Embed Size (px)
Citation preview

, . 183: 109–115 (1997)
THE EFFECTS OF MATERNAL PROTEINDEPRIVATION ON THE FETAL RAT PANCREAS:MAJOR STRUCTURAL CHANGES AND THEIR
RECUPERATION
. . 1*, . 2, . . 1, . 1, . 1, . . 2 . . 1
1Department of Morbid Anatomy and Histopathology, The Medical and Dental School of St. Bartholomew’s and the Royal LondonSchool of Medicine and Dentistry, The Royal London Hospital, London E1 1BB, U.K.
2Department of Clinical Biochemistry, Addenbroke’s Hospital, Hills Road, Cambridge CB2 2QR, U.K.
SUMMARY
There is evidence that low birth weight and poor growth in early life cause a long-term predisposition to non-insulin-dependentdiabetes. Morphological changes were assessed in fetal rat pancreas subjected to both pre- and post-natal maternal protein deprivation(LP). Further groups were subjected to purely prenatal maternal protein deprivation (preLP) and purely postnatal maternal proteindeprivation (postLP), as well as a control group. The results show that the LP and postLP groups had fewer but larger islets than thecontrol group, while the preLP group had more numerous, smaller islets. All three low protein groups had more irregularly shaped isletsthan the control group. There was a reduction in the amount of beta cells within each islet in all three protein-deprived groups. The LPand postLP groups showed a reduction in the percentage of islet tissue and beta cells per pancreas, but the percentage of islet tissueexpressed per unit body weight was similar in all four groups. These results show that in maternal protein deprivation, homeostaticmechanisms ensure a constant amount of pancreatic endocrine tissue per unit of body weight. However, there remain major structuralchanges in the size, shape, and composition of the islets. These results support the theory that early development profoundly affects thestructure of the pancreas and may play a role in the later development of adult diseases, such as non-insulin-dependent diabetes mellitus.? 1997 by John Wiley & Sons, Ltd.
J. Pathol. 183: 109–115, 1997.No. of Figures 4. No. of Tables 0. No. of References 21.
KEY WORDS—pancreas; protein; development; diabetes; endocrine
INTRODUCTION
There is evidence that events in pregnancy andearly postnatal life affect long-term predisposition todiabetes, acting by an indirect mechanism affectinggrowth.1–3 The hypothesis under test is that this mech-anism involves changes in pancreatic development,producing metabolic defects evident in later life,leading to a ‘thrifty phenotype’.4 Rats exposed to amaternal low protein diet in utero show changes in thepancreas5 and liver6 which suggest a severe develop-mental disturbance and long-term alteration of isletactivity and of liver enzyme activities associatedwith glucose metabolism. Ultrastructural changes inpancreatic beta cells have also been described.7 Weconsidered that the morphological characterization ofthese abnormalities would allow identification of thecomponents of the glands which had been affected bythe treatment.
There are established links between non-insulin-dependent diabetes mellitus (NIDDM), hypertension,and death from cardiovascular disease, and furtherassociations of impaired glucose tolerance (IGT) withraised blood pressure and increased risk of ischaemicheart disease. This interrelationship (one-third of sub-jects with IGT go on to develop NIDDM within 10years1) and the former associations suggest that theadverse effects of early malnutrition on pancreatic devel-opment and insulin secretion may be crucial in determin-ing the adequacy of production of insulin in later life.Children who have suffered severe protein malnutritionin early life have an irreversible impairment of insulinsecretion.8,9 Birth weight and weight at 1 year areinversely related to the percentage of subjects with IGTand the concentration of 32–33 split pro-insulin inplasma, in men of age 64 years.1We studied the growth and differentiation of
pancreatic islets in healthy rat pups and in rats bredfrom dams fed a protein-deficient diet during gestationand during nursing. We also studied a prenatal low-protein group, in which the rats born to dams fed aprotein-deficient diet were nursed by dams fed anormal diet. A fourth group included rat pups whichwere nursed by dams on a protein-deficient diet. Amaternal low-protein diet leads to major changes in thedeveloping pancreas in the offspring.
*Correspondence to: D. M. Berney, Department of MorbidAnatomy and Histopathology, The Medical and Dental School of St.Bartholomew’s and the Royal London School of Medicine andDentistry, The Royal London Hospital, London E1 1BB, U.K.
