Embed Size (px)
Citation preview

598 G. C. WOOD AND M. K. KEECH 1960Gross, J. (1956). J. biophy8. biochem. Cytol. 2, 261.Gross, J. (1958). J. exp. Med. 108, 215.Gross, J., Highberger, J. H. & Schmitt, F. 0. (1955). Proc.
nat. Acad. Sci., Wa8h., 41, 1.Gross, J. & Kirk, D. (1958). J. biol. Chem. 233, 355.Hall, C. E. (1956). Proc. nat. Acad. Sci., Wa8h., 42, 801.Hall, C. E. & Doty, P. (1958). J. Amer. chem. Soc. 80, 1269.Hawn, C. v. Z. & Porter, K. R. (1947). J. exp. Med. 86, 285.Jackson, D. S. (1958a). Recent Advances in Gelatin and Glue
Re8earch, p. 50. Ed. by Stainsby, G. London: Per-gamon Press Ltd.
Jackson, D. S. (1958b). New. Engl. J. Med. 259, 514.Jackson, D. S. & Fessler, J. H. (1955). Nature, Lond., 176,
69.Jackson, D. S. & Neuberger, A. (1957). Biochim. biophy8.
Acta, 26, 638.Jackson, S. F. (1954a). Nature, Lond., 173, 950.Jackson, S. F. (1954b). Proc. Roy. Soc. B, 142, 536.Jackson, S. F. (1955). Proc. Roy. Soc. B, 144, 556.Jackson, S. F. (1956). Proc. Roy. Soc. B, 146, 270.King, E. J. (1932). Biochem. J. 26, 292.Lowry, 0. H., Rosebrough, N. J., Farr, A. I. & Randall,
R. J. (1951). J. biol. Chem. 193, 265.Mathews, M. B., Kulonen, E. & Dorfinan, A. (1954). Arch.
Biochem. Biophy8. 52, 247.'Meyer, K. (1946). Amer. J. Med. 1, 675.Meyer, K. (1955-56). Harvey Lect. 1, 97.Neuman, R. E. & Logan, M. A. (1950). J. biol. Chem. 184,
299.
Porter, K. R. (1951). Tran8. 2nd Conf. Josiah Macy Jr.Found., New York: Connective Tis8ues, p. 126.
Porter, K. R. & Hawn, C. v. Z. (1949). J. exp. Med. 90,225.Porter, K. R. & Vanamee, P. (1949). Proc. Soc. exp. Biol.,
N.Y., 71, 513.Randall, J. T., Booth, F., Burge, R. E., Jackson, S. F. &
Kelly, F. C. (1955). Symp. Soc. exp. Biol. 9, 127.Reed, R. (1957). Counc. int. Org. Med. Sci. Symp.:
Connective Tissue, p. 304. Ed. by Tunbridge, R. E.Oxford: Blackwell Scientific Publications Ltd.
Schmitt, F. 0. (1957). J. cell. comp. Phy8iol. 49, Suppl. 1,-85.
Schmitt, F. 0. (1959). Rev. mod. Phys. 31, 349.Schmitt, F. O., Gross, J. & Highberger, J. H. (1955a).Symp. Soc. exp. Biol. 9, 326.
Schmitt, F. O., Gross, J. & Highberger, J. H. (1955b).Exp. Cell Res. Suppl. 3, 326.
Schwartz, W. (1957). Counc. int. Org. Med. Sci. Symp.:Connective Tisue, p. 144. Ed. by Tunbridge, R. E.Oxford: Blackwell Scientific Publications Ltd.
Scott, T. A. & Melvin, E. H. (1953). Analyt. Chem. 25,1656.
Shulman, S. & Ferry, J. D. (1950). J. Phys. Chem. 54, 66.Vanamee, P. & Porter, K. R. (1951). J. exp. Med. 94, 255.Wassermann, F. (1954). Amer. J. Anat. 94, 399.Wood, G. C. (1958). Abstr. Comm. 4th int. Congr. Biochem.,
Vienna, p. 26, 2-92.Wood, G. C. (1960a). Biochem. J. 75, 598.Wood, G. C. (1960b). Biochem. .J. 75, 605.
