Embed Size (px)
Citation preview

Volume 20(2), 118- 125, 2016 JOURNAL of Horticulture, Forestry and Biotechnology www.journal-hfb.usab-tm.ro
118
The Effects of Taxus Baccata L. Aqueous Extracts on Germination, Seedling Growth and Physiological Parameters of Test Species Lobiuc A. 1,2, Cuibari Roxana 1, Frunzete Mădălina 1,*, Naela C.1, Marian Burducea1, Ardelean Mirela 3, Zamfirache Maria-Magdalena 1
1Departament of Biology, Faculty of Biology, “Alexandru Ioan Cuza” University of Iași, Bd. Carol I, No. 20 A,
code 700505, Iași, Romania; 2 “Universitatea Ștefan cel Mare”, Universitatii str. 13, Suceava, România
3“Vasile Goldiș” University, Revolutiei str. 94, Arad, Romania;
*Corresponding author. Email: [email protected];
Abstract The present paper aimed to assess the phytotoxic potential of Taxus baccata L. (yew) aqueous extracts on Raphanus sativus L. and Cucumis sativus L. germination, growth, chlorophyll content and chlorophyll fluorescence (ɸPSII) of the seedling.
In the case of Raphanus sativus the germination indices were negatively affected by the leaves and aril extracts, with the 5% leaves extract having the most pronounced effect on the germination percentage, energy and speed of germination and vitality index. Leaves and aril extracts significantly reduced the fresh mass (with up to 41%) and radicle length (with up to 71%) of the seedlings, while the length of the hypocotyl was reduced especially by the bark extracts. Inhibitory effects on chlorophyll contents were exerted by 5% aril and 1% leaves extracts while chlorophyll fluorescence (ϕPSII) was decreased, however not significantly, by leaves extracts.
In the case of Cucumis sativus, inhibitory effects on germination were recorded for leaves 1% and aril 5% extracts. Morphometric inhibition was especially observed for 5% aril extract, which reduced the mass, hypocotyl and radicle lengths and also chlorophyll content and fluorescence parameters. The registered effects can be associated with the high contents of phenolic in the extracts as determined by UV-Vis spectrometry and also to physico-chemical characteristics of extracts. Taxus baccata is a well-known species for the toxic effect of its constituent, taxol, on animal cells. The tested extracts displayed a certain degree of toxicity also on plants which may add to the uses of this specie. Further research may include optimization of the extraction process and more thorough identification of compounds present in extracts.
Key words yew, phytotoxicity, morphometry, chlorophyll, fluorescence
Taxus baccata L. (yew) is a glaciary relict coniferous
specie, belonging to the Taxaceae family. It is a specie
with a long standing use by humans, especially for its
wood, from which furniture is produced but also for the
dyeing properties of the bark [27]. Some extracts form
Taxus are used in cosmetics products, as well as in
traditional medicine, for treating headache, cold or
nervous system disordes [15]. More recently Taxus
baccata is used for extraction of the anti-proliferative
compound taxol, an alkaloid with potent activity [22].
These are the main reasons for over exploitation of this
specie in many countries [8]. More over the specie is
considered endangered in many countries being
included in several red lists, including in Romania [25,
26], while habitats with yew are listed in the Habitats
Annex II [3].
The main factors affecting the Taxus
population establishment and trends are considered the
available amount of light in relation with other species’
competition, deforestation, browsing and bark stripping
by herbivores [8]. The requirements of yew towards
abiotic ecological factors are relatively well know, the
specie growing well in higher humidity, on calcareous
soils, altitude between 100-400 m in regions with mean
temperature between 8-21° C [41,27].

119
However, significantly less data are available
concerning the ecology of Taxus in interaction with
individuals of other species. Also, it considered that
Taxus seedling are not competitive in establishing new
populations, on one hand due to grazing and on the
other hand, due to inhibition of growth induced by
shade [31,3]. The possibility of self-inhibition of
seedling growth by other Taxus individuals was,
however, not studied previously, while it is known that
this phenomenon occurs in several forest species [21].
Autotoxicity is reported to occur in both manipulated
and natural ecosystems, in both herbaceous and tree
species [18]. Toxic effects of leachates or exudates
involve the presence of chemical substances, such as
phenolics [35,46], which affect surrounding plants by
interference with various metabolic pathways [4].
Few studies have documented the allelopathic
effects of Taxus on other species, showing a certain
degree of toxicity towards other [47]. Moreover,
various Taxus tissues contain relatively high amounts
of phenolic substances [6,7], and alkaloids [28], which
may have phytotoxic activity [2,30,33]. Therefore, the
current study aims to test whether aqueous extracts of
Taxus organs may induce growth inhibition, by
subjecting seeds and individuals of fast-growing test
species to extracts treatments and assessing
morphological and physiological growth parameters.