Contract grant sponsors: British Diabetic Association; MedicalResearch Council; Parthenon Trust.
CCC 0022–3417/97/090109–07 $17.50 Received 31 May 1996? 1997 by John Wiley & Sons, Ltd. Accepted 12 March 1997

MATERIALS AND METHODS
Virgin female Wistar rats from the Dunn NutritionalLaboratory were housed individually in a room main-tained at 22)C and 45 per cent humidity under acontrolled 12 h light–dark cycle. They were mated withnormal males and day 0 of gestation was taken as theday on which vaginal plugs were expelled. The pregnantrats were allocated to two groups. One group of ratsreceived a diet containing 20 per cent protein and theother group received an isocaloric diet containing 8 percent protein. Casein was used as a source of protein.Details of the diet and method are given in ref. 10. Allthe animals were fed their diets ad libitum from thebeginning of pregnancy and during the entire gestationaland lactational period.Spontaneous delivery took place on day 22 of
pregnancy, after which, at 3 days of age, large litterswere reduced to eight pups, thus ensuring that thecontrol and its pair on a low protein diet had equalnumbers of pups to nurse. These pups were thenfollowed in the four groups listed below until 21 days:
- Group 1: a control group (C) consisting of theoffspring of mothers fed a control diet, nursed bymothers fed a control diet. n=22 (12 pregnancies).
- Group 2: a low-protein (LP) group consisting ofthe offspring of mothers fed a low-protein diet,nursed by a mother fed a low-protein diet. n=23(12 pregnancies).
- Group 3: a postnatal low-protein (postLP) groupconsisting of the offspring of mothers fed a controldiet, nursed by mothers fed a low-protein diet. n=23(12 pregnancies).
- Group 4: a prenatal low-protein (preLP) groupconsisting of mothers fed a low-protein diet,but nursed by mothers fed a control diet. n=24 (12pregnancies).
At 21 days, the body weights were recorded and thenthe rats were killed and the pancreases were dissectedout, weighed, and fixed in formalin. After at least 24 hfixation, they were embedded in wax. Two represen-tative 5 ìm sections were then cut at more than 50 ìmapart and immunohistochemically stained for insulinand glucagon. Using morphometric techniques in whichthe main observers were blinded as to the groups, thepancreatic tissue was assessed for the total tissue area,the area and perimeter of the islets, and the area thatwas positive for insulin and glucagon. Overall, a total of4970 islets were measured.All morphometry was carried out using a Magiscan
M2 semi-automatic image analysis system (AppliedImaging International, Sunderland, Tyne and Wear,U.K.). Areas of interest, which were darkly stainedagainst a paler background, were detected automaticallyon the basis of a predetermined intensity level. The totaltissue area in each section was measured at a magnifica-tion of 26·7 ìm per image pixel, corresponding to a fieldarea of 186·9 mm2. The area and perimeter of all isletson each section, as well as all alpha and beta cells in eachislet, were measured at a higher power (2·28 ìm perimage pixel, giving a field area of 1·36 mm2).
The following variables were measured: IA=islet area;BA=beta-cell area in one islet; AA=alpha-cell area inone islet; TTA=total tissue area; PW=pancreaticweight; BW=rat body weight; OA=area of each islet;OP=perimeter of each islet.The results were calculated using the following
expressions:
% Beta cell per islet=(ÓBA/ÓIA)#100
where Ó signifies the sum of each measured variable persection
% Alpha cell per islet=(ÓAA/ÓIA)#100Weight of islet tissue in each pancreas=
(ÓIA/TTA)#PWWeight of beta-cell tissue in each pancreas=
(ÓBA/TTA)#PWWeight of alpha-cell tissue in each pancreas=
(ÓAA/TTA)#PW% Islet tissue in each animal=
{((ÓIA/TTA)#PW)/BW}#100% Beta-cell tissue in each animal=
{((ÓBA/TTA)#PW)/BW}#100% Alpha-cell tissue in each animal=
{((ÓAA/TTA)#PW)/BW}#100
The results were compared with the control groupusing Student’s t-test. Prior to statistical testing, the isletarea values were transformed to their logarithms inorder that their distribution should be normal. All othervariables were normally distributed.
