Embed Size (px)
Citation preview

Records of the Western Australian Museum Supplement No. 58: 335-347 (2000).
The evolution, radiation and biogeography of early spiriferid brachiopods
Remy Gourvennec
Laboratoire de Paleontologie. UMR 6538 "Domaines oceaniques". UFR Sciences and Techniques.Universite de Bretagne Occidentale, Av. Le Gorgeu, F-29285 BREST Cedex, France
e-mail: [email protected]
Abstract - Spiriferid brachiopods originated in South-East Asia during theLate Ordovician and subsequently became geographically widespread duringthe Silurian. This study deals with the ways this took place, the routes takenduring the expansion of this group and the implications to palaeogeography.During the Silurian, two peaks of diversification are recognised. The principalevolutionary trends evident within different lineages are outlined.
INTRODUCTION
Until recently, one of the most prolific group ofbrachiopods, the spiriferids, was thought to haveappeared in South China during the Early Silurian,but it has been now shown they originated in theLate Ordovician (Rong et al. 1994; Rong and Zhan1996). This investigation probes the mode ofdevelopment and geographical dispersal patterns ofspiriferids during the Silurian. In order to obtainreasonable precision, a comprehensive database onSilurian spiriferid species was compiled usingpublished literature.
In the course of the revision of the Treatise onInvertebrate Paleontology, Brachiopoda, somerepeated, but incorrect, suffixes (e.g. «idid») createdsome difficulties for informal useage. All theauthors endorsed the following suggestion: use «id»in place of «idid», etc., with lower case letters forinformal use and, in all formal uses, uppercase firstletter and termination in accordance to the ICZNcode. This principle is applied herein. The familiesdiscussed below are those defined and recognisedby Carter et al. (1994).
METHODS
The database for this investigation was compiledusing recent published works (species listspublished after 1980) in order to avoidnomenclatural confusion. Most works includerevised lists of previously published material.Where recent papers where unavailable for certainregions, older data was used. In such cases, wherenecessary, original designations were re-evaluatedto fit modern taxonomic concepts. The range of eachspecies was compared to current graptolitezonation, where available, so as to obtain detailed,homogeneous age control. Silurian spiriferidsconsist of more than 360 species in 46 genera, 25°!c)
of which are in open nomenclature. Spacelimitations preclude publication of the entire specieslist but requests for this information can be sent tothe author.
ORIGIN OF SPIRIFERIDS
Both monophyletic and polyphyletic origins havebeen suggested for the spire-bearing brachiopods.Copper (in Copper and Gourvennec, 1996), insuggesting a monophyletic origin, points out thatthis option would require the presence of acomplete jugum in the early eospiriferids. In thepolyphyletic theories put forward, the spiriferidsare derived either from an impunctate orthid,possibly a platyorthid (Wright 1979), or from arhynchonellid stock (Grunt 1982). These differentoptions are reviewed in Copper and Gourvennec(1996) who conclude that «new morphologicaldiscoveries in the oldest species from each of thethree orders [respectively Atrypida, Athyrida andSpiriferida] will probably dictate the ultimatephylogenetic scenario».
In their recent work, Rong and Zhan (1996)identified a ventro-laterally directed spiraliumand a small jugal process, but no jugum, in theearly spiriferids, Eospirifer and Striispirifer. A reexamination of the monophyletic theory, in termsof parsimony, is therefore warranted. Focusing onthe early stages of development of thebrachidium, Rong and Zhan (1996) suggestedpossible relationships between early spiriferidsand atrypoids and a common ancestor for the twogroups. Although most of the early atrypoidspossess a jugum, one of them, Cyclospira, lacksthis structure and is reminiscent of Eospirifer butpresents significative differences with other earlyatrypoids. Thus Rong and Zhan suggested acommon ancestor for Eospirifer and Cyclospira.
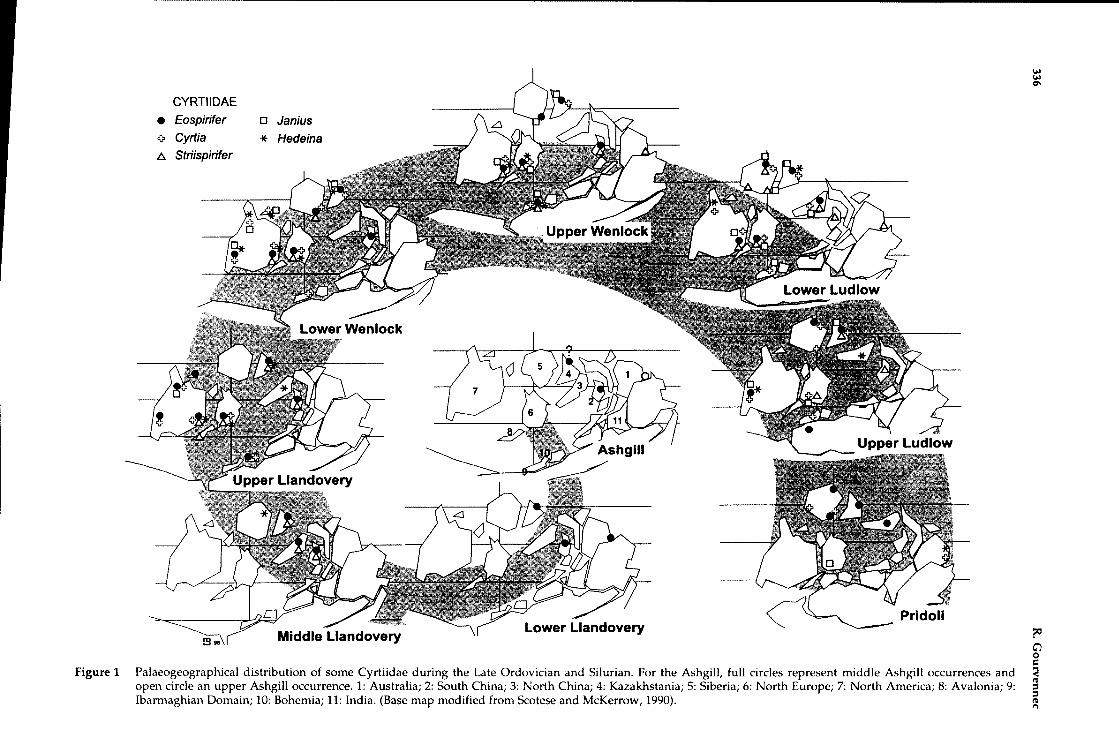
Figure 1 Palaeogeographical distribution of some Cyrtiidae during the Late Ordovician and Silurian. For the Ashgill, full circles represent middle Ashgill occurrences andopen circle an upper Ashgill occurrence. 1: Australia; 2: South China; 3: North China; 4: Kazakhstania; 5: Siberia; 6: North Europe; 7: North America; 8: Avalonia; 9:Ibarmaghian Domain; 10: Bohemia; 11: India. (Base map modified from Scotese and McKerrow, 1990).