Materials and Methods
Germination and growth conditions
The effects of yew extracts on germination
and initial growth were assessed using two test species,
frequently used for their fast germination and growth
and sensitivity towards external factors, Raphanus
sativus L. and Cucumis sativus L.
Petri dishes, sterilized, lined with sterile filter
paper were used for placing 16 radish and 10 cucumber
seeds per plate, subsequently wetting the paper with
distilled water for control plates or aqueous extracts
obtained from different yew organs. Plates were treated
initially with 3 ml of solution, then the filter paper was
kept moist when needed. The plates were placed in a
controlled environment growth chamber (Snijders
Scientific), with a 14:10 h photoperiod, 22-26°C
temperature and 70% relative humidity. Three plates
were used for each treatment variant, placed in a
random fashion in the growth chamber. The total
duration of the experiment was 120 hours.
Extract preparation
Aqueous extracts were prepared from arils,
leaves and bark of yew. From each organ, 1 g or 5 g of
tissue was grinded and suspended in 99 or 95 ml of
distilled water, in order to obtain 1% and 5% extracts
from each organ. The mixtures were placed on a heated
water bath and kept 150 minutes at 50° C to facilitate
compound extraction. Subsequently, the mixtures were
filtered and stored in refrigerated conditions before use.
Measurements of germination
For assessing the influence of extracts on the
germination process, four germination-related indices
were determined, namely the germination percentage
(GP), the speed of germination (SG), the energy of
germination (EG) and the vitality index (VI). These
two indices are among the most widely used for
assessing germination performance [5,23] and
calculated as follows.
GP =
EG =
VI =
Morphometric assessments
Effects of external factors on plant growth are
frequently evaluated by measurements of plant organs,
as growth stimulation or inhibition directly are directly
reflected in the length of these organs. Therefore, the
length of the hypocotyl, of the radicle and the mass of
fresh plants were measured after 120 hours from the
beginning of the experiment. A number of 8 radish
plantlets or 5 cucumber plantlets per dish were
measured, or the maximum available number of
plantlets per dish.
Evaluation of chlorophyll content
The content of assimilatory pigments in leaves
was estimated using a portable chlorophyll meter,
CCM-200 plus, Opti-Sciences Inc., UK. The device
measures the chlorophyll content by relating the
amount of light transmitted through leaf tissue to the
amount of pigments. A number of 8 radish plants or 5
120
cucumber plants or the maximum available number of
plants per dish were measured.
Chlorophyll fluorescence measurements
The influence of the extracts on the yield of
the photosynthetic apparatus were evaluated by
measuring the PSII quantum yield parameter using a
portable chlorophyll fluorescence meter, FMS2,
HansaTech Ltd., UK. Measurements were done on 5
leaves per dish per test specie, and the Fm’, Fs and
ɸPSII indices were recorded.
Total phenolic contents of extracts
The content of phenolic compounds in Taxus
extracts was evaluated using the spectrophotometric
method described in [10]. Briefly, ethanolic (50%)
extracts were prepared, at a concentration of 5 g fresh
material (leaves, bark and aril) per 100 ml ethanol
solution. An aliquot of 0.1 ml of each extracts was
incubated with 0.1 ml of Folin reagent and 1 ml of
distilled water. After 5 minutes 0.8 ml of 7.5%
Na2CO3 solution was added and incubated for further
90 min. The absorbances of the mixture were red at
760 nm using an UVmini-1240 Shimadzu
spectrophotometer. Results were expressed as gallic
acid equivalent per gram of fresh material, calculating
the means and standard errors of three replications per
each type of extract.
