Embed Size (px)
Citation preview

INVITED REVIEW
The force-from-lipid (FFL) principle of mechanosensitivity,at large and in elements
Jinfeng Teng & Stephen Loukin & Andriy Anishkin &
Ching Kung
Received: 10 April 2014 /Revised: 29 April 2014 /Accepted: 5 May 2014# Springer-Verlag Berlin Heidelberg 2014
Abstract Focus on touch and hearing distracts attention fromnumerous subconscious force sensors, such as the vital controlof blood pressure and systemic osmolarity, and sensors innonanimals. Multifarious manifestations should not obscureinvariant and fundamental physicochemical principles. Weadvocate that force from lipid (FFL) is one such principle. Itis based on the fact that the self-assembled bilayer necessitatesinherent forces that are large and anisotropic, even at life’sorigin. Functional response of membrane proteins is governedby bilayer force changes. Added stress can redirect theseforces, leading to geometric changes of embedded proteinssuch as ion channels. The FFL principle was first demonstrat-ed when purified bacterial mechanosensitive channel of largeconductance (MscL) remained mechanosensitive (MS) afterreconstituting into bilayers. This key experiment has recentlybeen unequivocally replicated with two vertebrate MS K2p
channels. Even the canonical Kv and the Drosophila canoni-cal transient receptor potentials (TRPCs) have now beenshown to be MS in biophysical and in physiological contexts,supporting the universality of the FFL paradigm. We alsoreview the deterministic role of mechanical force during stemcell differentiation as well as the cell-cell and cell-matrixtethers that provide force communications. In both the ear hair
cell and the worm’s touch neuron, deleting the cadherin ormicrotubule tethers reduces but does not eliminate MS chan-nel activities. We found no evidence to distinguish whetherthese tethers directly pull on the channel protein or a surround-ing lipid platform. Regardless of the implementation, pullingtether tenses up the bilayer. Membrane tenting is directlyvisible at the apexes of the stereocilia.
Keywords Force-sensing .Mechanosensitivity .
Lipid bilayer . K2p. Touch . Hearing
Introduction
“Never use the words higher or lower”—CharlesDarwin [21].
For this special issue, the editors’ instruction is as follows:“Encouraged is the presentation of the author’s personal pointof view in the context of current literature, controversies etc.”We comply. A small portion here covers our recent work, butthe bulk deals with broader issues raised by current literatureat large and in elements. By “at large,” we mean to reviewforce sensors beneath human consciousness, in plants andmicrobes, inside or between cells, and during development.This coverage is intended to balance the emphasis on touchand hearing in current research, reflected in this special issue.By “in elements,” we mean to assert that the force-from-lipid(FFL) paradigm applies to both pro- and eukaryotic ionchannels and applies even to channels traditionally classifiedas voltage-gated or ligand-gated. We argue that FFL is uni-versal. We view the tethers for “higher” mechanosensors asaccessories, necessitated by multicellularity. Ultimately, allchannels work at a “lower” level obeying the physics of lipidbilayers.
J. Teng : S. LoukinLaboratory of Molecular Biology, University of Wisconsin,Madison, WI 53706, USA
A. AnishkinDepartment of Biochemistry and Center for ComputationalProteomics at the Huck Institute of Life Sciences, Pennsylvania StateUniversity, University Park, State, College, PA 16802, USA
C. Kung (*)Laboratory of Molecular Biology and Department of Genetics,University of Wisconsin, Madison, WI 53706, USAe-mail: [email protected]
Pflugers Arch - Eur J PhysiolDOI 10.1007/s00424-014-1530-2

The gears below deck
Being conscious sensations,touch and hearing are near anddear to us. Far more vital, however, are the mechanical forcesensors below our consciousness.
Blood pressure is among the first measures taken when onevisits a western clinic. Heartbeats and the resulting pressureare too essential to be put under the whim of consciousness.Blood pressure is subjected to baroreflex, a rapid negativefeedback that makes short-term adjustments. The sensors,called “baroreceptors,” are not molecules but aremechanosensitive (MS) nerve endings that are excited by thestretch of major arteries. Despite its importance and the hugemedical literature on long-term implications of blood pressure,molecular studies on how stretch excites baroreceptor are rare.Acid-sensing ion channel 2 (ASIC2) of the DEG/epithelialsodium channel (ENaC) superfamily is expressed in barore-ceptors and their ganglia. ASIC2 knockout mice show poorbaroreflex and suffer hypertension but live [56]. Transientreceptor potential vanilloid 1 (TRPV1) appears to beexpressed in all parts of the baroreceptive neuron and phar-macological blockage of TRPV1 blunts baroreflex [89].Others found task-1−/− mice to have heart symptoms andelectrophysiology consistent with a diminished baroreflex[76]. There is no consensus at the moment on which chan-nel(s) conduct the MS receptor current(s) of baroreceptors.
Stretching smooth muscles causes depolarization and con-traction, observed in arteries and arterioles that supply skin,lungs, bladder, stomach, intestines, etc. This myogenic re-sponse, also known as the Bayliss effect, has been examinedin assorted arteries, but the force sensor(s) has not beenidentified. An MS cation conductance on many mammaliancells is well known [27] but is of unknown molecular identity.Canonical TRP 1 (TRPC1), TRPC6, TRPM5, TRPV4, ENaC,and Cl− channel have all been implicated [6]. See Sharif-Naeini et al. for a critical review [85].
Further below conscious attention is the homeostasis ofsystemic osmolality. Humans maintain extracellular fluid at288 mOsm/kg, a set point that is strictly limited to within±1 %. Deviations cause problems from headaches to death.Osmotic homeostasis, found in all vertebrates and in insects,evolved long before homeothermy. This early appearance inanimals reflects the importance of osmolarity and membraneturgor. In vertebrates, plasma osmolality is gauged by“osmoreceptors” in the hypothalamus. Again, these are notmolecules but neuron clusters. Plasma hyperosmolarity depo-larizes these neurons to discharge, causing the release of theantidiuretic vasopressin (VP) [84, 9]. Such neurons fire whenthe bath is made hypertonic or when a deflating suction isapplied. Cell-attached patches showed a cation-nonspecificunitary conductance that is stretch-inactivated [71]. Some ofthese neurons express a shorter N-terminal splice variant ofTRPV1. TRPV1−/− mice show serum hyperosmolarity and
poor VP responses to osmotic challenge, and their neuronsrespond to hyperosmolarity poorly [83]. Nonetheless, thesemice survive and mature, showing that TRPV1 cannot be thesole sensor for systemic osmolality. There are evidences in-cluding those from TRPV4−/− mice [49] that indicate the par-ticipation of other TRPV channels [84]. Some osmoreceptorneurons also express a variety of two-pore domain K+ channels(K2p’s)[9].
VP regulates the function of the kidney, where plasma isfiltered into urine across the slit diaphragm between podocytes(glomerular epithelial cells) [79]. Podocytes express compo-nents akin to those discovered in the genetic dissection oftouch in Caenorhabditis elegans. An essential component ofthe worm MS channel complex is MEC-2, an SPFH domainmembrane protein that recruits cholesterol [25]. Podocytesexpresses its counterpart, podosin, which complexes withthe TRPC6 channel [35]. The latter is activated by hypo-osmolarity or by prodding [2]. Cholesterol-rich thickenedlipid domain, partly constructed with these SPFH domainproteins, has been proposed to provide proper platforms tostage MS channels [3]. SLP3−/− mice, lacking one of theseSPFH proteins, show very poor touch sensation [93], muchlike the MEC-2−/− worm.
It takes an astronaut 3 years to gain back the bone lost in a3-month space flight. Mature bones constantly measure loadto remodel themselves, a remarkable type of mechanical forcesensing, though not in the timescale of hearing and touch.Unloading osteoporosis places bed-ridden patients at risk ofbone fracture. Unloading-induced bone loss is suppressed inTRPV4−/− mice [64]. Over 50 alleles of TRPV4 mutations arenow known to cause developmental bone diseases rangingfrom dwarfism to neonatal or prenatal death [68]. We haveshown that TRPV4 responds to hypo-osmolarity whenexpressed in yeast [55] and opens directly to stretch force inpatches excised from expressing Xenopus oocytes [54].Disease mutations seem to all be gain-of-function alleles,meaning that the mutant channels have higher basal openprobability (Po) than that of the wild type. Further, the severityof the bone disease appears to parallel the basal Po, indicatingthat channel leakage and Ca2+ poisoning may be the cause[53]. The mechanism of how bone senses load remains un-clear. TRPV4 appears to have a role in the maturation ofosteoclasts [61]. A major puzzle in this field is the fact thatTRPV4−/− mice, with a complete loss of TRPV4 function,develop almost normally.
