Embed Size (px)
Citation preview

Louisiana State UniversityLSU Digital Commons
LSU Historical Dissertations and Theses Graduate School
1977
The Genetic Behavior of Resistance in Soybeans tothe Wartelle Race of Root-Knot Nematode.John Kennedy SaichukLouisiana State University and Agricultural & Mechanical College
Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_disstheses
This Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion inLSU Historical Dissertations and Theses by an authorized administrator of LSU Digital Commons. For more information, please [email protected].
Recommended CitationSaichuk, John Kennedy, "The Genetic Behavior of Resistance in Soybeans to the Wartelle Race of Root-Knot Nematode." (1977). LSUHistorical Dissertations and Theses. 3165.https://digitalcommons.lsu.edu/gradschool_disstheses/3165

INFORMATION TO USERS
This material was produced from a microfilm copy of the original document. While the most advanced technological means to photograph and reproduce this document have been used, the quality is heavily dependent upon the quality of the original submitted.
The following explanation of techniques is provided to help you understand markings or patterns which may appear on this reproduction.
1. The sign or "target" for pages apparently lacking from the document photographed is "Missing Page(s)". If it was possible to obtain the missing page(s) or section, they are spliced into the film along with adjacent pages. This may have necessitated cutting thru an image and duplicating adjacent pages to insure you complete continuity.
2. When an image on the film is obliterated with a large round black mark, it is an indication that the photographer suspected that the copy may have moved during exposure and thus cause a blurred image. You will find ja good image of the page in the adjacent frame.
3. When a map, drawing or chart, etc., was part of the material being photographed die photographer followed a definite method in "sectioning" the material. It is customary to begin photoing at the upper left hand corner of a large sheet and to continue photoing from left to right in equal sections with a small overlap. If necessary, sectioning is continued again — beginning below the first row and continuing on until complete.
4. The majority of users indicate that the textual content is of greatest value, however, a somewhat higher quality reproduction could be made from "photographs" if essential to the understanding of the dissertation. Silver prints of "photographs" may be ordered at additional charge by writing the Order Department, giving the catalog number, title, author and specific pages you wish reproduced.
5. PLEASE NOTE: Some pages may have indistinct print. Filmed as received.
University Microfilms International300 North Zeeb RoadAnn Arbor, Michigan 48106 USASt. John’s Road, Tyler’s GreenHigh Wycombe, Bucks, England HP10 QHR

78-7556SAICHUK, John Kennedy, 1950- THE GENETIC BEHAVIOR OF RESISTANCE IN SOYBEANS TO THE WARTELLE RACE OF ROOT-KNOT NEMATODE.
The Louisiana State University andAgricultural and Mechanical College, Ph.D., 1977Agronomy
University Microfilms International , Ann Arbor, Michigan 40106

THE GENETIC BEHAVIOR OF RESISTANCE IN SOYBEANS TO THE WARTELLE RACE
OF ROOT-KNOT NEMATODE
A Dissertation
Submitted to the Graduate Faculty of the Louisiana State University and
Agricultural and Mechanical College in partial fulfillment of the requirements for the degree of
Doctor of Philosophy
in
The Department of Agronomy
byJohn Kennedy Saichuk
B.S., University of Southwestern Louisiana, 1972 M.S., Texas A and M University, 1974
December, 1977

ACKNOWLEDGMENTS
The accomplishments that this dissertation represents could not
have been achieved without the assistance of others.
For their understanding and encouragement during the past three
years, I thank my wife, my family, and my friends.
Special appreciation is extended to Louisiana State University
and the Louisiana Soybean Promotion Board who provided the assistant-
ship which made this endeavor possible.
I am indebted to my advisory committee: Dr. D. F. Gilman, major
advisor; Dr. W. Birchfield, minor advisor; Dr. J. E. Jones, member;
Dr. J. E. Sedberry, member; Dr. M. T. Henderson, member; Dr. B. G.
Harville, member; Dr. J. N. Henshaw, member; and Dr. C. Williams,
former advisor; for their assistance in planning this research pro
ject and compilation of this dissertation.
For their assistance in the field and greenhouse work I thank
Allen Arceneaux, Mike Bickham, Don Fontenot, George Hackman, Warner
Hall, Randall Jemison, Kevin Kilpatrick, and Kilren Vidrine. For
photographic assistance I thank Chuck Greene and Dale Reed. Thanks
also to Bridget Lemoine for assistance with typing.
To my wife, Susette, and my son, John, I dedicate this
dissertation.

TABLE OP CONTENTS
Page
LIST OF TABLES..........................................................iv
LIST OF F I G U R E S ........................................................ v
ABSTRACT.................................................................vi
INTRODUCTION............................................................. 1
LITERATURE R E V I E W ...................................................... 2
MATERIALS AND M E T H O D S .................... 19
RESULTS AND DISCUSSION................................................. 30
Performance of Parental Lines................................... 30
Performance of Cross Combinations...............................33
Bragg X D69-6344........................................... 33
Hill X D69-6344 ........................................... 35
Delmar X D69-6344 ........................................ 35
D69 -6344 X Pickett 7 1 ......................................38
Delmar X Pickett 7 1 ........................................38
Bragg X Pickett 71.......................................... 41
Hill X Pickett 7 1 .......................................... 43
Delmar X Bragg...............................................45
Genetic Model Development........................................45
SUMMARY................... . . 51
LITERATURE CITED........................................................53
VITA..................................................................... 61
i i i

LIST OF TABLES
Table
1
2
3
4
5
6
7
8
9
10
11
12
13
14
Cross Combinations Evaluated
Backcrosses Used in the Study
Nematode Population and Date That Each Cross Combination Was Evaluated
The Number of Observations of Each Population in Each Experiment
Summarized Performance of Parental Lines
Reaction of Parents, F- , and Segregating Generations From the Cross Bragg X D69-6344 to the Wartelle Race of Root-knot Nematode
Reaction of Parents, F , , and Segregating Generations From the Cross Hill X D69-6344 to the Wartelle Race of Root-knot Nematode
Reaction of Parents, F-p and Segregating Generations From the Cross Delmar X D69-6344 to the Wartelle Race of Root-knot Nematode
Reaction of Parents, F-. , and Segregating Generations From the Cross D69-6344 X Pickett 71 to the Wartelle Race of Root-knot Nematode
Reaction of Parents, F , and Segregating Generations From the Cross Delmar X Pickett 71 to the Wartelle Race of Root-knot Nematode
Reaction of Parents, F^, and Segregating Generations From the Cross Bragg X Pickett 71 to the Wartelle Race of Root-knot Nematode
Reaction of Parents, F , and Segregating Generations From the Cross Hill X Pickett 71 to the Wartelle Race of Root-knot Nematode
Reaction of Parents, F , , and Segregating Generations From the Cross Delmar X Bragg to the Wartelle Race of Root-knot Nematode
Proposed Parental Genotypes
iv
Page
24
25
28
29
32
34
36
37
39
40
42
44
46
49

LIST OF FIGURES
Figure Page
1 Parentage of D69-6344 and Hill 20
2 Parentage of Bragg 22
3 Parentage of Delmar 23
4 Response of Parent Lines to Nematode Population 31
v

ABSTRACT
The genetic behavior of resistance in soybeans (Glycine max (L.)
Merrill) to the Wartelle race of root-knot nematode (Meloidogyne
incognita (Kofoid and White) Chitwood was studied by screening F^,
F2 » and backcross populations of eight crosses between selected
parental soybean lines. These lines, 'Bragg1, 'Delmar', 'Hill',
'Pickett 71', and D69-6344, were chosen on the basis of their ancestry
and their reaction to root-knot nematode. Root systems of individual
seedlings, grown in naturally nematode infested soil in a greenhouse,
were examined and given a subjective rating for degree of resistance
to this organism. Segregation patterns were examined and tested for2goodness of fit to expected ratios by X analysis.
Results indicated that susceptibility was dominant to resistance
and that inheritance of reaction was conditioned by genes at two loci
which acted in a duplicate-dominant manner. Where reciprocal Fj's
were tested there was no indication of cytoplasmic or maternal effects.
Variance estimates suggested that the environment or minor genes or
both may influence disease reaction, but this should not prevent
selection for resistance to the Wartelle race of root-knot nematode
in conventional soybean breeding programs.
Delmar, Bragg, and Hill were reported to possess genes for
resistance to other forms of root-knot nematode, but each was sus
ceptible to the Wartlelle race which suggested that they may have
common genes for susceptibility. Delmar and Bragg segregated similarly
when crossed with D69-6344 and did not segregate when crossed with
vi

each other indicating identical genotypes. Resistance in D69-6344 is
thought to have been derived from both Hill and 'Laredo1, a soybean
cultivar reported to be resistant to root-knot nematode.
vii

INTRODUCTION
The root-knot nematode (Meloidogyne incognita (Kofoid and White)
Chitwood) has been a recognized pest of soybeans (Glycine max (L.)
Merrill) since 1882. The nematode has been the greatest threat in
the lighter textured soils of the warmer regions of the United States,
especially the Southeastern United States. Plants damaged by this
organism have reduced growth, fewer and smaller leaves, excessive
wilting in warm weather, and reduction in quantity and quality of
fruit.
In the past, efforts to control this pest have been centered on
the use of cultural practices, such as fallow plowing, rotation to
non-host crops, and the application of nematicides. Recently, more
emphasis has been placed on the development of resistant varieties.
The results generally have been favorable, but in some instances, only
temporary. The existence of new nematode races has shown the need for
the development and release of new resistant varieties.
In 1973 a root-knot nematode was collected on the Wartelle farm
in St. Landry Parish, Louisiana that was identified as Meloidogyne
incognita and was later designated a new race, the Wartelle race,
because of tis inability to reproduce on 'Centennial' sweetpotatoes,
(Ipomea batatis (L.) Lam.), a common host of M. incognita. This race
was investigated because of its ability to parasitize certain soybean
cultivars previously regarded as resistant to root-knot nematode.
This study was designed to determine the number and behavior of genes
involved in inheritance of resistance to the Wartelle race of root-
knot nematode.

