Embed Size (px)
Citation preview

Substance P is a major representative of the family ofneurokinin peptides in the mammalian nervous system.Involvement of substance P and other neurokinins in themechanisms of motivational behavior has been demonstrat-ed in models of self-administration [3], especially of alco-hol. Substance P decreases alcohol consumption after bothcentral and peripheral administration [1, 2]. It has been sug-gested that these properties result from the close interactionwith the mesocorticolimbic system of the brain, whose
dopaminergic neurons carry substance P receptors [10] or thepossession of intrinsic reinforcing activity by substance P.Substance P and neurokinin receptors are widely distribut-ed in a number of brain structures directly involved in emo-tional responses, such as the amygdala, hippocampus, andanterior cingulate gyrus [12, 18].
However, studies of substance P in a conditioned placepreference test showed that its reinforcing propertiesdepend on the route of administration of the peptide. Thereinforcing action was seen only after peripheral adminis-tration, while central administration either did not producethis effect or produced aversive influences [20]. Thus, theneurochemical mechanisms of action of substance P aftercentral administration require further study.
Apart from the fact itself of changes in the activity ofone or another neurochemical system, the dynamics ofchanges in neurotransmitter levels are also important,
Neuroscience and Behavioral Physiology, Vol. 34, No. 7, 2004
The Effects of Substance P After Central Administration on the Activity of the Mesolimbic System of the Rat Brain as Studied by Microdialysis
S. V. Nikolaev,1 A. A. Lebedev,2 E. R. Bychkov,1
A. V. Oblyapin,1 S. A. Dambinova,1 and P. D. Shabanov3
0097-0549/04/3407-0743©2004 Plenum Publishing Corporation
743
Translated from Rossiiskii Fiziologicheskii Zhurnal imeni I. M. Sechenova, Vol. 89, No. 2, pp. 123–128,February, 2003. Original article submitted October 24, 2002.
In vivo microdialysis was used to study the effects of substance P on dopamine, dihydroxyphenylaceticacid, and homovanillic acid levels in the nucleus accumbens in rats. Each animal received sequential injec-tions of physiological saline, 0.1 µg of substance P, and 1 µg of substance P into the lateral ventricle overthree days. Dialysates showed increases in dopamine levels in response to neuropeptide, by 41% for the0.1 µg dose and 71% for the 1 µg dose. The dynamics of these changes also depended on the concentra-tion of the agent. Administration of 1 µg of substance P gave a peak dopamine level at 50 min; the neu-rotransmitter level remained significantly elevated 75 min after dosage with substance P. The dopaminelevel was increased only at 75 min when the 0.1 µg dose of neuropeptide was used. Changes in metabo-lite levels were also dose-dependent. After the 1 µg dose, the dihydroxyphenylacetic acid level increasedby 28%, while the 0.1 µg dose produced no significant change in the level of this metabolite. Thehomovanillic acid level did not respond to administration of substance P at either dose. These data sup-port the suggestion that the influence of substance P on the internal compensation system is to a signifi-cant extent mediated by dopaminergic mechanisms and provides a possible explanation for the effects ofthe neuropeptide seen in a conditioned place preference reflex.
Key words: substance P, microdialysis, dopamine, nucleus accumbens, metabolism.
1 Institute of the Human Brain, Russian Academy of Sciences,12a Academician Pavlov Street, 197376 St. Petersburg, Russia.
2 Institute of Experimental Medicine, Russian Academy ofMedical Sciences, 12 Academician Pavlov Street, 197376St. Petersburg, Russia.
3 Military Medical Academy, 6 Lebedev Street, 194175St. Petersburg, Russia.

though it should be noted that traditional methods can onlybe used to obtain overall data on neurotransmitter contentsin tissues,without consideration of synaptic release. Theactivity of a system in this sense needs to take account ofthe levels of metabolites of a compound of interest. Micro-dialysis allows the activity of a neurochemical system to bestudied directly, by measuring the levels of the extracellularneurotransmitter pool and following changes in the activityof the system over time in a single animal.
The method is based on the ability of biologicalmolecules to penetrate semipermeable membranes locatedin the brain structure of interest,crossing a concentrationgradient. A continuous flow of solvent moving on the otherside of the membrane becomes enriched with componentsof the extracellular medium and can be collected in volumessuitable for analytical techniques. The concentration of thestudy compound in each fraction reflects its relative level inthe brain structure during a defined period of time.