RESULTS
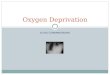
Individual islet morphologyThe size and shape of the islets differed significantly
between the four groups (Fig. 1a). The LP and postLPgroups had a significantly larger mean islet area than thecontrol group (P<0·05), while the preLP group had asmaller mean islet area (P<0·01). The logarithm of theislet area was tested, as this conformed to a normaldistribution. There were fewer islets in the LP andpostLP groups than in the control group (P<0·01), butmore islets in the preLP group (P<0·05).The shape of the islets was assessed by the ratio of the
area of each islet to its perimeter; thus, the islets with amore irregular shape have a smaller ratio. This showsthat the control group is strikingly different from theother three groups (Fig. 1b), which all show greaterirregularity in islet shape (P<0·001).
Beta cells and alpha cells in each islet
The percentage of beta cells per islet revealed all threematernal protein-deprived groups to have a smallerpercentage of beta cells than the control group (P<0·05).The differences were most marked in the LP groupand were intermediate in the preLP and postLP groups(Fig. 2).The percentage of alpha cells per islet suggests a
tendency for the maternal protein-deprived groups to
110 D. M. BERNEY ET AL.
? 1997 by John Wiley & Sons, Ltd. , . 183: 109–115 (1997)
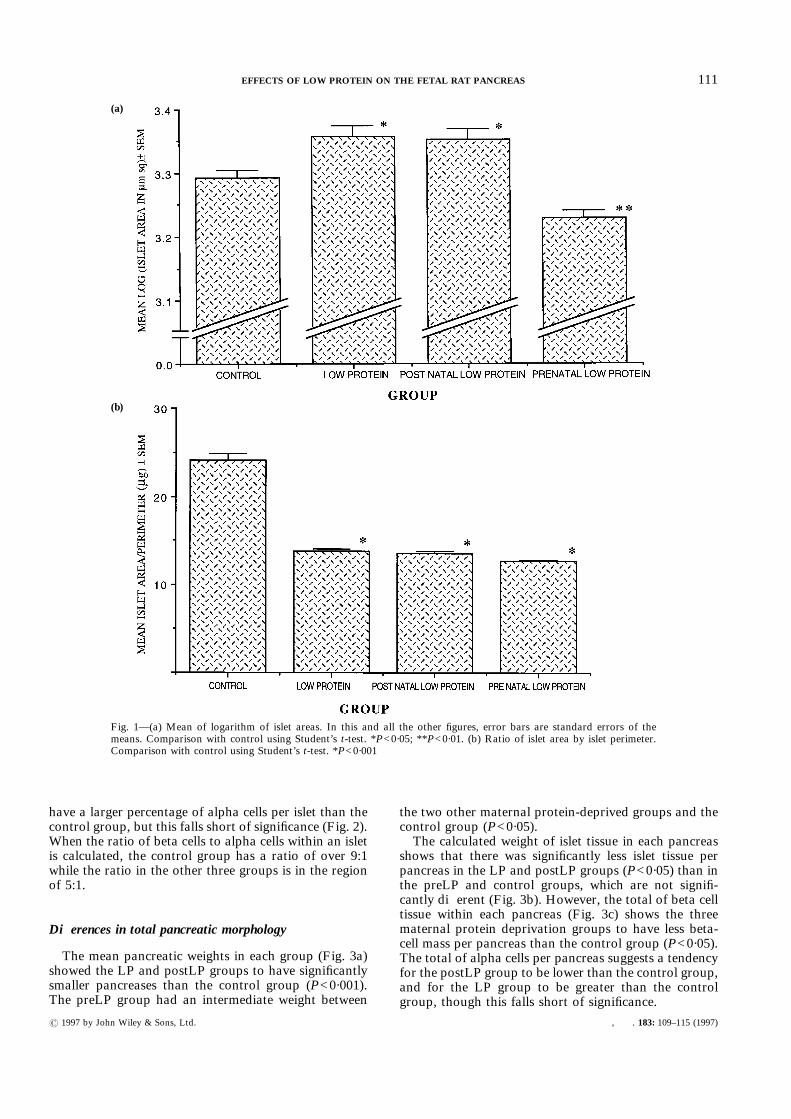
have a larger percentage of alpha cells per islet than thecontrol group, but this falls short of significance (Fig. 2).When the ratio of beta cells to alpha cells within an isletis calculated, the control group has a ratio of over 9:1while the ratio in the other three groups is in the regionof 5:1.
