Embed Size (px)
Citation preview

1 3
Oecologia (2016) 181:971–983DOI 10.1007/s00442-015-3544-7
HIGHLIGHTED STUDENT RESEARCH
The invasive annual cheatgrass releases more nitrogen than crested wheatgrass through root exudation and senescence
Kendalynn A. Morris1,2 · John M. Stark1,2 · Bruce Bugbee3 · Jeanette M. Norton2,3
Received: 25 December 2015 / Accepted: 28 December 2015 / Published online: 21 January 2016 © Springer-Verlag Berlin Heidelberg 2016
occurred despite lower root abundance (7.0 vs. 17.3 g dry root kg−1 soil) and N concentration (6.0 vs. 7.6 g N kg−1 root) in cheatgrass vs. crested wheatgrass. We propose that increases in soil N pool sizes and transformation rates under cheatgrass are caused by higher rates of root exuda-tion or release of organic matter containing relatively large amounts of labile N. Our results provide the first evidence for the underlying mechanism by which the invasive annual cheatgrass increases N availability and establishes positive plant-soil feedbacks that promote its success in western rangelands.
Keywords Bromus tectorum · Invasive grass · Plant-soil feedbacks · Root turnover · Mineralization
Introduction
A large body of literature has established that plant invasion alters ecosystem processes (Vitousek 1990; D’Antonio and Vitousek 1992; Ehrenfeld 2003). Despite calls for experi-ments which elucidate the mechanisms by which these impacts are realized (Kulmatiski et al. 2008; Jeschke et al. 2012; van der Putten et al. 2013), few studies examine the mechanisms for competition or plant-soil feedback effects on invasion success. One of the main reasons for this dis-parity is that many of the proposed mechanisms take place in the soil, making them inherently difficult to study. Soil-based mechanisms for invasive plant success include pro-duction of allelopathic compounds (Callaway and Ridenour 2004; Bongard 2012), alteration of soil microbial com-munities (Belnap and Phillips 2001; Bongard 2012), and shifts in soil nutrient cycling (Ehrenfeld 2003; Levine et al. 2003). These kinds of interactions have been documented in invasive plants varying from long-lived trees (Meinhardt
Abstract Plant-soil feedbacks are an important aspect of invasive species success. One type of feedback is alteration of soil nutrient cycling. Cheatgrass invasion in the western USA is associated with increases in plant-available nitrogen (N), but the mechanism for this has not been elucidated. We labeled cheatgrass and crested wheatgrass, a common per-ennial grass in western rangelands, with 15N-urea to deter-mine if differences in root exudates and turnover could be a mechanism for increases in soil N. Mesocosms contain-ing plants were either kept moist, or dried out during the final 10 days to determine the role of senescence in root N release. Soil N transformation rates were determined using 15N pool dilution. After 75 days of growth, cheatgrass accu-mulated 30 % more total soil N and organic carbon than crested wheatgrass. Cheatgrass roots released twice as much N as crested wheatgrass roots (0.11 vs. 0.05 mg N kg−1 soil day−1) in both soil moisture treatments. This
Communicated by Tim Seastedt.
Our results provide the initial evidence for the underlying mechanism by which cheatgrass increases soil N availability. This manuscript represents the thesis work of the first author.
Electronic supplementary material The online version of this article (doi:10.1007/s00442-015-3544-7) contains supplementary material, which is available to authorized users.
* Kendalynn A. Morris [email protected]
1 Department of Biology, Utah State University, Logan, UT 84322, USA
2 Ecology Center, Utah State University, Logan, UT 84322, USA
3 Department of Plants, Soils, and Climate, Utah State University, Logan, UT 84322, USA

972 Oecologia (2016) 181:971–983
1 3
and Gehring 2011) to annual forbs (Batten et al. 2006). Invasive annual grasses in particular are well documented in their ability to alter soil processes (D’Antonio and Vitousek 1992; Hawkes et al. 2005; Parker and Schimel 2010; Perkins and Nowak 2013). A major reason for this may be the change in plant life history characteristics com-pared to those of native perennial vegetation (Ehrenfeld 2003). However, it is not clear if differences in life history characteristics (i.e., annual vs. perennial) can explain many of the effects on soil processes or if other plant characteris-tics may play a role.
Cheatgrass (Bromus tectorum L.) is an invasive annual grass in the western USA that appears to alter soil nitrogen (N) cycling (Bolton et al. 1990; Evans et al. 2001; Blank and Morgan 2011). The direction of this shift is not always consistent, with a few studies finding that cheatgrass soils had lower inorganic N pools and cycling rates (Bolton et al. 1990; Concilio et al. 2015), and many others find-ing increases in pools and cycling rates (Booth et al. 2003; Norton et al. 2004; Adair and Burke 2010). However, these variable findings may be related to the soil conditions pre-sent prior to cheatgrass establishment or to the amount of time passed since invasion (Germino et al. 2015). While in some cases it is likely that cheatgrass has invaded sites that had large and rapidly cycling N pools prior to establish-ment, a recent paper describing a 24-year-long experiment where plant species were randomly assigned to replicated field plots provides the strongest evidence that cheatgrass increases soil N pool sizes and cycling rates relative to native vegetation (Stark and Norton 2014). Several studies hypoth-esize that these changes are caused by the unique chemistry of cheatgrass inputs to soil (Saetre and Stark 2005; Hooker and Stark 2008), and that these inputs establish a soil legacy that benefits future cheatgrass generations (Blank and Mor-gan 2013; Hamman and Hawkes 2013) because cheatgrass’s competitive ability increases with increasing N availabil-ity (Mangla et al. 2011). Due to the limited contribution of aboveground litter to belowground nutrient dynamics in arid systems (Austin and Vivanco 2006), the driver of these changes likely lies in some aspect of cheatgrass roots or root exudation. Understanding the mechanisms that allow cheat-grass to establish positive plant-soil feedbacks will improve our ability to model soil processes in invaded systems and possibly limit future invasions.
Crested wheatgrass (Agropyron cristatum L. Gaertn) is an introduced grass species that has been planted exten-sively as part of pasture improvement projects in sagebrush steppe ecosystems (Lesica and DeLuca 1996; Vaness and Wilson 2007; Fansler and Mangold 2011). As a perennial bunchgrass, crested wheatgrass superficially resembles the native grass species of the USA Intermountain West; however, crested wheatgrass alters soil carbon (C) and N in ways that differ from cheatgrass and native vegetation
(Chen and Stark 2000; Krzic et al. 2000). For example, studies in the Great Basin of western USA have found con-centrations of soil C and N under crested wheatgrass to be intermediate between those of cheatgrass and sagebrush-native perennial grass soils (Hooker et al. 2008), but soil respiration rates were lower beneath crested wheatgrass (Norton et al. 2012). Recent work by Blank et al. (2015) has demonstrated the importance of soil nutrients in com-petitive interactions between cheatgrass and crested wheat-grass, and crested wheatgrass is capable of out-competing cheatgrass in some circumstances (Francis and Pyke 1996; Blank et al. 2015). Therefore the relative ability of these two species to modify soil characteristics and create either positive or negative plant-soil feedbacks is of interest.
The goal of this study was to evaluate the mechanisms by which cheatgrass and crested wheatgrass modify soil N cycling. We examined two potential mechanisms: differ-ences between plant species in the quality of root-derived substrates released and available to soil microbes, and dif-ferences in rates of plant root senescence in response to soil drying. In the first case, root-derived substrates with a greater lability or lower C:N ratio could lead to greater N mineralization (release of excess N by soil microbes), which increases N availability to plants. In the second mechanism, greater root turnover following soil drying may increase the total quantity of plant material released to the soil. Since decomposition of plant roots frequently stimulates net N mineralization regardless of root C:N (Par-ton et al. 2007; Hooker and Stark 2008), greater root senes-cence should also increase microbial activity and N-cycling rates. Under field conditions, cheatgrass senesces as soils dry and eventually 100 % of the root mass turns over. In contrast, perennial plants maintain a substantial portion of their root system throughout the dry season, and only a por-tion of the root mass turns over. Therefore, root senescence in annuals should stimulate soil microbial activity and N cycling to a greater degree than in perennials.
To quantify the contribution of rhizosphere inputs and root senescence to N cycling in soils under cheatgrass and crested wheatgrass, we grew individuals of both species in mesocosms and labeled them with a 15N tracer. Two soil moisture treatments, a continuously moist treatment and a late-onset drying treatment meant to stimulate root senes-cence, were applied in order to separate release of root N in exudates from release of root N through senescence. The isotope tracer allowed us to measure the mass of N that flowed into soil from root exudation or turnover. In addi-tion, we used an ammonium (NH4
+) 15N pool dilution assay to measure soil N transformation rates and microbial respiration, and estimate the C:N of microbial substrates.