The FFL paradigm
Besides those above, the monitored forces in our body alsoinclude shear stress on ciliated epithelia, intraocular pressure,let alone those in the electromechanical feedbacks of thepounding heart, the peristalsis of the digestive tract, and many
Pflugers Arch - Eur J Physiol
more. Beyond the obvious, physical forces also play para-mount roles during development and cell-cell communication(below). Further, microbes and plants display their own formsof mechanosensitivity (below). Mechanosensitivity is as per-vasive as it is mysterious, peculiarly so in comparison to ourdeep knowledge of ligand and light sensing. Manifest diver-sities, however, should not obscure the common fundamentalprinciples behind. All inheritance, from pea seed color tosickle cell anemia, traces back to DNA. The H+-gradient-based chemiosmotic theory unifies oxidative phosphorylationand photosynthesis, which energize all creatures, pink orgreen. Occam’s razor aside, we believe that there is a funda-mental unifying principle for the myriad force sensations.
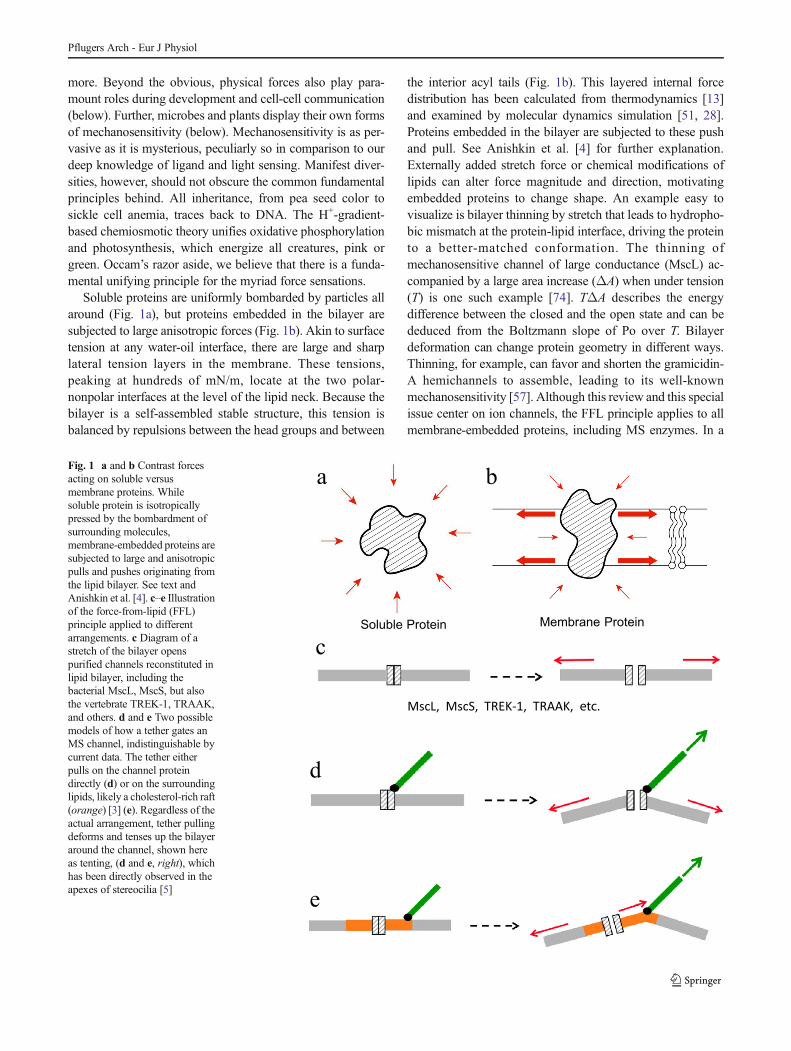
Soluble proteins are uniformly bombarded by particles allaround (Fig. 1a), but proteins embedded in the bilayer aresubjected to large anisotropic forces (Fig. 1b). Akin to surfacetension at any water-oil interface, there are large and sharplateral tension layers in the membrane. These tensions,peaking at hundreds of mN/m, locate at the two polar-nonpolar interfaces at the level of the lipid neck. Because thebilayer is a self-assembled stable structure, this tension isbalanced by repulsions between the head groups and between
the interior acyl tails (Fig. 1b). This layered internal forcedistribution has been calculated from thermodynamics [13]and examined by molecular dynamics simulation [51, 28].Proteins embedded in the bilayer are subjected to these pushand pull. See Anishkin et al. [4] for further explanation.Externally added stretch force or chemical modifications oflipids can alter force magnitude and direction, motivatingembedded proteins to change shape. An example easy tovisualize is bilayer thinning by stretch that leads to hydropho-bic mismatch at the protein-lipid interface, driving the proteinto a better-matched conformation. The thinning ofmechanosensitive channel of large conductance (MscL) ac-companied by a large area increase (ΔA) when under tension(T) is one such example [74]. TΔA describes the energydifference between the closed and the open state and can bededuced from the Boltzmann slope of Po over T. Bilayerdeformation can change protein geometry in different ways.Thinning, for example, can favor and shorten the gramicidin-A hemichannels to assemble, leading to its well-knownmechanosensitivity [57]. Although this review and this specialissue center on ion channels, the FFL principle applies to allmembrane-embedded proteins, including MS enzymes. In a
Soluble Protein Membrane Protein
MscL, MscS, TREK-1, TRAAK, etc.
a b
c
d
e
Fig. 1 a and b Contrast forcesacting on soluble versusmembrane proteins. Whilesoluble protein is isotropicallypressed by the bombardment ofsurrounding molecules,membrane-embedded proteins aresubjected to large and anisotropicpulls and pushes originating fromthe lipid bilayer. See text andAnishkin et al. [4]. c–e Illustrationof the force-from-lipid (FFL)principle applied to differentarrangements. c Diagram of astretch of the bilayer openspurified channels reconstituted inlipid bilayer, including thebacterial MscL, MscS, but alsothe vertebrate TREK-1, TRAAK,and others. d and e Two possiblemodels of how a tether gates anMS channel, indistinguishable bycurrent data. The tether eitherpulls on the channel proteindirectly (d) or on the surroundinglipids, likely a cholesterol-rich raft(orange) [3] (e). Regardless of theactual arrangement, tether pullingdeforms and tenses up the bilayeraround the channel, shown hereas tenting, (d and e, right), whichhas been directly observed in theapexes of stereocilia [5]
Pflugers Arch - Eur J Physiol

membrane, essentially any change in protein shape will exactsome energetic price, as some parts will move along or againstthe strong components of the pressure/tension profile.Therefore, any external condition that change the lipid forceswill change the energy cost of the conformational change,rendering membrane proteins MS to various extent, physio-logically relevant or not.
We have also argued that life’s origin requires enclosingRNA, etc. with an envelope of amphipaths, likely abioticlipids. Even this primordial bilayer comes with a set of phys-ical properties, including its anisotropic forces describedabove [4]. In the capricious primordial soup, one of its earlyfunctions is likely to deal with the fluctuation in the concen-tration of water, the solvent of life. Thus, stretching the bilayeris likely an “original sense” [4] and a mechanism that has beencontinuously employed throughout evolution. Perhaps not aserratic, but environmental water concentration still continuallychanges today. Water concentration is measured by the totalparticle bombardment on the bilayer, i.e., the osmotic force.As water being the universal biochemistry solvent, sensing ofosmotic and other forces can be said to be “solvent senses” asopposed to the “solutes senses” that measures ligands by lock-and-key bindings [45].