Differences in total pancreatic morphology
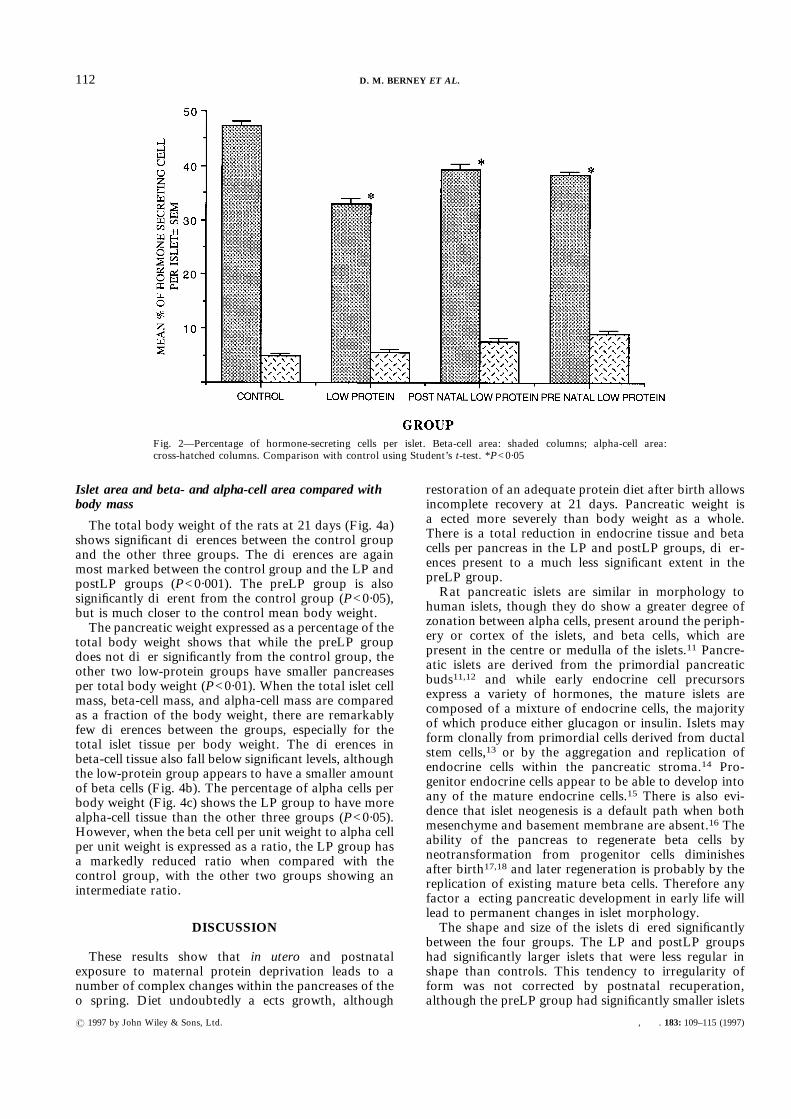
The mean pancreatic weights in each group (Fig. 3a)showed the LP and postLP groups to have significantlysmaller pancreases than the control group (P<0·001).The preLP group had an intermediate weight between
the two other maternal protein-deprived groups and thecontrol group (P<0·05).The calculated weight of islet tissue in each pancreas
shows that there was significantly less islet tissue perpancreas in the LP and postLP groups (P<0·05) than inthe preLP and control groups, which are not signifi-cantly different (Fig. 3b). However, the total of beta celltissue within each pancreas (Fig. 3c) shows the threematernal protein deprivation groups to have less beta-cell mass per pancreas than the control group (P<0·05).The total of alpha cells per pancreas suggests a tendencyfor the postLP group to be lower than the control group,and for the LP group to be greater than the controlgroup, though this falls short of significance.