CYRTIIDAE• Eospirifer<c> CyrtiaA Striispirifer
o Janius
* Hedeina
Middle Llandovery
\o:l\o:l0\
?'C'lo=~/'ll::s::s/'ll,.,
DELTHYRIDIDAE_ Howellella
t:::.. Delthyris
9 Rufispirifer
\\~
~/ ~~<J
/-/75//---Lower Wenlock
[998
.Upper Wenlock
Lower Ludlow
;;-
Pridoli
mllJ....~
'J;
"9.::!.'""'"....c:0-....llJ,.,::ro
"0oQ..'J;
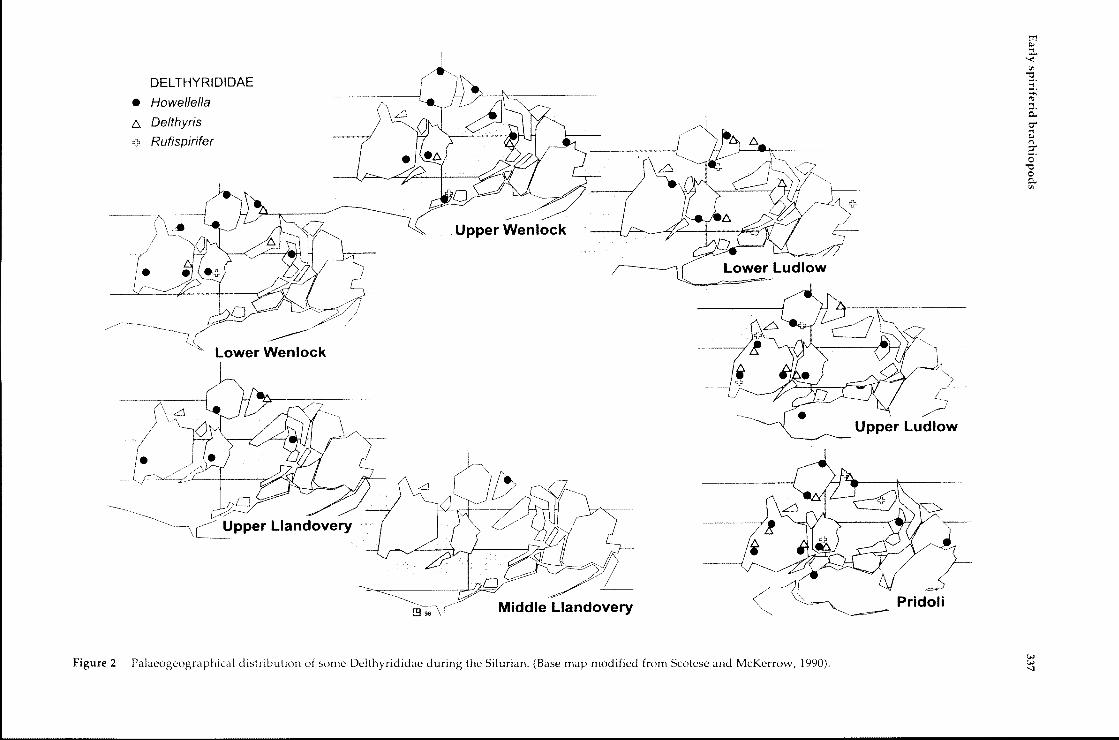
Figure 2 Palaeogeographical distribution of some Delthyrididae during the Silurian. (Base map modified from Scotese and McKerrow, 1990).ww'I

338
GEOGRAPHICAL DISTRIBUTION OF THEEARLY SPIRIFERIDS
The origin of the very early spiriferids in LateOrdovician sequences of South China has beendocumented by Rong et al. (1994) and Rong andZhan (1996) who provided a detailed overview ofprimitive eospiriferine species and attempted toplace them in an evolutionary perspective.
Eospirifer first appeared in the Ashgill ChangwuFormation, Jiangshan area, SE China (Rong et al.1994) and soon spread to neighbouring areas. Duringthe lower Llandovery, Eospirifer reached Kazakhstan(Modzalevskaya and Popov 1995) where anAshgillian occurrence is reported but not ascertained,and Tasmania (Sheehan and Baillie 1981) (Figure 1).From Kazakhstan Eospirifer dispersed to Central Asiaduring the Middle Llandovery and thence to Europeand North America during the Late Llandovery.During this time it also occurred in North China (Fu1982). Significantly, Eospirifer is not reported from theRussian platform and is only sporadically present inthe Altai Range and neighbouring areas. In SouthChina, Eospirifer more or less survived the entireSilurian and is probably the centre of origin for thesubsequent colonisation of Australia in the EarlyLochkovian.
The endemic Yingwuspirifer also appeared inSouth China in the Early Llandovery, but did notsurvive the Llandovery-Wenlock transition. Thecatch-all genus «Nikiforovaena» also remainedendemic to South China until the LlandoveryWenlock boundary where it apparently«disappeared». However, it suddenly «reappeared»in the Lower Ludlow with a widespreaddistribution including South and North China, theSiberian platform and North-Western Europe.Rather than an example of the Lazarus effect, thereappearence is more likely due to the fact thatearliest «Nikiforovaena» probably belong to anothergenus (see below).
South China remained a spiriferid nursery in theMiddle Llandovery, since it is also the centre oforigin of Striispirifer, a genus which becomesgeographically Widespread during the UpperLlandovery with occurrences recorded in NorthChina, Europe and North America (Rong andYang1981; Boucot 1963; Figure 1). Striispirifer is alsosporadically reported in Central Asia (Rong et al.1985) and the Russian platform from Wenlock toLudlow sequences (Modzalevskaja 1990).Striispirifer reached Australia during the UpperWenlock (Strusz 1985) but disappeared during theLower Ludlow. In Europe and North America, itsurvived until the Late Silurian.
Another early spiriferid nursery is represented bysequences in Kazakhstan with the documentationof Eospirifer in the early Llandovery(Modzalevskaya and Popov 1995). Eospirifer is alsoquestionably reported from the Ashgill Ulkuntas
R. Gourvennec
Horizon by Sapelnikov and Rukavischnikova(1975). The first occurrence of Howellella is alsoreported from this region, possibly from MiddleLlandovery beds. The origin of Howellella is thusfairly well constrained, with the only possibleancestor present in the region at this time beingEospirifer. In the Upper Llandovery, Howellellarapidly dispersed all over the world withoccurrences reported almost everywhere, Australiaand North Africa excepted (Figure 2). Hedeinaoriginated in Tuva approximately at the same time(Middle Llandovery). Again the only possibleancestor is Eospirifer. Hedeina became extinct at theend of the Llandovery in Tuva, but spread to theRussian platform, Europe and North Americaduring the Upper Llandovery. In addition to theabove mentioned genera, early endemic taxaappeared in Kazakhstan (the short lived Espella,Middle Llandovery) and on the Russian platform(Eohowellella, extinct in the Lower Ludlow).
