Embed Size (px)
Citation preview

Environmental and Experimental Botany, Vol. 32, No. 3, pp. 273 280, 1992 0098-8472]92 $5 00 + 0.00 Printed in Great Britain. rl:-~ 1992 Pergamon Press Ltd
THE INFLUENCE OF BENZYLADENINE AND N I T R O G E N SOURCES ON RESPONSES OF BEAN PLANTS
TO N I T R O G E N D I O X I D E
H. S. SRIVASTAVA,* D. P. ORMROD~" and B. A. HALE1
* Department of Plant Science, Rohilkhand University, Bareilly 243005, India and I Department of Horticultural Science, University of Guelph, Guelph, Ontario, Canada N 1G 2W 1
(Received 26 July 1991; accepted in final revisedJbrm 22 January 1992)
SR1VASTAVA H. S . , ORMROD D. P. and HALE B. A. The influence oJbenzyladenine and nitrogen source~ on response.~ of bean plants to nitrogen dioxide. ENVmONMENTAL AND EXPERIMENTAL BOTANY 32, 273--280, 1992. -Bean seedlings (Phaseolus vulgaris cv. Kinghorn Wax) were grown in nutrient solutions containing three different N sources or no added N and exposed to NO2 and a cytokinin, benzyladenine (BA). Cytokinins are known effectors of nitrate reduetase activity (NRA) and could stimulate the assimilation of nitrate derived from nutrient N sources or from NO2, decreas- ing NO, injury to plant tissue. There was no consistent effect of BA on any of the response variables. Exposure to NO~ at 0.3 ppm retarded shoot growth but not root growth. The N source greatly affected plant growth with NH,NO~ providing the greatest growth ibllowed by KNO~, NHtC1 and no added N. Shoot and root tissue N were greater with N salts supplied than without and generally higher in shoots but not roots after exposure to 0.3 ppm NO 2. Visible NO., injury occurred at the highest NO.., concentration only in NHgCl-grown plants. NRA was generally increased by NO,_, and was also highest in the NH4Cl-grown plants. While there were many significant interactions of treatment factors, including BA, the expected effect of BA in increasing NRA did not occur. This study demonstrated that BA treatment does not markedly affect NO~ assimilation or growth retardation by NO,.
Ke~, wor&: Inorganic nitrogen, nitrogen dioxidc, benzyladenine, Phaseolus vulgaris, bean, growth, nitrate reductase activity, air pollution.
INTRODUCTION
INHIBITION of p lan t growth by NO,,, is well known, ~'~'~ a l though s t imula t ion of p lan t growth at lower levels of the pol lu tan t has been repor ted. ~~'~24' The s t imula t ion of p lan t growth is appa ren t ly due to assimilat ion of n i t ra te and ni t ra te genera ted from NO2 inside the plants, resulting in an increase in total and organic nitro- gen contents. 17.23, The assimilat ion of NO~ has been demons t ra ted ~8.10, and appa ren t ly involves the G S / G O G A T pathway. 'L7 Al though the exact mechanism of NO2 phyto toxic i ty is not known,
accumula t ion of toxic levels of nitri te and ammon ium, and deplet ion of the essential cotRc- tors A T P and N A D P H may be assumed to con- t r ibute to the process. Thus, the rate and efficiency of NO2 assimilat ion will de te rmine the extent of positive or negative responses of plants to the pol lu tant . Cytokinins general ly increase ni t rogen assimilat ion by increasing ni t ra te reductase act ivi ty (NRA), ~4'7'12' and they are known to modit}¢ the responses of plants to O,~. '~'9'22~ This is believed to be due to the scav- enging of free radicals p roduced dur ing 03 exposure. 'q' The source of inorganic ni t rogen can
273

274 H . S . SRIVASTAVA et al.
influence the nitrogen assimilation process, ~a>' the rate of NO.2 uptake, ' >' and perhaps the overall assimilation of NO, as well. The current inves- tigation was undertaken to determine the influ- ence of the benzyladenine (BA) and nitrogen sources on the response of bean plants to NO.,. The hypothesis under test was that BA would increase N R A activity and assimilation of NOe and decrease NO., injury to tissue and that this would be influenced by the nutrient N source available to the plants. Confirmation of this hypothesis would provide independent cor- roboration of the metabolic thte of NO., and additional insights into the mechanism of NO,2 toxicity. A practical application of such a result could be the utilization of BA as a chemical pro- tectant against NO~ injury to plants in NO.e-prone locations.