Results and Discussion
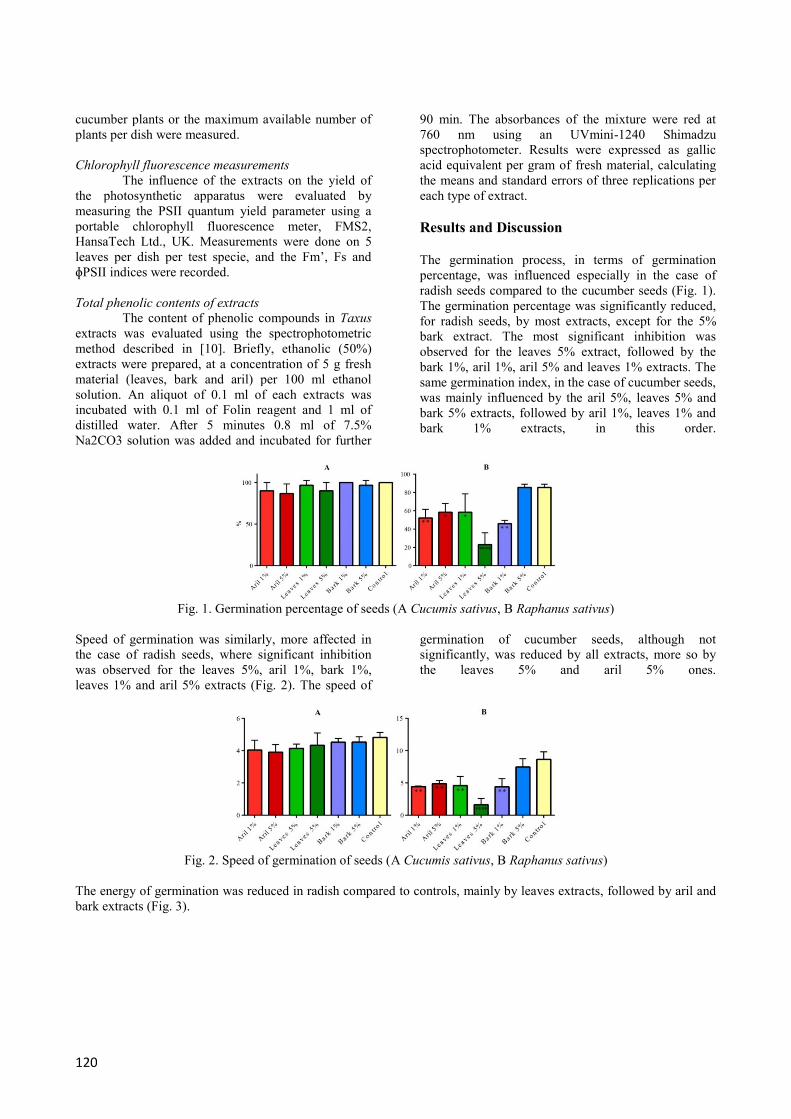
The germination process, in terms of germination
percentage, was influenced especially in the case of
radish seeds compared to the cucumber seeds (Fig. 1).
The germination percentage was significantly reduced,
for radish seeds, by most extracts, except for the 5%
bark extract. The most significant inhibition was
observed for the leaves 5% extract, followed by the
bark 1%, aril 1%, aril 5% and leaves 1% extracts. The
same germination index, in the case of cucumber seeds,
was mainly influenced by the aril 5%, leaves 5% and
bark 5% extracts, followed by aril 1%, leaves 1% and
bark 1% extracts, in this order.
Fig. 1. Germination percentage of seeds (A Cucumis sativus, B Raphanus sativus)
Speed of germination was similarly, more affected in
the case of radish seeds, where significant inhibition
was observed for the leaves 5%, aril 1%, bark 1%,
leaves 1% and aril 5% extracts (Fig. 2). The speed of
germination of cucumber seeds, although not
significantly, was reduced by all extracts, more so by
the leaves 5% and aril 5% ones.
Fig. 2. Speed of germination of seeds (A Cucumis sativus, B Raphanus sativus)
The energy of germination was reduced in radish compared to controls, mainly by leaves extracts, followed by aril and
bark extracts (Fig. 3).
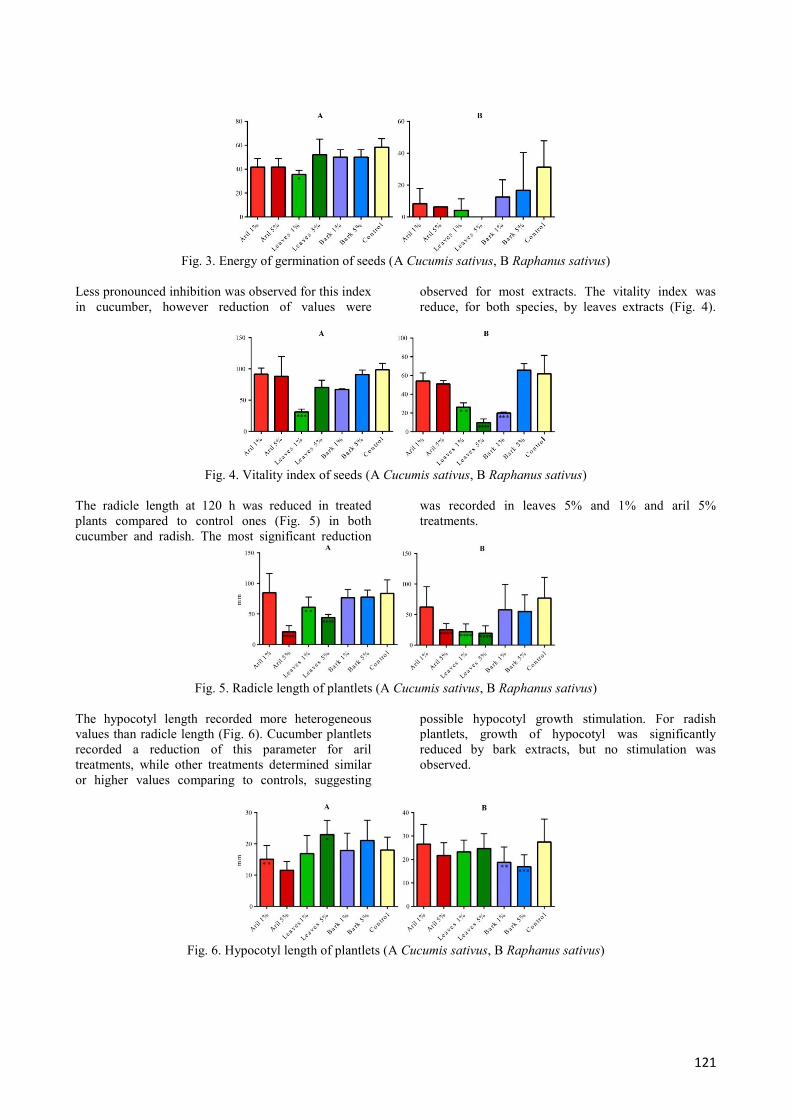
121
Fig. 3. Energy of germination of seeds (A Cucumis sativus, B Raphanus sativus)