Biochem. J. (1960) 75, 598
The Formation of Fibrils from Collagen Solutions2. A MECHANISM OF COLLAGEN-FIBRIL FORMATION
BY G. C. WOODNuffield Gerontological Re8earch Unit, Department of Medicine, Univer8ity of Leed8
(Received 22 June 1959)
In the preceding paper (Wood & Keech, 1960)evidence was adduced for the hypothesis that pre-cipitation of collagen fibrils from collagen solutionsoccurs in two steps, the first occurring during thelag period, the second being represented by thesigmoid precipitation curve. This suggestion hasalso been put forward by Bensusan & Hoyt (1958).The ultimate fibril width appears to be determinedduring the first step. It is the purpose of thepresent paper to see how the experimental resultsmay be interpreted quantitatively in terms of atwo-step mechanism.
Freshly prepared collagen precipitates can beredissolved by altering the pH or temperature andthen reprecipitated by returning these variables totheir original values (Jackson, 1957; Gross, 1958).
It seems reasonable therefore to regard the forma-tion of collagen fibrils as precipitation of the pro-tein, in an ordered form, from solution in which itis supersaturated under the particular conditions ofpH, ionic strength, temperature, etc. One mightexpect therefore some similarity between the pre-cipitation of collagen and the precipitation ofsparingly soluble inorganic salts, in crystallineform, from supersaturated solution.The kinetics of the latter process has been widely
studied and in many cases it has been recognizedthat the process takes place in two steps: (a)aggregation of the particles in solution to formnuclei, i.e. clusters of particles of just sufficientsize to be stable as a separate phase: (b) growth ofthese nuclei, by accretion of soluble material, to

MECHANISM OF COLLAGEN-FIBRIL FORMATION
form a visible precipitate. Equations describingthe kinetics of these two steps have been derivedand applied with some success to a number ofsystems (e.g. Johnson & O'Rourke, 1954; Duke &Brown, 1954; Nielsen, 1955, 1958; Collins &Leinweber, 1956).The mechanism of protein precipitation is
probably a good deal more complicated than that ofinorganic salts. This is likely to be particularly soin the formation of collagen fibrils, where highlyasymmetric soluble particles aggregate in a veryspecific manner with their axes parallel to theultimate fibril axis (Schmitt, Gross & Highberger,1955a, b; Gross, 1956; Schmitt, 1959). Neverthe-less, the same general principles should be applic-able. Waugh (1957) has, in fact, applied thenucleation-growth concept to the precipitation ofinsulin fibrils, which, like collagen fibrils, are theresult of aggregation of asymmetric particles.
It is of interest therefore to see how far the ideaaccounts for the rate of formation of collagenfibrils.
Following Waugh's treatment of the problem,the rate of formation of nuclei,
dndt =k1Cx, (1)
where C is concentration of collagen, n is concentra-tion of nuclei, x is a constant and k1 is a rateconstant. This equation implies that x collagenparticles must come together simultaneously toform a nucleus. The rate of growth of each pre-cipitated particle,
dmdt = k2AC, (2)
where m is mass of the precipitated particle, A isaggregating area and k2 is a rate constant. A isassumed to be proportional to the surface area andthe further assumption is made that the shape ofthe precipitated particle remains unchanged duringits growth. Then Aocml, and equation (2) becomes
dtThe rate of growth is thus assumed to be controlledby the condensation of the dissolved collagen on tothe precipitate and not by its diffusion to thesurface of the precipitated particles.
If the growth process is reversible, equation (3)must be modified to
dmd= k3mI(C-CS), (4)
where C8 is the value of C at equilibrium, i.e. thesolubility of the collagen.
In the very early stages of precipitation, when
C - Co (CO, initial collagen concentration) equation(4) may be integrated:
k3(CO-C8) t327
Introducing equation (1),
mdn= k3k dt(Co-C8)3tdt27
therefore
(Co-C) = (nftmdn = k1k3Co(Co-C8)3t4Jo 108
Introducing the extent of precipitation,
p = (Co-C)/(Co-aC),and putting (CO-C)/Co =
108
where
(5)
(6)
KI = klk3-This relationship will clearly result in an apparentlag period such as is actually observed (Bensusan &Hoyt, 1958; Wood & Keech, 1960).As the collagen concentration falls appreciably
below C0 the rate of nucleation as given byequation (1) falls rapidly due to the term CO. Atthe same time the rate of growth of each precipi-tated particle as given by equation (4) increases asthe particle grows, due to the term mn', untileventually the fall of collagen concentration slowsgrowth down too. Thus the major part of precipi-tation will occur by growth of those nuclei formedearly in the reaction, i.e. during the lag period. Thecontribution due to growth of nuclei formed laterwill be small, its actual extent depending on therelative values of k1, k3 and x. Assuming that theircontribution is negligible, we have at the end of thelag period a set of small particles whose subsequentgrowth is responsible for all further precipitation.