LITERATURE REVIEW
Plant damage from root-knot nematodes was first recorded in 1855
by Berkeley (13) who described "Vibrio" attacking the roots of cucum
ber (Cucumis sativus L.) plants growing in a greenhouse in London.
The disease had acquired the names root-knot, beaded root-knot, root-
gall, and big root by 1911. It had been found throughout the United
States, particularly in the southern states, and it was common in
greenhouses throughout the world (14). Neal (76) reported that the
earliest knowledge of root-knot damage in Florida was around 1805,
although the first records did not appear until 1857. Several authors
(37, 66, 67, 68) have reported root-knot nematode in Louisiana,
especially in the lighter textured soils of the state.
The root-knot nematode has a wide host range, encompassing some
2000 plant species. Although all the plant species are not highly
susceptible, they do act as hosts for one or more nematode species.
Often the host is susceptible to one nematode species but is resis
tant to other species (2, 23). According to Good (44), soybeans
(Glycine max (L.) Merrill) was added to the list of host species by
Frank, who discovered root-knot nematodes on roots of soybean plants
grown in a greenhouse in Germany in 1882. Today, root-knot damage is
a potential problem wherever soybeans are grown, and is of considerable
economic importance in the southern United States.
Initial attempts to identify root-knot nematodes were often dis
organized and inconsistent. New names were applied to previously de
scribed nematodes, genera names were changed, and species were shifted

from one genus to another as root-knot damage was observed on addition
al plant species. Atkinson (6) published a list of root-knot nematodes
that had been identified by 1889 which included: Heterodera schachtii,
described by Schacht in Europe in 1859; H. radicicola, first recorded
in 1872 as Anguillula radicicola by Greeff but transferred to the genus
Heterodera by Muller in 1884; H. javanica, found in Java in sugarcane
(Saccharum spp.) roots by Treub; and Meloidogyne spp., found in 1878
in roots of coffee (Coffea spp.) by Goeldi.
In 1919, Kofoid and White (62) described a nematode found in
human fecal samples and called it Oxyuris incognita. According to
Wright (108) this organism was synonymized with H. radicicola by
Sandground. Goodey (45) suggested that the new species H. marioni
had priority due to its earlier application. Chitwood (21) revised
the genus Meloidogyne in 1949 and included M. incognita (Kofoid and
White, 1919), with the former name being preferred for the species.
Other workers (7, 22), acknowledging the earlier work of Goodey as
being correct, referred to H. marioni as the root-knot nematode.
Wester (104), working with root-knot nematodes of lima beans (Phaseolus
lunatus L.), concluded "... it (root-knot) was formerly attributed to
a single species H. marioni (Cornu) Goodey , but it is now recognized
to be caused by a group of species of a separate genus (Meloidogyne
spp.)." Today the revision of the genus Meloidogyne by Chitwood (21)
is still accepted and root-knot nematodes are considered members of
the genus Meloidogyne.
The identity of the nematode itself was often the culprit when
conflicting reports were published. This was demonstrated by the
results of Winstead and Riggs (107) who reported that Bessey (1911)

found 'Striped Blue Ribbon' watermelon (Citrullus vulgaris Schrader ex.
Ecklon and Zeyher) resistant to M. incognita acrita (Chitwood), but in
their studies, watermelon was susceptible to M. incognita acrita.
Winstead and Riggs concluded that Bessey had been working with M. hapla
(Chitwood) or a different population of M. incognita acrita. The prob
lem was recognized by Smith and Taylor (95) who suggested that data
obtained in studies of a particular race of root-knot nematode be
applied only to that nematode rather than to root-knot nematodes in
general.
The identification of root-knot nematode species is based primar
ily on morphological characteristics of the adult female (100).
Ibrahim et al. (55) were able to identify M. javanica and M. incognita
by perineal patterns. In an earlier study Lordello (64) was unable to
distinguish a nematode, whose female perineal patterns closely resem
bled M. incognita, from M. incognita. He used morphological charac
teristics of larvae and males to distinguish the group of nematodes
from M. incognita and concluded that the nematode was a new species.
Dropkin (34) studied varietal response of soybeans to Meloidogyne spp.
using galls and egg mass variation, differences in giant cell formation,
and sex ratios of nematode populations. He found that variation in
galls and egg masses and differences in giant cell formation were
suitable criteria, but that sex ratios were not sufficiently reliable
to distinguish nematode populations. Sasser (82) also used perineal
patterns of adult females to identify nematodes, but indicated that
this method was reliable only in the hands of a specialist. He then
proposed a procedure for identifying unknown nematode populations by
utilizing a series of four test species: peanuts (Arachis hypogaea L.),

watermelon or wheat (Tritlcum aestivum L.) or barley (Hordeum vulgare
L.) or corn (Zea mays L.), pepper (Capsicum spp.), and the Peruvian
tomato (Lycopersicon peruvianum (L„) Miller). Christie (23) also
suggested that vague morphological characteristics were not always
reliable in distinguishing nematode subspecies or races and suggested
a technique like that developed by Sasser as the only sound means of
nematode identification.
Evidence of physiological races of root-knot nematodes has been
reported by several authors (4, 24, 63, 66, 67, 68, 82, 106). Christie
and Albin (24) studied the reaction of 10 host species to 14 nematode
populations that could not be distinguished morphologically. Hosts
varied in reaction and in type of galling to different nematode pop
ulations, Martin (66) described differences in parasitism among
isolates of M. incognita and M. incognita acrita on cotton (Gossypium
hirsutum L.) which ranged from no parasitism to severe parasitism.
Using morphological characteristics of anal plates, Allen (4) found
that four nematode populations isolated in California were all
M. incognita acrita. However, cotton cultivars reacted differently to
the different nematode populations suggesting the existence of races.
Allen hypothesized that the development of different races resulted
from natural selection imposed on nematode populations by various crops.
Martin and Birchfield (67) demonstrated that physiological races of
M. incognita existed which were pathogenic on cotton, sweetpotatoes
(Ipomea batatis (L.) Lam.), lima bean, okra (Hibiscus esculentus L-),
tomato (Lycopersicon esculentum Miller), Crowder pea (Pisum sativum
L.), Amaranthus spp., and gardenia (Gardenia thunbergia L.)„

Cooperative research between Martin and Birchfield (68) and
Williams et al. (106) resulted in isolation of a nematode that caused
severe yield reduction in the soybean cultivar ’Bragg' , which previ
ously had been considered resistant to root-knot nematode. The new
nematode was identified as M. incognita, but it failed to develop
mature females on a susceptible sweetpotato cultivar, so it was des
ignated the Wartelle race for the name of the landowner on whose farm
it was first collected. Boquet et al. (18) studied four populations
of nematodes from Louisiana, including the Wartelle race. Based on
differential reactions of several soybean cultivars, they concluded
that at least two physiological races existed. Of the two'races, the
Wartelle race was the most pathogenic.
Plants infected with root-knot nematodes exhibit symptoms which
are visible in all plant parts. Symptoms include reduced plant growth,
fewer and smaller leaves, excessive wilting in warm weather, and
reduction in quantity and quality of fruit (2). These symptoms are
consequences of destruction of root tissue which prevents normal
uptake of water and minerals. Necrosis of root tissue and prolifera
tion of lateral roots at the point of invasion are associated with
nematode invasion. Internally, the invasion of roots by root-knot
nematodes results in production of giant cells which appear as
abnormal swellings of root tissue (i. e. knots or galls) thus producing
the most characteristic symptom of the disease.
Formation of giant cells was confirmed by Peacock (77) to be a
vital part of the host-parasite relationship involving root-knot
nematodes. Giant cells develop as a result of a nematode secretion
and provide a source of nutrients for feeding nematodes.

Crittenden (30) reported that susceptible host plants generally had the
following characteristics: giant cells surrounding the head of the
nematode, many giant cells present, large giant cell area, dense cyto
plasm in giant cells, many enlarged nuclei in each giant cell, and
enlargement of pericycle with giant cells in this region. Character
istics of resistant hosts were essentially the opposite of susceptible
hosts.
Dropkin and Nelson (36) revealed that galls of soybeans are com
posed of parenchyma originating in the pericycle. They grouped giant
cells into four classes and associated only one class with suscepti
bility. The host-parasite interaction was classed tolerant if both
host and parasite growth was good. The interaction was considered
intolerant if both parasite and host growth was poor. Plants were
considered susceptible when parasite growth was good and host growth
was poor, and resistant when the opposite occurred.
Tyler (104) defined resistance as the ability to obstruct the
invasion of a parasite, and susceptibility as the condition of being a
suitable host for a given parasite. Rohde (78) defined resistance as
a set of characteristics of the host plant which act more or less to
the detriment of the parasite.
Sasser (82) rated fifty plant species and cultivars for reaction
to root-knot nematodes. He defined infection as invasion of the plant
by larvae of root-knot nematodes and immunity as the ability to prevent
infection with the effect of no disease development (i. e. total resis
tance). Resistance to infection was highly variable and was charac
terized by reduced nematode invasion of plant tissues.

In an early study, Sasser and Taylor (84) pointed out three forms
of larval activity which could be equated with resistance. Larvae
could fail to enter roots, enter in reduced numbers and fail to reach
maturity, or enter in large numbers with varying degrees of develop
ment. Resistance was reported to vary from plant to plant because of
these conditions.
Dean and Struble (33) studied plant reaction to root-knot nema
todes in tomatoes and sweetpotatoes and found resistance in sweetpota-
toes to be different from resistance in tomatoes. When root-knbt
larvae invaded either of the two crops, root necrosis was evident as
a mechanism of resistance, but in tomatoes, there was also a reduction
in numbers of invading larvae. There was no substantial difference in
number of larvae entering resistant or susceptible sweetpotato tubers.
Dropkin (35) combined initiation of larval growth, induction of cell
necrosis, and gall formation as criteria for resistance or suscepti
bility of tomatoes to M. incognita. He noticed that when cytokinins
were supplied the response of resistant plants was shifted toward a
susceptible reaction. Sawhney and Webster (86) studied effects of
plant growth hormones on resistance of tomato plants to root-knot and
showed that plant growth hormones played some role in resistance, but
were not the only factor involved. Hutton et. al. (54) noted the
possibility of chemotoxic effects of the roots of 'Siratro' (Phaseolus
atropurpureus D. C.) when lines were screened for root-knot resistance.
Barrons (11) found no significant difference in rate of larval
entry in resistant or susceptible hosts whether the hosts were in
adult or seedling stage. He proposed that root-knot resistance is
due to substance(s) synthesized by the plant that counteract the

giant cell inducing effect of salivary secretions of nematode larvae.
Whether the phenomenon of host specialization was due to genetic dif
ferences in the chemical nature of the salivary secretions of the
nematode rather that to difference in ability of the nematode to enter
the host was not determined.
Minton (72) investigated factors influencing cotton reaction to
root-knot nematodes and reported that resistance was not due to mor
phological differences or toot barriers which prevented penetration,
but to root tip hypersensitivity to penetrating larvae and failure of
root cells to respond to nematodes.
In a review of several papers concerning the nature of nematode
resistance in plants, Rohde (78) concluded: "A resistant plant is
resistant usually for several different reasons, and no one mechanism
can be designated as most important. However, it appears that in most
cases resistance results from upsets in the delicate balance of
reactions that occur between a parasite and its host.1'
According to Rohde (78) resistance became more common as complex
ity of host parasite interaction increased. Therefore, resistance to
root-knot and cyst nematodes, which require the formation of giant
cells in order to feed, was much more common than resistance to ecto-
parasitic nematodes. Malloch (65) felt that plant breeders had two
alternatives in breeding for resistance, find a source of resistance
and make crosses or look for transgressive segregation. Although the
work of Bailey (8) indicated good possibilities for resistant variety
development, he felt a conservative attitude should be taken toward
selection of highly root-knot tolerant plants due to the ability of
nematodes to adapt to a new host species.