The FFL principle was established some 25 years ago. Thisbegan in the first patch-clamp survey of E. coli membrane,when Boris Martinac encountered large MS unitary conduc-tances [60] and also found them activated by amphipaths [59].Eventually, purifiedMscLwas reconstituted into lipid bilayersand found to retain its mechanosensitivity [88]. As an exerciseof ultimate reductionism, this experiment excludes any tetherproteins or other channel subunits, leaving the bilayer as theonly source for the forces that gate MscL. In fact, the MscLprotein was isolated by tracking the reconstituted MS activityin column fractions starting from E. coli lysate, much likeenzyme purification [87]. Much crystallographic, biochemi-cal, genetic, and biophysical work onMscL and its analogMSchannel of small conductance (MscS) followed, as reviewedby Cox et al., by Rees and their coworkers in this issue andelsewhere. However, there has always been the notion thatgating directly by bilayer stretch is restricted to the “lowly”bacteria. A current review [22] states “Two primary modelshave been proposed for mechano-gating: the lipid bilayerstretch model evidenced by microbial MS channels and themore sophisticated tether model of eukaryotes by whichtethers pull open the transduction channel.” Before we dealwith the “more sophisticated tether” below, we wish to pointout that the FFL principle also applies to animal channels.Membrane blebs, apparently devoid of cytoskeleton, retain theactivities of the native MS channels in Xenopus oocyte [30,95]. Doping membrane with various amphipaths activatessuch MS channels [59, 34]. Lipids with polyunsaturated fattyacids (PUFAs) enhance and are likely required for theC. elegans touch response [92]. Dissociating subcortical
cytoskeleton liberates rather than inhibits MS channels (seebelow). Nevertheless, these experiments were performed oncomplex cell membranes. It is again the ultimate reductionismof reconstituting purified animal MS channels below thatproves the broad, if not universal, application of the FFLprinciple.
The “proto-behavior” of "eu-channels"
Enriched fractions of NMDA receptor channel [43] or TRPC1[58] have been reconstituted showing mechanosensitivity.While we await replications and extensions of these works,two recent reports [8, 12] demonstrated incontrovertibly thattwo-pore-domain K+ channels (K2p’s) retained theirmechanosensitivity upon reconstitution into bilayers, likeMscL and MscS. The molecular biology and electrophysiol-ogy of the K2p TREK-1 (TWIK-related K+-1) and its homologTRAAK (TWIK-related arachidonic acid-stimulated K+) havebeen extensively studied by Patel, Honoré, and coworkers [73,34], laying a solid foundation for further investigation. TREK-1 current is suppressed by hyperosmolarity and activated bypressure in either direction, and by acidic phospholipids,lysophospholipids, PUFAs, or volatile general anesthetics[73, 34]. Atomic structures of two K2P’s [63, 11, 10] arenow known. TRAAK has features not found in KcsA, Kv,Kir, GIRK, or BK. It has a twofold symmetry, and its secondtransmembrane helix from each of the two subunits is long,with a kink that makes the inner portion lying almost flat. Thisportion is amphipathic, placing the two peptides at themembrane-cytoplasm interface of the inner leaflet, rich inacidic lipids in vivo. Interestingly, the inner transmembranehalf of the K2P channel is fenestrated. A sizable portion ofTRAAK’s inner half is not enclosed by protein, leaving a 5-Å-wide gap extending from the bottom of the filter to thelower edge of the channel, presumably filled with lipidsin vivo [11].
Berrier et al. [8] have recently reconstituted an enrichedfraction of the mouse TREK-1 into liposomes and examinedpatches excised from them. They observed the ~80-pS K+-specific outward-rectifying chlorpromazine-sensitive conduc-tance. Surprisingly, these channels displayed spontaneousactivities that could not be further increased by pipet suctions.Positive pipet pressure pulses, however, could proportionallyclose the channels. These authors reasoned that Po has beenmaximized by inherent tension in the patches [8]. Such atension is likely generated during the gigaohm seal formation,bonding the bilayer lipids to the pipet glass surface [72, 81,80] (see below).
In a recent rigorous study, Brohawn et al. [12] purifiedzebrafish TREK-1 and human TRAAK to homogeneity, asevidenced by the monodispersion of column elution profiles(Fig. 2a) and also reconstituted them into liposomes made of
Pflugers Arch - Eur J Physiol
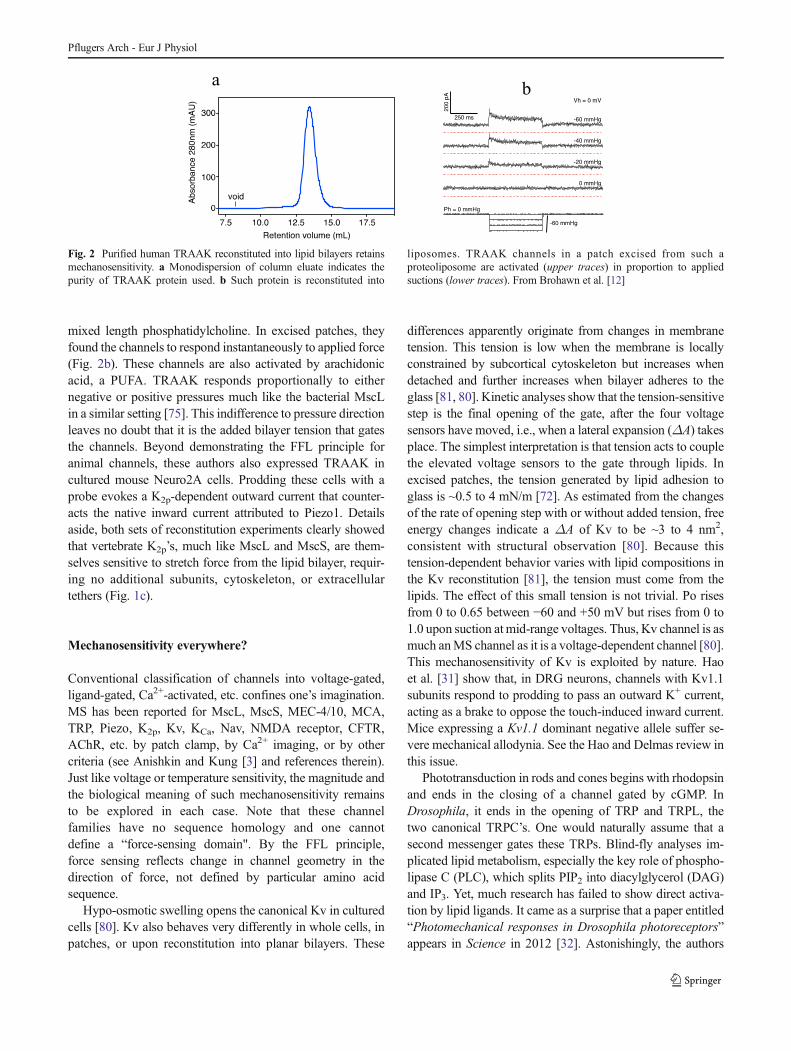
mixed length phosphatidylcholine. In excised patches, theyfound the channels to respond instantaneously to applied force(Fig. 2b). These channels are also activated by arachidonicacid, a PUFA. TRAAK responds proportionally to eithernegative or positive pressures much like the bacterial MscLin a similar setting [75]. This indifference to pressure directionleaves no doubt that it is the added bilayer tension that gatesthe channels. Beyond demonstrating the FFL principle foranimal channels, these authors also expressed TRAAK incultured mouse Neuro2A cells. Prodding these cells with aprobe evokes a K2p-dependent outward current that counter-acts the native inward current attributed to Piezo1. Detailsaside, both sets of reconstitution experiments clearly showedthat vertebrate K2p’s, much like MscL and MscS, are them-selves sensitive to stretch force from the lipid bilayer, requir-ing no additional subunits, cytoskeleton, or extracellulartethers (Fig. 1c).
Mechanosensitivity everywhere?