We expected that cheatgrass roots would exude com-pounds with greater N content when compared with crested wheatgrass, and that soils under cheatgrass would exhibit

973Oecologia (2016) 181:971–983
1 3
faster gross N mineralization rates and have larger inorganic N pools than those under crested wheatgrass. We also expected that in dry soils root senescence would increase N mineraliza-tion rates for soils under both plant species because substrate availability exerts a dominant control on mineralization rates (Booth et al. 2005), but that this would happen to a greater degree in cheatgrass soils. Lastly, we expected that senescing roots would have lower C:N ratios than healthy roots due to root respiration decreasing C content of root tissues.
Materials and methods
Mesocosms
Plants were grown in mesocosms constructed of untreated pine with Plexiglas fronts and backs (Fig. S1) and drainage holes drilled into the bottom. Within each mesocosm, a fine mesh partition (US 100 mesh, 0.14-mm openings) vertically separated a 3-cm-thick layer of soil from the rest of the mes-ocosm. Soil was divided in this way to prevent roots from growing into this section of soil and provide data on bulk soil N cycling for each mesocosm; however, fine roots pene-trated the mesh divider in mesocosms of both plant species. Because the main goal of this study was to measure effects of the two plant species and soil moisture treatments on soil N cycling rather than the effect of root abundance, only the data from soil in the main compartment are presented.
Mesocosms were filled with soil composed of a 9:1 mix-ture (by volume) of a baked clay aggregate and soil from a local sagebrush steppe ecosystem. The sagebrush soil was collected from the Green Canyon Ecology Research Center in North Logan, Utah, in April 2012, sieved (2-mm mesh) and stored, field moist at 4 °C until use. This soil is a rocky Mollisol (loamy-skeletal, carbonatic, mesic Typic Haplox-eroll) formed on alluvial fan material (Southard et al. 1978) and provided an inoculum of soil microbes and a small amount of organic matter (~0.14 g organic C kg−1 soil). The baked-clay aggregate Profile (Table S1; Profile Prod-ucts LLC, Buffalo Grove, IL), which comprised the major-ity of the mesocosm soil, is a common greenhouse growth medium (Adams et al. 2014) and was used in this study to provide high cation exchange capacity and sufficient aera-tion. Prior to mixing with the inoculant soil, the aggregate was rinsed with tap water and soaked in a concentrated nutrient solution (Table S2) following the recommendations of Adams et al. (2014) to minimize differences between 50-lb bags and reduce potentially high manganese loads.
Plants
Cheatgrass seeds were hand-collected from a population in Cache County, Utah (41°46′07″N, 111°47′11″W). Crested
wheatgrass (Kirk variety) seeds were purchased from Gran-ite Seed (Lehi, UT). Seeds of both species were stored at 4 °C for 2 weeks until mesocosms were ready for plant-ing. Storing seeds in this manner encourages cheatgrass to flower when grown in a greenhouse (Meyer et al. 2004). Seeding and germination of the two species was staggered with crested wheatgrass seeded 4 May 2012 and germi-nated 10 May and cheatgrass seeded 26 May and germi-nated 31 May. Greenhouse growing conditions were 25 °C day, 20 °C night, and a natural light photoperiod (Logan, UT). Mesocosms were grouped by plant species but rand-omized by soil moisture treatment within a 2 × 4-m area. Seedlings were thinned so that each mesocosm contained six plants. Plants were watered with 250 mL of dilute nutri-ent solution (Table S2) every 2 weeks. Fertilization was used to aid plant growth because of the very low organic matter content and nutrient supplying capacity of the soil mixture used. Fertilizer contained only NH4
+ as N to maxi-mize the detectability of nitrification.
Watering
To ensure that the plants and soil microbes in each meso-cosm experienced similar moisture conditions, mesocosms were watered every 1–3 days based on the mass lost by evapotranspiration. To do this, an estimate of mesocosm mass with the soil at maximum water-holding capacity (container capacity) was made for each unit. Mass at maxi-mum water-holding capacity was determined by watering mesocosms until water flowed freely from the bottom and then allowing them to drain for 12 h before weighing. Each day mesocosm mass was recorded and subtracted from the maximum capacity for that specific mesocosm. Mesocosms were watered up to maximum capacity whenever at least 250 g (about 10 % of the total water held) of mass had been lost. To adjust for accumulating plant biomass, mass at maximum capacity was determined three times during the course of the experiment. Soil at maximum water-holding capacity held roughly 0.65 kg H2O kg−1 dry soil.
Soil drying treatment
To examine the effect of soil drying and root senescence on N cycling, a drying treatment was applied to five ran-domly selected mesocosms of each plant species. The dry treatment was initiated 10 days prior to harvesting (i.e., after 65 days of plant growth). Because of the method used to water mesocosms (described above), daily water loss due to evapotranspiration was known for each mesocosm. These data were used to calculate the mass lost (g H2O) to evapotranspiration in 48 h. This value was averaged across the previous 2 weeks to give an average 48-h evapotranspi-ration loss, which ranged from 260 to 560 g of H2O lost

974 Oecologia (2016) 181:971–983
1 3
48 h−1. For the drying treatment, half of this mass (130–280 g H2O) was added to mesocosms every other day. This meant that dry-treatment mesocosms were only receiving half of their daily water requirement for 10 days prior to harvesting. Therefore each day would have resulted in an increasing water deficit. When dry-treatment mesocosms were watered, half of the water was injected into the soil about 10 cm below the surface to allow the soil to gradu-ally dry throughout the mesocosm. This was done using a syringe and an 18-gauge double side-port needle.
15N isotopic labeling of plants
A 15N tracer was used to track the flow of plant N into soil N pools. Plants in both soil moisture treatments were labeled with 15N using the cotton wick method (Russell and Fillery 1996). After 68 days of plant growth, cotton wicks (cotton polyester blend thread) of 0.8 mm diameter (1/32 inch) were passed through one culm of each plant using a large sewing needle (size 110/18, used for sewing leather or heavy denim). The ends of the thread on either side of the culm were passed through small diameter rubber tubing to prevent evaporative losses. Tape was wrapped around the culm to prevent splitting and was also used to help secure tubing. Both ends of the thread were submerged in a small vial containing a 6.01 mM 15N-urea solution. Urea is com-monly used in the cotton wick method because it is read-ily transported throughout the plant (Palta et al. 1991). The cotton wick method has previously been shown to produce detectable amounts of 15N within roots and rhizosphere soil (Hertenberger and Wanek 2004; Yasmin et al. 2006).
Because of the relative novelty of this technique for labe-ling belowground N pools, we performed a trial experiment comparing the efficacy of solutions of potassium nitrate (KNO3) and urea with equal enrichment (98 atom % 15N) and N concentration (0.1 mM N). The urea solution led to greater 15N enrichment in plant roots and surrounding soil, therefore we used a 98 atom % 15N urea solution for the main experiment. The cotton wicks were left in place for 3 days to allow for sufficient quantities of label to enter the plants. Mesocosms were harvested 4 days after the end of labeling (7 days after the start of labeling) because previous experiments found that it takes up to a week for the label to reach its maximum distribution throughout the plant (Palta et al. 1991).
Background 15N enrichment of soil N pools and plant tissues was obtained from four unlabeled, control meso-cosms (two of each grass species) that were established at the start of the experiment. These were identical to meso-cosms with moist-soil watering regimes, with the excep-tion that when labeling began, plants in control meso-cosms received a natural abundance urea solution (0.365 % 15N). These control mesocosms were sampled in the same
manner as treatment mesocosms but data from these mes-ocosms were used solely to determine background 15N enrichment values.
Sampling
Mesocosms were harvested 7 days after the start of 15N labeling and 10 days after the start of the drying treatment. Within each mesocosm, buffer strips of soil were excluded from sampling along the sides (3 cm), top (2 cm), and bot-tom (10 cm) to reduce variability from edge effects such as chemical leaching from the wood, surface drying, and satu-rated conditions at the bottom of the mesocosm.
After the mesocosm sides had been detached, above-ground plant biomass was clipped at the soil surface. Plant tissues were washed clean of excess soil and oven dried for 48 h at 65 °C. Soils were sieved (2 mm) to remove roots and subsamples of soil were used to determine gravimetric water content, extractable organic and inorganic N pools, microbial biomass N, and N transformation rates. The loose texture of the soil lent itself readily to sieving and root separation, but any visible root material passing through the sieve was removed by hand. To determine gravimetric water content, subsamples of soil were dried at 105 °C for 24 h. One set of subsamples was extracted immediately in 0.5 M potassium sulphate (K2SO4; 60 g field-moist soil in 200 mL of extractant) for determining soil N pools. Soil extracts were filtered through pre-leached Whatman no. 4 filter paper. An additional 60-g sample was fumigated with ethanol-free chloroform for 3 days and extracted to deter-mine the microbial flush N pool size and 15N enrichment (Robertson et al. 1999).
