certain types of osteoporosis and is
often overlooked.686
Newman et al. (1995)581 reviewed the application of the ovariectomised sheep model to the study of
osteoporosis. Sheep represent a promising model as they have similar oestrus and hormonal cycles
to women, with the exception of a short period of anoestrus during the winter months. Turner and
coworkers have characterised this model in extensive studies.127,290,581,840-845 Most bone
histomorphometric parameters change slowly, with only a slight decrease in bone mass at six
months post-ovariectomy, accompanied by a slight increase in trabecular separation and decrease in
wall thickness.845
Plasma osteocalcin is potentially a useful measure of osteoblast function in sheep,217,581 though it is
reduced by low dietary phosphate,156 pregnancy,217 and long term administration of
adrenocorticotropic hormone (ACTH) or corticosteroids.157,236 Plasma values in both growing and
mature animals are at least 6-10 times that of equivalent human subjects, probably reflecting the
genetically selected higher growth rate in these production animals.157 In contrast to humans, there
is no significant circadian variation157 or rapid pulsatile variation of circulating osteocalcin in

70.
sheep.156 However, plasma alkaline phosphatase has limited value as a marker of ovine bone
formation.156
1.1.6 IN VITRO INVESTIGATION OF SYNOVIAL TISSUE
1.1.6.1 Culture Methods of Synovial Tissue in vitro
In the past, researchers used a variety of in vitro culture methods (reviewed by Henderson (1981)337)
to examine the metabolic changes evident in synovial tissue in different disease states. Fraser
(1965)241 first described an enzymatic method of harvesting synovial cells, involving intra-articular
injection and aspiration of trypsin solution. However, he found that the cells so isolated do not grow
well in culture, demonstrating a heavy serum requirement and large losses at subculture. It is now
recognised this method yields predominantly Type A synoviocytes,59,242 as demonstrated by their
failure to produce hyaluronan.166 Type A cells do not persist in prolonged culture or serial
passage.1,176,242 Using this method, it has been shown that the number of Type A cells declines with
time in culture (falling after about 4 days), and drops sharply after subculture procedures (<4% after
1st passage, <1% after 2nd passage).242
Other researchers have used synovial organ culture methods involving biopsy specimens,715
weighed dissected synovial portions,133,394,627, intact synovial villi,565,570 or minced synovium.223,714
Such techniques have the advantage that they contain a representative mixture of type A and B
synoviocytes, together with cells normally present in the subsynovial layer. However, one problem
with organ culture methods is that weight-for-weight comparisons are invalid due to the variably
increased cellularity of the synovial lining layer in inflammatory states.337 If synovial organ cultures
are cultured for long periods in such a manner that they are not disturbed, marginal cells adhere to
the floor of the culture vessel and proliferate as a cell monolayer. This monolayer initially contains

71.
both cell types, but synovial fibroblasts quickly overgrow Type A cells.438 This explant outgrowth
method has been widely used to isolate synoviocytes.27,302
The majority of recent synovial studies have employed a method whereby minced synovial tissue is
subjected to sequential enzymatic digestion to yield a cellular suspension. Most utilise a sequential
trypsin/collagenase digestion similar to that described by Smith and Ghosh (1987).173,193,250,263,539,770
While most studies use subcultured cells, McInnes et al. (1996)532 used this method to derive a
primary single cell suspension that was assayed directly. Several authors suggest that Type A cells
adhere more rapidly than fibroblasts, and it is possible to select for them on this basis.730 Without
selection procedures, Type A cells are rapidly eliminated from synoviocyte monolayers.128,377 Even
synovial fibroblasts will eventually become senescent after 10-12 passages in monolayer culture.231
Georgescu et al. (1988)250 studied the effect of continued passage on lapine synovial fibroblasts, and
showed that the growth rate slowed after the 4th passage, and was minimal after 8-12 population
doublings.
Recent advances in synoviocyte culture technique have included the use of three-dimensional
culture systems, such as collagen gels.810 Other studies have utilised co-cultures of synovial
fibroblasts and macrophages, in attempts to simulate the in vivo interactions of Type A and B
synoviocytes.334,382,617,705,740 Such co-culture techniques certainly cause more rapid and more
extensive release of catabolic factors in vitro.617
1.1.6.2 Culture Requirements of Synoviocytes
Synoviocytes are undemanding in their culture requirements. Addition of serum is not essential, but
has a stimulatory effect on the biosynthesis of collagen, total protein, and glycosaminoglycans.503,654
Similarly, refreshing serum-containing culture medium has been shown to stimulate HA
synthesis.706 However, cells derived from inflamed synovium may be less sensitive to serum

72.
depletion.27 Moderate variations in media pH have no effect on cell growth, but may affect HA
production.145 Early investigations of rheumatoid synovial tissue showed a preference for relatively
alkaline (pH 7.8) culture conditions.27,648 Trypsinization of cellular monolayers transiently induces
collagenase and plasminogen activator secretion, but this effect is reduced by presence of serum.892
Authors examining the effect of trypsin have shown that little cell damage occurs, and that recovery
is rapid on the addition of serum-containing media.59,503
1.1.6.3 Differences Between Normal and Arthritic Synovium in Culture
Smith and Hamerman (1969)767 were among the first authors to note persistent differences between
rheumatoid and normal synoviocytes in culture, showing altered morphology, growth, and cartilage
catabolic activity.766 Other studies have noted greater glucose consumption, lactate formation, and
HA synthesis,119 lower serum requirement in culture,503, and altered responses to drugs347 in cells
derived from arthritic joints. Some authors have shown that such metabolic differences in inflamed
synovia persist many generations in culture.276,377,908 However, others have suggested that while the
production of mediators such as PGE2 is increased in cultures derived from inflamed synovia
relative to healthy tissues, this difference has disappeared after repeated media changes (i.e. with
the loss of the non-adherent cell population).530,531,689,705 This process parallels a change in cellular
morphology - cells in primary culture are large (sometimes multinucleate), stellate, and granular in
appearance, gaining a more fibroblastic appearance as culture progresses.435,767 Castor et al.
(1971)119 first suggested that soluble “activating factors" were capable of inducing these changes in
normal synovial fibroblasts, and that increased autocrine production of such factors may account for
most of the in vitro changes observed. The typical stellate (or ‘dendritic’) morphology of activated
cells is probably related to a prostaglandin–induced increase in intracellular cAMP.55,334,435 PGE2
production is massive in primary synovial fibroblast cultures, especially those of rheumatoid origin
(e.g. 1 g/106 cells/day),172 and declines with time in culture.176,276,689

73.
It currently seems unlikely that rheumatoid synoviocytes are inherently different from normal
cells.530 There are no ultrastructural features characteristic of rheumatoid cells.431,917 One study
showed no difference in glucose uptake or glycosaminoglycan synthesis.26 Others have concluded
quantitative rather than qualitative differences.764 For example, production of IL-1-like activity is
increased in RA synoviocytes, but is similar to cells derived from other arthritic conditions per unit
of DNA.97 However, rheumatoid synoviocytes do exhibit growth characteristics distinct from those
of OA-derived cells; for example, only the former are capable of monoclonal expansion on soft
agarose gels.266,369
A cell type intermediate of Type A and B cells has been described in early synovial
cultures.531,613,753 These intermediate / ’dendritic’ / ‘transformed-appearing’ cells are nominally
associated with the cartilage and bone destruction which occurs in RA, and are major producers of
degradative products such as collagenase in culture.915 These cells, commonest in rheumatoid
cultures, are characterised by dense rough endoplasmic reticulum, large irregular pale nuclei with
prominent nucleoli, and abundant cytoplasm. There is some confusion in the literature over the
origins of this cell type, and whether it represents a distinct cell type or an overstimulated Type A or
B cell. Goto et al. (1987)277 established clonal colonies of all three cell types and found that all
retained distinct morphologies in long-term culture. However, Hendler et al. (1985)342 observed
direct transition of dendritic cells to fibroblasts and vice versa. It is interesting to note that in the
former study, ‘dendritic cell’ clones maintained higher spontaneous IL-1 production – suggesting
that autocrine cytokine stimulation may be responsible for their characteristic morphology.277

74.
The following metabolic changes are characteristic of ‘activated’ or chronically cytokine-stimulated
synoviocytes:
i) strong expression of oncogene products such as c-fos and c-jun.420,693
ii) reduced growth rate and shortened life span.654,767 However, Myers and Brandt (1987)567 noted
that synovial cultures from arthritic dogs (8-12wks post ACLT) proliferated more rapidly than
control specimens. It has been shown that IL-1 is mitogenic to synovial fibroblasts, but only
when induction of PGE2 is blocked (e.g. by indomethacin).172,304
iii) increased synthesis of glycosaminoglycan, particularly hyaluronic acid.504 MCM,117,168,654
purified IL-1,375,410,921,922 TNF- ,922 PGE2,763 and bacterial lipopolysaccharide99 all increase
synovial glycosaminoglycan synthesis, possibly via an increase in the key enzyme hyaluronic
acid synthetase.654 However, HA produced by inflamed synoviocytes is of abnormally low
molecular weight and reduced viscosity.116,654 Expression of CD44 (the principal receptor for
HA) is reduced in rheumatoid synovia.341
iv) decreased synthesis of type I, III, and VI collagen.62,168,683 This response marks another
distinction between synoviocytes and tissue fibroblasts, in which these cytokines would
normally stimulate fibrosis.
v) increased synthesis of other connective tissue proteins such as fibronectin 453
vi) increased synthesis of prostaglandins, predominantly PGE2.172,276
vii) increased synthesis of matrix metalloproteases.138 Synovial fibroblast production of MMPs
(92kD gelatinase, collagenase, stromelysin) wanes with time and passage in culture,415,825,847
but remains very sensitive to re-induction by IL-1 and TNF- .173,174,176,480,595,847 In dermal
fibroblasts, collagenase production has been shown to be reliant on mechanical stress within
the supporting collagen framework, partially explaining the low resting expression in cells
cultured on rigid substrates, and its induction by trypsinization.447
viii) increased spontaneous autocrine IL-1 production.276 IL-1 receptors are also up-regulated in
OA synoviocytes but this difference does not persist in culture.709

75.
ix) dramatically increased IL-6 production.296 Studies of clonal rheumatoid synovial fibroblast
populations have shown that some clones are high IL-6 producers while others low producers,
with production independent of the presence of other cell types or autocrine IL-1ß / TNF-
stimulation.64 However, another study showed no difference in IL-6 production between
clones of fibroblastic, dendritic, or macrophage-like synoviocytes.547
x) increased nitric oxide production.532 However Stefanovic-Racic et al. (1994)795 showed
obvious differences between individual rabbit synovial fibroblast cultures with respect to NO
production , with up to 10% showing no detectable production.
1.1.7 PHARMACEUTICAL MANAGEMENT OF OSTEOARTHRITIS
1.1.7.1 Non-steroidal anti-inflammatory drugs (NSAIDs)
Non-steroidal anti-inflammatory drugs remain the mainstay for the pharmacological management of
osteoarthritis symptoms. This is despite evidence that many common NSAIDs do not reduce joint
degeneration in OA, and may even hasten it.259 NSAIDs have traditionally been considered
analgesic and anti-inflammatory drugs, without involvement in the predominantly cytokine-driven
pathways of cartilage degradation in OA. Given recent revelations that COX-2 is superinduced in
cartilage,22 and that prostaglandins do indeed interact directly with cytokine actions and matrix
metabolism,24 NSAID usage is being re-examined in a new critical light. It is increasingly being
recognised that despite their functional grouping, there is considerable variation in the mode of
action of different NSAIDs. While long thought to act peripherally in inflamed tissues, there is also
evidence that some of their analgesic action may be mediated centrally.181
The recent discovery of the inducible COX-2 isoform of cyclooxygenase has been of particular
importance to NSAID pharmacology. The two COX isoforms share only 60% homology, though
much of the difference in at the carboxy and amino termini, while the central portion is highly

76.
conserved.861 Thus the proteins are structurally very similar, though key amino acid differences
allow selective pharmaceutical targeting of the induced isoform. This discovery lead to
development and promotion of a new generation of NSAIDs which are reported to more selectively
inhibit COX-2, and subsequently possess reduced gastrointestinal toxicity.861 The COX selectivity
of various NSAIDs has been investigated in dogs; meloxicam was found to have a COX-2:COX-1
ratio of approximately 12:1, while for carpofen the ratio was approximately 1.75:1.413 However,
recent evidence also suggests an important role for COX-2 in the kidney, where it may be involved
in renin release in salt-depleted and volume-depleted states.575 Hence evidence is emerging that
COX-2 selective inhibitors are not entirely free of renal toxicity in human or veterinary clinical
practice.91,489,690
As their main mode of action is thought to be the inhibition of cyclooxygenase, the action of
NSAIDs in the OA joint is dependent on the net role of prostaglandins, especially PGE2, in joint
tissues. Past evidence suggests that PGE2 is largely supportive of cartilage matrix synthesis, and
opposes some of the actions of IL-1ß in synovial tissue. This has lead to the proposal that NSAIDs
may accelerate the progression of osteoarthritis. Certainly this is the case with many of the previous
generation of NSAIDs. Indomethacin, aspirin, naproxen, and phenylbutazone have all been shown
to suppress proteoglycan biosynthesis in cartilage, in a variety of species.198,259,343,661
Phenylbutazone is especially interesting in this regard as it is not a COX-inhibitor but specifically
inhibits formation of PGE2. Similarly, indomethacin and ketoprofen have been shown to suppress
chondrocyte collagen synthesis.274 Moreover, aspirin and indomethacin have been shown to
accelerate cartilage degeneration in animal models.610,641 However, newer NSAIDs have been
shown not to have deleterious effects on matrix synthesis. Rather, aceclofenac and tenidap actually
increase glycosaminoglycan synthesis in human cartilage.81,198 Carprofen also increases synthesis
and reduces release of proteoglycan from equine and canine cartilage,37,734 though inhibition of
synthesis was seen at higher doses in the canine study. Numerous studies have shown meloxicam to
have no detrimental effect on proteoglycan synthesis in human, porcine, or canine cartilage,81,662,663

77.
and no effect on DNA or type II collagen synthesis by human chondrocytes.61 Similarly, piroxicam
has no demonstrated effect on cartilage synthetic activity.198,661 The effect of etodolac on
glycosaminoglycan synthesis is less clear; while it has been shown to decrease synthesis in rabbit
chondrocytes,674 it had no effect on human chondrocytes in a three-dimensional culture system.343
These newer NSAIDs have also been attributed chondroprotective properties unrelated to their
effects on proteoglycan synthesis. For example, etodolac prevents IL-1ß-induced suppression of
chondrocyte collagen synthesis.274,519 The chondroprotective label has also been applied to
piroxicam228,347,348 and tiaprofenic acid.259,262,626
The specific influence of NSAIDs on MMP activity requires further clarification. Sadowski and
Steinmeyer (2001)710 recently showed that a range of NSAIDs inhibited IL-1ß-induced collagenase
activity in bovine articular chondrocyte cultures, while indomethacin and meloxicam additionally
inhibited proteoglycan degradation. However, clear differences in their modes of action suggested
that these effects were independent of cyclooxygenase inhibition. Arner et al. (1998)43 found a
range of NSAIDs to have no effect on aggrecanase activity in cartilage explant cultures.
While the cartilage effects of COX inhibitors are well studied, their effects on synovial metabolism
are surprisingly poorly known. Meloxicam was found to have no effect on IL-1ß release, but
increased IL-6 production by human synovial explants.663 In other studies, tenidap was shown to
reduce synovial expression of IL-1 receptors and collagenase,513 while nimesulide and naproxen
reduce urokinase and IL-6 synthesis, and increase PAI-1 production in human OA synovial
fibroblasts.636 Meloxicam was found to attenuate the acute inflammatory synovial fluid changes
provoked by intra-articular injection of contrast agent in dogs.853 However in a separate but similar
study, meloxicam prevented the pain but not the synovial fluid responses induced by intra-articular
injection of calcium pyrophosphate crystals.662 Etodolac reduces the arthritic changes associated
with adjuvant arthritis514 and type-II collagen-induced arthritis373 in mice. These observations are

78.
interesting in the light of revelations that homozygous COX-2-deficient mice display essentially
normal inflammatory responses to antigen-induced arthritis.371
As the in vivo effects of the modern generation of NSAIDs do not fit those predicted from the
known, mainly protective effects of PGE2 in joint tissues, COX-2 selectivity alone cannot explain
the improved safety profile of these agents in cartilage. Evidence of a chondroneutral or even
chondroprotective status for these drugs increasingly refutes the existence of a functional inhibitory
feedback loop between PGE2 and IL-1ß. The observation that the stimulatory effect of aceclofenac
on cartilage proteoglycan synthesis is dependant on the presence of serum, and that serum can be
replaced in this role by IGF-1 alone,198 seems to specifically contradict the positive interactions of
PGE2 and the IGF-1 axis proposed by Pelletier and co-workers.186,187,190 It seems increasingly likely
that the interaction between PGE2 and NO is important in the actions of modern NSAIDs, and
PGE2-mediated augmentation of chondrocyte NO production as observed by Onga et al. (2000)599
provides a possible link between prostaglandin production and cartilage degradation. Such
discussion is seriously hampered by our incomplete understanding of the mode(s) of action of
modern NSAIDs such as carprofen and meloxicam. L-NAME has been shown to block the anti-
nociceptive effects of meloxicam, nimesulide, ketorolac, and dipyrone in pain models in rats,
suggesting involvement of a NO-cyclic GMP-mediated pathway in peripheral pain
transmission.6,283,374 Similarly, NO-releasing NSAIDs are being developed by the addition of
nitroxybutyl moieties to existing NSAIDs such as aspirin, with the aim of reduced gastrointestinal
toxicity.818 These have also been found to possess increased anti-inflammatory activities.137,230,883
Given the strong evidence of a role for PGE2 in bone metabolism, COX inhibitors may also actively
modify bony tissues. To date, studies suggest this effect is certainly possible but inconsistent.
Meloxicam prevents alveolar bone loss in experimental periodontitis,75 yet phenylbutazone
decreases the rate of mineral apposition and healing of cortical defects in horses.692 Pelletier et al.
(1999)633 found that carprofen, while preventing cartilage erosion following canine ACLT, also

79.
reduced subchondral bone loss and normalised subchondral osteoblast metabolism. On this basis,
the authors proposed that the protective actions of the drug might partly relate to its stabilising
effect on subchondral bone. However, reduced bone loss might alternatively be explained by
increased weight-bearing in this unilateral model, and bone morphology was not examined at other
sites in this study.
1.1.7.2 Corticosteroids
Corticosteroids inhibit phospholipase A2, thereby reducing production of both cyclooxygenase and
lipoxygenase end-products. As well as their potent anti-inflammatory action, corticosteroids are
capable of decreasing cartilage degradation, and importantly are potent inhibitors of MMP
activity.710 However, they also suppress chondrocyte proteoglycan and collagen synthesis, by a
mechanism that is poorly understood.255 In synovial fibroblasts, hydrocortisone similarly reduces
anabolic activity, particularly hyaluronic acid synthesis,365,566,706-708,922 but also suppresses release of
catabolic mediators such as plasminogen activator.303 In vivo, both beneficial and deleterious effects
have been demonstrated, a discrepancy possibly related to differences in dosage.497,555,623 Current
evidence suggests that occasional use of corticosteroids to manage acute episodes of joint pain may
be beneficial, but prolonged use and high doses should be avoided.389
1.1.7.3 Sulphated Heparinoids
Pentosan Polysulphate
Pentosan polysulphate (PPS) is produced by sulphation of xylanopyranose chains isolated from
beech tree hemicellulose. The chondroprotective properties of PPS have been extensively reviewed
by Ghosh (1999).257 PPS has been shown both in vitro and in vivo to promote chondrocyte anabolic
activity, and to limit cartilage catabolic events, particularly by inhibiting MMP activity. Its mode of
action is not fully known, though many of its actions are mediated at the level of gene expression.

80.
PPS increases synovial fibroblast hyaluronan synthesis, and increases the mean size of hyaluronan
molecules synthesized,365 thereby improving the viscoelastic and lubricating properties of synovial
fluid. It has also been shown to up-regulate synovial release of TIMP-3,819 and modulate release of
nitric oxide and cytokines (IL-6) in rabbit synovial fibroblasts.772,774 Due to anti-thrombotic and
fibrinolytic activity presumably related to its heparinoid structure, PPS may improve capillary
microcirculation in inflamed synovium and subchondral bone.239,863 Numerous trials of PPS in
rabbit, canine, and ovine models of OA have demonstrated a chondroprotective and anti-
inflammatory effect.18,239,257,670,772,774
Polysulphated-glycosaminoglycan
Polysulphated glycosaminoglycan (PS-GAG), like PPS, contains highly sulphated
glycosaminoglycan repeats (mainly chondroitin sulphate) but is sourced from animal rather than
plant sources. It has been shown to have similar anabolic and anti-catabolic effects to PPS,151
though experimentally its effects in cartilage are somewhat inconsistent.101 It has been shown to be
protective in various arthritis models including rabbit immobilization,271 intra-articular injection of
iodoacetate in horses,929,930 and canine meniscectomy313 and ACLT models.19 As with PPS, PS-
GAG increases both the rate of synthesis and molecular weight of hyaluronan produced by synovial
fibroblasts.238,584 It has been shown to increase the level and polymerisation state of synovial fluid
HA after intra-articular injection in humans.866
1.1.7.4 Hyaluronan Preparations
Various purified preparations of hyaluronic acid (HA) are available for intra-articular use. Its mode
of action is poorly understood; though its immediate effect is viscosupplementation of HA-depleted
joint fluid, it is rapidly cleared from the joint cavity hence any lasting effects cannot be due to a
simple replacement function. Its primary action may via an anti-inflammatory and anti-catabolic
effect on synovial fibroblasts. Smith and Ghosh (1987)770 showed that synovial fibroblast HA

81.
synthesis is markedly influenced by the nature (especially molecular weight) of HA in the
extracellular environment. HA has been shown to reduce the release of catabolic mediators such as
IL-1ß by synovial fibroblasts in vitro.816 Animal models suggest HA is most protective where
cartilage degradation is linked to synovial inflammation, and is less effective where overloading is
the primary cause.256,259 Some studies have found that the protective effect of HA is influenced by
the molecular weight of the preparation.39 However, others have found this is not important.751
1.1.7.5 Oral Supplements
The majority of oral nutritional supplements or "nutraceuticals" promoted for the treatment of
osteoarthritis contain glucosamine or chondroitin sulphate, either as a crude extract or in some
purified form. Traditional extracts such as shark cartilage235 and green-lipped mussel extract863
contain similar active ingredients. The widespread use of such compounds has been based on the
premise that nutritional supply of glycosaminoglycan precursors in supraphysiological quantities
will promote cartilage repair processes. Glucosamine has been shown to increase proteoglycan
synthesis in IL-1ß-stimulated rat chondrocytes.278 It has also been shown to suppress in vitro release
of phospholipase A2, PGE2, collagenase, stromelysin, and NO by chondrocytes.278,643 Other authors
have suggested that the primary action of glucosamine may be via promotion of synovial HA
production, thus providing a mechanism for the rapid clinical improvement often observed.527
Though evidence for in vivo efficacy has been largely anecdotal, increasing evidence of a beneficial
effect is emerging from controlled clinical trials.357,829
Ginger extracts787 and essential fatty acid supplements544 are other oral preparations reported to be
beneficial in osteoarthritis. One problem with evaluating such compounds is the magnitude of the
placebo effect in human patients. For example, even patient interaction as slight as monthly
telephone calls have been shown to have a significant effect on functional indices of OA.679

82.
1.1.7.6 Potential Future Therapies
Anti-cytokine Therapies
Increased understanding of the importance of cytokines in OA has lead to the development of
therapies that target their induction or activity. Inhibitors of TNF- -converting enzyme (TACE) and
IL-1ß-converting enzyme (ICE) are promising agents currently in development.621 Diacerein (4,5-
diacetoxy-9,10-anthroquinone-2-carboxylic acid) and its metabolite rhein also show promise as
anti-IL-1 therapies. In both OA cartilage and synovium in vitro, diacerein reduces IL-1 synthesis,
IL-1 receptor levels, ICE activity, and MMP expression.511,622 Diacerein may also modify
subchondral bone responses in OA.368,622 Diacerein has been shown to effectively reduce OA
symptoms in human638 and ovine368 trials; however, gastrointestinal side-effects were observed
which may limit its usefulness. More specific molecular targeting of cytokines may be possible in
the near future. Gene transfer of IL-1Ra was successful in suppressing both joint inflammation and
cartilage erosion in an animal model.492 However, approaches targeting TNF-R55 have proved
disappointing.557
MMP Inhibitors
Agents inhibiting MMP activity have been the focus of intensive research over the last decade.
Given the predominant role of MMPs in cartilage degradation, it is likely that such agents would be
of significant benefit in OA. However, the low cartilage permeability of MMP inhibitors so far
developed has limited their efficacy.639,871 Tetracycline and its synthetic derivatives possess potent
MMP inhibitory activity independent of their antimicrobial activities.23,871 Doxycycline inhibits
activity and synthesis of MMP-1, -8, and -13 in cell culture,311,754,755 and has been shown to reduce
the progression of OA in the canine ACLT model.931 However, no disease-modifying effect was
found in a clinical trial of doxycycline in RA patients.790

83.
Nitric Oxide Synthase Inhibitors (NOSI)
As both physiological and pathological roles have been identified for NO, the in vivo effects of
NOS inhibitors (NOSI) on the degradative and inflammatory changes of arthritis are difficult to
predict.381 To date, experimental studies in animal models of arthritis have consistently shown
NOSI to confer a beneficial effect. NOSI suppress the progression of arthritis, particularly synovial
inflammation, in acute inflammatory models such as rat adjuvant arthritis,792 streptococcal cell wall
arthritis,525 and rabbit antigen-induced arthritis.608 As NO is a principal effector of immune-
mediated vascular injury and modulator of leucocyte function,215 these findings are to be expected.
However in the zymosan-induced arthritis model, cartilage from iNOS knockout mice showed
reduced pathology, markedly less inhibition of proteoglycan synthesis, and normal IGF-1
responsiveness, while joint inflammation was similar to that of wild-type mice.854,856 NOSI were
also effective in restoring proteoglycan synthesis after intra-articular administration of IL-1ß in
rats.649 This suggests that NOSI might possess chondroprotective capabilities separate from any
anti-inflammatory effects.
Convincing evidence that NOSI might possess disease-modifying properties in the less
inflammatory, more slowly progressive disorder of osteoarthritis is still lacking, though suppression
of NO production has been implicated in the chondroprotective effect of many existing OA
therapies, such as tetracyclines.23,53,235 Using the canine ACLT model, Pelletier and coworkers have
shown that the selective iNOS inhibitor L-N-iminoethyl-L-lysine (L-NIL) reduces development of
cartilage and synovial pathology, synovial fluid IL-1ß, and chondrocyte immunostaining for iNOS,
nitrotyrosine, MMP-1 and -3, and caspase 3, a marker of apoptosis. No evidence of toxic effects
was recorded.630,631,633,634 However, these studies were short (10-12 weeks of treatment), and grading
of joint pathology was somewhat subjective. In addition, ACLT was performed by a blind stab
incision, a technique criticised for creating significant intra-articular haemorrhage and subsequent

84.
synovial inflammation.569 Nevertheless, this data is encouraging and further trials of NOSI in other
OA models should ensue. Issues of potential toxicity (in processes as varied as cardiovascular
regulation, wound repair, and immune response) need to be better addressed before clinical trials
are attempted.359 Such toxic effects will be lessened, but not completely eliminated, by using
selective inhibitors of iNOS. Other interactions might be predicted from known actions of nitric
oxide. For example, NOS inhibition may reduce PGE2 production within the OA joint. Preliminary
evidence from the canine ACLT model suggests this is indeed the case, as L-NIL treatment reduced
canine synovial fluid PGE2 content and cartilage COX-2 expression.630 However, NOSI may
accentuate local bone resorptive processes in arthritis, as well as potentially influencing peripheral
bone density.199,405,906
1.2 STUDY BACKGROUND
Osteoporosis is the most common metabolic bone disease in the developed world, and represents a
substantial and escalating burden to healthcare services.615 The disorder is characterised by reduced
bone mineral density, associated with bone fragility and an increased risk of fractures. Whilst
hormone replacement therapy of oestrogen supplementation is an effective treatment, compliance is
often poor due to concern over potential hormonal side-effects such as withdrawal bleeding, weight
gain, breast tenderness, and increased risk of uterine and breast cancer.204,615,720 Despite the potential
of newer therapies such as bisphosphonates and selective oestrogen receptor modulators
(SERMs),466,720 an acceptable but inexpensive therapy for osteoporosis (one which restores bone
density and trabecular connectivity with minimal side-effects) remains a major goal in
pharmacological research.615
As recent research has suggested that the skeletal and cardiovascular protective effects of oestrogen
are largely mediated by nitric oxide, nitric oxide donors might therefore be novel and effective

85.
therapies for osteoporosis, eliminating the concomitant side-effects associated with the use of
oestrogen replacement therapy. Wimalawansa and co-workers have demonstrated that glyceryl
trinitrate prevents both ovariectomy- and corticosteroid-induced bone loss in rats,904,906 and have
begun trials in human patients.902 Early results of clinical trials have shown a significant protective
effect against bone loss following surgical menopause, equivalent to that of oestrogen
supplementation.902 Similarly, intermittent use of nitrates by elderly women has been
epidemiologically associated with increased bone mineral density in the hip and heel.380
However, NO is suspected to be a key mediator involved in the inflammatory and catabolic changes
in cartilage in arthritic disorders such as RA and OA.381,789 NO production is elevated within OA
joints,25,49,538,609,637,676,791 and NO has been shown to have multiple effects on joint tissues in vitro,
including reduction in articular cartilage proteoglycan synthesis,349,393,801,822 increased MMP
synthesis,561 and extensive interactions with other inflammatory mediators.346,532,637 Experiments
using iNOS inhibitors in two rodent models of inflammatory arthritis525,792 and a canine OA
model619 have shown that such compounds reduce the progression of arthritic changes. However,
knowledge of the net effects of NO-donors on cartilage metabolism in vivo is presently unknown
and requires urgent clarification given not only their proposed therapeutic use in preventing
osteoporosis, but also their current routine use in the treatment of angina and other cardiac disorders
in elderly patients, who may also be osteoarthritic.
The in vivo effects of NO-donors also need to be considered in relation to subchondral bone, given
the hypothesis by some authors that subchondral changes may be central to the early pathogenesis
of OA.184,618,659 It might be proposed that NO may influence the progression of arthritis by
promoting subchondral bone sclerosis, increasing subchondral bone stiffness and the resultant
stresses transmitted to chondrocytes. Furthermore, by promoting angiogenesis,937 NO may play a
role in the neovascularization of the calcified cartilage zone that accompanies subchondral
remodelling as a histological hallmark of OA.346,791

86.
Any evaluation of the effects of nitric oxide needs to encompass study of its interactions with
oestrogen status. Oestrogen levels and NO synthesis are closely linked in many
tissues,36,406,666,676,884,890 and plasma levels of NO and oestrogen are closely correlated.703 Oestrogen
is itself suspected to play an important role in joint tissue homeostasis.36,666,676,884,890 There is
abundant but conflicting epidemiological evidence that oestrogen plays a role in OA, most
consistent of which is the rise in OA incidence at the time of menopause,577,578,782 and the protective
effect of oestrogen replacement therapy.577,719,783,934 Experimental evidence is conflicting: while
administration of oestrogen has been shown to cause cartilage degeneration in animal
studies,699,701,834,837 ovariectomy of ewes induces biomechanical deterioration of articular cartilage
preventable by oestrogen replacement.841
The importance of oestrogen in OA may be primarily due to its effects on bone.226 The role of
oestrogen in maintaining bone mass is well known. Therefore its relative role in the OA process
depends on whether subchondral bone attrition or sclerosis is relatively more important in the
pathogenesis of OA, a factor which is currently unknown. It has been suggested that the protective
effect of oestrogen replacement therapy in OA is related to a reduction of bone turnover and
stabilisation of subchondral bone remodelling.578 Oestrogen withdrawal (or menopause) might
therefore be expected to worsen concurrent osteoarthritis by increasing subchondral bone loss.
However, osteoporosis and OA are negatively correlated epidemiologically.312,779
In order to investigate the subtle structural changes in joint tissues induced by pharmacological
interventions, a method of objectively quantifying these changes is required. Recent improvements
in the processing power of personal computers, and the availability of sophisticated image analysis
software, has lead to the application of computerised image analysis methodologies to structural
assessments in arthritis.126,299,384,455,750 However published methods vary widely, and no consensus
yet exists regarding the best ways to apply this technology.

87.
1.3 AIMS AND HYPOTHESES
The broad aims of this project were to:
i) to develop and apply novel and published computerised image analysis methods for
histomorphometric analysis of structural changes in cartilage and subchondral bone in the
ovine meniscectomy model of osteoarthritis.
ii) to further develop understanding of the OA process in this well-established surgical model,
particularly with respect to the role of synovial membrane and subchondral bone changes
within the overall pathogenesis.
iii) to extend earlier studies367,611,769 demonstrating significant structural and biochemical
alterations in joint tissues following ovariectomy in sheep.
iv) to assess the influence of prior ovariectomy on the development of cartilage, synovial, and
subchondral bone lesions following ovine meniscectomy.
v) to investigate the effects of an exogenous nitric oxide donor, glyceryl trinitrate, on the
structural and metabolic integrity of joint tissues in normal, ovariectomised, and
osteoarthritic (meniscectomised) sheep.
The specific hypotheses to be tested were:
i) computerised image analysis methodologies would be more sensitive to structural
alterations in joint tissues in the ovine meniscectomy model of OA, compared to traditional
semi-quantitative scoring methods.
ii) the gross synovitis thought to occur in this OA model474 would be accompanied by
demonstrable microscopic pathology, and persistent metabolic alterations in synoviocytes
in vitro.

88.
iii) ovariectomy of aged ewes would induce structural and metabolic alterations in femoro-
tibial articular cartilage, as well as synovial and subchondral bone tissue.
iv) prior ovariectomy of ewes would worsen and/or accelerate the progression of joint changes
following meniscectomy.
v) the topical use of an exogenous nitric oxide donor, glyceryl trinitrate (GTN), would reduce
the loss of bone mineral density following ovariectomy in aged ewes, as has been shown in
other species.902,904,906
vi) the same dose of GTN which produces the above skeletal effects would adversely affect the
structural and metabolic integrity of normal articular cartilage, in keeping with the known
degradative effects of NO on chondrocytes.78,140,322,802,803,822
vii) given the net effect of NO varies in physiological and pathological conditions381,552 and NO
is a key mediator of oestrogen activity,36,329,330,406,666,676,884,890,906 that the response of joint
tissues to GTN treatment would vary between normal animals, ovariectomised animals, and
animals with surgically-induced OA.
viii) given nitric oxide synthase inhibitors reduce the progression of experimental
arthritis,525,619,792 that GTN treatment would worsen and/or accelerate the progression of
joint changes in the ovine meniscectomy model of OA.

89.
CHAPTER 2:
DEVELOPMENT OF NOVEL METHODS OF HISTOLOGIC
ASSESSMENT OF CARTILAGE AND SYNOVIUM, IN A TRIAL OF
ORAL AVOCADO-SOYA UNSAPONIFIABLES
2.1 INTRODUCTION
Traditionally, histological analysis of cartilage pathology has been qualitative. Whilst histological
grading systems have been used, such as that developed by Mankin,498 these are at best semi-
quantitative, subjective, and prone to inter-user variability. Recent applications of image analysis
systems to quantitative analysis of cartilage changes in arthritis demonstrate the potential and
acceptance of this technology.126,299,384,750,927 Shimizu et al. (1997)750 demonstrated that
determination of cartilage area and safranin-O staining intensity by computerised image analysis
allowed reliable and precise evaluation of cartilage deterioration in a rabbit model of osteoarthritis.
In the past, management of OA has primarily focussed on the relief of symptoms, using agents such
as analgesics and NSAIDs. Such an approach has been criticised for failing to prevent continued
articular cartilage degeneration, and in the case of certain NSAIDs, exacerbating the process by
inhibiting proteoglycan synthesis.197 Identifying agents which are capable of preventing, slowing, or
reversing the structural and pathological alterations which occur in OA joints is an important
research objective. Such agents, termed disease-modifying osteoarthritis drugs (DMOADs), are
difficult to evaluate in clinical trials, due to the slow progression of OA in humans and the
challenge of assessing joint changes by non-invasive methods.259,463 The use of animal models

90.
permits the timing of onset to be known, producing a homogeneous, reliable model of the OA
process for the evaluation of disease-modifying therapies.258,260
Avocado-soya unsaponifiables (ASU) is a novel anti-arthritic agent derived from unsaponifiable
residues of avocado and soya oils, mixed in the ratio of 1:2 respectively. ASU have been shown in
various in vitro systems to partially reverse the effects of IL-1ß on cultured chondrocytes,
stimulating collagen synthesis and inhibiting the production of matrix metalloproteases, IL-6, IL-8
and PGE2.317,345,517,518 A similar effect has also been demonstrated in rheumatoid synoviocytes.517,518
Boumediene et al. (1999)86 recently reported that ASU enhanced the expression of transforming
growth factor-ß (TGF-ß) in cultured articular chondrocytes, providing the first clues as to its
possible mode of action. The active ingredients of the plant extract are unknown, but synergism
between the avocado and soya components, and their relative ratios, appears to be important.345,491
The only published animal study found oral ASU significantly prevented the occurrence of cartilage
lesions in a rabbit post-contusive model.522 While its symptomatic efficacy was recently
demonstrated in double blind, placebo-controlled prospective clinical trials in patients with
osteoarthritis,82,491 including evidence of reduced joint space narrowing,464 its ability to also protect
the articular cartilage of osteoarthritic joints in vivo remains to be established.
Many clinical studies have used qualitative or semi-quantitative scoring systems to grade
synovitis.434,468,471,569,613,697 While these studies have mostly examined rheumatoid synovia, several
have examined the synovial changes accompanying OA.470,568,682,768 Smith et al. (1997)768 found that
in patients with early OA, a significant increase in synovial thickness, vascularity, inflammatory
cell infiltration, and cytokine production was consistently detected even in early stages of the
disease. While synovitis is known to occur in the ovine meniscectomy model of OA,474
histopathological changes have not been systematically examined.

91.
The purpose of this trial was to: (1) examine the effect of ASU on the histological progression of
joint changes in the ovine lateral meniscectomy model of OA; (2) compare traditional qualitative
methods of assessing cartilage histological changes, with a novel computerised histomorphometric
method; and (3) assess the level of microscopic synovitis developed in this model, using a
qualitative scoring system.
2.2 MATERIALS AND METHODS
2.2.1 TRIAL PROTOCOL
2.2.1.1 Source of Animals
All animal procedures were approved by the Murdoch University Animal Ethics Committee. Forty-
eight mature (three year old) Merino wethers were obtained from a single source. Animals were
selected for uniformity of size, conformation, soundness of health, and for absence of lameness. On
introduction to the Murdoch University farm, the sheep were ear-tagged and vaccinated against
clostridial diseases and Corynebacterium pseudotuberculosis [Glanvac 6, CSL, Australia]. All
sheep were treated prophylactically for endo- and ectoparasites [Ivomec Liquid for Sheep
(ivermectin), MSD Agvet, Australia; Vanquish, (alphamethrin 50 g/L), SmithKline Beecham,
Australia] and to prevent footrot [Footrite (zinc sulphate 224 g/L), Hardman, Australia].
2.2.1.2 Meniscectomy
Animals were assigned randomly to one of six treatments groups. In four of the groups of 8 sheep
(n=32), bilateral lateral meniscectomy was performed. Surgery on both left and right joints was
performed within a single anaesthetic procedure. The day before surgery, three 50 g/hr fentanyl
patches [Durogesic, Janssen-Cilag, Belgium] were applied to a clipped area of skin in the mid-

92.
thoracic area, and held in place with a canvas band. These patches remained in place for 3 days and
have been shown to provide effective analgesia in sheep.203 After deprivation of food and water
overnight, the sheep were anaesthetised with intravenous thiopentone sodium (50mg/ml) [Thiobarb,
Pitman-Moore, Australia]. Anaesthesia was maintained with inhaled halothane [Rhône Mérieux,
Australia] using a Fluotec 3 vaporiser [Cyprane Keighley, UK] and a circle anaesthetic circuit. Both
hindlimbs were clipped and aseptically prepared, and 1g of cephazolin sodium [David Bull
Laboratories, Australia] was injected intravenously immediately prior to surgery. A 10cm
parapatellar incision was made on the lateral aspect of the femoro-tibial joint, and dissection
continued through incision of the fascia lata, crural retinaculum, and joint capsule to expose the
lateral meniscus. Bleeding was controlled throughout surgery using electrocoagulation. All
peripheral capsular attachments of the meniscus were stripped using a meniscotome, before
severing the cranial tibio-meniscal ligaments using a number 15 scalpel blade. The cranial pole of
the meniscus was grasped and pushed caudally under the collateral ligament and popliteus tendon.
With caudal retraction of the skin and fascia lata, a second fascial incision was made caudal to the
collateral ligament to expose the translocated cranial pole of the meniscus. This was grasped with a
Backhaus towel clip and retracted caudolaterally. With tension applied to the meniscus, a
meniscotome was used to locate and sever the femoro-meniscal ligament and any remaining caudal
tibial attachments, allowing removal of the meniscus. The joint was then lavaged with sterile saline
before closure of the the retinacular and fascial incisions using 3 metric polydioxanone sutures
[PDS II, Ethicon, USA] in a continuous pattern. Subcuticular tissues were apposed in two layers
using 3 metric poliglecaprone 25 sutures [Monocryl, Ethicon, USA], and the skin closed using 2
metric braided polyglactin 910 [Vicryl, Ethicon, USA] in a buried intradermal pattern. Following
surgery, all animals received 100mg of flunixin meglumine by intramuscular injection [Ilium
Flunixin, Ilium, Australia]. The sheep were maintained under close observation for 3-5 days in pens
within a covered barn area, before return to pasture. Complications were infrequent, and were
mostly limited to transient lameness or seroma formation at the surgical site.

93.
2.2.1.3 Trial Protocol
Following a brief recovery period, sheep were maintained on irrigated pasture and supplemented
with lupin grain daily. From approximately one month postoperatively, meniscectomised groups
were given orally administered capsules of ASU (900mg/weekday) [Piascledine 300®] or placebo
(paraffin oil) both provided by Pharmascience [Laboratoires Pharmascience, Courbevoie, France].
Dosing was not performed at weekends. This dose, higher than the usual daily human dose, was
used to compensate for the weekday-only administration protocol and the possibility of partial
degradation of the compound in the rumen. Body weights were recorded at approximately monthly
intervals. Treatment groups were therefore: meniscectomy plus ASU (MX+ASU), meniscectomy
plus placebo (MX), or non-operated control (NOC). Eight sheep of each treatment type were
euthanased at approximately three months and six months postoperatively (except the six-month
MX group, which contained only seven animals, due to the death of one animal from neurological
symptoms of unknown origin).
2.2.1.4 Sacrifice and Tissue Processing
At sacrifice, the sheep were killed by intravenous injection of 20ml of pentobarbitone sodium (300
mg/ml) [Valabarb, Pitman-Moore, Australia]. Joints were immediately dissected and the soft tissues
removed. Histological slides were prepared from each of the following regions: medial femoral
condyle (MFC), lateral femoral condyle (LFC), medial tibial plateau (MTP), lateral tibial plateau
(LTP), and trochlear groove (TG) (six months post-op only). Osteochondral slices (approximately
3mm thick) were taken from each region by bandsaw cuts in the medio-lateral plane, perpendicular
to the articular surface. Cuts were positioned in the femoral condyles at the point of tibial contact in
normal stance, and in the tibial plateaux at the level of the intercondylar eminences. Bone slices
were fixed in 10% neutral buffered formalin for 48 hours, then decalcified in 10% formic acid / 5%
formaldehyde for 7 days. The specimens were then washed, dehydrated in increasing alcohol

94.
concentrations (70-100%), and double-embedded in methyl benzoate/celloidin and paraffin wax.
4 m sections were cut on a rotary microtome and adhered to treated glass slides [Superfrost Plus,
Menzel Gläser, USA].
Toluidine blue staining was performed in batches under constant conditions. Sections were
deparaffinised in 70% alcohol for 15 minutes, stained in 0.04% toluidine blue / 0.1M sodium
acetate buffer (10 minutes) then 0.1% fast green (2 minutes), dehydrated in isopropanol then
xylene, and coverslipped.
2.2.2 GROSS PATHOLOGY
At six months, colour slide photographs were taken of the opened joints. Four joint regions (MFC,
LFC, MTP and LTP) were later scored for cartilage damage and osteophyte formation, by a single
blinded operator, according to the following scale:39
Cartilage: Osteophytes:
0 = normal 0 = none 1 = roughened 1 = slight osteophyte development 2 = fibrillated / fissured 2 = moderate osteophyte development 3 = small erosion (<5mm) 3 = marked osteophyte development 4 = large erosion (>5mm)
Gross synovial pathology was noted at the time of joint dissection, according to the following scale:
Synovium (all regions):
0 = normal 1 = slight abnormality (e.g. redness or fibrosis) 2 = synovitis

95.
2.2.3 CARTILAGE HISTOPATHOLOGICAL SCORING
Semi-quantitative histopathological scoring was performed on 4 m toluidine blue-stained sections
according to a published modification474,926 of Mankin’s grading system.498 In one section from each
of the four joint regions, four zones were scored: (1) outer (abaxial) margin; (2) outer perilesional
zone; (3) lesional zone (or spatially equivalent zone in controls); and (4) inner perilesional zone.
Scoring was done by a single observer, according to a five point scale (Figure 2.1), and a mean
aggregate score determined as the average of these four zonal values (maximum possible score =
26).
2.2.4 HISTOMORPHOMETRIC IMAGE ANALYSIS
Computer-assisted histomorphometric image analysis was conducted in a manner similar to that
described by Shimizu et al. (1997).750 Sections were mounted in a slide cartridge customised to hold
glass slides, scanned directly using a slide scanner [Microtek Slidescanner 35t plus (Model PTS-
1950), Microtek Lab Inc., USA] and acquired at 400 dpi resolution using standard graphics
software [ScanWizard v3.0.7, Microtek Lab Inc., USA; Adobe Photoshop v3.0, Adobe Systems,
USA]. Stored image files were analysed using image analysis software [Image Pro Plus v3.0.1,
Media Cybernetics, USA] on a personal computer [Power Macintosh 7600/200, Apple Computer
Inc., USA]. Spatial calibration was performed by scanning a 10x10mm high precision graticule, and
the intensity scale was calibrated to give a positive linear response to increasing optical density
(white=0, black=255).

96.
Figure 2.1: The cartilage histopathological scoring system used in this study (after Little et al. (1997)474) I Structure
0 normal 1 slight surface irregularities 2 moderate surface irregularities 3 severe surface irregularities 4 clefts / fissures into transitional zone (1/3rd depth) 5 clefts / fissures into radial zone (2/3rds depth) 6 clefts / fissures into calcified zone (full depth) 7 fibrillation and/or erosion to transitional zone (1/3rd depth) 8 fibrillation and/or erosion to radial zone (2/3rds depth) 9 fibrillation and/or erosion to calcified zone (full depth) 10 fibrillation and/or erosion to subchondral bone
II Cellularity
0 normal 1 increase or slight decrease 2 moderate decrease 3 severe decrease 4 no cells present
III Chondrocyte Cloning
0 normal 1 several doublets 2 many doublets 3 doublets and triplets 4 multiple cell nests
IV Territorial Toluidine Blue Staining
0 normal 1 increase or slight decrease 2 moderate decrease 3 severe decrease 4 no staining
V Interterritorial Toluidine Blue Staining
0 normal 1 loss of staining to tangential zone (1/3rd depth) 2 loss of staining to transitional zone (2/3rds depth) 3 loss of staining to radial zone (full depth) 4 no staining
Maximum aggregate score = 26

97.
Using the image analysis software, markers were applied such that the cartilage length was divided
into three zones each comprising approximately 1/3rd of the total arc length, termed the inner,
middle, and outer zones (Figure 2.2). The middle zone included the usual point of erosion in the
lateral compartment of meniscectomised animals. In each of the three zones, the intensity of
toluidine blue staining, an index of proteoglycan content,253 was determined as the mean pixel
optical density after conversion from colour to greyscale (Figure 2.4). Uncalcified cartilage and
subchondral bone plate thickness (mm) were determined from Masson’s Trichrome-stained sections
with similarly defined zones, using a radial thickness algorithm within the software to generate
mean thickness measurements between lines delimiting the cartilage surface, most advanced
tidemark, and limit of cortical subchondral bone (intertrabecular spaces < 500 m). Areas of obvious
osteophyte growth were excluded from analyses.
Figure 2.2: Schematic representation of the femoro-tibial joint in cross-section, showing division
of the articular cartilage and subchondral bone plate of each condyle into inner (I), middle (M), and
outer (O) zones, for the purposes of histomorphometric analysis. LFC, lateral femoral condyle; LTP,
lateral tibial plateau.

98.
Reproducibility of this method was assessed by repeat scanning and analysis of ten sections (data
not shown), and was found to be high. Mean variation of repeated thickness analyses was ±5%, and
that of mean staining intensity measurements was ±2%.
2.2.5 SYNOVIAL HISTOPATHOLOGICAL SCORING
Specimens of synovium were obtained from the suprapatellar fold at the time of sacrifice and fixed
in 10% neutral buffered formalin. Histopathological scoring was performed on 4 m haematoxylin
and eosin-stained sections by a single blinded observer, according to the following scale (modified
from Smith et al. (1997)768) (Figure 2.3):
Figure 2.3: The synovial histopathological scoring system used in this study A: Intimal hyperplasia 0 normal (1-2 layers)
1 mild-moderate, focal thickening (3-4 layers) 2 mild-moderate, diffuse thickening 3 moderate-marked, diffuse thickening (>5 layers)
B: Lymphocytic / plasmacytic 0 none infiltration 1 mild-moderate, focal infiltrate
2 mild-moderate, diffuse infiltrate 3 moderate-marked, diffuse infiltrate
C: Subintimal / perivascular fibrosis 0 none 1 slight fibrosis 2 moderate fibrosis 3 marked fibrosis
D: Vascularity 0 normal 1 slight increase in vascular elements 2 moderate increase in vascular elements 3 marked increase in vascular elements

99.
2.2.6 STATISTICAL ANALYSIS
Statistical comparisons were generated using specialist software [Statview 5.0, SAS Institute Inc.,
USA], performing one-way analysis of variance (ANOVA) to analyse variance across treatment
groups, and Fisher’s analysis of least significant difference (Fisher’s PLSD) to compare treatment
group means, except where indicated. A significance level of p=0.05 was used throughout this
study. The medial compartment of the joint was found to be significantly affected by lateral
meniscectomy, preventing the use of the medial compartment as an internal control.

100.
2.3 RESULTS
2.3.1 BODY WEIGHT
There was no significant difference in body weights between the three groups before the trial
commenced (Figure 2.7). Animals were in moderately lean body condition. After surgery, mean
body weights of meniscectomised animals were significantly below those of non-operated control
animals. Mean body weights of animals receiving placebo capsules were consistently lower than
those of animals receiving ASU, significantly so at 5-6 months post-operatively.
2.3.2 GROSS PATHOLOGY
Meniscectomised joints showed gross pathological changes comparable with early human OA, with
articular cartilage erosion in the lateral compartment (milder changes in the medial compartment),
and moderate-marked osteophyte development (Figure 2.5). In control animals, gross changes were
restricted to occasional cartilage roughening, and/or slight osteophyte formation, in the medial
compartment. Joints of animals treated with ASU tended to have lower cartilage scores than non-
treated (Table 2.1) though this difference was not statistically significant. Osteophyte values were
similarly high in both groups. MX+ASU sheep showed a tendency for a greater degree of gross
synovitis than MX, which approached statistical significance (p=0.066).
2.3.3 CARTILAGE HISTOPATHOLOGICAL SCORING
Articular cartilage sections from meniscectomised animals showed central areas of fibrillation and
erosion, with loss of chondrocytes and toluidine blue staining. Adjacent regions showed
chondrocyte cloning and increased toluidine blue staining intensity (Figure 2.6). Mean
histopathological scores were higher in meniscectomised animals compared to those of NOC. This

101.
difference was significant only in the lateral compartment at three months (data not shown), but in
all regions at six months post-op (Table 2.2). There were no significant differences between
aggregate scores of ASU-treated animals and those of the placebo group, at three or six months.
MX+ASU animals showed a tendency for lower scores than MX in the LFC at six months which
approached statistical significance (p=0.0504).
2.3.4 HISTOMORPHOMETRIC IMAGE ANALYSIS
Intensity of Toluidine Blue Staining
At three months post-op (Figure 2.8), the mean intensity of cartilage toluidine blue staining tended
to increase in the medial compartment, but decrease in the lateral compartment, primarily due to
loss of stain in the area of central erosion. At six months post-op (Figure 2.9), loss of toluidine blue
stain predominated, except in the trochlear groove. MX+ASU animals showed either a greater
increase or decreased loss of stain relative to MX in all regions, statistically significant in the MTP
(90±9 vs 78±11, p=0.015), LTP (81±4 vs 67±9, p=0.001), and TG (104±9 vs 93±7, p=0.023;
NOC=89±6, p=0.011) at six months post-op.
Uncalcified Cartilage Thickness
Mean uncalcified cartilage thickness tended to increase following meniscectomy. At three months
post-op (Figure 2.8, left) increased cartilage thickness was prominent in the MFC, and outer zones
of the LFC and LTP. At six months post-op (Figures 2.10 and 2.11, above), cartilage thicknesses
tended to be higher in MX+ASU than MX, especially in the middle (lesion) zone. In the middle
zone of the LFC, MX+ASU cartilage was significantly thicker than MX (p=0.0181) or NOC. In the
middle zone of the LTP, MX cartilage tended to be thinner than both NOC and MX+ASU
(p=0.0506) values.