10
The interaction between root-knot nematode and wilt-inducing
fungi has long been studied. Smith (94) found an association between
resistance to both root-knot nematode and wilt in cotton. Shepherd
(90) also reported that high root-knot nematode resistance appeared to
provide high resistance to Fusarium wilt in cotton. The level of re
sistance to Fusarium wilt in soybeans grown in the Southeastern United
States was not significantly reduced by root-knot nematodes, but cyst
nematodes caused reduction in resistance according to research by Ross
(80). Rhizoctonia solani Kuhn, has been reported to react with
M. hapla and M. javanica by Taylor and Wyllie (97) and with
M. incognita by Golden and Gundy (42). In all instances, the fungus-
nematode complex had greater detrimental effects than either the fungus
or nematode when inoculated separately. Wyllie and Taylor (109) inoc
ulated 'Harosoy' soybeans with Phytophthora megasperma Drechs. var.
sojae A. A. Hildeb. and M. hapla and obtained more severe damage than
when either pathogen was used alone. When Agarwal and Goswami (1)
investigated the interrelationships between a fungus (Macrophomina
phaseoli (Maubl.) Ashby) and M. incognita in soybeans, significant
synergistic effects were noted when nematode inoculation preceeded
fungus inoculation by three weeks. The nematode predisposed the
fungus making the host much more susceptible than when plants were
infected with the fungus alone. Goswami et. al. (46) also reported a
synergistic virus-nematode (M. incognita) interaction with cowpea
(Vigna unguiculata ssp. unguiculata (L.) Walp.).
Sayre (87) observed the larvae of an amoeba (Theratromyxa weberi)
engulfing M. incognita and introduced another problem for scientists
studying plant nematode reactions. In this study, during the winter

11
of 1975-76, a protozoan identified by Birchfield and Antonopoulos (16)
as Dubosquia penetrans, parasitized some of the root-knot larvae. In
this instance, the parasite did not provide a means of biological con1*
trol, but it could have affected the outcome of the experiment by
reducing the vigor of some nematodes.
Methods for effectively controlling the root-knot nematode have
been sought for many years. As early as 1889, Neal (76) recommended
drainage, frost, fire, sterile soils (in greenhouses), nonuse of land,
nonuse of easily infected crops, insect pathogens, use of vermicide
fertilizers, and use of non-infected stocks as methods of control.
Christie (23), in 1959, listed trap crops, flooding, fallow and dry
tillage, rotations and cover crops, chemicals, and nematode resistant
plants as methods for controlling root-knot nematodes. Sasser (83),
however, pointed out that changes in environment which are sufficient
ly severe to control nematodes often are detrimental for the host
plant. He recommended using a multiple integrated approach which
included good cultural practices, chemicals, and resistant varieties.
Chemicals are often recommended to control root-knot nematodes in
both field and greenhouse conditions (2, 23, 100). However, the use
of chemicals provide an added expense and the benefits derived from
chemical application are usually transitory, requiring subsequent
annual applications for effective control of the nematode population,
"The development of plants possessing genetically controlled
resistance to nematodes has been recognized as one of the most effec
tive and economical means of reducing the losses caused by these pests
especially the root-knot nematodes1' (58), Crittenden (29) suggested
the use of a crop rotation system involving resistant crops for two

12
consecutive years as the best method for controlling M. incognita
acrita. Resistant cultivars used either continuously or in a crop
rotation system also has been suggested by others (14, 15). Good (43)
stated that resistant varieties offered the least expensive and often
only practical means of controlling some nematodes, but he acknowledged
that pathogenic races may result from continued use of resistant
varieties. Sasser (83) felt that a system which incorporated several
means of nematode control would produce the most lasting beneficial
effects since it should reduce the development of physiological races
which often overcome resistant varieties. Kehr (58) suggested that
race development might be reduced by the use of multipte-gene resis
tance coupled with rotations of susceptible, resistant, and non-host
plants.
Resistance is valuable to plant breeders only if it can be trans
mitted from parent to progeny. Weimer and Harter (102) found varietal
resistance in sweetpotatoes to H. radicicola in California in 1925.
In 1931, Isbell (56) discovered two varieties of pole snap beans
(Phaseolus vulgaris L.) which were highly resistant to nematodes.
Frazier and Dennett (38) studied resistance in L. esculentum
lines using gall formation as an expression of resistance. Dominance
of resistance in the was considered high, and resistance in seed
lings was highly correlated with resistance in mature plants.
Lider (63) investigated inheritance of resistance in Vitis spp.
to M. incognita acrita. He reported that resistance was dominant and,
in at least one cross, monogenic in nature. Hare (47) examined F^, F 2 ,
F ^ , and backcross populations of crosses between susceptible bell
peppers and resistant hot peppers. He concluded resistance to

13
M. incognita in pepper was controlled by a single dominant gene and
this same gene also controlled resistance to M. incognita acrita and
possibly M. javanica and M. arenaria (Neal) Chitwood .
Collins and Hagan (25) used pineapple (Ananas sativus Schult.)
root number and length as indicators of resistance to H. radicicola.
No immune varieties were identified. Root number was not influenced by
presence of nematodes, however, root length was reduced. Varying
degrees of resistance were found, which suggested that resistance was
conditioned by several genes and/or environmental influences.
Cordner et al, (26) classified F^ offspring of sweetpotatoes
for reaction to root-knot nematode. In crosses of resistant X suscep
tible plants approximately one third of the progeny were susceptible,
one third intermediate, and one third were resistant. In resistant X
resistant crosses, the ratio of resistant, intermediate, and suscep
tible plants was approximately 5:3:2. When susceptible X susceptible
crosses were made, ten percent of the progeny were resistant, twenty-
five percent were intermediate, and sixty-five percent were susceptible.
Weinberger et al. (103) examined roots of open pollinated peach
(Prunus persica (L.) Batsch.) seedlings after one year of growth in
nematode infested soil. They found that F^ progeny of susceptible X
resistant crosses were resistant, indicating that resistance was
dominant. Resistance to M. incognita and M. javanica in peaches
depends upon different genes according to Sharpe et al. (88) who
reported resistance to M. incognita is monofactorial and dominant.
They seemed uncertain concerning resistance to M. javanica, reporting
only that two or more genes are involved.

14
Kochba and Speigel-Roy (60) reported a high degree of resistance
to root-knot nematode in some bitter almond (Prunus amygdalus Batsch.)
progenies. They suggested that either resistance was dominant to
susceptibility or a cytoplasmic factor conditioned the character. Two
susceptible almond X resistant peach F^ progenies showed almost com
plete dominance of resistance to M. javanica. In a separate study they
found all progenies from four of five reciprocal crosses of resistant
bitter almond X susceptible sweet almond were completely resistant (61).
Progenies of resistant peach X susceptible almond cultivars segregated,
thus they suggested one mode of inheritance of resistance in peach and
another in almond.
Inheritance of root-knot resistance in cotton was investigated
by Wright (108). Using a visual estimate of the proportion of root
tissue damaged by nematodes, he evaluated parents, Fpt and F-j popula
tions of a cross between 'Clevewilt 6 ’ (moderately resistant) and
'Deltapine 15' (susceptible) varieties. He concluded that resistance
was partially dominant and quantitative in nature. Resistance to root-
knot nematode also was reported to be dominant in cotton by Wiles (105).
Shepherd (89) found that inheritance of root-knot resistance in pro
geny of G. barbadense X G. hirsutum appeared to be polygenic.
McFarlane et. al. (69), working with interspecific tomato hybrids,
reported that resistance to the root-knot nematode was dominant and
was controlled by a few factors. Watts (101) used L. peruvianum as a
source of nematode resistance in tomatoes and reported resistance in
early stages of plant growth was controlled by two dominant factors.
Gilbert and McGuire (39) found variations in degree of galling, appar
ently because of wide differences in severity of root-knot infesta

15
tion of commercial tomatoes in Hawaii. In other studies with tomatoes,
resistance was reported to be dominant and conditioned by one major
gene (9, 10, 40, 51, 98).
Sidhu and Webster (91) identified three genes for resistance to
M. incognita in tomato. Resistance was attributed to a single dominant
gene in two cultivars and to a single recessive gene in one cultivar.
There was no evidence for allelism. In a later study of root-knot
nematode - wilt-fungus complex in tomatoes (92), resistance to each
disease was expressed as a single dominant gene which segregated
independently. However, expression of fungus resistance was dependent
on nematode resistance, Sikora et. al, (93) noted that one or more
genes may directly or indirectly affect root-knot nematode resistance
in tomatoes. They proposed screening cultivars in specific areas
before making recommendations for the given area.
Amosu and Frankowiak (5) examined F 2 populations of crosses
involving four susceptible and two resistant cowpea cultivars. They
concluded that resistance to root-knot nematode populations from
Nigeria was governed by a single dominant factor and was augmented by
modifying gene(s).
Based on F 2 segregation ratios of an 'Alabama No. lf (resistant)
X 'Kentucky Wonder' (susceptible) pole bean cross Barrons (12) reported
that three or more genes of equal action condition the reaction to
root-knot nematode. He indicated that a certain minimum number of
genes for susceptibility was necessary before all resistance was lost.
Allard (3), in greenhouse experiments with lima beans, observed
gradation from high resistance to high susceptibility , which indicated
that either several genes, environmental influences, or both, condi

16
tioned resistance to root-knot nematode. In another study with lima
beans, McGuire et. al, (71) indicated that only a few genes were in
volved in root-knot nematode resistance. Because of the presence of
susceptible progeny of resistant X resistant crosses, genes govern
ing resistance were thought to differ from plant to plant.
Boquet e t . al. (18, 19) reported that inheritance of resistance
to M. incognita in soybeans was due to one or two major genes or
one major gene and at least one modifying gene, and that susceptibility
was partially dominant to resistance. These conclusions were based on
screening trials with F^, F 2 , and Fg progeny of a cross between resis
tant D69-6344 and susceptible D69-8178 soybeans.
At least 26 soybean cultivars and 14 strains have been listed as
resistant to M. incognita (18, 19, 27, 28, 29, 32, 41, 44, 49, 55, 59,
74, 75, 81, 96, 99, 106). These genotypes represent several maturity
groups and are an excellent source of germplasm for breeding for root-
knot nematode resistance in soybeans. According to Williams et. al.
(106), resistance in soybeans to the Wartelle race of root-knot nema
tode may be derived from several sources, including 'Hill', Bragg, and
D69-6344. In addition, the cultivar 'Delmar1 has been reported to be
resistant to root-knot nematode (32) and may provide a distinct source
of resistance to root-knot nematode.
Although several authors (31, 84) have found field studies for
resistance in soybeans to various nematodes to be quiet effective,
nematode populations in the field commonly are not uniform, resulting
in questionable results. Romshe (79), for example, had difficulty in
field tests of nematode resistance in tomatoes due to a low nematode
population. Wester (104) compared greenhouse and field methods for

17
evaluating lima beans for resistance to root-knot nematode and con
cluded that the greenhouse method was more rapid and severe than
the field test. He also obtained higher root-knot indices in the
greenhouse test than in the field test. Greenhouse screening was
reported by Shepherd (90) to be more effective than field screening
for transgressive segregation for root-knot nematode resistance in
cotton. McGuire and Allard (70) were even more emphatic when they
stated, "Infestation by root-knot nematodes (Meloidogyne spp.) is
rarely uniform throughout a field. Population densities also tend to
vary from season to season and from year to year in local areas
within the same field. Dependable tests of resistance to root-knot
nematodes are therefore difficult to obtain in field trials." They
attributed the success of their greenhouse tests to favorable climate,
freedom from other pathogens, and uniform infestation.
Several methods have been developed to classify plants for reac
tion to root-knot nematode. Sasser (85) and Boquet (17) preferred
using egg mass indices as a means of classifying resistant and suscep
tible plants. Brodie (20) combined gall development with root necrosis
and the ability of nematodes to reach full maturity to distinguish
between resistant and susceptible upland cotton seedlings. Tyler (99)
was aware of the possibility of escapes in this type of technique and
suggested that the lack of gall formation should not be used as the
sole criterium for resistance. Holston and Crittenden (53) felt that
gall ratings might indicate tolerance and not resistance because
heavily galled and lightly galled plants contained the same nematode
population in their roots. This was sustained by Minton and Parker
(73), but they also reported that root-knot indices were more accurate

than nematode population as an estimator of damage. Gall indices are
often preferred because galls represent the formation of giant cells
which are necessary for nematodes to parasitize the host (30, 72, 78).