It can now be seen how the observed two-stepnature of precipitation (Wood & Keech, 1960)arises in terms of this mechanism. The first step,occupying the lag period, consists mainly of theformation of nuclei whereas the second consists ofthe growth of these nuclei into fibrils. The fibrillarnature of the precipitate results from the asym-metry of the nuclei and of the collagen macro-molecules themselves. In the closed system studiedexperimentally the size of the fibrils is determinedby the number and the shape of the nuclei and ishence decided during the lag period. It is notaffected by change in rate of growth after the lagperiod, provided that the manner in which growthtakes place does not change at the same time. Themechanism thus accounts qualitatively for theobservation that although precipitation is acceler-ated by increasing the temperature either before or
Vol. 75 599
G. C. WOODafter the lag period, fibril width is altered only ifthe temperature is increased before the lag periodand is not affected by increasing the temperatureafter the lag period.For low values of p the predicted course of pre-
cipitation is given by equation (6). The size distri-bution and hence the growth characteristics, at theend of the lag period, of those particles fermedduring the lag period might be calculated fromequation (5) and the integrated form of equation (1):
n = kCOO t. (7)
The time course of precipitation after the lag I ,riodcould then be calculated by summing the contribu-tion of all the species of particle. The calculationcan be simplified by replacing the distribution ofparticles by a concentration, ne, of hypoth ticalidentical particles. The value of ne and the sze ofthese particles is determined by the conditions thattheir total surface area and total mass must be thesame as those of the actual distribution of particles.Then from equation (4)
dmnne d = k3n4m*C,
which may be transformed into
dP = K2 , CO*P*('1-P)' (8)
where K2 = k2ni. On integration this becomes:
K2flICit p( ,I, (9)
where
(R -1)2 1 + 2R-1
and R = pi.
Application of equation8 (6) and (9)to experimental data
The experimental data are those given in thepreceding paper (Wood & Keech, 1960). In all theexperiments the collagen was almost completelyprecipitated and therefore , B 1.
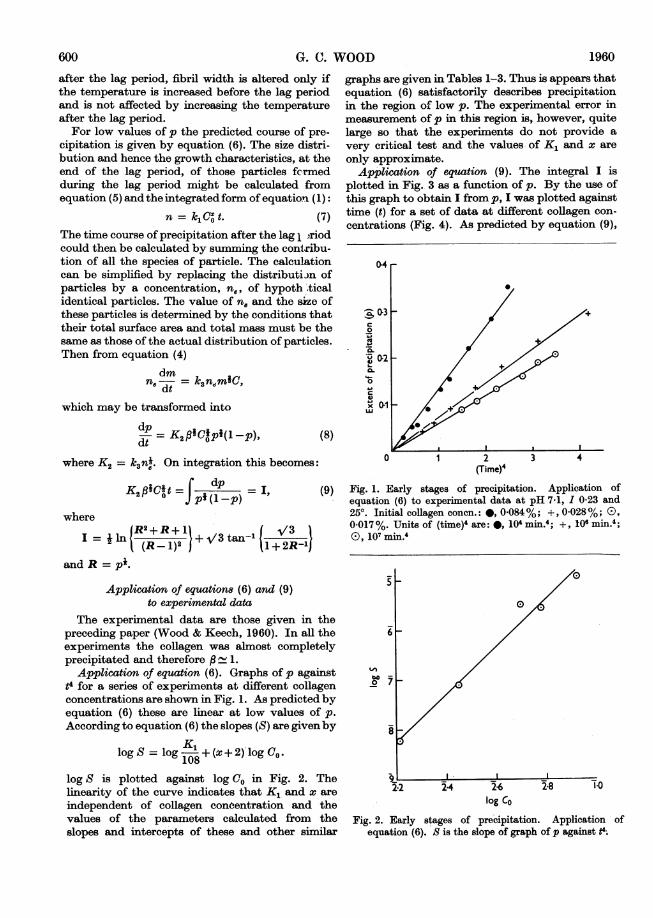
Application of equation (6). Graphs of p againstt4 for a series of experiments at different collagenconcentrations are shown in Fig. 1. As predicted byequation (6) these are linear at low values of p.According to equation (6) the slopes (S) are given by
logS = log K52z+(x+2) log Co.logS = 108
log S is plotted against log CO in Fig. 2. Thelinearity of the curve indicates that K, and x areindependent of collagen conoentration and thevalues of the parameters calculated from theslopes and intercepts of these and other similar
graphs are given in Tables 1-3. Thus is appears thatequation (6) satisfactorily describes precipitationin the region of low p. The experimental error inmeasurement of p in this region is, however, quitelarge so that the experiments do not provide avery critical test and the values of K, and x areonly approximate.