102.
Subchondral Bone Plate Thickness
Subchondral bone plate (SCP) thickness (Figures 2.10 and 2.11, below) tended to increase in the
lateral compartment following meniscectomy. This increase was greatest in the outer zone, but was
significant in all zones. In the inner zone of the LTP, significantly lower SCP thickness was found
in MX+ASU compared with MX animals (p=0.049).
2.2.5 SYNOVIAL HISTOPATHOLOGICAL SCORING
Meniscectomised sheep showed a significant increase in lymphoplasmocytic infiltrate, subintimal
fibrosis, and subintimal vascularity relative to control sheep (Table 2.3). MX+ASU sheep showed a
significantly greater degree of fibrosis (p=0.010) and sum of scores (p=0.042) relative to MX
animals.

103.

104.
Figure 2.5: Gross appearance of the femoral condyles (left) and tibial plateaux (right)
of non-operated control (above) and meniscectomised (below) joints. Note cartilageerosion and extensive osteophyte formation six months after meniscectomy. MED,medial; LAT, lateral.
MED
MED MED
MEDLAT
LAT LAT
LAT
Figure 2.6: Histological appearance of toluidine blue-stained articular cartilage in thetibial plateaux of non-operated control (left) and meniscectomised (right) sheep. Notesurface fibrillation and fissure formation, with regions of proteoglycan depletion andchondrocyte death adjacent to regions of attempted hypertrophic repair.

105.
Figure 2.7: Body weights (mean ± SE) of non-operated control (NOC), meniscectomised (MX),
and ASU-treated meniscectomised (MX+ASU) animals, by date (approximately monthly intervals).
* MX+ASU differs significantly from MX, p<0.05
Table 2.1: Gross pathology scores (mean ± SD) at six months in control (NOC), meniscectomised
(MX), and ASU-treated meniscectomised (MX+ASU) animals.
* differs significantly from NOC (p<0.05) ; a: p=0.066 (MX vs MX+ASU)
Treatment Group Cartilage
fibrillation
(max = 16)
Osteophyte
formation
(max = 12)
Gross
Synovitis
(max = 2)
NOC 0.56±0.73 0.37±0.62 0.06±0.25 MX 6.86±2.11* 6.43±1.91* 0.71±0.91* MX+ASU 6.19±0.36* 6.37±1.54* 1.19±0.75*a
Start Wt 13.3.98 15.4.98 13.5.98 1.6.98 29.6.98 27.7.98 26.8.9845
47
49
51
53
55
57
59
61
63
65
67
MX+ASU
MX
NOC
Bo
dy W
eig
ht
(kg)
*
*

106.
Table 2.2: Mean aggregate histopathology scores (mean ± SD) at six months, in the lateral and
medial femoral condyles (LFC, MFC) and tibial plateaux (LTP, MTP) of control (NOC),
meniscectomised (MX), and ASU-treated meniscectomised (MX+ASU) animals.
* differs significantly from NOC (p<0.05), b: p=0.0504 (MX vs MX+ASU); maximum score = 26
Treatment Group LFC MFC LTP MTP
NOC 3.12±1.00 2.53±0.94 4.43±1.81 3.91±1.57 MX 9.07±0.45 4.32±1.46* 9.04±1.81 5.36±1.48 MX+ASU 8.03±1.22*b 4.00±0.94* 9.00±1.37* 5.97±1.77*
Table 2.3: Synovial membrane histopathology scores (six months post-meniscectomy) of control
(NOC), meniscectomised (MX), and ASU-treated meniscectomised (MX+ASU) animals.
* differs significantly from NOC (p<0.05), † MX+ASU differs from MX (p<0.05); maximum score = 3;
maximum sum = 12
Treatment
Group
Intimal
hyperplasia
Cellular
infiltrate
Subintimal
fibrosis
Vascularity SUM
NOC 1.50±0.53 0.50±0.76 0.87±0.64 0.75±0.71 3.62±2.07
MX 1.00±0.82 1.43±0.53* 1.00±0.58 1.14±1.21 4.57±2.76
MX+ASU 1.50±0.92 2.00±0.76* 2.12±0.83*† 1.87±0.64 7.50±2.39*†

107.
Figure 2.8: Uncalcified cartilage thickness (left) and proteoglycan content (right) (mean greyscale pixel intensity; white=0, black=255) of toluidine blue-stained 4µm histological sections from normal sheep (NOC), meniscectomised sheep (MX), and meniscectomised sheep treated with avocado-soya unsaponifiables (MX+ASU), in the inner, middle and outer zones of the lateral and medial femoral condyles (LFC, MFC) and lateral and medial tibial plateaux (LTP, MTP) three months post-operatively. * differs significantly from NOC (p<0.05); error bars represent one standard error.

108.
Figure 2.9: Cartilage proteoglycan content, measured as mean greyscale pixel intensity (white=0, black=255) of toluidine blue-stained 4µm histological sections from normal sheep (NOC), meniscectomised sheep (MX), and meniscectomised sheep treated with avocado-soya unsaponifiables (MX+ASU), in the inner, middle and outer zones of the lateral and medial femoral condyles (LFC, MFC) and lateral and medial tibial plateaux (LTP, MTP), and medial, middle, and lateral zones of the trochlear groove (TG) six months post-operatively. * differs significantly from NOC (p<0.05), † MX+ASU differs significantly from MX (p<0.05); error bars represent one standard error.

109.
Figure 2.10: Thickness (mm) of uncalcified cartilage (above) and the subchondral bone plate (below) in normal sheep (NOC), meniscectomised sheep (MX), and meniscectomised sheep treated with avocado-soya unsaponifiables (MX+ASU), in the inner, middle and outer zones of the lateral femoral condyle (LFC) (left) and the medial femoral condyle (MFC) (right) six months post-operatively. * differs significantly from NOC (p<0.05), † MX+ASU differs significantly from MX (p<0.05); error bars represent one standard error.

110.
Figure 2.11: Thickness (mm) of uncalcified cartilage (above) and the subchondral bone plate (below) in normal sheep (NOC), meniscectomised sheep (MX), and meniscectomised sheep treated with avocado-soya unsaponifiables (MX+ASU), in the inner, middle and outer zones of the lateral tibial plateau (LTP) (left) and the medial tibial plateau (MTP)(right) six months post-operatively. * differs significantly from NOC (p<0.05), † MX+ASU differs significantly from MX (p<0.05) a : p=0.0506 (MX vs MX+ASU); error bars represent one standard error.

111.
2.4 DISCUSSION
Hypothesis: that computerised image analysis methodologies would be more
sensitive to structural alterations in joint tissues in the ovine meniscectomy model
of OA, compared to traditional semi-quantitative scoring methods.
As shown in previous studies using this model, lateral meniscectomy induced thinning, fibrillation,
and erosion of articular cartilage secondary to altered joint congruity and load distribution in the
meniscus-deficient lateral compartment.258,260,264,474 In cartilage adjacent to these lesions, an increase
in cartilage thickness was evident, due to a combination of over-hydration of disrupted cartilage and
hypertrophic repair responses. In the early hypertrophic stages of experimental OA, increased
chondrocyte biosynthetic activity usually precedes the inhibition of synthesis or degradation of
proteoglycans, resulting in a period of increased proteoglycan concentration in the matrix.499,554,869
This early response is generally coupled with damage to the collagen network, particularly in the
superficial region, allowing over-hydration of aggrecan and volume expansion of articular cartilage.
The increase in cartilage thickness seen here following meniscectomy may be the result of cartilage
swelling, marginal osteochondral remodelling, and/or hypertrophic matrix production
accompanying attempted cartilage repair.635,869 However, focal cartilage erosion occurred
simultaneously, partially reversing this increase and giving meniscectomised animals mean
uncalcified thickness values in eroded zones similar to those of controls. The tendency for greater
cartilage thickness observed in the middle cartilage zone of ASU-treated animals, statistically
significant in the LFC (Figure 2.10), is therefore difficult to interpret but may be the result of
reduced cartilage erosion, and/or increased matrix production, in the presence of a disrupted
collagen network.
Quantitation of toluidine blue staining provides further evidence of partial preservation of articular
cartilage integrity by ASU following cartilage insult. Loss of toluidine blue staining was
significantly reduced by ASU treatment in both tibial plateaux at 6 months post-op (Figure 2.9). In

112.
the trochlear groove at six months post-op, toluidine blue staining of MX+ASU animals was
significantly greater than either MX or NOC animals. Again, the tendency for increased (or reduced
loss of) cartilage proteoglycan content in ASU-treated animals may be the result of decreased
catabolism and/or increased anabolism. The exaggerated compensatory increase in proteoglycan
production in areas distant to the insult, such as the trochlear groove, is suggestive of a pro-anabolic
effect. These findings are consistent with published in vitro studies showing ASU are capable of
stimulating matrix production and reducing the deleterious effects of IL-1ß on articular chondrocyte
metabolism.317,345,517,518 It was recently suggested that many of the actions of ASU may be mediated
by an increase in expression of TGF-ß,86 a potent stimulator of chondrocyte matrix production and
antagonist of the deleterious effects of IL-1.653,672,673,754,852 The results of this trial appear consistent
with such a mechanism of action.
Subchondral bone plate changes reflected altered joint mechanics resulting from meniscectomy, in
that SCP thickness decreased in the outer zone of the medial compartment (reduced load-bearing),
and increased in the lateral compartment, especially in zones previously protected by the meniscus.
Significantly reduced subchondral sclerosis in the inner zone of the LTP in ASU-treated animals
suggests modification of OA-stimulated subchondral events by this drug. However, as SCP
thickness changes were more pronounced in other zones, in which the drug showed no effect, the
importance of this effect is unknown.
The use of a synovial histopathological scoring system was found to be useful in demonstrating the
effects of ASU. While the synovial changes observed were modest, ASU-treated animals were
found to have a significantly greater degree of synovial fibrosis relative to placebo-treated animals.
This effect may again be consistent with its reported influence on TGF-ß expression. Experimental
studies indicate an important role for TGF-ß in promoting the fibrosis, lining cell hyperplasia, and
inflammatory cell infiltration that accompanies synovitis.16,446,563,687,877,878 When experimentally
injected directly into joints, TGF-ß causes fibrosis and increased synovial cellularity, due to

113.
fibroblast proliferation and monocyte recruitment.16 TGF-ß is thought to act synergistically with IL-
1 in inducing fibroblast proliferation.878 While it is immunosuppressive, suppressing T-cell and
macrophage function,446,805,806,878 TGF-ß is also markedly chemotactic, stimulating recruitment of
leucocytes.16,852,857 Intra-articular administration of anti-TGF-ß antibodies greatly reduces
inflammatory cell accumulation and tissue pathology in rat streptococcal cell wall-induced
arthritis.877 It is therefore conceivable that ASU may accentuate synovial fibrosis, if acting through a
TGF-ß-related mechanism as proposed by Boumediene et al. (1999).86 However, synovial
hyperplasia was found not to be a significant sequelae of meniscectomy in this trial.
Little clinical data was collected in this trial. However, body weight changes provide possible
clinical evidence for an improvement of animal well-being with ASU treatment. Decreased weight
gain of meniscectomised animals is likely to be a result of post-surgical stress and subclinical
lameness (affecting grazing behaviour, therefore lowering food intake). This effect appears to be
lessened in animals treated with ASU, though a direct effect of the drug on weight gain cannot be
ruled out from the data. It is also possible that ASU may confer analgesic properties independent of
any chondroprotective effects. Patient non-steroidal anti-inflammatory drug usage was evaluated as
the primary efficacy criteria in one placebo-controlled clinical trial,82 and was found to be
significantly reduced by ASU treatment. However, an increase in exercise has been shown to
exacerbate the cartilage degeneration seen in the similar ovine medial meniscectomy model.40,265
Any analgesic effect attributable to ASU in this trial would therefore be expected to have lessened
the beneficial effects seen on the progression of osteoarthritic lesions.
The results obtained confirmed the advantages of computerised, quantitative methodologies for
assessment of joint changes in arthritis. Scores obtained by gross visual assessment and a published
histopathological scoring method474,926 failed to demonstrate any significant effect of ASU
treatment. However, results obtained by computerised image analysis revealed subtle but
statistically significant effects of the drug, in maintaining articular cartilage proteoglycan content

114.
and thickness, and modifying subchondral sclerosis. Most studies of potential DMOADs in animal
models have used qualitative or semi-quantitative outcome measures, such as the size of cartilage
erosions or osteophytes.632 Such measures are unreliable and difficult to reproduce due to inter-
operator error. There are relatively few published reports applying computerised digital imaging
techniques to the quantification of cartilage damage. Chang et al. (1997)126 used video image
analysis to quantify Indian ink staining of lapine articular surfaces. Hacker et al. (1997)299 applied
an image analysis method to measure surface roughness and various dimensional parameters
associated with repair of osteochondral defects. Intra- and inter-observer precision was assessed and
was found to be high. Image analysis has also been used to determine loss of cartilage area and
surface roughness following ACLT in rabbits.750,927 Shimizu et al. (1997)750 assessed proteoglycan
content following safranin-O staining, as both the percentage area stained (using a density-slice
function) and the mean greyscale intensity of stained areas.
The method used here showed several advantages to those of previous reports. Firstly, the use of a
slide scanner provided a simple, low-cost alternative to complex digital imaging systems. By
adapting a slide cartridge to hold glass slides, a standard slide scanner is converted into a useful
densitometer, with a simple constant light source, and a charge-coupled device (CCD) detector free
of interposed apertures, lenses, or automatic gain controls. Secondly, toluidine blue staining was
quantified rather than safranin-O. A comparison of the two staining methods has shown toluidine
blue to be preferable for demonstrating loss of cartilage metachromasia.253 However, as the degree
of staining might be expected to be influenced by variables such as staining time, all sections need
to cut and stained using stringent protocols, to minimise this source of variability. Thirdly, mean
greyscale pixel intensity was used rather than a thresholding approach defining “stained area”.750
Greyscale intensity of stained regions (defined by density slice) as measured by Shimizu et al.
(1997)750 was found to be a less useful parameter, due to the non-parametric nature of the data
obtained (results not shown). Le Roux et al. (2000)455 similarly used a whole-area mean greyscale
approach to quantify safranin-O staining following canine ACLT. Fourthly, the use of more

115.
sophisticated image analysis software allowed easy determination of mean radial thickness
measures, which were found to be more sensitive subtle cartilage changes than cruder cross-
sectional area measures (data not shown). The use of thickness rather than area determinations also
removes the need to first measure and define a region of cartilage for analysis.
One complication in interpreting thickness and proteoglycan-staining parameters in this model is
the presence of simultaneous catabolic and anabolic responses in articular cartilage. Areas of
fibrillation and proteoglycan loss are bounded by regions of hypertrophic repair and increased
proteoglycan content. Similarly, cartilage thickness is increased (by over-hydration and swelling) at
the same time as focal areas are eroded. Previously reported image analysis methods have mostly
utilised the rabbit ACLT model.126,299,750 In this rapidly progressive model, severe cartilage erosion
and disruption predominate. In the present ovine meniscectomy model, the histopathological
changes induced are comparatively mild, and cartilage erosion is relatively focal. Therefore,
histomorphometric methods which quantify changes over the entire articular cartilage area may be
more sensitive to the surrounding compensatory and hypertrophic events, thus complicating
interpretation of results. The division of the cartilage area into inner, middle, and outer zones aided
assessment of focal events such as cartilage erosion.

116.
CHAPTER 3
EFFECTS OF GLYCERYL TRINITRATE, OVARIECTOMY, AND
MENISCECTOMY ON FEMORO-TIBIAL JOINT TISSUES OF AGED
EWES
3.1 INTRODUCTION
Glyceryl trinitrate (propane-1,2,3-triol trinitrate) (Figure 3.1) is a lipophilic organic nitrate which is
rapidly cleaved in vivo to inorganic nitrite, then to nitric oxide. As an efficient nitric oxide donor, it
has been used for many years as a vasodilator and symptomatic treatment for angina pectoris,
hypertension, and congestive heart failure. A variety of delivery systems are available, including
buccal tablets, oral sprays, topical ointments, slow-release discs, and intravenous preparations.2 The
pharmacokinetics of organic nitrates are complex as they are highly lipophilic and widely
distributed throughout the body. Extensive metabolism occurs within erythrocytes, vascular
endothelium and liver cells.2
Wimalawansa and co-workers have used GTN extensively to study the effects of nitric oxide donors
on bone metabolism in rats903,904,906,907 and humans.902 Preliminary studies compared various nitric
oxide donors (GTN, molsidomine, sodium nitrite, L-arginine) and found GTN to be the most
effective.901 The GTN dosage used in the following trial was extrapolated directly from these rat
studies, to give equivalent dosage on a mg per week basis. However, the frequency of
administration was lower, due to labour constraints. The pharmacokinetics of GTN in sheep have
not been studied. Plasma levels are very low (sub-nanogram) and difficult to assay due to the rapid
decomposition in plasma.60,160,725,920 Determination of plasma GTN levels following topical

117.
administration in sheep was therefore outside the scope of this study. However, a pilot study was
performed in four sheep, which showed a distinct peak in plasma nitrite levels 1-2 hours after
administration in three of the four sheep (data not shown), as has been shown after human usage.60
No side effects or behavioral changes were observed and the absence of methaemoglobinaemia was
confirmed by potassium ferricyanide assay213 (data not shown).
Figure 3.1: Structure of glyceryl trinitrate.
O
O
O
N N
N
O
O-
O O-
O
O-

118.
3.2 MATERIALS AND METHODS
3.2.1 TRIAL PROTOCOL
3.2.1.1 Source Of Animals
Aged Merino ewes (Peppin strain) were obtained from a farm near Cataby, Western Australia.
These animals were born in 1993 making them six and a half years old at the start of the trial. All
animals were bred on the property and subsequently managed together, and therefore represented
stock of genetically and environmentally homogenous background. Experimental subjects were
selected from a larger group by size, weight, and body condition criteria (50-60 kg, body score 1+
or above) and to exclude those animals which were obviously lame, unthrifty, or appeared never to
have lambed.
On introduction to the Murdoch University farm, the sheep were ear-tagged and vaccinated against
clostridial diseases and Corynebacterium pseudotuberculosis [Glanvac 6, CSL, Australia]. All
sheep were treated with an internal parasiticide [Cydectin Oral Drench for Sheep, Fort Dodge,
Australia; moxidectin 1 g/L], an external parasiticide [Vanquish Long Wool, Coopers Animal
Health, Australia; alpha-cypermethrin 50 g/L], and a preventive footrot preparation [Footrite,
Merial Australia, Australia; zinc sulphate 224 g/L]. In each sheep, both stifles joints were palpated
and medio-lateral radiographs obtained, to rule out pre-existing arthropathy. To ensure that they
were non-pregnant and actively cycling, all non-ovariectomised ewes received 125 g of
cloprostenol [Juramate, Jurox, Australia] by intramuscular injection. A vasectomised (teaser) ram
with a raddle harness was used to detect ewes coming into oestrus; the few which did not were
excluded from the trial.

119.
3.2.1.2 Surgery
3.2.1.2.1 Ovariectomy
Twenty-four sheep were ovariectomised by midline laparotomy. After deprivation of food and
water overnight, the sheep were anaesthetised with intravenous thiopentone sodium (50mg/ml)
[Jurox, Australia]. Anaesthesia was maintained with inhaled halothane [Rhône Mérieux, Australia]
using a Fluotec 3 vaporiser [Cyprane Keighley, UK] and a circle anaesthetic circuit. Ovariectomy
was performed, under full sterile conditions, via a 5cm midline laparotomy incision placed
approximately 5cm forward of the cranial border of the mammary gland. The ovaries were
exteriorised from the body cavity, the ovarian pedicle was clamped and ligated, and all ovarian
tissue was removed. The excised tissue was visually inspected to confirm the entire ovary had been
removed. All sheep showed signs of recent ovarian activity (enlarging follicles or corpora lutea).
One sheep was found to be in early pregnancy; in this ewe, full ovariohysterectomy was performed
via an enlarged midline incision. The linea alba incision was closed using 3 metric polydioxanone
suture [PDS II, Ethicon, USA], the subcutaneous tissues were apposed using 3 metric
poliglecaprone 25 suture [Monocryl, Ethicon, USA], and the skin sutured with 3 metric
polypropylene [Prolene, Ethicon, USA] in a simple interupted pattern. To provide analgesia, all
ewes received 100mg of flunixin meglumine [Ilium Flunixin, Ilium, Australia] by intramuscular
injection postoperatively. Cutaneous sutures were removed 9 days after surgery.
3.2.1.2.2 Meniscectomy
Bilateral lateral meniscectomy was performed on 24 sheep. Twelve of these sheep had been
ovariectomised 5 weeks previously, as described in Chapter 2. As in the previous trial,
complications and visible lameness were minimal. One ewe showed extensor weakness suggestive
of peroneal nerve damage, which resolved spontaneously within one week.

120.
3.2.1.3 Glyceryl Trinitrate Treatment
Animals assigned to GTN-treated groups were identified by coloured ear tags. Treatment was
performed twice per week (i.e. 3-4 day intervals). Calculated doses (0.7 mg/kg) of 2% glyceryl
trinitrate ointment [Nitro-Bid®, Hoechst Marion Roussel, Australia] (Figure 3.2) were individually
dispensed by weight onto waxed paper squares immediately prior to treatment. At scheduled
treatment times, the entire mob was brought into a sheep shed and animals to be treated were
drafted into a race. With the sheep standing in the race, the ointment was applied with gloved hands
to the hairless region of skin on the medial aspect of the upper thigh, an area of approximately 250
cm2 (Figure 3.4), and gently rubbed until the ointment was absorbed. No behavioural abnormalities
or other signs were observed to indicate any side-effects associated with GTN treatment.
3.2.1.4 Animal Maintenance And Sampling
The sheep were maintained as a single mob, and rotated between 1-2 hectare paddocks on the
University farm. Pastures were irrigated and consisted mainly of kikuyu, perennial white and
strawberry clover (minimally oestrogenic varieties), and perennial ryegrass. Sheep were weighed at
approximately monthly intervals. Maintenance was limited to regular foot trimming and
endoparasiticidal treatment [Closantel (closantel 37.5 g/L), West Stock Distributors, Australia;
Ivomec Liquid for Sheep (ivermectin 0.8 g/L), Merial, Australia]. Toe or sole abscesses were an
occasional problem and were treated by corrective trimming, intramuscular injection of procaine
penicillin / streptomycin [Ilium, Australia], and short-term confinement in a dry shed if necessary.
Faecal eggs counts were tested periodically to monitor endoparasite burdens.
Urine was collected at various time points, either during anaesthesia, by free catch following
induction of urination by nasal occlusion, or at the time of sacrifice. Animals were sacrificed in a

121.
staggered, blocking design over a period of four months. Two GTN-treated animals were sacrified
simultaneously with two sheep from the relevant control group. Periods of post-meniscectomy,
post-menisctomy, and of GTN treatment are indicated in Table 3.1.
Table 3.1: Treatment intervals (weeks) at time of sacrifice, by treatment group.
Group Weeks post-ovariectomy Weeks post-meniscectomy Duration of GTN treatment
NOC - - 26-28 OVX 28-30 - 26-28 MX - 27-29 25-27 OVX+MX 32-34 27-29 23-25
To label mineralizing bone, all animals received intravenous injections of calcein (15 mg/kg)
thirteen days before sacrifice, and xylenol orange (15 mg/kg) three days before sacrifice.729 Calcein
[Sigma, USA] was prepared as a 40 mg/ml solution in water for injection. The solution was
adjusted to pH 7.4 with sodium hydroxide, filtered through a 0.2 m syringe filter, and injected
slowly intravenously via a 19 gauge butterfly catheter in the lateral cephalic vein. Xylenol orange
[ICN Biomedicals, USA] was prepared as a 40 mg/ml solution, adjusted to pH 7.4 with
hydrochloric acid, and administered similarly.
3.2.1.5 Sacrifice And Tissue Processing
At time of sacrifice, jugular blood samples were collected from each sheep before euthanasia by
intravenous injection of 20ml of pentobarbitone sodium (300 mg/ml) [Valabarb, Pitman-Moore,
Australia]. Serum for osteocalcin assay was separated within 2 hours, and frozen immmediately at -
80°C. Urine was collected and both hindlimbs were skinned and removed intact. The first lumbar
vertebrae was identified and removed for storage in 70% ethanol. One randomly selected leg (left or
right) was sealed in plastic bags, stored in crushed ice, and transported overnight by air to the
Raymond Purves Bone and Joint Laboratory (Institute of Bone and Joint Research, University of

122.
Sydney at the Royal North Shore Hospital, St. Leonards, NSW). This limb was used for
biomechanical testing, intact bone densitometry, and tibial plateau decalcified histology. The
contralateral limb remained at Murdoch University for tissue culture procedures. After tissue had
been harvested for culture, a band-saw was used to remove osteochondral slabs for histologic and
biochemical assessment. Two 2-3 mm thick, mid-transverse sections were taken : (i) one across the
femoral condyles, to include the point of tibial contact in usual stance, placed into formalin (for
decalcified histology); and (ii) one across the tibial plateau, at the level of the intercondylar
eminences, placed into 70% ethanol (for cubic bone density assessment and undecalcified bone
histology).
3.2.2 GROSS PATHOLOGY
Gross pathological assessment was recorded as the joint was opened prior to tissue culture, using
the scale detailed previously (see 2.2.2).
3.2.3 DYNAMIC BIOMECHANICAL TESTING
The biomechanical properties of tibial articular cartilage were assessed by Dr. Richard Appleyard
(Orthopaedic Research Institute, University of New South Wales at the St. George Hospital, NSW),
using a recently developed hand-held indentation device.35 Briefly, the device incorporated a handle
and a 150mm long stainless steel tube (5mm external diameter) (Figure 3.5). Located at one end of
the tube was a vibration unit consisting of a flexing beam, held in a cantilevered configuration, with
strain gauges attached. A small cylindrical probe (0.5mm diameter) was attached to the free end of
the beam and extended out the side of the side of the tube through a narrow slot. A long shielded
wire connected the indenter to a strain gauge amplifier unit that was connected to a computer, via an
A-to-D card. A single frequency waveform (20Hz), applied to the instrument, caused the probe to
vibrate in an up-and-down motion with respect to the tube opening. The amplitude of oscillation

123.
was set at ±0.03mm. Both the supply and response (strain gauge) signals were monitored
simultaneously, and these time domain signals converted into the frequency domain using Fast
Fourier Transform (FFT) techniques.
Biomechanical testing was performed on intact tibial plateaux, which had been frozen at –20°C
awaiting analysis. Freeze-thawing of cartilage has been shown to have no effect on biomechanical
properties.47 On the day of testing, the samples were immersed in fresh phosphate-buffered saline
(PBS) and equilibrated to 20°C for 45 minutes. Using a grid projected onto the articular surface by
a modified slide projector, a 3 x 3 array was marked on the cartilage surface of the lateral and
medial tibial plateaux with Picro Sirius red stain (Figure 3.6). At each of these locations (n=18), the
indentation probe was pressed against the surface with moderate pressure. Dynamic stiffness and
phase lag were continuously displayed on a computer monitor. Pressing a button on the handle of
the indentation device instructed collection of data. On completion of biomechanical testing,
articular cartilage thickness was determined at each location using a needle penetration method.807
Accuracy and precision trials using the indenter on standard rubbers and normal ovine tibial
plateaux have demonstrated excellent correlations.35
The dynamic shear modulus, G*, was calculated using the theory published by Hayes et al.
(1972),331 where:
G* = P * (1- )
H * 4a
and a is the indenter radius, P*/H* is the dynamic force divided by the dynamic displacement
(dynamic stiffness), s the Poisson’s ratio and is the theoretical correction function. Hayes et al.
(1972) tabulated the variation in with alteration in Poisson’s ratio and aspect ratio, a/h, where h is
the thickness of articular cartilage beneath the indenter.331

124.
3.2.4 HISTOLOGICAL ASSESSMENTs
3.2.4.1 Cartilage Histology
Specimens were processed for decalcified histology as described in Chapter 2. Computerised image
analysis was completed as described previously, with the following exceptions: (i) femoral condylar
sections were cut at 5 m instead of 4 m, (ii) quantification of toluidine blue staining (mean
greyscale pixel intensity) was performed only in the femoral condyles, as freezing may have caused
artifactual loss of proteoglycan in the tibial plateaux; and (iii) greyscale pixel intensity was
measured by the image analysis software NIH Image v.1.62 [National Institutes of Health, USA]
instead of Image Pro. In addition, cross-sectional osteophyte area (mm2) was determined in each
region in meniscectomised sheep. Calcified cartilage pathology was evaluated according on a scale
of 0-4 (below), at each of the four zones scored for histopathology. Calcified cartilage vascularity
was measured as the number of blood vessels invading the length of the calcified cartilage layer
(Figure 3.9), excluding marginal regions of osteophytic remodelling.367 Chondrocyte number was
also quantified in the middle zone of each femoral condyle, by counting the number of distinct
nuclei in a 250x250 m grid using an eyepiece graticule positioned midway between the surface and
tidemark layers, at three adjacent locations.
CALCIFIED CARTILAGE SCORE
0: Intact subchondral bone plate and single tidemark 1: Intact subchondral bone plate with multilayered tidemark 2: Blood vessels penetrating into calcified cartilage layer 3: Blood vessels penetrating through calcified cartilage layer to tidemark 4: Tidemark penetrated by blood vessels

125.
3.2.4.2 Collagen Birefringence
Quantitative analysis of collagen birefringence was performed in 4 m sections from the tibial
plateaux (intact osteochondral sections) and the LFC (full-thickness cartilage plug) of non-
meniscectomised sheep only. Sections were treated with bovine testicular hyaluronidase [ICN
Biomedicals, Australia] to remove proteoglycans, then stained with Picro Sirius red using a
modification of the published method of Junqueira et al. (1979).396 Birefringence intensity was
determined using methods similar to those of Király et al. (1997)421 and Arokoski et al. (1996).45
Stained sections were viewed through a Leica DMLB polarisation microscope [Leica, Germany]
under monochromatic ( 550nm) light supplied by an interference filter [OG 550nm, Schott,
Germany]. With the cartilage surface oriented at 45° to the lower polarisation filter, greyscale
digital images were collected using a digital camera. A full-thickness birefringence intensity profile
was generated using image analysis software [Image Pro Plus v3.0.1, Media Cybernetics, USA].
The profile plot was then normalised by depth (0= cartilage surface, 1= calcified cartilage) to allow
comparative analysis of cartilage of different thickness (Figure 3.7).
3.2.4.3 Synovial Histopathology
Synovial pathology was scored subjectively, as detailed in Chapter 2 (see 2.2.5). In addition,
cellularity was assessed. Using an eyepiece graticule [Olympus, Japan] defining a 250 m-square
grid (50 m divisions) at x200 magnification, positioned over relatively flat segments of synovia,
the number of cell nuclei in a 250 m long strip of intima (to a depth of 50 m) was counted at five
randomly selected points of each specimen.

126.
3.2.4.4 Undecalcified Bone Sections
Lateral tibial plateau bone slabs (stored in 70% ethanol) were dehydrated by sequential exposure to
90% and 100% ethanol. Slabs were then exposed to two changes of methyl methacrylate (MMA)
monomer [Fluka Chemike, Switzerland] prior to embedding. Monomer was washed prior to use to
remove hydroquinone inhibitors of spontaneous polymerisation, by washing three times with an
equal volume of 5% w/v NaOH and three times with ultrapure water. Monomer was then dewatered
by overnight addition of granular anhydrous CaCl2 followed by filtration. Bone specimens were
then embedded in MMA monomer activated by the addition of 30g/L benzoyl peroxide [Sigma,
USA], in small capped gass jars. The jars were placed in an evacuated dessicator within a 37°C
oven for 4-5 days. When polymerisation was complete, the jars were broken and the blocks were
trimmed to a small field (approximately 10x10mm, in the central zone of LTP) using a hacksaw and
a low-speed diamond saw [Buehler Isomet, USA].
Undecalcified sections (5 m) were cut on a Model 1140 Reichert-Jung Autocut microtome using a
“D”-profile tungsten carbide microtome knife [Austral Scientific, Australia]. Sections were floated
onto an 80°C water bath and collected onto silanized glass slides, which were then clamped
between sheets of plastic and baked at 60°C overnight. Sections were stained with solochrome
cyanin to differentiate osteoid (Figure 3.11).607 Briefly, slides were stained with solochrome cyanin
(1% w/v solochrome cyanin [BDH Chemicals, UK], 2% v/v glacial acetic acid, 0.4% ammonium
ferric sulphate, in ultrapure water) for 20 minutes, rinsed in warm water for 2-4 minutes until bone
appeared blue, dehydrated through increasing alcohol concentrations, cleared in xylene and
mounted with DPX.

127.
Surface parameters were calculated in cancellous bone by manual point-counting, using a Weibel II
eyepiece graticule with 21 parallel bars [Graticules Ltd, UK]. Where a bar crossed from marrow
space to bone, the surface was recorded as quiescent surface, osteoid surface (stained red-pink), or
eroded surface (Howship’s lacunae present). Three to five hundred surfaces were counted in each
section. Surface parameters were calculated as a percentage using total surface as the referent in all
cases. Trabecular volume was calculated by image analysis of scanned sections [NIH Image v.1.62;
National Institutes of Health, USA], and calculated as a percentage using total volume as the
referent.
Unstained undecalcified sections were examined using a fluorescent microscope [Olympus Vanox;
Olympus Optical Co., Japan] with blue filters (BG-12 exciter filter, DM-500 and O-515 dichroic
mirror, O-515 barrier filter). Using the above point-counting technique, surfaces were counted as
either labelled or unlabelled according to the presence of a distinct line of calcein fluorescence
(Figure 3.12). Xylenol orange labelling was faint and indistinct, so was excluded from analyses.
The reasons for this are unknown, though xylenol orange has been shown to fade with prolonged
storage or exposure to solvents.660 Labelled surface was calculated as a percentage of total surface.
The small sample and density of the cortical bone prevented accurate calculation of surface-
labelling parameters in the subchondral bone plate. Instead, calcein labelling was scored
qualitatively according to the following scale:
SUBCHONDRAL BONE PLATE CALCEIN SCORE
0: No calcein-labelled osteons 1: Few calcein-labelled osteons (<25%) 2: Multiple calcein-labelled osteons (25-50%) 3: Many calcein-labelled osteons (50-75%) 4: Most osteons calcein-labelled (>75%)

128.
3.2.5 BONE DENSITOMETRY
Bone mineral density of the dissected first lumbar vertebra was determined by dual-energy X-ray
absorptiometry (DEXA) using a Hologic QDR 1000-W densitometer [Hologic Inc., USA].
Vertebrae were anatomically positioned on a perspex support and immersed in a 10cm water bath.
The region of interest (ROI) was defined manually to include only the vertebral body itself.
BMD was determined in the LTP, MTP, LFC, MFC, femoral head and neck, and femoral and tibial
diaphyses using a Hologic QDR 4500-W absorptiometer [Hologic Inc., USA]. A bed of rice of
constant thickness (5cm) was used to both position the specimen and to simulate soft-tissue density.
Scanning of subchondral sites was conducted such that the weight-bearing articular surface was
parallel to the beam. Bone density (gm/cm2) was determined as the mean of three 5x3 mm ROI, in
the inner, middle and outer subchondral zones of the tibial plateau, and three 3x3 mm ROI in the
corresponding zones of the femoral condyles.368 The dissected femoral head and tibial and femoral
shafts were scanned in normal anatomical position (anteroposterior). Five ROI were analysed in the
proximal femur: femoral neck (23x32 mm), Ward's triangle (10x10mm), two 5x5mm ROI in the
subchondral weight-bearing region, and the fovea capitus (5x5cm). In the tibia and femur, ROI
included approximately 3cm of the entire shaft excised from the mid-diaphyseal region.
BMD was also determined in tibial bone slabs prior to undecalcified histology, using a Hologic
QDR 4500-W absorptiometer [Hologic Inc., USA] (Figure 3.3). The slabs were placed flat on a
perspex plate overlaying a bed of rice (5cm). ROI were identifed as for the intact condyles. The
thickness of the slab was determined using a digital caliper, and the bone density reported in
gm/cm3 as described by Armstrong (1993).39

129.
3.2.6 BLOOD AND URINE ASSAYS
3.2.6.1 Plasma 17ß-Oestradiol
Plasma 17ß-oestradiol concentrations were measured using the method of Webb et al. (1985),887
involving an affinity chromatography extraction procedure followed by double-antibody
radioimmunoassay (RIA). All antibodies used were those described by Webb et al. (1985),887 kindly
provided by Prof. Graeme Martin (Dept. of Animal Science, University of Western Australia). A
known volume of serum (2-5 ml) was diluted to 10 ml with ultrapure water and gently mixed for 2
hours at room temperature with antibody-sepharose 4B beads (sheep anti-oestradiol-17ß-6-
carboxymethil-oxime serum). Beads were recovered by passing mixture through baked glass
columns containing sintered glass filters (porosity 1) and rinsing twice with water. Bound steroid
was eluted with 3ml of 90% methanol and evaporated to dryness. A standard preparation of 2,4,6,7-
3H-17ß-oestradiol [Amersham Pharmacia Biotech, UK] in sheep plasma was included in each batch
to evaluate extraction efficiency. Samples were then resuspended in gelatin-phosphate buffer (0.1%
gelatine in PBS) and dispensed in duplicate into 12 x 75mm glass tubes. Aliquots of radioiodinated
steroid (17ß-oestradiol-11 -tyrosinemethyester) and first antibody (rabbit anti-17ß-oestradiol-11ß-
succinyl-BSA serum, 1:240,000 dilution) were added prior to incubation for 4 hours at room
temperature. 100 l normal rabbit serum, diluted in gelatin-phosphate buffer containing 0.1M
ethylenediaminetetraacetic acid (EDTA), and 100 l of second antibody (donkey anti-rabbit serum)
were then added as precipitating agents. Following a 36 hour incubation at 4°C, 1.5ml of ice-cold
polyethylene glycol (2% in PBS) was added and the tubes centrifuged at 3000 rpm for 25 minutes.
The supernatant was immediately aspirated and the radioactivity in the precipitates counted by
routine gamma counting. Concentrations of oestradiol were computed by ‘AssayZap Universal
Assay Calculator’ [Elsevier-Biosoft, Cambridge, UK] against a standard curve (0-100 pg/ml)
assayed simultaneously, and corrected for extraction efficiency (65.4 ± 3.2 %) and original plasma
volume.

130.
3.2.6.2 Serum Osteocalcin
Osteocalcin was assayed by Graeme Worth at Sir Charles Gairdner Hospital, Perth, Australia using
a competitive radioimmunoassay. Briefly, 125I-bovine osteocalcin tracer, rabbit anti-bovine
osteocalcin serum, and normal rabbit serum were combined with the test sample or bovine
osteocalcin standard and incubated for 48 hours at 4°C. Antibody complexes were precipitated by
addition of an anti-rabbit gamma globulin precipitating sera and centrifugation. Precipitates were
washed once, and counted by routine gamma counting. Antisera raised against bovine osteocalcin
have been shown to cross-react with both human and ovine osteocalcin.157
3.2.6.4 Urine Collagen Crosslinks
Pyridinoline and deoxypyridinoline assays were kindly performed by Nick Avery (Collagen
Research Group, University of Bristol) on air-freighted frozen urine samples. Collagen crosslinks
have been shown to be extremely stable in frozen and freeze-thawed samples.251 Collagen crosslinks
were detected using a published high-performance liquid chromatography (HPLC) method.761
Briefly, urine was mixed with 6N hydrochloric acid and to 115°C for 24 hours. Hydrolysates were
then freeze-dried and pre-chromatographed on a Whatman CF-1 cellulose powder column.
Following vacuum concentration and resuspension in 1% trifluoroacetic acid, HPLC was performed
using a 4.6 x 100mmm Shandon Hypercarb-S graphitic carbon column. With a flow rate of
1ml/minute and 0.25%/ml, crosslink peaks were resolved using a linear gradient from 0-15%
tetrahydrofuran in water, both containing 0.5% trifluoroacetic acid. Resulting peaks were quantified
by comparison to known standards prepared in the laboratory.
Urine collagen crosslink content was corrected for urine creatinine concentration. Creatinine was
assayed in urine diluted 1:100 in ultrapure water, using a modified Jaffe reaction.333 50 l of dilute

131.
urine, 0.5ml 0.04M picrate, 0.5ml 0.75M NaOH, and 950 l H20 were combined in a disposable
cuvette. The absorbance at 500nm was determined on a spectrophotometer [Beckamn DU-650;
Beckman, USA] and compared to standard solutions (0-20 g/ml) of creatinine.
3.2.7 CELL CULTURE PROCEDURES AND ASSAYS ON CELL CULTURE
CONDITIONED MEDIA (CM)
After skinning and removal of the hindlimb, the distal crus and excess muscle tissue were removed
and the joint sprayed with 70% ethanol. The joint was then transferred to a laminar flow cabinet,
after which point it was handled only with full sterile procedure. After further trimming of all tissue
overlying the joint capsule, the joint was carefully dissarticulated and synovial, cartilage, and bone
tissue removed to labelled petri dishes containing sterile PBS [Trace Scientific, Australia] for
subsequent tissue culture.
All cell culture and bioassay procedures used tissue culture-treated plastic multiwells manufactured
by TPP [Techno Plastic Products, Switzerland], unless otherwise specified. Water for all assay and
culture procedures was purified by ultrafiltration [MilliQ Ultrapure Water System, Millipore
Australia]. All cell culture incubation was conducted in a 37°C humidified 5% CO2 incubator
[Forma Scientific, USA]. Foetal calf serum (FCS) was obtained from Trace [Trace Scientific,
Australia]. Ovine recombinant IL-1ß743 was kindly supplied by the Centre for Animal
Biotechnology, University of Melbourne, Australia. [5'-3H] thymidine (1mCi/ml) was obtained
from Amersham Pharmacia Biotech, UK. All ß-counting was conducted using a Beckman LS 3801
Liquid Scintillation counter [Beckman, USA] after addition of 3ml of scintillation fluid [EcoLite
(+), ICN Biomedicals, USA] per scintillation tube.

132.
3.2.7.1 Patellar Cartilage Explant Culture
Four cartilage explant discs were removed from the proximal articular surface of the patella using a
disposable 3mm biopsy punch [Stiefel, Germany]. After rinsing in sterile PBS, the explants were
transferred to individual wells of 24-well multiwell culture plates. Explants were cultured in 500 l
of Hams F12 medium [Sigma, USA] supplemented with 1mM L-glutamine [Sigma, USA], 50 g/ml
gentamicin [David Bull Laboratories, Australia], and 10% v/v foetal calf serum, pH 7.2. After 24
hours of culture, media was replaced by 500 l of fresh Hams F12/10% FCS supplemented with
4 Ci/ml Na35SO4 [Amersham Pharmacia Biotech, UK]. 100pg/ml ovine recombinant IL-1ß was
added to two of the wells. After a further 48 hours of incubation, the explants (patted dry) and
aliquots of conditioned media were immediately frozen at -20°C. Media for PGE2 assay (NOC and
MX sheep only) was stored at -80°C.
3.2.7.2 Subchondral Bone Explant Culture
Explant culture of subchondral bone was performed using the method of Hilal et al. (1998).350 Two
subchondral bone explants were removed from the anteromedial portion of the lateral tibial plateau
using a 5mm Michele bone biopsy trephine. After excision of attached cartilage and cancellous
bone using a No. 11 scalpel blade, the explants were rinsed thoroughly in PBS and transferred to
individual wells of a 24-well culture plate. 1ml of serum-free BGJb medium (Fitton-Jackson
modification) [Sigma, USA] supplemented with 50 g/ml gentamicin was added to each well. After
incubation for 95 hours, aliquots of conditioned media were frozen at -20°C. The explants were
patted dry with tissue and weighed on a 0.00001g balance [Sartorius, Germany]; subsequent assay
results are expressed as activity per mg wet weight of tissue.