MATERIALS AND METHODS
Four commercial soybean cultivars and one advanced breeding line
were selected for this study on the basis of their different reaction
to root-knot nematode. Delmar, Hill, Bragg, and D69-6344, an ad
vanced breeding line selected at Stoneville, Mississippi, have been
reported to be resistant to one or more species of root-knot nematode
{18, 19, 32, 44, 50, 52, 57, 59, 74, 81, 96, 106), whereas,
'Pickett 71' has consistently been reported to be highly susceptible
to the organism (44, 50, 59, 81, 106).
Source of resistance to the root-knot nematode also was a cri
terion for selection of the resistant strains. Bragg, D69-6344,
Hill, and Delmar were selected because they represented different
sources of resistance to some forms of root-knot nematode. Since
1 Haberlandt', one of the grandparents of Hill, has been reported to be
resistant (27, 28, 99), then Hill must have derived its resistance
from Haberlandt (Figure 1). Both Hill and 'Laredo', grandparents of
D69-6344, have been reported to be resistant to root-knot nematode
(19, 27, 41, 44, 49, 57, 75, 96, 99, 106). Since D69-6344 was report
ed to be more resistant to the Wartelle race than either Hill or
Laredo it was likely that both cultivars contributed to the resis
tant characteristic of D69-6344 as was postulated by Williams et. al.
(106) (Figure 1).
Neither D49-2491 nor its parents, CNS and S-100, have been
reported to be resistant to root-knot nematode. 'Tokyo' and
PI 54,610, parents of 'Volstate', which was a parent of 'Jackson',
19

Figure I. Parentage of
D 63-6094
D 69-6344
D 6 9 -6 3 4 4 and Hill
Dunfield
D632-I5
Haberlondt
H ill
D49-2525
S-IQQ
PI 171.442CNS
D 4 9 -2 4 9 I ^ 03-100
Laredo

21
also are not reported to be resistant to root-knot nematode. However,
'Palmetto' and Jackson are reported to be resistant (49, 52, 106).
Hence, Bragg must have obtained its resistance from Palmetto via
Jackson (Figure 2).
Delmar was an Fg out of the cross FC 33243 X C 799 (32). Neither
C 799 nor any of its progenitors have been reported to be resistant to
root-knot nematode. However, FC 33243, a selection out of 'Anderson',
has been reported to be resistant (49) and, therefore, is probably
the source of the genes for resistance to root-knot nematode in
Delmar (Figure 3).
The parental lines were crossed in all possible combinations
with the exception of Hill X Bragg, which was not attempted due to
the absence of a suitable genetic marker (Table 1). Plants selected
at random from each F^ population were backcrossed to the line pos
sessing a dominant genetic marker (Table 2). These and other F^
plants also provided F 2 seed used in the study. F^ Delmar X Hill
plants did not survive, therefore, backcross and F2 seed from this
combination were not obtained. All crossing was accomplished in the
field at the Perkins Road Agronomy Farm in Baton Rouge, Louisiana
during the summers of 1974, 1975, and 1976. Reciprocal F^ seed of
D69-6344 X Pickett 71, Hill X Pickett 71, and Hill X D69-6344
(Table 1) were obtained from Dr. D. J. Boquet of the Northeast
Louisiana Experiment Station, St. Joseph, Louisiana.
In the fall of both 1975 and 1976, Gallion very fine sandy loam
soil naturally infested with the Wartelle race of root-knot nematode
was collected from a field in St. Landry Parish, Louisiana. The soil
was stored in bins in the greenhouse both years. The root-knot

Figure 2. Parentage of
Volstate
Jackson
VolstateBra a a
CNS
D49-249I
Bragg
Tokyo-PI 8 4 2 4 - I t a Mame
54610
PI from China

Figure 3. Parentage of Delmar
FC 33243 Anderson
DelmarPotokg sib PI 70-218-2-19-3
C 799
Lincoln

24
Table 1. Cross combinations evaluated.
Female Male Genetic Marker
Bragg X D69-6344 purple hypocotyl
D69-6344 X Pickett 71* purple hypocotyl
Delmar X D69-6344 purple hypocotyl, brown pubescence
Hill X D69-6344* purple hypocotyl, brown pubescence
Delmar X Pickett 71 purple hypocotyl
Bragg X Pickett 71 purple hypocotyl
Hill X Pickett 71* purple hypocotyl
Delmar X Bragg brown pubescence
*Reciprocal F ' 1 s also evaluated

Table 2. Backcrosses used in the study.
Female Parent Male Parent
Bragg X (F^ Bragg X D69-6344)
Hill X (Ft Hill X D69-6344)
Delmar X (Fx Delmar X D69-6344)
D69-6344 X (Fx D69-6344 X Pickett 71)
Delmar X (F^ Delmar X Pickett 71)
Bragg X (F- Bragg X Pickett 71)
Hill X (Fx Hill X Pickett 71)
Delmar X (F^ Delmar X Bragg)

26
susceptible soybean cultivar, Pickett 71, was grown in the soil during
the fall and winter of each year to maintain an adequate nematode
population.
Each cross combination was evaluated in the same manner. Nema
tode infested soil was mixed and placed in 7.6 cm X 7.6 cm X 6.2 cm
plastic pots. A greenhouse bench was filled with sand, then the pots
were placed in the sand to within 1.5 cm of the top of the pot to
minimize soil temperature and moisture fluctuations. Seeds were
planted at the rate of three per pot for parental- lines and one per
pot for the F-p F 2 , and backcross generations. Pots containing
parental lines were thinned to one plant per pot after emergence. ■ A
minimum soil temperature of 21°C was maintained with electrical heat
ing coils. A minimum air temperature of approximately 27°C was main
tained. Supplemental lighting was provided by three 40 watt flores-
cent lights to increase light quality and intensity.
Two 1000 ml soil samples from each test were evaluated for root-
knot nematode population level by the USDA Nematology Research lab at
Louisiana State University in Baton Rouge. Approximately 500 ml of
soil were mixed with 1000 ml of water and stirred vigorously. The
suspension was strained to remove organic matter, large soil parti
cles, and other soil debris and allowed to stand briefly so larger
sand particles could settle out. The supernatant was strained
through four 300 mesh screens into a petri dish containing water.
The remaining water which contained the nemas was poured into a 50 ml
beaker and standardized to 50 ml, A 10 ml aliquot was placed in a
Syracuse watch glass. Nematodes on one-fourth of the surface area of
the dish were counted using a dissecting microscope. The nematode

27
population was computed by multiplying the actual count by 20. The
nematode densities for each test are shown in Table 3.
Plants were rated for reaction to the root-knot nematode 30 days
after planting using a previously described technique (81). Briefly,
the plants were removed from the pots, washed carefully to remove
adhering soil and other debris from the root systems, and examined
under a 7X dissecting microscope. Plants were rated subjectively
using a scale of 1 to 6, where: 1 = 0-10%, 2 = 11- 20%, 3 = 21-30%,
4 = 31-40%, 5 = 41-50%, and 6 = 51-100% of the roots galled. This
classification scale is a modification of a system used to classify
plant reactions to root-knot nematode by numerous researchers (3, 5,
48, 55, 59, 93, 95, 106). The number of plants examined for each
cross combination are reported in Table 4.
Segregation ratios from each cross combination were evaluated for2goodness of fit to various genetic models using X analysis.

Table J. Nematode population and date that each cross combination was evaluated.
Cross Combination Nemas/500 ml soil Date Examined
Bragg X D69-6344 1980 11/18/76
Hill X D69-6344 6800 10/18/76
Delmar X D69-6344 3240 10/27/76
D69-6344 X Pickett 71 440 02/29/76
Delmar X Pickett 71 1780 12/16/76
Bragg X Pickett 71 2200 12/08/76
Hill X Pickett 71 1410 04/05/76
Delmar X Bragg 5700 11/09/76

Table 4. The number of observations of each population in each experiment.
Cross CombinationFemaleParent
MaleParent *1
Population Recip. Fj Backcross
Bragg X D69-6344 12 10 2 0 340 7
Hill X D69-6344 8 8 10 8 324 27
Delmar X D69-6344 10 8 14 0 321 36
D69-6344 X Pickett 71 10 10 11 10 292 31
Delmar X Pickett 71 12 10 2 0 340 7
Bragg X Pickett 71 9 10 13 0 334 31
Hill X Pickett 71 10 8 8 10 236 31
Delmar X Bragg 8 9 14 0 309 58

RESULTS AND DISCUSSION
Root-gall ratings of all genotypes tended to be larger at higher
nematode densities than at lower densities (Figure 4), as was previ
ously reported (17). D69-6344, for example, consistently rated 1.0
at the lower inoculum levels, but rated slightly above 1.0 at the
higher inoculum levels. Because of the increase in plant infection
at higher nematode population densities, a threshold of approximately
2000 larvae per 500 ml soil was established. At nematode densities
below 2000 per 500 ml soil, only plants with a root-gall rating of 1.0
were classed as resistant; whereas, plants with a rating of both 1.0
and 2.0 were considered resistant at nematode populations above 2000
per 500 ml soil.
Performance of Parental Lines
As an average over all cross combinations, D69-6344 had the low
est mean root-gall index (1.1) and Pickett 71 had the highest (5.9)
(Table 5). These results indicated that D69-6344 was resistant and
Pickett 71 was highly susceptible to the Wartelle race of root-knot
nematode, which agreed with previous reports (17, 18, 19, 81, 106).
Mean root-gall ratings for both Bragg and Hill were 5.5, indicating
that both cultivars were susceptible to the Wartelle race. Root-gall
ratings for Delmar plants ranged from 1.0 to 6.0, however, 23 of the
27 plants evaluated were classed as susceptible (Table 5). The mean
root-gall index for Delmar was 3.8 which indicated that Delmar could
have some genes for resistance to the Wartelle race.
30

Nem
as
/ 50
0 m
l So
il
31
Figure 4. Response of Parent Lines to Nematode Population
7 0 .
65
60 .oo
55.x
35
30
25
20
Root-Gall Index0 6 9 - 6 3 4 4 - # - Bragg-A—
Pickett 71 - o - Hill - □ -Delmar — # —

Table 5, Summarized performance of parental lines.
Parental LineRoot-Gall Indices Classification
Mean Variance1 2 3 4 5 6 S R
Bragg 1 1 7 19 27 1 5.5 1.04
Pickett 71 1 1 36 38 5.9 0.36
D69-6344 29 4 33 1.1 0.33
Hill 2 1 1 14 18 5.5 1.04
Delmar 2 CO 2 6 5 23 4 3.8 1.60
uro