NH4+-15N pool dilution was used to determine gross N
cycling rates (Hart et al. 1994). Briefly, 7 mL of 15N labeled solution {80 mg N L−1 of ammonium sulfate [(NH4)2SO4] at 98 atom % 15N} was added to 200 g of sieved mesocosm soil. To that same soil, additional water was added to wet the soil to 0.65 kg water kg−1 dry soil (approximate water-holding capacity). The 15N solution and water were thor-oughly mixed into the soil and the soil was immediately divided into three separate subsamples. One approximately 60-g subsample was immediately extracted in 200 mL of 0.5 M K2SO4 (as described previously) to determine time-zero NH4
+ pool size and 15N enrichment. The other two subsamples were put into plastic cups that were sealed into 1-L mason jars with rubber septa in the lids and incubated in the dark at 23 °C for 2 days. At the end of the second day, headspace gas samples were collected using a syringe and analyzed for CO2 concentration (by gas chromatogra-phy with a thermal conductivity detector) to determine C mineralization rates over 2 days. One soil subsample was extracted in 0.5 M K2SO4 to determine the size and 15N enrichment of the NH4
+ pool after 2 days, and the second

975Oecologia (2016) 181:971–983
1 3
subsample was fumigated with chloroform to determine the microbial flush N pool size and 15N enrichment at 2 days. The size of the microbial N flush (a proxy for microbial biomass N) was determined by subtracting total extractable N (organic N plus inorganic N) of non-fumigated samples from total extractable N in chloroform-fumigated samples (Robertson et al. 1999).
Laboratory analysis
Soil extracts were analyzed for inorganic N [NH4+ and
nitrate (NO3−)] and organic N concentrations. Extractable
NH4+ and NO3
− were measured using a Lachat Quick-Chem 8500 Series 2 Colorimetric Analyzer (Lachat Instru-ments, Milwaukee, WI) by the indophenol and cadmium-reduction methods, respectively. To determine extractable organic N, extracts were persulfate digested as described in Cabrera and Beare (1993) followed by colorimetric analysis.
Soil extracts were prepared for analysis of 15N abun-dance using a diffusion procedure (Stark and Hart 1996). This method uses acidified filter paper “traps” enclosed in Teflon tape to capture ammonia gas from alkalized soil extracts in a solid form. To diffuse soil extracts for NO3
−, Devarda’s alloy was added to convert NO3− to
NH4+ after diffusion for NH4
+ was complete. The filter papers were then analyzed for N content and 15N enrich-ment by continuous-flow direct-combustion mass spec-trometry using a Europa 20-20 system (PDZ, Crewe, UK).
Because mass spectrometry results are most accu-rate when the sample mass is greater than 20 µg N, some samples with low concentrations of N were supplemented or “spiked” with additional N mass. The mass and 15N content of the spike is known and therefore can be sub-tracted from the measured data to determine the original sample N content and 15N enrichment. The majority of samples that were diffused for NO3
− were of sufficiently low mass (<12 µg N in the total volume of extract) that a spike was used. The spike was 20 µg depleted N (99.99 atom % 14N) from (NH4)2SO4 in a 2.5 µg µL−1 solution. Additional samples were diffused containing either KNO3 standards, (NH4)2SO4 spikes, or both. These samples were used to check data for potential differential recovery dur-ing diffusion of NH4
+ and NO3− in spiked diffused sam-
ples. Extracts that had been persulfate digested had very low recoveries following diffusion, and as a result it was not possible to determine the 15N enrichment of extractable organic N pools before and after chloroform fumigation.
Whole, dry soils were ground to a fine powder using a mortar and pestle. Calcium carbonate was removed using the acid fumigation procedure of Harris et al. (2001), by wetting 60-mg dried samples of soil with 50 µL of water
and fumigating them with concentrated hydrochloric acid for 6 h. Randomly selected subsamples of dry plant tissues were ground to a fine powder using a mortar and pestle. Plant and soil samples were analyzed for C and N content and 15N enrichment, and diffused filter papers were ana-lyzed for N content and 15N enrichment by continuous-flow direct-combustion mass spectrometry. Additional dry plant tissue samples were sent to Dairy One Cooperative (Ithaca, NY) for acid detergent analysis of lignin and cellulose concentrations.
Root N export model
A 15N tracer model was developed in Microsoft Office Excel 2010 (Microsoft, Redmund, WA) to estimate the flow of N from roots into soil N pools over the 7 days between the start of 15N labeling of plants and the time of harvest. This model used the measured 15N enrichment (atom % 15N excess) of plant roots and the amount of 15N above back-ground in soil from each mesocosm to calculate the rate at which N moved from roots to soil N pools. Because our goal was to model movement of root N into all soil pools, we first considered using the 15N content of the total soil N pool; however, the sensitivity of this measure is low, and in some cheatgrass mesocosms total soil N was not detectably enriched in 15N (see “Results”). A more sensitive measure of the amount of 15N moving into soil is the extractable N following chloroform fumigation, since this includes extractable organic and inorganic N plus a portion of the microbial N pool. However, low recoveries during diffusion of persulfate digests prevented us from obtaining the 15N enrichment of this pool. Instead, we used the 15N from the NO3
− pool and the NH4+ pool following chloroform fumi-
gation. Previous experiments have shown that NH4+ pools
increase two- to threefold following fumigation, likely due to the release of organic N by deaminase enzymes.
All 15N data were converted to atom % 15N excess by subtracting background 15N enrichments (from non-15N-la-beled mesocosms) from measured enrichments. In our model, we assumed that N exported from roots had the same 15N enrichment as the whole root. This assumption is consistent with other studies in the field (Janzen 1990; Jensen 1996; Arcand et al. 2013). We also assumed that the increase in enrichment of roots over time followed the same pattern in both grass species, increasing linearly from background 15N levels to the final measured 15N enrich-ment over 3 days, then holding steady at the final measured enrichment for the remaining 4 days. This assumption was tested using a trial experiment where the shape and slope of the line tracking relative enrichment of root tissue over time was found to be statistically identical between the two grass species (data not shown). Using these assumptions a tracer model was fit (Stark 2000). This model uses the root

976 Oecologia (2016) 181:971–983
1 3
enrichment but not root mass and thus results are independ-ent of root abundance. In the tracer model, flow of N from the root to the soil sink at each time step was expressed as:
where Sm is the sink pool mass (mg N kg−1 soil), Se is the sink pool enrichment (atom % 15N excess), and Re is the root pool enrichment. We initialized the model at time-zero with root and soil N at 0 atom % 15N excess and allowed N to move from plant roots to soil N pools at an arbitrar-ily chosen rate in 1-h time steps for 168 h. We adjusted the rate at which N left the roots until the final mass of 15N in the sink pool estimated by the model matched the mass of 15N measured in the post-fumigation inorganic N pool. This was done using an iterative procedure (command Goal Seek) that tested values until the difference between model-predicted 15N mass and measured 15N mass was zero. Final rates were expressed in milligrams N/kilogram soil per day. These rates were also converted to a root mass basis by dividing by the mass of dry roots per kilogram soil.
Calculation of net and gross N transformation rates
Net and gross soil N transformation rates, measured in laboratory assays of subsamples collected from all meso-cosms, were calculated using the following equations. Net rates of ammonification, nitrification, and N mineralization were calculated as:
where Pt is the pool size (mg N kg−1 soil) of either NH4+
(for calculation of net ammonification), NO3− (for net nitri-
fication), or NH4+ plus NO3
− (for net N mineralization) at time-t (2 days in this study) and P0 is the correspond-ing pool size at time-zero. Gross rates were calculated using pool sizes and enrichments as described in Hart et al. (1994). These were:
where P0 is the NH4+ pool size at time-zero, Pt is the pool
size at time-t, I0 is the 15N excess enrichment of the NH4+
pool at time-zero, and It is the 15N excess enrichment at time-t. Because soils received some 15N from labeled plant roots, background enrichment was not equal to 0.365 % 15N and differed for each mesocosm. To adjust for this, “background” enrichments measured in NH4
+ pools from 15N-urea labeled mesocosms prior to adding additional 15NH4
+ were used in gross rate calculations.
NFlux =Sm × Se
Re
Net Flux =Pt − P0
t
Gross N Mineralization Rate (GMR) =P0 − Pt
t×
log I0It
log P0Pt
and
Statistical analysis
ANOVA for a 2 × 2 factorial design was used to test com-binations of the two plant species of interest (cheatgrass and crested wheatgrass) and two soil moisture conditions (moist and dry). Soil N and C pools, ratios, and transfor-mation rates were compared between the two plant species and soil moisture treatments. Treatments had five repli-cates for a total of 20 mesocosms, but a single outlier was dropped from the moist soil × crested wheatgrass treat-ment in the final model. This outlier was removed because most values were two to three orders of magnitude greater than other values for the same treatment; its removal had little effect on significant vs. non-significant results, except for some loss of interactions. In the ANOVA model, plant species and soil moisture were treated as fixed factors using the PROC MIXED procedure (SAS version 9.3, Cary, NC). Statistical differences were considered significant for α = 0.05.