Fig. 1—(a) Mean of logarithm of islet areas. In this and all the other figures, error bars are standard errors of themeans. Comparison with control using Student’s t-test. *P<0·05; **P<0·01. (b) Ratio of islet area by islet perimeter.Comparison with control using Student’s t-test. *P<0·001
(a)
(b)
111EFFECTS OF LOW PROTEIN ON THE FETAL RAT PANCREAS
? 1997 by John Wiley & Sons, Ltd. , . 183: 109–115 (1997)
Islet area and beta- and alpha-cell area compared withbody mass
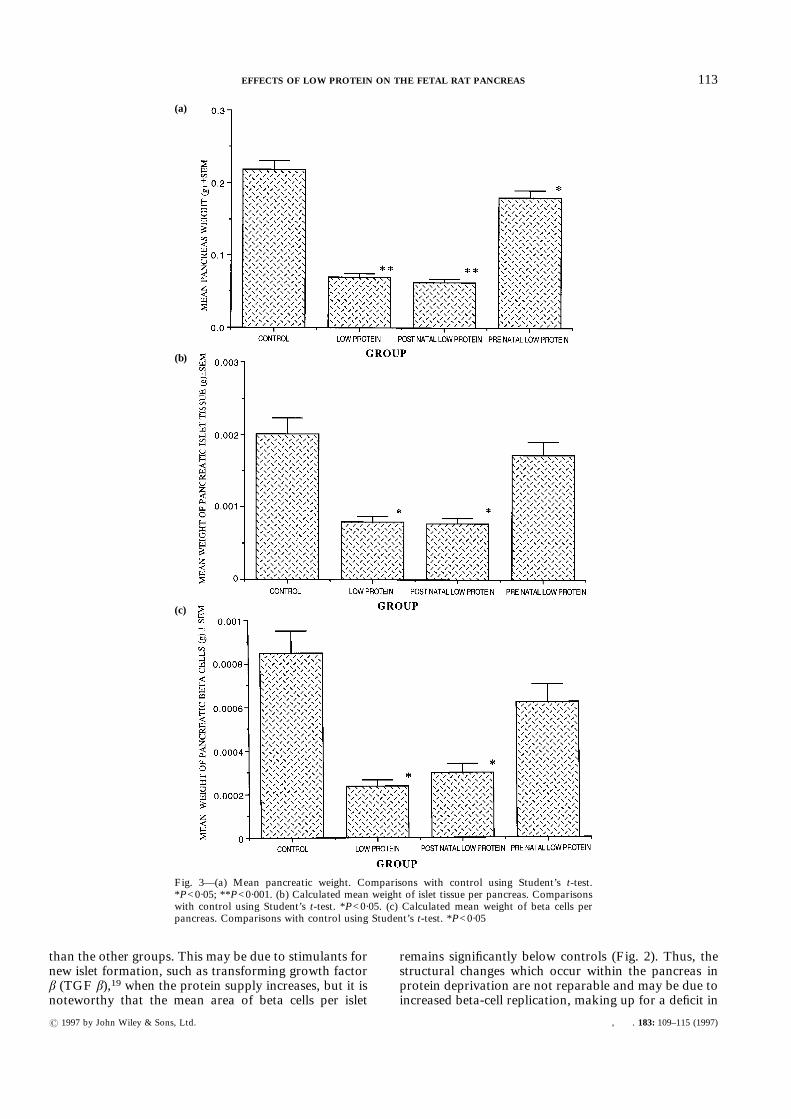
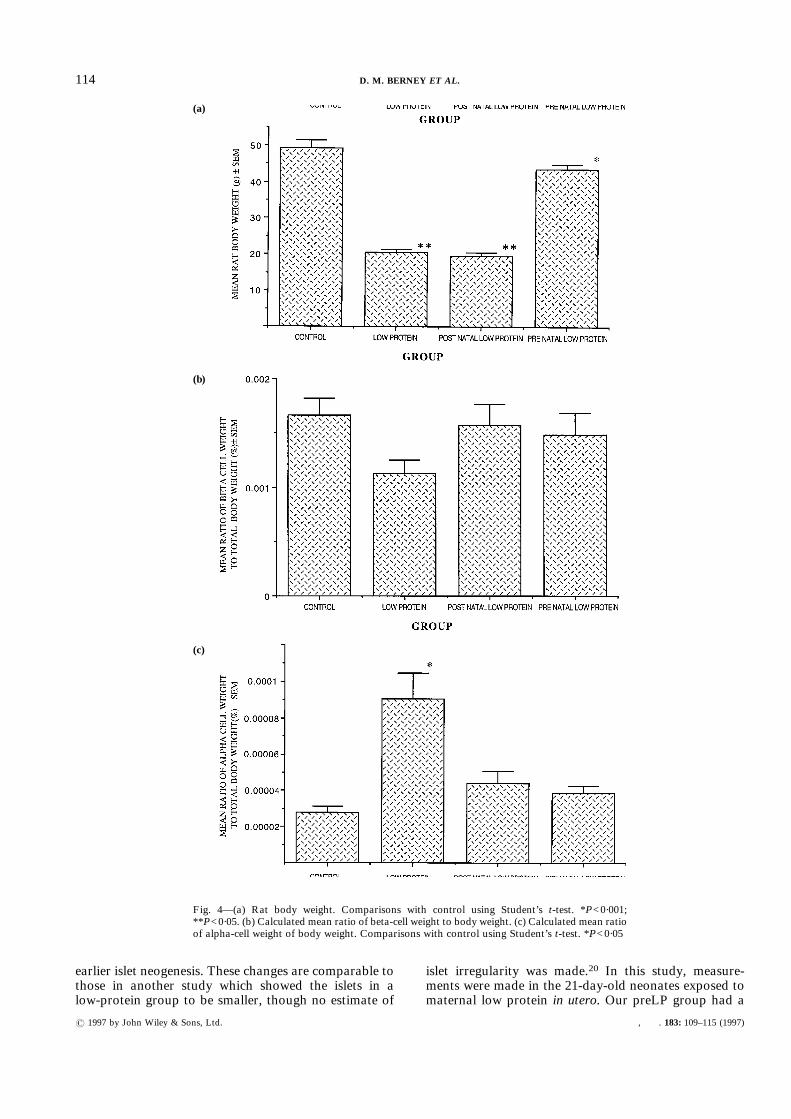
The total body weight of the rats at 21 days (Fig. 4a)shows significant differences between the control groupand the other three groups. The differences are againmost marked between the control group and the LP andpostLP groups (P<0·001). The preLP group is alsosignificantly different from the control group (P<0·05),but is much closer to the control mean body weight.The pancreatic weight expressed as a percentage of the
total body weight shows that while the preLP groupdoes not differ significantly from the control group, theother two low-protein groups have smaller pancreasesper total body weight (P<0·01). When the total islet cellmass, beta-cell mass, and alpha-cell mass are comparedas a fraction of the body weight, there are remarkablyfew differences between the groups, especially for thetotal islet tissue per body weight. The differences inbeta-cell tissue also fall below significant levels, althoughthe low-protein group appears to have a smaller amountof beta cells (Fig. 4b). The percentage of alpha cells perbody weight (Fig. 4c) shows the LP group to have morealpha-cell tissue than the other three groups (P<0·05).However, when the beta cell per unit weight to alpha cellper unit weight is expressed as a ratio, the LP group hasa markedly reduced ratio when compared with thecontrol group, with the other two groups showing anintermediate ratio.
DISCUSSION
These results show that in utero and postnatalexposure to maternal protein deprivation leads to anumber of complex changes within the pancreases of theoffspring. Diet undoubtedly affects growth, although
