Embed Size (px)
Citation preview

Colloids and Surfaces, 15 (1985) 239- 247 Elsevier Science Publishers B.V., Amsterdam - Printed in The Netherlands
239
THE ELECTROPHORETIC PROPERTIES OF DIPALMITOYLPHOSPHATIDYLCHOLINE-PHOSPHATIDYLINOSITOL VESICLES IN THE PRESENCE OF LANTHANUM IONS
M.D. REBOIRAS* and M.N. JONES
Department of Biochemistry, University of Manchester, Manchester, Ml3 9PL (United Kingdom)
(Received 21 January 1985; accepted in final form 19 March 1985)
ABSTRACT
Large multilamellar vesicles have been prepared from dipalmitoylphosphatidylcholine
(DPPC)- phosphatidylinositol (PI) mixtures over the complete range of composition.
The electrophoretic mobilities of the vesicles have been measured in a buffer at pH 6.6
and as a function of lanthanum ion concentration over a range covering the zero point
of charge. The zeta potentials of the vesicles increase with PI content and the degree of
ionisation of the PI molecules lies in the range 0.16 to 0.23. The zeta potentials have
been interpreted in terms of La3+ ion binding using a Gouy-Chapman-Stern model
as proposed by Ottewill and co-workers. Apparent and intrinsic binding constants and
the number of binding sites have been determined. The Gibbs energy of ion adsorption
(AGoads ) has been found to obey the linear relationship:
AGoads (kJ mall’) = (66.9 (* 1.7)~~~ + 41.2 (? 0.8)~~~~~)
which demonstrates that each phospholipid molecule binds La3+ ions independently.
INTRODUCTION
The interaction of inorganic ions with phospholipids either in the form of monolayers or dispersed in bilayer systems has been investigated in some detail [l-13]. The studies have been predominantly concerned with the binding of ions to surfaces consisting of a single phospholipid, notably the phosphatidylcholines (PC) or phosphatidylserines (PS), although ion binding to binary phospholipid systems of PC plus PS and PC plus phosphatidyl- ethanolamine (PE) covering a limited composition range have been inves- tigated [l] . The biochemical significance of phosphatidylinositol (PI) in membranes, particularly with regard to its role in ion transport and the influence of hormones and neurotransmitters on its metabolism [ 14-161, has stimulated work on the properties of the binary system PC plus PI [S, 17, 181. These studies include the binding of calcium and magnesium ions
*Present address: Department0 de Electroquimica, Facultad de Ciencias, Universidad
Autonoma, Canto Blanco, Madrid 34, Spain.
0166.6622/85/$03.30 G 1985 Elsevier Science Publishers B.V

240
to vesicle surfaces and it has been found that Ca2’ binds more strongly than
Mg” to PC-PI vesicles [ 181. Microelectrophoresis has been used by a number of investigators to study
ion-vesicle interactions [l, 3-6, 8, 9, 17, 181 and the results have generally been interpreted in terms of the Gouy-Chapman-Stern theory. In some cases, theoretical fits to zeta potantial-log ion concentration data have required the introduction of several parameters such as the distance of the plane of shear from the phospholipid head-group plane and the area per phospholipid in the surface as well as binding constants [l, 33. The theory of Ottewill and co-workers [19, 201 developed to interpret electrophoretic data on the adsorption of surfactant ions on colloidal particles assumes a Stern layer but requires no further assumptions other than that, in the region of the zero point of charge, the measured zeta potential (5) can be equated to the potential at the outer Helmholtz plane. Here we have applied the theory of Ottewill and co-workers to interpret the effect of lanthanum ions on the electrophoretic mobilities of PC-PI multilamellar vesicles cover- ing the complete composition range. Lanthanum ion interaction with PC bilayers [4, 7, 211 has been studied and La3’ has been used as a specific Ca*’ substitute or antagonist in biological systems [22, 231.