Results
Plants and soils
After 75 days of growth, the two plant species differed in root characteristics. Cheatgrass mesocosms had roughly half the root abundance of mesocosms with crested wheat-grass (Table 1). Root abundance was also marginally differ-ent between soil moisture treatments, with dry soils trend-ing towards greater root abundance (p = 0.08; Table 2). Crested wheatgrass roots had greater N concentrations and lower C:N than cheatgrass roots (Table 1). There was a sig-nificant interaction between plant species and soil moisture treatments in root C:N ratios. While crested wheatgrass roots were unaffected by moisture (C:N = 46 ± 2 in moist soils, 48 ± 2 in dry soils) cheatgrass roots had lower C:N in dry soil (56 ± 3) than in moist (66 ± 2, pinteraction = 0.012, F = 8.01,15). Cheatgrass roots had what appeared to be lower lignin and slightly more cellulose than crested wheatgrass roots; however, the differences were not signifi-cant. Soil gravimetric water content at the time of harvest differed between soil moisture treatments (0.58 ± 0.01 kg H2O kg−1 soil for moist soils vs. 0.44 ± 0.01 for dry-down soils, p < 0.0001), but not between plant species.
Gross NH+
4 Consumption Rate = GMR
− Net Ammonification Rate
Gross Inorganic N Immobilization Rate = GMR
− Net N Mineralization Rate.
977Oecologia (2016) 181:971–983
1 3
15N tracer experiment
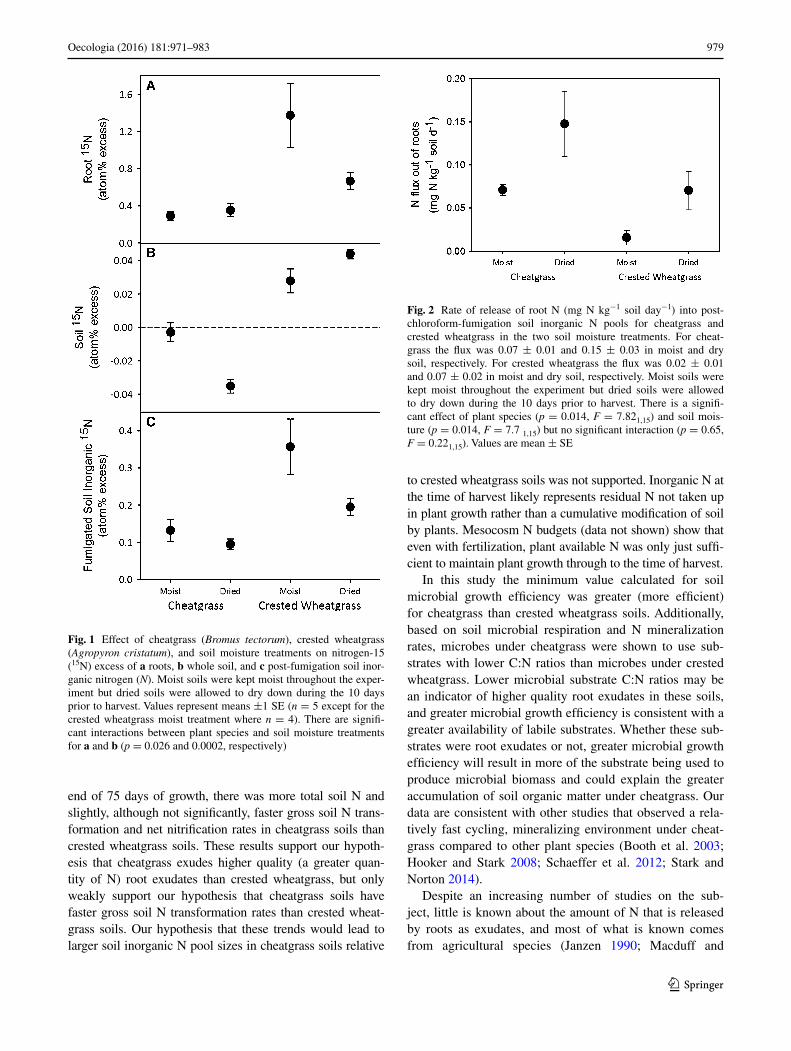
The cotton wick method successfully labeled plant roots and many soil N pools to levels above background 15N enrichment. Cheatgrass roots had lower 15N enrichment than crested wheatgrass roots (p = 0.0004, Fig. 1a). Cheat-grass root enrichment was unaffected by soil moisture treat-ments, but drying lowered enrichment of crested wheat-grass roots (pinteraction = 0.026). Whole soil 15N enrichment differed by plant species and soil moisture treatment. Crested wheatgrass soils had higher 15N enrichment than cheatgrass soils (p < 0.0001, Fig. 1b). In crested wheatgrass mesocosms, the soil dry-down treatment increased whole soil 15N enrichment relative to soils that were kept moist, while in cheatgrass soils, enrichment in both moisture treat-ments was below background enrichment determined from unamended mesocosms.
Because the sensitivity of total soil N measurement was not great enough to detect 15N enrichments in most cheatgrass soils, we used the 15N enrichments of the post chloroform-fumigation inorganic N pools to model the flow of N from roots into soil N pools during the 7-day
labeling period (Fig. 1c). In contrast to total soil N pools, 15N enrichments in the post chloroform-fumigation inor-ganic N pool were significantly enriched for both plant species and moisture treatments. In spite of crested wheat-grass soils having more 15N, the tracer model showed that cheatgrass roots released approximately twice as much N into these pools (0.110 ± 0.021 mg N kg−1 soil day−1) as crested wheatgrass roots (0.046 ± 0.015 mg N kg−1 soil day−1, p = 0.0135; Fig. 2). This outcome is due to the fact that cheatgrass roots had substantially lower 15N enrichments, and thus more root N had to be released to account for the 15N found in the soil pools. The tracer model also showed that roots in moist soils released less N (0.047 ± 0.010 mg N kg−1 soil day−1) than roots in dried soils (0.109 ± 0.023 mg N kg−1 soil day−1, p = 0.0142; Fig. 2).
Plant effects on soil pools and transformation rates
Soils under cheatgrass had nearly one third more total soil N and organic C than soils under crested wheatgrass (Table 1). The C:N ratio of soil organic matter was the
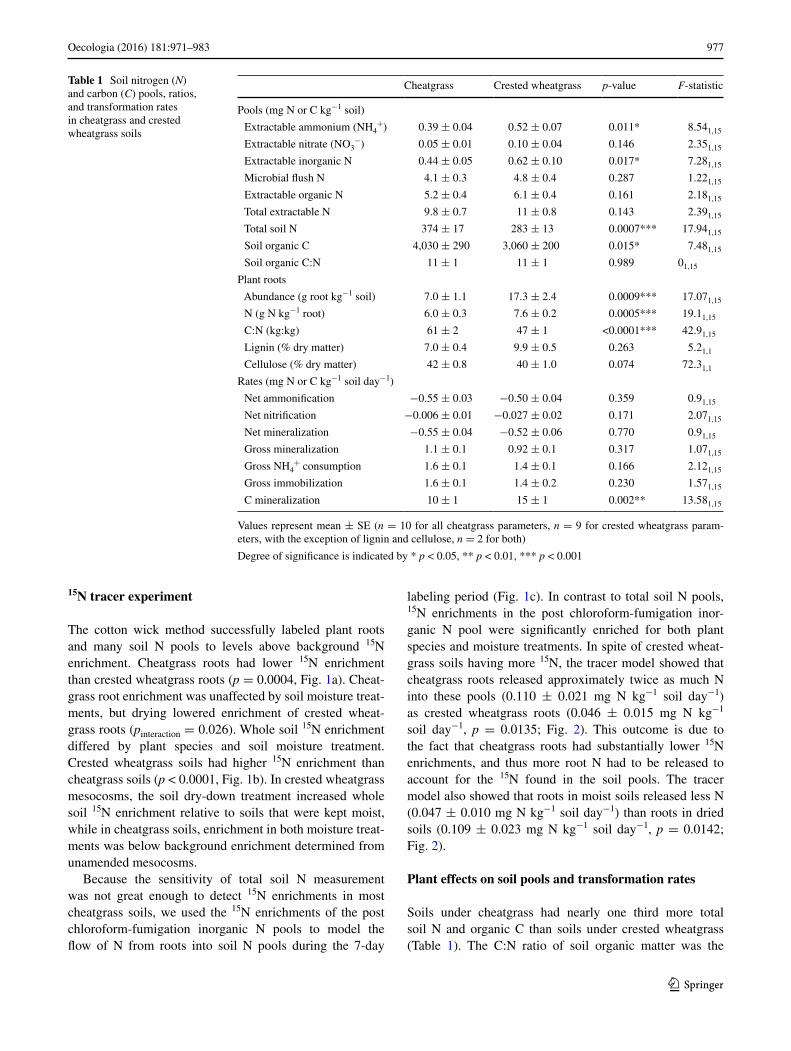
Table 1 Soil nitrogen (N) and carbon (C) pools, ratios, and transformation rates in cheatgrass and crested wheatgrass soils
Values represent mean ± SE (n = 10 for all cheatgrass parameters, n = 9 for crested wheatgrass param-eters, with the exception of lignin and cellulose, n = 2 for both)
Degree of significance is indicated by * p < 0.05, ** p < 0.01, *** p < 0.001
Cheatgrass Crested wheatgrass p-value F-statistic
Pools (mg N or C kg−1 soil)
Extractable ammonium (NH4+) 0.39 ± 0.04 0.52 ± 0.07 0.011* 8.541,15
Extractable nitrate (NO3−) 0.05 ± 0.01 0.10 ± 0.04 0.146 2.351,15
Extractable inorganic N 0.44 ± 0.05 0.62 ± 0.10 0.017* 7.281,15
Microbial flush N 4.1 ± 0.3 4.8 ± 0.4 0.287 1.221,15
Extractable organic N 5.2 ± 0.4 6.1 ± 0.4 0.161 2.181,15
Total extractable N 9.8 ± 0.7 11 ± 0.8 0.143 2.391,15
Total soil N 374 ± 17 283 ± 13 0.0007*** 17.941,15
Soil organic C 4,030 ± 290 3,060 ± 200 0.015* 7.481,15
Soil organic C:N 11 ± 1 11 ± 1 0.989 01,15
Plant roots
Abundance (g root kg−1 soil) 7.0 ± 1.1 17.3 ± 2.4 0.0009*** 17.071,15
N (g N kg−1 root) 6.0 ± 0.3 7.6 ± 0.2 0.0005*** 19.11,15
C:N (kg:kg) 61 ± 2 47 ± 1 <0.0001*** 42.91,15
Lignin (% dry matter) 7.0 ± 0.4 9.9 ± 0.5 0.263 5.21,1
Cellulose (% dry matter) 42 ± 0.8 40 ± 1.0 0.074 72.31,1
Rates (mg N or C kg−1 soil day−1)