Application of equation (9). The integral I isplotted in Fig. 3 as a function of p. By the use ofthis graph to obtain I from p, I was plotted againsttime (t) for a set of data at different collagen con-centrations (Fig. 4). As predicted by equation (9),
04
0 2(TiMe)4
3 4
Fig. 1. Early stages of precipitation. Application ofequation (6) to experimental data at pH 7-1, 1 0-23 and250. Initial collagen conen.: *, 0-084%; +, 0-028%; 0),0-017%. Units of (time)4 are: *, 104 min.4; +, 106 min.4;(D, 107 min.4
8
0
Fi ~0
2-2 2-4 2-6 2-8 1-0log Co
Fig. 2. Early stages of precipitation. Application ofequation (6). S is the slope of graph of p against t4.
600 1960
I .- I I

MECHANISM OF COLLAGEN-FIBRIL FORMATIONthe curves are linear over most of their course. Theslopes are proportional to K2, whereas the inter-cepts, t, mark the ends of the lag periods. Thedeviation from linearity at low values ofp might beexpected, due to overlap of the nucleation andgrowth phases, whereas deviation at values of p
Table 1. Effect of collagen concentration oparameters for nucleation and growth
Experimental conditions: 250; pH 7-1; I 0 idefinitions of Co, x, K1, t4 and K2, see text. The refor two different preparations of collagen solution.
co(%)
0-0840-0610O0500-0280-017,0 0500-0340.017
tcx K1 (min.)
4-75*8
2-7 227 5-414-025-53-8
2-5 100 7-214-5
n the
approaching unity might be due to the sensitivityof I to errors in p in this region. In spite of thesedeviations equation (9) describes the shape of theprecipitation curves quite well, as shown in Fig. 5,where the smooth curves were calculated fromvalues of K2 and t, obtained from straight linessuch as those shown in Fig. 4. Similar results wereobtained for a wide range of conditions and thevalues of K2 and t, are given in Tables 1-3.
23. For Fibril widthsults are The successful application of equation (9) to
precditation curves and the fact that the mech-anismii accounts qualitatively for the observation
K2 that fibril size is determined during the lag period1-49 suppbrts the assumption that only those nuclei1-32 forn-d during the lag period contribute signifi-0-76 cantly to precipitation.0 49 According to the simple mechanism, at any stage0.99 of precipitation these nuclei will have grown into0-580 49
Table 2. Effect of pH and ionic strength on theparameters for nucleation and growth
Experimental conditions: 250; C0 0.05 %. For definitionsof x, K1, tc and K2, see text. Each set ofresults was obtainedon a different preparation of collagen solution.
pH 16-0 0-237.1 0-237-5 0-238-0 0*236-0 0-237*1 0-238-0 0-237*0 0*137*0 0-237.0 0-317-0 0-137*0 0-237*0 0-316-0 0-136-0 0-236-0 0-318*0 0-138-0 0-238-0 0-31
x
2-52-73*0
1-81*83-3
K1
30 5001 4003 100
68 0001 400540
tc(min.)2-07*07.56-82*05-04-71*02-5
26-31-050
10-01-02-01.03-16-8
27-0
I
K24-421*030*961-336-121-772-287-441-180-459*281770-819-364-227-224-571-320-48
Table 3. Effect of temperature on the parametersfor nucleation and growth
Experimental conditions: pH 7; I 0-23; C0 0.05 %. Fordefinitions of x, K1, K2 and (K2). see text.