Published data on microdialysis and relating to theeffects of substance P on the dopaminergic system havebeen obtained mainly in conditions of peripheral orintrastructural administration of the neuropeptide. In theformer case, indirect effects can occur, as substance P hasmarked peripheral actions [7]; in the latter case, we do notyet have a complete picture of the central effects. In addi-tion, some of the data were obtained in acute experiments,when the effects of anesthesia might influence the studyresults [4],and massive release of neurotransmitters fromdamaged nervous tissue masks the physiological variationsin their levels. The aim of the present work was to assess theactivity of the dopaminergic mesolimbic system of the brainusing microdialysis during intraventricular administrationof substance P in animals in conditions of free behavior.
METHODS
Studies were performed using six male Wistar rats.Cannulae were implanted into the right ventricle (AP = –0.9,L = 1.4, H = 3.5) of all rats under Nembutal anesthesia.Microdialysis cannulae for analyzing the extracellular fluidwere simultaneously implanted into the nucleus accumbens(AP = 2.0,L = 1.5,H = 7.0). After attaching the body of thecannula to the animal’s head with acryloxide, animals wereplaced in individual cages.
Microdialysis cannulae were concentric, a designcausing less damage to brain tissue than others [21].Cannulae were made of hollow dialysis membrane (Hospal,France) with an external diameter of 300 µm and a molec-ular threshold of 20 kDal,membranes being fixed into steeltubes (diameter 500 µm) with epoxide glue such that thelength of the free part of the membrane was 2.5 mm. Apolythene tube of length 1.2 m was attached to the endopposite the membrane, for collecting the dialysate. Anopening was made in the wall of the polythene tube 5 mm
from the junction; a flexible capillary with an externaldiameter of 150 µm was inserted through this opening todeliver fluids. The capillary was passed along the steel tubeto the membrane such that the distance between the capil-lary and the edge of the membrane was no greater than1.5mm. All components were attached with epoxide glue.
Microdialysis studies were performed 48 h after surgery.Given that experiments were performed in conditions offree behavior, the pharmacological effects could interferewith the physiological changes in the activity of thedopaminergic system,particularly reactions to novelty [6].Animals therefore remained in the experimental chamberthroughout the period leading up to microdialysis to ensurefamiliarity with the apparatus. During the experiment, thedelivery cannula was attached to a 1-ml syringe filled withmodified Ringer’s solution containing 146 mM NaCl,4 mM KCl, and 2.2 mM CaCl2. A precision pump was usedto deliver fluid into the microdialysis chamber at a constantvolume rate of 2.5 µl/min. The collecting cannula wasinserted into a 500-µl microtube containing 5 µl ofhydrochloric acid as preservative. Dialysate was collectedfor 25 min,after which the tube containing dialysate wasreplaced and immediately frozen in liquid nitrogen.Neurokinin was given into the lateral ventricle 5 min beforethe end of the period for collection of the fourth dialysatesample. Over a period of three days, each animal receivedsequentially: physiological saline on day 1, 0.1 µg of sub-stance P dissolved in 5 µl of physiological saline on day 2,and 1 µg of substance P on day 3. Doses were given usinga microdoser over 1.5 min. A further four fractions werecollected after dosage. Samples were kept at –70°C prior toanalysis. Dopamine, dihydroxyphenylacetic acid (DOPAA),and homovanillic acid (HVA) concentrations in dialysatesamples were measured by HPLC with electrochemicaldetection. Dialysate (40 µl) from each sample was injectedinto the chromatograph without prior purif ication.
The positions of the microdialysis tubes in brain struc-tures were confirmed morphologically. After experiments,animals were sacrif iced, cannulae were removed carefully,and brains were harvested and placed in 10% formalin.Brains were then frozen and a microtome was used to cutseries of frontal sections every 250 µm.
Data were analyzed by two-factor dispersion analysis(ANOVA) with time and pharmacological treatment as thefactors. Basal levels (taken as 100%) were mean neuro-transmitter and metabolite concentrations in the three sam-ples collected immediately before dosage.