Conventional classification of channels into voltage-gated,ligand-gated, Ca2+-activated, etc. confines one’s imagination.MS has been reported for MscL, MscS, MEC-4/10, MCA,TRP, Piezo, K2p, Kv, KCa, Nav, NMDA receptor, CFTR,AChR, etc. by patch clamp, by Ca2+ imaging, or by othercriteria (see Anishkin and Kung [3] and references therein).Just like voltage or temperature sensitivity, the magnitude andthe biological meaning of such mechanosensitivity remainsto be explored in each case. Note that these channelfamilies have no sequence homology and one cannotdefine a “force-sensing domain". By the FFL principle,force sensing reflects change in channel geometry in thedirection of force, not defined by particular amino acidsequence.
Hypo-osmotic swelling opens the canonical Kv in culturedcells [80]. Kv also behaves very differently in whole cells, inpatches, or upon reconstitution into planar bilayers. These
differences apparently originate from changes in membranetension. This tension is low when the membrane is locallyconstrained by subcortical cytoskeleton but increases whendetached and further increases when bilayer adheres to theglass [81, 80]. Kinetic analyses show that the tension-sensitivestep is the final opening of the gate, after the four voltagesensors have moved, i.e., when a lateral expansion (ΔA) takesplace. The simplest interpretation is that tension acts to couplethe elevated voltage sensors to the gate through lipids. Inexcised patches, the tension generated by lipid adhesion toglass is ~0.5 to 4 mN/m [72]. As estimated from the changesof the rate of opening step with or without added tension, freeenergy changes indicate a ΔA of Kv to be ~3 to 4 nm2,consistent with structural observation [80]. Because thistension-dependent behavior varies with lipid compositions inthe Kv reconstitution [81], the tension must come from thelipids. The effect of this small tension is not trivial. Po risesfrom 0 to 0.65 between −60 and +50 mV but rises from 0 to1.0 upon suction at mid-range voltages. Thus, Kv channel is asmuch anMS channel as it is a voltage-dependent channel [80].This mechanosensitivity of Kv is exploited by nature. Haoet al. [31] show that, in DRG neurons, channels with Kv1.1subunits respond to prodding to pass an outward K+ current,acting as a brake to oppose the touch-induced inward current.Mice expressing a Kv1.1 dominant negative allele suffer se-vere mechanical allodynia. See the Hao and Delmas review inthis issue.
Phototransduction in rods and cones begins with rhodopsinand ends in the closing of a channel gated by cGMP. InDrosophila, it ends in the opening of TRP and TRPL, thetwo canonical TRPC’s. One would naturally assume that asecond messenger gates these TRPs. Blind-fly analyses im-plicated lipid metabolism, especially the key role of phospho-lipase C (PLC), which splits PIP2 into diacylglycerol (DAG)and IP3. Yet, much research has failed to show direct activa-tion by lipid ligands. It came as a surprise that a paper entitled“Photomechanical responses in Drosophila photoreceptors”appears in Science in 2012 [32]. Astonishingly, the authors
ab
Fig. 2 Purified human TRAAK reconstituted into lipid bilayers retainsmechanosensitivity. a Monodispersion of column eluate indicates thepurity of TRAAK protein used. b Such protein is reconstituted into
liposomes. TRAAK channels in a patch excised from such aproteoliposome are activated (upper traces) in proportion to appliedsuctions (lower traces). From Brohawn et al. [12]
Pflugers Arch - Eur J Physiol

showed that light induces a near-micrometer shrinkage of theommatidia (units of the compound eye, comprisingsupporting and photoreceptor cells), which is directly visibleunder a light microscope or quantified with an atomic forcemicroscope. The contraction precedes the channel current.Deleting the PLC removes the contraction; deleting the chan-nels does not. It is modeled that beheading PIP2 to make thesmaller DAG increases tension in the inner membrane leafletto pull open channels. Gramicidin A [57], the well-establishedMS channel, was added to the photoreceptor cell and found topass current proportional to light intensity. Note that thevertebrate retina also has intrinsically photosensitive ganglioncells that use a homologous pathway for circadian entrainment[36]. The melanopsin there is far more similar to the insectrhodopsin than vertebrate rhodopsin [82]. Melanocyte [7] andkeratinocyte [66] of the skin appear to have similar pathways.Small microvilli, cilia, filopodia, or dendritic spines are com-mon, but unlike the ommatidium with thousands of stackedmicrovilli that house the phototransduction complex, physicalmovement cannot easily be detected in these isolated smallstructures. These structures, however, often have signal-transduction pathways with PLC and channels. The currentdominant paradigm for these pathways ends in protein phos-phorylations due to PLC’s activation of PKC. The PLC-induced changes in bilayer tension reviewed here may in factparticipate in these pathways [50].
The jungle
Our sense of self-importance should not blind us to the vast-ness and richness of the biological world. Humans, verte-brates, and indeed all animals together are only a minor partof the biosphere. Microbes outweigh animals in number, invariety, and in total mass. E. coli is undoubtedly smaller andless complex than Homo sapiens, but it is just as modern andsuccessful. Another common misunderstanding is that mi-crobes are ancient relics and use outmoded machineries.Billion years of divergence have led to different life-styles and therefore different implementations of thebasic molecular ingredients. Yet, the fundamental ma-chineries necessarily evolved early and remain compul-sory for all. Universal is the DNA-RNA-protein infor-mation flow, ATP synthesis and hydrolysis, the lipidbilayers wrapping, etc.
We have pointed out the need to face up to osmotic changeseven for the primordial protocells. Whether it is the osmoticforce or a physical impact, the membrane deforms according-ly. A Newton is a Newton. The sense of touch must haveevolved early since it is seen in both unicellular and multicel-lular forms. Bacterial flagella respond to the external mechan-ical force—slowing flagellum rotation is sensed, possibly asthe change in the charge flow through the rotor [39].
Paramecium, a large eukaryotic unicell, reverses its ciliarybeat when impacted at its anterior end to perform an “avoidingreaction” observed in the nineteenth century [37]. Intracellularrecording shows that impact triggers a Ca2+ action potential inthe cilia [23], which can be erased by deciliation [70] or bygenetic means [78] to reveal a cation-nonspecific touch recep-tor potential originated from the soma. In multicellular ani-mals, primary cilia sense force and are pivotal in development.They also become sensory cilia, including the kinocilia of theinner ear. See review by A. Patel in this issue.
Although some might think that the only reason to studybacteria is to find ways to kill them, microbes in fact contrib-ute to our own survival and to biological research in importantways. Their lifestyle obviates the paraphernalia needed formulticellular existence, which complicates research. For ex-ample, in hearing research, various accessories have beenidentified, but the strenuous and perennial search for the truehair-cell transduction channel remains inconclusive, as is ev-ident from the reviews in this issue.
The entire central dogma was erected from the study ofbacteria and bacterial phages, leading to the slogan “What istrue to E. coli is true to the elephant” during the romanticperiod of molecular biology revolution in the 1950s. Wewould argue that the revolution is not yet over, and somebasic principles are yet to be discovered. Microbes are wide-spread and have explored essentially all organic and inorganicniches on earth where water is available. In so doing, far moremolecular fabrications have been explored by microbes thanby the animals. Their wide expeditions resulted in materialswe can exploit as tools. Recall the restriction endonucleasesand Taq or Pfu polymerase, without which, few of today’sresearch laboratories or biotech companies can operate. Thereare still many microbial tricks we have yet to employ. Abrand-new tool is called the CRISPR-Cas system (clusteredregulatory interspaced short palindromic repeats-CRISPR-associated), which is a bacterial and archaeal antiviralimmune system (yes, they have one). Since 2012 [38]CRISPR-Cas has been advancing research on microbes,worm, fly, fish, mouse, monkey, and plant, i.e., revolu-tionizing the entire fields of genetics and genomics atthis very moment [90].
Plants form another sizable part of the biosphere. Whentouched,Mimosa pudica droops within our attention span andtherefore draws human attention. This, however, is an excep-tional adaptation. Vascular plants detect the forces of rain,wind, predators, etc. by thigmomorphogenesis, i.e., alteringgrowth pattern “slowly” by human standard. They also sensegravity to direct the shoots to grow up and the roots to godown (gravitropism). Darwin described in detail these re-sponses in 1881 [20]. Current research showed that theseprocesses employ Ca2+-binding proteins, Ca2+, and thereforeCa2+-passing channels, including MscS homologs [65]. Seereview by Cox et al. in this special issue [19].