MATERIALS AND METHODS
Plant culture Snap bean (Phaseolus vulgaris L. cv. Kinghorn
Wax) seeds were sown in 20 cm diameter plastic pots filled with medium-sized washed (deionized water) vermiculite. The plants were watered once a day with modified hall'-strength Hoagland 's solution containing no nitrogen. Seedlings were raised in a controlled environment growth chamber under PPF density of approximately 300 /~mol m -2 sec I with a 14 hr photoperiod and continuous temperature of 25~C. On the seventh day after sowing, the seedlings were thinned to two per pot. On the eighth day, 10 plants were harvested and growth and nitrogen contents of roots and shoots, and N R A in the ti'esh pr imary (bitbliate) leaves were measured.
£~perimental treatments On the eighth day after sowing, the exper-
imental plants were segregated into tbur groups and transferred to 75x 75 x 75 cm Plexiglass chambers. Plants were irrigated once a day with half-strength Hoagland 's solution containing either no nitrogen or KNO~ (5 mM), NHeC1 (5 raM) or NH4NO3 (2.5 mM) as nitrogen source. Each nutrient solution was balanced for K + and pH was maintained at 6.0 by adding K O H or HC1. Half an hour before exposing these plants to either 0.1 or 0.3 ppm NO~, the uppersides of
the leaves were sprayed with either water or 10 /~M BA (dissolved in a tkw drops of 0.1 N K O H ) . The exposure to charcoal-filtered air ( - NO.,) or to NO~ continued tbr 7 days, with a 30 min break each day for irrigating. The leaves of BA-treated plants were sprayed again with BA on the eleventh day. During NO2 exposure and growth periods, the plants were subjected to a ten> perature of 25 4- 2~C and PPFD of approximately 300/lnlol m 2 sec i during a 14-hr photoperiod. The pollutant was introduced continuously into the exposure chamber from a compressed gas cyl- inder and its concentration monitored with a Thermoelectron model 12T chemiluminescent N O 2 analyzer and maintained at the desired level.
The exposure was terminated on day 15 and tissue samples were collected immediately tbr analysis. The experiment was repeated in its entirety to provide a second replication (Block).
~.leasurements Shoots were weighed as soon as possible after
their excision (SFW). Roots were removed care- fully, washed with tap water and blotted with paper towels belbre weighing (RFW). Dry weights of roots (RDW) and shoots (SDW) were determined after drying in a forced draft oven at 70 80°C for 48 hr.
In vitro nitrate reductase activity (NRA) in the freshly harvested pr imary (bifoliate) leaves was measured as described earlier, la, Briefly, the enzyme was extracted from fresh leaf tissue below 4°C with a pestle and mortar in a medium con- sisting of 0.2 M sodium phosphate buffer (pH 7.5), 2 mM EDTA, 1 mM cysteine, and 0.5°o casein. The clear supernatant obtained after cen- trifugation at 1-4°C for 10 rain at 20,000 g was used as the enzyme preparation. The assay mix- ture consisted of 2.3 ml of 0.2 M sodimn phos- phate buffer (pH 7.8), 0.2 ml of 0.2 M KNO3, 0.3 ml of 2 m M N A D H , and 0.3 ml of enzyme preparation. The mixture was incubated at 26°C tbr 1 hr. The reaction was terminated by the addition of 1.5 ml of 1°~ sulfanilamide (in 1.5 N HC1) and 1.5 ml of0.02°o naphthyl ethyldiamine dichloride.
Total organic nitrogen (N) in dry root and shoot samples was determined by the mic- rokjeldahl method. An aliquot of the powdered root and shoot samples was digested with cone.

BA, N AND NO, ON BEAN PLANTS 275
H2SO 4 and 30~/o H=,O., and analyzed in a nitrogen autoanalyzer.
The data were analyzed as a completely ran- domized three-factor study, examining the effects of BA, nitrogen salts, and concentrations of NO2 tbr main effects, as well as all two-way interactions and the three-way interaction. The data pre- sented are the means of 12 plants (from two sep- arate experiments) for growth, and of four deter- minations for N R A and total nitrogen.