Less pronounced inhibition was observed for this index
in cucumber, however reduction of values were
observed for most extracts. The vitality index was
reduce, for both species, by leaves extracts (Fig. 4).
Fig. 4. Vitality index of seeds (A Cucumis sativus, B Raphanus sativus)
The radicle length at 120 h was reduced in treated
plants compared to control ones (Fig. 5) in both
cucumber and radish. The most significant reduction
was recorded in leaves 5% and 1% and aril 5%
treatments.
Fig. 5. Radicle length of plantlets (A Cucumis sativus, B Raphanus sativus)
The hypocotyl length recorded more heterogeneous
values than radicle length (Fig. 6). Cucumber plantlets
recorded a reduction of this parameter for aril
treatments, while other treatments determined similar
or higher values comparing to controls, suggesting
possible hypocotyl growth stimulation. For radish
plantlets, growth of hypocotyl was significantly
reduced by bark extracts, but no stimulation was
observed.
Fig. 6. Hypocotyl length of plantlets (A Cucumis sativus, B Raphanus sativus)
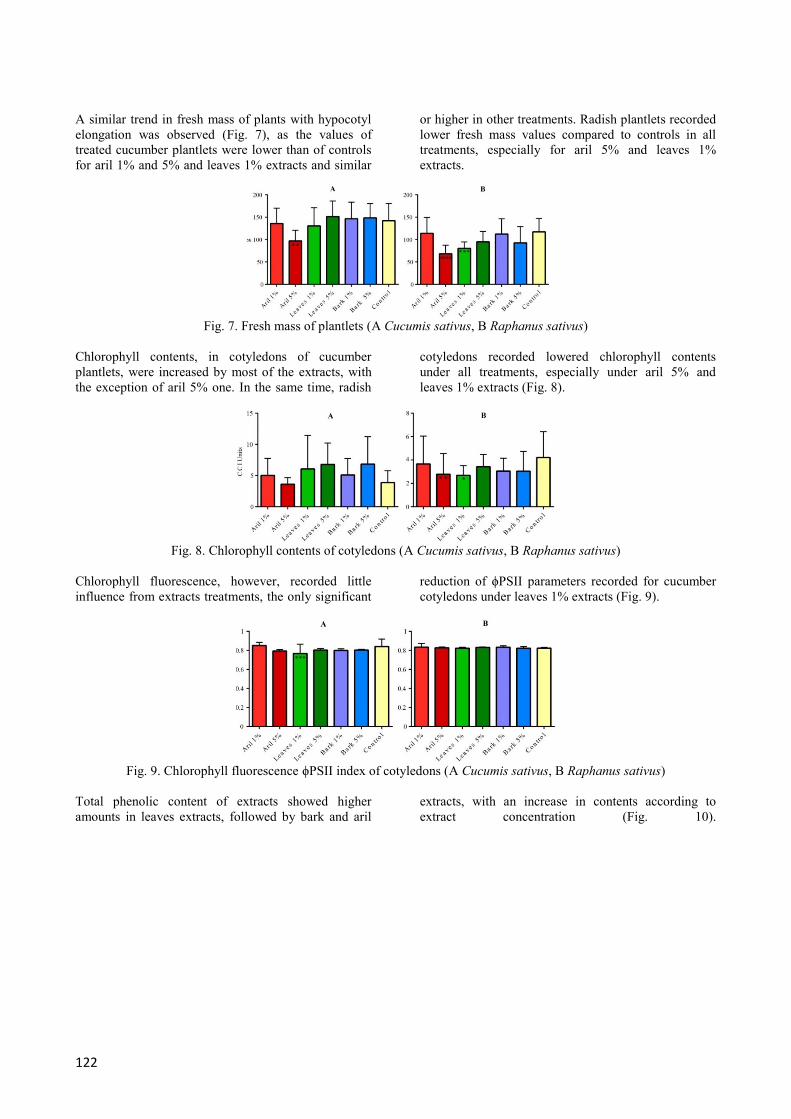
122
A similar trend in fresh mass of plants with hypocotyl
elongation was observed (Fig. 7), as the values of
treated cucumber plantlets were lower than of controls
for aril 1% and 5% and leaves 1% extracts and similar
or higher in other treatments. Radish plantlets recorded
lower fresh mass values compared to controls in all
treatments, especially for aril 5% and leaves 1%
extracts.