RESULTS
The basal dopamine level in the nucleus accumbenswas 4.56 ± 0.35 pg; that of DOPAA was 236 ± 17.9 pg, andthat of HVA was 203 ± 12.4 pg per 10 µl of sample. The rel-ative contents of dopamine and its metabolites at different
Nikolaev, Lebedev, Bychkov, et al.744
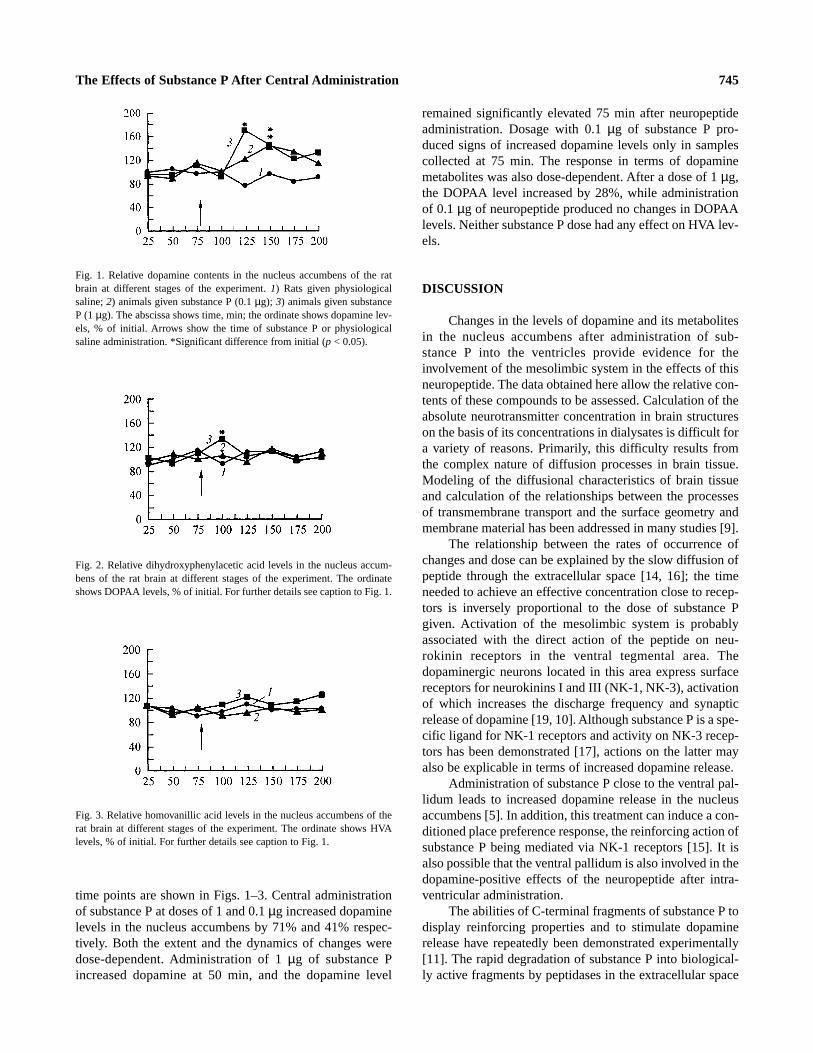
time points are shown in Figs. 1–3. Central administrationof substance P at doses of 1 and 0.1 µg increased dopaminelevels in the nucleus accumbens by 71% and 41% respec-tively. Both the extent and the dynamics of changes weredose-dependent. Administration of 1 µg of substance Pincreased dopamine at 50 min, and the dopamine level
remained significantly elevated 75 min after neuropeptideadministration. Dosage with 0.1 µg of substance P pro-duced signs of increased dopamine levels only in samplescollected at 75 min. The response in terms of dopaminemetabolites was also dose-dependent. After a dose of 1 µg,the DOPAA level increased by 28%,while administrationof 0.1 µg of neuropeptide produced no changes in DOPAAlevels. Neither substance P dose had any effect on HVA lev-els.
DISCUSSION
Changes in the levels of dopamine and its metabolitesin the nucleus accumbens after administration of sub-stanceP into the ventricles provide evidence for theinvolvement of the mesolimbic system in the effects of thisneuropeptide. The data obtained here allow the relative con-tents of these compounds to be assessed. Calculation of theabsolute neurotransmitter concentration in brain structureson the basis of its concentrations in dialysates is difficult fora variety of reasons. Primarily, this difficulty results fromthe complex nature of diffusion processes in brain tissue.Modeling of the diffusional characteristics of brain tissueand calculation of the relationships between the processesof transmembrane transport and the surface geometry andmembrane material has been addressed in many studies [9].