In the Upper Llandovery new genera appearedfor the first time outside of South-East Asia. Amongthem is the ubiquitous appearance of Cyrtia. Thisgenus occurs synchronously in sequences fromNorthern Europe, Turkey, Northern and easternAmerica (Figure 1). With its peculiar external shapeand internal morphology this genus is unmistakablein the fossil record, so that its rapid widespreaddistribution, like Howellella, is somewhat puzzling.
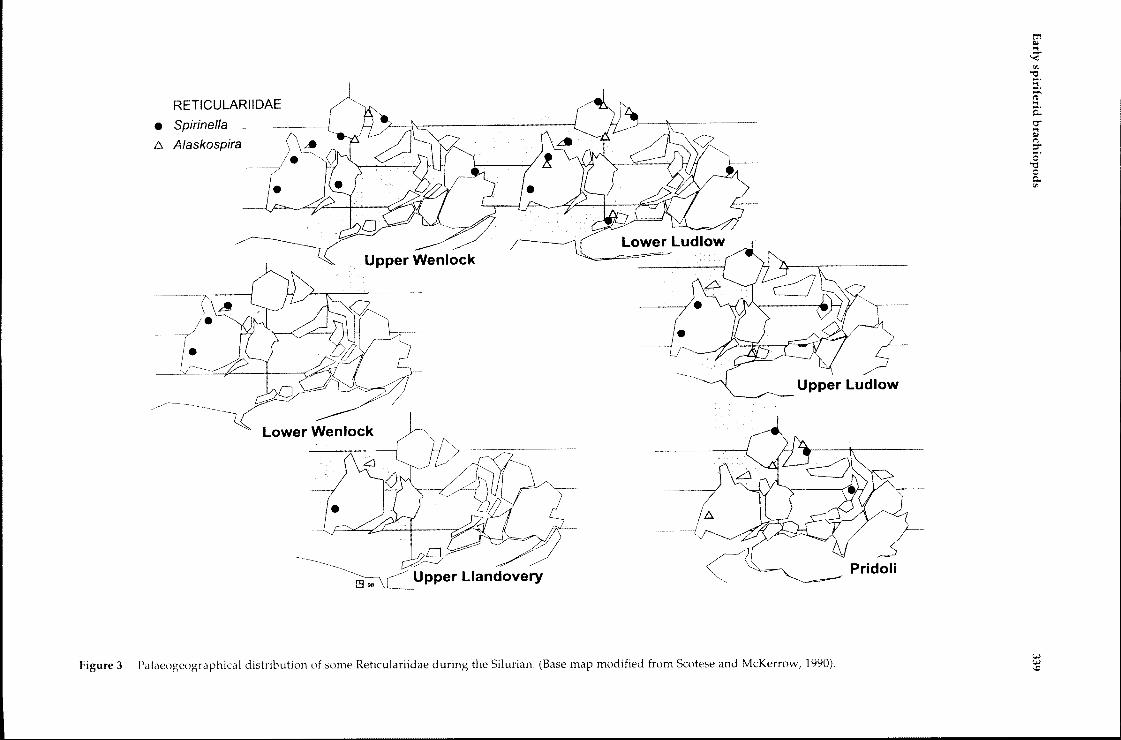
The appearance of endemic taxa in the UpperLlandovery slowly increased the diversity rate ofspiriferids. Mictospirifer from central North Americabecame extinct at the end of the Llandovery andMyriospirifer occurs for the first time in the Balticarea (though this identification is questionable,since it is not reported from Upper Wenlock toLower Devonian sequences). Another significantgenus is Spirinella which first appears in NorthAmerica (Great Basin) in the Llandovery, andsubsequently expands to the Canadian and RussianArctic regions during the Wenlock, and thence toBohemia, Mongolia, South China and Australia inthe Ludlow (Figure 3).
No major evolutionary events occurred duringthe Lower Wenlock. Besides the stock of existingspiriferids, new taxa, most of them endemic, arosein Bohemia (Araspirifer), Arctic Canada(Dicoelospirifer, Plicocyrtia and Eoplicoplasia, anotherLazarus genus), and the eastern Urals (Eoreticularia,Pavdenia). Special mention is made of the genusDelthyris that appeared in Central Asia in the upperLlandovery, and underwent a successful expansionfirst towards North China and North America(lower Wenlock), and then towards Europe andsouth China (upper Wenlock). The presence ofSpurispirifer in the Lower Wenlock of the ArcticCanada (Zhang 1989) is highly questionable: allother known occurrences are from Upper Wenlockto Early Devonian sequences in Bohemia.
RETICULARIIDAE
• Spirinel/a
l::,. Alaskospira
Upper Wenlock
Lower Wenlock
~ ~CJ~~erL1andovery
Figure 3 Palaeogeographical distribution of some Reticulariidae during the Silurian. (Base map modified from Scotese and McKerrow, 1990).
