Embed Size (px)
Citation preview

Neuron, Vol. 14, 1213-1222, June, 1995, Copyright © 1995 by Cell Press
The Integrin Receptor Mediates Interactions of Embryonic Chick Motor and Sensory Neurons with Tenascin-C Barbara Varnum-Finney,*§ Kristine Venstrom,t Ulrich Muller,* Robert Kypta,* Carey Backus,* Matthias Chiquet,$ and Louis F. Reichardt*t *Neuroscience Program Department of Physiology ~Howard Hughes Medical Institute University of California, San Francisco San Francisco, California 94143-0724 ~Department of Physical Chemistry Biocentrum University of Basel Ch-4056 Basel Switzerland
Summary
This paper identifies a neuronal receptor for tenas- cin-C (tenascin/cytotactin), ~n extracellular matrix protein that has previously been detected in devel- oping sensory and motor neuron pathways and has been shown to regulate cell migration in the devel- oping CNS. Antibodies specific for each subunit of the integrin ~ are used to demonstrate that o ~ medi- ates neurite outgrowth of embryonic sensory and mo- tor neurons on this extrecellular matrix protein. In ad- dition, expression of o~ in K562 cells results in surface expression of a.13~ heterodimers that are shown to pro- mote attachment of this cell line to tenascin. The major domain in tenascin that mediates neurite outgrowth is shown to be localized to fibronectin type III re- peats 6-8.
Introduction
Interactions between neurons and cell- and extracellular matrix (ECM)-associated proteins are essential for regu- lating development of the nervous system (reviewed in Goodman and Shatz, 1993). Tenascin-C (tenascin/cyto- tactin) is a member of a small family of ECM-associated glycoproteins that have been proposed to have many dis- tinct functions in neural development (reviewed in Rei- chardt and Tomaselli, 1991; Riou et al., 1992). In cell cul- ture, substrate-bound tenascin has been observed to inhibit migration of neural crest cells and axon growth by hippocampal, cerebellar, and mesencephalic neurons (Tan et al., 1987; Halfter et al., 1989; Faissner and Kruse, 1990). In vivo, its distribution in regions of the central ner- vous system, such as barrel fields in the somatosensory cortex, is consistent with the possibility that it delineates barriers that restrict axon growth (e.g., Steindler et al., 1989; Bartsch et al., 1992). In the peripheral nervous sys- tem, tenascin is found in migratory pathways of neural crest cells and along routes followed by motor and sensory
§Present address: Department of Pediatric Oncology, Fred Hutchin- son Cancer Research Center, Seattle, Washington 98109.
neurons, suggesting it might promote and direct neural crest migration and axon outgrowth (e.g., Wehrle and Chi- quet, 1990; Riou et al., 1992). Consistent with this, sub- strate-bound tenascin promotes neurite outgrowth by sen- sory and motor neurons in vitro (e.g., Wehrle and Chiquet, 1990; Wehrle-Haller and Chiquet, 1993).
The diverse responses of neurons to tenascin seem likely to be mediated by different receptors interacting with separate domains in the tenascin molecule. Tenascin is a hexamer whose individual subunits consist of linear arrays of several epidermal growth factor (EGF)-Iike re- peats, 8-14 fibronectin type III (FNIII) domains, and a C-terminal fibrinogen-like domain (see Figure 3A). Differ- ential splicing of exons encoding FNIII domains generates several isoforms of tenascin. Previous work indicated that several domains of tenascin interact with cells. The third FNIII domain in chicken tenascin contains an RGD se- quence, similar to cell binding sites in fibronectin and vitronectin. The RGD-containing domain in tenascin has been shown to mediate attachment by at least some non- neural cells (Prieto et al., 1992). Both monoclonal and poly- clonal antibodies to FNIII domains 6-7 strongly inhibit neu- rite outgrowth, implying that neurons recognize this portion of the molecule (Husmann et al., 1992; Wehrle- Hailer and Chiquet, 1993). Other domains of tenascin have also been shown to promote cell attachment (e.g., Hus- mann et al., 1992; Prieto et al., 1992). Inhibitory or "anti- adhesive" domains of tenascin have been localized tenta- tively to the EGF repeat region and to alternatively spliced FNIII domains (e.g., Spring et al., 1989).
Several proteins, including an immunoglobulin-class cell adhesion molecule named F l l , two proteoglycans named syndecan and cytotactin-binding proteoglycan, and integrins, have been proposed to function as recep- tors for tenascin (Zisch et al., 1992; Salmivirta et al., 1991; Hoffman et al., 1988; Bourdon and Ruoslahti, 1989; Srira- marao et al., 1993; Prieto et al., 1993). Both av and J31 subunit-containing integrins have been implicated as re- ceptors. Inhibitory effects of integrin-specific antibodies implicate ~2131 and avl~3 as receptors for endothelial cells (Sriramarao et al., 1993). The astrocytoma U-251 MG cell line has been shown to attach to the third, RGD-containing fibronectin repeat of tenascin via the integrin avl~3 (Prieto et al., 1993). ~v associated with another 13 subunit may also function on other cells as a tenascin receptor (Prieto et al., 1993).
13~ integrins also appear to be important in mediating cell attachment to tenascin domains (Prieto et al., 1993). In addition, I~1 integrins have recently been implicated as neuronal receptors for tenascin (Wehrle-Haller and Chi- quet, 1993). Neurite outgrowth by sensory neurons is strongly inhibited by a monoclonal antibody (MAb), JG22, to the integrin I~1 subunit.
Bossy et al. (1991) isolated cDNAs encoding the integrin ~8 subunit by virtue of homology with other integrin a sub- units. These investigators observed strong expression of the integrin cza subunit by subclasses of neurons in both
Neuron 1214
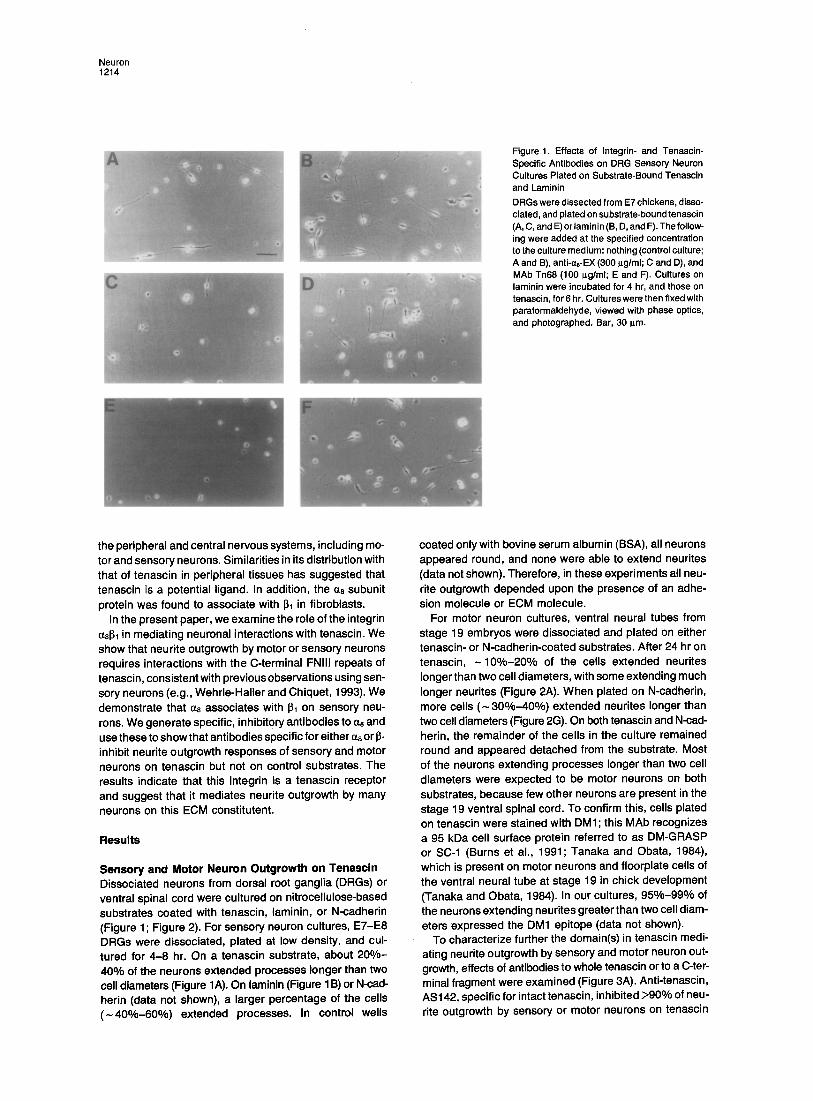
Figure 1. Effects of Integrin- and Tenascin- Specific Antibodies on DRG Sensory Neuron Cultures Plated on Substrate-Bound Tenascin and Laminin DRGs were dissected from E7 chickens, disso- ciated, and plated on substrate-bound tenascin (A, C, and E) or laminin (B, D, and F). The follow- ing were added at the specified concentration to the culture medium: nothing (control culture; A and B), anti-aB-EX (300 I~g/ml; C and D), and MAb Tn68 (100 p.g/ml; E and F). Cultures on laminin were incubated for 4 hr, and those on tenascin, for 6 hr. Cultures were then fixed with paraformaldehyde, viewed with phase optics, and photographed. Bar, 30 I~m.
the peripheral and central nervous systems, including mo- tor and sensory neurons. Similarities in its distribution with that of tenascin in peripheral tissues has suggested that tenascin is a potential ligand. In addition, the ct8 subunit protein was found to associate with I~1 in fibroblasts.