Fig. 7. Fresh mass of plantlets (A Cucumis sativus, B Raphanus sativus)
Chlorophyll contents, in cotyledons of cucumber
plantlets, were increased by most of the extracts, with
the exception of aril 5% one. In the same time, radish
cotyledons recorded lowered chlorophyll contents
under all treatments, especially under aril 5% and
leaves 1% extracts (Fig. 8).
Fig. 8. Chlorophyll contents of cotyledons (A Cucumis sativus, B Raphanus sativus)
Chlorophyll fluorescence, however, recorded little
influence from extracts treatments, the only significant
reduction of ϕPSII parameters recorded for cucumber
cotyledons under leaves 1% extracts (Fig. 9).
Fig. 9. Chlorophyll fluorescence ϕPSII index of cotyledons (A Cucumis sativus, B Raphanus sativus)
Total phenolic content of extracts showed higher
amounts in leaves extracts, followed by bark and aril
extracts, with an increase in contents according to
extract concentration (Fig. 10).
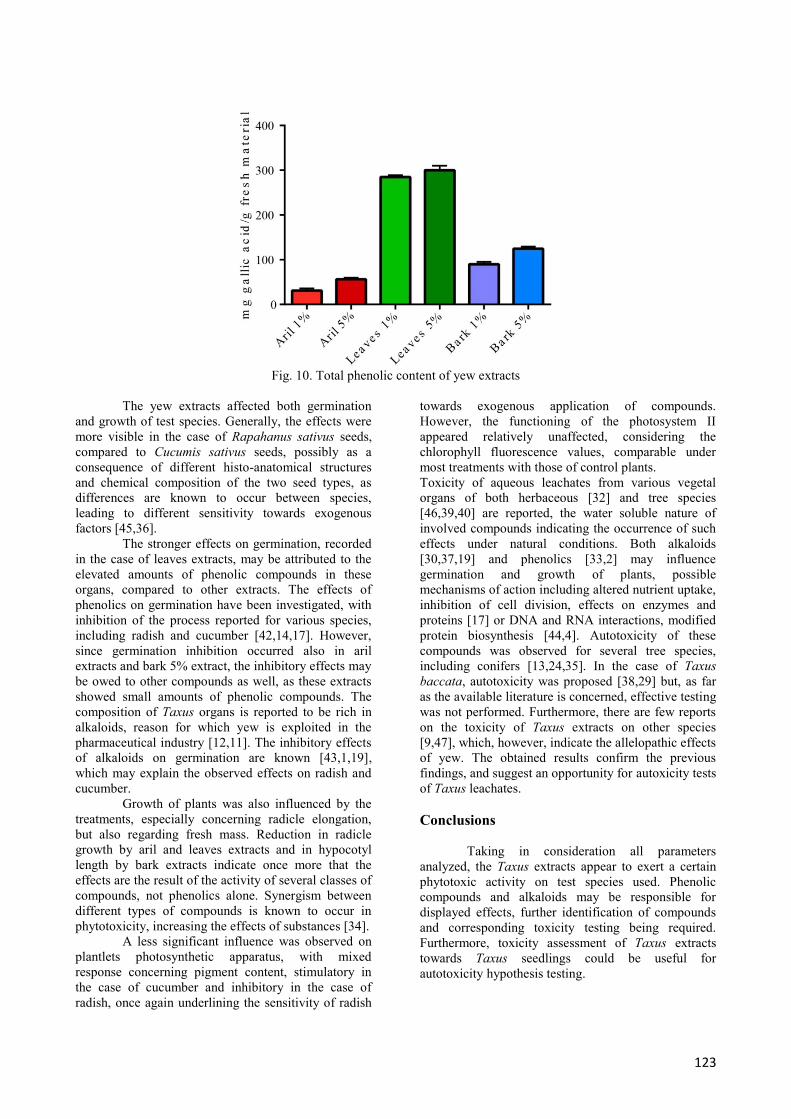
123
Fig. 10. Total phenolic content of yew extracts
The yew extracts affected both germination
and growth of test species. Generally, the effects were
more visible in the case of Rapahanus sativus seeds,
compared to Cucumis sativus seeds, possibly as a
consequence of different histo-anatomical structures
and chemical composition of the two seed types, as
differences are known to occur between species,
leading to different sensitivity towards exogenous
factors [45,36].