33
Possible sources of the variation associated with Delmar included
seed contamination, although no off-type plants were observed in
Delmar monocultures; susceptibility to a soil pathogen that the other
parental lines were resistant to; and the possibility that Delmar was
heterogeneous for genes conditioning reaction to this race of the
nematode. Since variation among Delmar plants was considerably larger
than that among plants of the other parental lines even though all
plants were exposed to similar environmental conditions, another
likely explanation for the variation was that Delmar simply was more
sensitive to environmental influences than the other genotypes studied.
Performance of Cross Combinations
Bragg X D69-6344. The mean root-gall index for Bragg was 5.2
and for D69-6344 was 1.0 (Table 6). One Bragg plant had a rating of
1.0. This plant presumably was an escape, and was responsible for the
large variance estimate for Bragg (Table 6),
The Fi plants were all highly susceptible, with a mean root-gall
rating of 5.93 (Table 6). The mean response of the population was
not significantly different from that of the susceptible parent, indi
cating that susceptibility to the Wartelle race of root-knot nematode
is dominant to resistance.
Root-gall ratings of the F 2 plants ranged from 1.0 to 6.0, but
were skewed toward the high infection level. When plants rating 1.0
and 2.0 were considered resistant and all other plants were considered
susceptible, the F 2 segregation closely fitted a digenic model (Table
6). Plants from a backcross to the susceptible parent were suscepti
ble. The results indicated that reaction to the Wartelle race in this
cross was conditioned by two genes that acted in a duplicate-dominant

Table 6. Reaction of parents, F^, and segregating generations from the cross Bragg X D69-6344 tothe Wartelle race of root-knot nematode.If
Population Mean Variance
Mo. plants per root-gall index 2/
1 2 3 4 5 6
No. plants grouped 3/
S RExpectedRatio X 2 P
Bragg 5.20 2.40 1 3 6 9 1
D69-6344 1.00 0.00 7 7
Fi 5.93 0.07 1 13 14
F2 5.51 1.49 12 7 12 3 28 262 305 19 15:1 .08 .77
Bragg X F-l 5.83 0.22 1 3 25 29
1/ Nematode population = 1980 nemas/500 ml soil 2/ 1 - 1-10% roots galled, 6 = 51-100% roots galled3/ Plants rated 1 & 2 resistant, 3-6 susceptible

35
manner. The classification of F 2 plants in intermediate infection
classes not shared by parental or plants, which resulted in a
continuous-type F 2 distribution, presumably resulted from either minor
genes, environmental conditions, or both.
Hill X P69-6344. Hill had a mean root-gall index of 5.88;
whereas, the mean index for 069-6344 was 1.13 (Table 7). The F^
population had a mean root-gall index of 6.00, which was not signif
icantly different from that of the susceptible parent. Mean reactions
of the F^ and reciprocal F^ populations did not differ significantly,
indicating no maternal or cytoplasmic effects.
Of the 324 F 2 plants evaluated, 49 were classed as resistant and
275 were classed as susceptible. This segregation pattern did not fit
any of the conventional ratios. The nematode population at which this
cross was evaluated was extremely high (6800 larvae per 500 ml soil),
which undoubtedly resulted in an underestimation of resistant plants.
Although no conventional segregation ratio was detectable, the vari
ation apparent among the F^ plants indicated that the parental lines
differ in reaction to the Wartelle race of root-knot nematode by at
least one gene. There was no segregation in the backcross to the
susceptible parent.
Delmar X D69-6344. Root-gall ratings for Delmar ranged from 2.0
to 6.0 and averaged 4.40 (Table 8). D69-6344 had a mean root-gall
index of 1.38. The mean root-gall index for the F^ population was
5.93.
Root-gall ratings of the F 2 plants ranged from 1.0 to 6.0, but
were skewed toward the high infection level (Table 8). When plants
rating 1.0 and 2.0 were considered resistant and all other plants

Table 7. Reaction of parents, F,, and segregating generations from the cross Hill X D69-6344to the Wartelle race or root-knot nematode.1}
No. plants per root-gall index 2/
No. plants grouped 3/ Expected
Ratio X PPopulation Mean Variance 1 2 3 4 5 6 ' s rHill 5.88 0.13 1 7 8
D69-6344 1.13 0.13 7 1 8
*1 6.00 0.00 10 10
Recip. Fj 5.75 0.21 2 6 8
*2 5.14 3.09 44 5 3 0 28 244 275 49 3:1 16.86 0
Hill X Fx 5.93 0.07 2 25 27
1/ Nematode population = 6800 netnas/500 ml soil 2/ 1 = 1-10% roots galled, 6 = 51-100% roots galled3/ Plants rated 1 Sc 2 resistant, 3-6 susceptible

Table 8. Reaction of parents, Fi, and segregating populations from the cross Delmar X D69-6344to Wartelle race of root-knot nematode.1/
Population
No. plants per root-gall index 2/
Mean Variance 1 2 3 4 5 6
No. plants grouped 3/
S RExpected Ratio X P
Delmar 4.40 2.93 0 2 2 0 2 4 8 2
D69-6344 1.38 0.27 5 3 8
F1 5.93 0.07 1 13 14
*2 5.64 1.30 13 5 3 3 14 283 303 18 15:1 .23 .63
Delmar X 5.64 1.15 1 0 2 0 2 31 35 1
1/ Nematode population 3240 nemas/500 ml soil 2/ 1 = 0-10% of roots galled, 6 = 51-100% roots galled3/ Plants rated 1 Sc 2 resistant, 3-6 susceptible

38
were considered susceptible, the F 2 segregated in a 15:1 ratio for
susceptibility and resistance (Table 8). There was no detectable
segregation pattern in the population from the backcross to Delmar.
The results indicated that Delmar and D69-6344 differed in reaction to
the Wartelle race at two loci.
D69-6344 X Pickett 71. The mean root-gall rating for D69-6344
was 1.00 and for Pickett 71 was 5.60 (Table 9). One Pickett 71 plant
had a root-gall index of 3.0, however, galls present on the plant had
large egg-masses, which indicated the plant was susceptible, but that
the low inoculum level (440 larvae per 500 ml soil) prevented a reli
able disease classification.
Mean root-gall ratings of the F^’s and their reciprocals were not
significantly different and were not different from that of the sus
ceptible parent. The results indicated that susceptibility was dom
inant to resistance and that there were no maternal or cytoplasmic
effects.
Of the 292 F2 plants rated, 226 were classed as susceptible and
66 were classed as resistant, which gave a reasonably good fit to a
3:1 ratio (Table 9). The backcross to the resistant parent segregat
ed in a 1:1 ratio for susceptibility and resistance. Results indi
cated that reaction to the Wartelle race in this cross was conditioned
by one major gene pair. Variation apparent in the F 2 generation in
dicated that minor genes, environmental conditions, or both, also were
involved in the disease reaction.
Delmar X Pickett 71 . The mean root-gall rating for Delmar was
3.42 and for Pickett 71 was 6.00 (Table 10). All ten of the
Pickett 71 plants had root-gall ratings of 6,00. The Delmar plants,
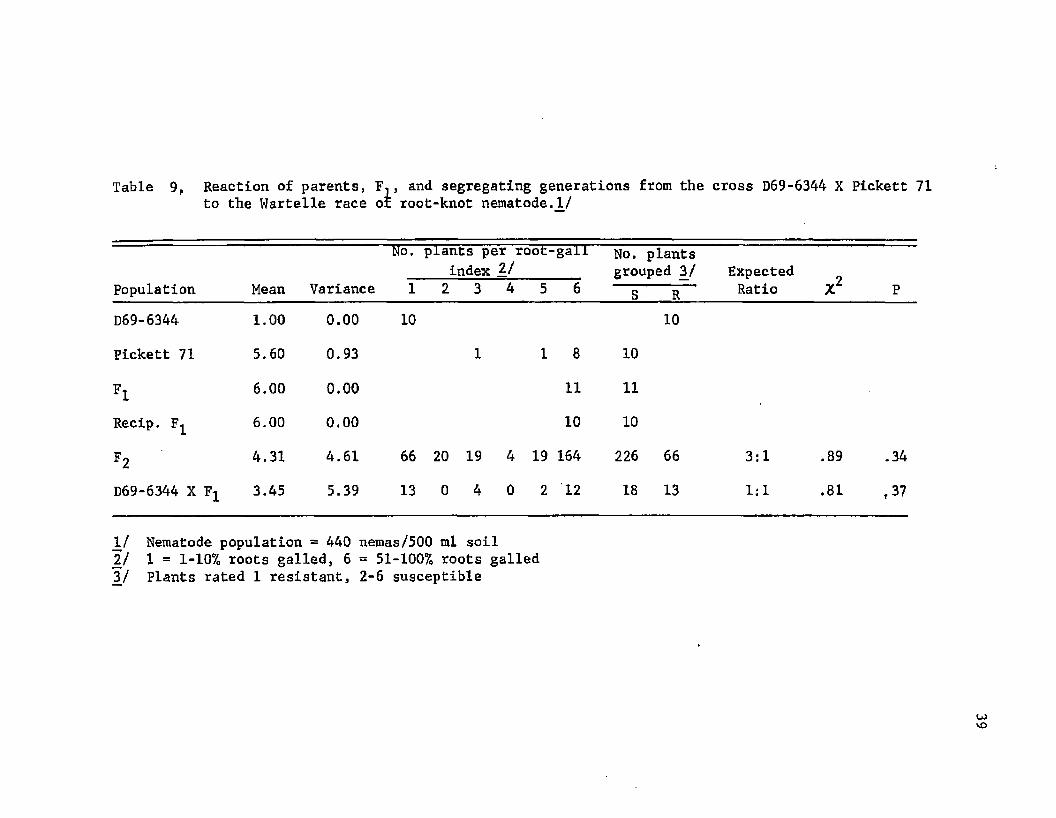
Table 9, Reaction of parents, F,, and segregating generations from the cross D69-6344 X Pickett 71to the Wartelle race or root-knot nematode.1/
No. plants per root-galT index 2f
No. plants grouped 3/ Expected
Population Mean Variance 1 2 3 4 5 6 S R Ratio x2 P
D69-6344 1.00 0.00 10 10
Pickett 71 5.60 0.93 1 1 8 10
F1 6.00 0.00 11 11
Recip. F 6.00 0.00 10 10
F2 4.31 4.61 66 20 19 4 19 164 226 66 3:1 .89 .34
D69-6344 X Fjl 3.45 5.39 13 0 4 0 2 12 18 13 1:1 .81 ,37
I f Nematode population = 440 nemas/500 ml soili f 1 = 1-10% roots galled, 6 = 51-100% roots galled3/ Plants rated 1 resistant, 2-6 susceptible
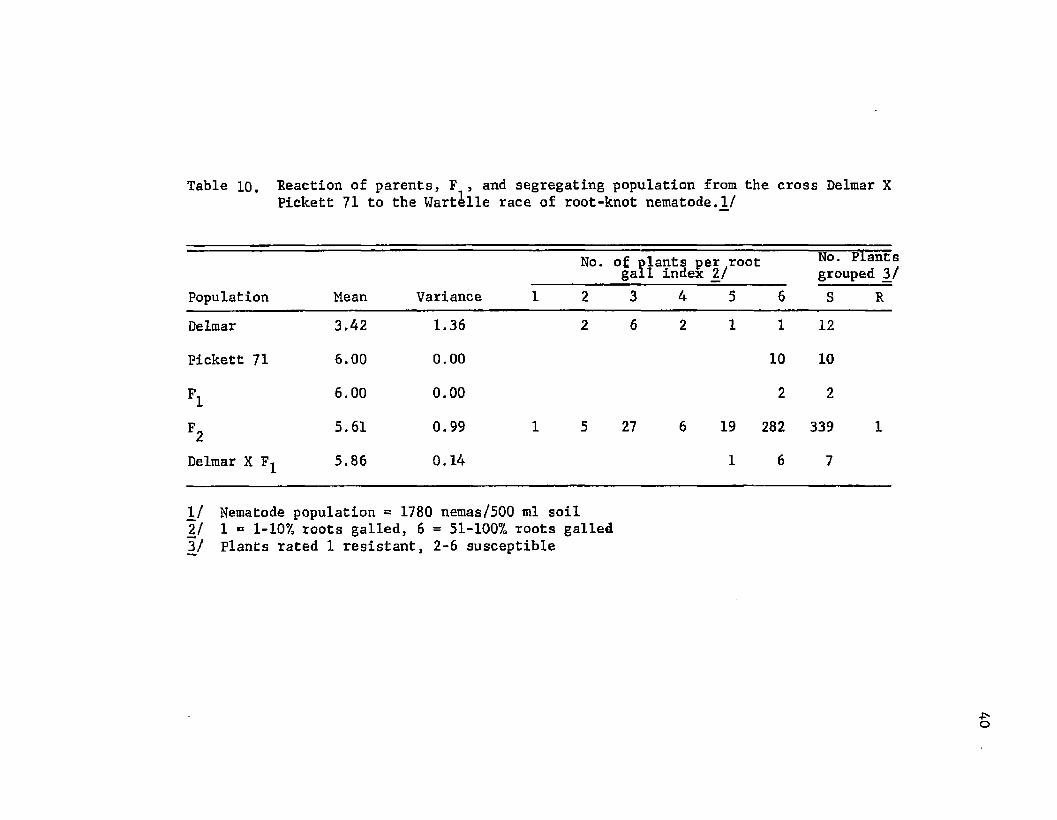
Table 10. Reaction of parents, F , and segregating population from the cross Delmar XPickett 71 to the Wartille race of root-knot nematode.If
Population Mean Variance
No. of plants per root gall index 2/No. Plants grouped 3/
1 2 3 4 5 6 S RDelmar 3.42 1.36 2 6 2 1 1 12
Pickett 71 6.00 0.00 10 10
Fi 6.00 0.00 2 2
F2 5.61 0.99 1 5 27 6 19 282 339 1
Delmar X F^ 5.86 0.14 1 6 7
1/ Nematode population = 1780 nemas/500 ml soil 2/ 1 ** 1-10% roots galled, 6 = 51-100% roots galled3/ Plants rated 1 resistant, 2-6 susceptible