Temp. x K1 K2 (K2)g200 - - 0-18 0-1725 2*5 100 0*99 1-0230 1X9 440 2X25 1.5537 1X6 28 000 11-35 4*85
I 1.0 02 04 0-6 &8
Extent of precipitation (p)
I.1.0
Fig. 3. The integral I plotted as a function of p(see equation 9).
I
Time (min.)Fig. 4. Precipitation at pH 7-1, I 0-23 and 250 at threedifferent collagen concentrations: (1) 0.084%; (2) 0*028 %;(3) 0-017 %. Data are plotted in accordance with equation(9).
Vol. 75 601

G. C. WOOD
p 30-4 (a) 0c4 )(
02
50 100 150 200 0 20 40 60 80 100t (min.) t (min.)
10 5 0 12 0 0 o00
0.8 (3) t
0-6 0-6 (2)p 3
0-4 (b) 0.4(4) (d)
0-2 0-2
0 50 100 150 200 0 20 4060 80
t (min.) t (min.)
Fig. 5. Precipitation under different experimental condi-tions. The curves represent the best fit of equation (9) tothe experimental data. (a) Different collagen concentra-tions CO: (1) 0.084%; (2) 0-028%; (3) 0.017%. (b)Different temperatures: (1) 370; (2) 250; (3) 200. (c)Different ionic strengths, 1: (1) 0-13; (2) 0-23; (3) 0-31.(d) Different pH values: (1) pH 6-0; (2) pH 7-1; (3) pH 7-5;(4) pH 8-1. Except for the parameters stated in each set,conditions were: pH 7-1; I 0-23; Co 0.05%; temp. 250.
fibrils, the largest of which correspond to nucleiformed at zero time and the smallest to nucleiformed at t = ti. From equations (5) and (7),the distribution of fibril width will be such thatthere are equal numbers of fibrils of differentwidths, between the two extremes, if, as assumedearlier, the shape of the fibrils remains constantduring growth. The observed distribution is narrowbut does not conform precisely to this prediction(Wood & Keech, 1960), as may be seen from thehistograms reproduced in Figs. 6 and 7. Since it islikely, however, that the shape of the fibrils isdistorted during drying for electron microscopy(see Anderson, 1956), this does not necessarily in-validate the assumption of constant shape and, as
shown below, the observed distribution is not in-consistent with the mechanism.From equation (4) the mass ofeach ofthe smallest
fibrils (mnc) at any time, t, is given approximately by
k3 Cdt k30Co tf(1 )dt
Regarding each fibril as a long cylinder of radius r
and length 1,(rm.)t piTm(r0)3,
1960
where p is density, a( = I/r) is a constant and (rc)t is
radius of each of the smallest fibrils at time t.
Therefore (r,)t = j3Co (1 -p) dt. (10)
Similarly, the radius (ro) of the largest fibrils is
(fo)dt=
3p7 (I -p) dt =3(pnoc)* cte -P) dtA
(11)
-o. 1.0
D° 26
11
.0
0
0
v
W024,ILIA0~
10-3xfibril width (X)05 1-0 1-5. 2-0 2-5 3-0 3-5 4-0
Fig. 6. Comparison of calculated (broken lines) andobserved (full lines) fibril-width distributions. Data were
obtained at pH 7-1, C0 0-05% and 250. Ionic strengths, I:
(a) 0-31; (b) 0-23; (c) 0-13.
.0= 10
'lbc
o 20o
C
0~
10-3x fibril width (A)05 1-0 1-5 20
Fig. 7. Comparison of calculated (broken lines) andobserved (full lines) fibril-width distributions. Data were
obtained at pH 7-1, I 0-23; C0 0-05%. Temperatures:(a) 250; (b) 370.