In the present paper, we examine the role of the integrin a81~1 in mediating neuronal interactions with tenascin. We show that neu rite outgrowth by motor or sensory neurons requires interactions with the C-terminal FNIII repeats of tenascin, consistent with previous observations using sen- sory neurons (e.g., Wehrle-Haller and Chiquet, 1993). We demonstrate that a0 associates with ~1 on sensory neu- rons. We generate specific, inhibitory antibodies to a8 and use these to show that antibodies specific for either as or I~1 inhibit neurite outgrowth responses of sensory and motor neurons on tenascin but not on control substrates. The results indicate that this integrin is a tenascin receptor and suggest that it mediates neurite outgrowth by many neurons on this ECM constitutent.
Results
Sensory and Motor Neuron Outgrowth on Tenascin Dissociated neurons from dorsal root ganglia (DRGs) or ventral spinal cord were cultured on nitrocellulose-based substrates coated with tenascin, laminin, or N-cadherin (Figure 1; Figure 2). For sensory neuron cultures, E7-E8 DRGs were dissociated, plated at low density, and cul- tured for 4-8 hr. On a tenascin substrate, about 200/o - 40% of the neurons extended processes longer than two cell diameters (Figure 1A). On laminin (Figure 1 B) or N-cad- herin (data not shown), a larger percentage of the cells ( - 4 0 0 - - 6 0 o ) extended processes. In control wells
coated only with bovine serum albumin (BSA), all neurons appeared round, and none were able to extend neurites (data not shown). Therefore, in these experiments all neu- rite outgrowth depended upon the presence of an adhe- sion molecule or ECM molecule.
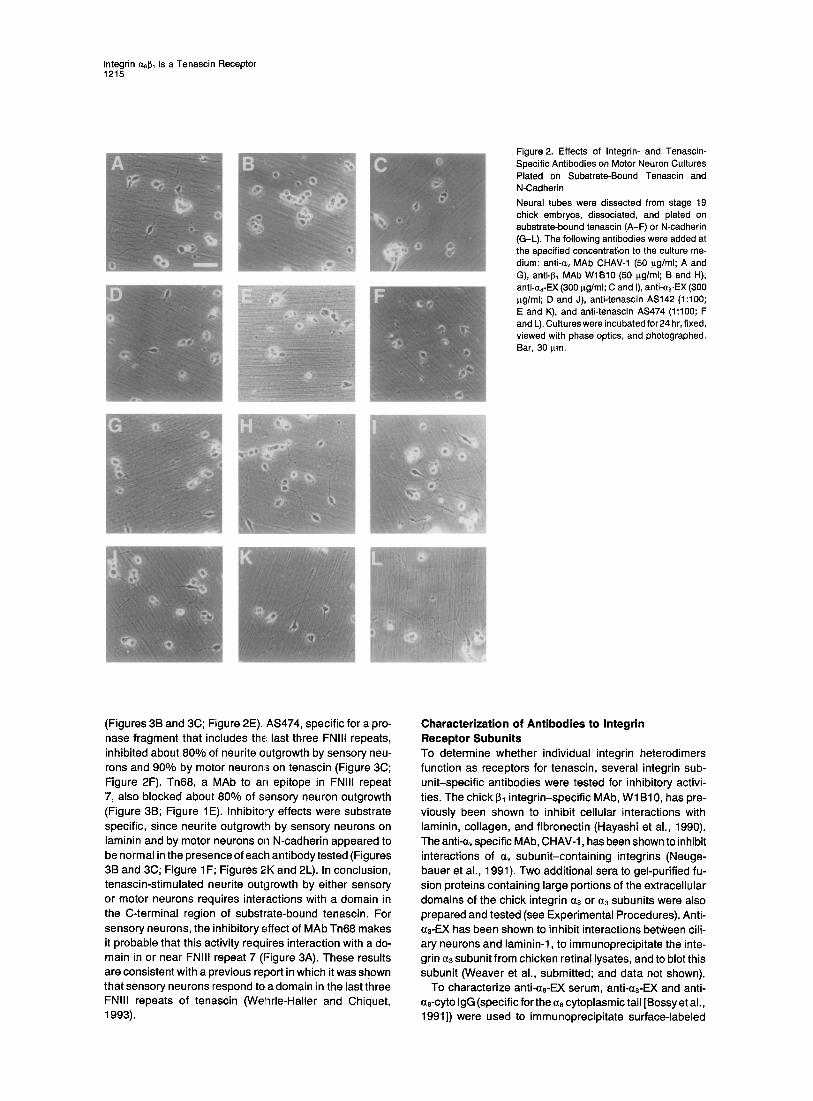
For motor neuron cultures, ventral neural tubes from stage 19 embryos were dissociated and plated on either tenascin- or N-cadherin-coated substrates. After 24 hr on tenascin, - 1 0 % - 2 0 % of the cells extended neurites longer than two cell diameters, with some extending much longer neurites (Figure 2A). When plated on N-cadherin, more cells ( - 30%-40%) extended neurites longer than two cell diameters (Figure 2G). On both tenascin and N-cad- herin, the remainder of the cells in the culture remained round and appeared detached from the substrate. Most of the neurons extending processes longer than two cell diameters were expected to be motor neurons on both substrates, because few other neurons are present in the stage 19 ventral spinal cord. To confirm this, cells plated on tenascin were stained with DM1; this MAb recognizes a 95 kDa cell surface protein referred to as DM-GRASP or SC-1 (Burns et al., 1991; Tanaka and Obata, 1984), which is present on motor neurons and floorplate cells of the ventral neural tube at stage 19 in chick development (Tanaka and Obata, 1984). In our cultures, 95%-99% of the neurons extending neurites greater than two cell diam- eters expressed the DM1 epitope (data not shown).
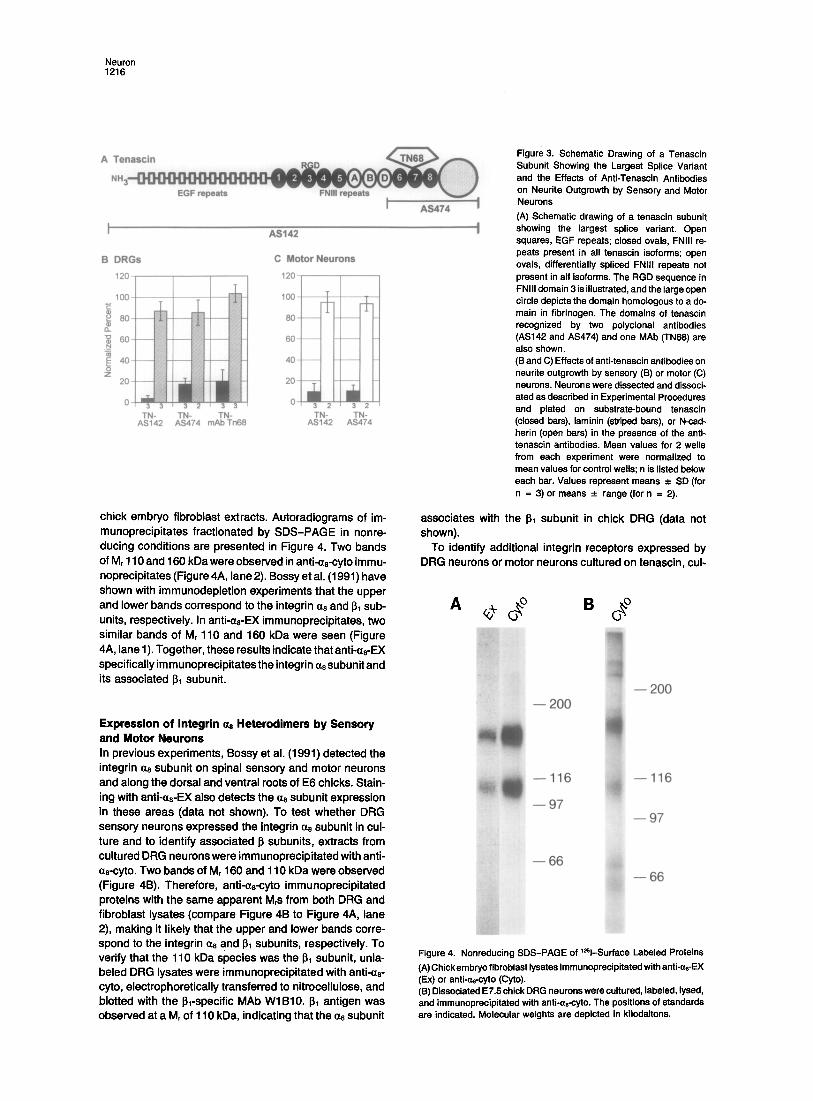
To characterize further the domain(s) in tenascin medi- ating neurite outgrowth by sensory and motor neuron out- growth, effects of antibodies to whole tenascin or to a C-ter- minal fragment were examined (Figure 3A). Anti-tenascin, AS142, specific for intact tenascin, inhibited >90% of neu- rite outgrowth by sensory or motor neurons on tenascin
Integrin a8131 Is a Tenascin Receptor 1215
Figure 2. Effects of Integrin- and Tenascin- Specific Antibodies on Motor Neuron Cultures Plated on Substrate-Bound Tenascin and N-Cadherin Neural tubes were dissected from stage 19 chick embryos, dissociated, and plated on substrate-bound tenascin (A-F) or N-cadherin (G-L). The following antibodies were added at the specified concentration to the culture me- dium: anti-c~v MAb CHAV-1 (50 p.g/ml; A and G), anti-J3~ MAb WlB10 (50 I~g/ml; B and H), anti-~8-EX (300 ilg/ml; C and I), anti-~3-EX (300 ~g/ml; D and J), anti-tenascin AS142 (1:100; E and K), and anti-tenascin AS474 (1:100; F and L). Cultures were incubated for 24 hr, fixed, viewed with phase optics, and photographed. Bar, 30 ~.m.