The relationship between the rates of occurrence ofchanges and dose can be explained by the slow diffusion ofpeptide through the extracellular space [14,16]; the timeneeded to achieve an effective concentration close to recep-tors is inversely proportional to the dose of substance Pgiven. Activation of the mesolimbic system is probablyassociated with the direct action of the peptide on neu-rokinin receptors in the ventral tegmental area. Thedopaminergic neurons located in this area express surfacereceptors for neurokinins I and III (NK-1,NK-3), activationof which increases the discharge frequency and synapticrelease of dopamine [19,10]. Although substance P is a spe-cific ligand for NK-1 receptors and activity on NK-3 recep-tors has been demonstrated [17],actions on the latter mayalso be explicable in terms of increased dopamine release.
Administration of substance P close to the ventral pal-lidum leads to increased dopamine release in the nucleusaccumbens [5]. In addition, this treatment can induce a con-ditioned place preference response, the reinforcing action ofsubstance P being mediated via NK-1 receptors [15]. It isalso possible that the ventral pallidum is also involved in thedopamine-positive effects of the neuropeptide after intra-ventricular administration.
The abilities of C-terminal fragments of substance P todisplay reinforcing properties and to stimulate dopaminerelease have repeatedly been demonstrated experimentally[11]. The rapid degradation of substance P into biological-ly active fragments by peptidases in the extracellular space
The Effects of Substance P After Centr al Administr ation 745
Fig. 1. Relative dopamine contents in the nucleus accumbens of the ratbrain at different stages of the experiment. 1) Rats given physiologicalsaline; 2) animals given substance P (0.1 µg); 3) animals given substanceP (1 µg). The abscissa shows time, min; the ordinate shows dopamine lev-els, % of initial. Arrows show the time of substance P or physiologicalsaline administration. *Significant difference from initial (p < 0.05).
Fig. 2. Relative dihydroxyphenylacetic acid levels in the nucleus accum-bens of the rat brain at different stages of the experiment. The ordinateshows DOPAA levels,% of initial. For further details see caption to Fig. 1.
Fig. 3. Relative homovanillic acid levels in the nucleus accumbens of therat brain at different stages of the experiment. The ordinate shows HVAlevels,% of initial. For further details see caption to Fig. 1.

[13] suggests that such fragments are of no small impor-tance for the changes seen in the levels of dopaminergictransmission.
Negative results observed in the conditioned placepreference test after central administration of substance Pcan be explained in terms of the dynamics of activation ofthe dopaminergic system after this means of neuropeptidetreatment. Substance P at a dose of 0.1 µg increased meandopamine levels only after 75 min which, considering themethod generally used in testing the conditioned place pref-erence response, may be critical for the formation of theconditioned reflex link between the reinforcing propertiesof the compound administered and the contextual stimuli.
On the other hand, realization of the reinforcing prop-erties of this neurokinin may not be restricted to dopamin-ergic mechanisms. The opiatergic system of the brain isinvolved in some of the effects of neurokinins; in particular,blockade of opiate receptors inhibits the development of theconditioned place preference reaction in response to admin-istration of substance P [8].
Thus, administration of substance P into the lateralventricle of the brain was followed by increases in the lev-els of dopamine and its metabolite DOPAA in the nucleusaccumbens. The onset time and extent of changes dependedon the neuropeptide dose given.
REFERENCES
1. E. R. Bychkov, A. A. Lebedev, and S. A. Dambinova, “Effects ofsubstance P on ethanol consumption in subchronically alcoholizedrats in a test based on restricted access to alcohol,” Zh. Vyssh. Nerv.Deyat., 51, No. 1,120–122 (2001).
2. E. R. Bychkov, S. V. Nikolaev, A. A. Lebedev, and S. A. Dambinova,“Effects of peripheral administration of substance P on alcohol con-sumption and the activity of the dopaminergic systems of the brain,”Psikhofarmakol. Biol. Narkol., 1, 43–46 (2001).
3. S. K. Sudakov, I. V. Figurina,O. F. Medvedeva,and I. V. Rusakova,“Effects of substance P on the intravenous morphine self-adminis-tration reaction in two rat lines,” Byull. Éksperim. Biol. Med., 131,No. 10,364–367 (2001).