THEORY
The change in surface charge density (A Ud) at the outer Helmholtz plane (OHP) due to adsorption of an ion of valency z at a concentration c is given by the Stern equation
-Aad = zeN,k,c
1 + k,c
where N, is the number of binding sites per unit area, e the electronic charge and k2 the apparent binding constant [19, 241. For a spherical particle (radius a), the change in charge (A&) is related to the change in potential (A $ d) at the OHP by the relations
A& - =-Aad= EoEr(l + Ka)A$‘d
4na2 (2)
a
where co and ey are the permittivity of free space and relative permittivity, respectively, K -’ is the Debye length. Assuming that A $ d fi A{, it follows that
A(=- a X zeN, k,c
EoEr(l + Ka) 1 + k2c (3)
and if Ka 9 1 then
-zeN1 k2c Al =
EOE~K(~ + k,c) (4)

241
If the zeta potential in the absence of adsorption is f” then { is given by
(={O-A{={O+ zeNlk2c
EOEyK(l + kzc)
At the zero point of charge* it can be shown that
= 2.303 co foerKt” - -1
f=o zeN,
(5)
(6)
and
1 zeN, -= k, --1 CO EoErS“K
where co is the ion concentration at the zero point of charge. Both N1 and k2 can be determined by solving Eqns (6) and (7) simultaneously given c as a function of log c.
EXPERIMENTAL
Materials
L-a-Dipalmitoylphosphatidylcholine (DPPC), molecular weight 734, was obtained from Sigma London Chemical Co., approximately 99% pure (product number P-6267). Phosphatidylinositol (PI) (sodium salt from soy- bean, molecular weight 846 [25] ) was Grade I from Lipid Products, South Nutfield, U.K. All other materials used in the preparation of solutions were of analytical grade.
Methods
Multilamellar vesicles were prepared by adding the required amounts of phospholipid stock solutions (5 mg crnm3 in chloroform) together with 50 cm3 of chloroform/methanol mixture (4:l v/v) to a l-l round-bottomed flask. The solvent was removed by rotary evaporation at a temperature of 50-55°C. The resulting lipid film was flushed with nitrogen and dispersed by shaking with 5-7 cm3 buffer of pH 6.6 and then incubated at 55-6O”C. After incubation the total volume was made up to 32 cm3 with buffer. The buffer (pH 6.6) was prepared from 20 r&f imidazole (pK, 6.953) plus hydrochloric acid giving an ionic strength of 0.0138 M.
Electrophoretic mobilities were measured using a Rank Bros. (Bottisham, Cambridge, U.K.) microelectrophoresis apparatus [26] with a flat cell and grey platinum electrodes. The time (t) of migration of the multilamellar
*Here and subsequent zero point of charge refers to vesicles having no net charge, i.e. the zero point of electrokinetic charge or the isoelectric point.

242
vesicles or small vesicle aggregates over a known distance (d) was determined and used to calculate their mobilities (u) from the equation
d Le u=-
tv
where L, is the effective cell length and at least 12 velocities were measured in polarity of the field. The zeta potentials chowski equation applicable when ~a S 1
*=Z eoer
(8)
V the applied voltage. Generally, each direction by reversing the were calculated from the Smolu-
(9)
where, for dilute aqueous media, we took the viscosity Q = 8.904 X 10W4 N s rnm2, er = 78.5, and e. = 8.854 X lo-l2 C2 J-l m-l at 25°C. For an ionic strength of 0.0138 M, K = 3.9 X lo8 m-‘, thus a must be > 2.6 nm for Eqn (9) to hold. Since the radii of DPPC-PI ‘sonicated’ vesicles exceed this radius [17] , Eqn (9) should apply to multilamellar vesicles. Addition of La3+ ions as’ La(N03)3 leads to aggregation of the vesicles; however, the mobilities of small clusters of multilamellar vesicles did not differ significantly from single vesicles. Table 1 gives results for DPPC-PI (50:50 wt%) vesicles to- gether with the errors in the zeta potentials. The percentage error in zeta potential was largest in the vicinity of the zero point of charge where the particles were moving slowest.
TABLE 1
Electrophoretic mobilities (u) and zeta potentials (r) for multilamellar DPPC-PI (50:50 wt%) vesicles at 25”C, pH 6.6
[La”] X 10’ (M) u X 10’ (m’s’ V’) f ? SDa (mV) nb
0.00 -5.09 -65.17 i 2.09 102 0.50 -4.42 -56.66 ? 1.74 44 1.00 -3.84 -49.27 * 1.62 44 2.00 -2.40 --30.74 ? 1.11 49 3.00 -1.75 -22.35 i 0.63 47 4.00 -1.14 -14.54 i 0.57 46 5.00 -0.88 -11.24 + 0.64 28 6.00 -0.58 -7.45 * 0.51 35 7.00 -0.52 -6.61 f 0.71 11 8.00 -0.39 -5.00 f 0.53 12
12.00 +0.44 +5.66 + 1.13 10 20.00 iO.78 +9.99 ?r 0.68 10
a Standard deviation. bNumber of particles observed.