Net ammonification −0.55 ± 0.03 −0.50 ± 0.04 0.359 0.91,15
Net nitrification −0.006 ± 0.01 −0.027 ± 0.02 0.171 2.071,15
Net mineralization −0.55 ± 0.04 −0.52 ± 0.06 0.770 0.91,15
Gross mineralization 1.1 ± 0.1 0.92 ± 0.1 0.317 1.071,15
Gross NH4+ consumption 1.6 ± 0.1 1.4 ± 0.1 0.166 2.121,15
Gross immobilization 1.6 ± 0.1 1.4 ± 0.2 0.230 1.571,15
C mineralization 10 ± 1 15 ± 1 0.002** 13.581,15
978 Oecologia (2016) 181:971–983
1 3
same for cheatgrass and crested wheatgrass soils (Table 1). Extractable inorganic N pools were extremely small in all soils. The concentration of NH4
+ was slightly but signifi-cantly higher in crested wheatgrass soils. Although mean net nitrification, gross mineralization, gross NH4
+ con-sumption, and gross immobilization rates all appeared faster in cheatgrass soils, no soil N transformation rates dif-fered between plant species (Table 1). Crested wheatgrass soils had significantly greater C mineralization rates than cheatgrass soils (Table 1).
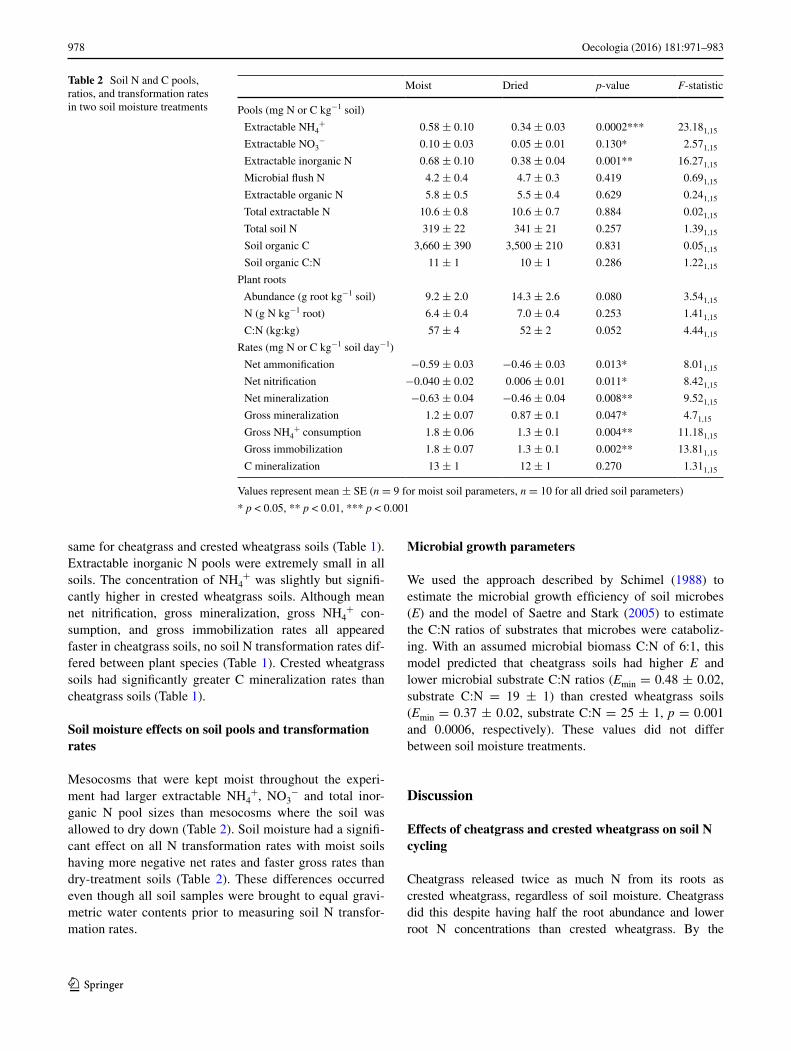
Soil moisture effects on soil pools and transformation rates
Mesocosms that were kept moist throughout the experi-ment had larger extractable NH4
+, NO3− and total inor-
ganic N pool sizes than mesocosms where the soil was allowed to dry down (Table 2). Soil moisture had a signifi-cant effect on all N transformation rates with moist soils having more negative net rates and faster gross rates than dry-treatment soils (Table 2). These differences occurred even though all soil samples were brought to equal gravi-metric water contents prior to measuring soil N transfor-mation rates.
Microbial growth parameters
We used the approach described by Schimel (1988) to estimate the microbial growth efficiency of soil microbes (E) and the model of Saetre and Stark (2005) to estimate the C:N ratios of substrates that microbes were cataboliz-ing. With an assumed microbial biomass C:N of 6:1, this model predicted that cheatgrass soils had higher E and lower microbial substrate C:N ratios (Emin = 0.48 ± 0.02, substrate C:N = 19 ± 1) than crested wheatgrass soils (Emin = 0.37 ± 0.02, substrate C:N = 25 ± 1, p = 0.001 and 0.0006, respectively). These values did not differ between soil moisture treatments.
Discussion
Effects of cheatgrass and crested wheatgrass on soil N cycling
Cheatgrass released twice as much N from its roots as crested wheatgrass, regardless of soil moisture. Cheatgrass did this despite having half the root abundance and lower root N concentrations than crested wheatgrass. By the
Table 2 Soil N and C pools, ratios, and transformation rates in two soil moisture treatments
Values represent mean ± SE (n = 9 for moist soil parameters, n = 10 for all dried soil parameters)
* p < 0.05, ** p < 0.01, *** p < 0.001
Moist Dried p-value F-statistic
Pools (mg N or C kg−1 soil)
Extractable NH4+ 0.58 ± 0.10 0.34 ± 0.03 0.0002*** 23.181,15
Extractable NO3− 0.10 ± 0.03 0.05 ± 0.01 0.130* 2.571,15
Extractable inorganic N 0.68 ± 0.10 0.38 ± 0.04 0.001** 16.271,15
Microbial flush N 4.2 ± 0.4 4.7 ± 0.3 0.419 0.691,15
Extractable organic N 5.8 ± 0.5 5.5 ± 0.4 0.629 0.241,15
Total extractable N 10.6 ± 0.8 10.6 ± 0.7 0.884 0.021,15
Total soil N 319 ± 22 341 ± 21 0.257 1.391,15
Soil organic C 3,660 ± 390 3,500 ± 210 0.831 0.051,15
Soil organic C:N 11 ± 1 10 ± 1 0.286 1.221,15
Plant roots
Abundance (g root kg−1 soil) 9.2 ± 2.0 14.3 ± 2.6 0.080 3.541,15
N (g N kg−1 root) 6.4 ± 0.4 7.0 ± 0.4 0.253 1.411,15
C:N (kg:kg) 57 ± 4 52 ± 2 0.052 4.441,15
Rates (mg N or C kg−1 soil day−1)
Net ammonification −0.59 ± 0.03 −0.46 ± 0.03 0.013* 8.011,15
Net nitrification −0.040 ± 0.02 0.006 ± 0.01 0.011* 8.421,15
Net mineralization −0.63 ± 0.04 −0.46 ± 0.04 0.008** 9.521,15
Gross mineralization 1.2 ± 0.07 0.87 ± 0.1 0.047* 4.71,15
Gross NH4+ consumption 1.8 ± 0.06 1.3 ± 0.1 0.004** 11.181,15
Gross immobilization 1.8 ± 0.07 1.3 ± 0.1 0.002** 13.811,15
C mineralization 13 ± 1 12 ± 1 0.270 1.311,15
979Oecologia (2016) 181:971–983
1 3
end of 75 days of growth, there was more total soil N and slightly, although not significantly, faster gross soil N trans-formation and net nitrification rates in cheatgrass soils than crested wheatgrass soils. These results support our hypoth-esis that cheatgrass exudes higher quality (a greater quan-tity of N) root exudates than crested wheatgrass, but only weakly support our hypothesis that cheatgrass soils have faster gross soil N transformation rates than crested wheat-grass soils. Our hypothesis that these trends would lead to larger soil inorganic N pool sizes in cheatgrass soils relative
to crested wheatgrass soils was not supported. Inorganic N at the time of harvest likely represents residual N not taken up in plant growth rather than a cumulative modification of soil by plants. Mesocosm N budgets (data not shown) show that even with fertilization, plant available N was only just suffi-cient to maintain plant growth through to the time of harvest.