m......--<
00
"2.:::1......11)...0:0";l'"::ro'
"t:loc-oo
(H(H\l:)
340
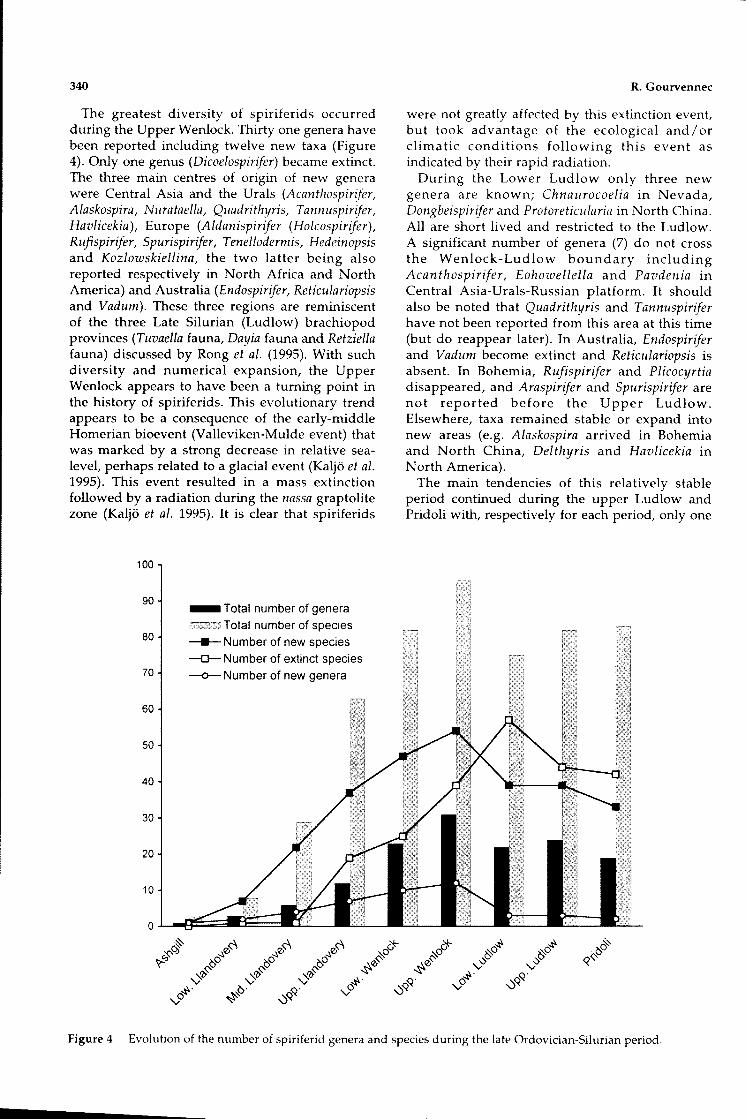
The greatest diversity of spiriferids occurredduring the Upper Wenlock. Thirty one genera havebeen reported including twelve new taxa (Figure4). Only one genus (Dicoelospirifer) became extinct.The three main centres of origin of new generawere Central Asia and the Urals (Acanthospirifer,Alaskospira, Nurataella, Quadrithyris, Tannuspirifer,Havlicekia), Europe (Aldanispirifer (Holcospirifer),Rufispirifer, Spurispirifer, Tenellodermis, Hedeinopsisand Kozlowskiellina, the two latter being alsoreported respectively in North Africa and NorthAmerica) and Australia (Endospirifer, Reticulariopsisand Vadum). These three regions are reminiscentof the three Late Silurian (Ludlow) brachiopodprovinces (Tuvaella fauna, Dayia fauna and Retziellafauna) discussed by Rong et al. (1995). With suchdiversity and numerical expansion, the UpperWenlock appears to have been a turning point inthe history of spiriferids. This evolutionary trendappears to be a consequence of the early-middleHomerian bioevent (Valleviken-Mulde event) thatwas marked by a strong decrease in relative sealevel, perhaps related to a glacial event (Kaljo et al.1995). This event resulted in a mass extinctionfollowed by a radiation during the nassa graptolitezone (Kaljo et al. 1995). It is clear that spiriferids
100
R. Gourvennec
were not greatly affected by this extinction event,but took advantage of the ecological and / orclimatic conditions following this event asindicated by their rapid radiation.
During the Lower Ludlow only three newgenera are known; Chnaurocoelia in Nevada,Dongbeispirifer and Protoreticularia in North China.All are short lived and restricted to the Ludlow.A significant number of genera (7) do not crossthe Wenlock-Ludlow boundary includingAcanthospirifer, Eohowellella and Pavdenia inCentral Asia-Urals-Russian platform. It shouldalso be noted that Quadrithyris and Tannuspiriferhave not been reported from this area at this time(but do reappear later). In Australia, Endospiriferand Vadum become extinct and Reticulariopsis isabsent. In Bohemia, Rufispirifer and Plicocyrtiadisappeared, and Araspirifer and Spurispirifer arenot reported before the Upper Ludlow.Elsewhere, taxa remained stable or expand intonew areas (e.g. Alaskospira arrived in Bohemiaand North China, Delthyris and Havlicekia inNorth America).
The main tendencies of this relatively stableperiod continued during the upper Ludlow andPridoli with, respectively for each period, only one
90
80
70
60
50
40
30
20
10
_ Total number of genera
Total number of species___ Number of new species
-4J- Number of extinct species
-0- Number of new genera
.~IQ<:A IQ<:A IQ<:A ~ ~ ~ ~ &~ ~o ~o ~o ~o i,,~q:' o-s. o-s. -s.
'?' ~t3: ~'t3 ?:P ~IQ~ ~IQ~ 'vv 'vv «..::>'If ..::>'If 'lt~ ~ .
v~~'V ~' v~~· 'v0
~' ~c' R' 'v0
'v0 v~
Figure 4 Evolution of the number of spiriferid genera and species during the late Ordovician-Silurian period,
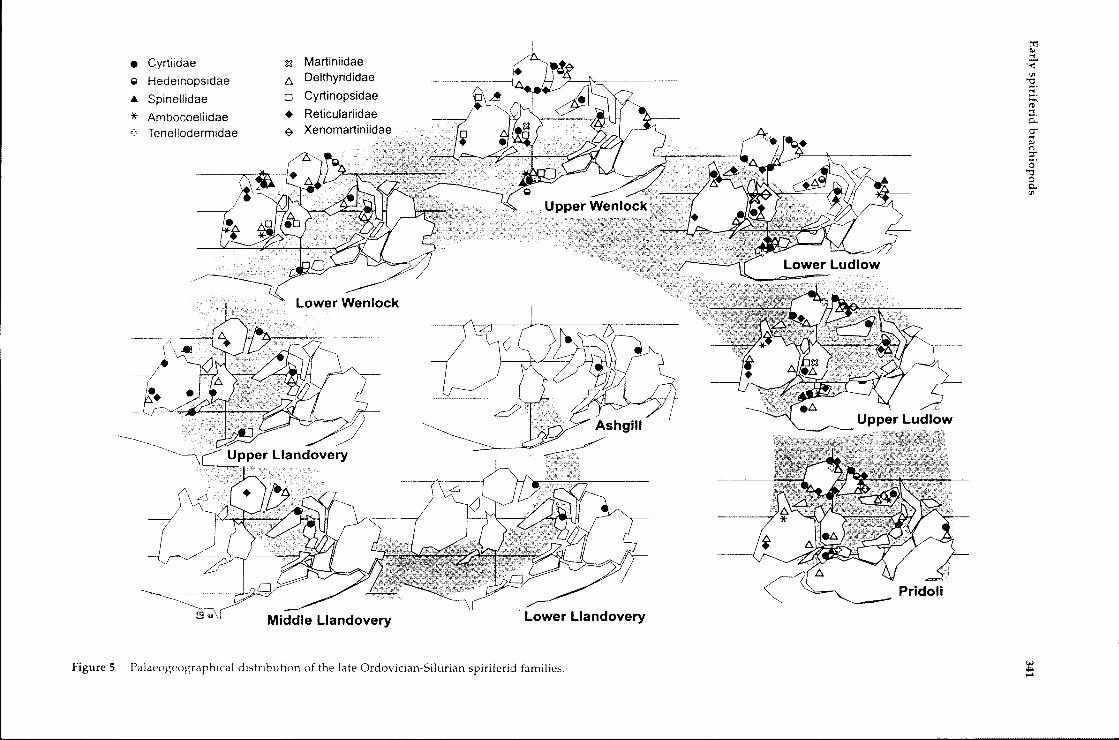
• Cyrtiidae
Q Hedeinopsidae
• Spinellidae
* AmbocoeliidaeSi Tenellodermidae
(\\
=«.,
[9 ..