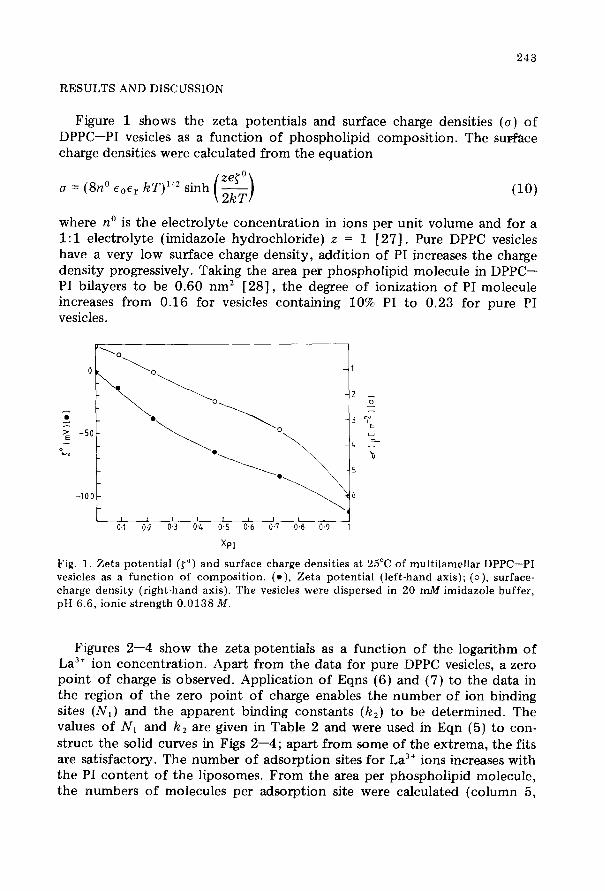
243
RESULTS AND DISCUSSION
Figure 1 shows the zeta potentials and surface charge densities (u) of DPPC-PI vesicles as a function of phospholipid composition. The surface charge densities were calculated from the equation
u = (Go eOer hT)“* sinh (10)
where no is the electrolyte concentration in ions per unit volume and for a 1: 1 electrolyte (imidazole hydrochloride) z = 1 [27]. Pure DPPC vesicles have a very low surface charge density, addition of PI increases the charge density progressively. Taking the area per phospholipid molecule in DPPC- PI bilayers to be 0.60 nm2 [28], the degree of ionization of PI molecule increases from 0.16 for vesicles containing 10% PI to 0.23 for pure PI vesicles.
-lOO-
I ,I I I 0.1 0.2 0.3 04 0.5 0.6 0.7 04 09 1
XPI
Fig. 1. Zeta potential (to) and surface charge densities at 25°C of multilamellar DPPC-PI vesicles as a function of composition. (e), Zeta potential (left-hand axis); (o), surface- charge density (right-hand axis). The vesicles were dispersed in 20 mA4 imidazole buffer, pH 6.6, ionic strength 0.0138 M.
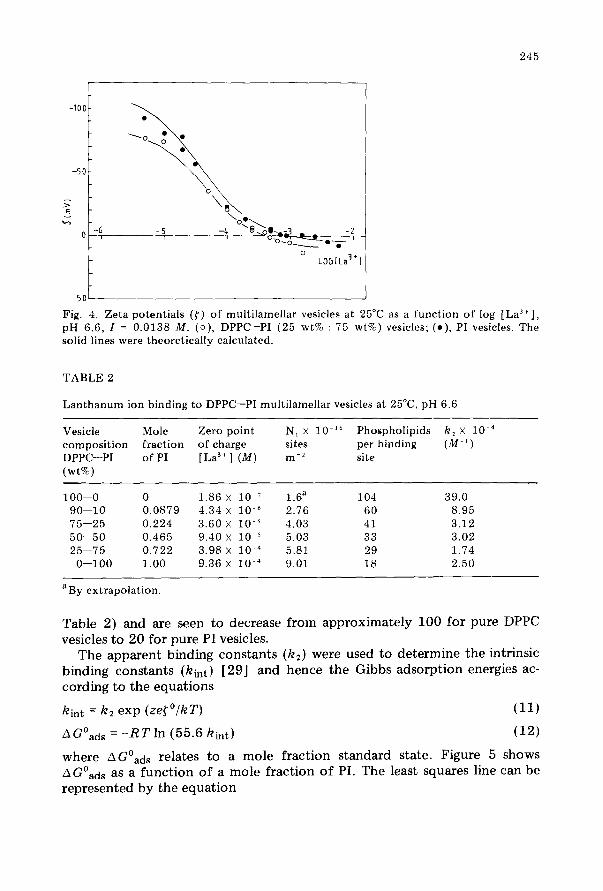
Figures 2-4 show the zeta potentials as a function of the logarithm of La3’ ion concentration. Apart from the data for pure DPPC vesicles, a zero point of charge is observed. Application of Eqns (6) and (7) to the data in the