133.
3.2.7.3 Synovial Fibroblast Culture
Synovial lining was harvested by sharp dissection with a No. 11 scalpel blade, from all regions of
the joint. Care was taken to exclude excessive subsynovial or fibrous tissue. The synovium was
placed into PBS, diced finely using two No. 11 scalpel blades in a scissoring action, and transferred
to a sterile plastic centrifuge tube. The tissue was washed by two cycles involving the addition of
excess PBS, centrifugation (10 mins, 2000 rpm, 20° C), and careful removal of excess PBS between
the floating and sediment layers using a plastic transfer pipette. An equal volume of 0.2% trypsin
solution [Gibco BRL, USA] in 0.1% EDTA was added to the tubes, which were then stored at 4°C
overnight. Leaving synovium at 4°C in normal saline overnight has been shown to have no apparent
effect on subsequent culture or viability.530 After incubation for 1 hour at 36°C, 1ml of FCS was
added, and the tubes centrifuged as before. Excess trypsin solution was removed to leave a volume
of 10ml, before addition of 40ml of Dulbecco's Modified Eagles Medium [Trace Scientific,
Australia] supplemented with 44mM NaHCO3, 15mM N-[2-hydroxyethyl]piperazine-N'-[2-
ethanesulfonic acid] (HEPES) [Sigma, USA], and 10% FCS, pH 7.2 (DMEM/10% FCS). After
centrifuging as before, excess solution was removed and an equal volume of 2 mg/ml collagenase
solution [Clostridial Type IA; Sigma, USA] in DMEM/10% FCS was added before incubation for 3
hours at 36°C. After centrifuging as before, the supernatant (including digested fat layer) was
removed to waste. The sediment was washed twice by gentle resuspension in fresh media,
centrifugation as before, and removal of supernatant. The sedimented cells were thoroughly
resuspended in 15ml of media, and filtered through a single layer of autoclaved sheer polyester
voile (mesh size 25 x 25 m) before mixing an aliquot with an equal volume of 0.4% trypan blue
[Sigma, USA] and counting cell yield and viability with an improved Neubauer haemocytometer
[Weber Scientific International Ltd., England]. Viability was always >95%. After removing aliquots
for primary cell culture, the remaining cell suspension was transferred to two 75cm2 canted neck
cell culture flasks [Corning Inc., USA] and cultured in DMEM/10% FCS containing 10 ml/L of an

134.
antibiotic / antimycotic mixture (10 000U penicillin, 10mg streptomycin, 25 g amphotericin B per
ml) [Sigma, USA].
Primary isolates were cultured for 8 days, with changes of media at days 3 and 6. During this
period, cultures proliferated to confluence, and changed from a sparse pavement-like appearance to
dense whorls of fibroblastic spindle-shaped cells (Figure 3.10). To subculture, media was removed
and the monolayer rinsed twice with sterile PBS. 1-2 ml of 0.2% trypsin in 0.1% EDTA was added
to each flask before incubation for 3-5 minutes. The loosened cell sheet was transferred to 15ml
centrifuge tubes and washed twice by suspension in excess DMEM/10% FCS, centrifugation (10
min, 2000rpm, 20°C), and removal of the supernatant to waste. After thorough resuspension in fresh
media, cell concentration was counted using a haemocytometer as before. Cells not needed for
further subculture were sedimented by centrifugation and resuspended in 200 l TEDG buffer
(10mM Tris HCl, 1.5mM EDTA, 2mM dithiothreitol, 10% v/v glycerol, pH 7.5) and frozen at -
20°C awaiting oestrogen receptor assay.
Cells from each sheep were plated into four wells of two tissue culture-treated 24-well culture
plates at an initial cell density of 2x105 cells/well. After 4 hours, media and any unattached cells
were removed and replaced with 500 l fresh DMEM/10% FCS. Two of the wells were
supplemented with 100 pg/ml ovine recombinant IL-1ß. After incubation for 48 hours, aliquots of
conditioned media were immediately frozen at -20°C pending biochemical assays (-80°C for PGE2
determinations). The remaining cell monolayer was drained and frozen at -20°C awaiting assay of
DNA content.
Passaged cells from each sheep were also seeded into pentiplicate wells of tissue culture-treated 96-
well culture plates [Falcon, USA] at a density of 1x104 cells/well (200 l/well) for separate
proliferation assay. Media was replaced after 4 hours to remove any unattached cells. After 44
hours of incubation, 50 l of 3H-thymidine solution (5 Ci/ml in DMEM/10%FCS) was added to

135.
each well. After a further 4 hour incubation, media was removed, and the cell monolayers were
rinsed twice with PBS. 100 l of 0.2% trypsin/0.1% EDTA was added before incubation at 37°C for
20 minutes. Using a cell harvester [PHD Cell Harvester, Cambridge Technology, USA], cells were
harvested onto glass filter paper discs [Whatman GF/C glass microfibre filters, Whatman
International, England] which were placed in scintillation vials and counted by routine ß-counting.
Primary isolates were also cultured for cytokine determinations. After filtering the digestate and
counting the resulting single cell suspension, cells were plated in quadruplicate into 24-well
multiwell culture plates at 5x105 cells/well. The media was carefully changed after 24 hours and
replaced with 500 l fresh DMEM/10% FCS with the addition of 1 g/ml lipopolysaccharide
[Escherichia coli Serotype O55:B5; Sigma, USA] to two of the wells. After a further 48 hours of
incubation, aliquots of conditioned media and the drained culture plates were immediately frozen at
-20°C. Careful removal of media was found not to disturb the cell layer, as the cells rapidly secreted
and became embedded in a loose glycosaminoglycan matrix.
3.2.7.4 DNA Assay
DNA was assayed using the fluorescent DNA binding agent Hoescht 33258 (bis-benzimide).417,442
Synovial culture wells were carefully emptied of all conditioned media. 500 l of papain solution
was added to each well (approximately 0.8 U/ml papain [Sigma, USA; from papaya latex] in PBS
with 0.005M EDTA and 0.01M cysteine added, pH 6.0) before incubation for 2 hours at 60°C.
Cartilage explants were papain digested in 650 l papain solution for 4 hours at 60°C. 200 l of the
digestate was combined with 300 l dye solution in a 10mm borex test tube (2 g/ml Hoescht 33258
[Sigma, USA] in 2M NaCl, 0.05M Na3PO4, pH 7.4) and mixed well. After standing for 5 minutes,
1ml of buffer was added and the fluorescence read on a Perkin Elmer LS-50 luminescence
spectrometer ( ex 350nm, em 450nm). DNA content ( g/well) was calculated versus a standard
curve of calf thymus DNA [Sigma, USA] assayed in parallel. As all synovial fibroblast monolayers

136.
were visibly confluent at the end of the culture period, the DNA content of each well is proportional
to confluent cell density ( g per 24-well microplate well).
3.2.7.5 Nitric Oxide
Nitric oxide release was assayed colorimetrically as the nitrite content of conditioned media using
the Greiss reaction.286,297 100 l of CM was combined in a disposable cuvette with 500 l of 1%
sulfanilamide [ICN Biomedicals, USA] in 5% H3PO4, and 500 l of 0.1% N-(1-
naphthyl)ethylenediamine dihydrochloride [ICN Biomedicals, USA]. After 5 minutes, the
absorbance at 540nm was measured on a spectrophotometer [Beckamn DU-650; Beckman, USA].
The appropriate media blank value (determined using 5% H3PO4 without sulfanilamide) was
subtracted from all absorbances. The nitrite content was detemined against a standard curve of 0-
50 M NaNO3 [Sigma, USA] assayed in parallel.
3.2.7.6 MMP Activity
MMP activity was assayed in CM from serum-free cultures. Both auto-activated and total (p-
aminophenylmercuric acetate (APMA)-activated) activities were determined using the fluorogenic
substrate [(7-methoxycoumarin-4-yl)acetyl](Mca)-Pro-Leu-Gly-Leu-[N-3-(2,4-dinitrophenyl)-L-
2,3-diaminopropionyl-](Dpa)-Ala-Arg-NH2 [Novabiochem, Switzerland, #03-32-5032].425 This
extremely sensitive substrate is cleaved primarily by gelatinases but also by (amongst others)
MMP-1, -3, and -7 and is suitable for assay of total MMP activity in crude preparations.66,425
The fluorescent substrate was stored frozen in dimethyl sulfoxide (DMSO) (1mg/ml) and further
diluted to 40 M in TC buffer (50mM Tris-HCl, 5mM CaCl2, pH 7.5) before use. 20 l of diluted
substrate was combined with an equal volume of CM and incubated for 2 hours (cartilage explant
CM) or 4 hours (synovial fibroblast CM) in a 37°C water bath. The reaction was terminated with

137.
1ml of cold 40mM EDTA and the fluorescence determined on a Perkin Elmer LS-50 luminescence
spectrometer ( ex 328nm, em 393nm). A blank consisting of substrate only was subtracted from all
results and the activity reported as F/min/ g DNA where F equals the change in fluorescence
units over the incubation period.
Total (APMA-activated) activity was determined after incubation (3hr, 37°C) of 50 l CM with 10 l
2.5mM APMA. APMA solution was prepared by 1:4 dilution of 10mM stock (3.5mg/ml in 0.1M
NaOH) in TTC buffer (50mM Tris-HCl, 5mM CaCl2, 0.05% (v/v) Triton X-100) and readjusted to
pH 7.5. 20 l of activated CM was assayed as above after 1 hr incubation.
3.2.7.7
35SO4-Labelled Proteoglycan Synthesis
35SO4-labelled proteoglycans were isolated by precipitation of free 35SO4 as insoluble BaS04.151
400 l of cartilage explant digestate was combined in a 12 x 75 mm borosilicate test tube with 200 l
of a solution containing equal volumes of 0.2M Na2SO4 and 50mg/ml chondroitin sulphate A (CSA)
[Sigma, USA; from bovine trachea], then vortexed. 100 l of 0.4M BaCl2 was added before
vortexing again. Tubes were then centrifuged in a Beckman J-6B refrigerated centrifuge at 1000 g
for 10 minutes. 400 l of the supernatant was transferred to a clean test tube without disturbing the
precipitate. 100 l of a 2:5 mixture of 50mg/ml CSA and 0.4M BaCl2 was added and the tubes
vortexed, before addition of 100 l 0.2M Na2SO4, vortexing, and centrifuging as before. 200 l
aliquots of the supernatant were carefully transferred to scintillation vials and the radioactivity
counted on a Beckman LS 3801 Liquid Scintillation counter [Beckman, USA]. Results are
expressed as DPM per g DNA in the explant.
35SO4-proteoglycan content was also determined in cartilage explant conditioned medium. 100 l of
CM was added to 400 l papain solution and digested as before (2 hours, 60°C). 400 l of the
digestate was assayed as above. Total 35SO4-proteoglycan synthesis was calculated as the total DPM

138.
of the explant plus the total DPM of the CM. Percentage release of newly synthesised proteoglycan
was determined as the DPM of CM x 100 / total DPM of CM + explant.
3.2.7.8 PGE2
Conditioned media samples for PGE2 assay were stored at -80°C for a maximum of six months
prior to assay. Assay was performed using a commercially available enzyme immunoassay (EIA)
[Cayman Chemical Company, USA] according to the manufacturers instructions. The 2B5
monoclonal antibody in this kit is highly specific for PGE2, though some cross-reactivity with PGE1
and PGE3 is documented in the product specifications. Briefly, CM samples were diluted 1:10 in
EIA buffer (1:100 dilution for IL-1ß-stimulated cartilage CM). 50 l of the diluted samples were
added to individual wells of a 96-well plate pre-coated with goat anti-mouse antibody. 50 l of
PGE2-acetylcholinesterase tracer and 50 l of murine PGE2 monoclonal antibody were added prior
to incubation for 18 hrs at 4°C. The plate was then rinsed five times with the supplied wash buffer
before addition of Ellman's reagent. After incubation for a further 150 minutes, the absorbance at
405nm was determined on a microplate spectrophotometer [Biorad Model 450 Microplate reader;
Biorad, USA]. After subtraction of non-specific binding values, the %B/B0 (% bound/maximum
binding) was determined for each sample and compared to a standard curve of 7.8 - 1000 pg/ml
PGE2 assayed in parallel.
3.2.7.9 Urokinase-Type Plasminogen Activator (uPA) Activity
uPA activity was measured in conditioned cell culture media using the chromogenic plasmin
substrate H-D-valyl-leucyl-lysine-p-nitroaniline dihydrochloride [S-2251™, Chromogenix, Italy] in
a modification of the Leprince et al. (1989)462 adaptation of the two-step assay of Coleman and
Green (1981).149 20 l of serum-free CM was combined in 96-well microwells with 20 l of
plasminogen solution (0.1g/L bovine plasminogen [Sigma, USA] in 100mM glycine, 100mM Tris-

139.
HCl, 0.5mg/ml bovine serum albumin, pH 8.5) and mixed well. Plasminogen-independent activity
was assessed by adding equivalent buffer only. After incubation (2 hrs, 37°C), 160 l of substrate
mixture was added (0.1mM H-D-Val-Leu-Lys-pNA in 200mM Na3PO4, 200mM NaCl, 0.25mM
DTNB [5,5'-dithio-bis(2-nitrobenzoic acid); Sigma, USA], 0.1% Triton X-100, pH 7.5). The
combination of dilution, pH reduction, and addition of NaCl and Triton X-100 effectively
terminates further plasminogen activation and maximises plasmin activity.149 After a further 15
minutes of incubation, absorbance at 405nm was read on microplate spectrophotometer [Biorad
Model 450 Microplate reader; Biorad, USA]. Plasminogen-independent activity was subtracted
from plasminogen-dependant wells, and the results compared to a range of human urokinase
standards (0-0.01 g/ml) [Sigma, USA] included in each microplate.
Alternative assay was conducted using the fluorogenic urokinase substrate benzyloxycarbonyl(Bzl)-
Gly-Gly-Arg-[7-amino-4-methylcoumarin](AMC) [Novabiochem, Switzerland].938 Briefly,
substrate was dissolved in DMSO and diluted to a 0.2mM working solution in 20mM Tris-HCl, pH
7.5. 20 l of CM (or uPA standard) and 20 l of substrate solution were combined in a glass test tube
and incubated for 24 hours at 37°C. Reaction was terminated with 1ml of 0.1M -amino-n-caproic
acid (EACA) [Sigma, USA] in 20mM Tris, pH 7.5, and the fluorescence read on a Perkin Elmer
LS-50 luminescence spectrometer ( ex 383nm, em 455nm).
3.2.7.10 IL-6 Bioassay
IL-6 content of CM from primary synovial cultures was assayed using a bioassay of IL-6-dependant
7TD1 hybridoma cells [supplied by the Walter and Elisa Hall Institute, Melbourne, Australia].358
7TD1 cells were grown in RPMI-1640 media [Sigma, USA], 10 M ß-mercaptoethanol, 10% v/v
FCS (RPMI-1640/10%FCS), supplemented with 100 ng/L IL-6 [human recombinant IL-6;
Boehringer Mannheim Biochemica, Germany]. For the bioassay, 25 l of primary CM was added to
duplicate wells of a 96-well culture plate [Techno Plastic Products, Switzerland] and diluted with

140.
75 l of RPMI-1640/10% FCS. Dilutions of recombinant human IL-6 served as a standard for
quantification. 7TD1 cells were harvested by centrifugation, washed twice in RPMI/10% FCS
(without IL-6), and added to the assay plates at a density of 10,000 cells/well (in 100 l). After
incubation for 72 hours at 37°C, 0.5 Ci of 3H-thymidine was added to each well before incubation
for a further 6 hours. Cells were harvested as described above (see 3.2.7.3). Radioactivity of the
discs was counted by routine liquid scintillation counting. Half-maximal stimulation was produced
by 10 pg/ml IL-6, similar to that reported in other studies358,583,875 and the product data sheet (ICN
Biomedicals, USA).
3.2.7.11 IL-1 Bioactivity (Lymphocyte Proliferation Assay)
IL-1 bioactivity of primary synoviocyte CM was assayed using an ovine lymphocyte proliferation
assay.743 The thymus of a 4-week old lamb was collected aseptically and minced coarsely before
being pushed through a stainless steel sieve (mesh size 150 m). The resulting material was diluted
in DMEM/10% FCS and filtered through a single layer of sheer polyester voile (mesh size 25 x 25
m) to yield a single cell suspension. After three wash cycles of centrifugation and resuspension in
fresh media, the cells were counted and added to 96-well tissue culture plates at 7x105 cells/well in
DMEM/10%FCS containing 4 g phytohaemagglutinin [from Phaseolus vulgaris; Sigma, USA]. An
equal volume of CM sample (diluted 1:5 in DMEM/10% FCS) or diluted ovine recombinant IL-1ß
was added before incubation for 48 hours. 3H-thymidine was added to each well (0.5 Ci/well)
before incubation for a further 16 hours. Cells were harvested onto glass filter paper using a cell
harvester, and the incorporated radioactivity counted by routine liquid scintillation counting. The
IL-1ß standard curve showed a linear proliferative response between 2 and 1000 pg/ml; limit of
detection was approximately 1 pg/ml.

141.
3.2.8 Oestrogen Receptor Assay
The presence of oestrogen receptors in ovine synovial fibroblasts was demonstrated by Scatchard
saturation analysis using hydroxyapatite precipitation.65 A single saturating dose (SSD) assay was
then used to compare receptor levels between treatment groups.776 For the saturation analysis, four
75cm2 flasks of synovial fibroblasts (second passage) were cultured to confluence and harvested by
brief trypsinization and centrifugation. Cells were resuspended in 1000 l TEDG buffer (10mM Tris
HCl, 1.5mM EDTA, 2mM dithiothreitol, 10% v/v glycerol, pH 7.5). Cells were disrupted by a
single freeze-thaw cycle followed by sonication (Braunsonic 1510 [Braun, Germany] sonicator,
50W, 2 bursts of 20 seconds). An aliquot of the homogenate was removed for DNA determination.
The homogenate was then centrifuged for 12 minutes at 29 psi (approximately 250,000 g) in an
airfuge [Beckman Air-driven Ultracentrifuge, Beckman, USA]. 100 l of cytosol was combined in a
glass test tube with 25 l of [2,4,6,7-3H]oestradiol [Amersham Pharmacia Biotech, UK] and 25 l of
either buffer (non-specific bonding) or unlabelled oestradiol solution (specific binding). A range of
3H-oestradiol concentrations (8,6,4,2, and 1nM) were used with or without a corresponding 100-
fold excess of 17ß-oestradiol [ICN Biomedicals, USA]. After mixing, the test tubes were covered
and incubated for 4 hours at 4°C. During incubation, a 10 l sample was taken from each tube to
determine the total radioactivity.
After incubation, 300 l of 10% washed hydroxyapatite slurry (0.1g/ml in TEDG buffer) [Biorad
DNA-grade HTTP; Biogel, Australia] was added. After incubation for a further 45 minutes at 4°C,
2ml of buffer was added before sedimenting the hydroxyapatite by centrifugation (1000g, 5 mins,
4°C), and complete removal of the supernatant (containing non-protein-bound steroid) to waste.
This washing step was repeated twice more, before transfer of the hydroxyapatite sediment to
scintillation vials for routine ß-counting. Specific binding was determined by subtracting non-
specific binding (determined by regression plot of non-specific binding versus total radioactivity
added) from total binding. For Scatchard analysis, results were plotted as bound/free (nM/nM)

142.
versus bound steroid (nM). For SSD assays, results are expressed as pmol bound per mg DNA in
the crude homogenate.
3.2.9 Statistical Methods
Statistical comparisons were generated using specialist software [Statview 5.0, SAS Institute Inc.,
USA], performing one-way analysis of variance (ANOVA) to analyse variance across treatment
groups, and Fisher’s protected least significant difference (Fisher’s PLSD) test to compare means of
all treatment groups. Only the following statistical comparisons were reported: all treated groups vs
NOC; OVX+GTN vs OVX; MX+GTN, OVX+MX, and OVX+MX+GTN vs MX; and
OVX+MX+GTN vs OVX+MX. When multiple zones or points were compared simultaneously,
zone (inner, middle, or outer) or point identifier was included as a second independent variable. In
comparing histopathological scores in multiple zones for each region, mean scores (for single
criteria) or mean aggregate scores (for sum of means) were tested statistically. Unless otherwise
described, correlations were tested using the Pearson correlation test. A significance level of p=0.05
was used throughout.

143.
3.3 RESULTS
3.3.1 BODY WEIGHTS
The mean body weight of all sheep (Figure 3.14) at the start of the trial was 56.1±5.2 kg, with no
significant differences between sheep later allocated to treatment groups. After meniscectomy,
sheep lost an average of 5.6 kg by the time of next weighing. Unoperated sheep gained weight
rapidly early in the trial period (corresponding to spring pasture growth), with lower weight gains in
the latter half of the trial. An average of 3.4 kg was lost by all sheep due to shearing of wool in the
middle of the trial period. GTN treatment had no significant effect on body weight.
OVX+MX+GTN sheep has consistently lower body weights than the OVX+MX sheep after
initiating treatment, but this difference was not statistically significant by repeated measures
analysis of variance (p=0.187).
3.3.2 GROSS PATHOLOGY
The joints of non-meniscectomised sheep showed minimal gross pathology, though some degree of
roughening or fissuring of cartilage was seen in the MTP in 83% of specimens, usually in the region
not covered by the meniscus. Mild cartilage lesions were less commonly observed in the LTP and
MFC, while no cartilage abnormalities were observed in the LFC. Four non-meniscectomised joints
showed minor osteophyte formation. The joints of all meniscectomised groups showed significantly
greater cartilage erosion and osteophyte formation (p<0.0001) (Figure 3.15). Gross erosion of
cartilage and marked osteophyte formation was mostly limited to the meniscectomised lateral
compartment, with the medial compartment showing minor changes. The cartilage of the MFC was
often grossly normal despite severe lateral pathology (54% of specimens). Complete erosion of LFC
and/or LTP cartilage exposing the subchondral bone was seen in a few animals. Ovariectomy and

144.
GTN treatment had no significant effect on the development of gross cartilage pathology or
osteophyte formation.
In non-meniscectomised animals, cartilage scores were higher in left knees when compared to right
knees (2.67±2.34 vs 1.25±0.75), though this difference was not statistically significant (p=0.059).
Osteophytic changes were observed only in left knees (p=0.004). However, there was no difference
in the gross pathology observed between left and right knees of meniscectomised sheep.
Nineteen of 24 of meniscectomised sheep (79%) showed some degree of synovial change (synovial
hyperaemia, fibrosis, or synovitis). Nine of 24 specimens (37%) were found to have overt synovitis.
The degree of gross synovitis was significantly correlated to both gross cartilage pathology
(p=0.021) and osteophyte formation (p=0.009) (Figure 3.16). Four non-meniscectomised sheep
(17%) showed slight synovial abnormalities and none demonstrated synovitis.
3.3.3 BIOMECHANICAL TESTING
Dynamic biomechanical testing of non-meniscectomised animals (Figure 3.17) revealed an overall
reduction in phase angle ( ) in tibial cartilage of both NOC+GTN (11.0±1.9°; p=0.0003) and OVX
(10.9±2.2°; p<0.0001) sheep relative to NOC (12.1±2.3°). Phase angle of OVX+GTN (12.6±2.7°)
was significantly greater than OVX (p<0.0001) and did not differ significantly from NOC.
Similarly, tibial cartilage thickness was reduced overall relative to NOC in NOC+GTN (p<0.0001)
and OVX (p=0.0002), but did not differ in OVX+GTN sheep. Thinning of cartilage was most
prominent in the LTP. Dynamic stiffness was found to be significantly increased overall in
NOC+GTN (6.31±4.76 N/mm; p=0.015) and OVX (6.20±5.52 N/mm; p=0.039) relative to NOC
(5.59±5.06 N/mm). However, when stiffness was corrected for cartilage thickness as the dynamic
shear modulus (G*) according to the theory of Hayes et al. (1972),331 no overall differences were
apparent, though regional differences were noted particularly in the inner regions of the LTP

145.
(Figure 3.17). Results for G* and were comparable with previous reports of the shear properties of
articular cartilage.745,746,935
After meniscectomy, cartilage thickness decreased in the inner regions of the tibial plateaux and
increased in the outer regions (Figure 3.18). MX+GTN had significantly thinner cartilage in the
eroded middle regions of the LTP relative to MX. OVX+MX sheep had significantly thicker
cartilage overall relative to MX (p=0.001); this greater thickness was evident in both inner (eroded)
and outer (intact) regions. However, cartilage thickness of OVX+MX+GTN sheep differed from
MX in the inner MTP only, and did not differ overall. Dynamic shear modulus (G*) was
significantly lower in all meniscectomised groups, primarily due to changes in the lateral tibial
cartilage; this reduction was significantly greater in OVX+MX and OVX+MX+GTN groups
relative to that of MX (p<0.0001). Phase changes in meniscectomised groups were variable; phase
was significantly lower overall relative to NOC in OVX+MX animals (p=0.029) but significantly
greater in OVX+MX+GTN sheep (p=0.024).
3.3.4 HISTOLOGICAL ASSESSMENTS
3.3.4.1 Cartilage Histology
3.3.4.1.1 Cartilage Histopathological Scores
OVX animals showed significantly greater aggregate histopathology scores (Table 3.2) in the MFC,
relative to NOC (p=0.009). Specifically, scores for abnormalities of structure (p=0.045) and
interterritorial staining (p=0.048) were significantly increased relative to NOC. OVX+GTN sheep
did not differ from NOC in the MFC, and showed significantly lower scores for structure and
interterritorial staining relative to OVX (p=0.043 and p=0.03 respectively). Both OVX and
OVX+GTN animals showed significant loss of territorial staining in the LTP (p=0.035 and

146.
p=0.024) relative to NOC.
Histopathology scores were increased in all regions in meniscectomised groups, though these
differences were not significant in the MTP. MX+GTN animals were found to have significantly
greater structural damage (p=0.033) and loss of interterritorial staining (p=0.003) in the MFC, and
greater scores for chondrocyte cloning in the LFC (p=0.015), relative to MX. OVX+MX and
OVX+MX+GTN animals had significantly lower aggregate scores in the LFC, relative to MX
(p=0.018 and p=0.034 respectively). In the LFC, OVX+MX showed lower scores for structure
(p=0.030), territorial staining (p=0.003) and interterritorial staining (p=0.002) relative to MX.
OVX+MX and OVX+MX+GTN animals also showed reduced loss of interterritorial staining
relative to MX in the MFC (p=0.007 and p=0.007 respectively).
Ovariectomy resulted in increased neovascularisation of the calcified cartilage layer in both
qualitative and quantitative indices. Scores for calcified cartilage pathology (Table 3.3) were
significantly increased in both OVX and OVX+GTN sheep when the whole joint was considered
(p=0.038 and p=0.007 respectively). Calcified cartilage vascularity (number of invading
vessels/tidemark length) (Figure 3.19) was also increased overall in OVX (p=0.0003), OVX+GTN
(p=0.002), and NOC+GTN (p=0.03) relative to NOC. Femoral condyle chondrocyte density did not
differ between non-meniscectomised groups (Table 3.4)
After meniscectomy, scores for both calcified cartilage pathology (Table 3.3) and vessel number
(Figure 3.19) were increased in all regions (p<0.0001), especially the lateral compartment.
MX+GTN sheep had a greater number of vessels invading calcified cartilage in the LFC relative to
MX (p=0.031). However, scores for pathology and vessel number were lower than MX sheep in the
MTP (p=0.023 and p=0.010 respectively). Vessel number was higher overall in OVX+MX and
OVX+MX+GTN groups overall relative to MX sheep (p=0.019 and p=0.034 respectively), though
scores for calcified cartilage pathology did not differ. Chondrocyte density was increased in the

147.
LFC in MX (p=0.013) and MX+GTN (p=0.010), but not OVX+MX or OVX+MX+GTN animals
(Table 3.4).
3.3.4.1.2 Computerised Histomorphometry
Uncalcified Cartilage Thickness
Mean uncalcified cartilage (UCC) thickness measures determined by image analysis (Figures 3.20 –
3.23, above) were comparable to results using the needle probe. Overall, NOC+GTN had
significantly thinner UCC in the MFC (p=0.035) and LTP (p<0.0001), with a similar trend in the
LFC. OVX sheep also had thinner UCC in the LTP (p=0.006) with a similar trend in the LFC
(p=0.07).
After meniscectomy, MX sheep showed thinner cartilage in the LFC and thicker cartilage in the
MFC, though these changes were not significant. Cartilage was significantly thinner in the eroded
middle (p<0.0001) and inner zones (p<0.0001) of the LTP and inner zone of the MTP (p=0.0003),
while cartilage thickness was significantly increased in the outer zone of the LTP (p=0.011).
OVX+MX animals showed significantly thicker UCC relative to MX in all locations, except the
inner and middle zones of the MFC. OVX+MX+GTN similarly showed thicker cartilage relative to
MX, except in the LTP, where UCC was significantly thinner than that of OVX+MX animals
(p=0.0008).
Subchondral Bone Plate Thickness
Subchondral bone plate (SCP) thickness [Figures 3.20 – 3.23, below] was increased in NOC+GTN
in the MTP (p=0.002), with a similar trend in the MFC (p=0.156). OVX sheep had significantly
thinner SCP in the LFC (p<0.01) and LTP (p=0.001), while OVX+GTN had significantly thicker
SCP relative to OVX in the MFC (p=0.04), LFC (p<0.0001) and LTP (p=0.002) and did not differ
significantly from NOC.

148.
After meniscectomy, the SCP of MX animals was significantly thicker in the LFC (p=0.035) and
the outer region of the LTP (p=0.008), while thinning of the SCP was found in the outer zone of the
MFC (p=0.034). MX+GTN animals showed significantly greater increase in SCP thickness relative
to MX in the middle zone of the LFC (p=0.024), and reduced loss of bone thickness in the outer
zone of the MFC (p=0.049). OVX+MX and OVX+MX+GTN groups did not differ significantly
from MX, though OVX+MX showed no increase in SCP thickness relative to NOC in the outer
zone of the LFC, as was seen in MX sheep.
Intensity Of Toluidine Blue Staining
The mean greyscale pixel intensity of toluidine blue-stained histological sections, an index of
proteoglycan content (Figure 3.24), was significantly lower in the MFC of NOC+GTN sheep
(p=0.011). Similarly, staining intensity was also significantly lower in the LFC and MFC of both
OVX (p=0.0005 and p=0.002 respectively) and OVX+GTN animals (p=0.040 and p<0.0001
respectively).
After meniscectomy, toluidine blue staining was significantly lower in all zones except the inner
zone of the LFC. Loss of staining was significantly greater in OVX+MX sheep relative to MX in
the LFC (p=0.021), with a similar trend in the MFC. Similarly, loss of staining in OVX+MX+GTN
sheep was greater than that of MX in both the LFC (p=0.021) and the MFC (p=0.005).
Toluidine blue staining intensity of patellar plugs (Figure 3.25) was significantly reduced in OVX
animals only (p=0.023). MX+GTN, OVX+MX and OVX+MX+GTN animals all showed lower
intensity of toluidine blue staining though this was not statistically significant.

149.
Osteophyte Size
Osteophyte size, measured as the cross-sectional area (mm2) (Figure 3.26), was greatest in the LFC
and to lesser extent the LTP. While no statistically significant differences were evident in each
region, pooling of the regional values showed that both MX+GTN (p=0.049) and OVX+MX
(p=0.022) sheep showed greater osteophyte growth relative to MX ewes.
3.3.4.2 Collagen Birefringence
Collagen birefringence profile plots revealed a distinct superficial peak in greyscale intensity in all
regions tested (Figures 3.27 – 3.29). Superficial collagen birefringence was significantly increased
in both NOC+GTN and OVX sheep (relative to NOC) in the LFC and middle zone of the LTP, with
a similar trend in other regions. NOC+GTN sheep showed significantly greater intermediate zone
birefringence in the outer zone of the LTP, and middle zone of the MTP, relative to NOC.
3.3.4.3 Synovial Histopathological Scores
Differences between treatment groups were modest. As GTN-treated animals did not differ
significantly from untreated animals in any criteria, results were pooled within each surgical group
(Table 3.5). MX±GTN animals showed greater scores for intimal hyperplasia (p<0.0001),
inflammatory infiltrate (p=0.012), vascularity (p=0.017), and greater sum of scores (p=0.005),
relative to NOC±GTN synovia. Similarly, OVX+MX±GTN sheep showed greater scores for intimal
hyperplasia (p=0.018), inflammatory infiltrate (p=0.031), fibrosis (p=0.008), vascularity (p=0.032),
and greater sum of scores (p=0.036), relative to NOC±GTN synovia. OVX+MX±GTN synovia
showed less intimal hyperplasia relative to MX±GTN sheep (p=0.037). Cellularity of the synovial
initima did not differ significantly between treatment groups (Figure 3.30).

150.
3.2.4.4 Undecalcified Bone Histomorphometry
Histomorphometry of undecalcified bone sections from the middle zone of the LTP revealed few
statistically significant differences between treatment groups. NOC+GTN sections showed less
eroded trabecular surface relative to NOC (p=0.0072) (Figure 3.31). Osteoid surface, trabecular
bone volume (Figure 3.32), and calcein-labelled surface did not differ significantly between groups.
SCP calcein score (see 3.2.4.4) was significantly increased in OVX+MX relative to NOC animals
(p=0.387), and reduced in OVX+GTN relative to OVX (p=0.0313) (Table 3.6).
3.3.5 BONE MINERAL DENSITY
Ovariectomised sheep had lower bone mineral density (BMD) than NOC sheep at all extra-articular
sites tested (Figure 3.34), though this difference was statistically significant only in the first lumbar
vertebra (p=0.045). Meniscectomised sheep also showed loss of BMD relative to NOC animals,
which was statistically significant in the first lumbar vertebra (p=0.045) and the femoral head and
neck sites (p=0.047).
Subchondral BMD (Figure 3.35) was significantly greater in the LFC of NOC+GTN (p=0.004) and
lower in both OVX and OVX+GTN sheep in the LTP (p=0.004, p=0.015 respectively) and MTP
(p=0.0002, p<0.0001 respectively), relative to NOC. After meniscectomy, subchondral BMD was
increased in the LFC (p=0.002) and LTP (p=0.018), and decreased in the MFC (p=0.0003) and
MTP (p<0.0001). MX+GTN sheep showed a greater increase in subchondral BMD in the LFC
(p=0.019) and LTP (p=0.011), and reduced loss of BMD in the MTP (p=0.023), relative to MX.
Subchondral BMD changes in OVX+MX sheep were similar to those in MX animals, though
OVX+MX+GTN sheep had lower subchondral BMD in the LFC (p=0.018) and MFC (p=0.021).

151.
Determinations of the 'cubic' BMD (gm/cm3) of tibial bone slabs (Figure 3.36) more clearly showed
the effect of ovariectomy on subchondral BMD. OVX sheep had lower BMD in the LTP (p=0.022)
and MTP (p=0.026), relative to NOC. Similarly, OVX+MX sheep had lower BMD in the LTP
(p=0.003) and MTP (p=0.013) relative to MX. Results were strongly correlated with subchondral
bone plate thickness as measured in the contralateral limb (p<0.0001, r2=0.141) (Figure 3.37).
Across all groups, bone mineral density was greater in the left stifle relative to the right, in both the
tibial plateaux (0.85±0.14 vs 0.82±0.16; p=0.032) and femoral condyles (1.01±0.17 vs 0.96±0.16;
p=0.0014). This difference was most pronounced in the medial compartment.
3.3.6 BLOOD AND URINE MARKERS
3.3.6.1 Plasma 17ß-Oestradiol
Plasma oestradiol levels (Figure 3.38) were relatively low in all sheep. Levels were significantly
reduced at 5 weeks and subsequent time points post-ovariectomy. In MX±GTN sheep, plasma
oestradiol levels were significantly increased at the time of sacrifice, relative to NOC animals
(p=0.004).
3.3.6.2 Serum Osteocalcin
At the time of sacrifice, serum osteocalcin levels (Figure 3.39) were increased in OVX+GTN
(p=0.047), MX (p=0.020), OVX+MX (p=0.040) and OVX+MX+GTN (p=0.005) sheep relative to
NOC animals. Levels in MX+GTN animals were significantly lower relative to MX (p=0.004), and
did not differ from those of NOC sheep.

152.
3.3.6.3 Urine Collagen Crosslinks
Urinary levels of collagen pyridinium crosslinks (Figure 3.40) did not differ significantly between
treatment groups. MX+GTN sheep showed a tendency towards lower levels of Pyr and Dpyr
relative to MX animals.
3.3.7 ASSAYS ON TISSUE-CULTURE CONDITIONED MEDIA
3.3.7.1 Patellar Cartilage Explants
Nitric Oxide
Interleukin-1ß-stimulated cultures produced approximately twice the amount of nitric oxide
(measured as nitrite / g DNA) (Figure 3.41) compared to unstimulated cultures (p<0.0001). Nitric
oxide production by OVX cultures was increased in both unstimulated (p=0.005) and IL-1ß-
stimulated (p<0.0001) wells relative to NOC. However, OVX+GTN cultures produced less NO
relative to OVX in both unstimulated (p=0.044) and IL-1ß-stimulated (p=0.010) wells, and did not
differ significantly from NOC cultures. In IL-1ß-stimulated wells, MX/MX+GTN cultures
produced less NO relative to NOC/NOC+GTN cultures (p=0.046)
35SO4-labelled Proteoglycan Synthesis
IL-1ß-stimulation caused a slight reduction (average -23%) in de novo (35SO4-labelled)
proteoglycan synthesis (Figure 3.42). Approximately 25% of the total 35SO4-labelled proteoglycan
was present in conditioned media., with no significant difference between groups in the proportion
released. Explants from OVX sheep produced more 35SO4-labelled proteoglycan relative to NOC in
unstimulated cultures (p=0.031). Similarly, OVX/OVX+GTN explants produced more 35SO4-
labelled proteoglycan relative to NOC/NOC+GTN under IL-1ß-stimulated conditions (p=0.010).

153.
MMP Activity
Auto-activated MMP levels (not shown) represented on average 30% of total MMP activity. Auto-
activated MMP activity was increased in unstimulated OVX cultures relative to NOC cartilage
(14.9±5.6 vs 10.6±2.6, p=0.019). IL-1ß-stimulation had no significant effect on auto-activated
levels but produced an approximately four-fold increase in total (APMA-activated) MMP activity
(p<0.0001) (Figure 3.43). In unstimulated cultures, there were no significant difference between
groups in total MMP activity. However, under IL-1ß-stimulated conditions, total MMP activity was
increased relative to NOC in both OVX (p<0.0001) and OVX+GTN (p=0.0002) cultures.
3.3.7.2 Subchondral Bone Explants
Nitric Oxide
There were few significant differences between groups (Figure 3.44). OVX+MX+GTN cultures
produced less NO per mg wet weight relative to OVX+MX (p=0.016).
PGE2
Prostaglandin E2 release (pg per mg wet weight) was increased in OVX+MX (p=0.001) and
OVX+MX+GTN (p=0.002) explants relative to NOC (Figure 3.45). OVX+MX/OVX+MX+GTN
sheep also produced greater levels of PGE2 relative to MX/MX+GTN explants (p=0.002), which
did not differ from NOC.
Urokinase-type Plasminogen Activator Activity
Urokinase-type plasminogen activator (uPA) activity was increased in conditioned media from
OVX (p=0.023), OVX+MX (p=0.0002), and OVX+MX+GTN (p<0.0001) explants relative to NOC
(Figure 3.46). Activity in media from MX explants did not differ from NOC cultures. Activity was
increased relative to MX in conditioned media from both OVX+MX (p=0.0002) and
OVX+MX+GTN (p<0.0001) cultures.

154.
3.3.7.3 Synovial Cultures
3.3.7.3.1 Passaged Synovial Fibroblast Cultures
Nitric Oxide
Addition of IL-1ß to synovial fibroblast cultures caused a slight but statistically significant
reduction in nitric oxide production (measured as nmol nitrite / g DNA) (p=0.005). NO release
was greater in MX cultures (Figure 3.47) relative to NOC, in both unstimulated (p=0.004) and IL-
1ß-stimulated (p=0.002) wells. However, OVX+MX cultures produced less NO relative to MX, in
both unstimulated (p=0.042) and IL-1ß-stimulated (p=0.036) wells.
PGE2
A wide variation between individual cultures with respect to PGE2 release. IL-1ß-stimulation
induced a 3-4 fold increase in PGE2 release (p<0.0001) (Figure 3.48). No statistically significant
differences were observed between treatment groups.
MMP Activity
IL-1ß-stimulated cultures demonstrated an increase in total MMP activity (mean increase 145% of
unstimulated cultures, p<0.0001), but showed no significant effect on auto-activated levels. Auto-
activated levels averaged 9% of total (APMA-activated) levels in unstimulated cultures and 6.6% of
IL-1ß-stimulated totals. No significant differences in auto-activated MMP activity were observed.
MX and OVX groups had higher total activity than NOC, though this difference was not significant.
OVX+MX and OVX+MX+GTN sheep possessed greater total MMP activity relative to NOC,
under both unstimulated (p=0.023 and p=0.003 respectively) and IL-1ß-stimulated (p=0.002 and
p=0.0004 respectively) conditions (Figure 3.49). Total IL-1ß-stimulated MMP activity of
OVX+MX sheep was greater than that of MX cultures (p=0.038). Similarly, total MMP activity of

155.
OVX+MX+GTN cultures were also elevated relative to MX, under both unstimulated (p=0.020)
and IL-1ß-stimulated conditions (p=0.009).
Urokinase-type Plasminogen Activator Activity
Urokinase-type plasminogen activator activity did not differ between IL-1ß-stimulated and
unstimulated synovial fibroblast cultures (Figure 3.50). uPA activity was increased in MX cultures
relative to NOC, in unstimulated wells (p=0.026) but not IL-1ß-stimulated wells. OVX+MX
cultures released significantly greater levels of uPA activity relative to NOC under both resting
(p=0.011) and IL-1ß-stimulated (p=0.005) conditions. Under IL-1ß-stimulated conditions, uPA was
increased relative to MX in both OVX+MX (p=0.004) and OVX+MX+GTN (p=0.048) cultures.
Assay using the fluorogenic substrate (results not shown) proved more time-consuming and less
sensitive than the former method, but uPA activities obtained in preliminary tests closely matched
those from the chromogenic substrate assay, confirming urokinase as the principal plasminogen
activator in these CM.
Proliferation Rate and Confluent Density
There were no significant differences in proliferation rate (3H-thymidine incorporation, DPM)
between treatment groups (Figure 3.51). Confluent cell density was approximated as DNA content
( g) per well in 24-well microplate cultures (Figure 3.52). Confluent cell density was on average
90% greater in wells containing 10% FCS compared to those with unsupplemented media (results
not shown). In FCS-supplemented cultures, confluent density was lower in meniscectomised groups
relative to non-meniscectomised animals in both unstimulated (3.05±0.13 vs 3.60±0.14 g
DNA/well; p=0.0076) and IL-1ß-stimulated wells (3.05±0.13 vs 3.63±0.15 g DNA/well;
p=0.0048).

156.
3.3.7.3.2 Primary Synovial Cultures
Interleukin-6 Bioactivity
Production of IL-6 bioactivity by primary synovial isolates was much lower than that reported by
previous authors (Figure 3.53).296,539,698 Addition of LPS (1 g/ml) caused an average 8-fold increase
in IL-6 bioactivity (p<0.0001). IL-6 bioactivity was greater in conditioned media from
OVX/OVX+GTN cultures relative to NOC/NOC+GTN wells (p=0.036).
Interleukin-1 Bioactivity
Spontaneous release of IL-1 bioactivity by primary synovial isolates was lower in OVX /
OVX+GTN cultures relative to NOC / NOC+GTN (p=0.022) (Figure 3.54). Similarly, OVX+MX /
OVX+MX+GTN cultures produced less IL-1 bioactivity relative to both NOC / NOC+GTN
(p=0.015) and MX / MX+GTN (p=0.041).
3.2.8 OESTROGEN RECEPTORS
Scatchard analyses performed on several of the cell lines confirmed the presence of specific
oestradiol receptors in ovine synovial fibroblast cultures, though only low levels were detected. A
single class of receptor appeared to be present (Figure 3.55A). Receptor levels (as detemined by
single point assay) were increased following ovariectomy and meniscectomy, though this difference
was not statistically significant (Figure 3.55B).

157.

158.

159.

160.

161.

162.

163.

164.

165.

166.

167.

168.

169.

170.
Table 3.2: Mean aggregate cartilage histopathology scores in the lateral and medial femoral condyles (LFC, MFC) and tibial plateau (LTP, MTP). * differs significantly from NOC (p<0.05), † differs significantly from MX (p<0.05), ‡ OVX+GTN differs significantly from OVX (p=0.023); maximum possible score = 26 (see 2.2.3)
Treatment Group LFC MFC LTP MTP
NOC 1.87±0.34 1.45±0.57 2.19±0.31 3.37±0.53 NOC+GTN 2.21±1.05 2.25±1.16 2.67±1.09 2.33±2.10 OVX 1.75±0.71 3.04±0.97* 2.00±0.64 1.83±0.95 OVX+GTN 1.92±1.03 2.12±0.79‡ 2.87±0.75 2.81±1.28 MX 12.67±1.84* 5.21±1.52* 14.54±1.62* 6.92±2.27 MX+GTN 12.92±4.10* 6.33±2.21* 14.12±2.87* 8.21±3.87 OVX+MX 9.54±2.03*† 4.08±1.62* 14.79±1.28* 7.75±3.96 OVX+MX+GTN 9.87±3.39*† 3.92±1.18* 15.37±4.20* 7.92±3.71
Table 3.3: Mean scores for calcified cartilage pathology in the lateral and medial femoral condyles (LFC, MFC) and tibial plateau (LTP, MTP). * differs significantly from NOC (p<0.05), † MX+GTN differs significantly from MX (p=0.023); maximum possible score = 4
Treatment Group LFC MFC LTP MTP
NOC 1.33±0.20 1.21±0.12 1.12±0.31 1.21±0.19 NOC+GTN 1.08±0.30 1.25±0.55 0.92±0.30 1.21±0.43 OVX 1.37±0.34 1.58±0.49 1.29±0.29 1.58±0.75 OVX+GTN 1.58±0.26 1.62±0.26 1.12±0.52 1.79±0.37* MX 2.17±0.49* 1.75±0.45* 1.79±0.48* 1.96±0.56* MX+GTN 2.21±0.64* 1.67±0.58 2.12±0.47* 1.25±0.39† OVX+MX 2.21±0.43* 1.50±0.45 1.75±0.32* 1.58±0.46 OVX+MX+GTN 2.04±0.46* 1.67±0.20 2.08±0.54* 1.87±0.75*
Table 3.4: Chondrocyte density (nuclei / 250 x 250 m) in the intermediate zone of the central weight-bearing regions of the lateral and medial femoral condyle (LFC, MFC). * differs significantly from NOC (p<0.02)
Treatment Group LFC MFC
NOC 54.7±6.2 46.2±5.8 NOC+GTN 54.0±10.7 48.1±5.4 OVX 52.8±14.1 42.3±4.9 OVX+GTN 49.8±13.3 46.2±13.5 MX 77.2±24.8* 49.7±13.9 MX+GTN 79.1±22.3* 46.9±8.3 OVX+MX 71.8±6.5 46.3±10.9 OVX+MX+GTN 68.7±13.0 44.9±8.4

171.

172.

173.

174.

175.

176.

177.

178.

179.

180.

181.
Table 3.5: Synovial membrane histopathology scores (results for GTN-treated and untreated animals are pooled). * differs significantly from NOC±GTN (p<0.05), † OVX+MX±GTN differs from MX±GTN (p=0.037); maximum score = 3; maximum sum = 12
Treatment Group Intimal
hyperplasia
Cellular
infiltrate
Subintimal
fibrosis
Vascularity SUM
NOC±GTN 0.92±0.63 1.00±0.60 1.58±0.56 1.17±0.72 6.21±3.09
OVX±GTN 0.92±0.67 1.21±0.84 1.87±0.64 1.25±0.69 6.50±2.28
MX±GTN 2.17±0.65* 1.83±0.98* 1.92±0.51 1.92±0.67* 9.29±2.50*
OVX+MX±GTN 1.58±0.70*† 1.71±0.62* 2.25±0.62* 1.83±0.86* 8.46±2.24*
Figure 3.30: Cellularity of the synovial intimal layer (nuclei per 250 mx50 mx4 m). No significant differences between treatment groups. Error bars represent one standard error.
NOC OVX MX OVX+MX0
10
20
30
40
50
60Untreated GTN-treated

182.

183.

184.

185.

186.

187.

188.

189.

190.

191.

192.

193.

194.

195.

196.

197.

198.
3.4 DISCUSSION
3.4.1 EFFECT OF LATERAL MENISCECTOMY ON FEMORO-TIBIAL JOINT
TISSUES OF AGED EWES
3.4.1.1 Cartilage Changes After Meniscectomy
The cartilage damage in this trial was more severe than that seen in the younger wethers used in
Chapter 2. In these aged ewes, cartilage erosions were larger and deeper, occasionally to eburnated
subchondral bone. In contrast to the previous trial, an increase in cartilage thickness was observed
only in the outer zone of the LTP. Loss of proteoglycan was also more severe, especially in
uneroded areas such as the medial femoral condyle. This contrasts with the hypertrophic repair
response seen in peripheral areas in the younger sheep of Chapter 2. The difference between the two
trials in the severity of the lesions produced by an identical meniscectomy procedure may be related
to genetic variation, gender, or the older age of the sheep used. Aged human cartilage has been
shown to have reduced capacity for repair responses.422,896
Cartilage lesions were most severe in the tibial plateau, probably because the femoro-tibial
articulation effectively involves a greater cartilage surface area in the femoral condyles, compared
to the tibial surface. Lesions were less severe in the medial compartment, and due to the high
frequency of mild fibrillation in the medial tibial plateau in control animals, few significant
differences between groups were observed in this region. Mild cartilage fibrillation in the medial
tibial plateau has been previously reported as a normal finding in other species.126
There were relatively few statistically significant differences in cartilage biomechanical properties
between meniscectomised and unoperated sheep. This was mostly due to aberrant readings from
severely damaged regions of cartilage, where erosion down to the level of calcified cartilage or

199.
subchondral bone yielded extremely high stiffness values and hence a high standard error. Loss of
dynamic stiffness (G*) was most dramatic in the outer portions of the LTP, i.e. the area previously
protected by the meniscus. A similar loss of dynamic stiffness has been reported in other
biomechanical studies of OA cartilage.455,745,746 However, a consistent increase in phase lag ( ) was
not evident in this study as reported previously.34 Reasons for this are unknown and require further
clarification. Le Roux et al. (2000)455 similarly found no phase change in tibial plateau cartilage
following canine meniscectomy.
The in vitro responses of ovine cartilage to IL-1ß (i.e. increased nitric oxide and latent MMP
synthesis, suppression of proteoglycan synthesis) were similar to those reported in other
species.106,125,130,349,684,712,791 However, few differences were evident between patellar cartilage
explants from normal and meniscectomised sheep. No significant difference in nitric oxide release,
35SO4-proteoglycan synthesis, or MMP activity was observed. This finding was unexpected given
numerous studies showing persistent metabolic alterations in OA cartilage.49,538,625,864 The patella
may not be a very representative site to sample, given that the proteoglycan content of patellar
cartilage was similar between normal and menisectomised sheep (Figure 3.25). However, this is
likely due to loss of previously increased (hypertrophic) levels, rather than being indicative of a
normal metabolic state. Appleyard et al. (1999)33 showed significant biomechanical and
immunohistochemical changes in patellar cartilage six months following ovine meniscectomy. It
appears that the metabolic changes associated with meniscectomy in this region are less consistent
in vitro.
3.4.1.1 Synovial Changes After Meniscectomy
Hypothesis: that the gross synovitis thought to occur in this OA model474
would be accompanied by demonstrable microscopic pathology, and persistent
metabolic alterations in synoviocytes in vitro.