In this study the minimum value calculated for soil microbial growth efficiency was greater (more efficient) for cheatgrass than crested wheatgrass soils. Additionally, based on soil microbial respiration and N mineralization rates, microbes under cheatgrass were shown to use sub-strates with lower C:N ratios than microbes under crested wheatgrass. Lower microbial substrate C:N ratios may be an indicator of higher quality root exudates in these soils, and greater microbial growth efficiency is consistent with a greater availability of labile substrates. Whether these sub-strates were root exudates or not, greater microbial growth efficiency will result in more of the substrate being used to produce microbial biomass and could explain the greater accumulation of soil organic matter under cheatgrass. Our data are consistent with other studies that observed a rela-tively fast cycling, mineralizing environment under cheat-grass compared to other plant species (Booth et al. 2003; Hooker and Stark 2008; Schaeffer et al. 2012; Stark and Norton 2014).
Despite an increasing number of studies on the sub-ject, little is known about the amount of N that is released by roots as exudates, and most of what is known comes from agricultural species (Janzen 1990; Macduff and
Fig. 1 Effect of cheatgrass (Bromus tectorum), crested wheatgrass (Agropyron cristatum), and soil moisture treatments on nitrogen-15 (15N) excess of a roots, b whole soil, and c post-fumigation soil inor-ganic nitrogen (N). Moist soils were kept moist throughout the exper-iment but dried soils were allowed to dry down during the 10 days prior to harvest. Values represent means ±1 SE (n = 5 except for the crested wheatgrass moist treatment where n = 4). There are signifi-cant interactions between plant species and soil moisture treatments for a and b (p = 0.026 and 0.0002, respectively)
Fig. 2 Rate of release of root N (mg N kg−1 soil day−1) into post-chloroform-fumigation soil inorganic N pools for cheatgrass and crested wheatgrass in the two soil moisture treatments. For cheat-grass the flux was 0.07 ± 0.01 and 0.15 ± 0.03 in moist and dry soil, respectively. For crested wheatgrass the flux was 0.02 ± 0.01 and 0.07 ± 0.02 in moist and dry soil, respectively. Moist soils were kept moist throughout the experiment but dried soils were allowed to dry down during the 10 days prior to harvest. There is a signifi-cant effect of plant species (p = 0.014, F = 7.821,15) and soil mois-ture (p = 0.014, F = 7.7 1,15) but no significant interaction (p = 0.65, F = 0.221,15). Values are mean ± SE

980 Oecologia (2016) 181:971–983
1 3
Jackson 1992; Jensen 1996; Paynel et al. 2001; Wichern et al. 2008). Roots exude photosynthetically derived C, in the form of organic acids and sugars, concurrently with N in the form of NH4
+, NO3−, and amino acids (Merbach
et al. 1999; Paynel et al. 2001). These compounds may help recruit beneficial soil microbes, increase nutrient solubility, and contribute to N mineralization (Cardon and Whitbeck 2007; Hinsinger et al. 2009). Thus, greater root exudation in cheatgrass would be likely contribute to greater nutrient availability in the cheatgrass rhizosphere.
Because total soil N was not detectably enriched in 15N and diffusion efficiencies of soil digests were very low, post chloroform-fumigation inorganic 15N was used to estimate the amount of 15N that moved from the plant into the soil. This pool only represents a portion of the total amount of 15N that moved out of the plant roots; however, there is no reason to believe that this proportion would be influenced by plant species. Chloroform fumigation solubilizes micro-bial membranes and other non-polar compounds, and thus releases microbial biomass N into the extractable organic N pool. However, it does not inactivate soil enzymes, and subsequent activity by these enzymes converts substan-tial quantities of organic N to NH4
+. These pools should have been more sensitive to plant effects than total soil N, but because all the possible soil organic N sinks were not accounted for, our rates of root N export must be inter-preted as minimum estimates. The N release rates reported in Fig. 2, showing that cheatgrass roots released twice as much N, are independent of root abundance. However, cheatgrass had substantially lower root abundance than crested wheatgrass, and thus, when N flux is expressed per unit root mass, the differences are even greater: cheat-grass released 15 times as much N (150 μg N g−1 root) as crested wheatgrass (10 μg N g−1 root) over the 7-day labe-ling experiment.
Our results suggest that root N flux was dominated by root exudates in moist soils and the contribution of root turnover was minimal. Estimates of root lifespan vary widely, but the fastest turnover rates are seen in fine roots (Bloomfield et al. 1996). Fine roots of grasses are thought to live between 1 and 20 weeks depending on environmen-tal conditions (Garwood 1967; Bloomfield et al. 1996). Our rate of root N release to soil is based on the isotope tracer that was incorporated into plant tissues over a 7-day period. It seems likely that the vast majority of the 15N label would have moved into physiologically active roots. Furthermore, because most roots live at least 1 week, most of the roots that received label would have remained physiologically active over the 7-day period before harvest.
Another important consideration is the potential re-uptake of exuded N by plant roots (Jones et al. 2005, 2009). The ability of cheatgrass and crested wheatgrass to do this may differ due to their different phenologies. However, the
amount of re-uptake was probably small for both grass spe-cies during the short time period of the tracer experiment, especially when considering that most of the exuded N was organic and plant uptake of organic N is small (Jones et al. 2005). The ability of these two species to take up N that was previously released by their roots may be more impor-tant in the long term: the annual cheatgrass will have no re-uptake at the end of the growing season; the amount for crested wheatgrass is unknown.
Few studies have examined mechanisms underlying alteration of soil N cycling by cheatgrass. A similar study examined rhizosphere-mediated effects of cheatgrass on soil using a mass balance approach to quantify what was called the “rhizosphere priming effect” of cheatgrass on soil N (Concilio et al. 2015). For this, the total N in soil inorganic N pools and plant biomass was compared with the quantity of inorganic N in soil without plants. The dif-ference between these values was assumed to represent the rhizosphere priming effect. Less N was found in the com-bined soil inorganic and plant biomass N pools than in soils without plants, and thus it was concluded that there was a negative rhizosphere priming effect and that belowground processes play a minor role in cheatgrass alteration of soil N dynamics. A similar approach in our study might have led to the same conclusion. However, net change in pool sizes is a poor indicator of gross fluxes (Stark and Hart 1997). A tracer approach would have allowed estimation of rates of N flow through pools and might have shown differ-ences in fluxes even though pool sizes did not substantially change.
Effects of drying on soil N cycling
Somewhat surprisingly, the dry treatment did not appear to stimulate significant amounts of root senescence: soils that were allowed to dry down actually had greater root abun-dance than soils that stayed moist throughout the experi-ment. This is likely because the plants began to experience water stress early in the soil-drying process and responded by producing more roots to increase water acquisition (Pas-sioura 1996). The increase in root abundance in dry soils may also explain why more N was released in soils that were allowed to dry down than in moist soils. However, despite the increase in root abundance, the soil drying treat-ment may have led to some root turnover due to senescence of older, less efficient roots, that was compensated for by new root production. Therefore, we cannot say for certain what proportion of the N flux came from root exudates vs. turnover. Based on overall plant health at the time of har-vest, it seems that 10 days was insufficient to stimulate the kind of senescence needed to test our hypotheses. However, the drying treatment did stimulate root N release. Further-more, as we expected, cheatgrass root C:N ratios fell with

981Oecologia (2016) 181:971–983
1 3
soil drying, possibly due to increased net root respiration and depletion of root C reserves. The combination of nar-rowing C:N ratios and complete root senescence in annual species at the end of the growing season could contribute to even greater N availability in cheatgrass soils.