41
however, ranged in reaction from 2.0 to 6.0, with a preponderance of
plants having ratings at the intermediate infection level.
The mean root-gall rating for the F- population was 6.00, which
was not significantly different from the mean rating of the suscep
tible parent. Even though both parents were considered susceptible,
there were detectable differences in degree of susceptibility between
the two cultivars. Galls on Pickett 71 roots were larger and more
numerous than those on Delmar. The resemblence of reaction of F^
seedlings to those of Pickett 71 seedlings suggested that genes for
susceptibility which were contributed by Pickett 71 were dominant to
gene(s) contributed by Delmar.
Root-gall ratings of the F 2 plants ranged from 1.0 to 6.0, but
were skewed toward the high infection level. Only one of the 340
F 2 plants was rated as resistant and that plant presumably was an
escape. There also was no segregation in the population which re
sulted from the backcross to Delmar.
Bragg X Pickett 71. Root-gall indices averaged 5.56 for Bragg
and 6.00 for Pickett 71 (Table 11). Although all Bragg and Pickett 71
plants were susceptible, differences in type and severity of infection
were apparent. Root-galls were much larger and more numerous on
Pickett 71 plants than on Bragg plants, which denoted a genotypic
difference between the two cultivars. This was not surprising since
Bragg has been reported to be resistant to many forms of root-knot
nematode (18, 44, 59, 74, 96), whereas Pickett 71 has consistently
been reported to be susceptible (44, 50, 59, 81, 106).

Table 11. Reaction of parents, F]_, and segregating generations from the cross Bragg XPickett 71 to the Wartelle race of root-knot nematode.1/
Population Mean Variance
iJo. plants per root Gall index 2/
No. Plants grouped 3/
1 2 3 4 5 6 S R
Bragg 5.56 0.53 1 2 6 9
Pickett 71 6.00 0.00 10 10
Fi 6.00 0.00 13 13
f2 5.99 0.00 1 333 334
Bragg X F1 6.00 0.00 31 31
1/ Nematode population « 2200 nemas/500 ml soil2/ 1 = 1-10% roots galled, 6 = 51-100% roots galled3 / Plants rated 1 & 2 resistant, 3-6 susceptible

43
The F^ plants were all susceptible, with a mean root-gall rating
of 6,00, and all plants resembled Pickett 71 in infection character
istics .
The and backcross generation seedlings were all susceptible,
but differences in reaction were observed within each population. The
F 2 plants segregated in a ratio of 3 Pickett 71 infection type to 19Bragg infection type (X = ,20, P = .66), The population which
resulted from the backcross to Bragg segregated in a ratio of 12Pickett 71 type to 1 Bragg type (X = .29, P = ,59). The results
indicated that Bragg and Pickett 71 differed in reaction to the
Wartelle race by one gene pair.
Hill X Pickett 71. Both Hill and Pickett 71 were susceptible,
with mean root-gall indices of 5.20 and 6.00, respectively (Table 12),
Again, there was a difference between the two cultivars in degree of
reaction, with Pickett 71 exhibiting a more susceptible reaction than
Hill.
The F^'s and their reciprocals were all susceptible and more
closely resembled Pickett 71 than Hill. This suggested that Pickett
71 possessed genes which played a greater role in expression of dis
ease reaction than Hill. The similar reactions of the F^ and recip
rocal seedlings indicated no maternal or cytoplasmic effects.
The backcross generation was completely susceptible, however,
there was variation in reaction among seed lings from the severity of
Pickett 71 to the mildness of Hill. The population ranged in
reaction from 1.0 to 6.0, but only one of the 236 plants evaluated had
a rating of 1.0. That plant presumably was an escape. Tn all

Table 12, Reaction of parents, F^, and segregating generations from the cross Hill X Pickett 71 to the Wartelle race of root-knot nematode.1/
No. plants per root Gall index 2/
No. Plants 3 /groupedPopulation Mean Variance 1 2 3 4 5 6 S R
Hill 5.20 1.32 2 1 7 10
Pickett 71 6.00 0.00 8 8
F1 6.00 0.00 10 10
Reciprocal 6.00 0.00 8 8
F2 5.88 0.34 1 1 2 5 4 223 235 1
Hill X 6.00 0.00 31 31
17 Nematode population - 1410 nemas/500 ml soil 2/ 1 = 1-10% roots galled, 6 = 51-100% roots galled_3/ Plants rated 1 resistant, 2-6 susceptible

- 45
likelihood a segregation pattern similar to that of the Fg population
of Bragg X Pickett 71 existed but these data were not obtained.
Delmar X Bragg. The mean root-gall indices for Bragg and Delmar
were 5.78 and 4.39, respectively (Table 13). Two Delmar seedlings
were rated as resistant but these seedlings had damaged root systems,
which might have prevented detection of the disease expression.
The F^ plants were all highly susceptible, with a mean root-gall
index of 6.0 (Table 13). Root-gall indices of the F2 population
ranged from 1.0 to 6.0, but only two of the 309 plants evaluated were
rated as resistant. These two plants were thought to be escapes. The
backcross population had a mean root-gall rating of 5.64. Three
plants from the backcross population had a root-gall rating of 1.0.
These plants also had damaged root systems and probably would have
had susceptible reactions if their root systems had been normal.
Genetic Model Development
The highly susceptible reaction of F^ plants in all cross combi
nations indicated that susceptibility to the Wartelle race of root-
knot nematode is dominant to resistance. Boquet et a l , (19) reported
that susceptibility to the Wartelle race in the soybean cross D69-6344 .
X D69-8178 was partially dominant to resistance. They found that F^
plants were moderately resistant at low nematode population densities,
moderately susceptible at medium densities, and highly susceptible at
higher densities. Nematode densities used in this study were, with
the exception of the cross D69-6344 X Pickett 71, comparable to or
larger than the highest nematode density used by Boquet et al. (19).
Therefore, F^ plant reactions which were determined at comparable

Table 13. Reaction of parents, Fj_, and segregating populations from the cross Delmar XBragg to the Wartelle race of root-knot nematode.1/
No . plants per r< Gall index 2/ DOt No. Plants grouped 3/Population Mean Variance 1 2 3 4 5 6 S RDelmar 4.38 2.13 2 3 3 2
Bragg 5.78 0.19 2 7 9
F1 6.00 0.00 13 13
F2 5.95 0.20 2 0 1 0 3 303 307 2
Delmar X 5.64 1.39 3 0 1 0 3 51 55 3
J./ Nematode population = 5700 nemas/500 ml soil2/ 1 = l-10?o roots galled, 6 = 51-100% roots galled3/ Plants rated 1 & 2 resistant, 3-6 susceptible

47
nematode densities were very similar in the two studies. Suscepti
bility to the Wartelle race appeared to be dominant to resistance
at nematode densities at which plants are often exposed to in the
field.
Data from the cross D69-6344 X Pickett 71 indicated that reaction
to the Wartelle race was conditioned by a single gene pair. The Fg
segregation pattern of the cross Hill X D69-6344 approximated that
expected for a single gene model, however, there was a deficiency
of plants in the resistant class, which was presumably a result of
the extremely high nematode population (6800 larvae per 500 ml soil)
at which the cross was evaluated. If the high nematode population
caused the smaller than expected number of plants in the resistant
class by causing some resistant plants to appear susceptible, as
has been previously reported (19), then the results indicated that
D69-6344 and Hill also differ in reaction to the Wartelle race by
genes at a single locus. Although progeny of the cross Hill X
Pickett 71 were susceptible, parental types were detected in both the
F2 and backcross populations. These results indicated that the genes
for susceptibility possessed by Hill and Pickett 71 were allelic.
Resistance possessed by D69-6344 was recessive to the susceptibility
of both Hill and Pickett 71, and the susceptible reaction of Hill was
recessive to the highly susceptible reaction of Pickett 71,
When both Bragg and Delmar were crossed with D69-6344, the F2
populations segregated in a 15:1 ratio for susceptibility and resis
tance (Tables 6 and 8, pages 34 and 37). This indicated that reaction
to the Wartelle race in these two crosses was conditioned by two gene
pairs. Although Delmar varied in disease reaction more than Bragg,

the preponderance of Delmar seedlings were as susceptible as Bragg,
which indicated that the two cultivars were identical in genotype for
this character. Progeny from the cross Delmar X Bragg were suscepti
ble and there was no evidence for parental type segregation in either
the F2 or backcross population, which gave additional evidence that
Bragg and Delmar had identical genes for reaction to the organism.
All progeny from the cross Bragg X Pickett 71 were susceptible, which
indicated that Bragg and Pickett 71 had a common gene for suscepti
bility. Based on the performance of the parental lines and the
segregation ratios obtained in the crosses, the following genotypes
were apparent: D69-6344 - aabb, Bragg and Delmar - AABB, Hill - AAbb,
and Pickett 71 - A'A'bb (Table 14).
Boquet et al. (19) reported that inheritance of reaction to the
Wartelle race of root-knot nematode was conditioned by one major gene
and at least one modifying gene. Results obtained in this study,
however, indicated that inheritance of reaction to the Wartelle race
was conditioned by major genes at two loci that act in a duplicate-
dominant manner. Three alleles appeared to exist at one locus and two
alleles at the other. Variation among progeny of several crosses
exceeded that of the parental lines, which indicated that minor genes,
environmental conditions, or both, also condition the disease reaction.
Bragg, Hill, Delmar, and D69-6344 were selected for the study,
in part, to represent different sources of resistance to root-knot
nematode. Resistance possessed by Delmar, Hill, and Bragg were thought
to have been derived from FC 33243, Haberlandt, and Palmetto, respect
ively, whereas, resistance possessed by D69-6344 had been suggested to
have been derived from both Hill and Laredo (106). The susceptible

Table 14. Proposed parental genotypes.
Parental Line Genotype Classification
Delmar AABB susceptible
Bragg AABB susceptible
Hill AAbb susceptible
Pickett 71 A ’A'bb highly susceptible
D69-6344 aabb resistant

disease reaction of Delmar and Bragg in this study indicated that
genes possessed by FC 33243 and Palmetto did not condition resistance
to the Wartelle race of root-knot nematode. Although Hill was suscep
tible to the Wartelle race, D69-6344, a second generation selection
from a cross involving Hill, was highly resistant. These results
indicated that the resistance derived from Haberlandt by Hill condi
tioned resistance to the Wartelle race when in some genetic back
grounds. The hypothesis that resistance possessed by D69-6344 appear
ed to be derived from Hill and Laredo was supported by the fact that
the cross between Hill and D69-6344 segregated for only one gene.
Since the results indicated that reaction to the Wartelle race was
conditioned by major genes at two loci, D69-6344 and Hill appeared to
have one gene for resistance in common.