602
I10 - [ (a)
10 _ C=; iriINPo0 rn-2'lIi f (b)O Ir' 1--'
20 _ (c)
n

MECHANISM OF COLLAGEN-FIBRIL FORMATIONWhen precipitation is complete the ratio of theradii of the largest and smallest fibrils is
(ro)oo I (1-p) dttc
The right-hand side may be computed fromexperimental precipitation curves and the resultsof doing this for several experiments are shown inTable 4. In Figs. 6 and 7 the calculated spread offibril width is compared with the observed distri-bution. One limit of the calculated distributionwas chosen so that the maximum number of fibrilsshould be included and in all cases more than 70%of the fibrils come within the predicted limits,indicating that the observed spread of fibril widthis consistent with the mechanism.
Separation of the nucleation and growth 8tep8Each of the parameters K, and K2, resulting
from analysis of precipitation curves, involves con-stants relating to both nucleation and growth. Theeffect of varying experimental conditions on eachof the steps cannot therefore be determined fromthe variation of K1 and K2 alone. The contributionof growth to K2 can in principle be estimated byanalysis of fibril-width distribution, to give ne, butthe existing electron-microscope data are probablynot sufficiently precise. Information about thegrowth step can, however, be obtained by analternative method. If, in a series of precipitations,nucleation (i.e. lag period) is allowed to proceedunder identical conditions in all experiments theeffect of any subsequent change of conditions mustbe attributed to its effect on the growth step.
Table 3 shows data for a series of experiments inwhich the temperature during nucleation was 250 inall experiments but in which subsequent growth wasallowed to proceed at different temperatures. (Atypical experiment of this type was described in thepreceding paper.)
Table 4. Effect of ionic strength and temperatureon distribution of fibril radius
Experimental conditions: pH 7-1; Co 0.05%. For defini-tions of p, t, (ro)w and (r,)n, see text. Each set of resultswas obtained on a different preparation of collagensolution.
Temp.25025252537
I0-130-230-310-230-23
f(l -p) dtte
1-02551
26-3516*51-4
Now K2 = ksn4, and since n, is constant underthese conditions
d ln (K2) d In k1_ EAd I/T d 1/T R'
where (K2), is the value of K2 when nucleationoccurs at 250, EA is the activation energy forgrowth, T the absolute temperature and R the gasconstant. In introducing EA it is assumed that theshape of the fibrils and the relation betweenaggregating area and surface area are independentof the temperature during growth. From thegraph of log (K2)g against 1/T shown in Fig. 8,EA= 27 kcal./mole.
Included in Table 3 are data for experiments inwhich the whole of the precipitations were carriedout at different temperatures. Under these condi-tions:dlnK2 dln k d lnn_ E1 d ln (r6)
=__ + -_d 1/T d1T d l/T R d 1/T
From the graph of log K2 against 11T shown inFig. 8 and the above value of EA the ratio of theradii of the equivalent fibrils (r.). at 250 and 370 iscalculated to be 1 6. It can be seen from Fig. 7,which shows the distributions of fibril width atthese temperatures, that this figure is of the rightorder of magnitude.
DISCUSSIONThe satisfactory manner in which the simple two-
step mechanism accounts for the shape of theprecipitation curves and the fact that the predic-tions about fibril width are consistent with theelectron-microscope data strongly suggest thatcollagen fibrils form in vitro by the mechanismoutlined above, i.e. by the aggregation of solublecollagen particles to form nuclei, followed by thegrowth of the nuclei into fibrils.
1
be0
1-0 0
o
t( (ro).o(min.) (rc)oo
l.f} 1.QQ 3-3 34 3.55l0 1 M 103/T
10-0 1F61 Fig. 8. Variations of K2 (+) and (K2)g (0) with absolute10-01*60 temperature, T. For definitions of these parameters see2-0 1-80 text.