RESULTS
Plant growth, nitrogen contents of" roots and shoots, and N R A in the ti'esh pr imary leaves on the eighth day after sowing are recorded in Table 1. As differential treatments were imposed at this time on other plants growing in the same con- ditions these data represent the starting values tbr each parameter under study. There was sig- nificant interaction ( P < 5 ° o ) between and among BA, nitrogen salts, and concentrations of NO2 (BAxNO._,, B A x N salt, N O 2 x N salt, B A x N O ~ - N salt) for all growth variables (RDW, RFW, SDW, SFW) suggesting that main effects could not be used to summarize plant responses to these factors (Table 2). The contrast determining whether the plant response to NO~ concentrat ion was linear was highly significant
Table I. Growth, Kjeldahl nitrogen and nitrate reductaae activity in bean planl,~ grown Jbr 8 da~s with nutrient solution
containing no nitrogen
Plant organ
Root
Shoot
Leaf
Parameter Values
Fresh weight, mg/root 541 + 66 Dry weight, mg/root 31.0+_3.7 Kjeldahl nitrogen, (' o 3.25 +_ 0.43 Fresh weight, rag/shoot 1356 +_ 80 Dry weight, mg/shoot 127,5+_ 18.0 Kjeldahl nitrogen, "~, 5.29+_0.02
Nitrate reductase 81 +- 20 activity, nmoles NOg • hr ' • g- i fresh weight
The growth data are the average of five replications (_S.D.) with 10 plants in each replication. The enzyme activity and Kjeldahl nitrogen data are the average of four replications (_ S.D.).
(Table 2). The simple means of each t reatment combinat ion with the standard error of the mean for each response variable, pooled over the treat- ments, indicate the complexity of these inter- actions (Tables 3 and 4).
Neither exposure to NO._, nor the application of BA consistently affected root growth (Table 31. Root t~esh and dry weights were greatest when NH4NO:~ was in the nutrient solution. Inclusion of KNO~ was generally less advantageous than NH4NO:~ to root growth with NH4C1 similar to a nitrogen-free nutrient solution. No consistent patterns of factor interaction could be identified in the root growth measurements. This was in contrast to effects on shoot g rowth /Tab l e 4). The application of 0.3 ppm NO., resulted in con- sistently less t?esh and dr?' weight of shoots than tbr plants not exposed to NO, or exposed to 0.1 ppm NO.,. No overall responses to BA were rec- orded. Shoot growth was strongly favored by the addition of nitrogen to the nutrient solution with greatest growth in NH~NO~, followed by KNO~ and NH4C1. Even NH4CI provided much more shoot growth than the nitrogen-t?ee nutrient solu- tion. As for root growth, consistent interactions of treatment factors could not be identified.
Root and shoot nitrogen content An analysis of variance indicated that tor shoot
and root nitrogen little interaction (only BA x NO., for shoot N) existed among the three factors (Table 5). Exposure to 0.3 ppm NO2 gen- erally increased shoot N but neither root nor shoot N was influenced by application of BA to the foliage (Table 6). Shoot and root N were gen- erally greatest when grown in NH4C1 tbllowed t)y NH4NO.~, KNO.~ and control, in that order.
Nitrate reductase activity, Nitrate reductase activity was stimulated by
exposure to NO2 with much higher N R A levels in exposed plants (Table 71). Application of BA did not consistently increase or decrease NRA. In contrast the different N sources resulted in very different N R A levels with NH4NO, ~ resulting in the lowest level. There was a striking NO,, x N salt interaction because 0.3 ppm NOe greatly stimulated N R A in NHaC1 compared with 0.1 ppm in contrast to the lack of a differential effect