Pflugers Arch - Eur J Physiol

From egg to chicken
Eukaryotes include unicellular fungi, algae, flagellates, cili-ates, amoebae, diatoms, etc. Multicellularity recurred inbrown algal, red algal, animal, fungal, and plant lineages. Itcould, for example, begin with the failure of daughter cells toseparate, a phenotype commonly observed after laboratorymutageneses of unicells. Adhered cells became specializedto gain selective advantage for the clonal colony, a processrecapitulated in embryonic differentiation.
Multicellularity presents two challenges. One is the devel-opment from the unicellular zygote. The current frantic re-search on mammalian stem cell differentiation defies a reviewhere. For our purpose, suffice it to say that embryonic, plurip-otent, or adult stem cells all respond strongly to mechanicalstimuli [94]. Though small diffusible growth factors or inhib-itors are needed, the fate of the stem cells, however, dependson the mechanical properties of the extracellular matrix. Thereare now chemically defined hydrogels of different stiffnessthat present peptides to bind the glycosaminoglycans of hu-man embryonic stem (hES) cells. Only stiff gels maintain theproliferation and pluripotency of hES cells [67].
Adult stem cells, such as the marrow-derivedmesenchymalstem cells (MSCs) , circulate, engraft, and differentiate intodifferent tissues. In identical serum condition, MSCs fate canbe directed by the elasticity of the collagen-containing poly-acrylamide surface, controlled by the degree of cross-linking.Matching the stiffness of the target tissue, MSCs becomeneurons on soft gel, become myoblasts on stiffer gel, andosteoblast on rigid gel [24]. Thus, developing cells activelyprobe their physical environment by an integrin-actomyosin-based force-feedback mechanism. Though some componentsare identified, the mechanism of how force mediates differen-tiation as a whole remains obscure [94] and even controversialin details [91]. Multiple K+, Ca2+, Cl−, or TRP channels arefound in stem cells and appear needed for proliferation [48].Lowering the substrate stiffness reduces the frequency and themagnitude of the MSC Ca2+ oscillation of unknown signifi-cance [41].
Plant growth is ultimately driven by turgor pressure,reminding us again the universal importance of osmotic pres-sure. See Hamant et al. [29] on how forces guide the tissuedifferentiation at the shoot meristem.
Links and tethers
The second complication of multicellularity is cell-cell com-munication in the adults. Within animal cells, stresses areborne by the cytoskeleton, which, however, is not a eukaryoticinnovation. It presumably existed in the common ancestors,since prokaryotes express FtsZ, MreB and crescentin, therespective homologs of actin, tubulin, and the subunit of
intermediate filament [15]. Prokaryotes use them in growth,morphogenesis, DNA partition, cell division, and motility.
The subcortical cytoskeleton of animal cells refers to thefishnet of actin fibers, which subtends and attaches to theplasma membrane. Rat TRPV4 remains responsive to hypo-osmolarity when expressed in yeast [55] and can be stretchedopen in Xenopus oocytes [54]. It is difficult to see how the ratchannel receives force by reconnecting with yeast or toadactins. Cytochalasins or latrunculin, which dissociate actinfibers, enhances rather than disrupt MS channel activities[27, 46]. Mutating β-spectrin (UNC-70), the link betweenmembrane and cytoskeleton, reduces the force needed to pullmembrane tether from the worm’s touch-receptor neurons[44]. The canonical Kv is progressively more active in wholecells, in patches, or upon reconstitution into planar bilayers(above) [80]. These and other observations indicate the sub-cortical actin filaments protect MS channels from openingrather than pull them open.
Animal cells adhere with neighbors through adherencejunctions and to matrix through focal adhesions. Projectinginternally from this adhesion foci are stress fibers, each com-prising 10–30 actin filaments with attached cross-linkers andmyosin motors. The actomyosin produces tensions in thestress fibers between anchors. Stationary focal adhesion sus-tains an ~2.5 pN stretch force [26]. Stretching actin stressfibers in human umbilical vein endothelial cells with an opti-cal tweezer activate Ca2+-passing channels of unknown iden-tity near focal adhesions. Piezoelectrically moving beads at-tached to focal adhesion activates current within 10 ms,peaking within 100 ms [33]. Pulling on magnetic microbeadsattached to integrin through an antibody activates a Ca2+
influx within 4 ms, attributed to TRPV4 [62]. Even though itis far from the speed of the hair-cell transduction channels[17], there is little double that stress fibers can relay force toopen channels. However, it is not known whether the channelis attached to the fiber directly, through linker proteins orthrough a lipid raft (see below and Fig. 1d, e).
Accessory structures accompanying the MS channel arefound in the two key animal models: the vertebrate hair cellsfor hearing and the C. elegans touch cells. In both cases,removing these structures reduces but does not eliminate theMS channel responses. Engineered mutations intoprotocadherin-15 or cadherin-23 remove the tip link, buttransduction currents can still be recorded from hair cells withreduced amplitude and, surprisingly, responding to the nega-tive phase of the sinusoidal force stimulus, in the direction ofmoving the hair bundle away, instead of toward, the kinocil-ium [1]. Mutants lacking Tmc1 and Tmc2 also display re-duced and reverse-phase transduction currents [40], as donascent hair cells in fish lateral-line organs before tip-linkformation [42]. Though there are other explanations, it seemspossible that such currents are through MS channels inuntethered membrane being flexed by vibrations. Prodding
Pflugers Arch - Eur J Physiol

the touch neuron in the dissected worm generates transientinward currents. As expected, mutants lacking channel pore-forming subunits (MEC-4, MEC-10) or the crucial cholesterol-gathering protein (MEC-2) do not show such currents.However, the mutant without the long microtubule due to a nullmutation in MEC-7, which encodes β-tubulin, still displayssuch currents, albeit smaller [69]. It seems possible that tetheringthe MS channels to the tip link or the microtubule relay, orient,or amplify the force signal, but the channels ultimately respondto membrane deformations by the incoming mechanical force(Fig. 1d, e), though the response would be greatly reducedwithout these accessories. See other reviews in this issue.
The lipid bilayer
Voltage, heat, and force are universal physical parameters. Thesensing of all three might all have originated from and stillrequires the lipid bilayer. It is certainly this insulator that erectsany electrochemical gradients, including the all-important H+
gradient. Live cells, from human to bacteria, all maintain aresting potential across this insulator. S4’s arginines dominatecurrent thinking in channel voltage sensitivity. However, all αhelices are dipoles, often even decorated with polar or chargedresidues, making essentially all membrane protein voltage-sensitive to differing degrees. Electrophysiological protocolsare applied to many ion channels not classified as beingvoltage-gated, such as Cl−channels, TRPs, etc. Heat governsall molecules and reactions. Though there are thermodynamicarguments [18], we currently find no convincing molecularmechanisms that explain the unusual heat or cold sensitivity ofcertain TRP channels. Involvement of surrounding and boundlipids seems inevitable. In the recent atomic structure, a den-sity, likely from an endogenous lipid, occupies the capsaicin-binding site of the closed TRPV1 [14].
Parallel to the “RNAworld” theory for the origin of life, thereis a “lipid world” theory that argues for the need and the earlyappearance of an amphipathic envelope that partitions the cellfrom the primordial sea [16]. Such abiotic amphipaths includefatty acids, which could have arrived frommeteorites and can begenerated from reactions simulated in the Miller-Urey experi-ments. Key to the lipid world notion is that these amphipaths, ontheir own, assemble into micelles, monolayers, bilayer frag-ments, or vesicles, through entropy-driven hydrophobic interac-tions, requiring no genetic or divine guidance. Note that aniso-tropic forces within even a naked lipid bilayer can respond todeformation, heat, pH, ions, or heat-induced chemical modifi-cations (see Anishkin et al. [4] for references). These primitivesenses afford a base for improvement by the inclusion ofembedded impurities as arsenal in evolutionary competitions.