200.
Results in this trial confirm that gross synovitis is a common feature of the ovine meniscectomy
model of osteoarthritis. Many (79%) of meniscectomised joints showed some gross evidence of
synovial change, and 37% showed overt synovitis. However, differences at the histological level
were more modest. While meniscectomised sheep were generally found to have a greater degree of
intimal hyperplasia, lymphoplasmocytic infiltration, and vascularisation, similar changes were
observed in some normal sheep. While synovial histopathological scores were found to be
significantly correlated with severity of degenerative joint changes, the predictive power of this
relationship (i.e. r2 values) was low. Intimal fibrosis was not found to be a significant sequelae of
meniscectomy in this aged ewe trial, as it was in the younger wethers used in Chapter 2. These
findings suggest firstly that the method used here to objectively score synovial histopathology lacks
sufficient sensitivity to discriminate the more subtle changes seen in this OA model, compared to
those seen in rheumatoid arthritis or long-standing human OA.470,568,682,768 Sampling at more than
one site may have improved evaluation of synovial changes. However, histological scoring of
human synovial biopsies shows little variation between multiple sites, and good correlation with
gross synovial pathology.469,697 Secondly, it suggests that the synovial membrane of normal sheep
rarely fits the classic description of healthy synovium, with a single layer of intimal cells and little
fibrous tissue. Synovial changes seem to be more common in aged sheep, perhaps due to
accumulated micro-injury over time. Pasquali-Ronchetti et al. (1992)613 investigated age-related
changes in human synovia, and similarly noted an increase in fibrosis and number of fibroblasts
with increasing age.
In vitro, synovial fibroblasts from meniscectomised sheep displayed greater release of nitric oxide
and plasminogen activator, as has been reported in other arthritis models.102,532,535,795 A modest
increase in synovial MMP activity was seen, as has also been demonstrated previously.138,323,366,536
However, meniscectomy had no apparent effect on synovial PGE2 production, in contrast to
numerous published studies, though these have principally examined rheumatoid
synovia.175,338,526,530,715 Synovial PGE2 production was much lower than that of patellar cartilage per

201.
g of DNA, especially after IL-1ß stimulation (results not shown). This is consistent with Amin et
al. (1997)22 who identified cartilage as the major source of prostaglandins within the OA joint,
following super-induction of COX-2 expression in chondrocytes.
The in vitro responses of ovine synovial fibroblasts to IL-1ß stimulation were also not entirely
consistent with those reported in other studies of arthritic synovia. While PGE2 production and
latent MMP activity were significantly increased by the addition of IL-1ß, urokinase activity did not
increase as has generally been reported.308,459,529 Ito et al. (1992)375 similarly found that recombinant
human IL-1 had no effect on urokinase or PAI-1 production by human rheumatoid synovial
fibroblasts. The growth characteristics of synovial fibroblasts in this model were consistent the
report of Smith and Hamerman (1969),767 which noted that rheumatoid synovial fibroblasts showed
decreased cell density when confluent.
The effect of IL-1ß on synovial fibroblast nitric oxide release was surprising, with a slight but
statistically significant reduction in NO production observed. This contrasts with results obtained
from cartilage explants, in which NO production was greater per g of DNA and was
approximately doubled by IL-1ß stimulation. This suggests that chondrocytes may be the major
source of IL-1 in the meniscectomised ovine joint. Other in vitro studies have found NO release by
synovial tissue to be inconsistent.539 Mentzel and Bräuer (1998)539 found that TNF- -induced nitric
oxide production was greater in normal synoviocytes than in arthritis-derived cells. Sakurai et al.
(1995)714 found that whilst NO was produced by freshly isolated human synoviocytes,
supplementation with IL-1ß alone produced minimal stimulation, and that passaged rheumatoid
synovial fibroblast lines produced no detectable NO. Similarly, Rediske et al. (1994)676 found
negligible NO production in cultured human synoviocytes. Species differences seem likely, as rats
and rabbits are the only species in which NO production by passaged synovial fibroblast lines has
been reported.539,795 Even in rabbit cultures, NO production is variable, with a small percentage of
culture lines failing to produce NO under any conditions.795 NO production could not be induced in

202.
the immortalised HIG-82 rabbit synovial fibroblast line.795 Immunohistochemical studies have also
found synovial fibroblasts to be quite heterogenous with respect to iNOS expression.454,532 It seems
likely that a more complex cytokine stimulus is required for the induction of iNOS in ovine
synovial fibroblasts.676 IL-1ß alone does not induce iNOS in human cardiac fibroblasts,752 human
foetal fibroblasts,717 or murine macrophages.894 Interferon- may be an important co-stimulus, by
increasing tetrahydrobiopterin synthesis (a cofactor of NO synthesis) as it does in murine dermal
fibroblasts.894
Also contrary to published reports, meniscectomy had no significant effect on the release of IL-6 or
IL-1 by primary synoviocyte isolates. IL-6 production was found to be several orders of magnitude
lower than that shown by similar bioassay methods in other synovial studies.296,539,698 This may have
been due to species-specific effects on the bioassay system used, as the human recombinant IL-6
used as a standard produced similar effects on hybridoma proliferation as in published reports.583
Elevated IL-6 production is a key defining feature of synovial fibroblast activation which was
apparently absent in this study. IL-1 bioactivity was also low and did not differ in meniscectomised
sheep. While IL-1 bioactivity has also been shown to be species-specific,28,456,521 the assay used in
this study utilised a published ovine lymphocyte assay and a recombinant ovine IL-1ß standard.
Brennan et al. (1989)92 found spontaneous IL-1 production by human OA synovia was very low,
using a similar assay.
3.4.1.3 Bone Responses After Meniscectomy
As has been shown previously,38,39 significant remodelling of subchondral bone was evident six
months after meniscectomy. The subchondral bone plate was thickened in the LFC but thinner in
the outer MFC, reflecting the altered pattern of loading in the meniscus-deficient joint. Changes in
the tibial plateau were limited to the outer zone of the LTP, the region normally protected by the
meniscus in the intact joint. As cartilage changes were most severe in the tibial plateau and mild in

203.
the femoral condyles, there appears to be a poor regional correlation between cartilage and
subchondral bone thickening. It has been proposed350,506,618,658 that the local bone sclerosis
accompanying OA results in a stiffer, less compliant subchondral plate, thereby increasing
compressive forces on the overlying chondrocytes and accelerating cartilage degeneration. The lack
of a clear regional correlation in this study suggests different stresses (mechanical or biochemical)
are responsible for the changes observed in cartilage and subchondral bone. Similarly, the lack of
subchondral bone sclerosis in the medial compartment, despite OA-like changes in the overlying
cartilage, shows that subchondral bone changes are not necessary for the progression of cartilage
lesions. A similar lack of correlation has been shown in the canine ACLT model.90,179,180
Changes in peri-articular bone mineral density were more consistent, and showed the same pattern
of increased BMD in the lateral compartment and relative osteopenia in the medial side of the joint.
This again reflects the changes in joint kinetics following meniscectomy, whereby external femoral
rotation and slight valgus displacement of the stifle increased loading in the lateral compartment.
Early loss of subchondral bone volume and density is common in surgical animal models of OA,
due to decreased loading of the joint.179,540,614 The use of a bilateral meniscectomy model
theoretically prevents unloading due to shift of weight to the the contralateral limb. However, the
loss of BMD from femoral head and neck sites (with a similar trend in the femoral and tibial
diaphyses) suggests that some compensatory gait change does occur. The pattern of changes in this
trial suggests that subchondral bone changes in this and other surgical models of OA are driven
largely by changes in joint loading, rather than being directly related to cartilage pathology.
However, the subchondral bone changes following meniscectomy may still be useful in the study of
the bone changes accompanying OA, as demonstrated by the modified bone responses seen in trials
of putative DMOADs.104,368
The observation that BMD changes were more prominent than altered SCP thickness is interesting,
as the classic concept of OA subchondral bone involves a sclerotic but hypomineralised matrix.39,293

204.
Karvonen et al. (1998)403 used a similar DEXA method to show a significant decrease in
periarticular BMD in patients with early knee OA. Results in this model show that in some regions
(e.g. LTP) a significant increase in subchondral BMD occurs, accompanied by only minor
thickenening of the subchondral plate. Increased subchondral bone density is seen in other animal
models of OA.54,89,659
Tidemark alterations and neovascularisation of the calcified cartilage layer were prominent
following meniscectomy in this trial. Possibly due to the advanced age of the subjects, multiple
tidemarks were present in nearly all sheep, including control animals. However, blood vessels were
seen to invade the calcified cartilage in greater number and to a greater depth in meniscectomised
animals, especially in the lateral joint compartment. Armstrong (1993)39 and Simpson (1994)760
found that calcified cartilage changes were not a distinct feature of the sheep meniscectomy model.
This discrepancy may be due to differences in responses to medial and lateral meniscectomy, the
age of the subjects, or (more likely) to differences between bilateral and unilateral surgery and the
loading-driven subchondral bone remodelling subsequently induced.
Femoro-tibial bone mineral density measurements revealed a distinct difference between the left
and right stifles. Left joints showed significantly higher BMD, and this mirrored a greater
prevalence of cartilage lesions and osteophytes in the left leg of unoperated sheep. The same trend
was found in studies by Armstrong.39,41 This suggests that the left hindleg bears greater forces
during normal activity than the right. This might be due to the uneven abdominal distribution of the
rumen, or due to preferential usage of a 'leading' leg. Normal dogs show as much as 4% difference
in bone morphologic parameters between the left and right hind legs.179
Serum osteocalcin levels were significantly increased in meniscectomised sheep. This suggests an
increase in bone formation (or bone turnover) in these animals, and was seen despite significant net
loss of bone density at several extra-articular sites (L1, femoral head and neck). Stress is unlikely to

205.
have caused this elevation, as it has been shown in sheep that while short-term stresses such as
handling or transport are unlikely to influence osteocalcin levels, long-term corticosteroid
administration reduces plasma levels. Human studies have shown both elevated240,350 and
reduced11,779,798 osteocalcin levels in OA patients. A trend towards increased urinary excretion of
deoxypyridinoline (Figure 3.40) was also evident. Bone histomorphometric analysis failed to
demonstrate a significant difference in bone turnover in the area examined (central LTP), though a
trend towards increased calcein-labelled surface was apparent (Figure 3.33). Armstrong et al.
(1994)38 also failed to demonstrate any significant change in histomorphometric variables in peri-
articular trabecular bone following ovine meniscectomy, though an increase in osteoid volume and
tetracycline labelling were demonstrated in the subchondral cortex. This suggests that even
immediately adjacent to the joint, cancellous bone is shielded from altered focal forces following
meniscectomy, by thickening of the overlying subchondral plate.
The significant elevation in serum oestradiol levels seen in MX animals at the time of sacrifice is of
great interest. While a link between circulating oestradiol levels and human OA has frequently been
postulated, multiple studies have failed to conclusively establish a link.226,578,778,784 However, Tsai et
al. (1992)838 found that synovial fluid oestradiol levels are elevated in OA patients when compared
to normal postmenopausal women, to levels proportionate with OA severity. This finding suggests
that local production of oestradiol within OA joints may represent a potential source of circulating
hormone. Appreciable levels of 17ß-hydroxysteroid dehydrogenase activity have been shown in
rabbit cartilage.68 The results of this trial demonstrate that elevation in serum oestradiol can occur
secondary to an induced OA-like process. Alternatively, stress may have influenced oestradiol
synthesis. Acute stress raises serum oestradiol levels in intact female rats.756 However in ewes,
stress (such as that from transport) has generally been shown to suppress fertility and follicular
oestradiol production, by interrupting LH secretion.200 The observation of a similar rise (not
statistically significant) in serum oestradiol of OVX+MX sheep suggests that if stress was a factor,
extra-ovarian sources of oestradiol must be involved. In ewes, a considerable proportion of

206.
circulating oestradiol is not of ovarian origin but comes from both adrenal secretion and peripheral
aromatization of weak oestrogen precursors.127 However, the ovine adrenal mainly produces weak
oestrogen precursors, and this production is suppressed by glucocorticoids (N. Adams, personal
communication). ACTH administration does not acutely raise serum oestradiol levels in
ovariectomised pigs.564 Other peripheral sources may therefore be more likely, and it is possible that
the meniscectomised joint itself may be involved.
3.4.2 EFFECT OF OVARIECTOMY ON FEMORO-TIBIAL JOINT TISSUES OF
AGED EWES
Hypothesis: that ovariectomy of aged ewes would induce structural and
metabolic alterations in femoro-tibial articular cartilage, as well as synovial and
subchondral bone tissue.
3.4.2.1 Cartilage Changes After Ovariectomy
The results of this study provide strong evidence that ovariectomy induces significant structural
changes in ovine articular cartilage and subchondral bone, as recently suggested by other
authors.367,611,769,841 Ovariectomised sheep were found to have thinner cartilage in the lateral joint
compartment, with a generalised reduction in tibial phase lag ( ), and increased shear modulus (G*)
in the inner regions of the LTP (Figure 3.17). While no changes in gross appearance were evident,
histological examination showed a deterioration of structure and loss of proteoglycan, as confirmed
by the loss of toluidine blue staining intensity in the femoral condyles and patella. Patterns of
collagen birefringence suggest that collagen structure was also altered, particularly in the superficial
zone.
Turner et al. (1997)841 demonstrated that the biomechanical integrity of cartilage was adversely
affected by ovariectomy in aged ewes, as evidenced by decreased aggregate modulus (compressive

207.
stiffness) and shear modulus, but not cartilage thickness or permeability. Studies in young rabbits667
and rats597,918,923 have shown an increase in cartilage thickness following ovariectomy, though due to
species and age differences the latter findings may not be applicable to the current study. In this
trial, femoro-tibial articular cartilage of ovariectomised animals was found to be thinner, especially
in the lateral compartment. In the absence of gross or microscopic erosion, cartilage thinning may
be due to two processes. Firstly, net reduction in matrix production (as suggested by the reduction
in toluidine blue staining of femoral cartilage) may have produced an ‘atrophic’ cartilage state.
Secondly, advancement of the tidemark may have lead to endochondral ossification of the lower
layers of articular cartilage. The increase in calcified cartilage neovascularisation certainly suggests
active remodelling of this layer, as shown by Hwa et al. (2001)367 in a related trial. However, the
simultaneous reduction in subchondral bone thickness shows that the net result of osteochondral
remodelling processes is a thinner subchondral plate. While not measured separately, the calcified
cartilage layer appeared to be of similar thickness to that of control sheep.
While cartilage compressive stiffness is proportional to proteoglycan content,47 resistance to
dynamic biomechanical properties such as resistance to shear (G*) may correlate more with
collagen content,52,455,935 and in this study G* shows a strong inverse relationship with thickness
(Figure 3.17). Appleyard et al. (submitted for publication) have examined topographical variation in
biomechanical and biochemical variables across the ovine tibial plateaux. Cartilage thickness was
found to be strongly negatively correlated with collagen content and shear modulus.
Glycosaminoglycan content was found to be positively correlated with thickness, but negatively
correlated to G* in unoperated sheep. The thinner, outer zones of the tibial plateaux therefore have
more collagen and less proteoglycan on a dry weight basis. This increased collagen content may
account for much of the increase in shear modulus in these areas, despite correction for thickness
according to the theoretical variation in as published by Hayes et al. (1972).331 Zhu et. al.
(1993)935 found a similar correlation between G* and collagen content in bovine cartilage discs.
Similarly, Le Roux et al. (2000)455 correlated G* with surface collagen organisation (birefringence)

208.
in canine cartilage, and concluded that collagen governs the dynamic (shear) behaviour of cartilage.
The increase in G* in the middle and inner LTP of ovariectomised animals is therefore likely to
reflect an increase in collagen content, in association with the decrease in cartilage thickness also
observed.
The reduction in cartilage phase lag seen in ovariectomised animals might also reflect collagen
changes. Lower phase values mark a change in viscoelastic behaviour towards a more elastic solid,
and reduced dissipation of shear forces. The relationship between the structural components of
cartilage and its phase properties is complex, but simplistically the collagen network provides the
elastic response while proteoglycan provides the viscous response. Zhu et al. (1993)935 found a
weak positive correlation between and uronic acid content. However, more complex modelling
recognises the importance of energy dissipation from both proteoglycan-collagen and proteoglycan-
proteoglycan interactions. Phase properties are therefore influenced by the quantity and molecular
size of proteoglycans, and the degree to which they form stable aggregates, as well as intactness of
the collagen network. Therefore, the reduction in phase lag in the OVX group may be due to either
reduced cartilage proteoglycan content, some change in proteoglycan quality or aggregation state,
and/or an increase in collagen content or organisation. Analysis of collagen birefringence (Figures
3.27 - 3.29) shows an increase in superficial birefringence in the LFC and middle and outer zones of
the LTP, with a similar trend in other regions. This again suggests an increase in collagen density
and/or organization in OVX animals, specifically localised to the superficial layer of cartilage.
Ovariectomy is known to induce changes in collagen structure in various tissues. For example,
ovariectomy of adult rats increases collagen content of the temporomandilar joint disc,4 and
reduces mean fibril diameter of type I collagen in bone and skin397 The altered birefringence
patterns observed here, together with regional alterations in and G*, are highly suggestive of
similar structural reorganisation of cartilage collagen following ovariectomy of sheep.

209.
This study confirm suggestions from a recent related study611,769 showing altered cartilage
proteoglycan synthesis following ovariectomy in sheep. Proteoglycan synthesis was elevated in
patellar cartilage explants from ovariectomised animals (Figure 3.41). However, greater induction
of latent MMPs was seen following IL-1ß stimulation (Figure 3.43), suggesting that proteoglycan
catabolism may also have been up-regulated. Quantitation of toluidine blue staining showed a net
loss of proteoglycan content in both patellar and femoral condyle articular cartilage (Figures 3.24
and 3.25). This suggests that despite an increase in proteoglycan turnover, net loss of proteoglycan
had occurred by six months post-ovariectomy. Ovariectomy induces cellular proliferation and
increased matrix synthesis in growth plate cartilage in adult rats, a phenomenon thought to relate to
a concurrent increase in serum IGF-1.811 In a previous similar trial, cultured patellar cartilage
explants from ovariectomised sheep showed decreased 35SO4-incorporation at three months post-
surgery.769 The reasons for these different results are unknown, but they suggest that chondrocytes
undergo a prolonged period of fluctuating metabolic disturbance following oestrogen withdrawal.
Patellar cartilage explants from ovariectomised sheep produced significantly more nitric oxide
compared to entire animals (Figure 3.41). This result is unexpected as oestradiol is supportive of
constitutive nitric oxide production in most tissues.36,330,884,890 However, Kauser et al. (1997)406
demonstrated that ovariectomised rats experience a greater increase in plasma nitrate levels
following endotoxin treatment than entire animals, and that this increase could be attenuated by pre-
treatment with oestradiol.406 This suggests that oestrogen may normally function to suppress iNOS
induction. As nitric oxide is thought to decrease proteoglycan content561,801,822 and alter collagen
synthesis,110 increased cartilage iNOS expression following ovariectomy may be a possible
mechanism for some of the cartilage changes observed in this study. Yoon et al. (2001)925 found a
similar increase in NOS expression in rabbit clitoris and vagina following ovariectomy, and this
was associated with an increase in collagen content in these tissues.

210.
Some debate exists in the literature as to whether the effects of the female menopause are created by
the absolute loss of oestrogen, or are due to fluctuations in the oestrogen/progesterone balance in
the perimenopausal period.782 Serial assay of plasma 17ß-oestradiol in this study suggests a rapid
and permanent decline in oestradiol levels following ovariectomy. Resting plasma oestradiol levels
are normally very low in sheep, with a peak at oestrus.103 Serum concentrations of oestradiol in
women are much higher - reported levels include 74.1±59.1 pg/ml in premenopausal women,796
53.4±31.0 pg/ml in the perimenopausal period,796 and 4.1 pg/ml in postmenopausal women.120 The
presence of metabolic abnormalities in cartilage, synovium and subchondral bone (elevated
urokinase activity) tissues at six months post-ovariectomy confirms that the metabolic influence of
ovariectomy is prolonged, and therefore more likely relates to long-term hormone withdrawal,
rather than a temporary ‘perimenopausal’ imbalance. Analysis of serum oestradiol concentrations
(Figure 3.38) suggests that it is largely the loss of the cyclical peaks which is responsible for the
drop in average concentration, rather than a decline in the (already low) resting levels. This suggests
that, at least in sheep, it is loss of the normal cyclical rises in oestrogen levels that disrupts cartilage
homeostasis. As the oestrus cycles of women and sheep are temporally similar, the same may be
true of menopausal women.
3.4.2.2 Synovial Changes After Ovariectomy
Ovariectomy had little effect on cultured synovial fibroblasts. A slight but not statistically
significant increase in MMP synthesis was observed (Figure 3.49). A similar increase in gelatinase
synthesis was demonstrated by gelatin zymography in a previous study of ovariectomised ewes (see
Appendix 1). Oestrogen and MMP synthesis are potentially linked via the common involvement of
the AP-1 promotor element.420,620 Oestrogen receptor levels were found to be very low (in fact
barely detectable) in cultured synovial fibroblasts, though levels were slightly higher in cells
derived from in ovariectomised or meniscectomised sheep (Figure 3.55). A similar modest increase
in oestrogen receptor expression was seen following ovariectomy in a previous trial, using a whole

211.
cell uptake method (see Appendix 1). Smith et al. (2000)775 recently reported upregulation of
synovial oestrogen receptors after meniscectomy in sheep, using both a competitive binding assay
and immunohistochemistry.
While having little effect on synovial fibroblasts, ovariectomy was found to significantly alter
cytokine release by primary synovial isolates. The increase in IL-6 bioactivity in ovariectomised
cultures seems contrary to the report by Kawasaki et al. (2000)411 showing that exogenous
oestradiol increased IL-6 production by rheumatoid synovial fibroblasts. However IL-6 activity was
generally very low. IL-1 bioactivity was reduced in cultures from ovariectomised sheep. This is
consistent with a report that oestrogen (over the range of 1-100 ng/ml) induces a dose-dependant
increase in IL-1 secretion by rat peritoneal macrophages, whilst ovariectomy leads to a reduction in
IL-1 synthesis reversible by oestrogen replacement.361 The greater response to ovariectomy in
primary over passaged synovial cultures is consistent with studies by Cutolo and co-
workers,161,163,165 suggesting that synovial macrophages are a more important target for modulation
by oestrogen than fibroblastic cells.
3.4.2.3 Bone Changes After Ovariectomy
The findings in this study confirm those of a recent report by Hwa et al. (2001)367 showing that
ovariectomy results in active remodelling of the subchondral bone region, with net loss of BMD and
subchondral plate thickness, and increased neovascularisation of the calcified cartilage layer. This
study shows loss of subchondral bone mineral density (MTP and LTP) and subchondral plate
thickness (LTP and LFC only). Loss of bone mass is of course expected as part of a generalised
skeletal loss secondary to oestrogen withdrawal, as is well characterised in the ovine ovariectomy
model.581,840,844,845 In this trial ovariectomy lead to reduced BMD at all extra-articular sites
measured, though this loss was statistically significantly only in first lumbar vertebrae, a site known
to be useful for measuring post-ovariectomy BMD responses in sheep.844 Lumbar vertebral sites are

212.
important sites for assessing postmenopausal bone loss in women; for example, lumbar BMD was
12% lower versus age-matched control subjects in one study.592 Turner et al. (1995)840 has shown
bone responses to ovariectomy occur slowly in sheep, and only early changes in bone mass are
observed six months post-operatively. In addition, seasonal variation581 and the effects of stress845
may mask the responses observed.
Other measured parameters failed to show a significant disturbance in bone turnover following
ovariectomy. Serum osteocalcin and urinary collagen crosslink levels were not significantly
increased by ovariectomy alone in this trial, as has been shown in other studies.127,387 Similarly,
ovariectomy induced no detectable changes in cancellous bone histomorphometric parameters.
Other ovine studies have reported a lack of cancellous bone responses to ovariectomy within a six
month experimental period.127,840,845 It is likely that a longer experimental period and larger sample
size would be required to demonstrate significant alterations in these parameters.
A significant increase in the number and depth of blood vessel incursions into the calcified cartilage
layer was observed following ovariectomy. This may be related to the general remodelling
processes and altered stresses across this layer, or may result from specific biochemical stimuli such
as cytokine or growth factor release. Several lines of evidence suggest a link between ovariectomy
and angiogenesis. Firstly, elevated levels of the gonadotropins luteinising hormone (LH) and
follicle-stimulating hormone (FSH) following menopause or ovariectomy have been shown to
promote angiogenesis in ovarian carcinomas, possibly by increasing VEGF expression.731 Secondly,
ovariectomy influences the IGF-1 axis,227,811 while IGF-1 also induces VEGF expression in many
cell types, including tumour cells,10,72 retinal pigment cells,655 and osteoblastic cell lines.269 Thirdly,
nitric oxide may again be involved, as it is also thought to promote angiogenesis via interactions
with growth factors.937

213.
3.4.3 EFFECT OF PRIOR OVARIECTOMY ON THE DEVELOPMENT OF
FEMORO-TIBIAL JOINT LESIONS AFTER MENISCECTOMY OF AGED
EWES
Hypothesis: that prior ovariectomy of ewes would worsen and/or accelerate the
progression of joint changes following meniscectomy.
3.4.3.1 Cartilage Changes After Ovariectomy and Meniscectomy
The development of cartilage pathology post-meniscectomy was found to be significantly modified
by prior ovariectomy, providing further evidence of the importance of ovarian hormones in
maintaining cartilage integrity. However, the variety of parameters measured in this study provide
conflicting information as to whether cartilage integrity was worsened or improved following
ovariectomy. On one hand, OVX+MX sheep had significantly thicker uncalcified cartilage at all
points measured, except the inner zones of the MFC (Figures 3.22 and 3.23). This difference in
thickness was particularly prominent in areas subject to erosion following meniscectomy,
suggesting that ovariectomised sheep were partially protected from fibrillation and loss of cartilage.
Lower histopathology scores, and the lack of a significant increase in chondrocyte density in the
femoral condyles, also suggest that ovariectomised animals showed greater structural integrity of
cartilage following meniscectomy.
On the other hand, the increase in cartilage thickness in uneroded areas is less clearly a beneficial
change, as it may represent increased swelling (i.e. water content) secondary to collagen damage,
rather than an anti-catabolic effect. There is evidence that OVX+MX sheep possessed
biomechanically inferior cartilage, with greater loss of dynamic stiffness in the tibial plateaux. Loss
of proteoglycan (toluidine blue staining) was also significantly greater in the LFC (Figure 3.24),
with a similar trend in other regions. This suggests that the cartilage of OVX+MX sheep, while

214.
thicker, was structurally inferior and does indeed suggest an increase in hydration. As discussed
above, explant culture of patellar cartilage was not particularly useful and did not resolve this issue.
It is worth noting that prior ovariectomy increased cartilage nitric oxide release as it did in non-
meniscectomised animals, though this difference was not statistically significant.
Quantitative analysis of cartilage changes in this OA model therefore suggests that while thicker,
cartilage of OVX+MX sheep showed greater loss of proteoglycan content, and greater disruption of
collagen integrity as suggested by the greater decrease in G*. Increased post-meniscectomy loss of
cartilage proteoglycan has also been shown in a previous trial of ovariectomised ewes, and further
this was reversed by transdermal 17ß-oestradiol supplementation (Parker et al., in press). This
evidence appears consistent with the known association between human OA and the female
menopause,578 and the protective effect of HRT.226,758 Numerous mechanisms might explain the
influence of ovariectomy on the OA process, including disturbance of growth factor levels,227,811
nitric oxide synthesis,406,906, cytokine networks,161,691 and subchondral bone responses. However,
greater histopathologic severity of erosive lesions was not demonstrated in OVX+MX sheep.
Chambers et al. (2001)122 similarly found that neither ovariectomy or orchietomy had any effect on
the development of lesions or aggrecan cleavage in the STR/ort mouse model of spontaneous OA. It
was therefore not possible to confirm the hypothesis that prior ovariectomy would worsen the
osteoarthritic lesions produced by meniscectomy in this model.
3.4.3.2 Synovial Changes After Ovariectomy and Meniscectomy
In vitro, synovial fibroblasts from OVX+MX sheep showed significantly greater production of
latent MMPs relative to MX under both resting and IL-1ß-stimulated conditions (Figure 3.49), as
well as increased IL-1ß-stimulated urokinase activity (Figure 3.50). This confirms the trend towards
greater MMP production as was seen in OVX cultures. However, synovial fibroblast NO production

215.
was lower relative to that of MX sheep (Figure 3.47). This result must be interpreted in conjunction
with knowledge that IL-1ß stimulation significantly reduced in vitro NO production in these cells.
3.4.3.3 Bone Changes After Ovariectomy and Meniscectomy
While ovariectomy induced subchondral bone loss in normal sheep, it had no significant effect on
subchondral bone thickness changes secondary to meniscectomy. Similarly subchondral BMD as
measured in intact bones did not differ, though 'cubic' BMD measured in tibial bone slabs (Figure
3.36) was lower in OVX+MX animals - the latter technique may be more sensitive to trabecular
rather than cortical bone changes. The comparative lack of post-ovariectomy bone changes in
meniscectomised relative to unoperated sheep suggests that different processes are responsible for
bone remodelling in normal and arthritic joints, and that bone responses to the dual stimuli of
altered force distribution and/or cytokine stimulation following meniscectomy are largely
independent of oestrogen. This is consistent with the findings of Fazzalari et al. (2001),219 who
showed that a different set of mediators may control turnover in OA bone compared to normal
tissue. However, calcified cartilage neovascularisation and osteophyte growth were significantly
greater in OVX+MX joints. These changes may relate to alterations in levels of growth hormones
following ovariectomy as has been shown in other species.227,811
While subchondral bone was structurally similar, bone explant culture showed that prior
ovariectomy had a significant effect on the metabolic responses of tibial subchondral bone to
meniscectomy. OVX+MX but not MX explants showed increased production of PGE2 (Figure 3.45)
and urokinase (Figure 3.46) relative to control cultures. Both of these mediators are thought to be
important modifiers of bone metabolism. Urokinase is thought to be important in bone resorption,695
while PGE2 has varied effects but is also predominantly associated with bone loss.24 Hilal et al.
(1998)350 used an identical bone explant culture system to demonstrate that tibial bone from OA
patients released greater levels of urokinase and IGF-1 compared to normal specimens. However,

216.
the OA samples in this study were derived mainly from aged women (3 men, 6 women), while
control samples were predominantly sourced from men (4 men, 1 woman). Massicotte et al.
(2000)515 recently showed that different levels of cytokine production by osteoblasts discriminate
two subgroups of OA patients, with a group termed ‘high OA Ob responders’ showing greatly
increased IL-6 and PGE2 production. PGE2 production was partially responsible for this IL-6
increase as it could be blocked by NSAIDs. Gender of subjects was not recorded in this abstract.
Results from ovine bone explants in this study suggest that oestrogen withdrawal has an important
influence on the metabolic responses of subchondral bone in OA. If extrapolated to human OA, the
female menopause may have a similar effect and should be considered as a variable in such studies.
3.4.4 EFFECT OF TOPICAL GLYCERYL TRINITRATE ON FEMORO-TIBIAL
JOINT TISSUES IN NORMAL AND OVARIECTOMISED SHEEP
Hypothesis: that the topical use of an exogenous nitric oxide donor, glyceryl
trinitrate (GTN), would reduce the loss of bone mineral density following
ovariectomy in aged ewes, as has been shown in other species.902,904,906
Hypothesis: that the same dose of GTN which produces the above skeletal effects
would adversely affect the structural and metabolic integrity of normal articular
cartilage, in keeping with the known degradative effects of NO on
chondrocytes.78,140,322,802,803,822
Hypothesis: given the net effect of NO varies in physiological and pathological
conditions381,552
and NO is a key mediator of oestrogen
activity,36,329,330,406,666,676,884,890,906
that the response of joint tissues to GTN
treatment would vary between normal animals, ovariectomised animals, and
animals with surgically-induced OA.

217.
The results of this study reveal that topical application of glyceryl trinitrate to normal sheep for six
months caused significant changes to the structure and biomechanical properties of articular
cartilage in the femoro-tibial joint. Uncalcified cartilage of GTN-treated animals was found to be
thinner, by both direct (needle penetration) (Figure 3.17) and histomorphometric assessment
(Figures 3.20 and 3.21), in all regions tested. The changes in biomechanical properties of
NOC+GTN sheep paralleled that of OVX animals, with a similar decrease in phase lag, and
increased G* in the inner zones of the LTP (Figure 3.17). As with the changes observed following
ovariectomy (see 3.4.2.1), these biomechanical changes suggest thinner cartilage with an increase in
collagen content or organisation, and/or a reduction in proteoglycan content or the viscous qualities
(aggregation) of proteoglycan. Superficial collagen birefringence was increased in the LFC and
middle zone of the LTP, with similar changes other areas (Figures 3.27 – 3.29), confirming
reorganization of cartilage collagen. The lower intensity of toluidine blue staining observed in the
MFC (Figure 3.24) shows that proteoglycan content was also reduced by GTN treatment. Again,
these changes suggest more an ‘atrophic’ than a degenerative effect, and were not accompanied by
any detectable gross or histopathological alterations (except neovascularisation of calcified cartilage
layer). In experimental animal models, osteoarthritis is generally associated with reduced cartilage
shear modulus and increased phase lag, which is the opposite of that observed here.33,745
Several of the known actions of nitric oxide may explain the effects observed. Firstly, NO has been
shown to have direct effects on chondrocyte metabolism, including suppression of proteoglycan
synthesis, increasing MMP synthesis, and decreasing the level of (and response to) endogenous
growth factors.349,561,801 The changes observed might be consistent with prolonged suppression of
chondrocyte proteoglycan synthesis, or increased loss of proteoglycan due to elevated MMP
activity. Secondly, nitric oxide is known to influence collagen synthesis in many cell types. NO
increases collagen synthesis in most connective tissues925 and dermal fibroblasts in vitro909, and is
generally thought to promote collagen production during wound healing.562,726 However, NO has
been shown to decrease collagen production in other tissues, including mouse artery585 and rabbit

218.
cruciate ligament.108 In the only published study to examine its effect on chondrocytes, Cao et al.
(1997)110 showed that NO reduced type II collagen production by lapine chondrocytes in monolayer
culture, an effect attributed to inhibition of the key enzyme prolyl-hydroxylase. Changes in collagen
birefringence observed in this study (Figure 3.27 – 3.29) suggest that GTN increased collagen
organisation in the superficial zone of articular cartilage in certain areas, though the collagen type
involved was not identified. GTN treatment had no significant effect on chondrocyte density in
femoral condylar cartilage (Table 3.4). This result suggests that the pro-apoptotic effects of nitric
oxide found by some researchers78,321,481 were not operative in this model.
Alternatively, GTN may have influenced articular cartilage via its effects on subchondral bone. NO
is known to affect bone metabolism, its dominant role in vitro being tonic inhibition of osteoclastic
bone resorption.211 GTN-treated sheep showed a trend towards increased thickness of the
subchondral bone plate (statistically significant in the MTP) and greater subchondral BMD
(significant in the LFC). Concurrent increase in calcified cartilage neovascularisation provides
evidence of active subchondral plate remodelling. Given the hypothesis by some authors that
subchondral bone sclerosis may be central to the early pathogenesis of OA,659 and that much
experimental and epidemiological evidence associates OA with increased bone density,184,580 the
sclerosing effects of GTN treatment on subchondral bone might have induced changes in the
overlying cartilage. However, as discussed in 3.4.1.3, a clear association between subchondral
sclerosis and cartilage degradation was absent in this study.
In ovariectomised animals, GTN treatment appeared to have an opposite effect, apparently
reversing of the effects of ovariectomy on cartilage thickness and phase lag (Figure 3.17). It is not
possible from the variables measured to determine whether this represents a protective effect, or a
worsened degradative effect from the combined treatments. The thicker cartilage in OVX+GTN
animals may be closer to the normal state, or might reflect cartilage swelling secondary to
disruption of the collagen network. Similarly, the increase in might represent a more normal state,

219.
or be due to increased energy dissipation secondary to loosening of the proteoglycan-collagen
matrix.745 Importantly, was significantly elevated in the outer zones of the LTP in OVX+GTN
animals, relative to NOC as well as to OVX sheep (Figure 3.17). Trypsin degradation of ovine
cartilage significantly increases phase lag (Appleyard et al., submitted for publication). An increase
in cartilage thickness and phase is also seen in animal models of OA.34,746 While OVX+GTN
cartilage did not show the increase in surface birefringence seen in OVX and NOC+GTN animals,
birefringence plots were significantly altered relative to controls, especially in the outer LTP where
a reduced surface peak and elevated intermediate zone bifringence suggested similar collagen
disruption to that seen in OA (Appleyard et al., unpublished data). Such cartilage disruption,
combined with loss of proteoglycan as seen in the femoral condyles (which was not reversed by
GTN treatment) (Figure 3.24), might reflect a more severe structural degeneration of cartilage
compared to OVX alone.
However, there is some evidence that GTN treatment did indeed partially reverse the changes
associated with ovariectomy. Cartilage histopathological scores were not increased in the MFC as
was seen in OVX animals (Table 3.2). The increase in patellar explant nitric oxide release seen in
OVX animals was significantly reduced by GTN treatment (Figure 3.41), though 35SO4-labelled
proteoglycan synthesis and MMP activity were not affected. There are several concievable
mechanisms by which GTN might exert a protective effect after ovariectomy. Firstly, ovariectomy
has been shown to increase IL-1ß activity in certain tissues.161,361,605 If this mechanism were
operating in cartilage, GTN may have an early protective effect similar to that attributed to NO in
some studies of IL-1ß-stimulated cartilage.327,793,794 Secondly, ovariectomy leads to disturbance of
the IGF-1 axis, in a manner that is not entirely clear and may vary between species.111,227,811,868 If
ovariectomy disrupts cartilage by inducing cartilage growth factors as is seen in rat growth plate
cartilage,811 GTN may be protective, as NO is thought to blunt chondrocyte responses to IGF-1 and
TGF-ß.801,803 In this sense, NO may act as a non-specific ‘brake’ on both anabolic and catabolic
responses.140

220.
Alternatively, the primary influence of ovariectomy may be in its effect on subchondral bone. In
this study ovariectomised animals were found to have reduced BMD (MTP and LTP) (Figure 3.35)
and a thinner subchondral bone plate (LFC and LTP) (Figures 3.20 and 3.21) compared to entire
controls, as well as a neovascularisation response in the calcified cartilage layer (Figure 3.19). GTN
treatment significantly attenuated some of these changes, increasing subchondral plate thickness in
the LFC and LTP. By preventing subchondral bone loss, GTN may have a ‘stabilising’ effect on
joint tissues in ovariectomised sheep. Similarly, a recent study in dogs attributed chondroprotective
properties to calcitonin, another agent preventing bone resorption.496 However, subchondral BMD
loss and neovascularisation responses were not prevented by GTN treatment. Rather, GTN
treatment (in common with ovariectomy) was found to significantly promote neovascularisation of
calcified cartilage. This is consistent with the proposed role of NO in both VEGF and FGF-2-
related angiogenesis.247,937
Ovariectomy-associated bone loss is well described in sheep.581,840,843 Wimalawansa and coworkers
have demonstrated that topical glyceryl trinitrate attenuates both prednisolone- and ovariectomy-
induced bone loss in rats.904,906 Early results of clinical trials suggest the same effect may occur in
women,902 as does the epidemiological observation that intermittent use of nitrates is associated
with increased bone mineral density in the hip and heel in elderly women.380 The variables
measured in this trial failed to demonstrate a significant effect by topical GTN application on
skeletal bone density. A trend towards reversal of ovariectomy-associated bone loss was seen in the
tibial and femoral diaphyses, though neither the effect of GTN or the effect of ovariectomy was
statistically significant. As discussed above (see 3.4.2.3), ovariectomy failed to induce significant
changes in other indices of bone turnover. It was therefore not possible to demonstrate any reversal
of such changes following GTN treatment. However, the reduction in trabecular eroded surface
seen in NOC+GTN relative to NOC sheep (Figure 3.31) is consistent with the proposed skeletal
effects of GTN. The lack of change in serum osteocalcin is also consistent with both the human902

221.
and rat903 studies of Wimalawansa, which found that GTN treatment failed to negate ovariectomy-
induced increases in serum osteocalcin, suggesting it may have a positive effect on bone formation
separate to any effect on osteoclasts. It is likely that a larger group size, and/or greater time period,
would be required to show consistent effect by GTN on bone indices in this sheep model.
Alternatively, the frequency and dose of GTN treatment may have been insufficient. Wimalawansa
et al. (2000)905 demonstrated the importance of dose frequency in their rat model, finding that while
once daily treatment prevented bone loss in ovariectomised rats, twice- or thrice-daily treatment
was ineffective. Efficacy trials in humans and rats have used once-daily treatment.902,904,906 The
GTN dosage used in this study was limited for practical reasons to twice weekly application. While
this dose of GTN was demonstrated to have significant effects on cartilage structure, and to
regionally increase subchondral bone plate thickness in both control and ovariectomised animals, a
consistent effect on bone mass was not demonstrated in this study.
3.4.5 EFFECT OF TOPICAL GLYCERYL TRINITRATE ON THE
DEVELOPMENT OF FEMORO-TIBIAL JOINT LESIONS AFTER OVINE
MENISCECTOMY
Hypothesis: given nitric oxide synthase inhibitors reduce the progression of
experimental arthritis,525,619,792
that GTN treatment would worsen and/or
accelerate the progression of joint changes in the ovine meniscectomy model of
OA.
Treatment with glyceryl trinitrate was found to significantly affect the structural alterations seen in
cartilage and bone following meniscectomy. While there was no grossly visible difference in
cartilage pathology, other measures suggest that the cartilage of GTN-treated animals was degraded
to a greater degree compared to untreated animals. MX+GTN animals showed thinner cartilage at
central testing points in the LTP by the needle penetration method (Figure 3.18), though this was
not confirmed by histomorphometric measures. This suggests greater severity of articular cartilage

222.
erosion. The cartilage was biomechanically inferior, with an overall increase in phase lag, and a
greater reduction in G* in the outer zones of the LTP. Further, histopathological scoring revealed
greater structural degeneration and loss of proteoglycan in the MFC, and a greater degree of
chondrocyte cloning in the LFC. Together, these results suggest that topical treatment with glyceryl
trinitrate worsened cartilage degeneration following lateral meniscectomy.
Glyceryl trinitrate particularly influenced the subchondral bone remodelling processes associated
with meniscectomy. GTN-treated animals showed a greater increase in subchondral bone plate
thickness in the middle (in-contact) zone of the LFC, and reduced loss of thickness in the outer zone
of the MFC, relative to MX sheep (Figure 3.22). Similarly, treated animals showed a greater
increase in subchondral bone density in the LFC and LTP, and less loss of BMD in the MTP
(Figure 3.35). Induction of endogenous NO production in bone is thought to be a protective
response against cytokine-stimulated bone resorption in pathological conditions.362,483,664,665 NO is
known to inhibit both osteoclast function,88,405,486 and osteoclast differentiation from precursor cell
types.88 GTN, as an exogenous donor of nitric oxide, appears to have had a similar bone-promoting
effect. It is interesting to note that the effects of GTN were more pronounced in the disturbed
subchondral bone of meniscectomised animals, compared to its slight influence on remodelling
following ovariectomy. This may again support the hypothesis of Fazzalari et al. (2001)219 that a
different set of mediators might predominate in the remodelling processes of OA subchondral bone,
compared to those operative during normal homeostatic turnover. It also suggests that endogenously
generated nitric oxide may be a prime mediator in the subchondral bone plate sclerosis that occurs
in OA joints.
GTN treatment also promoted osteophytic remodelling, increasing the cross-sectional area of
marginal osteophytes especially in the femoral condyles (Figure 3.26). Van den Berg et al. (1999)858
found reduced growth of osteophytes in iNOS-deficient mice, an effect possibly linked to TGF-ß
production. Serum osteocalcin levels were similar to NOC animals, and significantly lower than

223.
that of MX sheep. Urinary collagen crosslinks showed a similar trend, with GTN reversing a
meniscectomy-associated increase in deoxypyridinoline excretion (Figure 3.40), though this was not
statistically significant. This suggests that GTN treament attenuated the increase in bone turnover
observed following meniscectomy. In vitro, NO has been shown to reduce osteoblast proliferation,
activity, and viability.170,171,362,665 However, topical GTN treatment did not affect the ovariectomy-
induced increase in serum osteocalcin levels in human and rat trials.902,903
In the combined OVX+MX model, there is also some evidence that GTN-treated animals
experienced greater cartilage deterioration. The strong trend towards lower body weights in
OVX+MX+GTN sheep may be secondary to increased disability and joint pain, leading to altered
grazing behaviour, as was proposed in Chapter 2. This difference in body weight was unfortunately
a confounding factor on other variables, as cartilage thickness and bone mineral densities were
found to be positively correlated closely with body weight (data not shown). However,
OVX+MX+GTN animals showed a significant increase in phase lag not observed in OVX+MX
animals, as well as greater decrease in G* overall. As the dynamic biomechanical properties of
cartilage should be mostly independent of body weight, this suggests that the OVX+MX+GTN
group experienced greater cartilage degeneration relative to the OVX+MX control.

224.
CHAPTER 4
CONCLUSION AND SUMMARY
These doctoral studies make important contributions to the scientific literature in three main areas.
Firstly, understanding of the ovine meniscectomy model of osteoarthritis has been importantly
extended. Novel computerised image analysis methodologies were shown to provide improved
quantification of the structural changes affecting cartilage and subchondral bone in this model.
These methods offered distinct advantages compared to traditional qualitative or semi-quantitative
outcome measures, and were able to confirm subtle structural changes associated with various
interventions (ASU, GTN, OVX) where no gross changes were evident. Further, bilateral lateral
meniscectomy was shown to induce significant structural and metabolic changes in synovial and
subchondral bone tissues, in addition to the well-characterised cartilage changes. Both gross and
microscopic synovitis were confirmed to be a feature of this OA model, with an increase in intimal
hyperplasia, inflammatory cell infiltration, vascularity, and fibrosis demonstrated following
meniscectomy.
Meniscectomy was also found to induce a significant elevation in plasma oestradiol levels. The
observation that an increase in circulating oestradiol can occur secondary to an induced OA-like
process is potentially very important, given the suggestion by some authors68,838 that local
production of oestradiol within OA joints may represent a potential source of circulating hormone.
Further studies with this model should include analysis of both blood and synovial fluid levels of
oestradiol to further examine this possibility.
However, the in vitro responses of cultured ovine synoviocytes to both meniscectomy and IL-1ß-
stimulation were modest and showed some inconsistency with previous studies in other species. It

225.
seems likely that true species differences exist with regard to the in vitro responses of arthritic
synovial tissue. Alternatively, these results may reflect the inherent flaws of monolayer cell culture
as a model of investigating synovial tissue changes. Cultures of passaged synovial fibroblasts fail to
model the Type A/Type B synoviocyte interactions and cartilage/synovial interactions that are
likely to be important in the synovial responses in osteoarthritis. In addition, the extremely rapid in
vitro proliferation of intimal fibroblasts, a cell type which normally has a low mitotic rate, is highly
artifactual.337,339 Ex vivo methods such as immunohistochemistry or in situ hybridisation may have
yielded different results regarding the nature of synovial activation in this model.
Secondly, this study demonstrated significant changes in joint tissues secondary to oestrogen
withdrawal by surgical ovariectomy. Ovariectomised sheep were shown to have thinner articular
cartilage in some regions, with altered dynamic biomechanical properties related to reduction of
proteoglycan content and reorganisation of superficial collagen structure. Patellar cartilage explant
studies showed an increase in both proteoglycan synthesis and MMP activity, with a net loss of
proteoglycan. This confirms the findings of previous studies (see 1.1.4) showing that ovarian
hormones, principally oestrogen, are important regulators of chondrocyte metabolism and cartilage
homeostasis. Cartilage events may also have been influenced by subchondral bone processes
occurring simultaneously. While generalised skeletal bone loss was detected, bone remodelling was
particularly prominent in the subchondral region, with loss of subchondral plate thickness and
BMD. These changes were accompanied by neovascularization of the calcified cartilage layer as
described by Hwa et al. (2001).367 However despite regional thinning, cartilage of ovariectomised
sheep was not obviously biomechanically inferior to that of normal animals, and the changes
observed were more atrophic than osteoarthritic. More time may be required to observe any long-
term degenerative sequelae of these cartilage changes, though some early histopathological changes
were observed.

226.
Prior ovariectomy was also shown to significantly influence the development of joint changes
following meniscectomy. However, as the increase in cartilage thickness observed might be due to
reduced erosion, or due to increased disruption and swelling of cartilage, it was not possible to
conclude whether the cartilage degenerative process was improved or worsened by ovariectomy.
The greater biomechanical disruption and proteoglycan loss in some areas suggests that cartilage
degeneration was worsened in these animals. This is consistent with the observations that (i) prior
ovariectomy significantly increased synovial fibroblast MMP and urokinase activity following
meniscectomy; and (ii) prior ovariectomy significantly influenced the metabolic responses of
subchondral bone to meniscectomy, increasing urokinase and PGE2 release by bone explants.
However, prior ovariectomy had little effect on the structural bone alterations induced by
meniscectomy, though osteophyte growth and calcified cartilage neovascularisation were also more
pronounced in ovariectomised sheep.
Thirdly, this study is the first to show that the nitric oxide donor glyceryl trinitrate, a ubiquitous
human medication used for symptomatic treatment of angina pectoris, is capable of inducing
structural and biomechanical changes in ovine articular cartilage. Topical treatment of ewes with
GTN was found to produce cartilage changes similar to those induced by ovariectomy, with reduced
thickness and altered biomechanical properties of femoro-tibial articular cartilage. Regional loss of
cartilage proteoglycan, altered collagen distribution, and calcified cartilage neovascularisation were
observed. These findings are consistent with chronic derangement of chondrocyte metabolism, as
might be predicted from the known deleterious effects of nitric oxide in cartilage. GTN treatment
also induced calcified cartilage neovascularisation, and increased subchondral bone thickness and
BMD in some joint regions. If these findings were applied clinically, the safety of NO-donor
compounds may be questioned. Indeed, some epidemiological evidence of an association between
nitrate use and hip OA was recently presented.450 However, it should be noted that the cartilage
changes observed in this study after GTN treatment are not necessarily degenerative, since minimal

227.
evidence of cartilage pathology was observed. In experimental animal models, osteoarthritis is
generally associated with reduced cartilage shear modulus and increased phase lag, which is the
opposite of that observed here745.33,745 Additionally, ovine knee joint cartilage is less than half the
thickness of the equivalent human cartilage.47 Nevertheless, the current findings indicate the need
for further epidemiological investigation into the cartilage effects of glyceryl trinitrate and other
nitric oxide donors.
While the topical use of glyceryl trinitrate did not significantly reverse skeletal bone loss in
ovariectomised ewes as has been shown in rats906 and women,902 it did attenuate post-ovariectomy
thinning of the subchondral plate in some regions. In addition, GTN treatment increased
subchondral bone plate thickness and BMD during subchondral remodelling following
meniscectomy. GTN treatment was also found to slightly but significantly worsen cartilage
degeneration following meniscectomy. This suggests that clinical use of GTN may accelerate both
cartilage degeneration and subchondral bone sclerosis in patients with concurrent osteoarthritis.
In this combined trial, either ovariectomy or GTN treatment were found to induce very similar
structural and biomechanical changes in articular cartilage. The observation of increased in vitro
nitric oxide production by patellar cartilage from OVX sheep, presumably the result of iNOS
induction, suggests that this similarity may be due to the prime involvement of NO in both
processes. However, the apparent reversal of the effects of ovariectomy by GTN treatment suggests
otherwise. While there are plausible reasons why GTN might be protective in this case, this paradox
may in fact be an illusion created by greater structural deterioration of articular cartilage in the
combined OVX+GTN group. It was not possible to confirm whether articular cartilage was more
normal, or instead showed a greater degree of disruption with the combined treatment. Either way,
results confirm that ovarian status has the potential to influence cartilage responses to GTN
treatment. If extrapolated to human usage, these results suggest that oestrogen (i.e. menopausal)
status may influence cartilage responses to clinical nitric oxide donors.

228.
Further studies will be necessary to confirm some of the extended hypotheses in this study. In order
to confirm the biochemical basis of the cartilage changes observed, more direct biochemical assays
on cartilage tissue (e.g. collagen content (as hydroxyproline), proteoglycan content (as sulphated
glycosaminoglycan), water content) could be included in future trials. Immunohistochemical studies
might be used to confirm the induction of cartilage iNOS following ovariectomy, and to resolve the
exact nature of the association between ovariectomy and the IGF-1 axis in the sheep model.
Cartilage collagen changes should be investigated with particular reference to the collagen types
involved, as aberrant chondrocyte expression of types I and III collagen is noted in disease states
such as OA. Double immuno-staining of synovial tissue may confirm that macrophage-like synovial
cells are a more important target for oestrogen modulation than the (here relatively inert)
fibroblastic cells. In addition, this study intentionally used a fairly crude bone explant system to
examine changes in subchondral bone metabolism in an attempt to replicate the findings of Hilal et
al. (1998).350 To further these investigations, more sophisticated culture methods (e.g. osteoblast
culture) or immunohistological methods should be used in future studies.

229.
APPENDIX 1
SELECTED RESULTS FROM ANOTHER TRIAL OF
OVARIECTOMISED AND MENISCECTOMISED EWES
The following results were obtained from synovial fibroblast cultures derived from a previous,
related trial of ovariectomised and meniscectomised ewes conducted by the Institute of Bone and
Joint Research, University of Sydney.367,611,769,775 A number of aged (8-10 years old) Merino ewes
were used for this study. Six of these were ovariectomised by midline laparotomy (OVX), six were
subjected to bilateral lateral meniscectomy (MX), and six served as non-operated controls (NOC).
Animals were euthanased six months post-operatively. Synovial fibroblast cultures were derived
using identical techniques to those described in 3.2.7.3.
1. Synovial Fibroblasts – Oestrogen Receptors (Whole Cell Uptake Method)
Method: Oestrogen receptors were measured by a whole cell uptake method.208,704 Briefly, cells were cultured to confluence then incubated with 10nM [2,4,6,7-3H]17ß-oestradiol [Amersham, UK] in the presence or absence of a 100-fold excess of non-radioactive oestradiol. After incubation for 30 mins at 37°C, monolayers were gently washed three times with warm PBS, trypsinised, and harvested onto filter paper discs using a cell harvester. Specific binding was determined by subtracting non-specific from total binding, and expressed as molecules bound/cell.
NOC OVX MX0
2000
4000
6000
8000
10000
Specific bindings / cell

230.
2. Synovial Fibroblasts – MMP (Gelatinase) Levels
MMP-2 and -9 levels in conditioned media from confluent primary cultures, as determined by quantitative gelatin zymography391,424 (mean summed density of all relevant bands (as % of NOC mean) and percentage active form, n=4 per group) and corrected for cell number.
* differs significantly from NOC, p=0.027; † differs significantly from MX, p=0.019
GROUP
(n=4) Total MMP-2
(% of NOC) % active
MMP-2 Total MMP-9
(% of NOC) % active
MMP-9
NOC 100.0 ± 14.1 42.5% 100.0 ± 29.2 8.3% OVX 178.9 ± 95.6*† 33.0% 213.3 ± 157.1 5.5% MX 92.2 ± 17.0 41.8% 131.4 ± 125.8 7.7%
Method: Conditioned media were diluted 1:5 in dilution buffer (50mM Tris, 0.15M NaCl, pH 7.2) before mixing with an equal volume of Tris-glycine SDS PAGE application buffer (0.0625 M Tris-HCl, 10% glycerol(v/v), 2% SDS(w/v), pH 6.8 with 0.0025% (w/v) bromophenol blue) and incubated for 30 minutes at 37°C to allow formation of SDS-protein complexes. 20 l samples were loaded into 10% homogenous polyacrylamide gels containing 0.1% gelatin [Novex, USA]. A standard solution of MMP-2 (6U/ml) [Boehringer Mannheim, Germany; from human fibrosarcoma cells] and a molecular weight standard [SeeBlue, Novex, USA] was run in each gel. Electrophoresis [XCell II Mini-Cell, Novex, USA] was conducted at 125V for 120 minutes in Tris-glycine buffer (0.025M Tris-HCl, 0.192M glycine, 0.1% (w/v) SDS, pH 8.4).443 Following electrophoresis, SDS was removed during incubation in 2.5% Triton X-100 (30 minutes, room temperature), with gentle agitation on a platform rocker. Gels were then incubated for a further 30 minutes in 50mM Tris-HCl, 20% glycerol, pH 7.2 to promote protein renaturation. Digestion of the gelatin was allowed to proceed by incubation (18 hours, 37°C) in 50mM Tris-HCl, 100mM NaCl, 20mM CaCl2, pH 7.2. The gel was then stained for 1 hour in Coomasie R250 solution (0.1% (w/v) in 40% (v/v) ethanol and 10% (v/v) acetic acid) and subsequently destained in 10% (v/v) ethanol, 7.5% (v/v) acetic acid. To quantify the density of the digested bands, each gel was scanned at 300 dpi on a flat bed scanner. An inverted greyscale image was analysed with the gel plotting macro in NIH Image [NIH Image 1.62, National Institutes of Health, USA], using the areas under the density plot for each lane. Values were corrected for standard values and the results presented as percentage of control mean.
Sample gelatin zymogram.
Lane 1- MMP-2 control; Lane 2 - media blank; Lane 3,4 - NOC: Lane 5,6 - OVX
MMP-9
MMP-2
pro
active
pro
active
> > >
> > >
LANE 1 2 3 4 5 6

231.
REFERENCE LIST
1. Abgarowicz T, Koziorowska J. Cultures of cells lining the synovial membrane.
Reumatologia 1969; 7: 291-3.
2. Abrams J. Nitrate Delivery systems in perspective. A decade of progress. Am J Med 1984; 38-46.
3. Abramson SB. The role of COX-2 produced by cartilage in arthritis. Osteoarthritis Cartilage 1999; 7: 380-1.
4. Abubaker AO, Hebda PC, Gunsolley JN. Effects of sex hormones on protein and collagen content of the temporomandibular joint disc of the rat. J Oral Maxillofac Surg 1996; 54: 721-7.
5. Aeberhard EE, Henderson SA, Arabolos NS, Griscavage JM, Castro FE, Barrett CT, et al. Nonsteroidal anti-inflammatory drugs inhibit expression of the inducible nitric oxide synthase gene. Biochem Biophys Res Commun 1995; 208: 1053-9.
6. Aguirre-Banuelos P, Granados-Soto V. Evidence for the participation of the nitric oxide-cyclic GMP pathway in the antinociceptive action of meloxicam in the formalin test. Eur J Pharmacol 2000; 395: 9-13.
7. Ahlqvist J, Österlund K. Possible role of inducible nitric oxide synthase in articular chondrocytes in the pathogenesis of arthritic swelling . Arthritis Rheum 1995; 38: 1529-31.
8. Ahmadzadeh N, Shingu M, Nobunaga M. The effect of recombinant tumor necrosis factor-alpha on superoxide and metalloproteinase production by synovial cells and chondrocytes. Clin Exp Rheumatol 1990; 8: 387-91.
9. Aigner T, Bertling W, Stöss H, Weseloh G, von der Mark K. Independent expression of fibril-forming collagen I, II, and III in chondrocytes of human osteoarthritic cartilage. J Clin Invest 1993; 91: 829-37.
10. Akagi Y, Liu W, Zebrowski B, Xie K, Ellis LM. Regulation of vascular endothelial growth factor expression in human colon cancer by insulin-like growth factor-I. Cancer Res 1998; 58: 4008-14.
11. Al-Awadhi A, Olusi S, Al-Zaid N, Prabha K. Serum concentrations of interleukin 6, osteocalcin, intact parathyroid hormone, and markers of bone resorption in patients with rheumatoid arthritis. J Rheumatol 1999; 26: 1250-6.
12. Alaaeddine N, di Battista J, Pelletier J, Kiansa K, Cloutier J, Martel-Pelletier J. Inhibition of tumour necrosis factor -induced prostaglandin E2 production by the antiinflammatory cytokines interleukin-4, interleukin-10, and interleukin-13 in osteoarthritic synovial fibroblasts. Distinct targeting in the signalling pathways. Arthritis Rheum 1999; 42: 710-8.
13. Alaaeddine N, Dibattista J, Pelletier J-P, Cloutier J, Kiansa K, Dupuis M, et al. Osteoarthritic synovial fibroblasts possess an increased level of tumor necrosis factor-receptor 55 (TNF-F55) that mediates biological activation by TNF-alpha. J Rheumatol 1997; 24: 1985-94.
14. Alexander C. Osteoarthritis: A review of old myths and current concepts. Skeletal Radiol 1990; 19: 327-33.
15. Alla-Kokko L, Baldwin CT, Moskovitz RW, Weaver EJ, Malemud CJ, Pathria MN, et al. A single base mutation in the type II procollagen gene (Col 2A1) as a cause of primary osteoarthritis associated with mild chondrodysplasia. Proc Natl Acad Sci USA 1990; 87: 6565-8.
16. Allen J, Manthey C, Hand A, Ohura K, Ellingsw L, Wahl S. Rapid onset synovial inflammation

232.
and hyperplasia induced by transforming growth factor-beta. J Exp Med 1990; 171: 231-47.
17. Altman R, Alarcon G, Appelrouth D, Bloch D, Borenstein D, Brandt K, et al. The American College of Rheumatology criteria for the classification and reporting of osteoarthritis of the hand. Arthritis Rheum 1990; 33: 1601-10.
18. Altman RD. Calcium pentosan in a canine model of osteoarthritis. Osteoarthritis Cartilage 1999; 7 (Suppl. A): S18 (Abstract).
19. Altman RD, Dean DD, Muniz OE, Howell DS. Therapeutic treatment of canine osteo-arthritis with glycosaminoglycan polysulfuric acid ester. Arthritis Rheum 1989; 32: 1300-7.
20. Altman R, Gray R. Inflammation in osteoarthritis. Clin Rheum Dis 1985; 11: 353-65.
21. Altman RD, Hochberg M, Murphy WA, Wolfe F, Lequesne M. Altlas of individual radiographic features in osteoarthritis. Osteoarthritis Cartilage 1995; 3 (Suppl A): 3-70.
22. Amin AR, Attur M, Patel RN, Thakker GD, Marshall PJ, Rediske J, et al. Superinduction of cyclooxygenase-2 activity in human osteoarthritis-affected cartilage: Influence of nitric oxide. J Clin Invest 1997; 99: 1231-7.
23. Amin AR, Attur MG, Thakker GD, Patel PD, Vyas PR, Patel RN, et al. A novel mechanism of action of tetracyclines: effects on nitric oxide synthases. Proc Natl Acad Sci USA 1996; 93: 14014-9.
24. Amin AR, Dave M, Attur M, Abramson SB. COX-2, NO, and cartilage damage and repair . Curr Rheumatol Rep 2000; 2: 447-53.
25. Amin AR, Dicesare PE, Vyas P, Atur M, Tzeng E, Billar TR, et al. The expression and regulation of nitric oxide synthase in human osteoarthritis-affected chondrocytes: Evidence for up-regulated neuronal nitric oxide synthase. J Exp Med 1995; 182: 2097-102.
26. Anastassiades T, Ley J, Wood A. Glycosaminoglycan synthesis and glucose uptake by rheumatoid and nonrheumatoid fibroblastic cells in culture. Arthritis Rheum 1979; 22: 871-6.
27. Anastassiades TP, Ley J, Wood A, Irwin D. The growth kinetics of synovial fibroblastic cells from inflammatory and non-inflammatory arthropathies. Arthritis Rheum 1978; 21: 461-6.
28. Andrews AE, Barcham GJ, Brandon MR, Nash AD. Molecular cloning and characterization of ovine IL-1-alpha and IL-1-beta. Immunol 1991; 74: 453-60.
29. Andrews HJ, Edwards TA, Cawston TE, Hazleman BL. Transforming growth factor-beta causes partial inhibition of interleukin-1 - stimulated cartilage degradation in vitro. Biochem Biophys Res Commun 1989; 162: 144-50.
30. Andrews J, Ghosh P. Low molecular weight serine proteinase inhibitors of human articular cartilage - isolation, characterization, and biosynthesis. Arthritis Rheum 1990; 33: 1384-93.
31. Antoniades L, MacGregor AJ, Matson M, Spector TD. A cotwin study of the relationship between hip osteoarthritis and bone mineral density. Arthritis Rheum 2000; 43: 1450-5.
32. Appel A, Hopson C, Herman J. Modulation of cartilage proteoglycan synthesis by osteoarthritic synovium. J Rheumatol 1988; 15:(10)1515-1524.
33. Appleyard RC, Ghosh P, Swain MV. Biomechanical, histological and immunohistochemical studies of patellar cartilage in an ovine model of osteoarthritis induced by lateral meniscectomy. Osteoarthritis Cartilage 1999; 7: 281-294.