The stronger effects on germination, recorded
in the case of leaves extracts, may be attributed to the
elevated amounts of phenolic compounds in these
organs, compared to other extracts. The effects of
phenolics on germination have been investigated, with
inhibition of the process reported for various species,
including radish and cucumber [42,14,17]. However,
since germination inhibition occurred also in aril
extracts and bark 5% extract, the inhibitory effects may
be owed to other compounds as well, as these extracts
showed small amounts of phenolic compounds. The
composition of Taxus organs is reported to be rich in
alkaloids, reason for which yew is exploited in the
pharmaceutical industry [12,11]. The inhibitory effects
of alkaloids on germination are known [43,1,19],
which may explain the observed effects on radish and
cucumber.
Growth of plants was also influenced by the
treatments, especially concerning radicle elongation,
but also regarding fresh mass. Reduction in radicle
growth by aril and leaves extracts and in hypocotyl
length by bark extracts indicate once more that the
effects are the result of the activity of several classes of
compounds, not phenolics alone. Synergism between
different types of compounds is known to occur in
phytotoxicity, increasing the effects of substances [34].
A less significant influence was observed on
plantlets photosynthetic apparatus, with mixed
response concerning pigment content, stimulatory in
the case of cucumber and inhibitory in the case of
radish, once again underlining the sensitivity of radish
towards exogenous application of compounds.
However, the functioning of the photosystem II
appeared relatively unaffected, considering the
chlorophyll fluorescence values, comparable under
most treatments with those of control plants.
Toxicity of aqueous leachates from various vegetal
organs of both herbaceous [32] and tree species
[46,39,40] are reported, the water soluble nature of
involved compounds indicating the occurrence of such
effects under natural conditions. Both alkaloids
[30,37,19] and phenolics [33,2] may influence
germination and growth of plants, possible
mechanisms of action including altered nutrient uptake,
inhibition of cell division, effects on enzymes and
proteins [17] or DNA and RNA interactions, modified
protein biosynthesis [44,4]. Autotoxicity of these
compounds was observed for several tree species,
including conifers [13,24,35]. In the case of Taxus
baccata, autotoxicity was proposed [38,29] but, as far
as the available literature is concerned, effective testing
was not performed. Furthermore, there are few reports
on the toxicity of Taxus extracts on other species
[9,47], which, however, indicate the allelopathic effects
of yew. The obtained results confirm the previous
findings, and suggest an opportunity for autoxicity tests
of Taxus leachates.
Conclusions
Taking in consideration all parameters
analyzed, the Taxus extracts appear to exert a certain
phytotoxic activity on test species used. Phenolic
compounds and alkaloids may be responsible for
displayed effects, further identification of compounds
and corresponding toxicity testing being required.
Furthermore, toxicity assessment of Taxus extracts
towards Taxus seedlings could be useful for
autotoxicity hypothesis testing.

124
References
1.Aerts RJ, Snoeijer W, Meijden E, Verpoorte R
(1991). Allelopathic inhibition of seed germination by
Cinchona alkaloids? Phytochemistry 30(9):2947-2951.
2.Aliotta G, Cafiero G, Fiorentino A, Strumia S (1993).
Inhibition of radish germination and root growth by
coumarin and phenylpropanoids. Journal of Chemical
Ecology 19(2):175-183.
3.Casals P, Camprodon J, Caritat A, Rios AI, Guixe D,
Garcia-Marti X, Martin-Alcon S, Coll L (2015). Forest
structure of Mediterranean yew (Taxus baccata L.)
populations and neighbor effects on juvenile yew
performance in the NE Iberian Peninsula. Forest
Systems 24(3).
4.Cheng F, Cheng Z (2015). Research progress on the
use of plant allelopathy in agriculture and the
physiological and ecological mechanisms of
allelopathy. Front Plant Sci 6:1020.
5.Chiapusio G, Sánchez AM, Reigosa M.J., González
L, Pellissier F(1997). Do germination indices
adequately reflect allelochemical effects on
germination process? Journal of Chemical Ecology
23(11):2445-2453.
6.Das B, Anjani G, Kashinatham A, Venkataiah B, Rao
S (1998). Taxoids, lignans, and simple phenolic
compounds from a sample of the needles of Himalayan
Taxus baccata. Natural Products Sciences 4(2):78-83.