(Figures 3B and 3C; Figure 2E). AS474, specific for a pro- nase fragment that includes th~ last three FNIII repeats, inhibited about 80% of neurite outgrowth by sensory neu- rons and 90% by motor neurons on tenascin (Figure 3C; Figure 2F). Tn68, a MAb to an epitope in FNIII repeat 7, also blocked about 80% of sensory neuron outgrowth (Figure 3B; Figure 1E). Inhibito~ effects were substrate specific, since neurite outgrowth by sensory neurons on laminin and by motor neurons on N-cadherin appeared to be normal in the presence of each antibody tested (Figures 3B and 3C; Figure 1F; Figures 2K and 2L). In conclusion, tenascin-stimulated neurite outgrowth by either sensory or motor neurons requires interactions with a domain in the C-terminal region of substrate-bound tenascin. For sensory neurons, the inhibitory effect of MAb Tn68 makes it probable that this activity requires interaction with a do- main in or near FNIII repeat 7 (Figure 3A). These results are consistent with a previous report in which it was shown that sensory neurons respond to a domain in the last three FNIII repeats of tenascin (Wenrle-Haller and Chiquet, 1993).
Characterization of Antibodies to Integrin Receptor Subunits To determine whether individual integrin heterodimers function as receptors for tenascin, several integrin sub- unit-specific antibodies were tested for inhibitory activi- ties. The Chick 131 integrin-specific MAb, W1 B10, has pre- viously been shown to inhibit cellular interactions with laminin, collagen, and fibronectin (Hayashi et al., 1990). The anti-~v specific MAb, CHAV-1, has been shown to inhibit interactions of ~v subunit-containing integrins (Neuge- bauer et al., 1991). Two additional sera to gel-purified fu- sion proteins containing large portions of the extracellular domains of the chick integrin ~8 or a3 subunits were also prepared and tested (see Experimental Procedures). Anti- ~3-EX has been shown to inhibit interactions between cili- ary neurons and laminin-1, to immunoprecipitate the inte- grin ~3 subunit from chicken retinal lysates, and to blot this subunit (Weaver et al., submitted; and data not shown).
To characterize anti-as-EX serum, anti-~8-EX and anti- ~8-cyto IgG (specific for the a8 cytoplasmic tail [Bossy et al., 1991]) were used to immunoprecipitate surface-labeled
Neuron 1216
A Tenascin
EGF repeats FNnl repeats
I As474 I
AS142 i
B DRGs C Motor Neurons
120 120
= i o 80- 80
"1o .~ 60- 6 0 - - -
40- 40
Z 20 20
TN- TN- TN- TN- TN- AS142 AS474 mAb Tn68 AS142 AS474
chick embryo f ibroblast extracts. Autoradiograms of im- munoprecipi tates fract ionated by SDS-PAGE in nonre- ducing condit ions are presented in Figure 4. Two bands of Mr 110 and 160 kDa were observed in anti-a8-cyto immu- noprecipitates (Figure 4A, lane 2). Bossy et al. (1991) have shown with immunodeplet ion exper iments that the upper and lower bands correspond to the integrin a8 and 131 sub- units, respectively. In anti-as-EX immunoprecipi tates, two similar bands of Mr 110 and 160 kDa were seen (Figure 4A, lane 1). Together, these results indicate that anti<zs-EX specifically im munoprecipi tates the integrin ~z8 subunit and its associated ~ subunit.
Expression of In tegr in ~ Heterodimers by Sensory and Motor Neurons In previous exper iments, Bossy et al. (1991) detected the integrin ~z8 subunit on spinal sensory and motor neurons and along the dorsal and ventral roots of E6 chicks. Stain- ing with anti-as-EX also detects the a8 subunit expression in these areas (data not shown). To test whether DRG sensory neurons expressed the integrin c¢8 subunit in cul- ture and to identify associated ~ subunits, extracts from cultured DRG neurons were immunoprecip i tated with anti- as-cyto. Two bands of Mr 160 and 110 kDa were observed (Figure 4B). Therefore, anti-as-cyto immunoprecipi tated proteins with the same apparent M~s from both DRG and f ibroblast lysates (compare Figure 4B to Figure 4A, lane 2), making it l ikely that the upper and lower bands corre- spond to the integrin a8 and ~1 subunits, respectively. To verify that the 110 kDa sPecies was the I~1 subunit, unla- beled DRG lysates were immunoprecip i tated with anti-as- cyto, electrophoretical ly transferred to nitrocellulose, and blotted with the ~l-specific MAb WIB10. ~1 antigen was observed at a Mr of 110 kDa, indicating that the cx8 subunit
Figure 3. Schematic Drawing of a Tenascin Subunit Showing the Largest Splice Variant and the Effects of Anti-Tenascin Antibodies on Neurite Outgrowth by Sensory and Motor Neurons (A) Schematic drawing of a tenascin subunit showing the largest splice variant. Open squares, EGF repeats; closed ovals, FNIII re- peats present in all tenascin isoforms; open ovals, differentially spliced FNIII repeats not present in all isoforms. The RGD sequence in FNIII domain 3 is illustrated, and the large open circle depicts the domain homologous to a do- main in fibrinogen. The domains of tenascin recognized by two polyclonal antibodies (AS142 and AS474) and one MAb (TN68) are also shown. (B and C) Effects of anti-tenascin antibodies on neurite outgrowth by sensory (13) or motor (C) neurons. Neurons were dissected and dissoci- ated as described in Experimental Procedures and plated on substrate-bound tenascin (closed bars), laminin (striped bars), or N-cad- herin (open bars) in the presence of the anti- tenascin antibodies. Mean values for 2 wells from each experiment were normalized to mean values for control wells; n is listed below each bar. Values represent means -*- SD (for n = 3) or means _ range (for n = 2).
associates with the I~1 subunit in chick DRG (data not shown).
To identify addit ional integrin receptors expressed by DRG neurons or motor neurons cultured on tenascin, cul-
- - 200
- -116
- - 97
- - 6 6
- - 200
- -116
- - 97 i ! i
- - 6 6
Figure 4. Nonreducing SDS-PAGE of l~l-Surface Labeled Proteins (A) Chick embryo fibroblast lysates immunoprecipitated with anti-as-EX (Ex) or anti-aa-cyto (Cyto). (B) Dissociated E7.5 chick DRG neurons were cultured, labeled, lysed, and immunoprecipitated with anti-as-cyto. The positions of standards are indicated. Molecular weights are depicted in kilodaltons.
Integrin ~8~11s a Tenascin Receptor 1217
A DRGs 120
1oo
o a .