233.
34. Appleyard RC, Swain MV, Ghosh P. Modification of the dynamic shear modulus and phase lag properties of tibial plateau articular cartilage in an ovine model of osteoarthritis by oral administration of calcium pentosan polysulphate. Trans Orthop Res Soc 2000; 320 (Abstract)
35. Appleyard RC, Swain MV, Khanna S, Murrell GAC. The accuracy and reliability of a novel handheld dynamic indentation probe for analysing articular cartilage. Phys Med Biol 2001; 46: 541-50.
36. Armour K, Ralston S. Estrogen upregulates endothelial constitutive nitric oxide synthase expression in human osteoblast-like cells. Endocrinol 1998; 139: 799-802.
37. Armstrong S, Lees P. Effects of R and S enantiomers and a racemic mixture of carprofen on the production and release of proteoglycan and prostaglandin E2 from equine chondrocytes and cartilage explants. Am J Vet Res 1999; 60: 98-104.
38. Armstrong S, Read R, Ghosh P. The effects of intra-articular hyaluronan on cartilage and subchondral bone changes in an ovine model of early osteoarthritis. J Rheumatol 1994; 21:680-8.
39. Armstrong SJ. Studies on the articular cartilage and subchondral bone following unilateral medial meniscectomy in sheep. PhD thesis: Murdoch University, 1993.
40. Armstrong SJ, Read RA, Ghosh P, Wilson DM. Moderate exercise exacerbates the osteoarthritic lesions produced in cartilage by meniscectomy: a morphologic study. Osteoarthritis Cartilage 1993; 1: 89-96.
41. Armstrong SJ, Read RA, Price R. Topographical variation within the articular cartilage and subchondral bone of the normal ovine knee joint: A histological approach. Osteoarthritis Cartilage 1995; 5: 25-33.
42. Arner EC, Dimeo TM, Ruhl DM, Pratta MA. In vivo studies on the effects of human recombinant interleukin-1-beta on articular cartilage. Agents Actions 1989; 27: 254-7.
43. Arner EC, Hughes CE, Decicco CP, Caterson B, Tortorella MD. Cytokine-induced cartilage proteoglycan degradation is mediated by aggrecanase. Osteoarthritis Cartilage 1998; 6:214-28.
44. Arner E, Pratta M. Modulation of interleukin-1-induced alterations in cartilage proteoglycan metabolism by activation of protein kinase-C. Arthritis Rheum 1991; 34: 1006-13.
45. Arokoski JPA, Hyttinen MM, Lapveteläinen T, Takács P, Kosztáczky B, Módis L, et al. Decreased birefringence of the superficial zone collagen network in the canine knee (stifle) articular cartilage after long distance running training, detected by quantitative polarised light microscopy. Ann Rheum Dis 1996; 55: 253-64.
46. Arts J, Kuiper G, Janssen J, Gustaffsson J-Å, Löwik C, Pols H, et al. Differences in estrogen receptor and ß mRNA during differentiation of human osteoblast SV-HFO cells. Endocrinol 1997; 138: 5067-70.
47. Athanasiou KA, Rosenwasser MP, Buckwalter JA, Malinin TI, Mow VC. Interspecies comparisons of in situ intrinsic mechanical properties of distal femoral cartilage. J Orthop Res 1991; 9: 330-40.
48. Athanasou NA, Quinn J, Heryet A, Puddle B, Woods CG, McGee JO'D. The immunohistology of synovial lining cells in normal and inflamed synovium. J Pathol 1988; 155: 133-42.
49. Attur MG, Patel IR, Patel RN, Abramson SB, Amin AR. Autocrine production of IL-1ß by human osteoarthritis-affected cartilage and differential regulation of endogenous nitric oxide, IL-6, prostaglandin E2, and IL-8. Proc Assoc Am Physicians 1998; 110: 65-72.

234.
50. Attur MG, Patel R, Dicesare PE, Steiner GC, Abramson SB, Amin AR. Regulation of nitric oxide production by salicylates and tenidap in human OA-affected cartilage, rat chondrosarcomas and bovine chondrocytes. Osteoarthritis Cartilage 1998; 6: 269-77.
51. Attur MG, Patel RN, Abramson SB, Amin AR . Interleukin-17 up-regulation of nitric oxide production in human osteoarthritis cartilage. Arthritis Rheum 1997; 40: 1050-3.
52. Bader DL, Kempson GE, Egan J, Gilbey W, Barrett AJ. The effects of selective matrix degradation on the short-term compressive properties of adult human articular cartilage. Biochim Biophys Acta 1992; 1116: 147-54.
53. Badger AM, Cook MN, Lark MW, Newman-Tarr TM, Swift BA, Nelson AH, et al. SB 203580 inhibits p38 mitogen-activated protein kinase, nitric oxide production, and inducible nitric oxide synthase in bovine cartilage-derived chondrocytes. J Immunol 1998; 161: 467-73.
54. Bailey A, Mansell J. Do subchondral bone changes exacerbate or precede articular cartilage destruction in osteoarthritis of the elderly? Gerontology 1997; 43: 296-304.
55. Baker D, Dayer J-M, Roelke M, Schumacher H, Krane S. Rheumatoid synovial cell morphologic changes induced by a mononuclear cell factor in culture. Arthritis Rheum 1983; 26: 8-14.
56. Balavoine JF, De Rochemonteix B, Williamson K, Seckinger P, Cruchaud A, Dayer J-M. Prostaglandin-E2 and collagenase production by fibroblasts and synovial cells is regulated by urine-derived human interleukin-1 and inhibitor(s). J Clin Invest 1986; 78: 1120-4.
57. Bandara G, Georgescu HI, Lin CW, Evans CH. Synovial activation of chondrocytes: Evidence for complex cytokine interactions. Agents Actions 1991; 34: 285-8.
58. Barland P, Novikoff A, Hamerman D. Electron microscopy of the human synovial membrane. J Cell Biol 1962; 14: 207-20.
59. Barratt ME, Fell HB, Coombs RR, Glauert AM. The pig synovium, II. Some properties of isolated intimal cells. J Anat 1977; 123: 47-66.
60. Bashir A, Lewis M, Henderson A. Pharmacokinetic studies of various preparations of glyceryl trinitrate. Br J Clin Pharmac 1982; 14: 779-84.
61. Bassleer C, Magotteaux J, Geenen V, Malaise M. Effects of meloxicam compared to acetylsalicyclic acid in human articular chondrocytes. Pharmacol 1997; 54: 49-56.
62. Bathon J, Hwang J, Shin L, Precht P, Towns M, Horton W. Type VI collagen-specific messengerRNA is expressed constitutively by cultured human synovial fibroblasts and is suppressed by interleukin-1. Arthritis Rheum 1994; 37: 1350-6.
63. Bauer DC, Orwoll ES, Fox KM, Vogt TM, Lane NE, Hochberg MC, et al. Aspirin and NSAID use in older women: Effect on bone mineral density and fracture risk. Study of Osteoporotic Fractures Research Group. J Bone Miner Res 1996; 11: 29-35.
64. Baumann H, Kushner I. Production of interleukin-6 by synovial fibroblasts in rheumatoid arthritis. Am J Pathol 1998; 152: 641-4.
65. Beato, Fiegelson. J Biol Chem 1972; 247: 7890-6.
66. Beekman B, Drijfhout JW, Bloemhoff W, Ronday HK, Tak PP, te Koppele JM. Convenient fluorometric assay for matrix metalloproteinase activity and its application in biological media. FEBS Lett 1996; 390: 221-5.
67. Bellenger C, Pickles D. Load-bearing in the ovine medial tibial condyle: Effect of meniscectomy.

235.
Vet Comp Orthop Traumatol 1993; 6: 100-4.
68. Bellino F. Estrogen metabolism, not biosynthesis, in rabbit articular cartilage and isolated chondrocytes: A preliminary study. Steroids 1992; 57: 507-10.
69. Ben-Hur H, Thole H, Mashiah A, Insler V, Berman V, Shezen E, et al. Estrogen, progesterone and testosterone receptors in human fetal cartilaginous tissue: Immunohistochemical studies. Calcif Tissue 1997; 60: 520-6.
70. Bendele A, McAbee T, Sennello G, Frazier J, Chlipala E, McCabe D. Efficacy of sustained blood levels of interleukin-1 receptor antagonist in animal models of arthritis. Arthritis Rheum 1999; 42: 498-506.
71. Beresford J. Osteogenic stem cells and the stromal system of bone and marrow. Clin Orthop 1989; 240: 270-80.
72. Bermont L, Lamielle F, Fauconnet S, Esumi H, Weisz A, Adessi GL. Regulation of vascular endothelial growth factor expression by insulin-like growth factor-I in endometrial adenocarcinoma cells. Int J Cancer 2000; 85: 117-23.
73. Berse B, Hunt J, Diegel R, Morganelli P, Yeo K, Brown F, et al. Hypoxia augments cytokine (transforming growth factor-ß (TGF-ß) and IL-1)-induced vascular endothelial growth factor secretion by human synovial fibroblasts. Clin Exp Immunol 1999; 115: 176-182.
74. Bertazzolo N, Punzi L, Stefani M, Cesaro G, Pianon M, Finco B, et al. Interrelationships between interleukin (IL)-1, IL-6 and IL-8 in synovial fluid of various arthropathies. Agents Actions 1994; 41: 90-2.
75. Bezerra MM, de Lima V, Alencar VB, Vieira IB, Brito GA, Ribeiro RA, et al. Selective cyclooxygenase-2 inhibition prevents alveolar bone loss in experimental periodontitis in rats . J Periodontol 2000; 71: 1009-14.
76. Bird JLE, May S, Bayliss MT. Nitric oxide inhibits aggrecan degradation in explant cultures of equine articular cartilage. Equine Vet J 2000; 32: 133-9.
77. Blanco F, Guitian R, Moreno J, de Toro F, Galdo F. Effect of antiinflammatory drugs on COX-1 and COX-2 activity in human articular chondrocytes. J Rheumatol 1999; 26: 1366-73.
78. Blanco FJ, Lotz M. IL-1 induced nitric oxide inhibits chondrocyte proliferation via PGE2. Exp Cell Res 1995; 218: 319-25.
79. Blanco FJ, Ochs RL, Schwarz H, Lotz M. Chondrocyte apoptosis induced by nitric oxide. Am J Pathol 1995; 146: 75-85.
80. Blaschke UK, Eikenberg EF, Hulmes DJS, Galla HJ, Bruckner P. Collagen XI nucleates self-assemby and limits lateral growth of cartilage fibrils. J Biol Chem 2000; 275: 370-8.
81. Blot L, Marcelis A, Devogelaer JP, Manicourt DH. Effects of diclofenac, aceclofenac and meloxicam on the metabolism of proteoglycans and hyaluronan in osteoarthritic human cartilage. Br J Pharmacol 2000; 131: 1413-21.
82. Blotman F, Maheu E, Wulwik A, Caspard H, Lopez A. Efficacy and safety of avocado/soybean unsaponifiables in the treatment of symptomatic osteoarthritis of the knee and hip. A prospective, multicenter, three-month, randomized, double-blind, placebo-controlled trial. Rev Rhum [Engl Ed] 1997; 64: 825-34.
83. Boniface RJ, Cain PR, Evans CH. Articular responses to purified cartilage proteoglycans. Arthritis Rheum 1988; 31: 258-266.

236.
84. Borderie D, Hernvann A, Lemarechal H, Menkes CJ, Ekindjian O. Inhibition of the nitrosothiol production of cultured osteoarthritic chondrocytes by rhein, cortisol and diclofenac. Osteoarthritis Cartilage 2001; 9: 1-6.
85. Borghaei R, Rawlings P, Mochan E. Interleukin-4 suppression of interleukin-1-induced transcription of collaenase (MMP-1) and stromelysin-1 (MMP-3) in human synovial fibroblasts. Arthritis Rheum 1998; 41: 1398-406.
86. Boumediene K, Felisaz N, Bogdanowicz P, Galera P, Guillou GB, Pujol JP. Avocado/soya unsaponifiables enhance the expression of tranforming growth factor ß1 and ß2 in cultured articular chondrocytes. Arthritis Rheum 1999; 42: 148-56.
87. Boyle D, Fereydoun GS, Firestein G. Inhibition of synoviocyte collagenase gene expression by adenosine receptor stimulation. Arthritis Rheum 39: 932-40.
88. Brandi ML, Hukkanen M, Umeda T, Moradi-Bidhendi N, Bianchi S, Gross SS, et al. Bidirectional regulation of osteoclast function by nitric oxide synthase isoforms. Proc Natl Acad Sci USA 1995; 92: 2954-8.
89. Brandt KD. Animal models of osteoarthritis. In: Reginster J-Y, Pelletier J-P, Martel-Pelletier J, Henrotin Y (Eds). Osteoarthritis: Clinical and experimental aspects. Berlin : Springer-Verlag, 1999: 81-100.
90. Brandt KD, Myers SL, Burr D, Albrecht M. Osteoarthritic changes in canine articular cartilage, subchondral bone, and synovium fifty-four months after transection of the anterior cruciate ligament. Arthritis Rheum 1991; 34: 1560-70.
91. Brater DC, Harris C, Redfern JS, Gertz BJ. Renal effects of COX-2-selective inhibitors. Am J Nephrol 2001; 21: 1-15.
92. Brennan FM, Chantry D, Jackson A, Maini RN, Feldmann M. Inhibitory effect of the TNF-alpha antibodies on synovial cell interleukin-1 production in rheumatoid-arthritis. Lancet 1989; 65 :244-7.
93. Brennan F, Chantry D, Turner M, Foxwell B, Maini R, Feldmann M. Detection of transforming growth factor-beta in rheumatoid arthritis synovial tissue: lack of effect on spontaneous cytokine production in joint cell cultures. Clin Exp Immunol 1990; 81:278-285. Abstract.
94. Bridges A, Malone D, Jicinsky J, Chen M, Ory P, Engber W, et al. Human synovial mast cell involvement in rheumatoid arthritis and osteoarthritis. Relationship to disease type, clinical activity, and antirheumatic therapy. Arthritis Rheum 1991; 34: 1116-24.
95. Brinckerhoff C, Guyre P. Increased proliferation of human synovial fibroblasts treated with recombinant immune interferon. J Immunol 1985; 134: 3142-6.
96. Brinckerhoff C, McMillan R, Dayer J-M, Harris EJ. Inhibition by retinoic acid of collagenase production in rheumatoid synovial cells. New Engl J Med 1980; 303: 432-6.
97. Brown M, Hazleman B, Dingle J, Dandy D, Murley A. Production of cartilage degrading activity by human synovial tissues. Ann Rheum Dis 1987; 46: 319-23.
98. Bryson H, Bunning R, Feltell R, Kam C, Kerrigan J, Powers J, et al. A serine proteinase inactivator inhibits chondrocyte mediated cartilage proteoglycan breakdown occurring in response to proinflammatory cytokines. Arch Biochem Biophys 1998; 355: 15-25.
99. Buckingham R, Castor C, Hoag P. The effect of bacterial products on synovial fibroblast function: hypermetabolic changes induced by endotoxin. J Clin Invest 1972; 51: 1186-94.

237.
100. Bunning RAD, Crawford A, Richardson HJ, Opdenakker G, Van Damme J, Russell RGG. Interleukin-1 preferentially stimulates the production of tissue-type plasminogen-activator by human articular chondrocytes. Biochim Biophys Acta 1987; 924: 473-82.
101. Burkhardt D, Ghosh P. Laboratory evaluation of glycosaminoglycan polysulphate ester for chondroprotective activity. Curr Ther Res 1986; 40: 1034-53.
102. Busso N, Peclat V, So A, Sappino A. Plasminogen activation in synovial tissue: Differences between normal, osteoarthritis, and rheumatoid arthritis joints. Ann Rheum Dis 1997; 56: 550-7.
103. Cahill LP. Folliculogenesis and ovulation rate in sheep. In: Lindsay DR, Pearce DT (Eds.) Reproduction in sheep. Cambridge: Cambridge University Press, 1984: 92-8.
104. Cake MA, Read RA, Guillou B, Ghosh P. Modification of articular cartilage and subchondral bone pathology in an ovine meniscectomy model of osteoarthritis by avocado and soya unsaponifiables (ASU). Osteoarthritis Cartilage 2000; 8: 404-11.
105. Cameron M, Buchgraber A, Passler H, Vogt M, Thonar E, Fu F, et al. The natural history of the anterior cruciate ligament-deficient knee. Changes in synovial fluid cytokine and keratan sulfate concentrations. Am J Sports Med 1997; 25: 751-4.
106. Campbell IK, Piccoli DS, Butler DM, Singleton DK, Hamilton JA. Recombinant human interleukin-1 stimulates human articular cartilage to undergo resorption and human chondrocytes to produce both tissue-type and urokinase-type plasminogen activator. Biochim Biophys Acta 1988; 967: 183-94.
107. Cantatore FP, Acquista CA, Corrado A, Iannone F, Lapadula G. Extracellular matrix interactions with osteoblasts in osteoarthritis (Letter). J Rheumatol 2001; 28: 922-4.
108. Cao M, Stefanovic-Racic M, Georgescu HI, Fu FH, Evans CH. Does nitric oxide help explain the differential healing capacity of the anterior cruciate, posterior cruciate, and medial collateral ligaments? Am J Sports Med 2000; 28: 176-82.
109. Cao M, Stefanovic-Racic M, Georgescu HI, Miller LA, Evans CH. Generation of nitric oxide by lapine meniscal cells and its effect on matrix metabolism: stimulation of collagen production by arginine. J Orthop Res 1998; 16: 104-11.
110. Cao M, Westerhausen-Larson A, Niyibizi C, Kavalkovich K, Georgescu HI, Rizzo CF, et al. Nitric oxide inhibits the synthesis of type-II collagen without altering Col2A1 mRNA abundance: prolyl hydroxylase as a possible target. Biochem J 1997; 324: 305-10.
111. Cardim HJ, Lopes CM, Giannella-Neto D, da Fonseca AM, Pinotti JA. The insulin-like growth factor-I system and hormone replacement therapy. Fertil Steril 2001; 75: 282-7.
112. Carnovale DE, Fukuda A, Underhill DC, Laffan JJ, Breuel KF. Aspirin dose dependently inhibits the interleukin-1 beta-stimulated increase in inducible nitric oxide synthase, nitric oxide, and prostaglandin E(2) production in rat ovarian dispersates cultured in vitro. Fertil Steril 2001; 75: 778-84.
113. Carroll G. Measurement of sulphated glycosaminoglycans and proteoglycan fragments in arthritic synovial fluid. Ann Rheum Dis 1989; 48: 17-24.
114. Carroll G. Porcine catabolin stimulates prostaglandin E2 secretion but does not affect intracellular cyclic AMP production in pig synovial fibroblasts. Ann Rheum Dis 1985; 44: 631-6.
115. Case JP, Lafyatis R, Kumkumia GK, Remmers EF, Wilder RL. IL-1 regulation of transin stromelysin transcription in rheumatoid synovial fibroblasts appears to involve 2 antagonistic transduction pathways, an inhibitory, prostaglandin-dependent pathway mediated by CAMP, and a

238.
stimulatory, protein kinase-C-dependent pathway. J Immunol 1990;145: 3755-61.
116. Castor CW, Dorstewitz EL. Abnormalities of Connective tissue cells cultured from patients with rheumatoid arthritis I: Relative unresponsiveness of rheumatoid synovial cells to hydrocortisone. J Lab Clin Med 1966; 68: 300-13.
117. Castor C, Ritchie J, Scott M, Whitney S. Connective tissue activation XI: Stimulation of glycosaminoglycan and DNA formation by a platelet factor. Arthritis Rheum 1977; 20: 859-68.
118. Castor C, Roberts D, Hossler P, Bignall M. Connective tissue activation XXV: Regulation of proteoglycan synthesis in human synovial cells. Arthritis Rheum 1983; 26: 522-7.
119. Castor C, Smith S, Ritchie J, Dorstewitz E. Connective tissue activation II: Abnormalities of cultured rheumatoid synovial cells. Arthritis Rheum 1971; 14: 55-66.
120. Cauley JA, Gutal JP, Kuller LH, Powell JG. The relation of endogenous sex steroid hormone concentrations to serum lipid and lipoprotein levels in postmenopausal women. Am J Epidemiol 1990; 132: 884-94.
121. Cawston T, Rowan A. Prevention of cartilage breakdown by matrix metalloproteinase inhibition - A realistic therapeutic target? Br J Rheumatol 1998; 37: 353-6.
122. Chambers MG, Cox L, Chong L, Suri N, Cover P, Bayliss MT, et al. Matrix metalloproteinases and aggrecanases cleave aggrecan in different zones of normal cartilage but colocalize in the development of osteoarthritic lesions in STR/ort mice. Arthritis Rheum 2001; 44: 1455-65.
123. Chander C, Desa F. The effects of estrogens on cartilage degradation using in vivo and in vitro models. Agents Actions 1991; 34: 282-4.
124. Chandrasekhar S, Harvey AK. Transforming growth factor-beta is a potent inhibitor of IL-1 induced protease activity and cartilage proteoglycan degradation. Biochem Biophys Res Commun 1988; 157: 1352-9.
125. Chandrasekhar S, Harvey AK, Hrubey PS. Intra-articular administration of interleukin-1 causes prolonged suppression of cartilage proteoglycan synthesis in rats. Matrix 1992; 11: 1-10.
126. Chang D, Iverson E, Schinagl R, Sonoda M, Amiel D, Coutts R, et al. Quantitation and localization of cartilage degeneration following the induction of osteoarthritis in the rabbit knee. Osteoarthritis Cartilage 1997; 5: 357-72.
127. Chavassieux P, Garnero P, Duboeuf F, Vergnaud P, Brunner-Ferber F, Delmas PD, et al. Effects of a new selective estrogen receptor modulator (MDL 103,323) on cancellous and cortical bone in ovariectomised ewes: A biochemical, histomorphometric, and densitometric study. J Bone Miner Res 2001; 16: 89-96.
128. Chen V, Croft D, Purkis P, Kramer I. Co-culture of synovial fibroblasts and differentiated U937 cells is sufficient for high interleukin-6 but not interleukin-1-beta or tumor necrosis factor-alpha release. Br J Rheumatol 1998; 37: 148-56.
129. Cheng MZ, Zaman G, Rawlinson SC, Mohan S, Baylink DJ, Lanyon LE. Mechanical strain stimulates ROS cell proliferation through IGF-II and estrogen through IGF-I. J Bone Miner Res 1999; 14: 1742-50.
130. Chevalier X. Upregulation of enzymatic activity by interleukin-1 in osteoarthritis. Biomed Pharmacother 1997; 51: 58-62.
131. Chevalier X, Tyler JA. Production of binding proteins and the role of insulin-like growth factor I binding protein 3 in human articular cartilage explants. Br J Rheumatol 1996; 35: 515-22.

239.
132. Chomarat P, Rissoan M, Pin J, Banchereau J, Miossec P. Contribution of IL-1, CD14, and CD13 in the increased IL-6 production induced by in vitro monocyte-synoviocyte interactions. J Immunol 1995; 155: 3645-52.
133. Chomarat P, Vannier E, Dechanet J, Rissoan M, Banchereau J, Dinarello C, et al. Balance of IL-1 receptor antagonist/IL-ß in rheumatoid synovium and its regulation by IL-4 and IL-10. J Immunol 1995; 154: 1432-9.
134. Chow JWM, Fox SW, Lean JM, Chambers TJ. Role of nitric oxide and prostaglandins in mechanically produced bone formation. J Bone Miner Res 1998; 13: 1039-44.
135. Chowdbury TT, Bader DL, Lee DA. Dynamic compression inhibits the synthesis of nitric oxide and PGE2 by IL-1ß-stimulated chondrocytes cultured in agarose constructs. Biochem Biophys Res Commun 2001; 285: 1168-74.
136. Chu CQ, Field M, Abney E, Zheng RQH, Allard S, Feldmann M, et al. Transforming growth factor-beta-1 in rheumatoid synovial membrane and cartilage pannus junction. Clin Exp Immunol 1991; 86: 380-6.
137. Cicala C, Ianaro A, Fiorucci S, Calignano A, Bucci M, Gerli R, et al. NO-naproxen modulates inflammation, nociception and downregulates T cell response in rat Freund's adjuvant arthritis . Br J Pharmacol 2000; 1330: 1399-405.
138. Ciosek (Jr) C, Goldlust M. Rabbit synovial tissue in culture: influence of the inflammatory state in vivo on the secretion of collagenase. J Rheumatol 1981; 8: 367-79.
139. Cipolleta C, Jouzeau JY, Gegout-Pottie P, Presle N, Bordji K, Netter P, et al. Modulation of IL-1-induced cartilage injury by NO synthase inhibitors: A comparative study with rat chondrocytes and cartilage entities. Br J Pharmacol 1998; 124: 1719-27.
140. Clancy R. Nitric oxide alters chondrocyte function by disrupting cytoskeletal signalling complexes. Osteoarthritis Cartilage 1999; 7: 399-400.
141. Clancy RM, Rediske J, Tang X, Nijher N, Frenkel S, Philps M, et al. Outside-in signaling in the chondrocyte. Nitric oxide disrupts fibronectin-induced assembly of a subplasmalemmal actin/rho A/focal adhesion kinase signaling complex. J Clin Invest 1997; 100: 1789-96.
142. Clapper JA, Clark TM, Rempel LA. Serum concentrations of IGF-I, estradiol 17ß, testosterone, and relative amounts of IGF binding proteins (IGFBP) in growing boars, barrows, and gilts. J Animal Sci 2000; 78: 2581-8.
143. Clark CR. The cellular distribution of steroid hormone receptors: have we got it right? Trends Biochem Sci 1984; 9: 207-8.
144. Clark IM, Powell LK, Ramsey S, Hazleman BL, Cawston TE. The measurement of collagenase, tissue inhibitor of metalloproteinases (TIMP), and collagenase TIMP complex in synovial fluids from patients with osteoarthritis and rheumatoid arthritis. Arthritis Rheum 1993; 36: 372-9.
145. Clarris B, Fraser J, Muirden K, Malcolm L, Holmes M, Rogers K. Stimulation of glycosaminoglycan production and lysosomal activity of human synovial cells in culture by low environmental pH. Ann Rheum Dis 1984; 43: 313-9.
146. Clemmensen I, Holund B, Andersen R. Fibrin and fibronectin in rheumatoid synovial membrane and rheumatoid synovial fluid. Arthritis Rheum 1983; 26: 479-85.
147. Cohen S, Katsikis P, Chu C-Q, Thomssen H, Webb LR, Londei M, et al. High level of interleukin-10 production by the activated T cell population within the rheumatoid synovial membrane. Arthritis Rheum 1995; 38: 946-52.

240.
148. Coleman P, Scott D, Abiona A, Ashhurst D, Mason R, Levick J. Effect of depletion of interstitial hyaluronan on hydraulic conductance in rabbit knee synovium. J Physiol (Lond) 1998; 509: 695-710.
149. Coleman P, Green G. A coupled photometric assay for plasminogen activator. Methods Enzymol 1981; 80: 408-14.
150. Collier S, Ghosh P. The role of plasminogen in Interleukin-1 mediated cartilage degradation. J Rheumatol 1988; 15: 1129-37.
151. Collier S, Ghosh P. Evaluation of the effects of antiarthritic drugs on the secretion of proteoglycans by lapine chondrocytes using a novel assay procedure. Ann Rheum Dis 1989; 48: 372-81.
152. Colombo C, Butler M, Hickman L, Selwyn M, Chart J, Steinetz B. A new model of osteoarthritis in rabbits II Evaluation of anti-osteoarthritic effects of selected antirheumatic drugs administered systemically. Arthritis Rheum 1983; 26: 1132-9.
153. Cooper C, Cook P, Osmond C, Fisher L, Cawley M. Osteoarthritis of the hip and osteoporosis of the proximal femur. Ann Rheum Dis 1991; 50: 540-2.
154. Cooper C, Egger P, Coggon D, Hart D, Masud T, Cicuttini F, et al. Generalized osteoarthritis in women: Pattern of joint involvement and approaches to definition for epidemiologic studies. J Rheumatol 1996; 23: 1938-42.
155. Cooper R, Milgram J, Robinson R. Morphology of the osteon. J Bone Joint Surg 1966; 48A: 1239-71.
156. Corlett S, Care A. The effects of reduced dietary phosphate intake on plasma osteocalcin levels in sheep. Quarterly J Exp Physiol 1988; 73: 443-5.
157. Corlett S, Couch M, Care A, Sykes A. Measurement of plasma osteocalcin in sheep: Assessment of circadian variation, the effects of age and nutritional status and the response to perturbation of the adrenocortical axis. Exp Physiol 1990; 75: 515-27.
158. Cova M, Toffanin R, Frezza F, Pozzi-Mucelli M, Mlynarik V, Pozzi-Mucelli RS, et al. Magnetic resonance imaging of articular cartilage: Ex vivo study on normal cartilage correlated with magnetic resonance microscopy. Eur Radiol 1998; 8: 1130-6.
159. Croxford LJ. COX-2 in synovial tissues. Osteoarthritis Cartilage 1999; 7: 406-8.
160. Curry S, Kwon H-R, Perrin J, Culp J, Pepine C, Yu W, et al. Plasma nitroglycerin concentrations and haemodynamic effects of sublingual, ointment, and controlled release forms of nitroglycerin. Clin Pharmacol Ther 1984; 36: 765-72.
161. Cutolo M. Do sex hormones modulate the synovial macrophages in rheumatoid arthritis? Ann Rheum Dis 1997; 56: 281-4.
162. Cutolo M. The roles of steroid hormones in arthritis. Br J Rheumatol 1998; 37: 597-9.
163. Cutolo M, Accardo S, Villaggio B, Barone A, Sulli A, Coviello D, et al. Androgen and estrogen receptors are present in primary cultures of human synovial macrophages. J Clin Endocrinol Metab 1996; 81: 820-7.
164. Cutolo M, Sulli A, Barone A, Seriolo B, Accardo S. Macrophages, synovial tissue and rheumatoid arthritis. Clin Exp Rheumatol 1993; 11: 331-9.
165. Cutolo M, Sulli A, Barone A, Seriolo B, Accardo S. Sex hormones, proto-oncogene expression

241.
and apoptosis: their effects on rheumatoid synovial tissue. Clin Exp Rheumatol 1996; 14: 87-94.
166. Dahl I, Husby G. Hyaluronic acid production in vitro by synovial lining cells from normal and rheumatoid joints. Ann Rheum Dis 1985; 44: 647-57.
167. Dahlberg L, Friden T, Roos H, Lark M, Lohmander L. Longitudinal study of cartilage matrix metabolism in patients with cruciate ligament rupture: Synovial fluid concentrations of aggrecan fragments, stromelysin-1 and tissue inhibitor of metalloproteinase-1. Br J Rheumatol 1994; 33: 1107-11.
168. Daireaux M, Penfornis H, Langris M, Bocquet J, Pujol J-P, Beliard R, et al. Effect of a mononuclear cell culture medium on collagen and glycosaminoglycan production by synovial cells in culture. FEBS Lett 1981; 132: 93-7.
169. Damien E, Price JS, Lanyon LE. The estrogen-receptors involvement in osteoblasts adaptive response to mechanical strain. J Bone Miner Res 1998; 13: 1275-82.
170. Damoulis PD, Hauschka PV. Nitric oxide acts in conjunction with proinflammatory cytokines to promote cell death in osteoblasts. J Bone Miner Res 1997; 12: 412-22.
171. Danziger R, Zuckerbraun B, Pensler J. Role of nitric oxide in the regulation of osteoblast metabolism. Plast Reconstr Surg 1997; 100: 670-3.
172. Dayer J-M, Goldring S, Robinson D, Krane S. Effects of human monocellular cell factor on cultured rheumatoid synovial cells: interactions of prostaglandin E2 and cyclic adenosine 3'5'monophosphate. Biochim Biophys Acta 1979; 586:87-105.
173. Dayer J-M, Krane SM, Russell RGG, Robinson DR. Production of collagenase and prostaglandins by isolated adherent rheumatoid synovial cells. Proc Natl Acad Sci USA 1976; 73: 945-9.
174. Dayer J-M, Russell R, Krane S. Collagenase roduction by rheumatoid synovial cells: stimulation by a human lymphocyte factor. Science 1977; 195: 181-3.
175. Dayer JM, De Rochemonteix B, Burrus B, Demczuk S, Dinarello CA. Human recombinant interleukin-1 stimulates collagenase and prostaglandin E-2 production by human synovial cells. J Clin Invest 1986; 77: 645-8.
176. Dayer J, Passwell J, Schneeberg E, Krane S. Interactions among rheumatoid synovial cells and monocyte-macrophages production of collagenase-stimulating factor by human monocytes exposed to concanavalin A or immunoglobulin Fc fragments. J Immunol 1980; 124: 1712-20.
177. Dean DD. Proteinase-mediated cartilage degradation in osteoarthritis. Sem Arthritis Rheum 1991; 20: 2-11.
178. Dechanet J, Taupin J-L, Chomarat P, Rissoan M-C, Moreau J-F, Banchereau J, et al. Interleukin-4 but not interleukin-10 inhibits the production of leukemia inhibitory factor by rheumatoid synovium and synoviocytes. Eur J Immunol 1994; 24: 3222-8.
179. Dedrick DK, Goldstein SA, Brandt KD, O'Connor BL, Goulet RW, Albrecht M. A longitudinal study of subchondral plate and trabecular bone in cruciate deficient dogs with osteoarthritis followed up for 54 months. Arthritis Rheum 1993; 36: 1460-7:
180. Dedrick DK, Goulet RW, O'Connor BL, Brandt KD. Preliminary report: Increased porosity of the subchondral plate in an accelerated canine model of osteoarthritis. Osteoarthritis Cartilage 1997; 5: 71-4.
181. Dee JF. Medical management of osteoarthritis in senior dogs. Vet Econ 1998; (Suppl) 4-10.

242.
182. Deleuran B, Chu C, Field M, Brennan F, Katsikis P, Feldmann M, et al. Localization of interleukin-1-alpha, type-1 interleukin-1 receptor and interleukin-1 receptor antagonist in the synovial membrane and cartilage pannus junction in rheumatoid arthritis. Br J Rheumatol 1992; 31: 801-9.
183. Demaziere A, Leek R, Athanasou N. Histological distribution of the interleukin-4 receptor (IL4R) within the normal and pathological synovium. Rev Rhum Mal Osteoartic 1992; 59: 219-24.
184. Dequeker J, Mohan S, Finkelman RD, Aerssens J, Baylink DJ. Generalized osteoarthritis associated with increased insulin-like growth factor types I and II and transforming growth factor b in cortical bone from the iliac crest. Possible mechanism of increased bone density and protection against osteoporosis. Arthritis Rheum 1993; 36:1702-8.
185. Dequeker J, Mokassa L, Aerssens J. Bone density and osteoarthritis. J Rheumatol 1995; 22:98-100.
186. Di Battista JA, Doré S, Martel-Pelletier J, Pelletier J-P. Prostaglandin E2 stimulates incorporation of proline into collagenase digestible proteins in human articular chondrocytes: identification of an effector loop involving insulin-like growth factor I. Mol Cell Endocrinol 1996; 123: 27-35.
187. Di Battista JA, Dore S, Morin N, He Y, Pelletier JP, Martel-Pelletier J. Prostaglandin E2 stimulates insulin-like growth factor binding protein-4 expression and synthesis in cultured human articular chondrocytes: possible mediation by Ca++-calmodulin regulated processes. J Cell Biochem 1997; 65:408-19.
188. Di Battista JA, Mertel-Pelletier J, Pelletier JP. Suppression of tumor necrosis factor (TNF- ) gene expression by prostaglanding E2. Role of early growth response protein-1 (Egr-1). Osteoarthritis Cartilage 1999; 7: 395-8.
189. Diab M, Wu J-J, Eyre DR. Collagen type IX from human cartilage: A structural profile of intermolecular cross-linking sites. Biochem J 1996; 314: 327-32.
190. DiBattista JA, Dore S, Morin N, Abribat T. Prostaglanding E2 up-regulates insulin-like growth factor binding protein-3 expression and synthesis in human articular chondrocytes by a cAMP-independent pathway: Role of calcium and protein kinase A and C. J Cell Biochem 1996; 63:320-33.
191. DiBattista J, Martel-Pelletier J, Fujimoto N, Obata K, Zafarullah M, Pelletier J-P. Prostaglandins E2 and E1 inhibit cytokine-induced metalloprotease expression in human synovial fibroblasts. Lab Invest 1994; 71:270-8.
192. DiBattista J, Pelletier J-P, Zafarullah M, Fujimoto N, Obata K, Martel-Pelletier J. Coordinate expression of matrix metalloproteases and tissue inhibitor of metalloproteinase expression in human synovial fibroblasts. J Rheumatol 1995; 22:(Suppl 43): 123-8.
193. DiBattista J, Pelletier J-P, Zafarullah M, Iwata K, Martel-Pelletier J. Interleukin-1ß induction of tissue inhibitor of metalloproteinase (TIMP-1) is functionally antagonized by prostaglandin E2 in human synovial fibroblasts. J Cell Biochem 1995; 57: 619-29.
194. Dieppe P, Cushnaghan J, Young P, Kirwan J. Prediction of the progression of joint space narrowing in osteoarthritis of the knee by bone scintigraphy. Ann Rheum Dis 1993; 52: 557-63.
195. Ding M, Dalstra M, Linde F, Hvid I. Mechanical properties of the normal human tibial cartilage-bone complex in relation to age. Clin Biomech 1998; 13: 351-8.
196. Dingle JT. Catabolin - a cartilage catabolic factor from synovium. Clin Orthop 1981; 156: 219-31.
197. Dingle JT. Cartilage maintenance in osteoarthritis: Interaction of cytokines, NSAID and

243.
prostaglandins in articular cartilage damage and repair. J Rheumatol 1991; 18 (Suppl 28): 30-37.
198. Dingle JT. Non-steroidal anti-inflammatory drug administration in the treatment of osteoarthritis. In: Reginster J-Y, Pelletier J-P, Martel-Pelletier J, Henrotin Y (Eds.) Osteoarthritis: Clinical and experimental aspects. Berlin : Springer-Verlag, 1999: 370-86.
199. Diwan A, Wang M, Jang D, Zhu W, Murrell G. Nitric oxide modulates fracture healing . J Bone Miner Res 2000; 15: 342-51.
200. Dobson H, Tebble JE, Phogat JB, Smith RF. Effect of transport on pulsatile and surge secretion of LH in ewes in the breeding season. J Reprod Fertil 1999; 116: 1-8.
201. Dooley S, Herlitzka I, Hanselmann R, Ermis A, Henn W, Remberger K, et al. Constitutive expression of c-fos and c-jun, overexpression of ets-2, and reduced expression of metastatis suppressor gene nm23-H1 in rheumatoid arthritis. Ann Rheum Dis 1996; 55: 298-304.
202. Dore S, Abribat T, Rousseau N, Brazeau P, Tardif G, DiBattista JA, et al. Increased insulin-like growth factor 1 production by human osteoarthritic chondrocytes is not dependent on growth hormone action. Arthritis Rheum 1995; 38: 413-9.
203. Dowd G, Gaynor J, Alvis M, Salman M, Turner A. A comparison of transdermal fentanyl and oral phenylbutazone for postoperative analgesia in sheep. Proc Ann Mtg Am Coll Vet Anaes 1997: 13 (Abstract).
204. Eastell R. Treatment of postmenopausal osteoporosis . N Eng J Med 1998; 338: 736-46.
205. Edwards J. The nature and origins of synovium: Experimental approaches to the study of synoviocyte differentiation. J Anat 1994; 184: 493-501.
206. Eisenberg SP, Evares RJ, Arend WP, Verderber E, Brewer E, Hannum CH, et al. Primary structure and functional expression for complementary DNA of a human interleukin-1 receptor antagonist. Nature 1990; 343: 341-6.
207. Erdmann J, Storch S, Pfeilschifter J, Ochlich P, Ziegler R, Bauss F. Effects of estrogen on the concentration of insulin-like growth factor-I in rat bone matrix. Bone 1998; 22: 503-7.
208. Eriksen E, Colvard D, Berg N, Graham M, Mann K, Spelsberg T, et al. Evidence of estrogen receptors in normal human osteoblast-like cells. Science 1988; 84-6.
209. Evans CH. Cartilage loss. In: Keuttner KE, Goldberg VM (Eds.) Osteoarthritic Disorders. Rosemont: American Academy of Orthopaedic Surgeons, 1995: 271-9.
210. Evans CH, Mazzocchi RA, Nelson DD, Rubash HE. Experimental arthritis induced by intraarticular injection of allogenic cartilaginous particles into rabbit knees. Arthritis Rheum 1984; 27: 200-7.
211. Evans DM, Ralston SH. Nitric oxide and bone. J Bone Miner Res 1996; 11: 300-5.
212. Evêquoz V, Schnyder J, Trechsel U, Baggiolini M, Fleisch H. Influence of macrophage products on the release of plasminogen activator, collagenase, ß-glucoronidase and prostaglandin E2 by articular chondrocytes. Biochem J 1984; 219: 667-77.
213. Fairbanks UF. Hemoglobin, hemoglobin derivatives, and myoblobin. In: Teitz NW (Ed.) Fundamentals of Clinical Chemistry. Philadelphia: WB Saunders, 1976: 401-54.
214. Farahat M, Yanni G, Poston R, Panayi G. Cytokine expression in synovial membranes of patients with rheumatoid arthritis and osteoarthritis. Ann Rheum Dis 1993; 52: 870-5.

244.
215. Farrell AJ, Blake DR. Nitric oxide. Ann Rheum Dis 1996; 55: 7-20.
216. Farrell AJ, Blake DR, Palmer RMJ, Moncada S. Increased concentration of nitrite in synovial fluid and serum samples suggest increased nitric oxide synthesis in rheumatic diseases. Ann Rheum Dis 1992; 51: 1219-22.
217. Farrugia W, Fortune C, HEath J, Caple I, Wark J. Osteocalcin as an index of osteoblast function during and after ovine pregnancy. Endocrinol 1989; 125: 1705-10.
218. Farrugia W, Yates N, Fortune C, McDugall J, Scoggins B, Wark J. The effect of uninephrectomy on osteocalcin metabolism in sheep: a direct evaluation of renal osteocalcin clearance. J Endocrinol 1991; 130: 213-21.
219. Fazzalari NL, Kuliwaba JS, Atkins GJ, Forwood MR, Findlay DM. The ratio of messenger RNA levels of receptor activator of nuclear factor kappa ß ligand to osteoprotegerin correlates with bone remodelling indices in normal human cancellous bone but not in osteoarthritis. J Bone Miner Res 2001; 16: 1015-27.
220. Feldmann M, Brennan FM, Maini RN. Role of cytokines in rheumatoid arthritis. Ann Rev Immun 1996; 14: 397-440.
221. Fell HB. Synoviocytes. J Clin Pathol 1978; 12 (Suppl):14-24.
222. Fell H, Jubb R. The effect of synovial tissue on the breakdown of articular cartilage in organ cultures. Arthritis Rheum 1977; 20: 1359-71.
223. Fell HB, Lawrence CE, Bagga MR, Glauert AM. The promotion and inhibition of collagen-breakdown in organ cultures of pig synovium: the requirement for serum components and the involvement of cyclic adenosine 3':5'-monophosphate (cAMP). Collagen Rel Res 1986; 6: 51-75.
224. Felson DT. Epidemiology of hip and knee osteoarthritis. Epidemiol Rev 1988; 10: 1-28.
225. Felson DT, Nevitt MC. The effects of estrogen on osteoarthritis. Curr Opin Rheumatol 1998; 10: 269-72.
226. Felson D, Nevitt M. Estrogen and osteoarthritis: How do we explain conflicting study results? (Editorial). Prev Med 1999; 28: 445-8.
227. Fernihough JK, Richmond RS, Carlson CS, Cherpes T, Holly JMP, Loeser RF. Estrogen replacement therapy modulation of the insulin-like growth factor system in monkey knee joints. Arthritis Rheum 1999; 42: 2103-11.
228. Fibbi G, Serni U, Matucci A, Mannoni A, Pucci M, Anichini E, et al. Control of the chondrocyte fibrinolytic balance by the drug piroxicam: Relevance to the osteoarthritic process. J Rheumatol 1994; 21: 2322-28.
229. Field M, Chu C, Feldmann M, Maini RN. Interleukin-6 localization in the synovial membrane in rheumatoid arthritis. Rheumatol Int 1991; 11: 45-50.
230. Fiorucci S, Santucci L, Cirino G, Mencarelli A, Familiari L, Soldato PD, et al. IL-1 beta converting enzyme is a target for nitric oxide-releasing aspirin: new insights in the antiinflammatory mechanism of nitric oxide-releasing non-steroidal antiinflammatory drugs. J Immunol 2000; 165: 5242-54.
231. Firestein G. Invasive fibroblast-like synoviocytes in rheumatoid arthritis. Passive responders or transformed aggressors? Arthritis Rheum 1996; 39: 1781-90.
232. Firestein G, Paine M, Littman B. Gene expression (collagenase, tissue inhibitor of

245.
metalloproteinases, complement, and HLA-DR) in rheumatoid arthritis and osteoarthritis synovium - Quantitative analysis and effect of intraarticular corticosteroids. Arthritis Rheum 1991; 34: 1094-105.
233. Fischer E, Van Zee K, Marano M, Rock C, Kenney J, Poutsiaka D, et al. Interleukin-1 receptor antagonist circulates in experimental inflammation and in human disease. Blood 1992; 79: 2196-200.
234. Flannery CR, Little CB, Hughes CE, Curtis CL, Caterson B, Jones SA. IL-6 and its soluble receptor augment aggrecanase-mediated proteoglycan catabolism in articular cartilage. Matrix Biol 2000; 19: 549-53.
235. Fontenele JB, Araúijo GB, Alencar JW, Viana GSD. The analgesic and anti-inflammatory effects of shark cartilage are due to a peptide molecule and are nitric oxide (NO) system dependent. Biol Pharm Bull 1997; 20: 1151-4.
236. Fortune C, Farrugia W, Tresham J, Scoggins B, Wark J. Hormonal regulation of osteocalcin plasma production and clearance in sheep. Endocrinol 1989; 124: 2785-90.
237. Fraenkel L, Zhang Y, Trippel SB, McAlindon TE, LaValley MP, Assif A, et al. Longitudinal analysis of the relationship between serum insulin-like growth factor-I and radiographic knee osteoarthritis. Osteoarthritis Cartilage 1998; 6: 362-7.
238. Francis D, Hutadilok N, Kongtawelert P, Ghosh P. Pentosan polysulphate and glycosaminoglycan polysulphate stimulate the synthesis of hyaluronan in vivo. Rheumatol Int 1993; 13: 61-4.
239. Francis DJ, Read RA. Pentosan polysulfate as a treatment for osteoarthritis (degenerative joint disease) in dogs. Aust Vet Practit 1993; 23: 104-9.
240. Franck H, Ittel T. Serum osteocalcin levels in patients with psoriatic arthritis: an extended report. Rheumatol Int 2000; 19: 161-4.
241. Fraser J, McCall J. Culture of synovial cells: in vitro notes on isolation and propagation. Ann Rheum Dis 1965; 24: 351-9.
242. Fraser J, Robinson A, Clarris B. Brooks P, York J. Synovial cell interactions in vitro In: Brooks PM, York JR (Eds.). Rheumatology – 85. Elsevier Science Publishers, 1985: 65-71
243. Freemont A. The pathophysiology of cartilage and synovium. Br J Rheumatol 1996; 35 (Suppl 3): 10-3.
244. Freemont A, Hampson V, Tilman R, Goupille P, Taiwo Y, Hoyland J. Gene expression of matrix metalloproteinase-1, metalloproteinase-3, and metalloproteinase-9 by chondrocytes in osteoarthritic human knee articular cartilage is zone and grade specific. Ann Rheum Dis 1997; 56: 542-9.
245. Frisbie DD, Nixon AJ. Insulin-like growth factor 1 and corticosteroid modulation of chondrocyte metabolic and mitogenic activities in interleukin 1-conditioned equine cartilage. Am J Vet Res 1997; 58: 524-30.
246. Frost L, Ghosh P. Microinjury to the synovial membrane may cause disaggregation of proteoglycans in rabbit knee joint articular cartilage. J Orthop Res 1984; 2: 207-20.
247. Fukumura D, Gohongi T, Kadambi A, Izumi Y, Ang J, Yun CO, et al. Predominant role of endothelial nitric oxide synthase in vascular endothelial growth factor-induced angiogenesis and vascular permeability. Proc Natl Acad Sci USA 2001; 98: 2604-9.
248. Garnero P, Tsouderos Y, Marton I, Pelissier C, Varin C, Delmas PD. Effects of intranasal 17ß-

246.
estradiol on bone turnover and serum insulin-like growth factor I in postmenopausal women. J Clin Endocrinol Metab 1999; 84: 2390-7.
249. Gedikoglu O, Bayliss MT, Ali SY, Tuncer I. Biochemical and histological changes in osteoarthritic synovial membrane. Ann Rheum Dis 1986; 45: 289-92.
250. Georgescu HI, Mendelow D, Evans CH. HIG-82: an established cell line from rabbit periarticular soft tissue, which retains the "activatable" phenotype. In Vitro Cell Dev Biol 1988; 24:1015-22.
251. Gerrits M, Thijssen J, van Rijn H. Determination of pyridinoline and deoxypyridinoline in urine, with special attention to retaining their stability. Clin Chem 1995; 41: 571-4.
252. Gershuni D, Amiel D, Gonsalves M, Akeson W. The biochemical response of rabbit articular cartilage matrix to an induced talcum synovitis. Acta Orthop Scand 1981; 52: 599-603.
253. Getzy LL, Malemud CJ, Goldberg VM, Moskowitz RW. Factors influencing metachromatic staining in paraffin-embedded sections of rabbit and human articular cartilage: A comparison of the safranin O and toluidine blue techniques. J Histotechnol 1982; 5: 111-6.
254. Ghivizzani S, Kang R, Georgescu H, Lechman E, Jaffurs D, Engle J, et al. Constitutive intra-articular expression of human IL-1b following gene transfer to rabbit synovium produces all major pathologies of human rheumatoid arthritis. J Immunol 1997; 159: 3604-12.
255. Ghosh P. Anti-rheumatic drugs and cartilage In: Brooks PM (Ed.) Bailliere's Clinical Rheumatology International Practice and Research. London: Bailliere Tindall, 1988; 309-38.
256. Ghosh P. The role of hyaluronic acid (hyaluronan) in health and disease: Interactions with cells, cartilage and components of synovial fluid. Clin Exp Rheumatol 1994; 12: 75-82.
257. Ghosh P. The pathobiology of osteoarthritis and the rationale for the use of pentosan polysulfate for its treatment. Semin Arthritis Rheum 1999; 28: 211-67.
258. Ghosh P, Armstrong S, Read R, Numata Y, Smith S, McNair P, et al. Animal models of early osteoarthritis: Their use for the evaluation of potential chondroprotective agents. In: Van den Berg W, van der Kraan PM, van Lent PLEM (Eds.) Joint Destruction in Arthritis and Osteoarthritis Basel:Birkhaüser Verlag, 1993:195-206.
259. Ghosh P, Brooks P. Chondroprotection - Exploring the concept. J Rheumatol 1991; 18:161-6.
260. Ghosh P, Burkhardt D, Read R, Bellenger C. Recent advances in animal models for evaluating chondroprotective drugs. J Rheumatol 1991; 18:143-6.
261. Ghosh P, Collier S, Andrews J. Synovial membrane-cartilage interactions: The role of serine proteinase inhibitors in interleukin-l mediated degradation of articular cartilage. J Rheumatol 1987; 14:122-4.
262. Ghosh P, Holbert C, Read R, Armstrong S, Wilson D. Chondroprotection and NSAIDs. Lancet 1991; 337: 1612-3.
263. Ghosh P, Hutadilok N. Interactions of pentosan polysulfate with cartilage matrix proteins and synovial fibroblasts derived from patients with osteoarthritis. Osteoarthritis Cartilage 1996; 4: 43-53.
264. Ghosh P, Numata Y, Smith S, Read R, Armstrong S, Johnson K. The metabolic response of articular cartilage to abnormal mechanical loading induced by medial or lateral meniscectomy. In: Van den Berg W, van der Kraan PM, van Lent PLEM (Eds.) Joint Destruction in Arthritis and Osteoarthritis. Basel:Birkhaüser Verlag, 1993: 89-93.