region of the zero point of charge enables the number of ion binding sites (N,) and the apparent binding constants (k,) to be determined. The values of N1 and k, are given in Table 2 and were used in Eqn (5) to con- struct the solid curves in Figs 2-4; apart from some of the extrema, the fits are satisfactory. The number of adsorption sites for La3’ ions increases with the PI content of the liposomes. From the area per phospholipid molecule, the numbers of molecules per adsorption site were calculated (column 5,
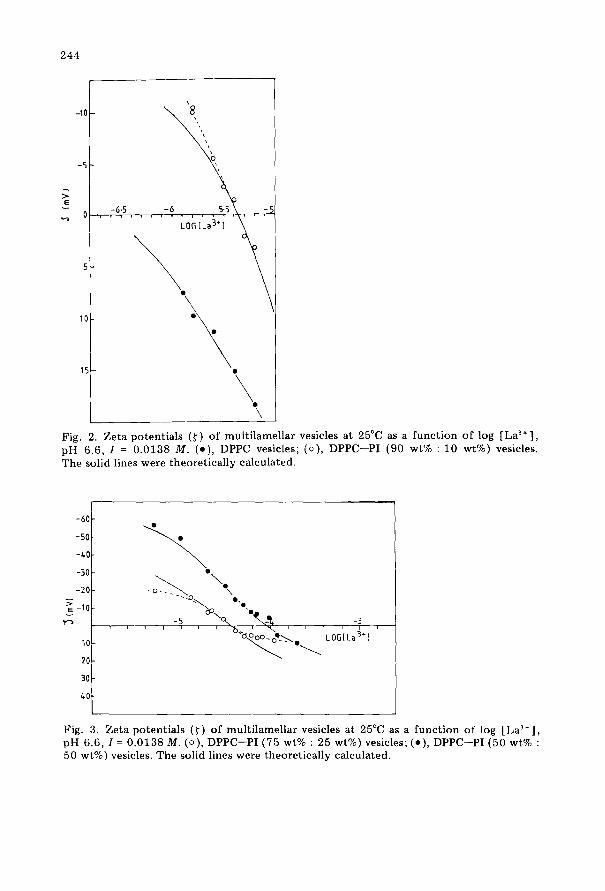
244
-10 -
-5 -
r; cJ I),,:‘;5 ,,,,.,
lo-
15- .
\
i
Fig. 2. Zeta potentials (f) of multilamellar vesicles at 25°C as a function of log [Las+], pH 6.6, I = 0.0138 M. (e), DPPC vesicles; (o), DPPC-PI (90 wt% : 10 wt%) vesicles. The solid lines were theoretically calculated.
-6O-
-so-
-LO-
-30 -
-2o-
5: E -lO-
ys -3 I I I / 1
lo- l \ LOG[La3+l
Fig. 3. Zeta potentials (r) of multilamellar vesicles at 25°C as a function of log [La3+], pH 6.6, I = 0.0138 M. (o), DPPC-PI (75 wt% : 25 wt%) vesicles; (.), DPPC-PI (50 wt% : 50 wt%) vesicles. The solid lines were theoretically calculated.
245
50
Fig. 4. Zeta potentials (<) of multilamellar vesicles at 25°C as a function of log [La3’], pH 6.6, I = 0.0138 M. (o), DPPC-PI (25 wt% : 75 wt%) vesicles; (a), PI vesicles. The solid lines were theoretically calculated.