-o 60 N
== 40-
z 2 0 -
0
B Motor Neurons 140
.L I
100 I 80-
40
20-
2 1 6 5 3 1 3 3 ~ 3 3 3 1 4 4 1 3 2 ~ 0 - 3 2 3 2 1 3 3 2 1 3 2 31-MAb c~8-EX c~8-EX c~8-EX a3-EX cw-MAb J~I-MAb c~8-EX c~8-EX ~8-EX ow-MAb W1B10 IgG aff. pur. Fab' I,qG CHAV-1 W1B10 IgG Fab' IgG CHAV-1
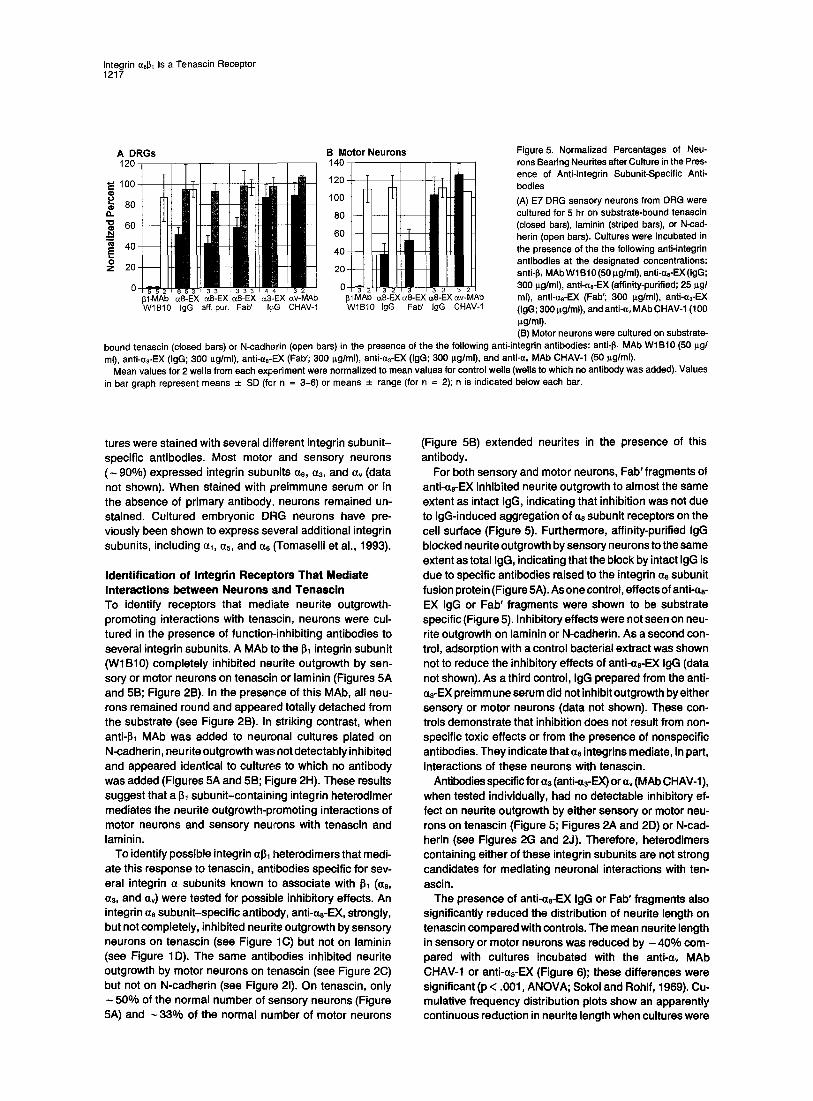
Figure 5. Normalized Percentages of Neu- rons Bearing Neurites after Culture in the Pres- ence of Anti-lntegrin Subunit-Specific Anti- bodies (A) E7 DRG sensory neurons from DRG were cultured for 5 hr on substrate-bound tenascin (closed bars), laminin (striped bars), or N-cad- herin (open bars). Cultures were incubated in the presence of the the following anti-integrin antibodies at the designated concentrations: anti-I~l MAb W1 B10 (50 i~g/ml), anti-as-EX (IgG; 300 p.g/ml), anti-as-EX (affinity-purified; 25 p.g/ ml), anti-as-EX (Fab'; 300 p.g/ml), anti-a3-EX (IgG; 300 I~g/ml), and anti-a, MAb CHAV-1 (100 p~g/ml). (B) Motor neurons were cultured on substrate-
bound tenascin (closed bars) or N-cadherin (open bars) in the presence of the the following anti-integrin antibodies: anti-I~l MAb WIB10 (50 p.g/ ml), anti-as-EX (IgG; 300 p.g/ml), anti-as-EX (Fab'; 300 i~g/ml), anti-a3-EX (IgG; 300 p.g/ml), and anti-av MAb CHAV-1 (50 p.g/ml).
Mean values for 2 wells from each experiment were normalized to mean values for control wells (wells to which no antibody was added). Values in bar graph represent means _+ SD (fcr n = 3-6) or means -+ range (for n = 2); n is indicated below each bar.
tures were stained with several different integrin subunit- specific antibodies. Most motor and sensory neurons ( - 90%) expressed integrin subunits as, a3, and av (data not shown). When stained with preimmune serum or in the absence of primary antibody,, neurons remained un- stained. Cultured embryonic DRG neurons have pre- viously been shown to express several additional integrin subunits, including al, as, and as (Tomaselli et al., 1993).
Identification of Integrin Receptors That Mediate Interactions between Neurons and Tenascin To identify receptors that mediate neurite outgrowth- promoting interactions with tenascin, neurons were cul- tured in the presence of function-inhibiting antibodies to several integrin subunits. A MAb to the I~1 integrin subunit (WlB10) completely inhibited neurite outgrowth by sen- sory or motor neurons on tenascin or laminin (Figures 5A and 5B; Figure 2B). In the presence of this MAb, all neu- rons remained round and appeared totally detached from the substrate (see Figure 2B). In striking contrast, when anti-I~t MAb was added to neuronal cultures plated on N-cadherin, neu rite outgrowth was not detectably inhibited and appeared identical to cultures to which no antibody was added (Figures 5A and 5B; Figure 2H). These results suggest that a I~ subunit-containing integrin heterodimer mediates the neurite outgrowth-promoting interactions of motor neurons and sensory neurons with tenascin and laminin.
To identify possible integrin al~l heterodimers that medi- ate this response to tenascin, antibodies specific for sev- eral integrin • subunits known to associate with 61 (as, a3, and av) were tested for possible inhibitory effects. An integrin as subunit-specific antibody, anti-as-EX, strongly, but not completely, inhibited neurite outgrowth by sensory neurons on tenascin (see Figure 1C) but not on laminin (see Figure 1D). The same antibodies inhibited neurite outgrowth by motor neurons on tenascin (see Figure 2C) but not on N-cadherin (see Figure 21). On tenascin, only
50% of the normal number of sensory neurons (Figure 5A) and - 3 3 % of the normal number of motor neurons
(Figure 5B) extended neurites in the presence of this antibody.
For both sensory and motor neurons, Fab' fragments of anti-as-EX inhibited neurite outgrowth to almost the same extent as intact IgG, indicating that inhibition was not due to IgG-induced aggregation of a8 subunit receptors on the cell surface (Figure 5). Furthermore, affinity-purified IgG blocked neurite outgrowth by sensory neurons to the same extent as total IgG, indicating that the block by intact IgG is due to specific antibodies raised to the integrin a8 subunit fusion protein (Figure 5A). As one control, effects of anti-as- EX IgG or Fab' fragments were shown to be substrate specific (Figure 5). Inhibitory effects were not seen on neu- rite outgrowth on laminin or N-cadherin. As a second con- trol, adsorption with a control bacterial extract was shown not to reduce the inhibitory effects of anti-as-EX IgG (data not shown). As a third control, IgG prepared from the anti- as-EX preimm une serum did not inhibit outgrowth by either sensory or motor neurons (data not shown). These con- trois demonstrate that inhibition does not result from non- specific toxic effects or from the presence of nonspecific antibodies. They indicate that a8 integrins mediate, in part, interactions of these neurons with tenascin.
Antibodies specific for a3 (anti-0~-EX) or a, (MAb CHAV-1), when tested individually, had no detectable inhibitory ef- fect on neurite outgrowth by either sensory or motor neu- rons on tenascin (Figure 5; Figures 2A and 2D) or N-cad- herin (see Figures 2G and 2J). Therefore, heterodimers containing either of these integrin subunits are not strong candidates for mediating neuronal interactions with ten- ascin.
The presence of anti-as-EX IgG or Fab' fragments also significantly reduced the distribution of neurite length on tenascin compared with controls. The mean neurite length in sensory or motor neurons was reduced by - 4 0 % com- pared with cultures incubated with the anti-av MAb CHAV-1 or anti-a3-EX (Figure 6); these differences were significant (p < .001, ANOVA; Sokol and Rohlf, 1969). Cu- mulative frequency distribution plots show an apparently continuous reduction in neurite length when cultures were
N e u r o n 1 2 1 8
A D R G s
80
60 ¢= o ._~ 40
20-
B Motor Neurons 1
0
aff. pur. CHAV-1 IgG aff. pur. CHAV-I
C D R G s D Motor Neurons 100 L - - , 100 , • •
o =~ t ' ~ - , I 60
o 0 36 76 116 156 43 83 123 163
Microns Microns
Figure 6. Effect of integrin-Specific Antibodies on Neurite Lengths in Sensory and Motor Neuron Cultures (A and C) DRG sensory neurons were cultured for 24 hr. Processes longer than 20 p,m were measured for 20 neurons in each well, and a mean length was calculated for each well. Mean values (A) represent the means _+ SEM for 4 wells from two experiments for the control and anti-Qs-EX, and the mean -+ range for 2 wells from a single experi- ment for anti-ev MAb CHAV-1. (C) shows a cumulative frequency distri- bution plot for the same neurons. The plot shows the percentage of neurons with neurites longer than a given length in micrometers (closed squares, control [no antibody]; closed circles, affinity-purified anti-~e-EX; closed triangles, anti-~v MAb CHAV-1). For sensory neu- rons, mean lengths (_+ SEM) were 92 _+ 8 p,m (control, anti-a3-EX), 64 - 6 pm (anti-(zs-EX), and 94 _+ 5 p,m (anti-~v). (B and D) Mean length and cumulative frequency distribution plots for motor neurons cultured for 7 hr. Motor neurons were incubated with anti-ct3-EX (control; closed squares), anti-~8-EX (closed circles), or anti- Qv MAb CHAV-1 (closed triangles). Mean values (B) represent the mean for 5 wells from three experiments. Cumulative frequency distribution plots (D) are for the same neurons as in (B). For motor neurons, mean lengths (_+ SEM)were 66 _ 3 p,m (control; no antibody), 44 _ 1 p.rn (anti-~ze-EX), and 74 _+ 5 p.m (anti-o~). For both sensory and motor neurons, differences in mean length between control or anti-~ versus anti-~s cultures were significant, as determined by ANOVA (p < .001; Sokol and Rohlf, 1969).
incubated with anti-czs-EX compared with anti-ev or no anti- body (Figure 6). Consistent with previous results demon- strat ing that the ae subunit is expressed by a lmost all motor neurons and DRG sensory neurons, this suggests that neuri te outgrowth on tenascin by the major i ty of neurons was inhibited by anti-es-EX IgG or Fab' f ragments in these
cultures.