247.
265. Ghosh P, Sutherland J, Bellenger C, Read R, Darvodelsky A. The influence of weight-bearing exercise on articular cartilage of meniscectomized joints - An experimental study in sheep. Clin Orthop 1990; 252: 101-13.
266. Ghosh S, Seshadri R. Expression of tumor cell properties in synovial cells in culture. Acta Cytologica 1986; 31: 77-80.
267. Girasole G, Jilka RL, Passeri G, Boswell S, Boder G, Williams DC. 17-beta-estradiol inhibits interleukin-6 production by bone marrow-derived stromal cells and osteoblasts in vitro-a potential mechanism for the antiosteoporotic effect of estrogen. J Clin Invest 1992; 89: 883-91.
268. Glansbeek HL, Vanbeuningen HM, Vitters EL, Van der Kraan PM, Van den Berg WB. Stimulation of articular cartilage repair in established arthritis by local administration of transforming growth factor beta into murine knee joints. Lab Invest 1998; 78: 133-42.
269. Goad DL, Rubin J, Wang H, Tashjian AH, Patterson C. Enhanced expression of vascular endothelial growth factor in human SaOS-2 osteoblast-like cells and murine osteoblasts induced by insulin-like growth factor I. Endocrinol 1996; 137: 2262-8.
270. Goldberg GI, Marmer BL, Grant GA, Eisen AZ, Wilhelm S, He CS. Human 72-kilodalton type-IV collagenase forms a complex with a tissue inhibitor of metalloproteases designated TIMP-2. Proc Natl Acad Sci USA 1989; 86: 8207-11.
271. Golding JC, Ghosh P. Drugs for osteoarthritis II: Glycosaminoglycan polysulphate ester (Arteparon) on proteoglycan loss and aggregation from articular cartilage of immobilized rabbit knee joints. Curr Therap Res 1983; 34: 67-80.
272. Goldring MB, Berenbaum F. Human chondrocyte culture models for studying cyclooxygenase expression and prostaglandin regulation of collagen gene expression. Osteoarthritis Cartilage 1999; 7: 386-8.
273. Goldring MB, Krane SM. Modulation by recombinant interleukin 1 of synthesis of types I and III collagens and associated procollagen mRNA levels in cultured human cells. J Biol Chem 1987; 262: 16724-9.
274. Goldring MB, Sohbat E, Elwell JM, Chang JY. Etodolac preserves cartilage-specific phenotype in human chondrocytes: effects on type II collagen synthesis and associated mRNA levels. Eur J Rheumatol Inflamm 1990; 10: 10-21.
275. Golds E, Santer V, Killackey J, Roughley P. Mononuclear cell factors stimulate the concomitant secretion of distinct latent proteoglycan, gelatin and collagen degrading enzymes from human skin fibroblasts and synovial cells. J Rheumatol 1983; 10: 861-71.
276. Goto M, Okamoto M, Sasano M, Yanagisawa S, Miyamoto T, Nishioka K, et al. Adherent synovial cells from nonrheumatoid arthritis do not release interleukin 1-beta and prostaglandin E2 spontaneously in longterm culture. J Rheumatol 1990; 17: 1299-302.
277. Goto M, Sasano M, Yamanaka H, Miyasaka N, Kamatani N, Inoue K, et al. Spontaneous production of an interleukin-1-like factor by cloned rheumatoid synovial-cells in long-term culture. J Clin Invest 1987; 80: 786-96.
278. Gouze JN, Bordji K, Gulberti S, Terlain B, Netter P, Magdalou J, et al. Interleukin-1ß down-regulates the expression of glucoronosyltransferase I, a key enzyme priming glycosaminoglycan biosynthesis: influence of glucosamine on interleukin-1ß-mediated effects in rat chondrocytes. Arthritis Rheum 2001; 44: 351-60.
279. Graabaek P. Characteristics of the types of synoviocytes in rat synovial membrane - an ultrastructural study. Lab Invest 1984; 50: 690-702.

248.
280. Graabek P. Ultrastructural evidence for two distinct types of synoviocytes in rat synovial membrane. J Ultras Res 1982; 78: 321-39.
281. Grabowski PS, England AJ, Dykhuizen R, Copland M, Benjamin N, Reid DM, et al. Elevated nitric oxide production in rheumatoid arthritis. Detection using the fasting urinary nitrate:creatinine ratio. Arthritis Rheum 1996; 39: 643-7.
282. Grabowski PS, Wright PK, Vanthof RJ, Helfrich MH, Ohshima H, Ralston SH. Immunolocalization of inducible nitric oxide synthase in synovium and cartilage in rheumatoid arthritis and osteoarthritis. Br J Rheumatol 1997; 36:651-5.
283. Granados-Soto V, Flores-Murrieta FJ, Castaneda-Hernadez G, Lopez-Munoz FJ. Evidence for the involvement of nitric oxide in the antinociceptive effect of ketorolac. Eur J Pharmacol 1995; 24: 281-4.
284. Gravallese EM, Darling JM, Ladd AL, Katz JN, Glimcher LH. In situ hybridization studies of stromelysin and collagenase messenger RNA expression in rheumatoid synovium. Arthritis Rheum 1991; 34: 1076-84.
285. Gravallese EM, Manning C, Tsay A, Naito A, Pan C, Amento E, et al. Synovial tissue in rheumatoid arthritis is a source of osteoclast differentiation factor. Arthritis Rheum 2000; 43: 250-8.
286. Green LC, Wagner DA, Glogowski J, Skipper PL, Wishnok JS, Tannenbaum SR. Analysis of nitrate, nitrite and [15N]nitrate in biological fluids. Anal Biochem 1982; 126: 131-8.
287. Greenfield EM. ODF/OPGL expression is increased by IL-1 and IL-6. J Bone Miner Res 1999; 14 (Suppl 1): S361 (Abstract).
288. Greisen H, Lust G, Summers B. A morphological study of the synovial membrane and articular cartilage in the early stages of osteoarthritis in canine hip joints. Semin Arthritis Rheum 1981; 11 (Suppl 1): 50-1.
289. Greisen HA, Summers BA, Lust G. Ultrastructure of the articular cartilage and synovium in the early stages of degenerative joint disease in canine hip joints. Am J Vet Res 1982; 43: 1963-71.
290. Grier SJ, Turner AS, Alvis MR. The use of dual-energy X-ray absorptiometry in animals. Invest Radiol 1996; 31: 50-62.
291. Grimbacher B, Aicher W, Peter H, Eibel H. Measurement of transcription factor c-fos and EGR-1 mRNA transcription levels in synovial tissue by quantitative RT-PCR. Rheumatol Int 1997; 17: 109-12.
292. Grisham MB, Jourd'Heuil D, Wink DA. Nitric oxide. I. Physiological chemistry of nitric oxide and its metabolites: Implications in inflammation. Am J Physiol 1999; 39: G315-21
293. Grynpas MD, Alpert B, Katz I, Lieberman I, Pritzker KP. Subchondral bone in osteoarthritis. Calcif Tissue Int 1991; 49: 20-6.
294. Guan Z, Buckman SY, Baier LD, Morrison AR. IGF-I and insulin amplify IL-1ß- induced nitric oxide and prostaglandin biosynthesis. Am J Physiol 1998; 274: F673-9.
295. Guerne PA, Carson DA, Lotz M. IL-6 production by human articular chondrocytes: modulation of its synthesis by cytokines, growth-factors, and hormones in vitro. J Immunol 1990; 144: 499-505.
296. Guerne P, Zuraw B, Vaughan J, Carson D, Lotz M. Synovium as a source of interleukin 6 in vitro - contribution to local and systemic manifestations of arthritis. J Clin Invest 1989; 83: 585-92.

249.
297. Guevara I, Iwanejko J, Dembi´nska-Kie´c A, Pankiewicz J, Wanat A, Anna P, et al. Determination of nitrite/nitrate in human biological material by the simple Greiss reaction. Clin Chim Acta 1998; 274: 177-88.
298. Gustafsson J. Therapeutic potential of selective estrogen receptor modulators. Curr Opin Chem Biol 1998; 2: 508-11.
299. Hacker S, Healey R, Yoshioka M, Coutts R. A methodology for the quantitative assessment of articular cartilage histomorphometry. Osteoarthritis Cartilage 1997; 5: 343-55.
300. Hamerman D, Smith C, Keiser HD, Craig R. Glycosaminoglycans produced by human synovial cell cultures. Coll Rel Res 1982; 2: 313-29.
301. Hamerman D, Wood D. Interleukin I enhances synovial cell hyaluronate synthesis. Proc Soc Exp Biol Med 1984; 177: 205-210.
302. Hamilton JA. Plasminogen activator activity of rheumatoid and nonrheumatoid synovial fibroblasts. J Rheumatol 1982; 9: 834-42.
303. Hamilton JA, Bootes A, Phillips PE, Slywka J. Human synovial fibroblast plasminogen activator: Modulation of enzyme activity by anti-inflammatory steroids. Arthritis Rheum 1981; 24: 1296-1303.
304. Hamilton JA, Butler DM, Stanton H. Cytokine interactions promoting DNA synthesis in human synovial fibroblasts. J Rheumatol 1994; 21: 797-803.
305. Hamilton J, Clarris B, Fraser J, Niall M. Peripheral blood mononuclear cells stimulate prostacyclin levels of human synovial fibroblast-like cells. Rheumatol Int 1985; 5: 121-5.
306. Hamilton J, Leizer T, Lingelbach S. The stimulation of human synovial fibroblast plasminogen activator activity. The involvement of cyclic AMP and cyclooxygenase products. Biochim Biophys Acta 1986; 886: 195-202.
307. Hamilton JA, Piccoli DS, Leizer T, Butler DM, Croatto M, Royston AKM. Transforming growth factor beta stimulates urokinase-type plasminogen activator and DNA synthesis, but not prostaglandin-E2 production, in human synovial fibroblasts. Proc Natl Acad Sci USA 1991; 88: 7180-4.
308. Hamilton J, Slywka J. Stimulation of human synovial fibroblast plasminogen activator production by mononuclear cell supernatants. J Immunol 1981; 126: 851-6.
309. Hamilton JA, Waring PM, Filonzi EL. Induction of leukemia inhibitory factor in human synovial fibroblasts by IL-1 and tumor necrosis factor-alpha. J Immunol 1993; 150: 1496-502.
310. Hamilton J, Zabriskie J, Lachman L, Chen Y-S. Streptococcal cell walls and synovial cell activation. J Exp Med 1982; 155: 1702-18.
311. Hanemaaijer R, Sorsa T, Konttinen YT, Ding YL, Sutinen M, Visser H, et al. Matrix metalloproteinase-8 is expressed in rheumatoid synovial fibroblasts and endothelial cells: Regulation by tumor necrosis factor alpha and doxycycline. J Biol Chem 1997; 272: 1504-9.
312. Hannan M, Anderson J, Zhang Y, Levy D, Felson D. Bone mineral density and knee osteoarthritis in elderly men and women: The Framingham study. Arthritis Rheum 1993; 36: 1671-80.
313. Hannan N, Ghosh P, Bellenger C, Taylor T. Systemic administration of glycosaminoglycan polysulphate (Arteparon) provides partial protection of articular cartilage from damage produced by meniscectomy in the canine. J Orthop Res 1987; 5: 47-59.

250.
314. Hardingham TE, Fosang AJ, Dudhia J. The structure, function and turnover of aggrecan, the large aggregating proteoglycan from cartilage. Eur J Clin Chem Clin Biochem 1994; 32: 249-57.
315. Hardingham TE, Muir H. The specific interactions of hyaluronic acid with cartilage proteoglycans. Biochim Biophys Acta 1972; 279: 401-5.
316. Hardingham TE, Venn G. Chondroitin sulphate/dermatan sulphate proteoglycans from cartilage: aggrecan, decorin and biglycan. In: Scott JE (Ed.) Dermatan Sulphate Proteoglycans. Chemistry, Biology, Chemical Pathology. London: Portland Press, 1993:207-7.
317. Harmand MF. Etude de l'action des insaponfiables d'avocat et de soja sur les cultures de chondrocytes articulaires. Gaz Med Fr 1985; 92: 1-3.
318. Hart D, Doyle D, Spector T. Incidence and risk factors for radiographic knee osteoarthritis in middle-aged women. Arthritis Rheum 1999; 42: 17-24.
319. Hart D, Mootoosamy I, Doyle D, Spector T. The relationship between osteoarthritis and osteoporosis in the general population: the Chingford study. Ann Rheum Dis 1994; 53: 158-62.
320. Hart T, Lamont A, Williams D. New drug targets in inflammation and immunomodulation. Drug Disc T 1998; 3: 516-21.
321. Hashimoto S, Setareh M, Ochs RL, Lotz M. Fas/Fas ligand expression and induction of apoptosis in chondrocytes. Arthritis Rheum 1997; 40: 1749-55.
322. Hashimoto S, Takahashi K, Amiel D, Coutts RD, Lotz M. Chondrocyte apoptosis and nitric oxide production during experimentally-induced osteoarthritis. Arthritis Rheum 1998; 41: 1266-74.
323. Hasty KA, Reife RA, Kang AH, Stuart JM. The role of stromelysin in the cartilage destruction that accompanies inflammatory arthritis. Arthritis Rheum 1990; 33: 388-97.
324. Hasunuma T, Kato T, Kobata T, Nishioka K. Molecular mechanism of immune-response, synovial proliferation and apoptosis in rheumatoid arthritis. Spr Sem Imm 1998; 20: 41-52.
325. Häuselmann HJ, Flechtenmacher J, Michal L, Thonar EJ-MA, Shinmei M, Kuettner KE, et al. The superficial layer of human articular cartilage is more susceptible to interleukin-1-induced damage than the deeper layers. Arthritis Rheum 1996; 39: 478-88.
326. Häuselmann HJ, Oppliger L, Michel BA, Stefanovic-Racic M, Evans CH. Nitric oxide and proteoglycan biosynthesis by human articular chondrocytes in alginate culture. FEBS Lett 1994; 352: 361-4.
327. Hauselmann HJ, Stefanovic-Racic M, Michel BA, Evans CH. Differences in nitric oxide production by superficial and deep human articular chondrocytes: Implications for proteoglycan turnover in inflammatory joint diseases. J Immunol 1998; 160: 1444-8.
328. Hay CW, Chu QL, Budsberg SC, Clayton MK, Johnson KA. Synovial fluid interleukin 6, tumor necrosis factor, and nitric oxide values in dogs with osteoarthritis secondary to cranial cruciate ligament rupture. Am J Vet Res 1997; 58: 1027-32.
329. Hayashi T, Abe E, Yamate T, Taguchi Y, Jasin HE. Nitric oxide production by superficial and deep articular chondrocytes. Arthritis Rheum 1997; 40: 261-9.
330. Hayashi T, Fukuto J, Ignarro L, Chaudhuri G. Basal release of nitirc oxide from aortic rings is greater in female rabbits than in male rabbits: implications for atherosclerosis. Proc Natl Acad Sci USA 1992; 89: 11259-63.
331. Hayes WC, Keer LM, Herrmann G, Mockros LF. A mathematical analysis for indentation tests of

251.
articular cartilage. J Biomech 1972; 5: 541-51.
332. Haylor J, Singh I, el Nahas AM. Nitric oxide synthesis inhibitor prevents vasodilation by insulin-like growth factor I. Kidney Int 1991; 39: 333-5.
333. Heinegård D, Tiderström G. Determination of serum creatinine by direct colorimetric method. Clin Chim Acta 1973; 43: 305-10.
334. Heino J, Viander M, Kouri T. Morphologic alterations in cultured human synovial fibroblasts induced by blood mononuclear cells. Exp Cell Biol 1984; 52: 347-54.
335. Hellio Le Graverand MP, Reno C, Hart DA. Influence of pregnancy on gene expression in rabbit articular cartilage. Osteoarthritis Cartilage 1998; 6: 341-50.
336. Hembry R, Bagga M, Reynolds J, Hamblen D. Immunolocalization studies on six matrix metalloproteinases and their inhibitors, TIMP-1 and TIMP-2, in synovia from patients with osteoarthritis and rheumatoid arthritis. Ann Rheum Dis 1995; 54: 25-32.
337. Henderson B. The biochemistry of the human synovial lining with special reference to alterations in metabolism in rheumatoid arthritis. Path Res Pract 1981; 172: 1-24.
338. Henderson B, Higgs G. Synthesis of arachidonate oxidation products by synovial joint tissue during the development of chronic erosive arthritis. Arthritis Rheum 1987; 30: 1149-56.
339. Henderson B, Pettipher E. The synovial lining cell - biology and pathobiology. Semin Arthritis Rheum 1985; 15: 1-32.
340. Henderson B, Pettipher ER. Arthritogenic actions of recombinant IL-1 and tumour necrosis factor a in the rabbit: evidence for synergistic interactions between cytokines in vivo. Clin Exp Immunol 1989; 75: 306-10.
341. Henderson KJ, Edwards JCW, Worrall JG. Expression of CD44 in normal and rheumatoid synovium and cultured synovial fibroblasts. Ann Rheum Dis 1994; 53: 729-34.
342. Hendler P, Lavoie P, Werb Z, Chan J, Seaman W. Human synovial dendritic cells: Direct observation of transition to fibroblasts. J Rheumatol 1985; 12: 660-4.
343. Henrotin Y, Bassleer C, Franchimont P. In vitro effects of etodolac and acetylsalicyclic acid on human chondrocyte metabolism. Agents Actions 1992; 36: 317-23.
344. Henrotin Y, De Groote D, Labasse A, Gaspar S, Zheng S-X, Geenen V, et al. Effects of exogenous IL-1ß, TNF- , IL-6, IL-8 and LIF on cytokine production by human articular chondrocytes. Osteoarthritis Cartilage 1996; 4: 163-73.
345. Henrotin Y, Labasse A, Jaspar J, Degroote D, Zheng S, Guillou G, et al. Effects of 3 avocado/soybean unsaponifiable mixtures on metalloproteinases, bytokines and prostaglandin E2 production by human articular chondrocytes. Clin Rheum 1998; 17: 31-9.
346. Henrotin YE, Zheng SX, Deby GP, Labasse AH, Crielaard JM, Reginster JY. Nitric oxide down-regulates interleukin-1-beta (IL-1-beta) stimulated IL-6, IL-8, and prostaglandin E2 production by human chondrocytes. J Rheumatol 1998; 25: 1595-601.
347. Herman JH, Appel AM, Hess EV. Modulation of cartilage destruction by select nonsteroidal antiinflammatory drugs - in vitro effect on the synthesis and activity of catabolism-inducing cytokines produced by osteoarthritic and rheumatoid synovial tissue. Arthritis Rheum 1987; 30: 257-65.
348. Herman J, Appel A, Khosla R, Hess E. In vitro effect of select nonsteroidal antiinflammatory

252.
drugs on the synthesis and activity of anabolic regulatory factors produced by osteoarthritic and rheumatoid synovial tissue. J Rheumatol 1989; 16: 75-81.
349. Hickery MS, Bayliss MT. Interleukin-1 induced nitric oxide inhibits sulphation of glycosaminoglycan chains in human articular chondrocytes. Biochim Biophys Acta 1998; 1425: 282-90.
350. Hilal G, Martel-Pelletier J, Pelletier J-P, Ranger P, Lajeunesse D. Osteoblast-like cells from human subchondral osteoarthritic bone demonstrate an altered phenotype in vitro: Possible role in subchondral bone sclerosis. Arthritis Rheum 1998; 41: 891-9.
351. Hilal G, Massicotte F, Martel-Pelletier J, Fernandes JC, Pelletier JP, Lajeunesse D. Endogenous prostaglandin E2 and insulin-like growth factor 1 can modulate the levels of parathyroid hormone receptor in human osteoarthritis osteoblasts . J Bone Miner Res 2001; 16: 713-21.
352. Hilliquin P, Borderie D, Hernvann A, Menkes C, Ekindjian O. Nitric oxide stimulates tumor necrosis factor production by synoviocytes in osteoarthritis. Osteoarthritis Cartilage 1997; 5:43-5
353. Hirai Y, Migita K, Honda S, Ueki Y, Yamasaki S, Urayama S, et al. Effects of nitric oxide on matrix metalloproteinase-2 production by rheumatoid synovial cells. Life Sci 2001; 68: 913-20.
354. Hochberg MC, Lethbridge-Cejku M, Scott WW, Reichle R, Plato CC, Tobin JD. Serum levels of insulin-like growth factor in subjects with osteoarthritis of the knee. Data from the Baltimore Longitudinal Study of Ageing. Arthritis Rheum 1994; 37: 1177-80.
355. Hofbauer LC, Heufelder AE, Erben RG. Osteoprotegrin, RANK and RANK ligand: The good, the bad, and the ugly in rheumatoid arthritis. J Rheumatol 2001; 28: 285-7.
356. Horowitz M. Cytokines and estrogen in bone: Anti-osteoporotic effects. Science 1993; 260: 626-7.
357. Houpt J, McMillan R, Wein C, Paget-Dellio S. Effect of glucosamine hydrochloride in the treatment of pain of osteoarthritis of the knee. J Rheumatol 1999; 26: 2423-30.
358. Houssiau FA, Devogelaer J-P, Van Damme J, de Deuxchaisnes CN, Van Snick J. Interleukin-6 in synovial fluid and serum of patients with rheumatoid arthritis and other inflammatory arthritides. Arthritis Rheum 1988; 31: 784-8.
359. Howe LM, Boothe HW. Nitric oxide: A review for veterinary surgeons. Vet Surg 2001; 30: 44-57.
360. Hrabak A, Vercruysse V, Kahan IL, Vray B. Indomethacin prevents the induction of inducible nitric oxide synthase in murine peritoneal macrophages and decreases their nitric oxide production. Life Sci 2001; 68: 1923-30.
361. Hu S-K, Mitcho Y, Rath N. Effects of estradiol on interleukin 1 synthesis by macrophages. Int J Immunopharmac 1988; 10: 247-52.
362. Hukkanen M, Hughes FJ, Buttery LD, Gross SS, Evans TJ, Seddon S, et al. Cytokine-stimulated expression of inducible nitric oxide synthase by mouse, rat, and human osteoblast-like cells and its functional role in osteoblast metabolic activity. Endocrinol 1995; 136: 5445-53.
363. Hulth A. Does osteoarthrosis depend on growth of the mineralized layer of cartilage? Clin Orthop 1993; 287: 19-24.
364. Hulth A, Lindberg L, Telhag H. Experimental osteoarthritis in rabbits. Acta Orthop Scand 1970; 41: 522-30.
365. Hutadilok N, Smith M, Cullis-Hill D, Ghosh P. Pentosan polysulphate stimulates hyaluronate and DNA synthesis in synovial fibroblasts and partially reduces the suppressive effect of

253.
hydrocortisone on fibroblast metabolism. Curr Ther Res 1988; 44: 845-60.
366. Hutchinson N, Lark M, Macnaul K, Harper C, Hoerrner L, Mcdonnell J, et al. In vivo expression of stromelysin in synovium and cartilage of rabbits injected intraarticularly with interleukin-1-beta. Arthritis Rheum 1992; 35: 1227-33.
367. Hwa S, Smith M, Burkhardt D, Ghosh P. Oestrogen depletion provokes significant vascular and matrix changes in the calcified cartilage and subchondral bone of the femoral-tibial joints of aged ewes. Trans Orthop Res Soc 2001: 373 (Abstract)
368. Hwa SY, Burkhardt D, Little C, Ghosh P. The effects of orally administered diacerein on cartilage and subchondral bone in an ovine model of osteoarthritis. J Rheumatol 2001; 28: 825-34.
369. Imamura F, Aono H, Hasunuma T, Sumida T, Tateishi H, Maruo S, et al. Monoclonal expansion of synoviocytes in rheumatoid arthritis. Arthritis Rheum 1998; 41: 1979-86.
370. Imthurn B, Rosselli M, Jaeger A, Keller P, Dubey R. Differential effects of hormone-replacement therapy on endogenous nitric oxide (nitrite/nitrate) levels in postmenopausal women substituted with 17ß-estradiol valerate and cyproterone acetate or medroxyprogesterone acetate. J Clin Endocrinol Metab 1997; 82: 388-94.
371. Iniguez MA, Pablos JL, Carreira PE, Cabré F, Gomez-Reino JJ. Detection of COX-1 and COX-2 isoforms in synovial fluid cells from inflammatory joint diseases. Br J Rheumatol 1998; 37:773-8.
372. Innes J, Sharif M, Barr A. Relations between biochemical markers of osteoarthritis and other disease parameters in a population of dogs with naturally acquired osteoarthritis of the genual joint. Am J Vet Res 1998; 59: 1530-6.
373. Inoue K, Motonaga A, Suzuka H, Yoshifusa H, Fujisawa H, Nishimura T, et al. Effect of etodolac on type-II collagen-induced arthritis . Agents Actions 1993; 39: 187-94.
374. Islas-Cadena M, Aguirre-Banuelos P, Granados-Soto V. Evidence for the participation of the nitric oxide-cyclic GMP pathway in the antinociceptive effect of nimesulide. J Pharmacol Toxicol Methods 1999; 42: 87-92.
375. Ito A, Itoh Y, Sasaguri Y, Morimatsu M, Mori Y. Effects of interleukin-6 on the metabolism of connective tissue components in rheumatoid synovial fibroblasts. Arthritis Rheum 1992; 35: 1197-201.
376. Izumi S, Takeya M, Takagi K, Takahashi K. Ontogenetic development of synovial A cells in fetal and neonatal rat knee joints. Cell Tissue Res 1990; 262: 1-8.
377. Jackson J, Minton J, Ho M, Wei N, Winkler J. Expression of vascular endothelial growth factor in synovial fibroblasts is induced by hypoxia and interleukin-1b. J Rheumatol 1997; 24: 1253-9.
378. Jacob T, Morrell M, Manzi S, Verdile V, Simmons RL, Peitzman A. Elevated nitrates in inflammatory joint disease: nitric oxide in the arthritides. Arthritis Rheum 1992; 35 (Suppl 5):R9 (Abstract)
379. Jamal OS, Conaghan PG, Cunningham AM, Brooks PM, Munro VF, Scott KF. Increased expression of human type IIa secretory phospholipase A2 antigen in arthritic synovium. Ann Rheum Dis 1998; 57: 550-8.
380. Jamal SA, Browner WS, Bauer DC, Cummings SR. Intermittent use of nitrates increases bone mineral density: The study of osteoporotic fractures. J Bone Miner Res 1998; 13: 1755-9.
381. Jang D, Murrell G. Nitric oxide in arthritis. Free Radic Biol Med 1998; 24: 1511-9.

254.
382. Janusz MJ, Hare M. Cartilage degradation by cocultures of transformed macrophage and fibroblast cell lines: A model of metalloproteinase-mediated connective tissue degradation. J Immunol 1993; 150: 1922-31.
383. Jarvinen TA, Moilanen T, Jarvinen TL, Moilanen E. Endogenous nitric oxide and prostaglandin E2 do not regulate the synthesis of each other in interleukin-1 beta-stimulated rat articular cartilage Inflammation 1996; 20: 683-92.
384. Jee WSS, Li XJ, Ke HZ, Li M, Smith RJ, Dunn CJ. Application of computer-based histomorphometry to the quantitative analysis of methylprednisolone-treated adjuvant arthritis in rats. Bone Miner 1993; 22: 221-47.
385. Jilka R, Hangoc L, Girasole Gea. Increased osteoclast development after estrogen loss: mediated by interleukin-6. Science 1992; 257: 88-91.
386. Johnson K, Jung A, Murphy A, Andreyev A, Dykens J, Terkeltaub R. Mitochondrial oxidative phosphorylation is a downstream regulator of nitric oxide effects on chondrocyte matrix synthesis and mineralisation. Arthritis Rheum 2000; 43: 1560-70.
387. Johnson RB, Gilbert JA, Cooper RC, Dai X, Newton BI, Tracy RR, et al. Alveolar bone loss one year following ovariectomy in sheep. J Periodontol 1997; 68: 864-71.
388. Johnston SA. Osteoarthritis: Joint anatomy, physiology, and pathobiology. Vet Clin North Am Small Anim Pract 1997; 27: 699-723
389. Johnston SA, Budsberg SC. Nonsteroidal anti-inflammatory drugs and corticosteroids for the management of canine osteoarthritis. Vet Clin North Am Small Anim Pract 1997; 27:841-62.
390. Jones G, Glisson M, Hynes K, Cicuttini. Sex and site differences in cartilage development. Arthritis Rheum 2000; 43: 2543-9.
391. Jouglin M, Robert C, Valette J-P, Gavard F, Quintin-Colonna F, Denoix J-M. Metalloproteinases and tumor necrosis factor-alpha activities in synovial fluids of horses: correlations with articular cartilage alterations. Vet Res 2000; 31: 507-15.
392. Jouvenne P, Vennier E, Dinarello C, Miossec P. Elevated levels of soluble interleukin-1 receptor type II and interleukin-1 receptor antagonist in patients with chronic arthritis. Correlations with markers of inflammation and joint destruction. Arthritis Rheum 1998; 41: 1083-9.
393. Jouzeau J-Y, Cippolletta C, Presle N, Netter P, Terlain B. Modulation of IL-1 effects on cartilage by NO synthase inhibitors. Osteoarthritis Cartilage 1999; 7: 382-5.
394. Jovanovic D, Pelletier J-P, Alaaeddine N, Mineau F, Geng C, Ranger P, et al. Effect of IL-13 on cytokines, cytokine receptors and inhibitors on human osteoarthritis synovium and synovial fibroblasts. Osteoarthritis Cartilage 1997; 6: 40-9.
395. Juma S, Munson ME, Malayer J, Svanborg A, Khalil DA, Arjmandi BH. Evidence for the presence of estrogen receptor beta but not alpha in human articular cartilage and meniscus. J Bone Miner Res 1999; 14 (Suppl 1): S452 (Abstract).
396. Junqueira LCU, Bignolas G, Brentani RR. Picrosirius staining plus polarization microscopy, specific method for collagen detection in tissue sections. Histochem J 1979; 11: 447-55.
397. Kafantari H, Kounadi E, Fatouros M, Milonakis M, Tzaphlidou M. Structural alterations in rat skin and bone collagen fibrils induced by ovariectomy. Bone 2000; 26: 349-53.
398. Kahlert S, Nuedling S, van Eickels M, Vetter H, Meyer R, Grohe C. Estrogen receptor alpha rapidly activates the IGF-1 receptor pathway. J Biol Chem 2000; 275: 18447-53.

255.
399. Kam GY, Leung KC, Baxter RC, Ho KK. Estrogens exert route- and dose-dependent effects on insulin-like growth factor (IGF)-binding protein-3 and the acid-labile subunit of the IGF ternary complex. J Clin Endocrinol Metab 2000; 85: 1918-22.
400. Kameda T, Mano H, Yuasa T, Mori Y, Miyazawa K, Shiokawa M, et al. Estrogen inhibits bone resorption by directly inducing apoptosis of the bone-resorbing osteoclasts. J Exp Med 1997; 186: 489-95.
401. Kaneko S, Satoh T, Chiba J, Ju C, Inoue K, Kagawa J. Interleukin-6 and interleukin-8 levels in serum and synovial fluid of patients with osteoarthritis. Cytokines Cell Mol Therapy 2000; 6:71-9
402. Kanematsu M, Ikeda K, Yamada Y. Interaction between nitric oxide synthase and cyclooxygenase pathways in osteoblastic MC3T3-E1 cells. J Bone Miner Res 1997; 12: 1789-96.
403. Karvonen RL, Miller PR, Nelson DA, Granda JL, Fernández-Madrid F. Periarticular osteoporosis in osteoarthritis of the knee. J Rheumatol 1998; 25: 2187-94.
404. Kassem M, Harris S, Spelsberg T, Riggs B. Estrogen inhibits interleukin-6 production and gene expression in a human osteoblastic cell line with high levels of estrogen receptors. J Bone Miner Res 1996; 11: 193-9.
405. Kasten TP, Collin-Osdoby P, Patel N, Krukowski M, Misko TP, Settle SL, et al. Potentiation of osteoclast bone-resorption activity by inhibition of nitric oxide synthase . Proc Natl Acad Sci USA 1994; 91: 3569-73.
406. Kauser K, Sonnenberg D, Tse J, Rubanyi G. 17b estradiol attenuates endotoxin-induced excessive nitrix oxide production in ovariectomized rats in vivo. Am J Physiol 1997; 42: H506-9
407. Kawaguchi H, Pilbeam CC, Harrison JR, Rasiz LG. The role of prostaglandins in the regulation of bone metabolism. Clin Orthop 1996; 313: 36-46.
408. Kawakami A, Eguchi K, Matsuoka N, Tsuboi M, Urayama S, Kawabe Y, et al. Inhibitory effects of interleukin-10 on synovial cells of rheumatoid arthritis. Immunol 1997; 91: 252-9.
409. Kawakami M, Suzuki K, Matsuki Y, Hidaka T, Ishizuka T, Konishi T, et al. Characterization of the newly established human type B synovial lining cell line, RAMAK-1: expression of adhesion molecules and cytokine production. Jap J Rheumatol 1998; 8: 401-9.
410. Kawakami M, Suzuki K, Matsuki Y, Ishizuka T, Hidaka T, Konishi T, et al. Hyaluronan production in human rheumatoid fibroblastic synovial lining cells is increased by interleukin 1-beta but inhibited by transforming growth factor beta-1. Ann Rheum Dis 1998; 57: 602-5.
411. Kawasaki T, Ushiyama T, Inoue K, Hukuda S. Effects of estrogen on interleukin-6 production in rheumatoid fibroblast-like synoviocytes. Clin Exp Rheumatol 2000; 18: 743-5.
412. Kawcak CE, McIllwraith CW, Norrdin RW, Park RD, James SP. The role of subchondral bone in joint disease: a review. Eq Vet J 2001; 33: 120-6.
413. Kay-Mugford P, Benn SJ, LaMarre J, Conlon P. In vitro effects of non-steroidal anti-inflammatory drugs on cyclooxygenase activity in dogs. Am J Vet Res 2000; 61: 802-10.
414. Kellgren JH, Lawrence JS. Radiological assessment of osteo-arthrosis. Ann Rheum Dis 1957; 16: 494-501.
415. Khalkhali-Ellis Z, Seftor E, Nieva D, Seftor R, Samaha H, Bultman L, et al. Induction of invasive and degradative phenotype in normal synovial fibroblasts exposed to synovial fluid from patients with juvenile rheumatoid arthritis: Role of mononuclear cell population. J Rheumatol 1997; 24: 2451-60.

256.
416. Kikuchi H, Tanaka S, Matsuo O. Plasminogen activator in synovial fluid from patients with rheumatoid arthritis. J Rheumatol 1987; 14: 439-45.
417. Kim Y-J, Sah RLY, Doong J-YH, Grodzinsky AJ. Fluorometric assay of DNA in cartilage explants using Hoechst 33258. Anal Biochem 1988; 174:(1)168-176.
418. Kimble R, Bain S, Pacifici R. The functional block of TNF but not of IL-6 prevents bone loss in ovariectomized mice. J Bone Miner Res 1997; 12: 935-41.
419. King WJ, Greene GL. Monoclonal antibodies localize oestrogen receptor in the nuclei of target cells. Nature 1984; 307: 745-7.
420. Kinne R, Boehm S, Iftner T, Aigner T, Vornehm S, Weseloh G, et al. Synovial fibroblast-like cells strongly express jun-B and C-fos proto-oncogenes in rheumatoid arthritis- and osteoarthritis. Scand J Rheumatol 1995; 101 (Suppl):121-5.
421. Király K, Hyttinen MM, Lapveteläinen T, Elo M, Kiviranta I, Dobai J, et al. Specimen preparation and quantification of collagen birefringence in unstained sections of articular cartilage using image-analysis and polarizing light microscopy. Histochem J 1997; 29: 317-27.
422. Kirkwood T. What is the relationship between osteoarthritis and aging? Bail Clin Res 1997; 11: 683-94.
423. Klareskog L, Forsum U, Kabelitz DPL, Sundstrom C, Nilsson K, Wigren A, et al. Immune functions of human synovial cells Phenotypic and T cell regulatory properties of macrophage like cells that express HLA-DR. Arthritis Rheum 1982; 25: 488-501.
424. Kleiner DE, Stetler-Stevenson WG. Quantitative zymography: detection of picogram quantities of gelatinases. Anal Biochem 1994; 218: 325-9.
425. Knight C, Willenbrock F, Murphy G. A novel coumarin-labeled peptide for sensitive continuous assays of the matrix metalloproteinases. FEBS Letters 1992; 296: 263-6.
426. Kobayashi K, Mishima H, Hashimoto S, Goomer RS, Harwood FL, Lotz M, et al. Chondrocyte apoptosis and regional differential expression on nitric oxide in the medial meniscus following partial meniscectomy. J Orthop Res 2001; 19: 802-8.
427. Kobayashi T, Okamoto K, Kobata T, Hasunuma T, Sumida T, Nishioka K. Tumor necrosis factor regulation of the Fas-mediated apoptosis-signalling pathway in synovial cells . Arthritis Rheum
1999; 42: 519-26.
428. Kobayashi Y, Ichijo Y, Nagatake H, Watanabe N, Nishizawa K, Goto M. Processing of interleukin-1b in synovial cells freshly isolated from patients with rheumatoid arthritis. Biochem Biophys Res Commun 1994; 199: 1035-40.
429. Koka S, Petro T, Reinhardt R. Estrogen inhibits interleukin-1-beta-induced interleukin-6 production by human osteoblast-like cells. J Interferon Cytokine Res 1998; 18: 479-83.
430. Konttinen Y, Michelsson J-E, Tolvanen E, Bergroth V. Primary inflammatory reaction in synovial-fluid and tissue in rabbit immobilization osteoarthritis. Clin Orthop 1990; 260: 280-6.
431. Konttinen Y, Saari H, Santavirta S, Antti-Poika I, Sorsa T, Nykanen P, et al. Synovial fibroblasts. Scand J Rheumatol 1988; 76 (Suppl.): 95-103.
432. Koolwijk P, Miltenburg AMM, van Erck MGM, Oudshoorn M, Niedbala MJ, Breedveld FC, et al. Activated gelatinase-B (MMP-9) and urokinase-type plasminogen activator in synovial fluids of patients with arthritis. Correlation with clinical and experimental variables of inflammation. J Rheumatol 1995; 22: 385-93.

257.
433. Kousteni S, Bellido T, Plotkin LI, O'Brien CA, Bodenner DL, Han L, et al. Nongenotropic, sex-nonspecific signalling through the estrogen or androgen receptors: Dissociation from transcriptional activity. Cell 2001; 104: 719-30.
434. Kraan M, Versendaal H, Jonker M, Bresnihan B, Post W, Thart B, et al. Asymptomatic synovitis precedes clinically manifest arthritis. Arthritis Rheum 1998; 41: 1481-8.
435. Krane S, Amento E. Cellular interactions and control of collagenase secretion in the syvovium. J Rheumatol 1983; 11 (Suppl): 7-12.
436. Krco CJ, Pawelski J, Harders J, McCormick D, Griffiths M, Luthra HS, et al. Characterization of the antigenic structure of human type II collagen. J Immunol 1996; 156: 2761-8.
437. Kreigsmann J, Keyszer G, Geiler T, Brauer R, Gay R, Gay S. Expression of vascular cell adhesion molecule-1 mRNA and protein in rheumatoid synovium demonstrated by in situ hybridization and immunohistochemistry. Lab Invest 1995; 72: 209
438. Krey P, Scheinberg M, Cohen A. Fine structural analysis of rabbit synovial cells. II. Fine structure and rosette-forming cells of explant and monolayer cultures. Arthritis Rheum 1976; 19: 581-92.
439. Kuiper GGJM, Lemmen JG, Carlsson B, Corton JC, Safe SH, Van der Saag PT, et al. Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor ß. Endocrinol 1998; 139: 4252-63.
440. Kuiper S, Joosten L, Bendele A, Edwards C, Arntz O, Helsen M, et al. Different roles of tumour necrosis factor and interleukin 1 in murine streptococcal cell wall arthritis. Cytokine 1998; 10: 690-702.
441. Kumkumian GK, Lafyatis R, Remmers EF, Case JP, Kim S-J, Wilder RL. Platelet-derived growth factor and IL-1 interactions in rheumatoid arthritis . J Immunol 1989; 143: 833-7.
442. Labarca C, Paigen K. A simple, rapid, and sensitive DNA assay procedure. Anal Biochem 1980; 102: 344-52.
443. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970; 227: 680-5.
444. Lafyatis R, Kim S, Angel P, Roberts A, Sporn M, Karin M, et al. Interleukin-1 stimulates and all-trans-retinoic acid inhibits collagenase gene expression through its 5' activator protein-1-binding site. Molecular Endocrinol 1990; 4:973-80.
445. Lafyatis R, Remmers E, Roberts A, Yocum D, Sporn M, Wilder R. Anchorage-independent growth of synoviocytes from arthritic and normal joints. Stimulation by exogenous platelet-derived growth factor and inhibition by transforming growth factor-beta and retinoids. J Clin Invest 1989; 83: 1267-76.
446. Lafyatis R, Thompson N, Remmers E, Flanders K, Roche N, Kim S-J, et al. Transforming growth factor-ß production by synovial tissues from rheumatoid patients and streptococcal cell wall arthritic rats. J Immunol 1989; 143: 1142-8.
447. Lambert CA, Lapiere CM, Nusgens BW. An interleukin-1 loop is induced in human skin fibroblasts upon stress relaxation in a three-dimensional collagen gel but is not involved in the up-regulation of matrix metalloproteinase 1. J Biol Chem 1998; 273: 23143-9.
448. Landino L, Crews B, Timmons M, Morrow J, Marnett L. Peroxynitrite, the coupling product of nitric oxide and superoxide, activates prostaglandin biosynthesis. Proc Natl Acad Sci USA 1996; 93: 15069-74.

258.
449. Lane N, Gore R, Cummings S, Hochberg M, Scott J, Williams E, et al. Serum vitamin D levels and incident changes of radiographic hip osteoarthritis. Arthritis Rheum 1999; 42 :854-60.
450. Lane N, Nevitt M, Williams E, Cummings S, Hochberg M. The association of nitrate use and incident radiographic hip OA in older white women. Proc Ann Mtg Am Coll Rheumatol 1999; Abstract 568 (Abstract).
451. Lapcik L, Lapcik L, De Smedt S, Demeester J, Chabrecek P. Hyaluronan: Preparation, structure, properties, and applications. Chem Rev 1998; 98: 2663-84.
452. Larbre J, Dasilva J, Moore A, James I, Scott D, Willoughby D. Cartilage contribution to gender differences in joint disease progression: A study with rat articular cartilage. Clin Exp Rheum 1994; 12: 401-8.
453. Lavietes B, Carsons S, Diamond H, Laskin R. Synthesis, secretion and deposition of fibronectin in cultured human synovium. Arthritis Rheum 1985; 28: 1016-26.
454. Lavnikova N, Laskin D. Unique patterns of regulation of nitric oxide production in fibroblasts. J Leukoc Biol 1995; 58: 451-8.
455. Le Roux MA, Arokoski J, Vail TP, Guilak F, Hyttinen MM, Kiviranta I, et al. Simultaneous changes in the mechanical properties, quantitative collagen organization, and proteoglycan concentration of articular cartilage following meniscectomy. J Orthop Res 2000; 18: 383-92.
456. Lederer JA, Czuprynski CJ. Species preference of bovine thymocytes and fibroblasts for bovine interleukin 1. Vet Immunol Immunopathol 1989; 23: 213-22.
457. LeGrand A, Fermor B, Fink C, Pisetsky DS, Weinberg JB, Vail TP, et al. Interleukin-1, tumor necrosis factor , and interleukin-17 synergistically up-regulate nitric oxide and prostaglandin E2 production in explants of human osteoarthritic knee menisci. Arthritis Rheum 2001; 44: 2078-83.
458. Leifke E, Gorenoi V, Wichers C, von zur Muhlen A, von Buren E, Brabant G. Age-related changes of serum sex hormones, insulin-like growth factor-1 and sex-hormone binding globulin levels in men:cross-sectional data from a healthy male cohort. Clin Endocrinol 2000; 53: 689-95.
459. Leizer T, Clarris BJ, Ash PE, van Damme J, Saklatvala J, Hamilton JA. Interleukin-1-beta and interleukin-1-alpha stimulate the plasminogen activator activity and prostaglandin E2 levels of human synovial cells. Arthritis Rheum 1987; 30: 562-6.
460. Leizer T, Hamilton JA. Plasminogen-activator and prostaglandin-E2 levels in human synovial fibroblasts - differential stimulation by synovial activator and other cytokines. J Immunol 1989; 143: 971-8.
461. Lembach K. Enhanced synthesis and extracellular accumulation of hyaluronic acid during stimulation of quiescent human fibroblasts by mouse epidermal growth factor. J Cell Physiol 1976; 89: 277-88.
462. Leprince P, Rogister B, Moonen G. A colorimetric assay for the simultaneous measurement of plasminogen activators and plasminogen activator inhibitors in serum-free conditioned media from cultured cells. Anal Biochem 1989; 177: 341-6.
463. Lesquesne M, Cadet C. Do chondroprotective agents exist in osteoarthritis treatment? Required proof. Therapie 1998; 53: 7-16.
464. Lesquesne M, Maheu E, Cadet C, Caspard H, Dreiser RL. Effect of avocado/soya unsaponifiables (ASU) on joint space loss in hip osteoarthritis (HOA) over 2 years. A placebo controlled trial. Arthritis Rheum 1996; 39 (Suppl 9): S227 (Abstract).

259.
465. Lettesjö H, Nordström E, Ström H, Nilsson B, Glinghammar B, Dahlstedt L, et al. Synovial fluid cytokines in patients with rheumatoid arthritis or other arthritic lesions. Scand J Immunol 1998; 48: 286-92.
466. Liberman U, Weiss S, Bröll J, Minne H, Quan H, Bell N, et al. Effect of oral alendronate on bone mineral density and the incidence of fractures in postmenopausal osteoporosis. N Engl J Med 1995; 333: 1437-43.
467. Lin C, Robbins P, Georgescu H, Evans C. Effects of immortalization upon the induction of matrix metalloproteinases in rabbit synovial fibroblasts. Exp Cell Res 1996; 223:117-26.
468. Lindblad S. Recent progress in the study of synovitis by macroscopic and microscopic examination - a review. Scand J Rheumatology 1988; 76 (Suppl.):27-32.
469. Lindblad S, Hedfors E. Intraarticular variation in synovitis. Local macroscopic and microscopic signs of inflammatory activity are significantly correlated . Arthritis Rheum 1985; 28:977-9
470. Lindblad S, Hedfors E. Arthroscopic and immunohistologic characterization of knee-joint synovitis in osteoarthritis. Arthritis Rheum 1987; 30: 1081-8.
471. Lindblad S, Hedfors E. The synovial membrane of healthy individuals - immunohistochemical overlap with synovitis. Clip Exp Immunol 1987; 69: 41-7.
472. Lindblad S, Klareskog L, Hedfors E, Forsum U, Sundstrom C. Phenotypic characterisation of synovial tissue cells in situ in different types of synovitis. Arthritis Rheum 1983; 26: 1321-32.
473. Lipowitz A, Wong P, Stevens J. Synovial membrane changes after experimental transection of the cranial cruciate ligament in dogs . Am J Vet Res 1985; 46: 1166-70.
474. Little C, Smith S, Ghosh P, Bellenger C. Histomorphological and immunohistochemical evaluation of joint changes in a model of osteoarthritis induced by lateral meniscectomy in sheep. J Rheumatol 1997; 24: 2199-209.
475. Little CB, Ghosh P, Bellenger CR. Topographic variation in biglycan and decorin synthesis by articular cartilage in the early stages of osteoarthritis: An experimental study in sheep. J Orthop Res 1996; 14: 433-44.
476. Lloyd ME, Hart DJ, Nandra D, McAlindon TE, Wheeler M, Doyle DV, et al. Relation between insulin-like growth factor-I concentrations, osteoarthritis, bone density, and fractures in the general population: the Chingford study. Ann Rheum Dis 1996; 55: 870-4.
477. Lohmander LS, Hoerrner LA, Lark MW. Metalloproteinases, tissue inhibitor, and proteoglycan fragments in knee synovial fluid in human osteoarthritis. Arthritis Rheum 1993; 36: 181-9.
478. Lohmander L, Neame P, Sandy J. The structure of aggrecan fragments in human synovial fluid: Evidence that aggrecanase mediates cartilage degradation in inflammatory joint disease, joint injury, and osteoarthritis. Arthritis Rheum 1993; 36: 1214-22.
479. Lohmander L, Roos H, Dahlberg L, Hoerrner L, Lark M. Temporal patterns of stromelysin-1, tissue inhibitor, and proteoglycan fragments in human knee joint fluid after injury to the cruciate ligament or meniscus. J Orthop Res 1994; 12: 21-8.
480. Lotz M, Guerne PA. Interleukin-6 induces the synthesis of tissue inhibitor of metalloproteinases-1/erythroid potentiating activity (TIMP-1/EPA). J Biol Chem 1991; 266: 2017-20.
481. Lotz M, Hashimoto S, Kühn K. Mechanisms of chondrocyte apoptosis. Osteoarthritis Cartilage 1999; 7: 389-91.