7.Das B, Takhi M, Srinivas KVNS, Yadav JS (1993).
Phenolics from needles of Himalayan Taxus baccata.
The International Journal of Plant Biochemestry in
Phytochemistry, 33(6):1489-1491.
8.Dhar A, Vacik H, Ruprech H, Klumpp R (2013).
Population dynamics of endangered English yew
(Taxus baccata L.): Implication for conservation and
management In: Endangered Species: Habitat,
Protection and Ecological Significance, Nova Science
Publishers, pp. 183.
9.Filova A, Rovna K, Kovar M (2007). Allelophatic
effects and secondary metabolites in life yew (Taxus
spp.). Acta Physiologiae Plantarum 29(suppl 1):S33-
S56.
10.Herald, T.J., Gadgil, P., Tilley, M. (2012). High-
throughput micro plate assays for screening flavonoid
content and DPPH-scavenging activity in sorghum
bran and flour. Journal of Agricultural and Food
Chemistry 92(11):2326-2331.
Itokawa H, Lee K (2003) Introduction. In: Itokawa H,
Lee K (Eds.) Taxus: The genus Taxus. Taylor &
Francis, pp. 1-18.
11.Jenniskens LHD, Rozendaal ELM, Beek TA (1996).
Identification of six taxine alkaloids from Taxus
baccata needles. Natural products 59(2):117-123.
12.Kong CH, Chen LC, Xu XH, Wang P, Wang SL
(2008). Allelochemicals and activities in a replanted
Chinese fir (Cunninghamia lanceolate (Lamb.)Hook)
tree ecosystem. Journal of Agricultural and Food
Chemistry 56(24):11734-11739.
13.Krogmeier MJ, Bremner JM (1989). Effects of
phenolic acids on seed germination and seedling
growth in soil. Biology and fertile of soils 8(2):116-
122.
14.Lamsal N, Jnawali P, Kawan S, Bhuju S (2013).
Population structure, habitat study and molecular
characterization of Taxus baccata subsp. Wallichiana
(ZUCC.) pilger: and endangered anticancerous plant of
Nepal. A project report submitted in the partial
fulfillment of requirements for the degree of Bachelor
of Tehnology in Biotechnology, Departament of
Biotechnology, School of Science, Kathmandu
University.
15.Li X, Yu M, Ruan X, Zhang Y, Wang Q (2014).
Phytoxicity of 4,8-dihydroxy-1-tetralone isolated from
Carya cathayensis Sarg. to various plant species.
Molecules 19:15452-15467.
16.Li Z, Wang Q, Ruan X, Pan C, Jiang D (2010).
Phenolics and plant allelopathy. Molecules 15:8933-
8952.
17.Liu Y, Zeng R, An M, Mallik A, Luo S (2008).
Autotoxicity in agriculture and forestry. In: Zeng, R.S.,
Mallik, A.U., Luo, S.M. (Ed.), Allelopathy in
Sustainable Agriculture and Forestry, Springer, pp.283-
301.
18.Lovett J, Ryuntyu M (1992). Alelopathy:
broadening the context In: Rizvi, S.J.H., Rizvi, V.
(Ed.), Alelopathy: Basic and applied aspects, Chapman
& Hall, London, pp.11-20.
19.Lovett JV, Hoult AHC (1998). Allelopathy in plants
In: Roberts, M.F., Wink, M. (Ed.), Alkaloids:
Biochemistry, Ecology, and Medicinal applications,
Springer Science + Business Media, LLC, 337-347.
20.Maclaren P (1983). Chemical welfare in the forest/
Review of Allelopathy with regard to New Zealand.
New Zealand Journal of Forestry:73-92.
21.Malik S, Cosido RM, Mirjalili MH, Moyano E,
Palazon J, Bonfill M (2011) . Production of the
anticancer drug in Taxus baccata suspension cultures:
a review. Process Biochemistry 46(1):23-34.
22.Mominul Islam AKM, Kato-Noguchi H (2014).
Phytotoxic activity of Ocimum tenuiflorum extracts on
germination and seedling growth of different plant
species. The Scientific World Journal
doi:10.1155/2014/676242.
23.Monnier Y, Vila B, Montes N, Bousquet-Melou A,
Prevosto B, Fernandez C (2011). Fertilization and
allelopathy modify Pinus halepensis saplings crown
acclimation to shade. Trees-Structure and Function
25(3):497-507.
24.Oltean M, Negrean G, Popescu A, Roman N,
Dihoru G, Sanda V, Mihailescu S (1994). Lista Rosie a
plantelor superioare din Romania [The Red list of
vascular plants of Romania]. Studii, sinteze,
documentatii de ecologie, Institutul de biologie,
Bucuresti, 1:1-52.