restoration of an adequate protein diet after birth allowsincomplete recovery at 21 days. Pancreatic weight isaffected more severely than body weight as a whole.There is a total reduction in endocrine tissue and betacells per pancreas in the LP and postLP groups, differ-ences present to a much less significant extent in thepreLP group.Rat pancreatic islets are similar in morphology to
human islets, though they do show a greater degree ofzonation between alpha cells, present around the periph-ery or cortex of the islets, and beta cells, which arepresent in the centre or medulla of the islets.11 Pancre-atic islets are derived from the primordial pancreaticbuds11,12 and while early endocrine cell precursorsexpress a variety of hormones, the mature islets arecomposed of a mixture of endocrine cells, the majorityof which produce either glucagon or insulin. Islets mayform clonally from primordial cells derived from ductalstem cells,13 or by the aggregation and replication ofendocrine cells within the pancreatic stroma.14 Pro-genitor endocrine cells appear to be able to develop intoany of the mature endocrine cells.15 There is also evi-dence that islet neogenesis is a default path when bothmesenchyme and basement membrane are absent.16 Theability of the pancreas to regenerate beta cells byneotransformation from progenitor cells diminishesafter birth17,18 and later regeneration is probably by thereplication of existing mature beta cells. Therefore anyfactor affecting pancreatic development in early life willlead to permanent changes in islet morphology.The shape and size of the islets differed significantly
between the four groups. The LP and postLP groupshad significantly larger islets that were less regular inshape than controls. This tendency to irregularity ofform was not corrected by postnatal recuperation,although the preLP group had significantly smaller islets
Fig. 2—Percentage of hormone-secreting cells per islet. Beta-cell area: shaded columns; alpha-cell area:cross-hatched columns. Comparison with control using Student’s t-test. *P<0·05