260.
482. Lotz M, Moats T, Villiger PM. Leukemia inhibitory factor is expressed in cartilage and synovium and can contribute to the pathogenesis of arthritis. J Clin Invest 1992; 90: 888-96.
483. Lowik CWGM, Nibbering PH, van de Ruit M, Papapoulos SE. Inducible production of nitric oxide in osteoblast-like cells and in fetal mouse bone explants is associated with suppression of osteoclastic bone resorption. J Clin Invest 1994; 93: 1465-72.
484. Lukoschek M, Schaffler M, Burr D, Boyd R, Radin E. Synovial membrane and cartilage changes in experimental osteoarthritis. J Orthop Res 1988; 6: 475-92.
485. Mabuchi K, Ujihira M, Sasada T. Relationship between the conformity and the lubrication ability of synovial joints. Clin Biomech 1998; 13: 250-5.
486. MacIntyre I, Zaidi M, Towhidul Alam ASM, Datta HK, Moonga BS, Lidbury PS, et al. Osteoclastic inhibition: An action of nitric oxide not mediated by cyclic GMP. Proc Natl Acad Sci USA 1991; 88: 2936-40.
487. Mackintosh D, Mason RM. Pharmacological actions of 17ß-oestradiol on articular cartilage chondrocytes and chondrosarcoma chondrocytes in the absence of oestrogen receptors. Biochim Biophys Acta 1988; 964: 295-302.
488. MacNaul K, Chartrain N, Lark M, Tocci M, Hutchinson N. Discoordinate expression of stromelysin, collagenase, and tissue inhibitor of metalloproteinases-1 in rheumatoid human synovial fibroblasts - synergistic effects of interleukin-1 and tumor necrosis factor-alpha on stromelysin expression. J Biol Chem 1990; 265: 17238-45.
489. MacPhail CM, Lappin MR, Meyer DJ, Smith SG, Webster CRL, Armstrong PJ. Hepatocellular toxicosis associated with administration of carprofen in 21 dogs. J Am Vet Med Assoc 1998; 212: 1895-901.
490. Maheu E, Dreiser R-L, Guillou GB, Dewailly J. Hand osteoarthritis patients characteristics according to the existence of a hormone replacement therapy. Osteoarthritis Cartilage 2000; 8 (Suppl A): S33-7.
491. Maheu E, Maziéres B, Valat J-P, Loyau G, Le Loët X, Bourgeois P, et al. Symptomatic efficacy of avocado/soybean unsaponifiables in the treatment of osteoarthritis of the knee and hip. A prospective, randomized, double-blind, placebo-controlled, multicenter clinical trial with a six-month treatment period and a two-month followup demonstrating a persistent effect. Arthritis Rheum 1998; 41: 81-91.
492. Makarov S, Olsen J, Johnston W, Anderle S, Brown R, Baldwin A, et al. Suppression of experimental arthritis by gene transfer of interleukin 1 receptor antagonist cDNA. Proc Natl Acad Sci USA 1996; 93: 402-6.
493. Malyak M, Guthridge J, Hance K, Dower S, Freed J, Arend W. Characterisation of a low molecular weight isoform of IL-1 receptor agonist. J Immunol 1998; 161:1997-2003.
494. Manfield L, Jang D, Murrell GAC. Nitric oxide enhances cyclooxygenase activity in articular cartilage. Inflamm Res 1996; 45: 254-8.
495. Manicourt D-H, Cornu O, Lenz ME, Druetz-van Egeren A, Thonar EJ-MA. Rapid and sustained rise in the serum level of hyaluronan after anterior cruciate ligament transection in the dog knee joint. J Rheumatol 1995; 22: 262-9.
496. Manicourt D, Altman R, Williams J, Devogelaer J, Dreutz-van Egeren A, Lenz M, et al. Treatment with calcitonin suppresses the responses of bone, cartilage, and synovium in the early stages of canine experimental osteoarthritis and significantly reduces the severity of the cartilage lesions. Arthritis Rheum 1999; 42: 1159-67.

261.
497. Mankin HJ, Conger KA. The acute effects of intra-articular hydrocortisone on articular cartilage in rabbits. J Bone Joint Surg 1966; 48A:,1383-8.
498. Mankin H, Dorfman H, Lippiello L, Zarins A. Biochemical and metabolic abnormalities in articular cartilage from osteoarthritic human hips. II Correlation of morphology with biochemical and metabolic data. J Bone Joint Surg 1971; 53A: 523-37.
499. Mankin HJ, Lippiello L. The glycosaminoglycans of normal and arthritic cartilage. J Clin Invest 1971; 50: 1712-9.
500. Mansell JP, Bailey AJ. Abnormal cancellous bone collagen metabolism in osteoarthritis. J Clin Invest 1998; 101: 1596-603.
501. Mapp PI. Innervation of the synovium. Ann Rheum Dis 1995; 54: 398-403.
502. Maroudas A, Bayliss M, Venn M. Further studies on the composition of human femoral head cartilage. Ann Rheum Dis 1980; 39:514-23.
503. Marsh J, Roback D, Ross G. In vitro conditions affecting the synthesis of sulphated proteoglycans by rheumatoid and normal synovial tissues in culture. Arthritis Rheum 1982; 25: 196-203.
504. Marsh J, Wiebkin O, Gale S, Muir H, Maini R. Synthesis of sulphated proteoglycans by rheumatoid and normal synovial tissues in culture. Ann Rheum Dis 1979; 38: 166-70.
505. Martel-Pelletier J, Di Battista J, Lajeunesse L. Biochemical factors in joint articular tissue degradation in osteoarthritis. In: Reginster J-Y, Pelletier J-P, Martel-Pelletier J, Henrotin Y, (Eds.) Osteoarthritis: Clinical and experimental aspects. Berlin : Springer-Verlag, 1999:156-187.
506. Martel-Pelletier J, Di Battista JA, Lajeunesse D, Pelletier J-P. IGF/IGFBP axis in cartilage and bone in osteoarthritis pathogenesis. Inflamm Res 1998; 47: 90-100.
507. Martel-Pelletier J, Faure M-P, McCollum R, Mineau F, Cloutier J-M, Pelletier J-P. Plasmin, plasminogen activators and inhibitor in human osteoarthritic cartilage. J Rheumatol 1991; 18: 1863-71.
508. Martel-Pelletier J, McCollum R, DiBattista J, Faure M-P, Chin JA, Fournier S, et al. The interleukin-1 receptor in normal and osteoarthritic human articular chondrocytes. Identification as the type I receptor and analysis of binding kinetics and biologic function. Arthritis Rheum 1992; 35: 530-40.
509. Martel-Pelletier J, McCollum R, Fujimoto N, Obata K, Cloutier J-M, Pelletier J-P. Excess of metalloproteases over tissue inhibitor of metalloprotease may contribute to cartilage degradation in osteoarthritis and rheumatoid arthritis. Lab Invest 1994; 70: 807-15.
510. Martel-Pelletier J, McCollum R, Pelletier J-P. The synthesis of IL-1 receptor antagonist (IL-1RA) by synovial fibroblasts is markedly increased by the cytokines TNF-alpha and IL-1 . Biochim Biophys Acta 1993; 1175: 302-5.
511. Martel-Pelletier J, Mineau F, Jolicoeur F-C, Cloutier J-M, Pelletier J-P. In vitro effects of diacerhein and rhein on interleukin 1 and tumor necrosis factor-a-systems in human osteoarthritic synovium and chondrocytes. J Rheumatol 1998; 25: 753-62.
512. Martel-Pelletier J, Mineau F, Jolicoeur FC, Pelletier J-P. Modulation of TNFsR55 and TNFsR75 by cytokines and growth factors in human synovial fibroblasts. J Rheumatol 1995; 22 (Suppl 43): 115-9.
513. Martel-Pelletier J, Mineau F, Tardif G, Fernandes J, Ranger P, Loose L, et al. Tenidap reduces the level of interleukin 1 receptors and collagenase expression in human arthritic synovial

262.
fibroblasts. J Rheumatol 1996; 23: 24-31.
514. Martel RR, Klicius J, Metcalf G. Effect of etodolac on articular and bone pathology associated with adjuvant arthritis in rats: a comparison with aspirin and naproxen. Agents Actions 1984; 14: 257-64.
515. Massicote F, Lajeunesse D, Benderbour M, Fernandes JC, Pelletier J-P, Martel-Pelletier J. Prostaglandin E2 and interleukin-6 production by human subchondral osteoblasts can discriminate two subgroups of osteoarthritic patients. Osteoarthritis Cartilage 2000; 8 (Suppl. A):S8 (Abstract).
516. Mauviel A, Chung K-Y, Agarwal A, Tamai K, Uitto J. Cell-specific induction of distinct oncogenes of the Jun family is responsible for differential regulation of collagenase gene expression by transforming growth factor-beta in fibroblasts and keratinocytes. J Biol Chem 1996; 271: 10917-23.
517. Mauviel A, Daireaux, Hartmann DJ, Galera P, Loyau G. Effets des insaponifiables d'avocat/soja (PIAS) sur la production de collagène par des cultures de synoviocytes, chondrocytes artiulaires et fibroblasts dermiques. Rev Rhum 1989; 56: 207-11.
518. Mauviel A, Loyau G, Pujol JP. Effet des insaponifiables d'avocat/soja (Piasclédine) sur l'activité collagènolytique de cultures de synoviocytes rhumatoïdes humains et de chondrocytes articulaires de lapin traités par l'interleukine-1. Rev Rhum 1991; 58: 241-5.
519. Mauviel A, Redini F, Loyau G, Pujol JP. Modulation of extracellular matrix metabolism in rabbit articular chondrocytes and human rheumatoid synovial cells by the non-steroidal anti-inflammatory drug etodolac. I: Collagen synthesis. Agents Actions 1990; 31: 345-52.
520. May NDS. The Anatomy of the Sheep: a dissection manual (3rd Ed.). Brisbane: University of Queensland Press, 1970
521. May SA, Hooke RE, Lees P. Species restrictions demonstrated by the stimulation of equine cells with recombinant human interleukin-1. Vet Immunol Immunopathol 1992; 30: 373-84.
522. Mazieres B, Tempesta C, Tiechard M, Vaguier G. Pathologic and biochemical effects of a lipidic avocado and soy extract (LASE) on an experimental post-contusive model of OA. Osteoarthritis Cartilage 1993; 1: 46-7 (Abstract).
523. McAlindon TE, Felson DT, Zhang YQ, Hannan MT, Aliabadi P, Weissman B, et al. Relation of dietary intake and serum levels of vitamin-D to progression of osteoarthritis of the knee among participants in the Framingham study. Ann Intern Med 1996; 125: 353-9.
524. McCachren SS. Expression of metalloproteinases and metalloproteinase inhibitor in human arthritic synovium. Arthritis Rheum 1991; 34: 1085-93.
525. McCartney-Francis N, Allen JB, Mizel DE, Albina JE, Xie Q-W, Nathan CF, et al. Suppression of arthritis by an inhibitor of nitric oxide synthase. J Exp Med 1993; 178: 749-54.
526. McCarty D, Cheung H. Prostaglandin (PG) E2 generation by cultured canine synovial fibroblasts exposed to microcrystals containing calcium. Ann Rheum Dis 1985; 44: 316-20.
527. McCarty M. Enhanced synovial production of hyaluronic acid may explain rapid clinical-response to high-dose glucosamine in osteoarthritis. Med Hypoth 1998; 50: 507-10.
528. McDonnell J, Hoerrner L, Lark M, Harper C, Dey T, Lobner J, et al. Recombinant human interleukin-1-beta-induced increase in levels of proteoglycans, stromelysin, and leukocytes in rabbit synovial fluid. Arthritis Rheum 1992; 35: 799-805.
529. McGuire-Goldring M, Meats J, Wood D, Ihrie E, Ebsworth N, Russell R. In vitro activation of

263.
human chondrocytes and synoviocytes by a human interleukin-1-like factor. Arthritis Rheum 1984; 27: 654-62.
530. McGuire MK, Meats JE, Ebsworth NM, Harvey L, Murphy G, Russell RG, et al. Properties of rheumatoid and normal synovial tissue in vitro and cells derived from them. Production of prostaglandins and collagenase in response to factors derived from cultured blood mononuclear cells and from synovium. Rheumatol Int 1982; 2: 113-20.
531. McGuire M, Meats J, Ebsworth N, Murphy G, Reynolds J, Russell R. Messenger functions of prostaglandins in cell to cell interactions and control of proteinase activity in the rheumatoid joint. Int J Immunopharm 1982; 4: 91-102.
532. McInnes LB, Leung BP, Field M, Wei XQ, Huang FP, Sturrock RD, et al. Production of nitric oxide in the synovial membrane of rheumatoid and osteoarthritis patients. J Exp Med 1996; 184: 1519-24.
533. Meats J, Elford P, Burning R, Russell R. Retinoids and synovial factor(s) stimulate the production of plasminogen activator by cultured human chondrocytes A possible role for plasminogen activator in the resorption of cartilage in vivo. Biochim Biophys Acta 1985; 838: 161-9.
534. Meats J, McGuire M, Russell R. Human synovium releases a factor which stimulates chondrocyte production of PGE and plasminogen activator. Nature 1980; 286: 891-2.
535. Medcalf R, Hamilton JA. Human synovial fibroblasts produce urokinase-type plasminogen activator. Arthritis Rheum 1986; 29:1397-401.
536. Mehraban F, Lark MW, Ahmed FN, Xu F, Moskowitz RW. Increased secretion and activity of matrix metalloproteinase-3 in synovial tissues and chondrocytes from experimental osteoarthritis. Osteoarthritis Cartilage 1998; 6: 286-94.
537. Mehraban F, Tindal M, Proffitt M, Moskowitz R. Temporal pattern of cysteine endopeptidase (cathepsin B) expression in cartilage and synovium from rabbit knees with experimental osteoarthritis: Gene expression in chondrocytes in response to interleukin-1 and matrix depletion. Ann Rheum Dis 1997; 56:108-15.
538. Melchiorri C, Meliconi R, Frizziero L, Silvestri T, Pulsatelli L, Mazzetti I, et al. Enhanced and coordinated in vivo expression of inflammatory cytokines and nitric oxide synthase by chondrocytes from patients with osteoarthritis. Arthritis Rheum 1998; 41:2165-74.
539. Mentzel K, Bräuer R. Matrix metalloproteinases, IL-6, and nitric oxide in rat antigen-induced arthritis. Clin Exp Rheumatol 1998; 16: 269-76.
540. Messner K, Fahlgren A, Ross I, Andersson B. Simultaneous changes in bone mineral density and articular cartilage in a rabbit meniscectomy model of knee osteoarthritis. Osteoarthritis Cartilage 2000; 8:197-206.
541. Messner K, Gao JZ. The menisci of the knee joint. Anatomical and functional characteristics, and a rationale for clinical treatment. J Anat 1998; 193: 161-78.
542. Meyer F, Yaron I, Yaron M. Synergistic, additive, and antagonistic effects of interleukin-1-beta, tumor necrosis-factor-alpha, and gamma-interferon on prostaglandin E, hyaluronic acid, and collagenase production by cultured synovial fibroblasts. Arthritis Rheum 1990; 33:1518-25.
543. Michel JB, Querterm T. Modulation of messenger-RNA levels urinary type and tissue-type plasminogen-activator and plasminogen-activator inhibitor-1 and inhibitor-2 in human-fibroblasts by interleukin 1. J Immunol 1989; 143:,890-5.
544. Miller WH, Scott DW, Wellington JR. Treatment of dogs with hip arthritis with a fatty acid

264.
supplement. Canin Pract 1992; 17: 6-8.
545. Milz S, Eckstein F, Putz R. The thickness of the subchondral plate and its correlation with the thickness of the uncalcified articular cartilage in human patella. Anat Embryol 1995; 192: 437-44.
546. Miwa M, Saura R, Hirata S, Hayashi Y, Mizuno K, Itoh H. Induction of apoptosis in bovine articular chondrocyte by prostaglandin E2 through cAMP-dependant pathway. Osteoarthritis Cartilage 2000; 8: 17-24.
547. Miyasaka M, Sato K, Hashimoto J, Kohsaka H, Yamamoto K, Goto M, et al. Constitutive production of interleukin-6/B cell stimulatory factor-2 from inflammatory synovium. Clin Immunol Immunopathol 1989; 52: 238-47.
548. Mochan E, Keler T. Plasmin degradation of cartilage proteoglycan. Biochim Biophys Acta 1984; 800: 312-5.
549. Mochan E, Uhl J, Newton R. Evidence that interleukin-1 induction of synovial cell plasminogen activator is mediated via prostaglandin-E2 and cAMP. Arthritis Rheum 1986; 29:1078-84.
550. Mohamed-Ali H. Influence of interleukin-1ß, tumour necrosis factor alpha and prostaglandin E2 on chondrogenesis and cartilage matrix breakdown in vitro. Rheumatol Int 1995; 14: 191-9.
551. Mohtai M, Smith RP, Schurman DJ, Tsuji Y, Torti FM, Hutchinson NI, et al. Expression of 92-KD type-IV collagenase gelatinase (gelatinase-B) in osteoarthritic cartilage and its induction in normal human articular cartilage by interleukin-1. J Clin Invest 1993; 92: 179-85.
552. Moncada S, Palmer RMJ, Higgs EA. Nitric oxide: Physiology, pathophysiology, and pharmacology. Pharmacol Rev 1991; 43: 109-42.
553. Morales-Ducret J, Wayner E, Elices MJ, Alvaro-Gracia JM, Zvaifler NJ, Firestein GS. 4/ß1 integrin (VLA-4) ligands in arthritis. J Immunol 1992; 149: 1424-31.
554. Morales TI, Hascall VC. Factors involved in the regulation of proteoglycan metabolism in articular-cartilage. Arthritis Rheum 1989; 32: 1197-201.
555. Moskowitz RW, Davis W, Sammarco J, Mast W, Chase SW. Experimentally induced corticosteroid arthropathy. Arthritis Rheum 1970; 13: 236-43.
556. Moskowitz R, Howell D, Goldberg V, Mankin H. Osteoarthritis-diagnosis and medical/surgical management. 2nd edition. WB Saunders Company, Philadelphia 1992
557. Müller-Ladner U, Evans C, Franklin B, Roberts C, Gay R, Robbins P, et al. Gene transfer of cytokine inhibitors into human synovial fibroblasts in the SCID mouse model. Arthritis Rheum 1999; 42: 490-7.
558. Müller-Ladner U, Gay R, Gay S. Structure and function of synoviocytes. In: Koopman W (Ed.). Arthritis and allied conditions. 13th edition.Baltimore: Williams and Wilkins 1997; 243-54
559. Muniyappa R, Walsh MF, Rangi JS, Zayas RM, Standley PR, Ram JL, et al. Insulin-like growth factor 1 increases vascular smooth muscle nitric oxide production. Life Sci 1997; 61: 925-31.
560. Murphy G, Hembry R. Proteinases in rheumatoid arthritis. J Rheumatol 1992; 19 (Suppl 32): 61-4.
561. Murrell GAC, Jang D, Williams RJ. Nitric oxide activates metalloprotease enzymes in articular cartilage. Biochem Biophys Res Commun 1995; 206: 15-21.
562. Muscara MN, McKnight W, Asfaha S, Wallace JL. Wound collagen deposition in rats: effects of an NO-NSAID and a selective COX-2 inhibitor. Br J Pharmacol 2000; 129: 681-6.

265.
563. Müssener Å, Funa K, Kleinau S, Klareskog L. Dynamic expression of transforming growth factor betas (TGF-ß) and their type I and type II receptors in the synovial tissue of arthritic rats. Clin Exp Immunol 1997; 107: 112-9.
564. Mwanza AM, Madej A, Kindahl H, Lundeheim N, Einarsson S. Plasma levels of cortisol, progesterone, oestradiol-17ß and prostaglandin F2 metabolite after ACTH (Synacthen Depot) administration in ovariectomised gilts. J Vet Med A Physiol Pathol Clin Med 2000; 47: 193-200.
565. Myers S. Canine platelet factors stimulate glycosaminoglycan synthesis by synovial cell and organ cultures. Connect Tissue Res 1985; 14: 41-9.
566. Myers S. Suppression of hyaluronic acid synthesis in synovial organ cultures by corticosteroid suspensions. Arthritis Rheum 1985; 28:(12)1275-1282.
567. Myers S, Brandt K. Studies of synovial hyaluronic acid synthesis in canine osteoarthritis. J Rheumatol 1987; 14: 1150-5.
568. Myers S, Brandt K, Ehlich J, Braunstein E, Shelbourne K, Heck D, et al. Synovial inflammation in patients with early osteoarthritis of the knee. J Rheumatol 1990; 17: 1662-9.
569. Myers S, Brandt K, O'Connor B, Visco D, Albrecht M. Synovitis and osteoarthritic changes in canine articular cartilage after anterior cruciate ligament transection - effect of surgical hemostasis. Arthritis Rheum 1990; 33: 1406-15.
570. Myers S, Christine T. Hyaluronate synthesis by synovial villi in organ culture. Arthritis Rheum 1983; 26: 764-70.
571. Myers S, Flusser D, Brandt K, Heck D. Prevalence of cartilage shards in synovium and their association with synovitis in patients with early and endstage osteoarthritis. J Rheumatol 1992; 19: 1247-51.
572. Nasatzky E, Schwartz Z, Boyan BD, Soskolne WA, Ornoy A. Sex-dependent effects of 17-beta-estradiol on chondrocyte differentiation in culture. J Cell Physiol 1993; 154: 359-67.
573. Nasu M, Sugimoto T, Kaji H, Chihara K. Estrogen modulates osteoblast proloferation and function regulated by parathyroid hormone in osteoblastic SaOS-2 cells: role of insulin-like growth factor (IGF)-I and IGF-binding protein-5. J Endocrinol 2000; 167: 305-13.
574. Nedelec E, Abid A, Cipolletta C, Presle N, Terlain B, Netter P, et al. Stimulation of cyclooxygenase-2-activity by nitric oxide-derived species in rat chondrocyte: lack of contribution to loss of cartilage anabolism. Biochem Pharmacol 2001; 61: 965-78.
575. Needleman P, Manning PT. Interactions between the inducible cyclooxygenase (COX-2) and nitric oxide synthase (iNOS) pathways: implications for therapeutic intervention in osteoarthritis. Osteoarthritis Cart 1999; 7:367-70.
576. Neidel J, Schulze M, Sova L. Insulin-like growth factor I accelerates recovery of articular cartilage proteoglycan synthesis in culture after inhibition by interleukin 1. Arch Orthop Trauma Surg 1994; 114: 43-8.
577. Nevitt M, Cummings S, Lane N, Hochberg M, Scott J, Pressman A, et al. Association of estrogen replacement therapy with the risk of osteoarthritis of the hip in elderly white women. Arch Intern Med 1996; 156: 2073-80.
578. Nevitt M, Felson D. Sex hormones and the risk of osteoarthritis in women: Epidemiologic evidence. Ann Rheum Dis 1996; 55: 673-6.
579. Nevitt MC, Felson DT, Williams EN, Grady D. The effect of estrogen plus progestin on knee

266.
symptoms and related disability in post-menopausal women: The heart and estrogen/progestin replacement study, a randomized, double-blind, placebo-controlled trial. Arthritis Rheum 2001; 44: 811-8.
580. Nevitt MC, Lane NE, Scott JC, Hochberg MC, Pressman AR, Genant HK, et al. Radiographic osteoarthritis of the hip and bone mineral density. Arthritis Rheum 1995; 38: 907-16.
581. Newman E, Turner AS, Wark JD. The potential of sheep for the study of osteopenia: current status and comparison with other animal models. Bone 1995; 16 (Suppl. 4): 277S-284S.
582. Ng KP, Datuin JP, Bern HA. Effects of estrogens in vitro and in vivo on cartilage growth in the Tilapia (Oreochromis mossambicus). Gen Comp Endocrinol 2001; 121: 295-304.
583. Nietfeld JJ, Wilbrink B, Helle M, van Roy JLAM, den Otter W, Swaak AJG, et al. Interleukin-1-induced interleukin-6 is required for the inhibition of proteoglycan synthesis by interleukin-1 in human articular cartilage. Arthritis Rheum 1990; 33: 1695-701.
584. Nishikawa H, Mori I, Umemoto J. Influences of sulphated glycosaminoglycans on biosynthesis of hyaluronic acid in rabbit knee synovium. Arch Biochem Biophys 1985; 240: 146-53.
585. Niu XL, Yang X, Hoshoao K, Tanaka K, Sawamura S, Koga Y, et al. Inducible nitric oxide synthase deficiency does not affect the susceptibility of mice to atherosclerosis but increases collagen content in lesions. Circulation 2001; 103: 1115-20.
586. Nixon AJ, Saxer RA, Brower-Trowland BD. Exogenous insulin-like growth factor-I stimulates an autoinductive IGF-I autocrine/paracrine response in chondrocytes. J Orthop Res 2001; 19: 26-32.
587. Norrdin RW, Jee WSS, High WB. The role of prostaglandins in bone in vivo. Prostagl Leuk Ess Fatty Acids 1990; 41: 139-49.
588. Notoya K, Jovanovic DV, Reboul P, Martel-Pelletier J, Mineau F, Pelletier JP. The induction of cell death in human osteoarthritis chondrocytes by nitric oxide is related to the production of prostaglandin E2 via the induction of cyclooxygenase-2. J Immunol 2000; 165: 3402-10.
589. Noyori K, Jasin H. Inhibition of human fibroblast adhesion by cartilage surface proteoglycans. Arthritis Rheum 1994; 37: 1656-63.
590. O'Connor BL, Visco DM, Heck DA, Myers SL, Brandt KD. Gait alterations in dogs after transection of the anterior cruciate ligament. Arthritis Rheum 1989; 32: 1142-7.
591. Ogawa O, Umegaki H, Sumi D, Hayashi T, Nakamura A, Thakur NK, et al. Inhibition of inducible nitric oxide synthase gene expression by indomethacin or ibuprofen in beta-amyloid protein-stimulated J774 cells . Eur J Pharmacol 2000; 408: 137-41.
592. Ohta H, Ikeda T, Masuzawa T, Makita K, Suda Y, Nozawa S. Differences in axial bone mineral density, serum levels of sex steroids, and bone metabolism between postmenopausal and age-and body size-matched premenopausal subjects. Bone 1993; 14: 111-6.
593. Okada Y, Gonoji Y, Nakanishi I, Nagase H, Hayakawa T. Immunohistochemical demonstration of collagenase and tissue inhibitor of metalloproteinases (TIMP) in synovial lining cells of rheumatoid synivium. Virchows Arch [Cell Pathol] 1990; 59: 305-12.
594. Okada Y, Shinmei M, Tanaka O, Naka K, Kimura A, Nakanishi I, et al. Localization of matrix metalloproteinase 3 (stromelysin) in osteoarthritic cartilage and synovium. Lab Invest 1992; 66: 680-90.
595. Okada Y, Takeuchi N, Tomita K, Nakanish I, Nagase H. Immunolocalization of matrix metalloproteinase-3 (stromelysin) in rheumatoid synovioblasts (B-cells) - correlation with

267.
rheumatoid-arthritis. Ann Rheum Dis 1989; 48: 645-53.
596. Okamoto H, Yamamura M, Morita Y, Harada S, Makino H, Ota Z. The synovial expression and serum levels of interleukin-6, interleukin-11, leukemia inhibitory factor, and oncostatin-M in rheumatoid arthritis. Arthritis Rheum 1997; 40: 1096-105.
597. Okuda T, Yasuoka T, Nakashima M, Oka N. The effect of ovariectomy on the temporomandibular joints of growing rats. J Oral Maxillofac Surg 1996; 54: 1201-10.
598. Oliveria S, Felson D, Klein R, Reed J, Walker A. Estrogen replacement therapy and the development of osteoarthritis. Epidemiol 1996; 7: 415-9.
599. Onga T, Saura R, Mizuno K, Itoh H. Effect of prostaglandin E2 on nitric oxide synthesis in articular chondrocytes. Kobe J Med Sci 2000; 46: 155-69.
600. Oreffo R, Kusec V, Virdi A, Flanagan A, Zambonin-Zallone A, Triffitt J. Estrogen receptor gene expression in osteoclasts - Loss with osteoclast maturation. J Bone Miner Res 1997; 12 (Suppl. 1): S435 (Abstract).
601. Otterness IG, Swindell AC, Zimmerer RO, Poole AR, Ionescu M, Weiner E. An analysis of 14 molecular markers for monitoring osteoarthritis: Segregation of the markers into clusters and distinguishing osteoarthritis at base-line. Osteoarthritis Cartilage 2000; 8: 180-5.
602. Oursler MJ. Estrogen regulation of gene expression in osteoblasts and osteoclasts. Crit Rev Eukaryot Gene Expr 1998; 8: 125-40.
603. Overall CM, Wrana JL, Sodek J. Independent regulation of collagenase, 72-kDa progelatinase, and metalloendoproteinase inhibitor expression in human fibroblasts by transforming growth-beta. J Biol Chem 1989; 264: 1860-9.
604. Paassilta M, Karjalainen A, Kervinen K, Savolainen MJ, Heikkinen J, Backstrom AC, et al. Insulin-like growth factor binding protein-1 (IGFBP-1) and IGF-I during oral and transdermal estrogen replacement therapy: relation to lipoprotein(a) levels. Atherosclerosis 2000; 149: 157-62.
605. Pacifici R, Brown C, Puscheck E, Friedrick E, Slatopolsky E, Maggio D, et al. Effect of surgical menopause and estrogen replacement on cytokine release from human mononuclear cells. Proc Natl Acad Sci USA 1991; 88: 5134-8.
606. Paech K, Webb P, Kuiper G, Nilsson S, Gustafsson J, Kushner P, et al. Differential ligand activation of estrogen receptors ER and ERß at AP1 sites. Science 1997; 277: 1508-10.
607. Page K. Bone and the preparation of bone sections. In: Bancroft J, Stevens A (Eds.) Theory and practice of histological techniques. Edinburgh: Churchill Livingstone, 1982
608. Palacios FA, Novaes GS, Guzzo ML, Laurindo IM, de Mello SB. Interrelationship of the kinin system, nitric oxide and eicosanoids in the antigen-induced arthritis in rabbits . Mediators Inflamm 1999; 8: 245-51.
609. Palmer RMJ, Hickery MS, Charles IG, Moncada S, Bayliss MT. Induction of nitric oxide synthase in human chondrocytes. Biochem Biophys Res Commun 1993; 193: 398-405.
610. Palmoski MJ, Brandt KD. In vivo effect of aspirin on canine osteoarthritic cartilage. Arthritis Rheum 1983; 26: 994-1001.
611. Parker D, Smith SM, Hwa SY, Smith MM, Ghosh P. The effects of ovariectomy and estrogen treatment on cartilage integrity in an ovine model of osteoarthritis. Osteoarthritis Cartilage 2000; 8 (Suppl.): S46 (Abstract).

268.
612. Parsons S, Watson S, Brown P, Collins H, Steele R. Matrix metalloproteinases. Br J Surg 1997; 84: 160-6.
613. Pasquali-Ronchetti I, Frizziero L, Guerra D, Baccarani-Contri M, Focherini M, Georgountzos A, et al. Aging of the human synovium: An in vivo and ex vivo morphological study. Semin Arthritis Rheum 1992; 21: 400-14.
614. Pastoreau PC, Chomel AC, Bonnet J. Evidence of early subchondral bone changes in the meniscectomized guinea pig. A densitometric study using dual-energy X-ray absorptiometry subregional analysis. Osteoarthritis Cartilage 1999; 7: 466-73.
615. Patel S. Current and potential future drug treatments for osteoporosis. Ann Rheum Dis 1996; 55: 700-14.
616. Paterson C. Osteoporosis and the individual patient. Scott Med J 1998; 43: 35-6
617. Peeters-Joris C, Vaes G. Degradation of cartilage proteoglycan and collagen by synovial cells. Stimulation by macrophages under activation by phagocytosis, lymphocyte factors, bacterial products or other inflammatory stimuli. Biochim Biophys Acta 1984; 804: 474-86.
618. Pelletier J-P. Pathophysiology of osteoarthritis . Osteoarthritis Cartilage 1998; 6:374-6.
619. Pelletier J-P, Caron JP, Evans C, Robbins PD, Georgescu HI, Jovanovic D, et al. In vivo suppression of early experimental osteoarthritis by interleukin-1 receptor antagonist using gene therapy. Arthritis Rheum 1997; 40: 1012-9.
620. Pelletier J-P, Faure M-P, DiBattista JA, Wilhelm S, Visco D, Martel-Pelletier J. Coordinate synthesis of stromelysin, interleukin 1, and oncogene proteins in experimental osteoarthritis: An immunohistochemical study. Am J Pathol 1993; 142: 95-105.
621. Pelletier J-P, Haraoui B, Fernandes JC. New and future therapies for osteoarthritis. In: Reginster J-Y, Pelletier J-P, Martel-Pelletier J, Henrotin Y (Eds.) Osteoarthritis: Clinical and experimental aspects. Berlin: Springer-Verlag, 1999: 387-407.
622. Pelletier J-P, Lajeunesse D, Reboul P, Mineau F, Fernandes JC, Sabouret P, et al. Diacerein reduces the excess synthesis of bone remodelling factors by human osteoblast cells from osteoarthritic subchondral bone. J Rheumatol 2001; 28: 814-24.
623. Pelletier J-P, Martel-Pelletier J. Protective effects of corticosteroids on cartilage lesions and osteophyte formation in the Pond-Nuki dog-model of osteo-arthritis. Arthritis Rheum 1989; 32: 181-93.
624. Pelletier J-P, Martel-Pelletier J, Ghandur-Mnaymneh L, Howell DA, Woessner JF. Role of synovial membrane inflammation in cartilage collagen matrix breakdown in the Pond-Nuki dog model of osteoarthritis. Arthritis Rheum 1985; 28: 554-61.
625. Pelletier J-P, Martel-Pelletier J, Howell DS, Ghandur-Mnaymneh L, Enis JE, Woessner JF. Collagenase and collagenolytic activity in human osteoarthritic cartilage. Arthritis Rheum 1983; 26: 63-8.
626. Pelletier J-P, McCollum R, Cloutier J, Martel-Pelletier J. Effects of tiaprofenic acid on plasminogen activators and inhibitors in human OA and RA synovium. Br J Rheumatol 1992; 31 (Suppl. 1): 19-26.
627. Pelletier J-P, McCollum R, Cloutier J, Martel-Pelletier J. Synthesis of metalloproteases and interleukin-6 (IL-6) in human osteoarthritis synovial membrane is an IL-1 mediated process. J Rheumatol 1995; 22 (Suppl. 43): 109-14.

269.
628. Pelletier J-P, Mineau F, Fernandes JC, Duval N, Martel-Pelletier J. Diacerhein and rhein reduce the interleukin-1-beta stimulated inducible nitric oxide synthesis level and activity while stimulating cyclooxygenase-2 synthesis in human osteoarthritic chondrocytes. J Rheumatol 1998; 25: 2417-24.
629. Pelletier J-P, Roughley P, Dibattis J, McCollum R, Martel-Pelletier J. Are cytokines involved in osteoarthritic pathophysiology? Semin Arth Rheum 1991; 20: 12-25.
630. Pelletier JP, Jovanovic D, Fernandes JC, Manning P, Connor JP, Currie MG, et al. Reduced progression of experimental osteoarthritis in vivo by selective inhibition of inducible nitric oxide synthase. Arthritis Rheum 1998; 41: 1275-86.
631. Pelletier JP, Jovanovic DV, Lascau-Coman V, Fernandes JC, Manning PT, Connor JR, et al. Selective inhibition of inducible nitric oxide synthase reduces progression of experimental osteoarthritis in vivo: possible link with the reduction in chondrocyte apoptosis and caspase 3 level. Arthritis Rheum 2000; 43: 1290-9.
632. Pelletier JP, Lajeunesse D, Hilal G, Jovanovic D, Fernandes, Mertel-Pelletier J. Carprofen reduces the structural changes and the abnormal subchondral bone metabolism of experimental osteoarthritis. Osteoarthritis Cartilage 1999; 7: 327-8.
633. Pelletier JP, Lajeunesse D, Jovanovic DV, Lascau-Coman V, Jolicoeur FC, Hilal G, et al. Carprofen simultaneously reduces progression of morphological changes in cartilage and subchondral bone in experimental dog osteoarthritis. J Rheumatol 2000; 27: 2893-902.
634. Pelletier JP, Lascau-Coman V, Jovanovic D, Fernandes JC, Manning P, Manning P, et al. Selective inhibition of inducible nitric oxide synthase in experimental osteoarthritis is associated with reduction in tissue levels of catabolic factors. J Rheumatol 1999; 26: 2002-14.
635. Pelletier JP, Martel-Pelletier J, Howell DS. Etiopathogenesis of osteoarthritis. In: Koopman WJ (Ed.) Arthritis and Allied Conditions. A textbook of Rheumatology. Baltimore: Williams & Wilkens, 1997: 1969-84.
636. Pelletier J, Mineau F, Fernandes J, Kiansa K, Ranger P, Martel-Pelletier J. Two NSAIDs, nimesulide and naproxen, can reduce the synthesis of urokinase and IL-6 while increasing PAI-1, in human OA synovial fibroblasts. Clin Exp Rheumatol 1997; 15: 393-8.
637. Pelletier JP, Mineau F, Ranger P, Tardif G, Martel-Pelletier J. The increased synthesis of inducible nitric oxide inhibits IL-1Ra synthesis by human articular chondrocytes: possible role in osteoarthritic cartilage degradation. Osteoarthritis Cartilage 1996; 4: 77-84.
638. Pelletier JP, Yaron M, Haraoui B, Cohen P, Nahir MA, Choquette D, et al. Efficacy and safety of diacerein in osteoarthritis of the knee. Arthritis Rheum 2000; 43: 2339-48.
639. Peng SX, Vonbargen EC, Bornes DM, Pikul S. Permeability of articular cartilage to matrix metalloprotease inhibitors. Pharm Res 1998; 15: 1414-8.
640. Persson L. On the Synovia in horses A Clinical and Experimental Study. Acta Vet Scand 1971; Suppl 35: 7-77.
641. Pettipher ER, Henderson B, Edwards JCW, Higgs GA. Effects of indomethacin on swelling, lymphocyte influx and cartilage proteoglycan depletion in experimental arthritis. Ann Rheum Dis 1989; 48: 623-7.
642. Pinus H, Ornoy A, Patlas N, Yaffe P, Schwartz Z. Specific ß estradiol binding in cartilage and serum from young mice and rats is age dependent. Connect Tissue Res 1993; 30: 85-98 .

270.
643. Piperno M, Reboul P, Hellio Le Graverand MP, Peschard MJ, Annefeld M, Richard M, et al. Glucosamine sulfate modulates dysregulated activities of human osteoarthritic chondrocytes in
vitro . Osteoarthritis Cartilage 2000; 8:207-12.
644. Pitsillides AA, Rawinson SCF, Suswillo RFL, Bourrin S, Zaman G, Lanyon LE. Mechanical strain-induced NO production by bone cells: A possible role in adaptive bone (re)remodeling? FASEB J 1995; 9: 1614-22.
645. Poli V, Balena R, Fattori E, Markatos A, Yamamoto M, Tanaka H, et al. Interleukin-6 deficient mice are protected from bone loss caused by estrogen depletion. EMBO J 1994; 13: 1189-96.
646. Pond , Nuki G. Experimentally induced osteoarthritis in the dog. Ann Rheum Dis 1973; 32:387
647. Poole CA. Articular cartilage chondrons: form, function and failure. J Anat 1997; 191: 1-13.
648. Poulter LW, Bitensky L, Cashman B, Chayen J. The maintenance of human synovial tissue in vitro. Virchows Archiv B Cell Pathol 1970; 4: 303-9.
649. Presle N, Cipolletta C, Jouzeau J-Y, Abid A, Netter P, Terlain B. Cartilage protection by nitric oxide synthase inhibitors after intraarticular injection of interleukin-1ß in rats. Arthritis Rheum 1999; 42: 2094-102.
650. Prins A, Lipman J, McDevitt C, Sokoloff L. Effect of purified Growth Factors on rabbit articular chondrocytes in monlayer culture. II: Sulfated proteoglycan synthesis. Arthr Rheum 1982; 25: 1228-38.
651. Prins A, Lipman J, Sokloff L. Effect of purified growth factors on rabit articular chondrocytes in monolayer culture. I: DNA synthesis. Arthritis Rheum 1982; 25: 1217-27.
652. Pu J, Nishida K, Inoue H, Asahara H, Ohtsuka A, Murakami T. Mast cells in osteoarthritic and rheumatoid arthritic synovial tissues of the human knee. Acta Med Okayama 1998; 52: 35-9.
653. Pujol JP, Galera P, Pronost S, Boumediene K, Vivien D, Macro M, et al. Transforming growth factor beta (TGF-beta) and articular chondrocytes. Ann Endocrinol (Paris) 1994; 55: 109-20.
654. Pukkli K, Vuorio E, Jalava S. The effect of rheumatoid synovial fluid macrophages on DNA, glycosaminoglycan and collagen synthesis by synovial fibroblasts. Rheumatol Int 1983; 3: 133-8.
655. Punglia RS, Lu W, Hsu J, Kuroki M, Tolentino MJ, Keough K, et al. Regulation of vascular endothelial growth factor expression by insulin-like growth factor I. Diabetes 1997; 46: 1619-26.
656. Radin EL. Subchondral bone changes and cartilage damage. Eq Vet J 1999; 31: 94-5.
657. Radin E, Paul I, Pollock D. Animal joint behaviour under excessive loading. Nature 1970; 226: 554-5.
658. Radin E, Paul I, Rose R. Role of mechanical factors in pathogenesis of primary osteoarthritis. Lancet 1972:519-22.
659. Radin EL, Rose RM. Role of subchondral bone in the initiation and progression of cartilage damage. Clin Orthop 1986; 213: 241-8.
660. Rahn BA, Perren SM. Xylenol orange, a fluorochrome useful in polychrome sequential labeling of calcifying tissues. Stain Technol 1971; 46: 125-9.
661. Rainsford K. Effects of antiinflammatory drugs on the release from porcine synovial tissue in vitro of interleukin 1 like cartilage degrading activity. Agents Actions 1987; 21: 337-40.

271.
662. Rainsford K, Skerry T, Chindemi P, Delaney K. Effects of the NSAIDs meloxicam and indomethacin on cartilage proteoglycan synthesis and joint responses to calcium pyrophosphate crystals in dogs. Vet Res Commun 1999; 23: 101-13.
663. Rainsford KD, Ying C, Smith FC. Effects of meloxicam, compared with other NSAIDs, on cartilage proteoglycan metabolism, synovial prostaglandin E2, and production of interleukin 1, interleukin 6 and interleukin 8, in human and porcine explants in organ culture. J Pharm Pharmacol 1997; 49: 991-8
664. Ralston SH, Ho LP, Helfrich MP, Grabowski PS, Johnston PW, Benjamin N. Nitric oxide: A cytokine-induced regulator of bone resorption. J Bone Miner Res 1995; 10: 1040-9.
665. Ralston SH, Todd D, Helfrich M, Benjamin N, Grabowski PS. Human osteoblast-like cells produce nitric oxide and express inducible nitric oxide synthase. Endocrinol 1994; 135: 330-6.
666. Ramsay B, Johnson M, Leone A, Steer P. The effect of exogenous oestrogen on nitric oxide production in women: a placebo controlled crossover study. Br J Obstet Gynaecol 1995; 102: 417-9.
667. Räsänen T, Mesner K. Articular cartilage compressive stiffness following oophorectomy or treatment with 17ß-estradiol in young postpubertal rabbits. Acta Obstet Gynecol Scand 1999; 78: 357-62.
668. Ratcliffe A, Doherty M, Maini R, Hardingham T. Increased concentrations of proteoglycan components in the synovial fluids of patients with acute but not chronic joint disease. Ann Rheum Dis 1988; 47: 826-32.
669. Ravaglia G, Forti P, Maioli F, Nesi B, Pratelli L, Cucinotta D, et al. Body composition, sex steroids, IGF-1, and bone mineral status in aging men. J Gerontol A Biol Med Sci 2000; 55: M516-21.
670. Read RA, Cullis-Hill D, Jones MP. Systemic use of pentosan polysulphate in the treatment of osteoarthritis. J Small Anim Pract 1996; 37: 108-14.
671. Reboul P, Pelletier J-P, Tardif G, Cloutier J-M, Martel-Pelletier J. The new collagenase, collagenase-3, is expressed and synthesized by human chondrocytes but not by synoviocytes. A role in osteoarthritis. J Clin Invest 1996; 97: 2011-9.
672. Redini F, Daireaux M, Mauviel A, Galera P, Loyau G, Pujol JP. Characterization of proteoglycans synthesized by rabbit articular chondrocytes in response to transforming growth factor-beta (TGF-beta). Biochim Biophys Acta 1991; 1093: 196-206.
673. Redini F, Galera P, Mauviel A, Loyau G, Pujol J-P. Transforming growth factor-beta stimulates collagen and glycosaminoglycan biosynthesis in cultured rabbit articular chondrocytes. FEBS Lett 1988; 234: 172-6.
674. Redini F, Mauviel A, Loyau G, Pujol JP. Modulation of extracellular matrix metabolism in rabbit articular chondrocytes and human rheumatoid synovial cells by the non-steroidal anti-inflammatory drug etodolac. II: Glycosaminoglycan synthesis. Agents Actions 1990; 31: 358-67.
675. Rédini F, Mauviel A, Pronost S, Loyau G, Pujol J-P. Transforming growth factor ß exerts opposite effects from interleukin 1ß on cultured rabbit articular chondrocytes through reduction of interleukin-1 receptor expression. Arthritis Rheum 1993; 36: 44-50.
676. Rediske JJ, Koehne CF, Zhang B, Lotz M. The inducible production of nitric oxide by articular cell types. Osteoarthritis Cartilage 1994; 2: 199-206.
677. Reitamo S, Remitz A, Tamai K, Uitto J. Interleukin-10 modulates type 1 collagen and matrix

272.
metalloprotease gene expression in cultured human skin fibroblasts. J Clin Invest 1994; 94:2489-92.
678. Remick D, DeForge L, Sullivan J, Showell H. Profile of cytokines in synovial fluid specimens from patients with arthritis: Interleukin 8 (IL-8) and IL 6 correlate with inflammatory arthritides. Immunol Invest 1992; 21: 321-7.
679. René J, Weinberger M, Muzzuca SA, Brandt KD, Katz BP. Reduction of joint pain in patients with knee osteoarthritis who have received monthly telephone calls from lay personnel and whose medical treatment regimens have remained stable. Arthritis Rheum 1992; 35: 511-5.
680. Renoux M, Hilliquin P, Galoppin L, Florentin I, Menkes CJ. Release of mast cells mediators and nitrites in to knee joint fluid in osteoarthritis - comparison with articular chondrocalcinosis and rheumatoid arthritis. Osteoarthritis Cartilage 1996; 4: 175-9.
681. Renoux M, Hilliquin P, Galoppin L, Florentin J, Menkes CJ. Cellular activation products in osteoarthritis synovial fluid. Int J Clin Pharmacol Res 1995; 15: 135-8.
682. Revell P, Mayston V, Lalor P, Mapp P. The synovial membrane in osteoarthritis - a histological study including the characterization of the cellular infiltrate present in inflammatory osteoarthritis using monoclonal antibodies. Ann Rheum Dis 1988; 47: 300-7.
683. Rezzonico R, Burger D, Dayer J-M. Direct contact between T lymphocytes and human dermal fibroblasts or synoviocytes down-regulates types I and III collagen production via cell-associated cytokines. J Biol Chem 1998; 273: 18720-8.
684. Richardson DW, Dodge GR. Effects of interleukin-1ß and tumor necrosis factor- on expression of matrix related genes by cultured equine articular chondrocytes. Am J Vet Res 2000; 61:624-30.
685. Richmond RS, Carlson CS, Register TC, Shanker G, Loeser RF. Functional estrogen receptors in adult articular cartilage: estrogen replacement therapy increases chondrocyte synthesis of proteoglycans and insulin-like growth factor binding protein 2. Arthritis Rheum 2000; 43:2081-90.
686. Rico H. The therapy of osteoporosis and the importance of cortical bone. Calcif Tissue Int 1997; 61: 431-2.
687. Roberts AB, Sporn MB, Assoian RK, Smith JM, Roche NS, Wakefield LM, et al. Transforming growth factor type-beta - rapid induction of fibrosis and angiogenesis in vivo and stimulation of collagen formation in vitro. Proc Natl Acad Sci USA 1986; 83: 4167-71.
688. Robinson D, Bastian D, Hamer P, Pichiarallo D, Stephenson M. Mechanism of stimulation of prostaglandin synthesis by a factor from rheumatoid synovial tissue. Proc Natl Acad Sci USA 1981; 78: 5160-4.
689. Robinson DR, Dayer JM, Krane SM. Prostaglandins and their regulation in rheumatoid inflammation. Annals New York Acad Sci 1979; 332: 279-294.
690. Rocha JL, Fernandez-Alonso J. Acute tubulointerstitial nephritis associated with the selective COX-2 enzyme inhibitor, rofecoxib. Lancet 2001; 357: 1946-7.
691. Rogers A, Eastell R. Effects of estrogen therapy of postmenopausal women on cytokines measured in peripheral blood. J Bone Miner Res 1998; 13: 1577-86.
692. Rohde C, Anderson DE, Bertone AL, Weisbrode SE. Effects of phenylbutazone on bone activity and formation in horses. Am J Vet Res 2000; 61: 537-43.
693. Roivainen A, Soderstrom K, Pirila L, Aro H, Kortekangas P, Merilahti-Palo R, et al. Oncoprotein expression in human synovial tissue: an immunohistochemical study of different types of arthritis.