The ever-increasing number of atomic structures of ionchannels greatly enhances our knowledge. Visualizing theamino acid arrangements, however, often led us to forget the
surrounding lipids. When included in the crystallization pro-cess of Kv, adjacent lipids can clearly be imaged. They are anintegral part of the channel structure, tightly bound to both theperipheral and the pore domains of Kv. These lipids retain anoverall semblance of a bilayer, but some are clearly distortedby specific binding to amino acids [52]. The surroundinglipids, often called annular lipids, are continuous with thebilayer, while those bound specifically to certain amino acidpockets can be regarded as channel cofactors [47]. The forcedistribution calculated from the pure lipid bilayer reviewedabove and elsewhere [45, 4] will certainly be distorted at thelipid-protein interface. When an embedded protein changesconformation, the associated lipids will have to reconfigure aswell. The free energy change associated with the bilayerdeformation is estimated to be comparable to that of thevoltage-dependent part of the total gating energy of voltage-gated channels [77].
Anishkin and Kung [3] reviewed evidences showing that,in animal cells, foci that are subjected to external forces aremembrane rafts enriched with cholesterol and sphingolipids.Such domains are found at the apex of hair cell [96], at theC. elegans that touch receptor complex, and at ordinaryadherens junctions between cells as well as the cell-matrixfocal adhesions. These more ordered lipid platforms functionto suppress mechanical noise, which is liable to cause leakagethrough sensitive MS channels. Such platforms also confineand/or redirect incoming force. These authors even speculatethat such lipid platforms can be dynamic and rearrange inresponse to the external force. Regardless of such lipid plat-forms, an unanswered question for animal force sensors iswhether the tether force is pulling the channel protein directly orpulling the surrounding lipids (Fig. 1d, e). Proteins function inaggregates: ribosome, lysosomes, chromosomes, endosomes,spliceosomes, etc. Ion channels can certainly be associated withvarious other proteins, but there is no clear evidence that theseassociations transmit force directly. The possibility of the tetherpulling on the lipid platform remains open.
Assuming that the ion channel is directly pulled by thetether, we still need to consider the fact that the channel isintimately glued to the annular lipids and beyond. As citedabove the free energy change in deforming the bilayer iscomparable to that associated with voltage gating of the Kvprotein [77]. In other words, when such a tether displaces anddeforms an MS channel, it necessarily deforms the surround-ing lipids. This is even directly visible, as tip-link leaningcauses tenting of the apical membrane of stereocilia [5](Fig. 1d, e, left vs right).
Retrospect and prospect
Mechanical force sensing is pervasive: above and below ourawareness, during development and in adult, in organs and in
Pflugers Arch - Eur J Physiol

cells, in bacteria, archaea, and eukaryotes (green or other-wise). Such extraordinarily broad manifestations should notblind us from basic molecular principles. We advocate thatFFL is the unifying paradigm. Due to entropy-driven hydro-phobic interactions, lipid bilayer self-assembles at the lifeorigin to enclose the first cytoplasm. It continues to be crucial,although often regarded as a simple barrier, cartooned as twostraight lines. Historically, tremendous efforts had beenwasted in the 1950s and 1960s hunting for “the” elusivehigh-energy intermediate for ATP synthesis, while the answerlays in the H+ gradient sustained by the bilayer. In recentyears, voltage sensing is gradually, if only grudgingly, accept-ed to involve bilayer lipids. We believe that the bilayer holdsother secrets, and some relate to its mechanical properties asdescribed here and elsewhere [3, 4].
We all begin our scientific education with the seventeenthcentury formula F=ma. Thus, it is a great irony that, in thetwenty-first century, we still do not have a clear molecularexplanation for how we, ourselves, feel F. Do not ask yourphysicians what measures blood pressure inside your body. Itwill only serve to embarrass them. Current research in the vastfield of mechanosensations is peculiarly retarded, compared tophotosensations or chemosensations. FFL could provide aparadigm for future research, but research will remain chal-lenging. Lipids are numerous and mixed. Membrane is aprotein-rich mosaic and not a lipid sea with protein icebergs.Lipid chemistry is unfamiliar, especially difficult in situ. Somecurrent biophysical tools, e.g., patch clamp, have inherentartifacts and complexities [86]. New tools, such as fluorescentdyes that gauge bilayer deformations, will be greatly wel-come. Meanwhile, anthropocentricity and the drumbeat fortranslational research aside, we encourage broader thinking toavoid the risk of degrading biology into a drug-discovery-fest.
Acknowledgments Work in our laboratories is supported by the HuckInstitute of Life Sciences (A.A.) and NIH grant GM096088 and the VilasTrust of the University of Wisconsin-Madison (to C.K.).
References
1. Alagramam KN, Goodyear RJ, Geng R, Furness DN, van Aken AF,Marcotti W, Kros CJ, Richardson GP (2011) Mutations inprotocadherin 15 and cadherin 23 affect tip links andmechanotransduction in mammalian sensory hair cells. PLoS One6(4):e19183
2. Anderson M, Kim EY, Hagmann H, Benzing T, Dryer SE (2013)Opposing effects of podocin on the gating of podocyte TRPC6channels evoked by membrane stretch or diacylglycerol. Am JPhysiol Cell Physiol 305(3):C276–C289
3. Anishkin A, Kung C (2013) Stiffened lipid platforms at mo-lecular force foci. Proc Natl Acad Sci U S A 110(13):4886–4892
4. Anishkin A, Loukin S, Teng J.-F, Kung C (2014) Feeling the hiddenmechanical forces in lipid bilayer is an original sense. Proc Natl AcadSci (in press)
5. Assad JA, Shepherd GM, Corey DP (1991) Tip-link integrity andmechanical transduction in vertebrate hair cells. Neuron 7(6):985–994
6. Baek EB, Kim SJ (2011) Mechanisms of myogenic response:Ca(2+)-dependent and -independent signaling. J SmoothMuscle Res 47(2):55–65
7. Bellono NW, Kammel LG, Zimmerman AL, Oancea E (2013) UVlight phototransduction activates transient receptor potential A1 ionchannels in human melanocytes. Proc Natl Acad Sci U S A 110(6):2383–2388
8. Berrier C, Pozza A, de Lacroix de Lavalette A, Chardonnet S,Mesneau A, Jaxel C, le Maire M, Ghazi A (2013) The purifiedmechanosensitive channel TREK-1 is directly sensitive to membranetension. J Biol Chem 288(38):27307–27314
9. Bourque CW (2008) Central mechanisms of osmosensation andsystemic osmoregulation. Nat Rev Neurosci 9(7):519–531
10. Brohawn SG, Campbell EB,MacKinnon R (2013) Domain-swappedchain connectivity and gated membrane access in a Fab-mediatedcrystal of the human TRAAKK+channel. Proc Natl Acad Sci U S A110(6):2129–2134
11. Brohawn SG, del Marmol J, MacKinnon R (2012) Crystal structureof the human K2P TRAAK, a lipid- and mechano-sensitive K+ ionchannel. Science 335(6067):436–441