112 D. M. BERNEY ET AL.
? 1997 by John Wiley & Sons, Ltd. , . 183: 109–115 (1997)
than the other groups. This may be due to stimulants fornew islet formation, such as transforming growth factorâ (TGF â),19 when the protein supply increases, but it isnoteworthy that the mean area of beta cells per islet
remains significantly below controls (Fig. 2). Thus, thestructural changes which occur within the pancreas inprotein deprivation are not reparable and may be due toincreased beta-cell replication, making up for a deficit in
Fig. 3—(a) Mean pancreatic weight. Comparisons with control using Student’s t-test.*P<0·05; **P<0·001. (b) Calculated mean weight of islet tissue per pancreas. Comparisonswith control using Student’s t-test. *P<0·05. (c) Calculated mean weight of beta cells perpancreas. Comparisons with control using Student’s t-test. *P<0·05
(a)
(b)
(c)
113EFFECTS OF LOW PROTEIN ON THE FETAL RAT PANCREAS
? 1997 by John Wiley & Sons, Ltd. , . 183: 109–115 (1997)
earlier islet neogenesis. These changes are comparable tothose in another study which showed the islets in alow-protein group to be smaller, though no estimate of
islet irregularity was made.20 In this study, measure-ments were made in the 21-day-old neonates exposed tomaternal low protein in utero. Our preLP group had a
Fig. 4—(a) Rat body weight. Comparisons with control using Student’s t-test. *P<0·001;**P<0·05. (b) Calculated mean ratio of beta-cell weight to body weight. (c) Calculated mean ratioof alpha-cell weight of body weight. Comparisons with control using Student’s t-test. *P<0·05
(a)
(b)
(c)
114 D. M. BERNEY ET AL.
? 1997 by John Wiley & Sons, Ltd. , . 183: 109–115 (1997)

similar period of exposure and in both cases the islet sizewas smaller.The cellular composition of the islets is also different
in offspring of maternal protein deprivation, with thechanges seen persisting in the preLP group. The differ-ences in beta-cell content and in the beta cell to alphacell ratio within islets are preserved in this group. Thesechanges therefore cannot be entirely rectified by a returnto normal diet, implying that these structural changesare permanent and may lead to a phenotype more at riskof developing NIDDM.The vascularity of the islets has not been measured
successfully in this study, since the fetal blood vessels aredifficult to visualize and we have been unable to identifyreliable markers for fetal rat endothelial cells. Islets ofLangerhans have been shown to have a complex bloodsupply; the afferent blood first encounters the islet betacells, before supplying the alpha cells. It is easy to seethat changes in islet shape, size, and cell distribution willaffect this blood supply pattern and some morphologicalstudies have suggested that this may be the case.20 It ispossible, however, that changes in the blood supply areprimarily responsible for the observed changes in isletmorphology.The percentage of islet tissue per rat is identical in all
three groups. Also, the percentage of beta cell tissue perrat is similar in all groups (Fig. 4b), though the alphacells are increased in the low-protein group (Fig. 4c).Thus, despite the striking morphological changes in thepancreas which occur in the maternal protein-deprivedgroups, there appear to be homeostatic mechanismswhich maintain a constant amount of endocrine tissueper unit of body weight. Weight changes in organs arenot always proportionate to body weight and asymmet-ric growth retardation with sparing of the brain has beenknown for many years in babies small for gestationalage.21 Similarly, the importance of pancreatic islet tissueto the development of the organism is highlighted by itspreservation in protein deficiency, but this preservationseems to be at the expense of fundamental changes inpancreatic structure and cellular composition. Thesechanges are partly preserved after recuperation, furtherhighlighting the importance of early growth to the finalphenotype of the organism. Other studies have shownimpaired insulin secretion in a similar protein-deprivedrat model.5In conclusion, we have found that after altered nutri-
tion during pregnancy, structural abnormalities occur inthe offspring which are still present after recuperation.These changes may lead to consequent alterations in thephysiological function of the islets of Langerhans.
ACKNOWLEDGEMENTS
We thank D. Hutt, A. Flack, A. Wayman, andL. Smith of Dunn Nutritional Laboratory for their
invaluable assistance. This work was supported by theBritish Diabetic Association, the Medical ResearchCouncil, and the Parthenon Trust.