A 1(562 Cell Line Expressing the Integrin o~ Subunit Exhibits Increased Attachment to Tenascin To provide addi t ional ev idence that the 0~8~1 heterodimer funct ions as a tenascin receptor, K562 cells (a human my- e logenous leukemia cell line) were t ransfected with a full- length chicken a0 cDNA in the vector pAW, which directs transcript ion f rom a CMV promoter. K562 cell l ines have been shown to express predominant ly a5131 and, to a lesser extent, ave5 integrins (Pasqual ini et al., 1993). Integrin as transfectants were screened for expression of the integrin a8 subunit, and those cell l ines express ing maximal
A B 110 110
~_ 7o 60 ~ eo 50 5O 40 ~ 40 3o ~ ao 20 2O
c~8-EX c~8-EX o¢8-EX 131-MAb c<v-ab
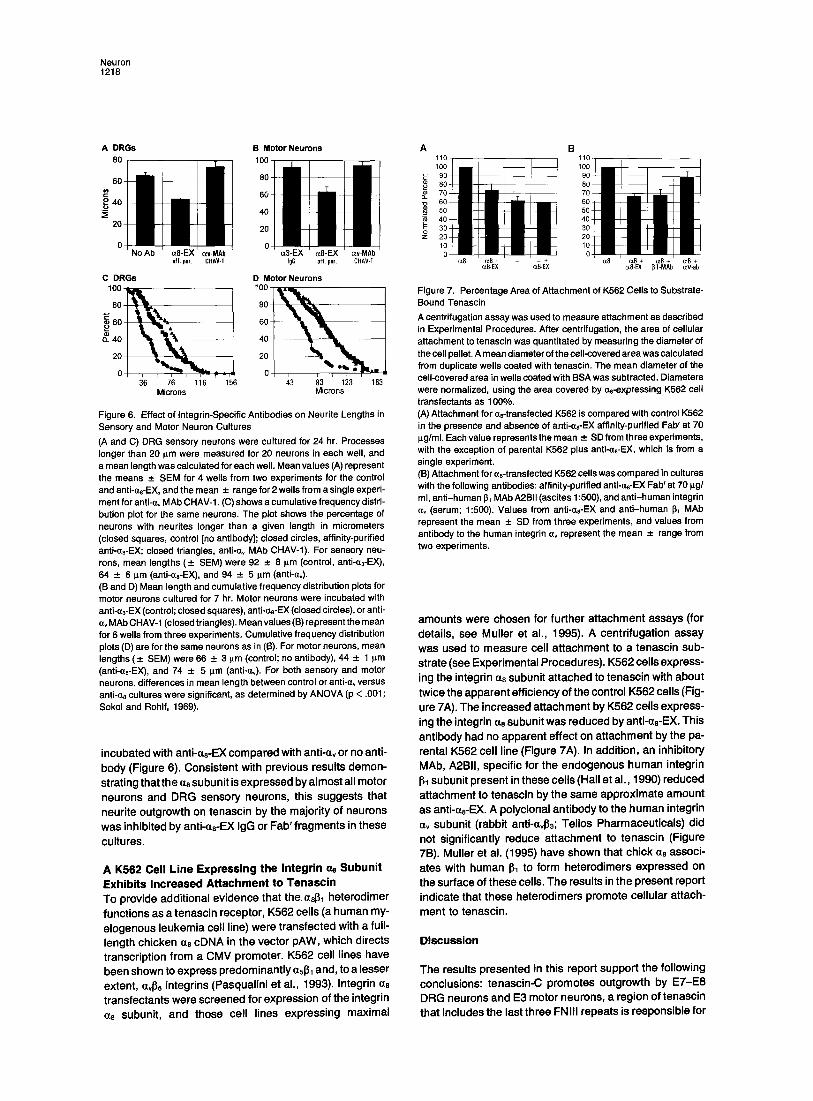
Figure 7. Percentage Area of Attachment of K562 Cells to Substrate- Bound Tenascin A centrifugation assay was used to measure attachment as described in Experimental Procedures. After centrifugation, the area of cellular attachment to tenascin was quantitated by measuring the diameter of the cell pellet. A mean diameter of the cell-covered area was calculated from duplicate wells coated with tenascin. The mean diameter of the cell-covered area in wells coated with BSA was subtracted. Diameters were normalized, using the area covered by aB-expressing K562 cell transfectants as 100%. (A) Attachment for ae-transfected K562 is compared with control K562 in the presence and absence of anti-as-EX affinity-purified Fab' at 70 p.g/ml. Each value represents the mean -+ SD from three experiments, with the exception of parental K562 plus anti-(ze-EX, which is from a single experiment. (B) Attachment for ~8-transfected K562 cells was compared in cultures with the following antibodies: affinity-purified anti-ae-EX Fab' at 70 pg/ ml, anti-human ~1 MAb A2.BII (ascites 1:500), and anti-human integrin ~v (serum; 1:500). Values from anti-czs-EX and anti-human ~1 MAb represent the mean _+ SD from three experiments, and values from antibody to the human integrin av represent the mean -*- range from two experiments.
amounts were chosen for further a t tachment assays (for details, see Mul ler et al., 1995). A centr i fugat ion assay was used to measure cell a t tachment to a tenascin sub- strate (see Exper imenta l Procedures). K562 cells express- ing the integrin a0 subunit a t tached to tenascin with about twice the apparent eff iciency of the control K562 cells (Fig- ure 7A). The increased a t tachment by K562 cells express- ing the integrin ms subunit was reduced by anti-as-EX. This ant ibody had no apparent effect on a t tachment by the pa- rental K562 cell l ine (Figure 7A). In addit ion, an inhibi tory MAb, A2BII, specif ic for the endogenous human integrin 131 subunit present in these cel ls (Hall et al., 1990) reduced at tachment to tenascin by the same approx imate amount as anti-ae-EX. A polyclonal ant ibody to the human integrin av subunit (rabbit anti-ct4~3; Tel ios Pharmaceut icals) did not signif icantly reduce at tachment to tenascin (Figure 7B). Mul ler et al. (1995) have shown that chick as associ- ates with human I~1 to form heterodimers expressed on the surface of these cells. The results in the present report indicate that these heterodimers promote cel lu lar attach-
ment to tenascin.
Discussion
The results presented in this report suppor t the fol lowing conclusions: tenascin-C promotes outgrowth by ET-E8 DRG neurons and E3 motor neurons, a region of tenascin that includes the last three FNIII repeats is responsible for

Integrin a8131 Is a Tenascin Receptor 1219
most neurite outgrowth-promoting effects, and the integrin receptor (z813~ functions as a tenascin receptor on these neurons.
Sensory and Motor Neuron Outgrowth on Tenascin At stage 19 of chick development, tenascin is found asso- ciated with the tips of motor and sensory neuron axons as they emerge from the neural tube and DRG, respec- tively, and grow through the anterior region of each somite (Wehrle and Chiquet, 1990). In this report, we show that neurite outgrowth by sensory and motor neurons is pro- moted by substrate-bound tenascin. Previously, it was re- ported that tenascin supports neurite outgrowth in spinal cord cultures from day 3 (stage 19) embryos and in sensory neurons from embyros at both day 4 (stage 20-22) and later developmental stages (Wehrle and Chiquet, 1990; Wehrle-Haller and Chiquet, 1993). Therefore, our results and those of other investigators show that in vitro tenascin has the potential to promote outgrowth from a subset of neurons through the anterior region of each somite, at stage 19 as well as later stages of the chick embryo devel- opment. The distribution of tenascin in vivo is consistent with the possibility that it functions similarly during em- bryogenesis.
Neurite outgrowth by sensory or motor neurons requires interactions with a domain of tenascin in the last three FNlll repeats. Antibodies raised to a pronase fragment of tenascin containing FNIII repeats 6-8 inhibited neurite outgrowth by motor ( - 9 0 % ) or sensory ( - 8 0 % ) neurons on substrate-bound tenascin. In addition, MAb Tn68, which recognizes an epitope in FNIII repeat 7, inhibited
80% of neurite outgrowth. These results are consistent with previous reports in which an antibody raised to the C-terminal fragment inhibited neurite outgrowth on tenas- cin from intact E6 DRG (Wehrle-Haller and Chiquet, 1993). Other investigators have also found that a MAb to this region inhibited neurite outgrowth on substrate-bound ten- ascin by dissociated cerebellar, hfppocampal, or mesen- cephalic neurons (Husmann et aL, 1992; Lochter et al., 1991). It is possible that an additional region in tenascin is also required for neurite outgrowth, since neurons from intact DRGs did not extend neurites on the 60 kDa C-ter- minal fragment of tenascin used to generate AS474 (Wehrle-Haller and Chiquet, 1993).