~ Martiniidael:. Delthyrididae
o Cyrtinopsidae
ReticulariidaeXenomartiniidae
Middle L1andovery . Lower Llandovery
t'T'I....,-.<
er>
'":1.'""11)
::1.0..cr"..,.."::ro·'"o0..
er>
Figure 5 Palaeogeographical distribution of the late Ordovician-Silurian spiriferid families. w~....

342
new taxon (Altajella, Ludlow, Salair; Grebenella,Pridoli, Russian Arctic) and three extinct taxa(Hedeinopsis, Nurataella, Protoreticularia, UpperLudlow; Araspirifer, Chnaurocoelia, Dongbeispirifer,Pridoli). The next radiation event occurred at theSiluro-Devonian boundary.
From these observations it is possible toreconstruct a preliminary model of the distributionof the first spiriferids. The group appeared in SouthChina and then followed a migration route viasouth central Asia (Kazakhstan), Europe and NorthAmerica. This counterclockwise colonisation isalmost completely restricted to the tropical belt.Later migrations occurred in many directions. Themajority of migrations clearly occurred in the UpperLlandovery (Figure 5). Diversity increased slowlyduring the Ashgill and Early Silurian, became morerapid up to the upper Wenlock, and finallydecreased until the Pridoli.
STRATIGRAPHIC DISTRIBUTION OF GENERA
Figure 6 shows that significant gaps occur in thestratigraphic distribution of early spiriferids. Thisphenomenon, known as the «Lazarus effect», mayhave different causes. Firstly, misidentification mayoccur when a taxon is poorly defined and/ordifficult to identify. This may be the case forNikiforovaena, for which the earliest occurrences arequestionable. For some Lazarus taxa lack ofsampling in poorly accessible areas may haveaffected the stratigraphic distribution. On the otherhand, some well defined taxa are apparentlymissing in some strata. This is true of Myriospirifer,which is recorded in the Upper Llandovery of theBaltic area and Lower Wenlock of Bohemia(Havlicek 1995). However, the next occurrence ofMyriospirifer is in Pragian sequences from Brittany(Gourvennec 1989) and possibly Moravia (Havlicekand Mergl 1990). The genus becomes more commonduring the Emsian in Bohemia, the Carnic Alps,Harz Mountains, and North Africa. Such a disjunctstratigraphic pattern (Upper Wenlock- Pragian) ispuzzling and cannot be satisfactorily explained atpresent. The traditional explanation (migration) isunlikely in this case. A similar situation prevailswith Reticulariopsis for which no occurrence areknown in the Late Silurian. Nevertheless, theunique lower Ludlow occurrence in Australia hasbeen verified (Gourvennec 1994).
In terms of evolutionary relationships, thissituation potentially results in serious problems ifthe «missing taxa» are not properly identified. Thefamily Cyrtinopsidae, for instance, has no knownrepresentative during the Lower Ludlow despitethe cosmopolitan distribution of Kozlowskiellina inthe upper Wenlock, so that the origin of theyounger genera belonging to this family remainsproblematic. At this stage, the family
R. Gourvennec
Ambocoeliidae similarly has no knownrepresentative during the Upper Wenlock. It couldbe that future studies, both on new collections andthe re-study of old collections, might fill the gaps.Otherwise it is unlikely that genera separated bywide geographic and/or time gaps may have beengenetically closely related. For example thesupposed links between Kozlowskiellina andMegakozlowskiella need to be revisited. The lastknown Kozlowskiellina is reported from the lowerLudlow of Gotland (Bassett and Cocks 1974) andthe first Megakozlowskiella appears in North America(and perhaps Salair) in the lower Lochkovian. Thisdisjunct pattern in both time and space precludes adirect link between the two taxa. A potential linkcould be represented by Araspirifer, but this genus isneither close to Kozlowskiellina nor toMegakozlowskiella but rather recalls Howellella inexternal morphology.
PROVINCIALISM AND COSMOPOLITANISM
Some of the cosmopolitan genera require moredetailed inquiry. Examination of the distribution ofgenera for each geological period, indicates thatsome taxa may temporarily become endemic e.g.Nikiforovaena, Kozlowskiellina or Alaskospira (Figure6). This may be due to either biological orpalaeogeographical reasons: the migration ordisappearance of a taxon in several areas may bethe result of unsuitable environmental conditionsand this may affect the distribution of benthiccommunities. It is well known that periods ofprovincialism alternate with periods ofcosmopolitanism (e.g. Boucot et al1967, 1969; Talent1972; Boucot 1983) and these fluctuations arerelated to major geological events. Only a fewgenera are not affected by global changes and theterm «cosmopolitan» should be restricted to generathat remain widespread both in time and space, i.e.during their entire lifetime, except the periodimmediately following their origination. In terms ofSilurian spiriferids, this represents about one thirdof the total number of genera for each period(Figure 6). This number is subject to revision sinceseveral endemic genera might represent minorvariations of more widely distributed genera and/or be identified as synonyms when furtherinvestigations are undertaken.
For example, if the distribution of Eospirifer isconsidered, it is obvious that its geographicalexpansion is constant from middle Ashgill toupper Wenlock. But, from the lower Ludlow, thegenus declined until its range was almost reducedto its original area of distribution in the LowerSilurian (South-eastern and Central Asia).Eospirifer finally became extinct in the LowerDevonian. However, there may be a gap in ourpresent knowledge of this genus. Eospirifer is

------------------------------------------ - -~-
Early spiriferid brachiopods 343
Ilvadum ~J i i [
Eo~ticul8ri8 ~~.. 1tt_d'~.Tmm_.A~~~Ni~.:Ii~'
Alaskospire i .'_$4__
ReucuJaoopsis ~m I '-'i~Protoreticu/aris' !
Reticulariidae
Cyrtinopsidae
XenomartiniidaeAfta!ell.
C ,. ,.~,.
~w,. ~I: !c: ,.,. "c&.~ Co c"
~~0." =..~ e,~ h". 0"
~ ~ . < • o. o..~ .. .~ n ~ ~ ~ ~ ~
.:< .:< .:<,..
Delthyrididae
Tenellodermidae
Cyrtiidae
Ambocoeliidae
Eospirifer
Yingwuspirifer
II
Number of species
_Endemic- « 3 regions)
Cosmopolitan- (~3 regions)
--No record
Figure 6 Diagram the stratigraphical distribution, number of species and level of endemism of the Silurianspiriferid genera.