25.Oprea A (2005). Lista critică a plantelor vasculare
din România [The critical list of vascular plants of

125
Romania], Ed. Universităţii „Alexandru Ioan Cuza”
Iaşi, 27.
26.Orwa C, Mutua A, Kindt R, Jamnadass R, Anthony
S (2009). Agroforesty Database: a tree reference and
selection guide version 4.0. Retrieved 2016 April 04
from
(http://www.worldagroforestry.org/sites/treedbs/treedat
abases.asp).
27.Parmar VS, Jha A (1998). Chemical constituents of
Taxus species. Studies in Natural Products Chemistry
20:79-134.
28.Perrin PM, Mitchell FJG (2013). Effects of shade
on growth, biomass allocation and leaf morphology in
European yew (Taxus baccata L.). European Journal of
Forest Research 132(2):211-218.
29.Petroski RJ, DornbosJr, DL, Powell RG (1990).
Germination and growth inhibition of annual ryegrass
(Lolium multiflorum L.) and alfalfa (Medicago sativa
L.) by loline alkaloids and synthetic N-acylloline
derivatives. Journal of Agricultural and Food
Chemistry 38(8):1716-1718.
30.Piovesan G, Saba EP, Biondi F, Alessandrini A,
Filippo A, Schirone B (2009). Population ecology of
yew (Taxus baccata L.) in the Central Apennines:
Spatial patterns and their relevance for conservation
strategies. Plant Ecology 205(1):23-46, Springer.
31.Read JJ, Jensen EH (1989). Phytotoxicity of water-
soluble substances from alfalfa and barley soil extract
on four crop species. Journal of Chemical Ecology
15(2):619-628.
32.Reigosa MJ, Souto XC, Gonz`lez L (1999). Effect
of phenolic compounds on the germination of six
weeds species. Plant Growth Regulation 28:83-88.
33.Rice E L (1984). Allelopathy, Second Edition. 356-
360, Academic press, Inc.
34.Ruan X, Li ZH, Wang Q, Pan CD, Jiang D, Wang
GG (2011). Autotoxicity and allelopathy of 3,4-
dihydroxycetophenome isolated from Picea
schrenkiana Needles. Molecules 16:8874-8893.
35.Sekutowski T (2011). Application of bioassays in
studies on phytotoxic herbicide residues in the soil
environment. Herbicides and Environment:253-272.
36.Shao H, Huang X, Zhang Y, Zhang C (2013). Main
alkaloids of Peganum harmala L. and their different
effects on dicot and monocot crops. Molecules
18(3):2623-2634.
37.Svenning J, Magard E (1998). Population ecology
and conservation status of the last natural population of
English yew Taxus baccata in Danmark. Biological
Conservation 88:173-182.
38.Teixeira da Silva J, Karimi J, Mohsenzadeh S,
Dobránszki J (2015). Allelopathic potential of select
gymnospermous trees. Journal of Forest and
Environmental Science 31(2):109-118.
39.Terzi I. (2001). Allelopathic effects of walnut leaf
extracts and juglone on seed germination and seedling
growth. The Journal of Horticulture Science and
Biotechnology 76(4):436-440.
40.Thomas PA, Polwart A. (2003). Taxus baccata L.
Journal of Ecology 91(3):489-524;
41.Williams R, Hoagland RE (1982). The effects of
naturally occurring phenolic compounds on seed
germination. Weed Science 30:206-212.
42.Wink M (1983). Inhibition of seed germination by
quinoliyidine alkaloids. Aspects if allelopathy in
Lupinus albus L. Planta 158:365-368.
43.Wink M, Latz-Bruning B (1995). Allelopathic
properties of alkaloids and other natural products.
Possible modes of action. In: Inderjit, Dakshini K M
M,., Einhellig F A (Eds.). Allelopathy: Organism,
Processes, and Application, American Chemical
Society, pp.117-126.
44.Xiangxiang G, Li M, Zongjun G, Zuowen S (2009).
Allelopathic effects of Conyza canadesis the
germination and growth of wheat, sorghum, cucumber,
rape and radish. Allelopathy Journal 23(2):287-296.
45.Yukiko I, Yasuo K, Minoru T (2001). Effects of
phenolic compounds on seed germination of
Shirakamba Bich, Betula platyphylla var. japonica.
Eurasian Journal of Forest Research 2:17-25.
46.Zhang Y, Lu S, Gao H (2010). Allelopathic effect
of different solvent extraction from seed of Taxus
chinensis var. mairei on cabbage seed germination and
seedling growth. Chinese Agricultural Science Bulletin
26:190-194.





