The Integrin as~ as a Receptor for Tenascin Experiments presented in this and previous reports indi- cate that neuronal outgrowth on a tenascin substrate re- quires ~-containing integrin heterodimers. In the present report, we show that an integrin 131-specific MAb (W1 B10) completely blocks neurite outgrowth by dissociated sen- sory or motor neurons on this substrate. This result corrob- orates a recently published report in which a different inte- grin ~-specif ic MAb (JG22) blocked neurite outgrowth from cultures of intact DRGs (Wehrle-Haller and Chiquet, 1993). Other investigations have demonstrated that an in- tegrin containing the 13~ subunit is a receptor for attach- ment of nonneuronal cell lines to non-RGD-containing re- gions of tenascin (Sriramarao et al., 1993; Prieto et al., 1993).
To identify integrin a subunits that mediate interactions with tenascin, we developed and tested several (z subunit- specific antisera. The antisera generated, anti-(zs-EX and anti-(~a-EX, efficiently immunoprecipitated their respective (z subunits and blocked ligand-receptor interactions. Anti- (zs-EX inhibited significantly, but not completely, neurite outgrowth by sensory and motor neurons on tenascin. The inhibitory effect was specific to tenascin and was not ob- served on laminin or N-cadherin. In addition, affinity- purified IgG and monovalent anti-(~8-EX Fab' were effec- tive. These results strongly implicate (z8131 as a tenascin receptor.
Since the block with anti-as-EX was incomplete, an addi- tional (~1~1 heterodimer may also help mediate neurite out- growth-promoting interactions. Alternatively, anti-as-EX may not inhibit (zsI3~-mediated interactions completely. Data in the present report suggest that the (z3 and the (~v subunit-containing heterodimers are not strong candi- dates to be tenascin receptors on these neurons, even though an (~v heterodimer has been shown to mediate in- teractions by nonneural cells with this protein (Prieto et al., 1993; Sriramarao et al., 1993). Neither (~3-specific nor (~v-specific antibodies detectably reduced neurite out- growth on tenascin, even though each has been shown to be inhibitory on other ECM substrates, e.g., laminin and vitronectin, respectively (Neugebauer et al., 1991; Weaver et al., submitted). ~213~ has been shown to mediate some interactions with tenascin by nonneuronal cells (Srira- marao et al., 1993) and is therefore a candidate to function as an additional tenascin receptor on motor and sensory neurons.
Experiments assaying cellular interactions with tenascin have produced a confusing literature. Substrate-bound or soluble tenascin has promoted or inhibited cell attachment or neurite outgrowth, depending on the cell type and/or method of plating tenascin. In immunohistochemical stud- ies on the expression of c(8 in developing chicks, Bossy et al. (1991) detected (~8 on subsets of neurons and axon tracts within the central and peripheral nervous systems. It seems likely that the presence or absence of (zs~ hetero- dimers is a major determinant of these strikingly different responses of neurons to tenascin.
Experimental Procedures
Purification of Substrate Proteins A tenascin purification was developed based on previously described methods (Huber et al., 1986; Vaughan et al., 1987). Sterna (40-60) were dissected from E17 chicks and immediately frozen with liquid N2. Sterna were weighed, and 1.0 ml of extraction buffer was added for each 0.1 g of sterna (1 M NaCI, 0.05 M Tris-HCI [pH 8.0], plus chymostatin, leupeptin, aprotinin, pepstatin [each at 5 i~g/ml], and 1 mM phenylmethylsulfonyl fluoride). Sterna were homogenized (Poly- tron) and extracted for 48 hr at 4°C. Extracted sterna were then spun at 12,000 x g for 15 min, and the supernatant was precipitated with 50% saturated ammonium sulfate for 1 hr. After spinning at 12,000 x g for 10 rain, the pellet was resuspended in PBS (8 mM Na2HPO4, 2 mM KH2PO4, 2 mM KCI, 0.136 M NaCI [pH 7.4]) containing protease inhibitors (see above) and dialyzed against PBS overnight. Tenascin was purified by affinity chromatography using the tenascin-specific MAb M 1 IgG coupled to CNBR Sepharose 4B (Pharmacia, Piscataway, NJ). The dialyzed protein solution was applied to the column. After extensive washing with PBS and PBS containing 0.1% Triton X-100,

Neuron 1220
tenascin was eluted with 80 mM diethylamine (pH 11.0). Eluted frac- tions were immediately neutralized with 1 M Na-HEPES (pH 7.0). Eluted material was pooled, dialyzed against PBS, and analyzed on polyacrylamide gels. The predominant species of tenascin in these preparations had a Mr of 190 kDa, indicating that it lacks the alterna- tively spliced FNIII repeats present in larger tenascin isoforms (de- picted in Figure 3). Purified tenascin was not reactive with an antibody to chicken fibrenectin (see Lefcort et al., 1992).
The extracellular domain of N-cadherin was expressed in COS cells and purified as described elsewhere (Kypta et al., unpublished data). Substrata-bound N-cadherin from these preparations promoted outgrowth from retinal neurons, which was inhibited with the anti- N-cadherin MAb NCD2.
Laminin was purified from Engelbreth-Holm-Swarm (EHS) sar- comatumors using published procedures (Kleinman et al., 1982; Timpl et al., 1982).
Substrate Preparation Linbre Titertek 96 well plastic dishes (Flow Laboratories, McLean, VA) were coated with nitrocellulose as described previously (Lagenauer and Lemmon, 1987). Purified tenascin (30-40 p.g/ml in PBS), N-cad- herin (30-70 i~g/ml in PBS plus 1 mM of CaCI2, 1 mM MgCI2), or laminin (10 p.g/mt in PBS) was added and incubated either for 15 rain at room temperature or overnight at 4°C.
Cell Culture Motor neuron cultures were prepared from stage 19 embryos ac- cording to procedures adapted from Henderson et al. (1984) and Wehrle and Chiquet (1990). Embryos were dissected from mem- branes, washed in PBS, and incubated in pancreatin (GIBCO, Grand island, NY) for 2 rain. Embryos were then placed in cold L-15 medium containing 5 mg/ml BSA (Sigma, St. Louis, MO), and neural tubes were isolated by dissection. Dorsal and ventral neural tube fragments were dissected with a glass needle. Ventral neural tubes were then placed in 0.25% trypsin for 5 min at 37°C, washed in L-15 containing 10 mg/ml BSA, centrifuged, and resuspended in complete motor neuron growth medium (F12 medium containing 0.5 mg/ml BSA [Serva, Ger- many]), brain-derived neurotrophic factor (diluted 1:1000; a generous gift from Dr. Patricia Olson, Chiron Corporation, Emeryville, CA), and 10 ng/ml basic flbroblast growth factor (Boehringer Mannheim, India- napolis, IN). Neural tubes were gentlytriturated four times. Dissociated cells were centrifuged, resuspended in growth medium to an appro- priate density (1 x 103 cells/well), plated in 100 p.I of growth medium, and incubated for 24 hr at 37°C with 5°/o CO2.
For sensory neuron cultures, E7-E8 chick DRGs were dissected and dissociated into single cells by incubation in 0.05% trypsin in 0.20/o versene, 0.10o/o glucose, 0.02°/0 EDTA, 0.0580/o NaHCO2 (UCSF Cell Culture Facility) for 10 min at 37°C, followed by trituration. Dissociated cells were collected by centrifugation and resuspended in F12 con- taining 10% fetal bovine serum. To enrich for neuronal cells, cell sus- pensions were plated onto 60 mm tissue culture dishes (Falcon) for 1-3 hr. Neurons were pipetted from the culture dishes, centrifuged, and resuspended in DRG growth medium (F12 containing 2% BSA [Serva] and 100 ng/ml nerve growth factor) at an appropriate density (1 x 102 cells/well). Fifty microliters of this cell suspension was added to the well of a 96 well culture dish containing antibody diluted to the appropriate concentration in 50 p.I of warmed and equilibrated growth medium. Neurons were gently centrifuged onto the dish and incubated for 4-7 hr at 37°C in a 5o/0 CO2 atmosphere.
Quantitation of Neurite Outgrowth For motor neurons cultured on tenascin or N-cadherin, neurons with neurites were defined as cells extending neurites longer than two cell diameters. For sensory neurons on tenascin, laminin, or N-cadherin, the percentage of neurons with neurites per total number of cells was determined. To measure neurite length of either motor or sensory neurons, cultures were viewed with an inverted Olympus IMT2 micro- scope using Nomarski optics. Images were collected with a cooled CCD camera (Photometric.s, series 200). Processes longer than 20 p.m were measured using the Prism program (Chen et al., 1989).
Tenascin and Integrin Antibodies Tenascin antibodies AS142 (Chiquet and Fambrough, 1984) and MAb
Tn68 (Chiquet-Ehrismann et al., 1988) are specific for whole tenascin and an epitope within FNIII repeat 7, respectively. Tenascin antibody AS474 is specific for a pronase-generated C-terminal fragment of ten- ascin containing FNIII repeats 6-8 plus the C-terminal fibrinogen-like module. These have been described previously (Wehrle-Haller and Chiquet, 1993).