-------

344
recorded in the upper Wenlock and upper Ludlowof North-western America but is apparently absentduring the lower Ludlow in this area. This is eithera sampling artefact or represents the real declineof the genus. The same pattern is apparent for theoccurrences in Turkey (upper Llandovery),Bohemia (upper Wenlock) and the Ibarmaghiandomain (upper Ludlow). In Bohemia and theIbarmaghian domain sampling bias cannot besuspected, but little is known concerning theTurkish domain. On the other hand, this situationmay reflect difficulty for Eospirifer to migrate alongthe North Gondwanan margin. According to mostreconstructions based on different groups ofinvertebrates or micro-organisms (e.g. Bergstrom1990; Rickards et al. 1990), the palaeocurrentsalong the northern margin of Gondwana circulatedin a counterclockwise direction, preventingmigration of larvae from east to west.Furthermore, during the greater part of theSilurian, this region was located beyond thetropical belt (300 5) and thus did not represent asuitable climatic environment for the developmentof spiriferids.
EVOLUTIONARY TRENDS
The relationships between the very earlyspiriferids have been discussed by Rong andZhan (1996). The first descendants of Eospirifer,namely Yingwuspirifer orientalis Rong and Xu 1974and "Nikiforovaena" primo,dialis Rong and Yang1981 are very closely related and are also close toEospirifer praecursor Rong et al. 1994. All thesespecies display a "primitive" morphology:reduced width (longitudinally extended tosubcircular shells), fold sharply delimited bydeep "sulci", resulting in a parasulcatecommissure and indicating a tendency toplication of flanks (e.g. Rong and Yang 1978, pI.1, fig. 8; Rong and Yang 1981, pI. 22, fig. 10 and12; Rong et al. 1994, fig. 9-12). I follow here theseauthors designation of the species primordialis to"Nikiforovaena" with quotation marks. Carter et al.(1994) only considered upper Silurian (Pridoli)and Devonian species and thus recognised thepresence of a ctenophoridium in Nikiforovaena.However, this structure is not present in "N".primordialis (cf. Rong and Yang 1981, fig. 59) andit seems that all Silurian species assigned to"Nikiforovaena" lack a ctenophoridium. Moreover,Nikiforovaena sensu stricto possess plicae on theflanks. So it seems likely that the Silurian"Nikiforovaena" belongs to another (probably new)genus. As far as is known, the type species ofNikiforovaena, N. ferganensis (see Nikiforova 1937)has not been revised and the inner structure ofthe dorsal valve is not known.
Spurispirifer (Pinguispiriferinae) and Tenellodermis
R. Gourvennec
(Tenellodermidae) are very close both in externalaspect and internal structure. They differ only inmicro-ornament. Havlicek (1971) suggested theexistence of a common ancestor for these twogenera. If only the external morphology isconsidered, Howellella may have been a possibleancestor, but the "advanced" aspect of its microornament precludes such a relationship. A commonancestor among the Cyrtiidae is more likely andStriispirifer with its "howellelliform" morphologyand lack of a ctenophoridium, may be a possiblecandidate.
Another evolutionary tendency in earlyspiriferids is the gradual loss of the fold/sulcus(Espella) accompanied by the emergence of complexinternal structures (cruralium). This stock isprobably by neotenic development, at the origin ofthe Ambocoeliidae, characterised by aberrant orcomplex structures, uncommon among otherspiriferids (e.g. reduction of interareas, bisulcatecommissures, often accompanied by the strong ortotal loss of crural or dental plates, etc.).Concurrently, the acquisition of a rudimentarydelthyrial plate in Nurataella is accompanied by anearly flat dorsal valve reminiscent of theambocoelids. A common ancestor with Espella mayhave been, by neotenic development, at the originof Eoplicoplasia and its descendants. Differentialdevelopment of the dorsal and ventral valves alsoexists in the lineage that leads from Hedeina to theAraspiriferinae, with a marked elevation of theventral interarea. This development reaches a peakin the Cyrtia-Plicocyrtia lineage. Nevertheless in thelatter case, the dorsal valve does not become flat orconcave as observed in the ambocoeliids.
Finally, Nurataella and Dongbeispirifer are close tothe ambocoelids and further study, particularly ofthe poorly known genus Dongbeispirifer, willperhaps result in referral to the Ambocoeliidae. Asmentioned above, Striispirifer is the probableancestor of Tenellodermis and Spurispirifer. It is alsothe ancestor of the plicate cyrtiids, by theacquisition of a well delimited, elevated interarea(Hedeina), then a rudimentary (Mictospirifer) or moredeveloped ctenophoridium (Havlicekia). In thislineage there is a tendency to reduce the lateralplications (Havlicek 1980).
The Cyrtia branch is characterised by a very high,hemipyramidal ventral valve. Such a morphologyappears sporadically and independently in diversefamilies during the evolution of the spiriferids. It iscommonly associated with a peculiar mode of life:shell freely reposing on the substrate by its ventralinterarea, suggesting a quiet environment. Thepresence of lateral plicae, that give rise to Plicocyrtia,is also repeated many times in spiriferid evolution.The relationship between Eospirifer and Cyrtiareinforces the "enigma" of the first appearance ofCyrtia. Cyrtia appears in the upper Llandovery of
~------

Early spiriferid brachiopods
the North American-North European domain whenEospirifer was formerly absent in this area. Eospirifermust have arrived in the region at this time andimmediately diversified to give Cyrtia. It is worthnoting that at the same time, and in the samedomains, Myriospirifer and Janius appeared. It isclear that the environmental conditions in the NorthAmerican-North European domain were veryfavourable for diversification at this time. Strongcompetition was prevalent resulting in a significantincrease in species disappearance on a global scale(Figures 4 and 5).
As noted previously by HavHcek (1980), thepresence of a particular micro-ornament in thegenus Myriospirifer is not of critical importance inthe systematics and evolution of the spiriferids sinceit has only been identified in unrelated genusHavlicekia. Apart from ornament, the genusMyriospirifer does not represent notableevolutionary progress compared to Eospirifer.Despite its long duration, Myriospirifer (still presentin the Devonian) has no known descendants. Janiusis characterised by the existence of secondary costaeand by the presence of ribs on the fold/sulcus insome species. Here again the genus (known in theEmsian) is long ranging but did not apparently giverise to a new phyletic branch during the Silurian.
Araspirifer and related genera (e.g. Kozlowskiellina)evolved an external morphology close to Hedeina,including an elevated interarea (the type species ofAraspirifer was first assigned to Hedeina). ButAraspirifer possesses a frilly ornament suggesting arelationship with the delthyridids, particularlyHowellella. Following the observations of Ivanova(1973), the appearance of free rhythmic growthlamellae corresponds to a fundamental change inthe evolution of the spiriferids. The Araspiriferinaeare related to the Delthyrididae despite theirmorphologic resemblance to the Cyrtiidae. Exceptfor Araspirifer, all Silurian araspiriferines possesseda ctenophoridium, a feature common to thedelthyridids. The "temporary" loss of thectenophoridium in Araspirifer is somewhatpuzzling, but may simply reflect imperfectpreservation.