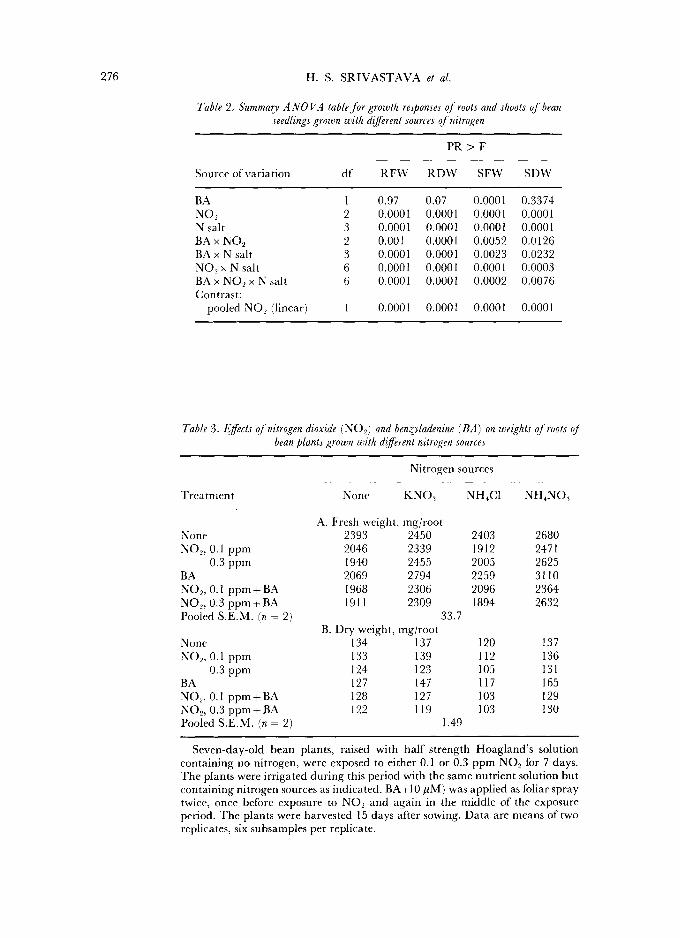
276 H. S. S R I V A S T A V A et al.
Table 2, Summary A NO VA table for growth responses of roots and shoots of bean seedlings grown with different sources of nitrogen
P R > F
Source of variation df R F W R D W SFW SDW
BA 1 0.97 0.07 0.0001 0.3374 NO-, 2 0.0001 0.0001 0.0001 0.0001 N salt 3 0.0001 0.0001 0.0001 0.0001 BA x NO., 2 0.001 0.0001 0.0052 0.0126 BAx N salt 3 0.0001 0.0001 0.0023 0.0232 NO., x N salt 6 0.0001 0.0001 0.0001 0.0003 BA x NO2 x N salt 6 0.0001 0.0001 0.0002 0.0076 Contrast:
pooled NO, (linear) 1 0.0001 0.0001 0.0001 0.0001
Table 3. Effects of nitrogen dioxide (NO2) and benzrladenine (BA) on weights of roots oJ bean plants grown with different nitrogen sources
Nitrogen sources
Treatment None KNO,~ NH~CI NH4NO.~
A. Fresh weight, mg/root None 2393 2450 2403 2680 NO2, 0.1 ppm 2046 2339 1912 2471
0.3 ppm 1940 2455 2005 2625 BA 2069 2794 2259 3110 NO2, 0.1 ppm + BA 1968 2306 2096 2364 NO2, 0.3 ppm + BA 1911 2309 1894 2632 Pooled S.E.M. (n = 2) 33.7
B. Dry weight, rag/root None 134 137 120 137 NO2, 0.1 ppm 133 139 112 136
0.3 ppm 124 123 105 131 BA 127 147 117 165 N O 2, 0. l p p m + B A 128 127 103 129 NO2, 0.3 p p m + B A 122 119 103 130 Pooled S.E.M. (n = 2) 1.49
Seven-day-old bean plants, raised with half strength Hoagland's solution containing no nitrogen, were exposed to either 0.1 or 0.3 ppm N O 2 for 7 days. The plants were irrigated during this period with the same nutrient solution but containing nitrogen sources as indicated. BA (10 pM) was applied as foliar spray twice, once before exposure to NO2 and again in the middle of the exposure period. The plants were harvested 15 days after sowing. Data are means of two replicates, six subsamples per replicate.