Anti-~8-EX was generated to an SDS-PAG E-purified GST-a8 fusion protein expressed in E. coiL The sequence included amino acids 62- 710 of the chick ~zs protein (with an internal, in-frame deletion of amino acids 150-185). This antibody is specific for chick integrin ~z8 as as- sessed by immunoblot or immunoprecipitation. It is more fully charac- terized elsewhere (M uller et al., 1995). Anti-a3-EX was generated to an SDS-PAGE-purified ~-galactosidase-as fusion protein that contained amino acids 17-558 of the chick a3 protein. This antibody is specific for chick integrin a3 as assessed by immunoblot and immunoprecipita- tion. It inhibits interactions of ciliary neurons with laminin-1 and is more fully characterized elsewhere (Weaver et al., submitted). Anti-as-cyto was prepared using a synthetic peptide corresponding to the cyto- plasmic domain of the chick ~8 subunit (Bossy et al., 1991).
IgG from rabbit sera or from anti-~l chick MAb W1 B10 ascites (Hay- ashi et al., 1990) was purified using MAC protein G disks (Amicon, Beverly, MA) according to the manufacturer's instructions. IgG from anti-chick ~v MAb CHAV-1 ascites (Neugebauer et al., 1991) was puri- fied from ascites using affinity chromatography on protein A-Sepha- rose CI-4B according to the manufacturer's instructions (Pharmacia). All IgG preparations were dialyzed extensively against PBS and steri- lized. The as and as IgG preparations were further dialyzed against F12 medium.
To affinity purify anti-as-EX IgG, 1 liter cultures of bacteria express- ing either the as-EX fusion protein or a control fusion protein were induced with 0.1 mM isoprepyl-~-thiogalactopyranoside for 4 hr. The bacterial pellets were collected, resuspended in 9.0 ml of PBS with pretease inhibitors, sonicated, and centrifuged for 10 rain at 10,000 x g. The resultant pellets were solubilized in 1.5% N-lauroylsarcosine (Sigma), 2% Triton X-100 (Sigma) in PBS and pretease inhibitors. This solution was extracted on ice for 1 hr and centrifuged for 10 min at 10,000 x g. A 10 x 2 cm piece of nitrocellulose membrane (Schleicher & Schuell, Keene, NH) was immersed in the supernatant for 1 hr. Before use for affinity purification of IgG, each membrane was washed extensively with PBS, pre-eluted with 0.1 M glycine (pH 2.7), equili- brated with PBS, and incubated with IgG for 1 hr. The anti-~8-EX IgG preparation was incubated first with a nitrocellulose membrane con- taining the immobilized control E. coil extract and then with a mem- brane containing the immobilized a8 fusion protein. After washing with PBS, affinity-purified as-EX was eluted from the ~z8 fusion protein- containing membrane with 0.1 M glycine (pH 2.7) and was immediately neutralized with 1 M Tris-CI (pH 8.5). This was dialyzed against PBS followed by F12 medium and was sterilized by filtration.
To generate Fab' fragments, IgG was dissolved in PBS and dialyzed against 100 mM Na-acetate (pH 5.5). Papain-agarose (Sigma) was incubated with the dialysate for 1.5 hr at 37°C. The reaction was stopped with 75 mM iodoacetimide (Sigma) for 30 min at room tempera- ture. The Fab' solution was then dialyzed against PBS and subse- quently against F12 medium.
Immunocytochemistry Cell cultures in 96 well dishes were fixed with 4% paraformaldehyde for 10 min, carefully washed, and incubated for 2 hr with integrin subunit antibodies (anti-e~-EX [16 ilg/ml IgG], anti-~3-EX [1:500 serum], or anti-~v MAb CHAV-1 [20 p_g/ml IgG]) diluted in PBS with 5°/o normal goat or horse serum. Preimmune controls were incubated at the same concentrations. Cultures were carefully washed and stained further according to instructions included in the Vectastain kits (Vector Labo- ratories).
Immunoprecipitations Chick embryo fibroblasts were prepared from E9 or El0 embryos ac- cording to Rein and Rubin (1968). Cells were resuspended in PBS and surface labeled in suspension with lactoperoxidase, H202, and 1251, as described below.
Dissociated DRG neurons were cultured for 24 hr on t 00 mm 2 Fal- con tissue culture dishes previously coated with a mixed solution of vitmnectin (1 p.g/ml; GIBCO) and fibronectin (20 p.g/ml; Calbiochem,

tntegrin (~81~ Is a Tenascin Receptor 1221
San Diego, CA) dissolved in PBS. After removal of growth medium and washing with PBS, cultures were surface labeled by adding lacto- peroxidase, H202, and ~2Sl in PBS for 15 rain. Lysates were prepared by adding 1.2 ml of lysis buffer containing 1% Triton X-100 in PBS and protease inhibitors to the cultures. Lysed material was then scraped from the dish with a cell scraper and centrifuged at 20,000 x g for 20 min at 4°C. After preclearing twice with 100 I~1 of protein A-Sepharose, supernatants were incubated for 4 hr with specific anti- bodies coupled to protein A-Sepharose (50 p.I of beads per culture; Pierce, Rockford, I L). Antibodies were coupled to protein A-sepharose with dimethylpimelimidate using the st~.ndard procedure (Harlow and Lane, 1988). Beads were then washed six times by adding 1.0 ml of 1% Triton X-100 in PBS and pelletinc beads at 134 x g for 10 s. Immunoprecipitated proteins were then separated with 6% SDS- PAGE in nonreducing conditions. Gels were dried and exposed to Kodak X Omat R film.
To identify protein species coprecipitating with (~8, unlabeled lysates prepared as above were immunoprecipitated with anti-(~8-EX or anti-~8- cyto. After fractionation by 6% SDS-P,Z~GE and electrophoretic trans- fer to nitrocellulose filters (Schleicher ~. Schuell), blots were blocked for 30 min with blocking buffer (5% Csrnation nonfat dry milk in 100 mM Tris-HCI [pH 7.5], 150 mM NaCI). The blots were then incubated with the anti-~l MAb Wl B10 (10 llg/m[! for 1 hr. After washing, blots were incubated for 1 hr with horseradish peroxidase-rabbit anti-mouse {gG (Zymed Corp.). After washing, the bands were visualized with electrochemical luminescence using a3 ECL kit and its enclosed in- structions (Amersham, Arlington Heigt"ts, IL).
Cell Attachment Assay A centrifugation assay was used to measure interactions of K562 cells with tenascin (Friediander et al., 1988). Wells of a Linbro Titertek 96 well round bottom dish (Linbro) were coated with 100 I11 of a nitrocellu- lose solution, washed with PBS, coated with 40 pl of 20-40 p.g/ml tenascin in PBS for 2 hr at 37°C, washed with PBS, and coated with a 10 mg/ml BSA solution for 1 hr at 37°C. Before use, wells were equilibrated with 24 mM Tris-CI (pH 7.5), 137 mM NaCI, 2.7 mM KCI, 2 mM glucose, 2 m M CaCI2, 2 mM MgCI;, 2 mg/ml BSA. An appropriate number of mid-log phase K562 cells were then removed from growth medium (RPMI plus 10% fetal calf serum), centrifuged at 500 x g for 1 min, and resuspended at 5 x 104 cells per 50 I~1 in the Tris-buffered BSA solution. After incubation with antibodies at 4°C for 20 min, 50 ~1 of cell suspension was then added to each well and was centrifuged immediately at 250 x g for 4 min.
After centrifugation, cell pellets were viewed with a dissection micro- scope using dark-field optics, imaged with a CCD camera, and pro- jected to a television screen. Diameters of cell pellets were measured to quantitate attachment (Pfieto et al. 1992). Unattached cells ap- peared as small, tight pellets in the center of the well, e.g., BSA-coated wells had pellet diameters of - 1 mm. Tightly attached cells were evenly spread over the entire diameter of the plated substrate and had a diameter of - 3 ram. The pellet diameter in the BSA-coated wells was subtracted from the measured diameter to quantitate cell- substrate interactions.
Acknowledgments
All correspondence should be addressed to L. F. R. We thank Dr. Blaise Bossy for assistance in preparing the ~8-transfected K562 cell lines and the anti-~8-EX antibody, Dr. Susannah Chang for the DM1 MAb, Dr. Karla Neugebauer for helping to develop the tenascin purifi- cation, and Ms. Cristina Weaver for reading the manuscript and for helpful discussions. This work was supported by National Institutes of Health grant PO1-16033. L. F. R. is an investigator of the Howard Hughes Medical Institute.
The costs of publication of this article were defrayed in part by the payment of page charges. This artcle must therefore be hereby marked "advertisement" in accordance with 18 USC Section 1734 solely to indicate this fact.
Received September 16, 1994; revisec March 21, 1995.
References
Bartsch, S,, Bartsch U., Dorries, U., Faissner, A., Weller, A., Ekblom,
P., and Schachner, M. (1992). Expression of tenascin in the developing and adult cerebellar cortex. J. Neurosci. 12, 736-748.
Bossy, B., Bossy-Wetzel, E., and Reichardt, L. F. (1991). Characteriza- tion of the integrin a8 subunit: a new integrin 131-associated subunit, which is prominently expressed on axons and on cells in contact with basal laminae in chick embryos. EMBO J. 10, 2375-2385.
Burns, F. R., von Kannen, S., Guy, L., Raper, J. A., Kamholz, J., and Chang, S. (1991). DM-GRASP, a novel immunoglobulin superfamily axonal surface protein that supports neurite extension. Neuron 7, 209- 220. Bourdon, M. A., and Ruoslahti, E. (1989). Tenascin mediates cell at- tachment through an RGD-dependent receptor. J. Cell Biol. 108, 1149-1155. Chen, H., Sedat, J. W., and Agard, D. A. (1989). Manipulation, display and analysis of three dimensional biological images. In The Handbook of Biological Confocal Microscopy, J. Pawley, ed. (Madison, Wiscon- sin: IMR Press), pp. 127-135.