REFERENCES
1. Hales CN, Barker DJP, Clark PMS, et al. Fetal and infant growth andimpaired glucose tolerance at age 64 years. Br Med J 1991; 303: 1019–1022.
2. Barker DJP, Hales CN, Fall CHD, Osmond C, Phipps K, Clark PMS.Type 2 (non-insulin dependent) diabetes mellitus, hypertension and hyper-lipidaemia (syndrome X): relation to reduced fetal growth. Diabetologia1993; 36: 62–67.
3. Barker DJP. Fetal and Infant Origins of Adult Disease. London: BritishMedical Journal Publications, 1992.
4. Hales CN, Barker DJP. Type 2 (non-insulin dependent) diabetes mellitus:the thrifty phenotype hypothesis. Diabetologia 192; 35: 595–601.
5. Dahri S, Snoek A, Reusens-Billen B, Remacle C, Hoet JJ. Islet function inoffspring of mothers on low protein diet during gestation. Diabetes 1991;40(Suppl): 2115–2120.
6. Desai M, Crowther NJ, Ozanne SE, Lucas A, Hales CN. Adult glucose andlipid metabolism may be programmed during fetal life. Biochem Soc Trans1995; 23: 331–335.
7. Perrier BH. Effects of prolonged maternal fast on the pancreas of 18–21 dayold foetal rats. Ultrastructural and morphometric study of the beta cells.Cell Tissue Res 1984; 237: 169–179.
8. James WPT, Coore HG. Persistent impairment of insulin secretion andglucose tolerance after malnutrition. Am J Clin Nutr 1970; 23: 386–389.
9. Milner RD. Metabolic and hormonal responses to oral amino acids ininfantile malnutrition. Arch Dis Child 1971; 46: 301–305.
10. Desai M, Crowther NJ, Lucas A, Hales CN. Organ-selective growth in theoffspring of protein-restricted mothers. Br J Nutr 1996; 76: 591–603.
11. Slack JMW. Developmental biology of the pancreas. Development 1995;121: 1569–1580.
12. Andrew A. An experimental investigation into the possible neural crestorigin of pancreatic APUD (islet) cells. J Embryol Exp Morph 1976; 35:577–593.
13. Bouwens L, Kloppel G. Islet cell neogenesis in the pancreas. Virchows Arch1996; 427: 553–560.
14. Deltour L, Leduque P, Paldi A, Ripoche M, Dubois P, Jami J. Polyclonalorigin of pancreatic islets in aggregation mouse chimaeras. Development1991; 112: 1115–1121.
15. Teitelman G, Alpert S, Polak JM, Martinez A, Hanahan D. Mouseendocrine pancreases coexpress insulin, glucagon and the neuronal proteinstyrosine hydroxylase and neuropeptide Y, but not pancreatic polypeptide.Development 1993; 118: 1031–1039.
16. Gittes GK, Galante PE, Hanahan D, Rutter WJ, Debas HT. Lineage-specific morphogenesis in the developing pancreas: role of mesenchymalfactors. Development 1996; 122: 439–447.
17. Kaung HL. Growth dynamics of pancreatic islet cell populations duringfetal and neonatal development of the rat. Dev Dyn 1994; 200: 163–175.
18. Wang RN, Bouwens L, Kloppel G. Beta-cell growth in adolescent and adultrats treated with streptozocin during the neonatal period. Diabetologia 1996;39: 548–557.
19. Sanvito F, Herrera PL, Huarte J, et al. TGF-beta 1 influences the relativedevelopment of the exocrine and endocrine pancreas in vitro. Development1994; 120: 3451–3462.
20. Snoeck A, Remacle C, Reusens B, Hoet JJ. Effect of a low protein dietduring pregnancy on the fetal rat endocrine pancreas. Biol Neonate 1990; 57:107–118.
21. Ogata ES, Bussey ME, LaBarbera A, Finley S. Altered growth, hypo-glycaemia, hypoalaninemia and ketonemia in the young rat: postnatalconsequences of intrauterine growth retardation. Pediatr Res 1985; 19:32–37.
115EFFECTS OF LOW PROTEIN ON THE FETAL RAT PANCREAS
? 1997 by John Wiley & Sons, Ltd. , . 183: 109–115 (1997)