Other studies investigating the contribution of root-derived N to soil N pools found similarly strong evidence for a positive N flux from roots (Janzen 1990; Jensen 1996). Due to differences in the plant species used, experi-mental design, and units of measure, it is not possible to make direct comparisons to our work. However, both stud-ies found evidence that exudates were readily degraded, leading to mineralization. In addition, Janzen (1990) also proposed that the flux of N was greatest at the end of the growing season, when cumulative root turnover would be highest.
Arid lands often have an accumulation of NO3− in soils
during the driest part of the summer (Jones and Woodman-see 1979; Booth et al. 2003; Sperry et al. 2006). In this study, we attempted to mimic the same decreasing soil moisture that would typically occur at the end of the grow-ing season. However, our drying treatment did not result in larger soil NO3
− pools. What we did observe was a positive net nitrification rate in dry-down soils. In fact, dry-down soils were the only treatment for which the net nitrification rate was positive. The drying treatment was only initiated 10 days before harvest, thus it is possible that net nitrifica-tion would have led to larger NO3
− pool sizes in dry-down soils if treatments had been maintained for a longer dura-tion. Interestingly, net N mineralization rates were also greater in the dry-down soils. Our isotope dilution meas-urements showed that this was not a result of greater gross N mineralization, but due to lower rates of inorganic N immobilization in the dry-down soils.
As an annual, cheatgrass has no need to store N in plant biomass for the next growing season. To facilitate growth of the next generation, N from dead biomass needs to be returned to plant-available forms. Invasive species often have faster growth rates than native vegetation (Ehren-feld 2003). For cheatgrass and other annuals, senescence or exudation of readily degraded tissue may facilitate faster growth by providing soil microbes with the materi-als needed to produce plant-available nutrients. However, the mechanisms by which invasive species modify soil biogeochemistry have not been clearly elucidated. Our results show that characteristics of root tissues resulting in increased rates of N exudation and release lead to increases in soil nutrient pools. Increased organic pools of nutrients will in turn lead to increased cycling rates and N availa-bility (Booth et al. 2005). The magnitude and direction of changes to soil nutrient cycling will vary not only as a con-sequence of the invasive species present, but also as a result of climate and soil properties.
Conclusion
Most invasive plant research focuses on comparing plant and soil performance under a particular set of conditions (competition, increased or decreased soil N, supplemen-tation of the microbial community), but very few studies investigate the mechanism(s) behind differential perfor-mance. In this study, we have identified a potential mecha-nism (i.e., root release of N) by which cheatgrass might increase N availability in soil and establish a positive plant-soil feedback. Although it is likely that cheatgrass more readily establishes in areas with inherently larger pools of plant-available N, multiple lines of evidence, including ours, support the conclusion that cheatgrass also modifies soil N cycling, making inorganic N pools larger and cycling rates faster. Additional research focused on ways of breaking this positive feedback loop may allow us to minimize the success of cheatgrass and other invasive species.
Acknowledgments This research was funded by grants from the Utah Agricultural Experiment Station (project UTAO 588, journal paper no. 8836), the US Department of Agriculture, Biology of Weedy and Inva-sive Plants (agreement no. 2008-35320-18668) and Utah State Univer-sity’s Ecology Center. Thanks goes to Tasha Prettyman and Amanda Stoudt for assistance with sample processing and instrumentation.
Author contribution statement K. M. and J. S. originally con-ceived the idea. K. M., J. S., B. B., and J. N. designed the experi-ment. K. M. performed the experiment and analyzed the data. K. M. wrote the manuscript and J. S., B. B., and J. N. provided editorial assistance.
References
Adair EC, Burke IC (2010) Plant phenology and life span influence soil pool dynamics: Bromus tectorum invasion of perennial C3–C4 grass communities. Plant Soil 335:255–269
Adams C, Jacobson A, Bugbee B (2014) Ceramic aggregate sorption and desorption chemistry: implications for use as a component of soilless media. J Plant Nutr 37:1345–1357. doi:10.1080/01904167.2013.837921
Arcand MM, Knight JD, Farrell RE (2013) Estimating belowground nitrogen inputs of pea and canola and their contribution to soil inorganic N pools using 15N labeling. Plant Soil 371:67–80
Austin AT, Vivanco L (2006) Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation. Nature 442:555–558
Batten KM, Scow KM, Davies KF, Harrison SP (2006) Two invasive plants alter soil microbial community composition in serpentine grasslands. Biol Invasions 8:217–230
Belnap J, Phillips SL (2001) Soil biota in an ungrazed grassland: response to annual grass (Bromus tectorum) invasion. Ecol Appl 11:1261–1275
Blank RR, Morgan T (2011) Evidence that invasion by cheatgrass alters soil nitrogen availability. Nat Resour Environ Issues 17:71–73
Blank RR, Morgan T (2013) Soil engineering facilitates downy brome (Bromus tectorum) growth-a case study. Invasive Plant Sci Man-age 6:391–400

982 Oecologia (2016) 181:971–983
1 3
Blank RR, Morgan T, Allen F (2015) Suppression of annual Bromus tectorum by perennial Agropyron cristatum: roles of soil nitro-gen availability and biological soil space. AoB Plants 7:plv006
Bloomfield J, Vogt K, Wargo PM (1996) Tree root turnover and senes-cence. In: Waisel Y, Eshel A, Kafkafi U (eds) Plant roots: the hid-den half, 2nd edn. Dekker, New York, pp 363–381
Bolton HJ, Smith J, Wildung R (1990) Nitrogen mineralization poten-tials of shrub-steppe soils with different disturbance histories. Soil Sci Soc Am J 54:887–891
Bongard C (2012) A review of the influence of root-associating fungi and root exudates on the success of invasive plants. NeoBiota 14:21–45
Booth MS, Stark JM, Caldwell MM (2003) Inorganic N turnover and availability in annual and perennial-dominated soils in a north-ern Utah shrub-steppe ecosystem. Biogeochemistry 66:311–330. doi:10.1023/b:biog.0000005340.47365.61
Booth MS, Stark JM, Rastetter E (2005) Controls on nitrogen cycling in terrestrial ecosystems: a synthetic analysis of literature data. Ecol Monogr 75:139–157
Cabrera ML, Beare MH (1993) Alkaline persulfate oxidation for determining total nitrogen in microbial biomass extracts. Soil Sci Soc Am J 57:1007–1012. doi:10.2136/sssaj1993.03615995005700040021x
Callaway RM, Ridenour WM (2004) Novel weapons: inva-sive success and the evolution of increased com-petitive ability. Front Ecol Environ 2:436–443. doi:10.1890/1540-9295(2004)002[0436:NWISAT]2.0.CO;2
Cardon ZG, Whitbeck JL (eds) (2007) The rhizosphere: an ecological perspective. Academic Press, San Diego
Chen J, Stark JM (2000) Plant species effects and carbon and nitrogen cycling in a sagebrush–crested wheatgrass soil. Soil Biol Bio-chem 32:47–57
Concilio A, Vargas T, Cheng W (2015) Rhizosphere-mediated effects of the invasive grass Bromus tectorum L. and native Elymus ely-moides on nitrogen cycling in Great Basin Desert soils. Plant Soil 393:245–257
D’Antonio CM, Vitousek PM (1992) Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annu Rev Ecol Syst 23:63–87
Ehrenfeld JG (2003) Effects of exotic plant invasions on soil nutrient cycling processes. Ecosystems 6:503–523
Evans R, Rimer R, Sperry L, Belnap J (2001) Exotic plant inva-sion alters nitrogen dynamics in an arid grassland. Ecol Appl 11:1301–1310
Fansler VA, Mangold JM (2011) Restoring native plants to crested wheatgrass stands. Restor Ecol 19:16–23. doi:10.1111/j.1526-100X.2010.00678.x
Francis MG, Pyke DA (1996) Crested wheatgrass-cheatgrass seed-ling competition in a mixed-density design. J Range Manage pp 432–438
Garwood E (1967) Seasonal variation in appearance and growth of grass roots. Grass Forage Sci 22:121–130
Germino MJ, Belnap J, Stark JM, Allen EB, Rau BM (2015) Ecosys-tem impacts of exotic annual invaders in the genus Bromus. In: Germino MJ, Chambers J, Brown CS (eds) Exotic brome-grasses in arid and semi-arid ecosystems of the Western US: causes, con-sequences, and management implications. Springer, New York