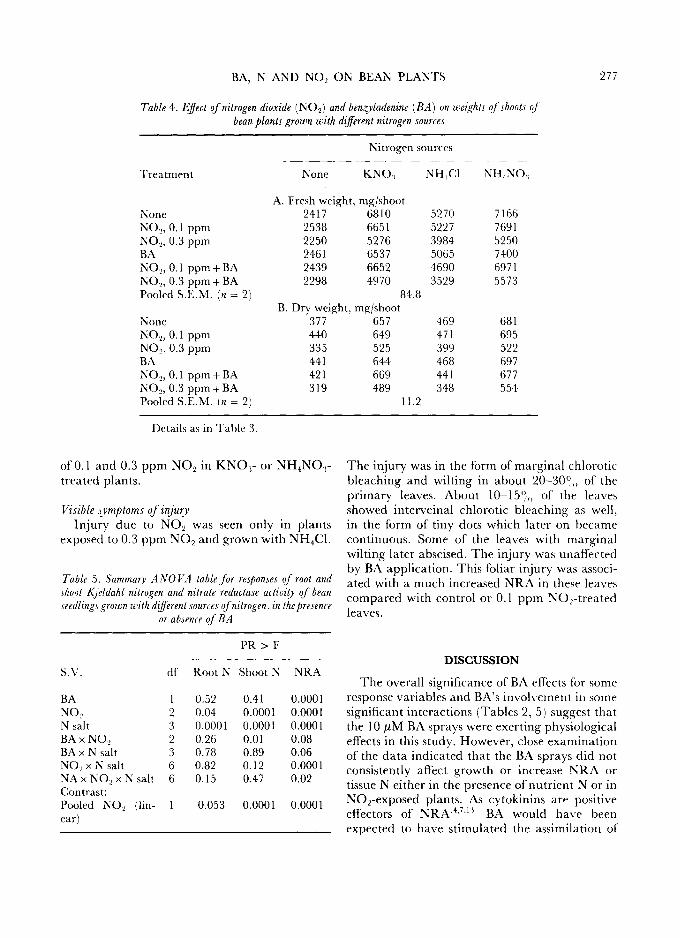
BA, N AND NO~ ON BEAN PLANTS 277
Table 4. Effect of nitrogen dioxide (NO2) and benzyladenine ( BA) on weights of shoots oJ bean plants grown with different nitrogen sources
Nitrogen sources
Treatment None KNO.~ NH,CI NH4NO.~
A. Fresh weight, rag/shoot None 2417 6810 5270 7166 NO..,, 0.1 ppm 2538 6651 5227 7691 NO..,, 0.3 ppm 2250 5276 3984 5250 BA 2461 6537 5065 7400 NO2, 0.1 ppm + BA 2439 6652 4690 6971 NO._,, 0.3 ppm + BA 2298 4970 3529 5573 Pooled S.E.M. (in = 2) 84.8
B. Dry weight, mg/shoot None 377 657 469 681 NO.2, 0.1 ppm 440 649 471 695 NO~, 0.3 ppm 335 525 399 522 BA 441 644 468 697 NO..,, 0.1 ppm + BA 421 669 441 677 NO,_,, 0.3 ppm + BA 319 489 348 554 Pooled S.E.M. (n = 2) 11.2
Details as in Table 3.
of 0.1 and 0.3 ppm NO2 in KNO3- or NH4NO~- t reated plants.
Visible +~rnptoms of injur), In ju ry due to NO~ was seen only in plants
exposed to 0.3 p p m NO2 and grown with NH4C1.
Table 5. Summary ANOVA table Jbr responses of root and shoot Kjeldahl nitrogen and nitrate reductase activity of bean seedlings grown with different sources of nitrogen, in the presence
or absence of BA
P R > F
S.\:. df Root N Shoot N NRA
BA 1 0.52 0.41 0.0001 NOe 2 0.04 0.0001 0.0001 N salt 3 0.0001 0.0001 0.0001 BA x NO._, 2 0.26 0.01 0.08 BA x N salt 3 0.78 0.89 0.06 NO._, x N salt 6 0.82 0.12 0.0001 NAx NO~ x N salt 6 0.15 0.47 0.02 Contrast: Pooled NO., (lin- 1 0.053 0.0001 0.0001 ear)
The injury was in the tbrm of marg ina l chlorotic bleaching and wil t ing in abou t 20-30t!o of the p r imary leaves. Abou t 10-15~}o of the leaves showed interveinal chlorotic b leaching as well, in the form of tiny dots which la ter on became continuous. Some of the leaves with margina l wil t ing la ter abscised. The injury was unaffected by BA appl icat ion. This foliar injury was associ- a ted with a much increased N R A in these leaves compared with control or 0.1 ppm NO2-treated leaves.