SUMMARY
The purpose of this investigation was to study the number and
behavior of genes for resistance in soybeans to the Wartelle race of
root-knot nematode.
Five parental lines were selected on the basis of their ancestry
and their reaction to root-knot nematode. Eight cross combinations
were made in the field and the resulting populations were grown in
nematode infested soil in a greenhouse. Root systems of individual
seedlings of these populations were examined and rated subjectively
for degree of resistance to this organism. The segregation patterns2were tested for goodness of fit to expected ratios by X analysis.
Results indicated that susceptibility to the Wartelle race of
root-knot nematode in soybeans was dominant to resistance. Inheri
tance of reaction to the Wartelle race was conditioned by genes at
two loci, which acted in a duplicate-dominant manner. Similar reac
tions of reciprocal F ^ 1s suggested that no maternal or cytoplasmic
effects were involved. Variance estimates suggested that minor genes
or environmental conditions or both may have contributed to the dis
ease reaction. However, the role of either the environment or minor
genes should not prevent selection for resistance in soybeans to the
Wartelle race of root-knot nematode in conventional breeding programs.
Delmar, Bragg, and Hill were all susceptible to the Wartelle
race and were thought to possess common genes for susceptibility even
though they were reported to derive genes for resistance to other
root-knot nematodes from separate sources. Delmar and Bragg were
51

thought to have identical genotypes while the common ancestry of
D69-6344 and Hill suggested that D69-6344 obtained genes for resis
tance from Hill, as well as, the root-knot resistant cultivar Laredi

LITERATURE CITED
1. Agarwal, D. K. and B. K. Goswami. 1973. Interrelationships between a fungus, Macrophomina phaseoli (Maubl.) Ashby and root- knot nematode Meloidogyne incognita (Kofoid and White) Chitwood in soybean Glycine max (L.) Merrill. Proc, Indian Natl. Sci. Acad. 39,B:701-704.
2. Agrios, G. N. 1969. Plant Pathology. Academic Press, New Yorkand London. 629 p.
3. Allard, R. W. 1954. Sources of root-knot nematode resistancein lima beans. Phytopath. 44:1-4.
4. Allen, M. W. 1952. Observations on the genus Meloidogyne.Proc. Helm. Soc, Wash. 19:44-51.
5. Amosu, J. 0. and J. D. Franckowiak. 1974. Inheritance of resistance to root-knot nematode in cowpea. Plant Dis. Reptr, 58:361-363.
6. Atkinson, G. F. 1889. A preliminary report upon the life history and metamorphosis of a root-gall nematode, Heterodera radicicola (Greeff) Mull., and the injuries caused by it upon the roots of various plants, Ala. Agr. Exp. Sta. Bull. No, 9:177-228.
7. Atkinson, R. E. 1944. Diseases of soybeans and peanuts in theCarolinas in 1943. Plant Dis. Reptr. Suppl. 148:254-259.
8. Bailey, D. M. 1941. The seedling test method for root-knotnematode resistance. Proc. Am. Soc. Hort. Sci. 38:573-575.
9. Barham, W. S. and J. N. Sasser. 1956. Root-knot nematoderesistance in tomatoes. Proc. Assoc, of South. Agr. Workers.53:150.
10. . and N. N. Winstead. 1957. Inheritance of resistance to root-knot nematodes in tomatoes. Proc. Am. Soc. Hort. Sci. 69:372-377.
11. Barrons, K. C. 1939. Studies of the nature of root-knot resistance. J. Agr. Res. 58:263-271.
12. . 1940. Root-knot resistance in beans. J. Hered.31:35-38.
13. Berkeley, J. J, 1855. Vibrio forming cysts on the roots of cucumbers. Gardner's Chronicle, April 7. p. 220.
53

5414.
15.
16.
17.
18.
19.
20 .
2 1.
22.
23.
24.
25.
26.
Bessey, E. A. 1911. Root-knot and its control. USDA Bur. of Plant Ind. Bull. No. 217.
______________ and L. P. Byars. 1915. The control of root-knot.USDA Farmer's Bull. No. 648.
Birchfield, W. and A. A. Antonopoulos. 1976. Scanning electronmicroscope observation of Dubosquia penetrans parasitizing root- knot larvae. J. Nematol. 8:272-273.
Boquet, D. J. 1974. Inheritance of resistance to the Wartelle race of root-knot nematode in soybeans. Unpublished manuscript of Doctor of Philosophy dissertation, Louisiana State University, Baton Rouge, Louisiana.
________________ C. Williams, and W. Birchfield. 1975. Resistance in soybeans to five Louisiana populations of the root-knot nematode. Plant. Dis. Reptr. 59:197-200.
______________ , C. Williams, and W. Birchfield. 1976. Inheritance of resistance to the Wartelle race of root-knot nematode in soybeans. Crop Sci. 16:783-785.
Brodie, B. B., L. A. Brinkerhoff, and F. B. Struble. 1960. Resistance to the root-knot nematode, Meloidogyne incognita acrita, in upland cotton seedlings. Phytopath. 50:673-677.
Chitwood, B. G. 1949. Root-knot nematodes - Part I. A revision of the genus Meloidogyne Goeldi, 1887. Proc. Helmintholog. Soc. Wash. 16:90-104.
Christie, J. R. 1936. The development of root-knot nematode galls. Phytopath. 26:1-22.
. 1959. Plant nematodes. Their bionomics andcontrol. H. and S. B. Drew Co., Jacksonville, Fla. 256 p.
_________________ and F. E. Albin. 1944. Host parasite relationships of the root-knot nematode, Heterodera marioni I. The question of races. Proc. Helmintholog. Soc. Wash. 11:31-37.
Collins, J. L. and H. R. Hagan. 1932. Nematode resistance of pineapples: Varietal resistance of pineapple roots to the nematode Heterodera radicicola (Greeff) Muller. J. Hered.23:503-508.
Cordner, H. B., F. B. Struble, and L. Morrison. 1954. Breeding sweetpotatoes for resistance to the root-knot nematode. Plant Dis. Reptr. Suppl. 227:92-93.
27. Crittenden, H. W. 1954. Factors associated with nematode resistance in soybeans. Phytopath. 44:388 (abstr.).

55
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
Crittenden, H. W. 1955. Root-knot nematode resistance of soybeans. Phytopath, 45:347 (abstr.).
__________________. 1956, Control of Meloidogyne incognita acritaby crop rotations. Plant Dis. Reptr. 40:977-980.
__________________. 1958. Histology and cytology of susceptibleand resistant soybeans infected with Meloidogyne incognita var. acrita. Phytopath. 48:461 (abstr.).
__________________. 1961. Studies of the host range ofMeloidogyne incognita acrita. Plant Dis. Reptr. 45:190-191.
____________________and R. H. Cole. 1967. Registration of Delmarsoybeans. Crop Sci. 7:279.
Dean, J. L. and F. B. Struble. 1953. Resistance and susceptibility to root-knot nematodes in tomato and sweetpotato. Phytopath. 43:290 (abstr.).
Dropkin, V. H. 1959. Varietal response of soybeans to Meloidogyne - A bioassay system for separating races of root- knot nematodes. Phytopath. 49:18-23.
________________, J. P. Helgeson and C. D. Upper. 1969. Thehypersensitivity reaction of tomatoes resistant to Meloidogyne incognita: Reversal by cytokinins. J. Nematol. 1:55-61.
________________ and P. E. Nelson. 1960. The histopathology ofroot-knot nematode infections in soybeans. Phytopath .50:442-447.
Fielding, M, J. and J. P. Hollis. 1956. Occurrence of plant parasitic nematodes in Louisiana soils. Plant Dis. Reptr. 40:403-405.
Frazier, W. A. and R. K. Dennett. 1949. Isolation of Lycopersicon esculentum type tomato lines essentially homozygous resistant to root-knot. Proc. Am. Soc. Hort. Sci. 54:225-236.
Gilbert, J. C. and D. C. McGuire. 1952. Root-knot resistance in commercial type tomatoes in Hawaii. Proc. Am. Soc. Hort. Sci. 60:401-411.
____________________________________ . 1956. Inheritance of resistance to severe root-knot from Meloidogyne incognita in commercial type tomatoes. Proc. Am. Soc. Hort. Sci. 68:437-442.
Godfrey, G. H. 1923. Root-knot: Its cause and control. USDA Farmer's Bull. 1345.

5642.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
Golden, J. K. and S. D. Van Gundy. 1975. A disease complex of okra and tomato involving the nematode, Meloidogyne incognita, and the soil-inhabiting fungus, Rhizoctonia solani. Phytopath. 65:265-273.
Good, J. M. 1972. Management of plant parasitic nematode populations. Tall Timbers Confer. Ecol. Aninv. Control Habitat Manage. 4:109-127.
___________ . 1973. Nematodes. In B. E. Caldwell (ed.) Soybeans:Improvement, production, and uses. Agronomy 16:527-543. Am.Soc. of Agron., Madison, W I .
Goodey, T. 1932. On the nomenclature of the root-gall nematodes. J. Helminthology. 10:21-28.
Goswami, B. K . , S. Singh, and V. S. Verma. 1974. Interaction of a mosaic virus with root-knot nematode Meloidogyne incognita in Vigna sinensis. Nematologica 20:366-367.
Hare, W. W. 1956. A major gene for resistance to root-knot nematodes in pepper. Phytopath. 46:14 (abstr.).
Hartmann, R. W. 1971. Inheritance of resistance to root-knot nematodes (Meloidogyne incognita) in beans (Phaseolus vulgaris). J. Am. Soc. Hort. Sci. 96:344-347.
Hartwig, E. E. 1973. Varietal development. In B. E„ Caldwell (ed.) Soybeans: Improvement, production, and uses. Agronomy 16:187-210. Am. Soc. of Agron., Madison, WI.
________________, J. M. Epps, and C. J. Edwards, Jr. 1971.Registration of Pickett 71 soybeans. Crop Sci. 11:603.
Hernandez, T. P., J. C. Miller, and M. J. Giamalva. 1965. Inheritance of resistance to root-knot nematodes, Meloidogyne incognita group, in tomatoes. Proc. Am. Soc. Hort. Sci. 87:412-414.
Hinson, K. and E. E. Hartwig. 1964. Bragg and Hardee soybeans. Crop Sci. 4:664.
Hoiston, E. M. and H. W. Crittenden. 1951. Resistance in soybeans to root-knot nematode. Phytopath. 41:562 (abstr.).
Hutton, E. M., W. T. Williams, and L. B. Beall. 1972. Reactions of lines of Phaseolus atropurpureus to four species of root-knot nematode. Aust. J. Agr. Res. 23:623-632.