Vol. 75 60aAV3

G. C. WOODAs discussed in the preceding paper, the nature
of the soluble collagen particles is uncertain; theymay be discrete rod-like collagen molecules oraggregates thereof. The values of the parameter xsuggest that two or three of these particles cometogether to form a nucleus. In view of the asym-metric nature of the collagen molecules the nucleiare probably filamentous; an important point,since the shape as well as the concentration ofnuclei plays an important part in determining therate of precipitation and the size of the ultimatefibrils. The turbidity method used to follow pre-cipitation is, however, unable to give detailedinformation about nucleation, whose precisenature therefore remains unknown.The rate of precipitation during the growth phase
is determined by the number of fibrils and the rateat which each grows. The rate of the latter processis controlled by the rate of aggregation and by thesurface area of the fibrils. Further quantitativeelectron-microscope observations, particularly atdifferent stages during precipitation, would prob-ably be useful in separating these three factors. Itmay be noted that the spread of fibril width, asgiven by equations (10) and (11), should be con-stant throughout precipitation and this conclusionmight also be tested by electron microscopy.The activation energy for growth under one set
of conditions is calculated to be 27 kcal./mole.Bensusan & Hoyt (1958) calculated activationenergies for the over-all precipitation process whichvaried from 23 to 51 kcal./mole according toexperimental conditions. The activation energycalculated by their method from the present data isabout 40 kcal./mole. If precipitation occurs by thetwo-step mechanism the precise meaning of Ben-susan & Hoyt's activation energies is obscure,since they are calculated from the complex result oftwo separate processes. The significance which theyattach to the fact that their activation energy isindependent of ionic strength in formulating amechanism for precipitation may not therefore bejustified. Bensusan & Hoyt observed that themaximum slope of graphs of extinction againsttime were proportional to the square of the collagenconcentration. It may be shown that this resultfollows from the equations for the two-step mech-anism if it is assumed that the final distribution offibril width is independent of collagen concentra-tion: from equation (8)
d-E Eoo(dp EEcoK2CO*P Mna(1 Prlia.)-( dt / \x dt,,/x.
It may be shown that pma.. = 0 4, therefore
(d) = constant x EO K2 CoE, k3 CO= constant x (pnoc)* (r,,)'-0
If (r,)., and hence EOOICO are independent of CO
(d) = constant x C(.
No electron-microscope data are available totest this but since, as described in the precedingpaper, the final extinction (E,,,) of the precipitate iscorrelated with fibril width, the fact that Eo,/Covaries little with collagen concentration supportsthe assumption.In deriving the equation for fibril growth it has
been assumed that reaction of the soluble collagenparticles at the surface of the precipitate is therate-controlling step. Several workers (Nielsen1955, 1958, 1959; Frisch & Collins, 1953; Collins &Leinweber, 1956; Ham, 1958) have derived equa-tions for the rate of precipitation when the diffusionof solute to the surface of the precipitate controlsthe rate and have found that the shape of the pre-cipitation curve is clearly distinguishable from thatobserved when reaction at the surface is rate-controlling. If, under certain experimental condi-tions, diffusion of the soluble collagen particles tothe surface of the fibril were the rate-controllingstep it should be possible to detect this.
SUMMARY
1. Fibril formation is regarded as the result oftwo processes: (a) nucleation, i.e. the aggregation ofsoluble collagen particles to form nuclei, thesmallest aggregates capable ofexisting as a separatephase; (b) growth of the nuclei into fibrils by accre-tion of further soluble collagen particles. It isassumed that reaction of the soluble collagenparticles with the surface of the growing fibrilscontrols the rate of growth and that the formerprocess is proportional to the surface area ofthe precipitate and to the collagen concentra-tion.
2. Equations are derived which qualitativelyaccount for the occurrence of a lag period in pre-cipitation, where the nucleation process predomi-nates over growth, and for the observation that thefinal distribution of fibril width is determinedduring the lag period.
3. The equation for growth gives a precisedescription ofthe shape of precipitation curves overa wide range of conditions. Predictions aboutfibril width are consistent with experimentalobservations.
4. The activation energy for growth is calculatedto be 27 kcal./mole (at pH 7-1, I 0-23, collagenconcentration 0.05 %).
The author wishes to thank Professor R. E. Tunbridgeand Dr D. A. Hall for their interest and encouragementthroughout this work.
604 1960

Vol. 75 MECHANISM OF COLLAGEN-FIBRIL FORMATION 605
REFERENCES
Anderson, T. F. (1956). In Physical Techniques in Bio-logical Research, vol. 3, p. 178. Ed. by Oster, G. &Pollster, A. W. New York: Academic Press Inc.
Bensusan, H. B. & Hoyt, B. L. (1958). J. Amer. chem. Soc.80, 719.
Collins, F. C. & Leinweber, J. P. (1956). J. phys. Chem. 60,389.