DISCUSSION
The overall significance of BA effects fbr some response var iables and BA's involvement in some significant interact ions (Tables 2, 5) suggest that the 10/~M BA sprays were exert ing physiological effects in this study. However , close examina t ion of the da ta indica ted that the BA sprays did not consistently affect growth or increase N R A or tissue N ei ther in the presence of nut r ien t N or in NO2-exposed plants. As cytokinins aro positive effectors of N R A L4'7'1'~' BA would have been expected to have s t imulated the assimilat ion of
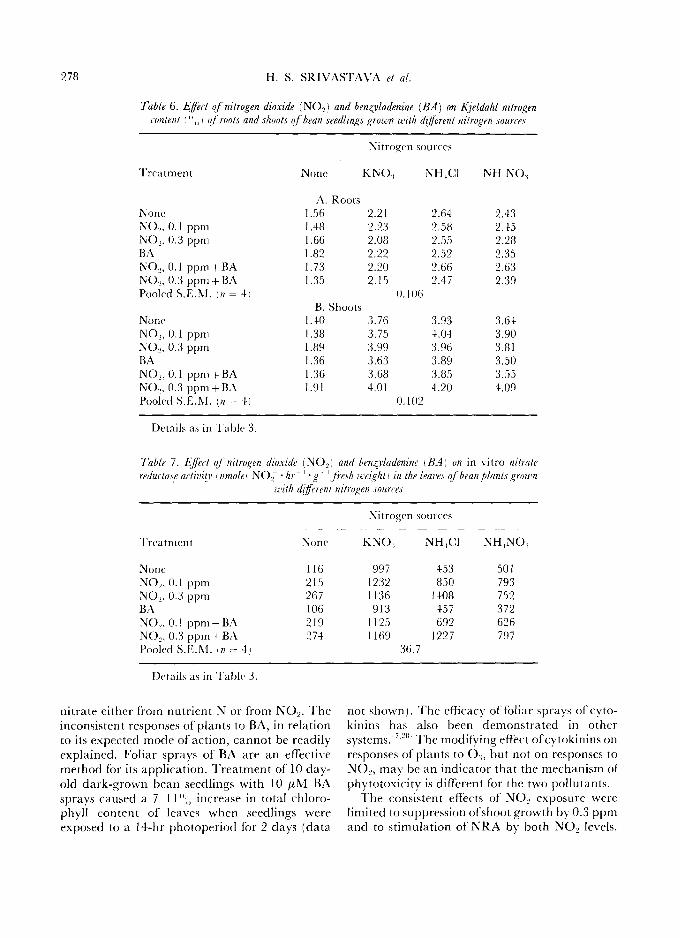
278 H . S . SRIVASTAVA et al,
Table 6. Effect o f nitrogen dioxide (NO,z) and ben4ffadenine (BA) on Kjeldahl nitrogen content I ",, ) o/root~ and shoot~ o f bean seedlings grown v:,ith di[Jerent nitrogo~ sources
Nitrogen sources
Treatment None KNO,~ NH~CI NH~NO,~
A. Roots None 1.56 2.21 2.64 2.43 NO~, 0.1 ppm 1.48 2.23 2.58 2.45 N()-,. 0.3 ppm 1.66 2.08 2.55 2.28 BA 1.82 2.22 2.52 2.35 NO-,, 0.1 ppm+BA 1.73 2.20 2.66 2.63 NO.,, 0.3 ppm+BA 1.35 2.15 2.47 2.39 Pooled S.E.M. In - 4/ 0.106
B. Shoots None 1.40 3.76 3.93 3.64 NO-,, 0.1 ppm 1.38 3.75 4.04 3.90 NO-,, 0.3 ppm 1.89 3.99 3.96 3.81 BA 1.36 3.63 3.89 3.50 NO_,, 0.1 ppm + BA 1.36 3.68 3.85 3.55 NO.,, 0.3 ppm+BA 1.91 4.01 4.20 4.09 Pooled S.E.M.I.n = 4) 0.102
Details as in Table 3.
Table 7. E~'ecl o j" nitrogen dioxide (NO..,) and benzrladenine I BA) on in vitro nitrate reducla,~.e aetivity I nmole~ NOe • hr ~ • g ~ ~ J're.~h ze'ezi~,hl'l in the leave.~ o f bean plants ,grown
u,ilh dijJi'rent nitrogen sources
Treatment
Nitrogen sources
None KNO; N H 4 C 1 NH4NO~
None 116 997 453 501 NO2, 0.1 ppm 215 1232 850 793 NO~,/).3 ppm 267 1136 1408 752 BA 106 913 457 372 NO-,, 0.1 ppm+BA 219 1125 692 626 NO._,, 0.3 ppm+BA 274 1169 1227 797 Pooled S.E.M. In = 4) 36.7
Details as in Table 3.
nitrate either ti'om nut r ient N or ti'om NO2. The inconsistent responses of plants to BA, in relation to its expected mode of action, cannot be readily explained. Foliar sprays of BA are an effective method for its application. T rea tmen t of 10 day- old dark-grown bean seedlings with 10 /.tM BA sprays caused a 7-11~},, increase in total chloro- phyll content of leaves when seedlings were exposed to a 14-hr photoperiod tbr 2 days (data
not shown). The efficacy of tbliar sprays of cyto- kinins has also been demonstrated in other
7 2 0 . . . . . . systems. ' The modlfymgeffect ofcytokmms on responses of plants to O.~, but not on responses to NO.,, may be an indicator that the mechanism of phytotoxicity is diff"rent for the two pollutants.