12. Brohawn SG, Su Z, Mackinnon R (2014) Mechanosensitivity ismediated directly by the lipid membrane in TRAAK and TREK1K+ channels. Proc Natl Acad Sci U S A 111(9):3614–3619
13. Cantor RS (1997) Lateral pressures in cell membranes: a mechanismfor modulation of protein function. J Phys Chem 101:1723–1725
14. Cao E, Liao M, Cheng Y, Julius D (2013) TRPV1 structures indistinct conformations reveal activation mechanisms. Nature504(7478):113–118
15. Celler K, Koning RI, Koster AJ, van Wezel GP (2013)Multidimensional view of the bacterial cytoskeleton. J Bacteriol195(8):1627–1636
16. Chen IA, Walde P (2010) From self-assembled vesicles to protocells.Cold Spring Harb Perspect Biol 2(7):a002170
17. Chris tensen AP, Corey DP (2007) TRP channels inmechanosensation: direct or indirect activation? Nat Rev Neurosci8(7):510–521
18. Clapham DE, Miller C (2011) A thermodynamic framework forunderstanding temperature sensing by transient receptor potential(TRP) channels. Proc Natl Acad Sci U S A 108(49):19492–19497
19. Cox CD, Nakayama Y, Nomura T, Martinac B (2014) The evolu-tionary ‘tinkering’ of MscS-like channels: generation of structuraland functional diversity. Pfluegers Archiv - Eur J Physiol (in press)
20. Darwin C, Darwin F (1881) The power of movement in plants.Appleton and Company, New York
21. Darwin C, Darwin F (1903) More letters of Charls Darwin. Hazell,Watson, and Viney, London
22. Delmas P, Coste B (2013) Mechano-gated ion channels in sensorysystems. Cell 155(2):278–284
23. Eckert R (1972) Bioelectric control of ciliary activity. Science 176:473–481
24. Engler AJ, Sen S, Sweeney HL, Discher DE (2006) Matrix elasticitydirects stem cell lineage specification. Cell 126(4):677–689
25. Goodman MB, Ernstrom GG, Chelur DS, O’Hagan R, Yao CA,Chalfie M (2002) MEC-2 regulates C. elegans DEG/ENaC channelsneeded for mechanosensation. Nature 415(6875):1039–1042
26. Grashoff C, Hoffman BD, Brenner MD, Zhou R, Parsons M, YangMT, McLean MA, Sligar SG, Chen CS, Ha T, Schwartz MA (2010)Measuring mechanical tension across vinculin reveals regulation offocal adhesion dynamics. Nature 466(7303):263–266
27. Guharay G, Sachs F (1984) Stretch-activated single ion channelsurrents in tissure-cultured embryonic chich skeletal muscle. JPhysiol 352:685–701
28. Gullingsrud J, Schulten K (2004) Lipid bilayer pressure profiles andmechanosensitive channel gating. Biophys J 86(6):3496–3509
Pflugers Arch - Eur J Physiol

29. Hamant O, Heisler MG, Jonsson H, Krupinski P, Uyttewaal M,Bokov P, Corson F, Sahlin P, Boudaoud A, Meyerowitz EM,Couder Y, Traas J (2008) Developmental patterning by mechanicalsignals in Arabidopsis. Science 322
30. Hamill OP, McBride DW Jr (1992) Rapid adaptation of singlemechanosensitive channels in Xenopus oocytes. Proc Natl AcadSci U S A 89(16):7462–7466
31. Hao J, Padilla F, Dandonneau M, Lavebratt C, Lesage F, Noel J,Delmas P (2013) Kv1.1 channels act as mechanical brake in thesenses of touch and pain. Neuron 77(5):899–914
32. Hardie RC, Franze K (2012) Photomechanical responses inDrosophila photoreceptors. Science 338(6104):260–263. doi:10.1126/science
33. Hayakawa K, Tatsumi H, Sokabe M (2008) Actin stress fiberstransmit and focus force to activate mechanosensitive channels. JCell Sci 121(Pt 4):496–503
34. Honore E (2007) The neuronal background K2P channels: focus onTREK1. Nat Rev Neurosci 8(4):251–261
35. Huber TB, Schermer B, Muller RU, Hohne M, Bartram M, CalixtoA, Hagmann H, Reinhardt C, Koos F, Kunzelmann K, Shirokova E,Krautwurst D, Harteneck C, Simons M, Pavenstadt H, Kerjaschki D,Thiele C,Walz G, Chalfie M, Benzing T (2006) Podocin andMEC-2bind cholesterol to regulate the activity of associated ion channels.Proc Natl Acad Sci U S A 103(46):17079–17086
36. Hughes S, Hankins MW, Foster RG, Peirson SN (2012) Melanopsinphototransduction: slowly emerging from the dark. Prog Brain Res199:19–40
37. Jennings HS (1908) The interpretation of the behavior of the lowerorganisms. Science 27(696):698–710
38. JinekM, Chylinski K, Fonfara I, HauerM, Doudna JA, Charpentier E(2012) A programmable dual-RNA-guided DNA endonuclease inadaptive bacterial immunity. Science 337(6096):816–821
39. Kawagishi I, Imagawa M, Imae Y, McCarter L, Homma M (1996)The sodium-driven polar flagellar motor of marine Vibrio as themechanosensor that regulates lateral flagellar expression. MolMicrobiol 20(4):693–699
40. Kim KX, Beurg M, Hackney CM, Furness DN, Mahendrasingam S,Fettiplace R (2013) The role of transmembrane channel-like proteinsin the operation of hair cell mechanotransducer channels. J GenPhysiol 142(5):493–505
41. Kim TJ, Seong J, OuyangM, Sun J, Lu S, Hong JP,WangN,WangY(2009) Substrate rigidity regulates Ca2+ oscillation via RhoA path-way in stem cells. J Cell Physiol 218(2):285–293
42. Kindt KS, Finch G, Nicolson T (2012) Kinocilia mediatemechanosensitivity in developing zebrafish hair cells. Dev Cell23(2):329–341
43. Kloda A, Lua L, Hall R, Adams DJ, Martinac B (2007) Liposomereconstitution and modulation of recombinant N-methyl-D-aspartatereceptor channels by membrane stretch. Proc Natl Acad Sci U S A104(5):1540–1545
44. Krieg M, Dunn AR, GoodmanMB (2014) Mechanical control of thesense of touch by beta-spectrin. Nat Cell Biol 16:224–233
45. Kung C (2005) A possible unifying principle for mechanosensation.Nature 436(7051):647–654
46. Lauritzen I, Chemin J, Honore E, Jodar M, Guy N, Lazdunski M,Jane Patel A (2005) Cross-talk between the mechano-gated K2Pchannel TREK-1 and the actin
47. Lee AG (2005) How lipids and proteins interact in a membrane: amolecular approach. Mol Biosyst 1(3):203–212
48. Li GR, Deng XL (2011) Functional ion channels in stem cells. WorldJ Stem Cells 3(3):19–24
49. Liedtke W, Friedman JM (2003) Abnormal osmotic regulation intrpv4-/- mice. Proc Natl Acad Sci U S A 100(23):13698–13703
50. Liman ER (2012) Cell signaling. Putting the squeeze onphototransduction. Science 338(6104):200–201
51. Lindahl E, Edholm O (2000) Spatial and energetic-entropic decom-position of surface tension in lipid bilayers from molecular dynamicssimulations. J Chem Phys 113:3882–3893
52. Long SB, Tao X, Campbell EB, MacKinnon R (2007) Atomicstructure of a voltage-dependent K+channel in a lipid membrane-like environment. Nature 450(7168):376–382
53. Loukin S, Su Z, Kung C (2011) Increased basal activity is a keydeterminant in the severity of human skeletal dysplasia caused byTRPV4 mutations. PLoS One 6(5):e19533
54. Loukin S, Zhou X, Su Z, Saimi Y, Kung C (2010) Wild-type andbrachyolmia-causing mutant TRPV4 channels respond directly tostretch force. J Biol Chem 285(35):27176–27181
55. Loukin SH, Su Z, Kung C (2009) Hypotonic shocks activate ratTRPV4 in yeast in the absence of polyunsaturated fatty acids.FEBS Lett 583(4):754–758
56. Lu Y, Ma X, Sabharwal R, Snitsarev V, Morgan D, Rahmouni K,Drummond HA, Whiteis CA, Costa V, Price M, Benson C, WelshMJ, Chapleau MW, Abboud FM (2009) The ion channel ASIC2 isrequired for baroreceptor and autonomic control of the circulation.Neuron 64(6):885–897