4. Y. U. Adachi, K. Watanabe, T. Satoh, and E. S. Vizi, “Halothanepotentiated the effect of methamphetamine and nomifensine onextracellular dopamine levels in rat striatum: a microdialysis study,”Brit. J. Anaesth., 86, No. 6,837–845 (2001).
5. F. Boix, P. Sandor J. C. Nogueira,J. P. Huston,and R. K. Schwarting,“Relationship between dopamine release in nucleus accumbens and
place preference induced by substance P injected into the nucleusbasalis magnocellularis region,” Neurosci., 64, No. 4, 1045–1055(1995).
6. M. G. Geenstra and M. H. Botterblom, “Rapid sampling of extracel-lular dopamine in the rat prefrontal cortex during food consumption,handling and exposure to novelty,” Brain Res., 742, No. 1–2,17–24(1996).
7. S. Harrison and P. Geppetti, “Substance P,” Int. J. Biochem. CellBiol., 33, 555–576 (2001).
8. R. U. Hasenöhri, P. Gerhardt, and J. P. Huston,“Naloxone blocksconditioned place preference induced by substance P and [pGlu6]-SP(6-11),” Regul. Pept., 35, 177–187 (1991).
9. J. K. Hsiao,B. A. Ball, P. F. Morrison,I. N. Mefford, and P. W. Bungay,“Effects of different semipermeable membranes on in vitro perfor-mance of microdialysis probes,” J. Neurochem., 54, 1449–1452(1990).
10. P. W. Kalivas,“Neurotransmitter regulation of dopamine neurons inventral tegmental area,” Brain Res. Rev., 18, 75–113 (1993).
11. S. Khan, N. Brooks, R. Whelpton, and A. T. Michael-Titus,“Substance P (1–7) and substance P (5–11) locally modulatedopamine release in rat striatum,” Eur. J. Pharmacol., 282, 229–233(1995).
12. H. Kuiama, H. Maeno,and M. Tohyama, “Substance P receptor(NK-1) in the central nervous system:possible functions from amorphological aspect,” Regul. Pept., 46, 114–123 (1993).
13. A. T. Michael-Titus, K. Fernandes,H. Setty, and R. Whelpton,“Invivo metabolism and clearance of substance P and co-expressedtachykinin in rat striatum,” Neurosci., 110, No. 2,277–286 (2002).
14. C. Nicholson and E. Sykova, “Extracellular space structure revealedby diffusion analysis,” Trends Neurosci., 21, 207–215 (1998).
15. S. Nikolaus,J. P. Huston,and R. U. Hasenöhri, “Reinforcing effectson neurokinin substance P in the ventral pallidum:mediation by thetachykinin NK-1 receptor,” Eur. J. Pharmacol., 370, 93–99 (1999).
16. W. M. Partridge, “Drug delivery to the brain,” J. Cereb. Blood FlowMetab., 17, 713–731 (1997).
17. D. Regoli, A. Boudon,and J. L. Fauchere, “Receptors and antago-nists for substance P and related peptides,” Pharmacol. Rev., 46,551–599 (1994).
18. A. Ribeiro-da-Silva and T. Hokfelt, “Neuroanatomical localizationof substance P in the CNS and sensory neurons,” Neuropeptides, 34,No. 5,256–271 (2000).
19. G. R. Seabrook, B. J. Bowery, and R. G. Hill, “Pharmacology oftachykinin receptors on neurons in the ventral tegmental area of ratbrain slices,” Eur. J. Pharmacol., 273, 113–119 (1995).
20. T. M. Tzschentke, “Measuring reward with the conditioned placepreference paradigm:a comprehensive review of drug effects,recentprogress and new issues,” Prog. Neurobiol., 56, No. 6, 613–672(1998).
21. B. H. C. Westerink and J. B. de Vries, “Characterization of in vivodopamine release as determined by brain microdialysis after acuteand subchronic implantations: methodological aspects,”J. Neurosci., 51, 683–687 (1988).
Nikolaev, Lebedev, Bychkov, et al.746






![Improvement of the Mechanical Stability of Microdialysis Catheters · 2016-02-09 · The next step in the development of microdialysis was when Urban Ungerstedt [3] introduced a hollow](https://img.pdfslide.net/doc/110x75/5f2bf162a4336743450d5a4a/improvement-of-the-mechanical-stability-of-microdialysis-2016-02-09-the-next-step.jpg)






