5755.
56.
57.
58.
59.
60.
61.
62.
63.
64.
65.
66.
67.
68.
Ibrahim, I. K. A., I, A. Ibrahim, and S. I. Massoud. 1972. Induction of galling and lateral roots on five varieties of soybeans by Meloidogyne javanica and M. incognita. Plant Dis. Reptr. 56:882-884.
Isbell, C. L. 1931. Nematode resistance studies with pole snap beans. J. Hered. 22:191-198.
Johnson, H. W. 1960. Registration of soybean varieties, VII. Agron. J. 52:659-660.
Kehr, A. E. 1966. Current status and opportunities for the control of nematodes by plant breeding. In Pest control by chemical, biological, genetic, and physical means. Agr. Res. Serv. 33-110:126-138.
Kinloch, R. A. and K. Hinson. 1973. The Florida program for evaluating soybean (Glycine max (L.) Merr.) genotypes for susceptibility to root-knot nematode disease. Soil Crop Sci. Soc. Fla., Proc. 32:173-176.
Kochba, J. and P. Speigel-Roy. 1972. Resistance to root-knot nematode in bitter almond progenies and almond X Okinawa peach hybrids. Hortsci. 7:503-505.
________________________________ . 1975. Inheritance of resistanceto the root-knot nematode (Meloidogyne javanica Chitwood) in bitter almond progenies. Euphytica 24:453-457.
Kofoid, C. A. and A. W. White. 1919. A new nematode infection of man. J. Am. Med, Assoc. 72:567-569.
Lider, L. A. 1954. Inheritance of resistance to a root-knot nematode (Meloidogyne incognita var. acrita Chitwood) in Vitis spp. Proc. Helmintholog. Soc. Wash. 21:53-60.
bordello, L. G. E. 1955. Nematodes attacking soybean in Brazil. Plant Dis. Reptr. 39:310-311.
Malloch, W. S. 1923. The problem of breeding nematode resistant plants. Phytopath. 13:436-450.
Martin, W. J. 1954. Parasitic races of Meloidogyne incognita and Meloidogyne incognita var acrita. Plant Dis. Reptr. Suppl. 227:86-89.
_______________ and W. Birchfield. 1955. Notes on plant parasiticnematodes in Louisiana. Plant Dis. Reptr. 39:3-4.
___________________________________. 1973. Further observations ofvariability in Meloidogyne incognita on sweetpotatoes. Plant Dis. Reptr. 57:199.

58
69.
70.
71.
72.
73.
74.
75.
76.
77.
78.
79.
80.
81.
82.
McFarlane, J, S., E. Hartzler and W. A. Frazier. 1946. Breeding tomatoes for nematode resistance and for high vitamin C content in Hawaii. Proc. Am. Soc. Hort. Sci. 47:262-270.
McGuire, D. C. and R. W. Allard. 1958. Testing nematode resistance in the field. Plant Dis. Reptr. 42:1169-1172.
____________________________________ and J. A. Harding. 1961.Inheritance of root-knot nematode resistance in lima beans.Proc. Am. Soc. Hort. Sci. 78:302-307.
Minton, N. A. 1962, Factors influencing resistance of cotton to root-knot nematodes (Meloidogyne spp.) Phytopath. 53:272-279.
______________ and M. B. Parker. 1974. Yields of six soybeancultivars following fumigation of soil infested with root-knot nematodes. Phytopath. 64:219-221.
__________________________________, and R. A. Flowers. 1975.Response of soybean cultivars to Meloidogyne incognita and to the combined effects of M. arenaria and Sclerotium rolfsii■ Plant Dis. Reptr. 59:920-923.
Morse, W. J. 1927. Soy beans: culture and varieties. USDA Farmer's Bull. 1520.
Neal, J. C. 1889. The root-knot disease of the peach, orange and other plants in Florida, due to the work of Anguillula.USDA Div. of Entom. Bull. No. 20.
Peacock, F. C. 1959. The development of a technique for studying the host/parasite relationship of the root-knot nematode Meloidogyne incognita under controlled conditions. Nematologica 4:43-55.
Rohde, R. A. 1965. The nature of resistance in plants to nematodes. Phytopath. 55:1159-1162.
Romshe, F. A. 1942. Nematode resistance test of tomatoes.Proc. Am. Soc. Hort. Sci. 40:423.
Ross, J. P. 1965. Predisposition of soybeans to fusarium wilt by Heterodera glycines and Meloidogyne incognita. Phytopath. 55:361-364.
Saichuk, J. K. , C. Williams, and W. Birchfield. 1976. A technique for screening soybeans for resistance to root-knot nematode. Plant Dis. Reptr. 60:868-870.
Sasser, J. N. 1954. Identification and host-parasite relationships of certain root-knot nematodes (Meloidogyne spp.). Univ. of Maryland Agr. Exp. Sta. Bull. A-77.

59
83.
84.
85.
86.
87.
88.
89.
90.
91.
92.
93.
94.
95.
96.
Sasser, J. N. 1972. Managing nematodes by plant breeding.Tall Timbers Conf. on Ecol. Anim. Control by Habitat Manage.Proc. 4:65-80.
and A. L. Taylor. 1952. Studies on the entry oflarvae of root-knot nematodes into roots of susceptible and resistant plants. Phytopath. 43:474 (abstr.).
and N. N. Winstead. 1956. Reaction of cucumbervarieties to five root-knot nematodes (Meloidogyne spp.).Plant Dis. Reptr. 40:272-275.
Sawhney, R. and J. M. Webster. 1975. The role of plant growth hormones in determining the resistance of tomato plants to the root-knot nematode, Meloidogyne incognita. Nematologica 21:95-103.
Sayre, R. M. 1973. Theratromyxa weberi, an amoeba predatory on plant parasitic nematodes. J. Nematol. 5:258-264.
Sharpe, R. H., C. 0. Hesse, B. F. Lownsbery, V. G. Perry, andC. J. Hansen. 1969. Breeding peaches for root-knot nematoderesistance. J. Am. Soc. Hort. Sci. 94:209-212.
Shepherd, R. L. 1974. Breeding root-knot resistant Gossypium hirsutum L. using a resistant wild G. barbadense L. Crop Sci. 14:687-691.
________________. 1974. Transgressive segregation for root-knotnematode resistance in cotton. Crop Sci. 14:872-875.
Sidhu, G. and J. M. Webster. 1973. Genetic control of resistance in tomato. I. Identification of genes for host resistance to Meloidogyne incognita. Nematologica 19:546-550.
______________________________ . 1974. Genetics of resistance inthe tomato to root-knot nematode - wilt-fungus complex. J.Hered. 65:153-156.
Sikora, R. A., K. Sitaramaiah and R. S. Singh. 1973. Reaction of root-knot nematode - resistant tomato cultivars to Meloidogyne javanica in India. Plant Dis. Reptr. 57:141-143.
Smith, A. L. 1941. The reaction of cotton varieties to Fusarium wilt and root-knot nematode. Phytopath. 31:1099-1107.
______________ and A. L. Taylor. 1947. Field methods of testingfor root-knot infestation. Phytopath. 37:85-93.
Stevenson, F. J. and H. A. Jones. 1953. Some sources of resistance in crop plants. Iri Yearbook of Agric. 192-216.

60
97. Taylor, D. P. and T. D. Wyllie. 1959. Interrelationship of root-knot nematodes and Rhizoctonia solani on soybean emergence. Phytopath. 49:553 (abstr.).
98. Thomason, I. J. and P. G. Smith. 1957. Resistance in tomato to Meloidogyne javanica and M. incognita acrita. Plant Dis. Reptr. 41:180-181.
99. Tyler, J. 1941. Plants reported resistant or tolerant to root- knot nematode infestation. USDA Misc. Publ. No. 406.
100. Walker, J. C. 1957. Plant pathology 3rd. ed. McGraw-Hill, New York, St. Louis, San Francisco, Toronto, London, Sydney. 819 p.
101. Watts, V. M. 1947. The use of Lycopersicon peruvianum as a source of nematode resistance in tomatoes. Proc. Am. Soc. Hort. Sci. 49:233-234.
102. Weimer, J. L. and L. L. Harter. 1925. Varietal resistance of sweetpotatoes to nematodes, Heterodera radicicola (Greeff) Mull, in California. Phytopath. 15:423-426.
103. Weinberger, J. H., P. C. Marth, and D. H. Scott. 1943. Inheritance study of root-knot nematode resistance in certain peach varieties. Proc. Am. Soc. Hort. Sci. 42:321-325.
104. Wester, R. E. 1950. A comparison of greenhouse and field methods for evaluating lima beans for resistance to root- knot nematodes. Proc. Am. Soc. Hort. Sci. 56:395-400.
105. Wiles, A. B. 1957. Resistance to root-knot nematode in cotton. Phytopath. 47:37 (abstr.).
106. Williams, C., W. Birchfield, and E. E. Hartwig. 1973. Resistance in soybeans to a new race of root-knot nematode. Crop Sci. 13:299-301.
107. Winstead, N. N. and R. D. Riggs. 1959. Reaction of watermelon varieties to root-knot nematodes. Plant Dis. Reptr. 43:909-912.
108. Wright, S. L. 1956. Inheritance of resistance to root-knot nematode in cotton. Unpublished manuscript of Master of Science thesis. Louisiana State University, Baton Rouge, Louisiana.
109. Wyllie, T. D. and D. P. Taylor. 1960. Phytophthora root rot of soybeans as affected by soil temperature and Meloidogyne hapla. Plant Dis. Reptr. 44:543-545.

VITA
John Saichuk, son of Mrs. Therese Saichuk, was born April 22,
1950, in New Orleans, Louisiana. In 1968 he completed his high school
education at Hanson Memorial High School in Franklin, Louisiana.
In the fall of 1968 he entered the University of Southwestern
Louisiana where he received a Bachelor of Science degree in Agriculture
in May of 1972. The following September he entered Texas A & M
University from which he received a Master of Science degree in
Agronomy in May of 1974. In September of that year he entered
Louisiana State University in pursuance of a Doctor of Philosophy
degree in Agronomy with a minor in Plant Pathology.
Shortly before completion of degree requirements he accepted a
teaching position at the University of Southwestern Louisiana in
Lafayette, Louisiana.
The author is married to the former Susette Anne Patin of
New Roads, Louisiana. They have one son, John.
Their permanent mailing address is: 301 Montrose Ave.
Lafayette, Louisiana 70503
61

E X A M I N A T I O N A N D THESIS R E P O R T
Candidate: John Kennedy Saichuk
Major Field: Agronomy
Title of Thesis: £he Genetic Behavior of Resistance in Soybeans to the Wartelle Raceof Root-Knot Nematode.
Approved:
ajor Professor ancP'Chairman
Y ~ * -Dean of the Graduatj? School
EXAMINING COMMITTEE:
Date of Examination:
Aagus.'f- / , / ? 7 7