57. Lundbaek JA, Collingwood SA, Ingolfsson HI, Kapoor R, AndersenOS (2010) Lipid bilayer regulation of membrane protein function:gramicidin channels as molecular force probes. J R Soc Interface7(44):373–395
58. Maroto R, Raso A, Wood TG, Kurosky A, Martinac B, Hamill OP(2005) TRPC1 forms the stretch-activated cation channel in verte-brate cells.[see comment]. Nat Cell Biol 7(2):179–185
59. Martinac B, Adler J, Kung C (1990) Mechanosensitive ion channelsof E. coli activated by amphipaths. Nature 348(6298):261–263
60. Martinac B, Buechner M, Delcour AH, Adler J, Kung C (1987)Pressure-sensitive ion channel in Escherichia coli. Proc Natl AcadSci U S A 84(8):2297–2301
61. Masuyama R, Vriens J, Voets T, Karashima Y, Owsianik G,Vennekens R, Lieben L, Torrekens S, Moermans K, Vanden BoschA, Bouillon R, Nilius B, Carmeliet G (2008) TRPV4-mediatedcalcium influx regulates terminal differentiation of osteoclasts. CellMetab 8(3):257–265
62. Matthews BD, Thodeti CK, Tytell JD, Mammoto A, Overby DR,Ingber DE (2010) Ultra-rapid activation of TRPV4 ion channels bymechanical forces applied to cell surface beta1 integrins. Integr Biol(Camb) 2(9):435–442
63. Miller AN, Long SB (2012) Crystal structure of the human two-poredomain potassium channel K2P1. Science 335(6067):432–436
64. Mizoguchi F,Mizuno A, Hayata T, Nakashima K, Heller S, Ushida T,Sokabe M, Miyasaka N, Suzuki M, Ezura Y, Noda M (2008)Transient receptor potential vanilloid 4 deficiency suppressesunloading-induced bone loss. J Cell Physiol 216(1):47–53
65. Monshausen GB, Haswell ES (2013) A force of nature: molecularmechanisms of mechanoperception in plants. J Exp Bot 64(15):4663–4680
66. Moore C, Cevikbas F, Pasolli HA, Chen Y, Kong W, Kempkes C,Parekh P, Lee SH, Kontchou NA, Ye I, Jokerst NM, Fuchs E,Steinhoff M, Liedtke WB (2013) UVB radiation generates sunburnpain and affects skin by activating epidermal TRPV4 ion channelsand triggering endothelin-1 signaling. Proc Natl Acad Sci U SA.110(34)E3225-3234
67. Musah S, Morin SA, Wrighton PJ, Zwick DB, Jin S, Kiessling LL(2012) Glycosaminoglycan-binding hydrogels enable mechanicalcontrol of human pluripotent stem cell self-renewal. ACS Nano6(11):10168–10177
68. Nilius B, Voets T (2013) The puzzle of TRPV4 channelopathies.EMBO Rep 14(2):152–163
69. O'Hagan R, Chalfie M, Goodman MB (2005) The MEC-4 DEG/ENaC channel of Caenorhabditis elegans touch receptor neuronstransduces mechanical signals. Nat Neurosci 8:43–50
Pflugers Arch - Eur J Physiol

70. Ogura A, Machemer H (1980) Distribution of mechanoreceptorchannels in the Paramecium surface membrane. J Comp Physiol135(3):233–242
71. Oliet SH, Bourque CW (1993) Mechanosensitive channels transduceosmosensitivity in supraoptic neurons. Nature 364(6435):341–343.doi:10.1038/364341a0
72. Opsahl LR, Webb WW (1994) Lipid-glass adhesion in giga-sealedpatch-clamped membranes. Biophys J 66(1):75–79
73. Patel AJ, Lazdunski M, Honore E (2001) Lipid and mechano-gated 2P domain K(+) channels. Curr Opin Cell Biol 13(4):422–428
74. Perozo E, Cortes DM, Sompornpisut P, Kloda A, Martinac B (2002)Open channel structure of MscL and the gating mechanism ofmechanosensitive channels. Nature 418(6901):942–948
75. Perozo E, Kloda A, Cortes DM, Martinac B (2002) Physical princi-ples underlying the transduction of bilayer deformation forces duringmechanosensitive channel gating. Nat Struct Biol 9(9):696–703
76. Petric S, Clasen L, van Wessel C, Geduldig N, Ding Z, SchullenbergM, Mersmann J, Zacharowski K, Aller MI, Schmidt KG, Donner BC(2012) In vivo electrophysiological characterization of TASK-1 de-ficient mice. Cell Physiol Biochem 30(3):523–537
77. Reeves D, Ursell T, Sens P, Kondev J, Phillips R (2008) Membranemechanics as a probe of ion-channel gating mechanisms. Phys Rev EStat Nonlinear Soft Matter Phys 78(4 Pt 1):041901
78. Satow Y, Murphy AD, Kung C (1983) The ionic basis of thedepolarising mechanoreceptor potential of Paramecium tetraurelia.J Exp Biol 103:253–264
79. Schermer B, Benzing T (2009) Lipid-protein interactions along theslit diaphragm of podocytes. J Am Soc Nephrol 20(3):473–478
80. Schmidt D, del Marmol J, MacKinnon R (2012) Mechanistic basisfor low threshold mechanosensitivity in voltage-dependent K+ chan-nels. Proc Natl Acad Sci U S A 109(26):10352–10357
81. Schmidt D, MacKinnon R (2008) Voltage-dependent K+ channelgating and voltage sensor toxin sensitivity depend on the mechanicalstate of the lipid membrane. Proc Natl Acad Sci U S A 105(49):19276–19281
82. Sexton T, Buhr E, Van Gelder RN (2012) Melanopsin and mecha-nisms of non-visual ocular photoreception. J Biol Chem 287(3):1649–1656
83. Sharif Naeini R, Witty MF, Seguela P, Bourque CW (2006) An N-terminal variant of Trpv1 channel is required for osmosensory trans-duction. Nat Neurosci 9(1):93–98
84. Sharif-Naeini R, Ciura S, Zhang Z, Bourque CW (2008) Contributionof TRPV channels to osmosensory transduction, thirst, and vasopres-sin release. Kidney Int 73(7):811–815
85. Sharif-Naeini R, Dedman A, Folgering JH, Duprat F, Patel A, NiliusB, Honore E (2008) TRP channels and mechanosensory transduc-tion: insights into the arterial myogenic response. Pflugers Arch456(3):529–540
86. Suchyna TM, Markin VS, Sachs F (2009) Biophysics and structureof the patch and the gigaseal. Biophys J 97(3):738–747
87. Sukharev SI, Blount P, Martinac B, Blattner FR, Kung C (1994) Alarge-conductance mechanosensitive channel in E. coli encoded bymscL alone. Nature 368(6468):265–268
88. Sukharev SI, Blount P, Mart inac B, Kung C (1997)Mechanosensitive channels of Escherichia coli: the MscL gene,protein, and activities. Annu Rev Physiol 59:633–657. doi:10.1146/annurev.physiol.59.1.633
89. Sun H, Li DP, Chen SR, Hittelman WN, Pan HL (2009) Sensing ofblood pressure increase by transient receptor potential vanilloid 1receptors on baroreceptors. J Pharmacol Exp Ther 331(3):851–859
90. Terns RM, Terns MP (2014) CRISPR-based technologies: prokary-otic defense weapons repurposed. Trends Genet 30(3):111–118
91. Trappmann B, Gautrot JE, Connelly JT, Strange DG, Li Y, OyenML,Cohen Stuart MA, BoehmH, Li B, Vogel V, Spatz JP,Watt FM, HuckWT (2012) Extracellular-matrix tethering regulates stem-cell fate.Nat Mater 11(7):642–649
92. Vasquez V, Krieg M, Lockhead D, Goodman MB (2014)Phospholipids that contain polyunsaturated fatty acids enhance neu-ronal cell mechanics and touch sensation. Cell Rep 6(1):70–80
93. Wetzel C, Hu J, Riethmacher D, Benckendorff A, Harder L, Eilers A,Moshourab R, Kozlenkov A, Labuz D, Caspani O, Erdmann B,Machelska H, Heppenstall PA, Lewin GR (2007) A stomatin-domain protein essential for touch sensation in the mouse. Nature445(7124):206–209
94. Yim EK, Sheetz MP (2012) Force-dependent cell signaling in stemcell differentiation. Stem Cell Res Ther 3(5):41
95. Zhang Y, Gao F, Popov VL, Wen JW, Hamill OP (2000)Mechanically gated channel activity in cytoskeleton-deficient plas-ma membrane blebs and vesicles from Xenopus oocytes. J Physiol523(Pt 1):117–130
96. Zhao H, Williams DE, Shin JB, Brugger B, Gillespie PG (2012)Large membrane domains in hair bundles specify spatially constrict-ed radixin activation. J Neurosci 32(13):4600–4609
Pflugers Arch - Eur J Physiol





