The consistent effects of NO 2 exposure were limited to suppression of shoot growth by 0.3 ppm and to st imulat ion of NRA by both NO., levels.

BA, N AND NO_, ON BEAN PLANTS 279
An increase in NRA caused by NO~ is considered to be due to nitrate build up in the tissue, as a result of dissolution of NO~ in apoplastic or cytoplasmic water. ''7'~"' The higher N R A in NH4Cl-grown plants may be due to a higher rate of NO., uptake in these plants. Fur thermore 0.3 ppm NO., applied to NH,Cl -g rown plants was the only treatment in which extensive foliar visible injury was noted. In an earlier study, involving short-term exposure of bean plants to NO.,, the rate of NO., uptake by the leaves of plants grown with urea was tbund to be substantially higher than that of nitrate-grown plants. 16~ However, NO., exposure did not greatly increase total N (Kjeldahl NIl with any of the N sources in the present study. Since NRA, which reduces nitrate to nitrite, was quite high in NH4Cl-grown plants, especially in those exposed to 0.3 ppm NO~ (Table 7) and showing visible injury, one would suspect that the further reduction of nitrite and/or assimilation of ammonium into amino acids was somewhat limited in these plants. Measurements of levels of N O , and NH4 ~ and/or the enzymes assimilating these species, in NO~-exposed plants and as affected by N sources are required to test this possibility. The accumulat ion of nitrite and axnmonium in NO.,-exposed plants has been con- sidered by some researchers ''-q2s~ to be the most important factor in pollutant toxicity.
Growth of bean seedlings, as measured by flesh and dry weights of shoots, was greatest when nutrient nitrogen was NH~NO:~ tbllowed by KNO:~ and NH4CI in that order. Superiority of nitrate over ammonium as a source of nitrogen has long been recognized. ~'¢f' The source of nutri- ent nitrogen modified the response to NO 2 only to a limited extent with one exception; the visible injury to the leaves by 0.3 ppm NO~ and the maximum increase in N R A by this concentrat ion of pollutant occurred in NH,Cl-supplied plants.
The data in this study did not support the mechanistic hypothesis underlying the experi- ments. Even though the known mode of action of BA, as a cytokinin, indicated that there would be a general impact of BA on plant response to NO2, the expected responses did not occur in situ. While the statistical analyses did indicate that BA was interacting with other treatment variables in some specific physiological ways, the over-riding effects of N R A and NO,, assimilation did not
occur, In spite of this negative response to BA applications, the study did reveal some important effects of N source in the rooting medium on plant growth, tissue N, N R A and visible injury. Differences between root and shoot responses to the treatment variables were also revealed. A par- ticular finding of interest was that the NO.,- induced visible injury and increase in N R A were maximal in NHaCl-grown plants exposed to 0.3 ppm NO.., suggesting a relationship between NO._, assimilation and ammonium nutrition that results in the accumulation of toxic nitrite and ammonium in tissue.
Acknowledgments The senior author was the recipient of a Natural Sciences and Engineering Research Coun- cil of Canada, International Scientific Exchange award during this study. The research was also supported by NSERC operating grant to D.P.O. We thank L. Pvear andJ. Zealand for technical assistance.
REFERENCES
1. ADEDIPE N. O. and ORMROD D. P. <19721 Hor- monal regulation of ozone phytotoxicity in Raphanus sativus. Z. Pflanzenp/9,.doL 68, 254 258.
2. CAPRON '1'. l~l. and MANSFIELD T. A. 11977! Inhibition of growth in tomato by air polluted with nitrogen oxides. J . exp. Bot. 28, 112 116.
3. FREER-SMiTh P. H. (1985> The influence of SO, and NO~ on the growth, development and gas exchange of Betula pendula Roth. New Ph~'lol. 99, 417 430.
4. KENDE H., HAHN H. and NAYS S. E. ',1971~ Enhancement of nitrate reductase activity by benzyladenine in :ty, ro.~temma o, itha¢,o. P/. Phv.~iol. 48, 702-706.
5. KIRKBY E. A. '. 1969) Ion uptake and ionic balance in plants in relation to the tbrm of nitrogen nutrition. Pages 215-235 in J. H. RORISON, ed. Ecological aspect.~ of mineral nutrition qf plant.~. Blackwell Seientitic Publication, Oxlbrd.