The Hedeinopsidae form a small groupcharacterised by the presencce of a delthyrial plate,an important, but not unique, innovation since it isalso found in numerous spiriferid groups. Thisfamily includes two genera (Tannuspirifer andHedeinopsis) and is also characterised by the loss ofcrural plates.
It is possible to consider the hypothesis that acommon ancestor of Howellella and Tannuspirifer,would have been characterised by plicate flanks, thepresence of a ctenophoridium and probably cruralplates shorter than in Eospirifer. However, no suchhas been described at present.
As mentioned above, the presence of regularly
345
spaced (rhythmic) growth lamellae, oftenterminating in spines, constitutes a major feature inthe spiriferid evolution. The first genus bearingsuch an ornament is Howellella. This evolutionarystage coincides with the appearance of thectenophoridium and lateral plications als9 occursduring this early Uandovery period simultaneouslyin Howellella and some cyrtiids (Striispirifer). Thegenus Howellella thus bears many innovations andconstitutes a potential source for innovation in laterdescendants. The origin of Delthyris is characterisedby the development of a ventral median septum. Itis also the ancestor of the RUfispirifer-"Ivanothyris"lineage by the loss of crural plates and apicalthickening in "Ivanothyris" chanxiensis (see Xian inXian and Jiang 1978). It is worth noting that theassignment of this species to Ivanothyris is doubtful.It is probably a homeomorph of Ivanothyris s. s.because all species of this genus are known onlyfrom Devonian series. Another lineage thatdeveloped from Howellella is distinguished by anincipient sinal plication (Acanthospirifer) and the lossof crural plates (Aldanispirifer).
The last important group of spiriferids is thereticulariids. The oldest genus of this group isEohowellella which is very similar to Howellella butdiffers in its internal features, with the presence of arudimentary spondylium. This makes any directrelation with Howellella highly improbable. Theremaining characters of the genus are poorly knownand its relationship with other genera is difficult toestablish. Eohowellella could be the ancestor of thefamily Xenomartiniidae which includes genera withrelatively complex internal structures. Two Silurianrepresentatives are known; Quadrithyris (with aventral septum) and Altajella (in which the septumand dental plates join to form a supportedspondylium).
Another group originating with Spirinella ischaracterised by the loss of lateral plications andthe fold and sulcus. Vadum is characterised by theloss of the ctenophoridium. This is followed by thedevelopment of a delthyrial plate and/or a strongapical thickening in conjunction with a septalium(Eoreticularia) or, on the other hand, with the loss ofapical plates (Protoreticularia, Chnaurocoelia).Although Alaskospira belongs to another subfamily(Obesariinae), for which it is the single Silurianrepresentative, it is morphologically very similar.
Concurrently, the conservation of the fold/ sulcus,sometimes exaggerated (Pavdenia), may beaccompanied by the neotenic development of lateralplications, first discrete (Reticulariopsis), then welldeveloped (Grebenella) in the late Silurian. Thisgroup of the Reticulariidae originated withSpirinella or from an unidentified common ancestorthat could have been at the origin of theReticularioidea. Many Reticularioidea are endemicwith short stratigraphic ranges: Vadum (Australia),

346
Protoreticularia (N. China; not to be confused withProreticularia), Chnaurocoelia (Nevada).
CONCLUSIONS
Migration pathways of genera, if they can bedeciphered, can provide better understanding ofevolutionary history. Howellella for example, clearlyoriginated in Kazakhstan during the earlyLlandovery, derived seemingly from Eospirifer inthe same region. Two different «strategies» ofexpansion are observed: 1) slow geographicmigration, not necessarily linked with the numberof species (e.g. Eospirifer), and 2) wide and rapidexpansion following emergence (e.g. Howellella).The most spectacular example of the latter is Cyrtia,becoming swiftly ubiquitous in the late Llandovery,a very favourable period for spiriferids. Significantevolutionary events occurred in the tropical belt. Itappears that the spiriferids migrated in a.counterclockwise direction, followingpalaeocurrents of that time.
During Ashgill and Early Silurian, the diversityincreased at a moderate rate until the lateLlandovery when a peak of diversificationoccurred (Figure 4). No major evolutionary eventoccured during the early Wenlock, but a newradiation event occurred in the late Wenlock. Thisevent probably represents the turning point in thehistory of spiriferids. It appears to have resultedfrom the early-middle Homerian bioevent. TheLudlow was a relatively stable period, though asignificant number of genera did not cross theWenlock-Ludlow boundary. Similar conditionsprevailed during the Pridoli. The next crisis andradiation events occurred at the Siluro-Devonianboundary.
Some genera are weakly affected by global events,remaining Widespread both in time and spaceduring their entire life-span (e.g. Howellella). Suchgenera represent about one third of the totalnumber of genera for each specified interval duringthe Silurian. Considering the wide distancesbetween their areas of distribution, and bearing inmind the length of larval life of present-daybrachiopods, the supposed links between somegenera are thought to be unlikely.
A relatively common problem is the «Lazaruseffect» apparent in many genera. It cannot beexplained as merely a reflection of inadequatesampling or misidentification; but is probably theresult of ambiguities in systematics when a familyhas no representatives for a significant timeinterval.
ACKNOWLEDGEMENTS
I am most grateful to Professors John Talent, RuthMawson, Tony Wright and Glenn Brock for their
R. Gourvennec
most helpful comments on and editing of thismanuscript. This is a contribution to IGCP Project421 North Gondwana mid-Palae020ic bioevent/biogeography patterns in relation to crustal dynamics.
REFERENCESBassett M.G. and Cocks L.R.M. (1974). A review of
Silurian brachiopods from Gotland. Fossils and Strata3: 1-56.
Bergstrom, S.M. (1990). Relations between conodontprovincialism and the changing palaeogeographyduring the Early Palaeozoic. In McKerrow W.S. andScotese c.R. (eds), Palaeozoic palaeogeography andbiogeography, Geological Society Memoir 12: 105-121.
Boucot AJ. (1963). The Eospiriferidae. Palaeontology 5:682-71l.
Boucot AJ. (1983). Does evolution take place in anecological vacuum? Journal ofPaleontology 57: 1-30.
Boucot, AM., Johnson, J.G. and Talent J.A (1967). Lowerand Middle Devonian faunal provinces based onbrachiopods. In Oswald, D.H. (ed.), Internationalsymposium on the Devonian sytstem Il, 1239-1254.Alberta Society for Petroleum Geology, Calgary.