TABLE 2
Lanthanum ion binding to DPPC-PI multilamellar vesicles at 25”C, pH 6.6
Vesicle Mole composition fraction DPPC-PI of PI (wt%)
100-0 0 1.86 x lo-’ 1.6a 104 39.0 90-10 0.0879 4.34 x 1o-6 2.76 60 8.95 75-25 0.224 3.60 x lo-’ 4.03 41 3.12 50---50 0.465 9.40 x 1om5 5.03 33 3.02 25-75 0.722 3.98 x 1O-4 5.81 29 1.74
O-100 1.00 9.36 x 10m4 9.01 18 2.50
Zero point N, x lo-l6 Phospholipids k, X 1O-4
of charge sites per binding (M-’ ) [La3+l (M) mm2 site
aBy extrapolation.
Table 2) and are seen to decrease from approximately 100 for pure DPPC vesicles to 20 for pure PI vesicles.
The apparent binding constants (k,) were used to determine the intrinsic binding constants (kht) [29] and hence the Gibbs adsorption energies ac- cording to the equations
hint = kz exp (ze[‘/kT)
AGoads = -RT In (55.6 kht)
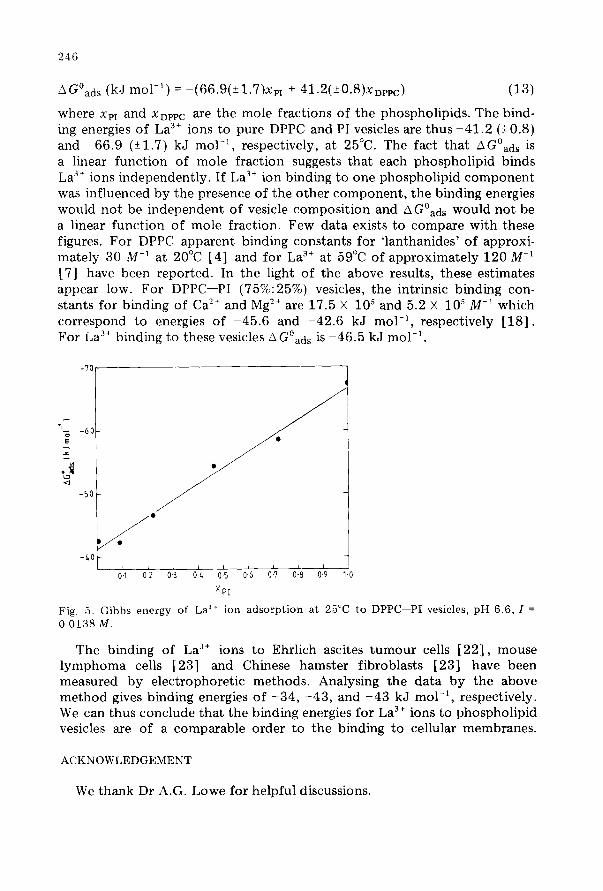
where AGoads relates to a mole fraction standard state. Figure 5
A Goads as a function of a mole fraction of PI. The least squares line represented by the equation
(11)
(12)
shows can be
246
A Goads (kJ mol-‘) = -(66.9(+1.7)x,, + 41.2(~0.S)xnppc) (13)
where xpI and xnppc are the mole fractions of the phospholipids. The bind- ing energies of La3’ ions to pure DPPC and PI vesicles are thus -41.2 (10.8) and -66.9 (21.7) kJ mol-‘, respectively, at 25°C. The fact that AGoads is a linear function of mole fraction suggests that each phospholipid binds La3’ ions independently. If La3’ ion binding to one phospholipid component was influenced by the presence of the other component, the binding energies would not be independent of vesicle composition and AGoads would not be a linear function of mole fraction. Few data exists to compare with these figures. For DPPC apparent binding constants for ‘lanthanides’ of approxi- mately 30 M-’ at 20°C [4] and for La3’ at 59°C of approximately 120 M-l [7] have been reported. In the light of the above results, these estimates appear low. For DPPC-PI (75%:25%) vesicles, the intrinsic binding con- stants for binding of Ca2’ and Mg2’ are 17.5 X lo5 and 5.2 X 10’ M-' which correspond to energies of -45.6 and -42.6 kJ mol-‘, respectively [18]. For ia3’ binding to these vesicles AGoad, is -46.5 kJ mol-‘.
XPI
Fig, 5. Gibbs energy of La” ion adsorption at 25°C to DPPC-PI vesicles, pH 6.6, I =
0.0138 M.
The binding of La3’ ions to Ehrlich ascites tumour cells [22], mouse lymphoma cells [23] and Chinese hamster fibroblasts [23] have been measured by electrophoretic methods. Analysing the data by the above method gives binding energies of -34, -43, and -43 kJ mol-‘, respectively. We can thus conclude that the binding energies for La3’ ions to phospholipid vesicles are of a comparable order to the binding to cellular membranes.