Hamman ST, Hawkes CV (2013) Biogeochemical and microbial lega-cies of non-native grasses can affect restoration success. Restor Ecol 21:58–66. doi:10.1111/j.1526-100X.2011.00856.x
Harris D, Horwáth WR, van Kessel C (2001) Acid fumigation of soils to remove carbonates prior to total organic carbon or carbon-13 isotopic analysis. Soil Sci Soc Am J 65:1853–1856. doi:10.2136/sssaj2001.1853
Hart SC, Stark JM, Davidson EA, Firestone MK (1994) Nitrogen mineralization, immobilization, and nitrification. In: Weaver R,
Angle S, Bottomley P, Bezdicek D, Smith S, Tabatabai A, Wol-lum A (eds) Methods of soil analyses. Part 2. Microbiological and biochemical properties, 3rd edn. Soil Science Society of America, Madison, pp 985–1018
Hawkes CV, Wren IF, Herman DJ, Firestone MK (2005) Plant invasion alters nitrogen cycling by modify-ing the soil nitrifying community. Ecol Lett 8:976–985. doi:10.1111/j.1461-0248.2005.00802.x
Hertenberger G, Wanek W (2004) Evaluation of methods to measure differential 15N labeling of soil and root N pools for studies of root exudation. Rapid Commun Mass Spectrom 18:2415–2425. doi:10.1002/rcm.1615
Hinsinger P, Bengough AG, Vetterlein D, Young IM (2009) Rhizos-phere: biophysics, biogeochemistry and ecological relevance. Plant Soil 321:117–152
Hooker TD, Stark JM (2008) Soil C and N cycling in three semiarid vegetation types: response to an in situ pulse of plant detritus. Soil Biol Biochem 40:2678–2685
Hooker TD, Stark JM, Norton U, Joshua Leffler A, Peek M, Ryel R (2008) Distribution of ecosystem C and N within contrast-ing vegetation types in a semiarid rangeland in the Great Basin, USA. Biogeochemistry 90:291–308
Janzen H (1990) Deposition of nitrogen into the rhizosphere by wheat roots. Soil Biol Biochem 22:1155–1160
Jensen ES (1996) Rhizodeposition of N by pea and barley and its effect on soil N dynamics. Soil Biol Biochem 28:65–71. doi:10.1016/0038-0717(95)00116-6
Jeschke J, Aparicio LG, Haider S, Heger T, Lortie C, Pyšek P, Strayer D (2012) Support for major hypotheses in invasion biology is uneven and declining. NeoBiota 14:1–20
Jones M, Woodmansee R (1979) Biogeochemical cycling in annual grassland ecosystems. Bot Rev 45:111–144
Jones DL, Healey JR, Willett VB, Farrar JF, Hodge A (2005) Dis-solved organic nitrogen uptake by plants—an important N uptake pathway? Soil Biol Biochem 37:413–423
Jones DL, Nguyen C, Finlay RD (2009) Carbon flow in the rhizo-sphere: carbon trading at the soil–root interface. Plant Soil 321:5–33
Krzic M, Broersma K, Thompson DJ, Bomke AA (2000) Soil proper-ties and species diversity of grazed crested wheatgrass and native rangelands. J Range Manage 53:353–358
Kulmatiski A, Beard KH, Stevens JR, Cobbold SM (2008) Plant–soil feedbacks: a meta-analytical review. Ecol Lett 11:980–992
Lesica P, DeLuca TH (1996) Long-term harmful effects of crested wheatgrass on Great Plains grassland ecosystems. J Soil Water Conserv 51:408–409
Levine JM, Vilà M, Antonio CMD, Dukes JS, Grigulis K, Lavorel S (2003) Mechanisms underlying the impacts of exotic plant invasions. Pro R Soc Lond Ser B Biol Sci 270:775–781. doi: 10.1098/rspb.2003.2327
Macduff J, Jackson S (1992) Influx and efflux of nitrate and ammo-nium in Italian ryegrass and white clover roots: compari-sons between effects of darkness and defoliation. J Exp Bot 43:525–535
Mangla S, Sheley R, James J, Radosevich S (2011) Intra and interspe-cific competition among invasive and native species during early stages of plant growth. Plant Ecol 212:531–542. doi:10.1007/s11258-011-9909-z
Meinhardt KA, Gehring CA (2011) Disrupting mycorrhizal mutu-alisms: a potential mechanism by which exotic tamarisk outcompetes native cottonwoods. Ecol Appl 22:532–549. doi:10.1890/11-1247.1
Merbach W, Mirus E, Knof G, Remus R, Ruppel S, Russow R, Gran-see A, Schulze J (1999) Release of carbon and nitrogen com-pounds by plant roots and their possible ecological importance. J Plant Nutr Soil Sci 162:373–383

983Oecologia (2016) 181:971–983
1 3
Meyer SE, Nelson DL, Carlson SL (2004) Ecological genetics of ver-nalization response in Bromus tectorum L. (Poaceae). Ann Bot 93:653–663. doi:10.1093/aob/mch088
Norton JB, Monaco TA, Norton JM, Johnson DA, Jones TA (2004) Soil morphology and organic matter dynamics under cheat-grass and sagebrush-steppe plant communities. J Arid Environ 57:445–466
Norton U, Saetre P, Hooker TD, Stark JM (2012) Vegetation and moisture controls on soil carbon mineralization in semiarid envi-ronments. Soil Sci Soc Am J 76:1038–1047
Palta JA, Fillery IR, Mathews EL, Turner NC (1991) Leaf feeding of N-15 urea for labeling wheat with nitrogen. Aust J Plant Physiol 18:627–636
Parker SS, Schimel JP (2010) Invasive grasses increase nitrogen avail-ability in California grassland soils. Invasive Plant Sci Manage 3:40–47
Parton W, Silver WL, Burke IC, Grassens L, Harmon ME, Currie WS, King JY, Adair EC, Brandt LA, Hart SC (2007) Global-scale similarities in nitrogen release patterns during long-term decom-position. Science 315:361
Passioura JB (1996) Drought and drought tolerance. Plant Growth Regul 20:79–83. doi:10.1007/BF00024003
Paynel F, Murray PJ, Cliquet JB (2001) Root exudates: a pathway for short-term N transfer from clover and ryegrass. Plant Soil 229:235–243
Perkins LB, Nowak RS (2013) Native and non-native grasses gen-erate common types of plant–soil feedbacks by altering soil nutrients and microbial communities. Oikos 122:199–208. doi:10.1111/j.1600-0706.2012.20592.x
Robertson GP, Coleman DC, Bledsoe CS, Sollins P (eds) (1999) Standard soil methods for long-term ecological research. Oxford University Press, New York
Russell C, Fillery I (1996) In situ 15N labelling of lupin below-ground biomass. Aust J Agric Res 47:1035–1046. doi:10.1071/AR9961035
Saetre P, Stark JM (2005) Microbial dynamics and carbon and nitro-gen cycling following re-wetting of soils beneath two semi-arid plant species. Oecologia 142:247–260
Schaeffer SM, Ziegler SE, Belnap J, Evans R (2012) Effects of Bro-mus tectorum invasion on microbial carbon and nitrogen cycling
in two adjacent undisturbed arid grassland communities. Biogeo-chemistry 111:427–441. doi:10.1007/s10533-011-9668-x
Schimel DS (1988) Calculation of microbial growth efficiency from 15N immobilization. Biogeochemistry 6:239–243. doi:10.1007/BF02182998
Southard AR, Wilson LM, Erickson AJ (1978) Chemical and physical properties of the soils of the Cache Valley Area and the Eastern Portion of Box Elder County. Utah, Utah Agric Exp Station
Sperry L, Belnap J, Evans R (2006) Bromus tectorum invasion alters nitrogen dynamics in an undisturbed arid grassland ecosystem. Ecology 87:603–615
Stark JM (2000) Nutrient transformations. In: Sala OE, Jackson RB, Mooney HA, Howarth RW (eds) Methods in ecosystem science. Springer, New York
Stark JM, Hart SC (1996) Diffusion technique for preparing salt solu-tions, kjeldahl digests, and persulfate digests for nitrogen-15 analysis. Soil Sci Soc Am J 60:1846–1855
Stark JM, Hart SC (1997) High rates of nitrification and nitrate turno-ver in undisturbed coniferous forests. Nature 385:61–64
Stark JM, Norton JM (2014) The invasive annual cheatgrass increases ntirogen availability in 24-year-old replicated field plots. Oecolo-gia. doi:10.1007/s00442-014-3093-5
van der Putten WH, Bardgett RD, Bever JD, Bezemer TM, Casper BB, Fukami T, Kardol P, Klironomos JN, Kulmatiski A, Sch-weitzer JA, Suding KN, Van de Voorde TFJ, Wardle DA (2013) Plant–soil feedbacks: the past, the present and future challenges. J Ecol 101:265–276. doi:10.1111/1365-2745.12054
Vaness BM, Wilson SD (2007) Impact and management of crested wheatgrass (Agropyron cristatum) in the northern Great Plains. Can J Plant Sci 87:1023–1028. doi:10.4141/CJPS07120
Vitousek PM (1990) Biological invasions and ecosystem processes: towards an integration of population biology and ecosystem studies. Oikos 57:7–13
Wichern F, Eberhardt E, Mayer J, Joergensen RG, Müller T (2008) Nitrogen rhizodeposition in agricultural crops: methods, esti-mates and future prospects. Soil Biol Biochem 40:30–48
Yasmin K, Cadisch G, Baggs EM (2006) Comparing 15 N-labelling techniques for enriching above- and below-ground compo-nents of the plant-soil system. Soil Biol Biochem 38:397–400. doi:10.1016/j.soilbio.2005.05.011





