Duke, F. R. & Brown, L. M. (1954). J. Amer. chem. Soc.76, 1443.
Frisch, H. L. & Collins, F. C. (1953). J. chem. Phys. 21,2158.
Gross, J. (1956). J. biophys. biochem. Cytol. 2, 261.Gross, J. (1958). J. exp. Med. 107, 265.Ham, F. S. (1958). J. Phys. Chem. Solids, 6, 335.
Jackson, D. S. (1957). Counc. int. Org. Med. Sci. Symp.:Connective Tissue, p. 69. Ed. by Tunbridge, R. E.Oxford: Blackwell Scientific Publications Ltd.
Johnson, R. A. & O'Rourke, J. D. (1954). J. Amer. chem.Soc. 74, 2124.
Nielsen, A. E. (1955). J. Colloid Sci. 10, 576.Nielsen, A. E. (1958). Acta chem. 8cand. 12, 951.Nielsen, A. E. (1959). Acta chem. 8cand. 13, 784.Schmitt, F. 0. (1959). Rev. mod. Phy8. 31, 349.Schmitt, F. O., Gross, J. & Highberger, J. H. (1955a).Symp. Soc. exp. Biol. 9, 326.
Schmitt, F. O., Gross, J. & Highberger, J. H. (1955b).Exp. Cell Re8. Suppl. 3, 326.
Waugh, D. F. (1957). J. cell. comp. Physiol. 49, Suppl. 1,145.
Wood, G. C. & Keech, M. K. (1960). Biochem. J. 75, 588.
Biochem. J. (1960) 75, 605
The Formation of Fibrils from Collagen Solutions3. EFFECT OF CHONDROITIN SULPHATE AND SOME OTHER NATURALLY
OCCURRING POLYANIONS ON THE RATE OF FORMATION
BY G. C. WOODNuffield Gerontological Research Unit, Department of Medicine, University of Leeds
(Received 22 June 1959)
The fact that mucopolysaccharides are abundantin developing connective tissue and the observa-tion that these substances precipitate collagenfibrils from collagen solutions have led to sugges-tions that they may be important in collagenfibrillogenesis (Meyer, 1946, 1955-56; Gross, 1956).Except for brief reports by Gross (1956, 1959)
and Gross & Kirk (1958) little attention has beenpaid to the effect of mucopolysaccharides on thekinetics of fibril formation. These workers foundthat, with two doubtful exceptions, a large numberof preparations of chondroitin sulphate, hyaluronicacid, keratosulphate and heparitin sulphate hadlittle or no effect on the rate of fibril formationunder the experimental conditions they used.Previous work has shown that the process ofcollagen-fibril formation may be divided into twophases (Bensusan & Hoyt, 1958; Wood & Keech,1960), which have been termed nucleation andgrowth (Wood, 1960). In this paper this work isextended to a study of the effect of the presence oflow concentrations of certain mucopolysaccharidesin the reaction mixture on the kinetics of fibrilformation. Particular attention is paid to theeffect of chondroitin sulphate on the overall pre-cipitation process and on the growth step alone. Ithas been suggested (Wood & Keech, 1960) that
different methods of preparing collagen solutionsand different methods of pretreating the samecollagen solution may result in starting materials,for precipitation experiments, of different degreesof molecular aggregation and therefore havingdifferent precipitation properties. A further aim ofthe present work was therefore to investigate thepossibility that the effect of chondroitin sulphate onrate of precipitation depends on the pretreatmentof the collagen solution.The ability to precipitate collagen fibrils is also
shown by the nucleic acids (Gross, 1956; Randall,Booth, Burge, Jackson & Kelly, 1955), which, likethe mucopolysaccharides, are polyanionic innature, and the effect of these substances on therate of fibril formation has therefore also beeninvestigated.
Part of this work has been reported briefly else-where (Wood, 1958).
MATERIALS AND METHODSCollagen solutiofs. For most of the experiments solutions
of collagen (0-1%) in 01M-NaCl containing 005M-sodiumacetate buffer, pH 4*2, were prepared from one sample ofpurified calf dermis (dermis X), as described previously(Wood & Keech, 1960). For a small number of experi-ments solutions were prepared in a similar way from another





