Boucot, AM., Johnson, J.G. and Talent J.A (1969). EarlyDevonian brachiopod zoogeography. Special Paper,Geological Society ofAmerica 119: 1-70.
Carter, J.L., Johnson, J.G., Gourvennec, R. and HouHong-fei (1994). A revised classification of thespiriferid brachiopods. Annals of the Carnegie Museum63: 327-374.
Copper, P. and Gourvennec, R. (1996). Evolution of thespire-bearing brachiopods (Ordovician-Jurassic). InCopper, P and Jin J. (eds), Brachiopods, Proceedings ofthe Third International Brachiopod congress, Sudbury,Ontario, Canada, September 1995: 81-88, Balkema.
Fu Li-pu (1982). Brachiopoda. In Paleontological atlas ofNW China, Shaanxi-Gansu-Ningxia volume 1:Precambrian and Early Paleozoic: 95-179.
Gourvennec, R. (1989). Brachiopodes Spiriferida duDevonien inferieur du massif armoricain.Systematique, paleobiologie, evolution,biostratigraphie. Biostratigraphie du PaIeozoique 9: 128l.
Gourvennec, R. (1994). Precisions nouvelles sur le gemeReticulariopsis Frederiks, 1916 (Brachiopoda,Spiriferida). Annales Societe geologique du Nord 3: 12313l.
Grunt, T. (1982). Mikrostruktura rakoviny brakhiopodotryada Athyrida. Paleontologicheskii Zhurna11982: 2135.
Havlfcek, V. (1971). Non-costate or weakly costateSpiriferidina (Brachiopoda) in the Silurian and LowerDevonian of Bohemia. Sbornik geologickych ved,Paleontologie 14: 7-34.
Havlfcek, V. (1980). New Eospiriferinae (Brachiopoda) inBohemia. Sbornik geologickych ved, Paleontologie 23: 748.
Havlfcek, V. (1995). New data on the distribution ofbrachiopods in the Motol and lowest KopaninaFormations (Wenlock, lower Ludlow, Prague Basin,Bohemia). Vestnik Ceskeho geologickeho ustavu 70: 47-63.

Early spiriferid brachiopods
Havlicek, V. and Mergl, M. (1990). Lower Devonianbrachiopods at Petrovice (Drahany Upland, Moravia).Casopis pro Mineralogii a Geologii 35: 225-238.
Ivanova, Ye. A (1973). Main features of spiriferid evolution(Brachiopoda). Paleontological Journal 6: 309-320.
Kalj6, D., Boucot, AJ., Corfield, RM., Le Herisse, A,Koren, T.N., Kriz, J., Mannik, P., Marss, T., Nestor,V., Shaver, RH., Siveter, D.J. and Viira, V. (1995).Silurian bio-events. In Walliser O.H. (ed.), Globalevents and event stratigraphy in the Phanerozoic: 173224, Springer Verlag, Berlin.
Modzalevskaya, T.L. (1990). Ludlovski brakhiopody izgraptolitovykh fatsii Novoi Zemli. EzhegodnikhVsesoyouznogo Paleontologicheskogo Obshchestva 33: 6478.
Modzalevskaya, T.L. and Popov, L.E. (1995). EarliestSilurian articulate brachiopods from centralKazakhstan. Acta Paleontologica Polonica 40: 399-426.
Nikiforova, 0.1. (1937). Brakhiopody verknego Silurasredneasiatskoi casti SSSR Monografii po paleontologiiSSSR 35(1): 1-94.
Rickards, B., Rigby, S. and Harris, J. H. (1990). Graptoloidbiogeography: recent progress, future hopes. InMcKerrow W.S. and Scotese C.R. (eds), Palaeozoicpalaeogeography and biogeography, Geological SocietyMemoir 12: 139-145.
Rong Jia-yu, Boucot, AJ., SU Yang-zheng and Strusz,D.L. (1995). Biogeographical analysis of Late Silurianbrachiopod faunas, chiefly from Asia and Australia.Lethaia 28: 39-60.
Rong Jia-yu, Xu Han-kui and Yang Xue-chang (1974).Silurian brachiopods. In Nanjing Institute of Geologyand Paleontology, Academia Sinica (eds), Handbook ofthe stratigraphy and paleontology in southwest China:195-208, Science Press, Beijing. (in Chinese)
Rong Jia-yu and Yang Xue-chang, (1978). Silurianspiriferoids from southwest China with specialreference to their stratigraphic significance. ActaPalaeontologica Sinica 17: 357-386.
Rong Jia-yu and Yang Xue-chang (1981). Middle andLate Early Silurian brachiopod faunas in Southwest
347
China. Memoirs of Nanjing Institute of geology andPalaeontology, Academia Sinica 13: 163-270.
Rong Jia-yu and Zhan Ren-bin (1996). B'rachidia of LateOrdovician and Silurian Eospiriferines (Brachiopoda)and the origin of the spiriferides. Palaeontology 39:941-977.
Rong Jia-yu, Zhan Ren-bin and Han Nai-ren (1994). Theoldest known Eospirifer (Brachiopoda) in theChangwu Formation (Late Ordovician) of WesternZhejiang, East China, with a review of the earliestspiriferoids. Journal ofPaleontology 68: 763-776.
Sapelnikov, V.P. and Rukavischnikova, T.B., (1975).Verkhneordovikskie, Siluriiskie i nijnedevonskiepentameridy Kazakhstana: 1-227, Nauka.
Scotese, C.R and McKerrow, W.S. (1990). Palaeozoicpalaeogeography and biogeography. Geological Soci.etyofLondon Memoir 12: 1-435..
Sheehan, P.M. and Baillie, P.W. (1981). A new species ofEospirifer from Tasmania. Journal of Paleontology 55:248-256.
Strusz D.L. (1985). Brachiopods from the Silurian ofFyshwick, Canberra, Australia. Bureau of MineralResources, Journal of Australian Geology and Geophysics9: 107-119.
Talent, J.A (1972). Provincialism and Australian EarlyDevonian faunas. Journal of the Geological Society ofAustralia 19: 81-97.
Wright, AD. (1979). The origin of the spiriferidinebrachiopods. Lethaia 12: 29-33.
Xian Si-yuan and Jiang Zong-Iong (1978): PhylumBrachiopoda. In Palaeontological Atlas of SouthwestChina, Guizhou Volume1 (Cambrian to Devonian): 251337, Geological Publishing House, Beijing. (inChinese)
Zhang, N. (1989). Wenlockian (Silurian) brachiopods ofthe Cape Phillips Formation, Baillie Hamilton Island,Arctic Canada. Part 1. Palaeontographica, Abt. A 206:49-97.
Manuscript received March 1999; accepted October 1999.





