6. KRESS L. W., SK~:LLV .J.M. and KINKELMANN K. H. (1982) Growth impact of O~, NO., and/or SO., on Plantanua occidentalia. Agric. Era'Jr. 7, 265 274.
7. L~r,J-]., ERTL.J.R. and CHE*'," C-re. ,'1990/ C~to- kinin enhancement of the light induction of nitrate reductase transcript levels in etiolated barh,y leaves. PI. runlet. BioL 14, 585-594.
8. ~IATSUMARU g., YONEYAMA W., TOTSUKA T. and SHIRATORI K. i 1979'1 Ahsorption of atmospheric N O 2 by plants and soils I. Quantitative estimation of absorbed NO,, in plants by I~N method. Soil 3'ci. Pl. Nutr. 25, 255 265.

280 H . S . SRIVASTAVA et al.
9. PAULS K. P. and THOMPSON J. E. (1982) Effects ofcytokinins and antioxidants on the susceptibility of membranes to ozone damage. Pl. (2ell Physiol. 23, 821-832.
10. ROGERS H. H., CAMPBELL J. C. and VOLK R. J. (1979) Nitrogen-15 dioxide uptake and incor- poration by Phaseolus vulgaris (L.). Science 206, 333- 335.
11. ROSNITSCHEK-SCHIMMEL I. (1982) Effect of ammo- nium and nitrate supply on dry matter production and nitrogen distribution in Urlica dioica. Z. Pflan- zenphysiol. 108, 329-341.
12. SAHULKA J. (1972) The effect of exogenous IAA and kinetin on nitrate reductase, nitrite reductase, and glutamate dehydrogenase activities in excised pea roots. Biol. Pl. 14, 330-336.
13. SCHMERDER B. and BORRISS H. (1986) Induction of nitrate reductase by cytokinin and ethylene in Agrostemma githago L. embryos. Planta 169, 589- 593.
14. SPIERINGS F. H. G. C. (1971) Influence oftumi- gation with NO~ on growth and yield of tomato plants. Neth. J. Pl. Pathol. 77, 194-200.
15. SRIVASTAVA H. S. (1980) Regulation of nitrate reductase activity in higher plants. Phytochemistr~ 19, 725-733.
16. SRIVASTAVA ['-I. S.,JOLLIFFE P. A. and RUNECKLES V. C. (1975) The influence of nitrogen supply during growth on the inhibition of gas exchange and visible injury in bean leaves by NO~. Envir. Pollul. 9, 35-42.
17. SRIVASTAVA H. S. and ORMROD D. P. (1984) Effects of nitrogen dioxide and nitrate nutrition on growth and nitrate assimilation in bean leaves. PI. Physiol. 76, 418 423.
18. SRIVASTAVA H. S., ORMROD D. P. and HALE MARIE B. (1990) Photosynthetic photon flux effects on bean response to nitrogen dioxide. Envir. exp. Bol. 30, 463-467.
19. SRIVASTAVA H. S. and SINGR R. P. (1987) Role and regulation of L-glutamate dehydrogenase activity in higher plants. Phytochemistry 26, 597 610.
20. STOBART A. K. and SHEWRY P. R. (1972) The effect ofkinetin on chlorophyll synthesis in ageing etiolated barley leaves exposed to light. Phy- lochemistry 11,571-577.
21. TAYLOR O. C. and EATON F. M. (1966) Sup- pression of plant growth by nitrogen dioxide. Pl. Physiol. 41, 132-135.
22. TOMLINSON H. and RICH S. (1973) Anti-senescent compounds reduce injury and steroid changes in ozonated leaves and their chloroplasts. Phytopath- olog~ 63, 903-906.
23. TROIANO J. J. and LEONE I. A. (1977) Changes in growth rate and nitrogen content of tomato plants after exposure to NO.,. Phylopatholog~ 67, 1130- 1133.
24. YONEYAMA T., TOTSUKA T., HAYAKAWA N. and YAZAK1 I. (1980) Absorption of atmospheric NO2 by plants and soils V. Day and night fumigation effect on plant growth and estimation of the amount of NO2 nitrogen absorbed by the plants. Res. Rep. natl Inst. Envir. Stud. 11, 31 50.
25. Yu S-W., LI L. and SHIMAZAKI K-I. (1988) Response of spinach and kidney bean plants to nitrogen dioxide. Envir. Pollul. 55, 1 13.
26. ZEEVART A. J. (1974) Induction of nitrate reductase by" NO~. Acta Bot. Neerl. 23, 345-346.





