Chiquet, M., and Fambrough, D. M. (1984). Chick myotendinous anti- gen. I. A monoclonal antibody as a marker for tendon and muscle morphogenesis. J. Cell Biol. 98, 1226-1236. Chiquet-Ehrismann, R., Kalla, P., Pearson, C., Beck, K., and Chiquet, M. (1988). Tenascin interferes with fibronectin action. Cell 53, 383- 390.
Faissner, A., and Kruse, J. (1990). J 1/Tenascin is a repulsive substrate for central nervous system neurons. Neuron 5, 627-637.
Friedlander, D. R., Hoffman, S., and Edelman, G. M. (1988). Functional mapping of cytotactin: proteolytic fragments active in cell-substrate adhesion. J. Cell Biol. 107, 2329-2340.
Goodman, C. S., and Shatz, C. J. (1993). Developmental mechanisms that generate precise patterns of neuronal connectivity. Cell 721Neu- ron 10 (Suppl.), 77-98.
Halfter, W., Chiquet-Ehrismann, R., and Tucker, R. P. (1989). The effect of tenascin and embryonic basal lamina on the behavior and morphology of neural crest cells in vitro. Dev. Biol. 128, 245-255.
Hall, D. E., Reichardt, L. F., Crowley, E., Holley, B., Moezzi, H., Son- nenberg, A., and Damsky, C. H. (1990). The (11/1~ and (z6/{}~ integrin heterodimers mediate cell attachment to dinstinct sites on laminin. J. Cell Biol. 110, 2175-2184.
Harlow, E., and Lane, D. (1988). Antibodies (Cold Spring Harbor, New York: Cold Spring Harbor Laboratory), pp. 521-523.
Hayashi, Y., Haimovich, B., Reszka, A., Boettiger, D., and Horwitz, A. (1990). Expression and function of chicken intregrin ~ subunit and its cytoplasmic domain mutants in mouse NIH3T3 cells. J. Cell Biol. 110, 175-184.
Henderson, C. E., Huchet, M., and Changeux, J.-P. (1984). Neurite- promoting activities for embryonic spinal neurons and their develop- mental changes in the chick. Dev. Biol. 104, 336-347.
Hoffman, S., Crossin, K. L., and Edelman, G. M. (1988). Molecular forms, binding functions, and developmental expression patterns of cytotactin and cytotactin-binding proteoglycan, an interactive pair of extracellular matrix molecules. J. Cell Biol. 106, 519-532.
Huber, S., van der Rest, M., Bruckner, P., Rodriguez, E., Winterhalter, K. H., and Vaughan, L. (1986). Identification of the type IX collagen polypeptide chains. The a2 (IX) polypeptide carries the condroitin sul- fate chain(s). J. Biol. Chem. 261, 5965-5968.
Husmann, K., Faissner, A., and Schachner, M. (1992). Tenascin pro- motes cerebellar granule cell migration and neurite outgrowth by differ- ent domains in the fibronectin type III repeats. J. Cell Biol. 116, 1475- 1486.
Kleinman, H. K., McGarvey, M. L., Liotta, L. A., Robey, P. G., Tryggva- son, K., and Martin, G. R. (1982). Isolation and characterization of type IV procollagen, laminin, and heparin sulfate proteoglycan from the EHS sarcoma. Biochemistry 21, 6188-6193.
Lagenauer, C., and Lemmon, V. (1987). An Ll-like molecule, the 8D9 antigen, is a potent substrate for neurite extension. Proc. Natl. Acad. Sci. USA 84, 7753-7757.
Lefcort, F., Venstrom, K., McDonald, J. A., and Reichardt, L F. (1992). Regulation of expression of fibronectin and its receptor (lsl~ during

Neuron 1222
development and regeneration of peripheral nerve. Development 116, 767-782.
Lochter, A, Vaughan, L., Kaplony, A., Prochiantz, A., Schachner, M., and Faissner, A. (1991). Jl/tenascin in substrete-bound and soluble form displays contrary effects on neurite outgrowth. J. Cell Biol. 113, 1159-1171.
Muller, U., Bossy, B., Venstrom, K., and Reichardt, L. F. (1995). Inte- grin ~aJ~l promotes attachment, cell spreading and neurite outgrowth on fibronectin. Mol. Biol. Cell 6, 433-448.
Neugebauer, K. M., Emmett, C. J., Venstrom, K. A., and Reichardt, L. F. (1991). Vitronectin and thrombospondin promote retinal neurite outgrowth: developmental regulation and role of integrins. Neuron 6, 345-358. Pasqualini, R., Bodorova, J., Ye, S., and Hemler, M, E. (1993). A study of the structure, function and distribution of J3s integrins using novel anti-~s antibodies. J. Cell Sci. 105, 101-105.
Prieto, A. L., Andersson-Fisone, C., and Crossin, K. L. (1992). Charac- terization of multiple adhesive and counteradhesive domains in the extracellular matrix protein cytotactin. J. Cell Biol. 119, 663-678. Prieto, A. L., Edelman, G. M., and Crossin, K. L. (1993). Multiple inte- grins mediate cell attachment to cytotactin/tenascin. Proc. Natl. Acad. Sci. USA 90, 10154-10158. Reichardt, L. F, and Tomaselli, K. J. (1991). Extracellular matrix mole- cules and their receptors: functions in neural development. Annu. Rev. Neurosci. 14, 531-570. Rein, A., and Rubin, H. (1968). Effects of local cell concentrations upon the growth of chick embryo cells in tissue culture. Exp. Cell Res. 49, 666-678. Riou, J.-F., Umbhauer, M., Shi, D. L., and Boucaut, J .-C. (1992). Tenas- cin: a potential modulator of cell-extracellular matrix interactions dur- ing vertebrate embryogenesis. Biol. Cell 75, 1-9. Salmivirta, M., Elenius, K., Vainio, S., Hofer, U., Chiquet-Ehrismann, F., Thesleff, I., and Jalkanen, M. (1991). Syndecan from embryonic tooth mesenchyme binds tenascin. J. Biol. Chem. 266, 7733-7739. Sokol, R. R., and Rohlf, F. J. (1969). Biometry (San Francisco, Califor- nia: W. H. Freeman and Co.), pp. 205-256. Spring, J., Beck, K., and Chiquet-Ehrismann, R. (1989). Two contrary functions of tenascin: dissection of the active sites by recombinant tenascin fragments. Cell 59, 325-334. Sriramarao, P., Mendler, M., and Bourdon, M. A. (1993). Endothelial cell attachment and spreading on human tenascin is mediated by ~2131 and av~. J. Cell Sci. 105, 1001-1012. Steindler, D. A., Cooper, N. G., Faissner, A., and Schachner, M. (1989). Boundaries defined by adhesion molecules during development of the cerebral cortex: the J 1 tenascin glycoprotein in the mouse somatosen- sory cortical barrel field. Dev. Biol. 131,234-260. Tan, S.-S., Crossin, K. L., Hoffman, S., and Edelman, G. M. (1987). Asymmetric expression in somites of cytotactin and its proteoglycan ligand is correlated with neural crest cell distribution. Proc. Natl. Acad. Sci. USA 84, 7977-7981. Tanaka, H., and Obata, K. (1984). Developmental changes in unique cell surface antigens of chick embryo spinal motor neurons and gan- glion cells. Dev. Biol. 106, 26-37. Timpl, R., Rohdl, H., Ristidi, L., Ott, A., Riber, P. G., and Martin, G. R. (1982). Laminin. Meth. Enzymol. 82, 831-838. Tomaselli, K. J., Doherty, P., Emmett, C. J., Damsky, C. H., Walsh, F. S., and Reichardt, L. F. (1993). Expression of ~1 integrins in sensory neurons of the dorsal root ganglion and their functions in neurite out- growth on two laminin isoforms. J. Neurosci. 13, 4880-4888. Vaughan, L., Huber, S., Chiquet, M., and Winterhalter, K. H. (1987). A major, six-armed glycoprotein from embryonic cartilage. EMBO J. 6, 349-353. Wehrle, B., and Chiquet, M. (1990). Tenascin is accumulated along developing peripheral nerves and allows neurite outgrowth in vitro. Development 110, 401-415. Wehrle-Haller, B., and Chiquet, M. (1993). Dual function of tenascin: simultaneous promotion of neurite growth and inhibition of glial migra- tion. J. Cell Sci. 106, 597-610.
Zisch, A. A., D'Alessandri, L., Ranscht, B., Falchetto, R., Winterhalter, K. H., and Vaughan, L. (1992). Neuronal cell adhesion molecule con- tactin/F11 binds to tenascin via its immunoglobulin-like domains. J. Cell Biol. 119, 203-213.
Note Added in Proof
The data referred to throughout as Weaver et al., submitted, are now in press: Weaver, C. D., Yoshida, C. K., de Curtis, I , and Reichardt, L. F. (1995). J. Neurosci., in press.





