273.
Br J Rheumatol 1996; 35: 933-42.
694. Ronday H, Breedveld F, Verheijen J. The plasminogen activation system in rheumatoid arthritis. Fibrinolysis 1996; 10 (Suppl. 2): 95-8.
695. Ronday H, Smits H, Quax P, Van der Pluijm G, Löwik C, Breedveld F, et al. Bone matrix degradation by the plasminogen activation system. Possible mechanism of bone destruction in arthritis. Br J Rheumatol 1997; 36: 9-15.
696. Roodman GD. Cell biology of the osteoclast. Exp Hematol 1999; 27: 1229-41.
697. Rooney M, Condell D, Quinlan W, Daly L, Whelan A, Feighery C, et al. Analysis of the histologic variation of synovitis in rheumatoid arthritis. Arthritis Rheum 1988; 31: 956-63.
698. Rosenbaum J, Cugnini R, Tara D, Hefeneider S, Ansel J. Production and modulation of interleukin 6 synthesis by synoviocytes derived from patients with arthritic disease. Ann Rheum Dis 1992; 51: 198-202.
699. Rosner I, Goldberge V, Moskowitz R. Estrogens and osteoarthritis. Clin Orthop 1986; 213: 77-83.
700. Rosner I, Kresina T, Boja B, Moskowitz R, Goldberg V. Tamoxifen in experimental immune synovitis: effects on pathology and prostaglandins. Trans Orthop Res Soc 1984: 314 (Abstract).
701. Rosner I, Malemud C, Goldberg V, Papay R, Getzy L, Moskowitz R. Pathologic and metabolic responses of experimental osteoarthritis to estradiol and an estradiol antagonist. Clin Orthop 1982; 117: 280-6.
702. Rosner I, Manni A, Malemud C, Boja B, Moskowitz R. Estradiol receptors in articular chondrocytes. Biochem Biophys Res Commun 1982; 106: 1378-81.
703. Rosselli M, Imthurn B, Macas E, Keller P, Dubey R. Circulating nitrite/nitrate levels increase with follicular development: Indirect evidence for estradiol mediated NO release. Biochem Biophys Res Commun 1994; 202: 1543-52.
704. Ruh M, Bi Y, Dalonzo R, Bellone C. Effect of estrogens on IL-1-beta promoter activity. J Steroid Biochem Mol Biol 1998; 66: 203-10.
705. Russell R, McGuire M, Meats J, Ebsworth N, Beresford J. Intercellular messengers in joint tissues in rheumatoid arthritis. How disturbed control mechanisms may contribute to tissue destruction and repair. Scand J Rheumatol 1981; 40(Suppl.): 75-87.
706. Saarni H, Tammi M. Time and concentration dependence of the action of the cortisol on fibroblasts in vitro. Biochim Biophys Acta 1978; 540: 117-26.
707. Saarni H, Tammi M, Vuorio E. Effects of cortisol on glycosaminoglycans synthesised by normal and rheumatoid synovial fibroblasts in vitro. Scand J Rheumatol 1977; 6:222-4.
708. Saarni H, Tammi M, Vuorio E, Penttinen R. Distribution of glycosaminoglycans in rheumatoid cultures and effects of cortisol on it. Scand J Rheum 1980; 9: 11-6.
709. Sadouk M, Pelletier J-P, Tardif G, Kiansa K, Cloutier J, Martel-Pelletier J. Human synovial fibroblasts coexpress interleukin-1 receptor type I and type II mRNA: the increased level of the interleukin receptor in osteoarthritic cells is related to an increased level of the type I receptor. Lab Invest 1995; 73: 347-55.
710. Sadowski T, Steinmeyer J. Effects of non-steroidal antiinflammatory drugs and dexamethasone on the activity and expression of matrix metalloproteinase-1, matrix metalloproteinase-3 and tissue inhibitor of metalloproteinases-1 by bovine articular chondrocytes. Osteoarthritis Cartilage 2001;

274.
9: 407-15.
711. Saito H, Yanaihara T. Steroid formation in osteoblast-like cells. J Int Med Res 1998; 26: 1-12.
712. Saklatvala J. Induction of cartilage proteoglycan breakdown by IL-1. J Leukocyte Biol 1985; 37: 738
713. Saklatvala J. Tumour necrosis factor-alpha stimulates resorption and inhibits synthesis of proteoglycan in cartilage. Nature 1986; 322: 547-9.
714. Sakurai H, Kohsaka H, Liu M-F, Higashiyama H, Hirata Y, Kanno K, et al. Nitric oxide production and inducible nitric oxide synthase expression in inflammatory arthritides. J Clin Invest 1995; 96: 2357-63.
715. Salmon J, Higgs G, Vane J, Bitensky L, Chayen J, Henderson B, et al. Synthesis of arachidonate cyclo-oxygenase products by rheumatoid and nonrheumatoid synovial lining in nonproliferative organ culture. Ann Rheum Dis 1983; 42: 36-9.
716. Salvemini D. Regulation of cyclooxygenase enzymes by nitric oxide. Cell Moll Life Sci 1997; 53: 576-82.
717. Salvemini D, Misko T, Masferrer J, Seibert K, Currie M, Needleman P. Nitric oxide activates cyclooxygenase enzymes. Proc Natl Acad Sci USA 1993; 90: 7240-4.
718. Salvemini D, Settle SL, Masferrer JL, Seibert K, Currie MG, Needleman P. Regulation of prostaglandin production by nitric oxide; an in vivo analysis. Br J Pharmacol 1995; 114: 1171-8.
719. Samanta A, Jones A, Regan M, Wilson S, Doherty M. Is osteoarthritis in women affected by hormonal changes or smoking? Br J Rheumatol 1993; 32:366-70.
720. Sambrook P. The treatment of postemenopausal osteoporosis (Editorial). New Engl J Med 1995; 333: 1495-6.
721. Sandell LJ. Molecular biology of collagens in normal and osteoarthritic cartilage. In: Keuttner KE, Goldberg VM (Eds.) Osteoarthritic Disorders. Rosemont: American Academy of Orthopaedic Surgeons, 1995: 131-46.
722. Sandmark H, Hogstedt C, Lewold S, Vingard E. Osteoarthrosis of the knee in men and women in association with overweight, smoking, and hormone therapy. Ann Rheum Dis 1999; 58: 151-5.
723. Sandy J, Flannery C, Neame P, Lohmander L. The structure of aggrecan fragments in human synovial fluid: Evidence for the involvement in osteoarthritis of a novel proteinase which cleaves the Glu 373-Ala 374 bond of the interglobular domain. J Clin Invest 1992; 89: 1512-6.
724. Sangha O. Epidemiology of rheumatic disease. Rheumatology 2000; 39 (Suppl. 2): 3-12.
725. Santoro A, Rovati LC, Follet M, Setnikar I, Caplain H, Gualano V. Plasma levels of glyceryl trinitrate and dinitrates during application of three strengths of a new glyceryl trinitrate transdermal patch. Arzneim-Forsch/Drug Res 2000; 50 (II): 786-94.
726. Schaffer MR, Tantry U, Thornton FJ, Barbul A. Inhibition of nitric oxide synthesis in wounds: pharmacology and effect on accumulation of collagen in wounds in mice. Eur J Surg 1999; 165: 2662-7.
727. Schalkwijk J, Joosten LAB, Van den Berg WB, Van Wyk JJ, Van de Putte LB. Insulin-like growth factor stimulation of chondrocyte proteoglycan synthesis by human synovial fluid. Arthritis Rheum 1989; 32: 66-71.

275.
728. Scheidegger KJ, Cenni B, Picard D, Delafontaine P. Estradiol decreases IGF-1 and IGF-1 receptor expression in rat aortic smooth muscle cells. Mechanisms for atheroprotective effects. J Biol Chem 2000; 275: 38921-8.
729. Schemitsch EH, Turchin DC, Kowalski MJ, Swiontkowski MF. Quantitative assessment of bone injury and repair after reamed and unreamed locked intramedullary nailing. J Trauma 1998; 45: 250-5.
730. Schenck P, Schneider S, Miehlke R, Prehm P. Synthesis and degradation of hyaluronate by synovia from patients with rheumatoid arthritis. J Rheumatol 1995; 22: 400-5.
731. Schiffenbauer YS, Abramovitch R, Meir G, Nevo N, Holzinger M, Itin A, et al. Loss of ovarian function promotes angiogenesis in human ovarian carcinoma. Proc Natl Acad Sci USA 1997; 94: 13203-8.
732. Schini-Kerth VB. Dual effects of insulin-like growth factor-I on the constitutive and inducible nitric oxide (NO) synthase-dependent formation of NO in vascular cells . J Endocrinol Invest 1999; 22 (Suppl.): S82-8.
733. Schini VB, Catovsky S, Schray-Utz B, Busse R, Vanhoutte PM. Insulin-like growth factor I inhibits induction of nitric oxide synthase in vascular smooth muscle cells . Circ Res 1994; 74:24-32.
734. Schneider TA, Budsberg SC. Plasma and synovial concentrations of carprofen in dogs with chronic osteoarthritis. Vet Comp Orthop Traumatol 2001; 14: 19-24.
735. Schneiderman R, Rosenberg N, Hiss J, Lee P, Liu F, Hintz RL, et al. Concentration and size distribution of insulin-like growth factor-I in human normal and osteoarthritic synovial fluid and cartilage. Arch Biochem Biophys 1995; 324: 173-88.
736. Schouten JS, Van den Ouweland FA, Valkenburg HA, Lamberts SW. Insulin-like growth factor-1: a prognostic factor of knee osteoarthritis. Br J Rheumatol 1993; 32: 274-80.
737. Schumacher H. Exploring the synovium in 1990 (Editorial). Br J Rheumatol 1990; 29: 3-5.
738. Schumacher HR. Synovial inflammation, crystals, and osteoarthritis . J Rheumatol 1995; 22 (Suppl. 43): 101-3.
739. Schwartz Z, Finer Y, Nasatzky E, Soskolne W, Dean D, Boyan B, et al. The effects of 17ß-estradiol on chondrocyte differentiation are modulated by vitamin D3 metabolites. Endocrine 1997; 7: 209-18.
740. Scott BB, Weisbrot LM, Greebwood JD, Bogoch ER, Paige CJ, Keystone EC. Rheumatoid arthritis synovial fibroblast and U937 macrophage/monocyte cell line interaction in cartilage degradation. Arthritis Rheum 1997; 40: 490-8.
741. Seitz M, Loetscher P, Dewald B, Towbin H, Ceska M, Baggiolini M. Production of interleukin-1 receptor antagonist, inflammatory chemotactic proteins, and prostaglandin E by rheumatoid and osteoarthritic synoviocytes - Regulation by IFN gamma and IL-4. J Immunol 1994; 152: 2060-5.
742. Seitz M, Loetscher P, Dewald B, Towbin H, Gallati H, Baggiolini M. Interleukin-10 differentially regulates cytokine inhibitor and chemokine release from blood mononuclear cells and fibroblasts. Eur J Immunol 1995; 25: 1129-32.
743. Seouw HF, Rothel JS, Wood PR. Expression and purification of recombinant interleukin-1 from Escherichia coli. Vet Immunol Immunopathol 1994; 41: 229-39.
744. Serni U, Fibbi G, Anichini E, Zamperini A, Pucci M, Mannoni A, et al. Plasminogen activator

276.
and receptor in osteoarthritis. J Rheumatol 1995; 22 (Suppl. 43): 120-2.
745. Setton LA, Elliott DM, Mow VC. Altered mechanics of cartilage with osteoarthritis: Human osteoarthritis and an experimental model of joint degeneration. Osteoarthritis Cartilage 1999; 7: 2-14.
746. Setton LA, Mow VC, Howell DS. Mechanical behavior of articular cartilage in shear is altered by transection of the anterior cruciate ligament. J Orthop Res 1995; 13: 473-82.
747. Sharif M, George E, Shepstone L, Knudson W, Thonar EJ-MA, Cushnaghan J, et al. Serum hyaluronic acid level as a predictor of disease progression in osteoarthritis of the knee. Arthritis Rheum 1995; 38: 760-7.
748. Shevde NK, Bendixen AC, Dienger KM, Pike JW. Estrogens suppress RANK ligand-induced osteoclast differentiation via a stromal cell independant mechanism involving c-Jun repression. Proc Natl Acad Sci USA 2000; 97: 7829-34.
749. Shigeru Kea. IL-17 in synovial fluids from patients with rheumatoid arthritis is a potent stimulator of osteoclastogenesis. J Clin Invest 1999; 103: 1345-52.
750. Shimizu C, Coutts R, Healey R, Kubo T, Hirasawa Y, Amiel D. Method of histomorphometric assessment of glycosaminoglycans in articular cartilage. J Orthop Res 1997; 15: 670-74.
751. Shimizu C, Kubo T, Hirasawa Y, Coutts R, Amiel D. Histomorphometric and biochemical effect of various hyaluronans on early osteoarthritis. J Rheumatol 1998; 25: 1813-9.
752. Shindo T, Ikeda U, Ohkawa F, Kawahara Y, Yokoyama M, Shimada K. Nitric oxide synthesis in cardiac myocytes and fibroblasts by inflammatory cytokines. Cardiovasc Res 1995; 29: 813-9.
753. Shiozawa S, Tokuhisa T. Contribution of synovial mesenchymal cells to the pathogenesis of rheumatoid arthritis. Semin Arthritis Rheum 1992; 21: 267-73.
754. Shlopov BV, Smith GN, Cole AA, Hasty KA. Differential patterns of response to doxycycline and transforming growth factor ß1 in the down-regulation of collagenases in osteoarthritic and normal human chondrocytes. Arthritis Rheum 1999; 42: 719-27.
755. Shlopov BV, Stuart JM, Gumanovskaya ML, Hasty KA. Regulation of cartilage collagenase by doxycycline . J Rheumatol 2001; 28: 835-42.
756. Shors TJ, Pickett J, Wood G, Paczynski M. Acute stress persistently enhances estrogen levels in the female rat. Stress 1999; 3: 163-71.
757. Shughrue P, Lane M, Scrimo P, Merchenthaler I. Comparative distribution of estrogen receptor-a (ER- ) and ß (ER-ß) mRNA in the rat pituitary, gonad, and reproductive tract. Steroids 1998; 63: 498-504.
758. Silberberg M, Silberberg R. Modifying action of estrogen on the evolution of osteoarthritis in mice of different ages. J Endocrinol 1963; 72: 449-51.
759. Simkin P. Physiology of normal and abnormal synovium. Semin Arthritis Rheum 1991; 21: 179-83.
760. Simpson DJ. The effect of total meniscectomy versus caudal pole hemimeniscectomy on the stifle joint of the sheep. MVetClinStud thesis, University of Sydney, 1994.
761. Sims TJ, Avery NC, Bailey AJ. Quantitative determination of collagen crosslinks. Methods Mol Biol 2000; 139: 11-26.

277.
762. Sinigaglia L, Varenna M, Binelli L, Bartucci F, Arrigoni M, Ferrara R, et al. Urinary and synovial pyridinium crosslink concentrations in patients with rheumatoid arthritis and osteoarthritis. Ann Rheum Dis 1995; 54: 144-7.
763. Sisson J, Castor C, Klavons J. Connective tissue activation XVIII: Stimulation of hyaluronic acid synthetase activity. J Lab Clin Med 1980; 96: 189-97.
764. Sloan TB, Martincic RR, Anderson B. Differences in antigen composition of rheumatoid arthritic (RA) and non-RA derived tissue cultured synovial cells as detected by monkey anti-RA synovial cell sera. J Rheumatol 1981; 8: 204-13.
765. Smiley J, Hoffman W, Moore S, Paradies L. The humoral immune response of the rheumatoid synovium [Review]. Semin Arthritis Rheum 1985; 14: 151-62.
766. Smith C. Properties of synovial cells in culture. J Exp Med 1971; 134 (Suppl.): S306-12.
767. Smith C, Hamerman D. Significance of persistant differences between normal and rheumatoid synovial membrane cells in culture. Arthritis Rheum 1969; 12: 639-45.
768. Smith M, Triantafillou S, Parker A, Youssef P, Coleman M. Synovial membrane inflammation and cytokine production in patients with early osteoarthritis. J Rheumatol 1997; 24: 365-71.
769. Smith MM, Engle JM, Hwa SY, Ghosh P. Estrogen replacement therapy restores proteoglycan synthesis in patellar cartilage of osteoarthritic sheep. Osteoarthritis Cartilage 2000; 8: S55-56 (Abstract).
770. Smith M, Ghosh P. The synthesis of hyaluronic acid by human synovial fibroblasts is influenced by the nature of the hyaluronate in the extracellular environment. Rheumatol Int 1987; 7: 113-22.
771. Smith M, Ghosh P. Osteoarthritis: current status and future directions. APLAR J Rheumatol 1998; 2: 27-52.
772. Smith M, Ghosh P, Numata Y, Bansal M. The effects of orally administered calcium pentosan polysulphate on inflammation and cartilage degradation produced in rabbit joints by intraarticular injection of a hyaluronate-polylysine complex. Arthritis Rheum 1994; 37: 125-36.
773. Smith M, Numata Y, Bowman L, Ghosh P. Calcium pentosan polysulfate given orally to rabbits with inflamed knee joints abrogates interleukin-1-induced antiproliferative effects and reduces nitric oxide but not interleukin-6 activity in ex vivo cultures of synovial fibroblasts. Arthritis Rheum 1995; 38: 1312 (Abstract).
774. Smith M, Numata Y, Bowman L, Ghosh P. Preservation of chondrocyte and synoviocyte metabolism (cytoprotection) by oral calcium pentosan polysulphate in a lapine model of inflammatory arthritis. Osteoarthritis Cartilage 1996; 4: 3ii-iii (Abstract).
775. Smith MM, Reichman LJA, Smith SM, Hwa S-Y, Ghosh P. Calcium pentosan polysulfate downregulates estrogen receptor presentation in synovial fibroblasts in an ovine model of osteoarthritis. Osteoarthritis Cartilage 2000; 8 (Suppl. B): S55 (Abstract)
776. Smyth CM, Reeve TS. Two aids in oestrogen receptor characterisation. Aust NZ J Surg 1982; 52: 161-4.
777. Somjen D, Weisman Y, Mor Z, Harell A, Kaye AM. Regulation of proliferation of rat cartilage and bone by sex steroid hormones. J Steroid Biochem Mol Biol 1991; 40: 717-23.
778. Sowers M, Hochberg M, Crabbe J, Muhich A, Crutchfield M, Updike S. Association of bone mineral density and sex hormone levels with osteoarthritis of the hand and knee in premenopausal women. Am J Epidemiol 1996; 143: 38-47.

278.
779. Sowers M, Lachance L, Jamadar D, Hochberg MC, Hollis B, Crutchfield M, et al. The associations of bone mineral density and bone turnover markers with osteoarthritis of the hand and knee in pre- and perimenopausal women. Arthritis Rheum 1999; 42:483-9.
780. Sowers M, Zobel D, Weissfeld L, Hawthorne VM, Carman W. Progression of osteoarthritis of the hand and metacarpal bone loss: a twenty-year followup of incident cases. Arthritis Rheum 1991; 34: 36-42.
781. Spector T, Brown G, Silman A. Increased rates of previous hysterectomy and gynaecological operations in women with osteoarthritis . Br Med J 1988; 297: 899-900.
782. Spector T, Campion G. Generalized osteoarthritis is a hormonally mediated disease. Ann Rheum Dis 1989; 48: 523-7.
783. Spector T, Nandra D, Hart D, Doyle D. Is hormone replacement therapy protective for hand and knee osteoarthritis in women ?: The Chingford study. Ann Rheum Dis 1997; 56: 432-4.
784. Spector TD, Perry LA, Jubb RW. Endogenous sex steroid levels in women with generalized osteoarthritis. Clin Rheumatol 1991; 10: 316-9.
785. Spreng D, Sigrist N, Jungi T, Busato A, Lang JH, Pfister H, et al. Nitric oxide metabolite production in the cranial cruciate ligament, synovial membrane and articular cartilage of dogs with cranial cruciate ligament rupture. Am J Vet Res 2000; 61: 530-6.
786. Spreng D, Sigrist N, Schweighauser A, Busato A, Schawalder P. Endogenous nitric oxide production in canine osteoarthritis: detection in urine, serum, and synovial fluid specimens. Vet Surg 2001; 30: 191-9.
787. Srivastava K, Mustafa T. Ginger (Zingiber officinale) in rheumatism and musculoskeletal disorders. Med Hypoth 1992; 39: 342-8.
788. Srivastava SK, Weitzmann MN, Chaudhari LR, Namba N, Pacifici R. Estrogen decreases the resposiveness of osteoblast precursors to OPGL by down regulating OPGL induced JNK activity. J Bone Miner Res 1999; 14 (Suppl. 1): S361 (Abstract).
789. St. Clair EW. Nitric oxide: Friend or foe in arthritis? J Rheumatol 1998; 25: 1451-3.
790. St. Clair EW, Wilkinson WE, Pisetsky DS, Sexton DJ, Drew R, Kraus VB, et al. The effects of intravenous doxycycline for rheumatoid arthritis. Arthritis Rheum 2001; 44: 1043-7.
791. Stadler J, Stefanovic-Racic M, Billiar TR, Curran RD, Mcintyre LA, Georgescu HI, et al. Articular chondrocytes synthesize nitric oxide in response to cytokines and lipopolysaccharide. J Immunol 1991; 147: 3915-20.
792. Stefanovic-Racic M, Meyers K, Meschter C, Coffey JW, Hoffman RA, Evans CH. N-Monomethyl arginine, an inhibitor of nitric oxide synthase, suppresses the development of adjuvant arthritis in rats. Arthritis Rheum 1994; 37: 1062-9.
793. Stefanovic-Racic M, Möllers MO, Miller LA, Evans CH. Nitric oxide and proteoglycan turnover in rabbit articular cartilage. J Orthop Res 1997; 15: 442-9.
794. Stefanovic-Racic M, Morales TI, Taskiran D, McIntyre LA, Evans CH. The role of nitric oxide in proteoglycan turnover by bovine articular cartilage organ cultures. J Immunol 1996; 156: 1213-20.
795. Stefanovic-Racic M, Stadler J, Georgescu H, Evans C. Nitric oxide synthesis and its regulation by rabbit synoviocytes. J Rheumatol 1994; 21: 1892-8.
796. Steinberg K, Freni-Titulaer L, dePuey E, Miller D, Sgoutas D, Coralli C, et al. Sex steroids and

279.
bone density in premenopausal and perimenopausal women. J Clin Endocrinol Metab 1988; 69: 533-9.
797. Stevens C, Blake D, Merry P, Revell P, Levick J. A comparative stidy by morphometry of the microvasculature in normal and rheumatoid synovium. Arthritis Rheum 1991; 34: 1508-13.
798. Stewart A, Black A, Robins SP, Reid DM. Bone-density and bone turnover in patients with osteoarthritis and osteoporosis. J Rheumatol 1999; 26: 622-6.
799. Stichtenoth DO, Fauler J, Zeidler H, Frolich JC. Urinary nitrate excretion is increased in patients with rheumatoid arthritis and reduced by prednisolone. Ann Rheum Dis 1995; 54: 820-4.
800. Strøm H, Svalastoga E. A quantitative assessment of the subchondral changes in osteoarthritis and its association to the cartilage degeneration. A histomorphometric investigation of normal and osteoarthritic canine hip joints. Vet Comp Orthop Traumatol 1993; 6: 198-201.
801. Studer R, Jaffurs D, Stefanovic-Racic M, Robbins PD, Evans CH. Nitric oxide in osteoarthritis. Osteoarthritis Cartilage 1999; 7: 377-9.
802. Studer RK, Georgescu HI, Miller LA, Evans CH. Inhibition of transforming growth factor-ß production by nitric oxide-treated chondrocytes: implications for matrix synthesis. Arthritis Rheum 1999; 42: 248-57.
803. Studer RK, Levicoff E, Georgescu H, Miller L, Jaffurs D, Evans CH. Nitric oxide inhibits chondrocyte response to IGF-1: inhibition of IGF-IRbeta tyrosine phosphorylation. Am J Physiol 2000; 279: C961-969.
804. Su S, DiBattista JA, Sun Y, Li WQ, Zafarullah M. Up-regulation of tissue inhibitor of metalloproteinases-3 gene expression by TGF-ß in articular chondrocytes is mediated by serine/threonine and tyrosine kinases. J Cell Biochem 1998; 70: 517-27.
805. Summers K, O'Donnell J, Heiser A, Highton J, Hart D. Synovial fluid transforming growth factor ß inhibits dendritic cell-T lymphocyte interactions in patients with rheumatoid arthritis . Arthritis Rheum 1999; 42: 507-18.
806. Summers K, O'Donnell J, Heiser A, Highton J, Hart DNJ. Synovial fluid transforming growth factor ß inhibits dendritic cell-T lymphocyte interactions in patients with chonic arthritis. Arthritis Rheum 1999; 42: 507-18.
807. Swann AC, Seedhom BB. Improved techniques for measuring the indentation and thickness of articular cartilage. Proc Inst Mech Eng [H] 1989; 203: 143-50.
808. Swierstra B. Experimental arthrosis: Early changes in synovial morphology and lysosomal enzyme activities. Acta Orthop Scand. 1983; 54: 317-21.
809. Szekanecz Z, Haines G, Koch A. Differential expression of the urokinase receptor (CD87) in arthritic and normal synovial tissues. J Clin Pathol 1997; 50: 314-9.
810. Taguchi K, Hirano T, Iwasaki K, Sugihara H. Reconstruction culture system simulating human synovium: A three-dimensional collagen gel culture of synoviocytes. Cell Struct 1997; 22: 443-53.
811. Tajima Y, Yokose S, Kawasaki M, Takuma T. Ovariectomy causes cell proliferation and matrix synthesis in the growth plate cartilage of the adult rat. Histochem J 1998; 30: 467-72.
812. Takagi M, Santavirta S, Ida H, Ishii M, Mandelin J, Konttinen YT. Matrix metalloproteinases and tissue inhibitors of metalloproteinases in loose artificial hip joints. Clin Orthop 1998; 35-45.
813. Takahashi K, Hashimoto S, Kubo T, Hirasawa Y, Lotz M, Amiel D. Hyaluronan suppressed nitric

280.
oxide production in the meniscus and synovium of rabbit osteoarthritis model. J Orthop Res 2001; 19: 500-3.
814. Takahashi S, Inoue T, Higaki M, Mizushima Y. Suppressive effects of the new antirheumatic drug KE-298 on TNF- -induced production of matrix metalloproteinases but not of tissue inhibitor-1 of metalloproteinases in human rheumatoid synoviocytes. Drugs Exptl Clin Res 1998; 24: 67-71.
815. Takahashi T, Kondoh T, Kamei K, Seki H, Fukada M, Nagai H, et al. Elevated levels of nitric oxide in synovial fluid from patients with temporomandibular disorders. Oral Surg Oral Med Oral Pathol Oral Radiol Endod 1997; 82: 505-9.
816. Takeshita S, Mizuno S, Kikuchi T, Yamada H, Namiki O, Kumagai K. The in vitro effect of hyaluronic acid on IL-1b production in cultured rheumatoid synovial cells. Biomed Res 1997; 18: 187-94.
817. Taketazu F, Kato M, Gobl A, Ichijo H, Ten Dijke P, Itoh J, et al. Enhanced expression of transforming growth factor-ßs and transforming growth factor-ß type II receptor in the synovial tissues of patients with rheumatoid arthritis. Lab Invest 1994; 70: 620-30.
818. Takeuchi K, Ukawa H, Konaka A, Kitamura M, Sugawa Y. Effect of nitric oxide-releasing aspirin derivative on gastric functional and ulcerogenic responses in rats: comparison with plain aspirin. J Pharmacol Exp Ther 1998; 286: 115-21.
819. Takizawa A, Ohuchi E, Yamanaka H, Nakamura H, Ikeda E, Ghosh P, et al. Production of tissue inhibitor of metalloproteinases 3 is selectively enhanced by calcium pentosan polysulfate in human rheumatoid synovial fibroblasts. Arthritis Rheum 2000; 43: 812-20.
820. Tardif G, Pelletier J, Dupuis M, Geng C, Cloutier J, Martel-Pelletier J. Collagenase 3 production by human osteoarthritic chondrocytes in response to growth factors and cytokines is a function of the physiological state of the cells. Arthritis Rheum 1999; 42: 1147-58.
821. Tardif G, Reboul P, Pelletier JP, Geng C, Cloutier JM, Martel-Pelletier J. Normal expression of type 1 insulin-like growth factor receptor by human osteoarthritic chondrocytes with increased expression and synthesis of insulin-like growth factor binding proteins. Arthritis Rheum 1996; 39: 968-78.
822. Taskiran D, Stefanovic-Racic M, Georgescu H, Evans C. Nitric oxide mediates suppression of cartilage proteoglycan synthesis by interleukin-1. Biochem Biophys Res Commun 1994; 200: 142-8.
823. Taylor D. Interleukin-4 (IL-4) induces down-modulation and shedding of the p55 tumour necrosis factor receptor and inhibits TNF 's effect on rheumatoid synovial fibroblasts. Rheumatol Int 1994; 14: 21-5.
824. Testa V, Capasso G, Maffulli N, Sgambato A, Ames P. Proteases and antiproteases in cartilage homeostasis. Clin Orthop 1994; 79-84.
825. Tetlow LC, Lees M, Ogata Y, Nagase H, Woolley DE. Differential expression of gelatinase B (MMP-9) and stromelysin-1 (MMP-3) by rheumatoid synovial cells in vitro and in vivo. Rheumatol Int 1993; 13: 53-9.
826. Theoret C, Barber S, Gordon J. The expression of IL-1, IL-6, and TGF-ß in the synovial fluid of horses with surgically-induced transient synovititis. Vet Comp Orthop Traumatol 1998; 11:141-5.
827. Tomita M, Sato EF, Nishikawa M, Yamano Y, Inoue M. Nitric oxide regulates mitochondrial respiration and functions of articular chondrocytes. Arthritis Rheum 2001; 44: 96-104 .

281.
828. Tortorella MD, Burn TC, Pratta MA, Abbaszade I, Hollis JM, Liu R, et al. Purification and cloning of aggrecanase-1: A member of the ADAMTS family of proteins. Science 1999; 284: 1664-6.
829. Towheed T, Anastassiades T. Glucosamine therapy for osteoarthritis (Editiorial). J Rheumatol 1999; 26: 2294-7.
830. Towle C, Hung H, Bonassar L, Treadwell B, Mangham D. Detection of interleukin-1 in the cartilage of patients with osteoarthritis: A possible autocrine/paracrine role in pathogenesis. Osteoarthritis Cartilage 1997; 5: 293-300.
831. Towle C, Lalanandham T, Pavia M, Treadwell B. Involvement of an interleukin-1 like factor in osteoarthritis. J Leukocyte Biol 1985; 37: 749
832. Trontzas P, Kamper EF, Potamianou A, Kyriazis NC, Kritikos H, Stavridis J. Comparative study of serum and synovial fluid interleukin-11 levels in patients with various arthritides. Clin Biochem 1998; 31: 673-9.
833. Truppe W, Basner R, von Figura K, Kresse H. Uptake of hyaluronate by cultured cells. Biochem Biophys Res Commun 1977; 78: 713-9.
834. Tsai C-L, Liu T-K. Inhibition of estradiol-induced early osteoarthritis changes by tamoxifen. Life Sci 1992; 50: 1943-51.
835. Tsai C-L, Liu T-K. Osteoarthritis in women: Its relationship to estrogen and current trends. Life Sci 1992; 50: 1737-44.
836. Tsai C-L, Liu T-K. Up-regulation of estrogen receptors in rabbit osteoarthritic cartilage. Life Sci 1992; 50: 1727-35.
837. Tsai C-L, Liu T-K. Estradiol-induced knee osteoarthrosis on ovariectomized rabbits. Clin Orthop 1993; 291: 295-302.
838. Tsai C, Liu T, Chen T. Estrogen and osteoarthritis: A study of synovial estradiol and estradiol receptor binding in human osteoarthritic knees. Biochem Biophys Res Commun 1992; 183: 1287-91.
839. Tsukahara H, Gordienko DV, Tonshoff B, Gelato MC, Goligorsky MS. Direct demonstration of insulin-like growth factor-I-induced nitric oxide production by endothelial cells. Kidney Int 1994; 45: 598-604.
840. Turner AS, Alvis M, Myers W, Stevens ML, Lundy MW. Changes in bone mineral density and bone-specific alkaline phosphatase in ovariectomized ewes. Bone 1995; 17:( 4 Suppl.): 395S-402S.
841. Turner A, Athanasiou K, Zhu C-F, Alvis M, Bryant H. Biomechanical effects of estrogen on articular cartilage in ovariectomized sheep. Osteoarthritis Cartilage 1997; 5: 63-9.
842. Turner A, Eckhoff D, Dewell R, Allen M, Nett T, Aberman H. Ovariectomy causes up-regulation of specific estrogen receptors in articular cartilage. Trans Orthop Res Soc 1998: 180 (Abstract).
843. Turner AS, Mallinckrodt CH, Alvis MR, Bryant HU. Dose-response effects of estradiol implants on bone mineral density in ovariectomized ewes. Bone 1995; 17 (4 Suppl.): 421S-427S.
844. Turner AS, Mallinckrodt CH, Alvis MR, Bryant HU. Dual-energy X-ray absorptiometry in sheep: Experiences with in vivo and ex vivo studies. Bone 1995; 17 (4 Suppl.): 381S-387S.
845. Turner AS, Villanueva AR, Alvis MR, Aberman HM. Unusual histomorphometric changes in the

282.
iliac crest in ovariectomized and sham-operated ewes. Vet Comp Orthop Traumatol 1995; 8: 184-90.
846. Uebelhart D, Thonar E, Pietryla D, Williams J. Elevation in urinary levels of pyridinium cross-links of collagen following chymopapain-induced degradation of articular cartilage in the rabbit knee provides evidence of metabolic changes in bone. Osteoarthritis Cartilage 1993; 1: 185-92. Abstract.
847. Unemori E, Hibbs M, Amento E. Constitutive expression of a 92-kD gelatinase (Type V collagenase) by rheumatoid synovial fibroblasts and its induction in normal human fibroblasts by inflammatory cytokines. J Clin Invest 1991; 88: 1656-62.
848. Urban J, Maroudas A, Bayliss MT, Dillon J. Swelling pressure of proteoglycans at the concentrations found in cartilaginous tissues. Biorheology 1979; 16: 447-64.
849. Ushiyama T, Inoue K, Nishioka J. Expression of estrogen receptor related protein (p29) and estradiol binding in human arthritic synovium. J Rheumatol 1995; 22: 421-6.
850. Ushiyama T, Ueyama H, Inoue K, Nishioka J, Ohkubo I. Estrogen receptor gene polymorphism and generalized osteoarthritis. J Rheumatol 1998; 25:134-7.
851. Ushiyama T, Ueyama H, Inoue K, Ohkubo I, Hukuda S. Expression of genes for estrogen receptors and ß in human articular chondrocytes. Osteoarthritis Cartilage 1999; 7: 560-6.
852. van Beuningen HM, van der Kraan PM, Arntz OJ, van den Berg WB. Protection from interleukin 1 induced destruction of articular cartilage by transforming growth factor ß: Studies in anatomically intact cartilage in vitro and in vivo. Ann Rheum Dis 1993; 52: 185-91.
853. Van Bree H, Justus C, Quirke J. Preliminary observations on the effects of meloxicam in a new model for acute intra-articular inflammation in dogs. Vet Res Commun 1994; 18: 217-24.
854. van de Loo FA, Arntz OJ, van Enckevort FH, van Lent PL, van den Berg WB. Reduced cartilage proteoglycan loss during zymosan-induced gonarthritis in NOS2-deficient mice and in anti-interleukin-1-treated wild-type mice with unabated joint inflammation. Arthritis Rheum 1998; 41: 634-46.
855. van de Loo FA, Joosten LAB, van Lent PLE, Arntz OJ, van den Berg WB. Role of interleukin-1, tumor necrosis factor a, and interleukin-6 in cartilage proteoglycan metabolism and destruction. Effect of in situ blocking in murine antigen- and zymosan-induced arthritis. Arthritis Rheum 1995; 38: 164-72.
856. van de Loo FAJ, Joosten LAB, van Lent PLEM, Arntz OJ, van den Berg WB. TNF- and IL-6 in cartilage proteoglycan metabolism and destruction. Arthritis Rheum 1995; 38: 164-72.
857. van den Berg WB. Joint inflammation and cartilage destruction may occur uncoupled. Springer Semin Immunopathol 1998; 20: 149-64.
858. van den Berg WB, van de Loo F, Joosten LAB, Arntz OJ. Animal models of arthritis in NOS2-deficient mice. Osteoarthritis Cartilage 1999; 7: 413-5.
859. van den Berg WB, van der Kraan PM, van Beuningen HM. Role of growth factors and cartilage repair. In: Reginster J-Y, Pelletier J-P, Martel-Pelletier J, Henrotin Y (Eds.) Osteoarthritis: Clinical and experimental aspects. Berlin : Springer-Verlag, 1999: 188-209.
860. van Linthoudt D, Beutler A, Clayburne G, Sieck M, Fernandes L, Schumacher H. Morphometric studies on synovium in advanced osteoarthritis: Is there an association between apatite-like material and collagen deposits? Clin Exp Rheumatol 1997; 15: 493-7.

283.
861. van Ryn J, Trummlitz G, Pairet M. COX-2 selectivity and inflammatory process. Curr Med Chem 2000; 7: 1145-61.
862. Vasilev V, Merker H-J, Vidinov N. Ultrastructural changes in the synovial membrane in experimentally-induced osteoarthritis of rabbit knee joint. Histol Histopath 1992; 7: 119-27.
863. Vaughan-Scott T, Taylor J. The pathophysiology and medical management of canine osteoarthritis. J SAfr Vet Assoc 1997; 68: 21-5.
864. Venn G, Billingham M, Hardingham T. Increased proteoglycan synthesis in cartilage in experimental canine osteoarthritis does not reflect a permanent change in chondrocyte phenotype. Arthritis Rheum 1995; 38: 525-31.
865. Venn G, Nietfeld J, Duits A, Brennan F, Arner E, Covington M, et al. Elevated synovial fluid levels of interleukin-6 and tumor necrosis factor associated with early experimental canine osteoarthritis. Arthritis Rheum 1993; 36: 819-26.
866. Verbruggen G, Veys E. Influence of an oversulphated heparinoid upon hyaluronate metabolism of the human synovial cell in vivo. J Rheumatol 1979; 6: 554-61.
867. Verschure P, Joosten L, Van de Loo F, Van den Berg W. IL-1 has no direct role in the IGF-1 non-responsive state during experimentally induced arthritis in mouse knee joints. Ann Rheum Dis 1995; 54: 976-82.
868. Vestergaard P, Hermann AP, Orskov H, Mosekilde L. Effect of sex hormone replacement on the insulin-like growth factor system and bone mineral: a cross-sectional and longitudinal study in 595 perimenopausal women participating in the Danish Osteoporosis Prevention Study. J Clin Endocrinol Metab 1999; 84: 2286-90.
869. Vignon E, Arlot M, Hartmann D, Moyen B, Ville G. Hypertrphic repair of articular cartilage in experimental osteoarthritis. Ann Rheum Dis 1983; 42: 82-8.
870. Villiger PM, Kusari AB, Ten Dijke P, Lotz M. IL-1ß and IL-6 selectively induce transforming growth factor-ß isoforms in human articular chondrocytes. J Immunol 1993; 151: 3337-44.
871. Vincenti M, Clark I, Brinckerhoff C. Using inhibitors of metalloproteinases to treat arthritis. Arthritis Rheum 1994; 8: 1115-26.
872. Vincenti MP, Coon CI, Brinckeroff CE. Nuclear factor kappa-ß P50 activates an element in the distal matrix metalloproteinase-1 promoter in interleukin-1-beta-stimulated synovial fibroblasts. Arthritis Rheum 1998; 41: 1987-94.
873. von Rechenberg B, Guenther H, McIlwraith CW, Leutenegger C, Frisbie DD, Akens MK, et al. Fibrous tissue of subchondral cystic lesions in horses produce local mediators and neutral metalloproteinases and cause bone resorption in vitro. Vet Surg 2000; 29: 420-9.
874. von Rechenberg B, McIllwraith CW, Akens MK, Frisbie DD, Leutenegger C, Auer JA. Spontaneous production of nitric oxide (NO), prostaglandin (PGE2) and neutral metalloproteinases (NMPs) in media of explant culture of equine synovial membrane and articular cartilage from normal and osteoarthritic joints . Equine Vet J 2000; 32: 140-50.
875. Waage A, Kaufmann C, Espevik T, Husby G. Interleukin-6 in synovial fluid from patients with arthritis. Clin Immunol Immunopathol 1989; 50: 394-8.
876. Wagner S, Fritz P, Einsele H, Sell S, Saal J. Evaluation of synovial cytokine patterns in rheumatoid arthritis and osteoarthritis by quantitative reverse transcription polymerase chain reaction. Rheumatol Int 1997; 16: 191-6.

284.
877. Wahl S, Allen J, Costa G, Wong H, Dasch J. Reversal of acute and chronic synovial inflammation by anti-transforming growth factor ß. J Exp Med 1993; 177: 225-30.
878. Wahl S, Allen J, Wong H, Dougherty S, Ellingsworth L. Antagonistic and agonistic effects of transforming growth factor-beta and IL-1 in rheumatoid synovium. J Immunol 1990; 145: 2514-9.
879. Walakovits LA, Moore VL, Bhardwaj N, Gallick GS, Lark MW. Detection of stromelysin and collagenase in synovial fluid from patients with rheumatoid arthritis and posttraumatic knee injury. Arthritis Rheum 1992; 35: 35-42.
880. Walker E, Boyd R, Wu D, Lukoschek M, Burr D, Radin E. Morphological and morphometric changes in synovial-membrane associated with mechanically induced osteoarthrosis. Arthritis Rheum 1991; 34: 515-24.
881. Walker GD, Fischer M, Gannon J, Thompson RC, Oegema TR. Expression of type-X collagen in osteoarthritis. J Orthop Res 1995; 13: 4-12.
882. Walker P, Dowson D, Longfield M, Wright V. "Boosted lubrication" in synovial joints by fluid entrapment and enrichment. Ann Rheum Dis 1968; 27: 512-20.
883. Wallace JL, Vergnolle N, Muscara MN, Asfaha S, Chapman K, McKnight W, et al. Enhanced anti-inflammatory effects of a nitric oxide-releasing derivative of mesalamine in rats. Gastroenterol 1999; 117: 557-66.
884. Wang H, Morris JF. Effects of oestrogen upon nitric oxide synthase NADPH-diaphorase activity in the hypothalamo-neurohypophysial system of the rat. Neurosci 1999; 88: 151-8.
885. Watson S, Tierney G. Matrix metalloproteinase inhibitors. A review. Biodrugs 1998; 9: 325-35.
886. Webb G, Westacott C, Elson C. Chondrocyte tumor necrosis factor receptors and focal loss of cartilage in osteoarthritis. Osteoarthritis Cartilage 1997; 5: 427-37.
887. Webb R, Baxter G, McBride D, Nordblom GD, Shaw MPK. The measurement of testosterone and oestradiol-17ß using iodinated tracers and incorporating an affinity chromatography extraction procedure. J Steroid Biochem 1985; 23: 1043-51.
888. Weinberg J, Pippen A, Greenberg C. Extravascular fibrin formation and dissolution in synovial tissue of patients with osteoarthritis and rheumatoid arthritis. Arthritis Rheum 1991; 34:996-1005.
889. Weinberger A, Simkin P. Plasma proteins in synovial fluids of normal human joints. Semin Arthritis Rheum 1989; 19: 66-76.
890. Weiner CP, Lizasoain I, Baylis SA, Knowles RG, Charles IG, Moncada S. Induction of calcium-dependent nitric oxide syntheses by sex hormones. Proc Natl Acad Sci USA 1994; 91: 5212-6.
891. Welshons WV, Lieberman ME, Gorski J. Nuclear localization of unoccupied oestrogen receptors. Nature 1984; 307: 747-8.
892. Werb Z, Aggeler J. Proteases induce secretion of collagenase and plasminogen activator by fibroblasts. Proc Natl Acad Sci USA 1978; 75: 1839-43.
893. Werb Z, Mainardi C, Vater C, Harris E. Endogenous activation of latent collagenase by rheumatoid synovial cells. Evidence for a role of plasminogen activator. New Eng J Med 1977; 296: 1017-23.
894. Werner-Felmayer G, Werner E, Fuchs D, Hausen A, Reibnegger G, Wachter H. Tetrahydrobiopterin-dependant formation of nitrite and nitrate in murine fibroblasts. J Exp Med 1990; 172: 1599-1607.

285.
895. Wernicke D, Seyfert C, Hinzmann B, Gromnica-Ihle E. Cloning of collagenase 3 from the synovial membrane and its expression in rheumatoid arthritis and osteoarthritis. J Rheumatol 1996; 23: 590-5.
896. Westacott C, Sharif M. Cytokines in osteoarthritis: Mediators or markers of joint destruction. Semin Arthritis Rheum 1996; 25: 254-72.
897. Westacott CI, Webb GR, Warnock MG, Sims JV, Elson CJ. Alteration of cartilage metabolism by cells from osteoarthritic bone. Arthritis Rheum 1997; 407: 1282-91
898. Westacott CI, Whicher JT, Barnes IC, Thompson D, Swan AJ, Dieppe PA. Synovial fluid concentration of 5 different cytokines in rheumatic diseases. Ann Rheum Dis 1990; 49: 676-81.
899. Wilkinson L, Edwards J, Poston R, Haskard D. Expression of vascular cell adhesion molecule-1 in normal and inflamed synovium. Lab Invest 1993; 68: 82-8.
900. Wilkinson LS, Pitsillides AA, Worrall JG, Edwards JCW. Light microscopic characterization of the fibroblast-like synovial intimal cell (synoviocyte). Arthritis Rheum 1992; 35: 1179-84.
901. Wimalawansa DR, Wimalawansa SJ. Comparison of the beneficial effects of various nitirc oxide donor compounds on ovariectomy-induced osteoporosis. J Bone Miner Res 1999; 14 (Suppl. 1): S533 (Abstract).
902. Wimalawansa SJ. Nitroglyerin therapy is as efficacious as standard estrogen replacement therapy (Premarin) in prevention of oophorectomy-induced bone loss: a human pilot clinical study. J Bone Miner Res 2000; 15: 2240-4.
903. Wimalawansa SJ. Restoration of ovariectomy-induced osteopenia by nitroglycerin. Calcif Tissue Int 2000; 66: 56-60.
904. Wimalawansa SJ, Chapa MT, Yallampalli C, Zhang R, Simmons DJ. Prevention of corticosteroid-induced bone loss with nitric oxide donor nitroglycerin in male rats. Bone 1997; 21: 275-80.
905. Wimalawansa SJ, Chapa T, Fang L, Yallampalli C, Simmons D, Wimalawansa S. Frequency-dependant effect of nitric oxide donor nitroglycerin on bone. J Bone Miner Res 2000; 15:1119-25.
906. Wimalawansa SJ, De Marco G, Gangula P, Yallampalli C. Nitric oxide donor alleviates ovariectomy-induced bone loss. Bone 1996; 18: 301-4.
907. Wimalawansa S, Simmons D, Yallampalli C. In vivo effects of nitric oxide donor, nitroglycerin, on bone . J Bone Miner Res 1999; 14: S198 (Abstract).
908. Winchester R, Su F, Ritchlin C. Alteration of synoviocytes by inflammation: The source of a persistent nonimmunological drive in synovitis: Analysis of levels of messenger-RNA expression by a simple multigene assay. Clin Exp Rheum 1993; 11 (Suppl. 8): S87-S90
909. Witte MB, Thornton FJ, Efron DT, Barbul A. Enhancement of fibroblast collagen synthesis by nitric oxide. Nitric Oxide 2000; 4: 572-82.
910. Witter J, Roughley P, Webber C, Roberts N, Keystone E, Poole A. The immunologic detection and characterization of cartilage proteoglycan degradation products in synovial fluid of patients with arthritis. Arthritis Rheum 1987; 30: 519-29.
911. Wluka AE, Cicuttini FM, Spector TD. Menopause, oestrogens and arthritis. Maturitas 2000; 35: 183-99.
912. Wolff J. Das gesetz der transformation. Transformation der knochen. Berlin: Hirschwald, 1892.

286.
913. Wood D, Ihrie E. Presence of interleukin-1 in human arthritis. J Leukocyte Biol 1985; 37: 753
914. Wood D, Ihrie E, Hamerman D. Release of interleukin-1 from human synovial tissue in vitro. Arthritis Rheum 1985; 28: 853-61.
915. Woolley D, Brinckerhoff C, Mainardi C, Vater CEJ, Harris EJ. Collagenase production by rheumatoid synovial fibroblasts: morphological and immunohistochemical studies of the dendritic cell. Ann Rheum Dis 1979; 38: 262-70.
916. Wright JK, Cawston TE, Hazleman BL. Transforming growth factor beta stimulates the production of the tissue inhibitor of metalloproteinases (TIMP) by human synovial and skin fibroblasts. Biochim Biophys Acta 1991; 1094: 207-10.
917. Wynne-Roberts C, Castor C. Ultrastructural comparison of rheumatoid and nonrheumatoid synovial fibroblasts grown in tissue culture. Arthritis Rheum 1972; 15: 65-83.
918. Yamashiro T, Takano-Yamamoto T. Differential responses of mandibular condyle and femur to oestrogen deficiency in young rats. Arch Oral Biol 1998; 43: 191-5.
919. Yanni G, Whelan A, Feighery C, Quinlan W, Symons J, Duff G. Contrasting levels of in vitro cytokine production by rheumatoid synovial tissues demonstrating different patterns of mononuclear cell infiltration. Clin Exp Immunol 1993; 93: 387-95.
920. Yap PSK, Fung HL. Pharmacokinetics of nitroglycerin in rats. J Pharm Sci 1978; 67: 584-6.
921. Yaron I, Meyer F, Dayer J-M, Yaron M. Human recombinant interleukin-1-beta stimulates glycosaminoglycan production in human synovial fibroblast cultures. Arthritis Rheum 1987; 30: 424-30.
922. Yaron I, Meyer F, Dayer J, Bleiberg I, Yaron M. Some recombinant human cytokines stimulate glycosaminoglycan synthesis in human synovial fibroblast-cultures and inhibit it in human articular-cartilage cultures. Arthritis Rheum 1989; 32: 173-80.
923. Yasuoka T, Nakashima M, Okuda T, Tatematsu N. Effect of estrogen replacement on temporomandibular joint remodelling in ovariectomised rats. J Oral Maxillofac Surg 2000; 58: 189-96.
924. Yelin E, Callahan LF. The economic cost and social and psychological impact of musculoskeletal conditions.National Arthritis Data Work Groups. Arthritis Rheum 1995; 38: 1351-62.
925. Yoon HN, Chun WS, Park YY, Shim BS, Han WS, Kwon SW. Effects of estrogen on nitric oxide synthase and histological composition in the rabbit clitoris and vagina. Int J Impot Res 2001; 13: 205-11.
926. Yoshimi T, Kikuchi T, Obara T, Yamaguchi T, Sakakibara Y, Itoh H, et al. Effects of high-molecular-weight sodium hyaluronate on experimental osteoarthrosis induced by the resection of rabbit anterior cruciate ligament. Clin Orthop 1994; 298: 296-304.
927. Yoshioka M, Coutts R, Amiel D, Hacker S. Characterization of a model of osteoarthritis in the rabbit knee. Osteoarthritis Cartilage 1996; 4: 87-98.
928. Young P, Stack M. Estrogen and glucocorticoid receptors in adult canine articular cartilage. Arthritis Rheum 1882; 25: 568-73.
929. Yovich JV, Trotter GW, McIlwraith CW, Norrdin RW. Effect of polysulfated glycosaminoglycan (Adequan) upon the healing of physical and chemical defects in equine articular-cartilage. Vet Surg 1986; 15: 138

287.
930. Yovich JV, Trotter GW, McIlwraith CW, Norrdin RW. Effects of polysulfated glycosaminoglycan on chemical and physical defects in equine articular cartilage. Am J Vet Res 1987; 48: 1407-14.
931. Yu LP, Burr DB, Brandt KD, O'Connor BL, Rubinow A, Albrecht M. Effects of oral doxycycline administration on histomorphometry and dynamics of subchondral bone in a canine model of osteoarthritis. J Rheumatol 1996; 23: 137-42.
932. Zafarullah M, Pelletier J-P, Cloutier J-M, Martel-Pelletier J. Elevated metalloproteinase and tissue inhibitor of metalloproteinase mRNA in human osteoarthritic synovia. J Rheumatol 1993; 20: 693-7.
933. Zhang Y, Lin J-X, Vilcek J. Synthesis of interleukin 6 (interferon-ß2/B cell stimulatory factor 2) in human fibroblasts is triggered by an increase in intracellular cyclic AMP. J Biol Chem 1988; 263: 6177-82.
934. Zhang YQ, Mcalindon TE, Hannan MT, Chaisson CE, Klein R, Wilson PWF, et al. Estrogen replacement therapy and worsening of radiographic knee osteoarthritis. Arthritis Rheum 1998; 41: 1867-73.
935. Zhu W, Mow VC, Koob TJ, Eyre DR. Viscoelastic shear properties of articular cartilage and the effects of glycosidase treatments. J Orthop Res 1993; 11: 771-81.
936. Ziche M. Role of nitric oxide in the angiogenesis of avascular tissue. Osteoarthritis Cartilage 1999; 7: 403-5.
937. Ziche M, Parenti A, Ledda F, Dell'Era P, Granger HJ, Maggi CA, et al. Nitric oxide promotes proliferation and plasminogen activator production by coronary venular endothelium through endogenous bFGF. Circ Res 1997; 80: 845-52.
938. Zimmerman M, Quigley JP, Ashe B, Dorn C, Goldfarb R, Troll W. Direct fluorescent assay of urokinase and plasminogen activators of normal and malignant cells: kinetics and inhibitor profiles. Proc Natl Acad Sci USA 1978; 75: 750-3.