ACKNOWLEDGEMENT
We thank Dr A.G. Lowe for helpful discussions.

247
REFERENCES
1
2 3
4
9
10 11 12
13
14 15
16
17
18
19
20 21
22 23 24
25
26
27
28 29
S. McLaughlin, N. Mulrine, T. Gresalfi, G. Vaio and A. McLaughlin, J. Gen. Physiol., 77 (1981) 445-473. J.S. Puskin and M.T. Coene, J. Membr. Biol., 52 (1980) 69-74. A. Lau, A. McLaughlin and S. McLaughlin, Biochim. Biophys. Acta, 645 (1981) 279-292. H. Hauser, M.C. Phillips, B.A., Levine and R.J.P. Williams, Eur. J. Biochem., 58 (1975) 133-144. H. Hauser, A. Darke and M.C. Phillips, Eur. J. Biochem., 62 (1976) 335-344. S.A. Tatulian, Biochem. Biophys. Acta, 736 (1983) 189-195. H. Akutsu and J. Seelig, Biochemistry, 20 (1980) 7366-7373. K. Okhi, T. Sekiya, T. Yamauchi and Y. Nozawa, Biochim. Biophys. Acta, 644 (1981) 165-174. H. Hauser, M.C. Phillips and M.D. Barratt, Biochim. Biophys. Acta, 413 (1975) 341-353. H. Oshima, Y. Inoko and T. Mitsui, J. Colloid Interface Sci., 86 (1982) 57-72. S. Ohki, N. Duzgunes and K. Leonards, Biochemistry, 21 (1982) 2127-2133. A. McLaughlin, C. Grathwohl and S. McLaughlin, Biochim. Biophys. Acta, 513 (1978) 338-357. H. Hauser, C.C. Hinckley, J. Krebs, B.A. Levine, M.C. Phillips and R.J.P. Williams, Biochim. Biophys. Acta, 468 (1977) 364-377. R.H. Michell, Trends Biochem. Sci., 4 (1979) 128-131. T.W. Honeyman, W. Strohsnitter, C.R. Scheid and R.J. Schimmel, Biochem. J., 212 (1983) 489-498. M.J. Merridge, R.M.C. Dawson, C.P. Downes, J.P. Heslop and R.F. Irvine, Biochem. J., 212 (1983) 473--482. K. Hammond, M.D. Reboiras, I.G. Lyle and M.N. Jones, Biochim. Biophys. Acta, 774 (1984) 19-25. K. Hammond, M.D. Reboiras, I.G. Lyle and M.N. Jones, Colloids Surfaces, 10 (1984) 143-153. R.H. Ottewill, M.C. Rastogi and A. Watanabe, Trans. Faraday Sot., 56 (1960) 854- 865. R.H. Ottewill and A. Watanabe, Kolloid Z., 170 (1960) 132-139. B.Z. Chowdhry, G. Lipka, A.W. Dalziel and J.M. Sturtevant, Biophys. J., 45 (1984) 633-635. T.C. Smith, J. Cell. Physiol., 87 (1976) 47-52. R.G. Greig, M.N. Jones and S.R. Ayad, Biochem. J., 160 (1976) 325-334. R.J. Hunter, Zeta Potential in Colloid Science, Academic, New York, NY, 1981, Ch. 8, page 309. L. Ter-Minassian-Saraga and G. Mandelmont, J. Colloid Interface Sci., 85 (1982) 375-388. A.D. Bangham, R. Flemans, D.H. Heard and G.V.F. Seaman, Nature, 182 (1958) 642-644. R.J. Hunter, Zeta Potential in Colloid Science, Academic, New York, NY, 1981, Ch. 2, p. 28. I.G. Lyle, K. Hammond and M.N. Jones, unpublished observations. C. Tanford, Physical Chemistry of Macromolecules, Wiley, New York, NY, 1961, Ch. 8, p. 535.



![A method for determining electrophoretic and …...[4,5]. Current techniques for measuring electrophoretic mo-bility include an electroacoustic method [6], electrophoretic light scattering](https://img.pdfslide.net/doc/110x75/5f08e22b7e708231d4242f99/a-method-for-determining-electrophoretic-and-45-current-techniques-for-measuring.jpg)

























